Embed Size (px)
Citation preview

ACTA NEUROBIOL. EXP. 1990, 50: 439-450
Symposium "Recovery from brain damage: behavioral and neurochemical approaches" 4 7 July, 1989, Warsaw, Poland
GANGLIOSIDES AS NEUROTROPHIC AGENTS: STUDIES ON THE MECHANISM OF ACTION
Robert W. LEDEEN I, Gusheng WU I, Mic~hael S. CANNELLA I,
Barbara ODERFELD-NOWAK and A. Claudio CUELLO
Department of Neurology and Biochemistry, Albert Einstein College of Medicine, 1300 Morris Park ,Avenue, F-140, Bronx, New York 10461, USA,
Department of Neurophysiology, Nencki Institute of Experimental Biology, 3 Pasteur St., 02-093 Warsaw, Poland and :Department
of Pharmacology and Therapeutics, McGill University, 3655 D r u m o n d St., Montreal, PQ H3G 1Y6, Canada
Key words: gangliosides, neurons, neurotrophic agents, neuro-%A neuroplastoma cells, PC12 pheochromocytoma cells, embryonic chick dorsal root ganglia, septo- hippocampal model, cortex-nucleus basalis magnocellularis model
Abstract. During the normal course of neuronal differentiation gan- gliosides undergo marked changes in both quantity and quality. These changes, i.e. several-fold increase in concentration and appearance of the gangliotetraose family, have been observed both in vivo and in neuronal cell cultures. In addition to these naturally occurring (endo- genous) manifestations, exogenously administered gangliosides have been observed to .exert neuritogenic and/or neuronotrophic effects on a variety of neuroblastoma cell lines and primary neuronal culture systems. Un- like the endogenous effects, which appear to require gangliotetraose structures, the structural specificity 01 the exogenous effect is quite broad. Thus, all of 11 different gangliosides proved neuritogenic with neuro-2A neuroblastoma cells. Furthermore, synthetic sialoglycolipids possessing a p-ketosidic sialic acid linkage and/or a glyceride-like moiety in place of ceramide all caused enhanced neurite outgrowth in neuro-2A, PC12, and embryonic chick dorsal root ganglia. A derivative of GM1 lacking the negative charg,e was also active in the same 3 systems.

440
These results point to general perturbation of the membrane, probably a physical-chemical effect, in a manner which triggers intracellular events leading to differentiation. In contrast to these in vitro results, use of the above GMl derivative in two in vivo models proved ineffective in maintaining the level of cholinergic markers that were preserved by GM1 in lesioned brains. These results point to fundamentally different me- chanism for the trophic effects of administered gangliosides in vivo and in vitro.
INTRODUCTION
The ganglioside changes accompanying neuronal differentiation are noteworthy for the remarkable increases in gangliotetraose structures which occur during the course of neurite outgrowth and synaptogenesis (24, 26). However, the specific functional roles of gangliosides in the developing and mature neuron are not well understood. Some studies have focussed on the behavior of the cell's natural (i.e. endogenous) gangliosides during these various phases and much has been learned through use of specific cell surface probes and interventive agents. The discovery that exogenously administered gangliosides have neuritogenic and neuronotrophic properties capable of influencing neuronal diffe- rentiation and survival (for review, see ref. 10, 11) has provided another general approach to this aspect of ganglioside function. As it turns out, the two models do not reflect identical phenomena and hence the re- sults obtained with exogenous gangliosides must be interpreted with caution in relation to the cellular function of endogenous gangliosides. Nevertheless, the exogenous effects are worthy of investigation owing to the significant pharmacological properties which administered gan- gliosides exhibit in many in vitro and in vivo systems.
In view of certain apparent similarities in trophic phenomena it has been thought likely that the in vivo and in vitro effects of exogenous gangliosides share a common mechanism, although direct evidence for this is lacking. Little is known, in fact, concerning the mechanistic role of gangliosides as trophic agents in general. One approach to this que- stion, applied thus far to cell culture systems (2, 3, 5, 6, 28), has been the use of different gangliosides and synthetic sialoglycolipids to assess the structural requirements for neuritogenic activity. We have found that a derivative of GM1, in which the negative charge has been re- moved, has proved especially useful in comparing in vivo and in vitro activities (4). We review here results obtained with these and other derivatives and the insight they help to provide concerning neuronotro- phic mechanisms.

STRUCTURAL REQUIREMENTS FOR NEURITOGENESIS
Beginning with reports that exogenous gangliosides added to the B104 neuronal cell linc acted as differentiation and survival factors (15), many laboratories have demonstrated similar ganglioside effects with a variety of neuronal cell lines and primary neuronal cultures. The neu-
Galoctore Glucose
Galactose Glucoae
HNAc
N- Acetylnsurominic acid
epi -GM3
Fig. 1. Structure of epi-GM3, in comparison to naturally occurring GM3. Note in- version of the sialic acid lretosidic linkage from the naturally occurring alpha (GM3)
to the neuraminidase-resistant beta configuration in epi-GM3.

ro-2A mouse neuroblastoma cell line has proved especially useful for this purpose, owing to the prolific outgrowth of processes induced by exogenous gangliosides at concentrations in the micromolar range (20). The physiological significance of this phenomenon was suggested by the formation under favorable conditions of mature synapse-like contacts resembling the mature postsynaptic thickenings found in Gray Type I synapses (21).
To determine the structural requirements for neuritogenic activity of this kind, we first compared the effects of 11 different naturally occurring gangliosides on neuro-2A cells (2). Despite wide variations in structure, ranging from GM, with two sugars to GQlb with nine, all 11 substances proved to be active neuritogenic agents. This was true even in the presence of serum which normally inhibits neurite outgrowth. Subsequent studies with synthetic sialoglycolipids, including "epi-GM3" with a 0-linked sialic acid (Fig. 1) (5) and a "glycero-ganglioside" con- taining sialic acid attached to a diglyceride-like backbone (Fig. 2) (3), revealed these also to be neuritogenic not only toward neuro-2A cells but also dorsal root ganglia and PC12 cells. In the latter case ganglio- sides or synthetic sialoglycolipids proved neuritogenic only in the pre- sence of nerve growth factor, as previously described (8). Additional synthetic sialoglycolipids were shown to enhance neurite outgrowth of neuro-2A cells (28).
The above studies demonstrated that a sphingosine-containing ce- ramide moiety is not necessary, nor is a complex oligosaccharide chain since sialic acid was the sole carbohydrate attached to the lipid backbone in the case of the glycero-ganglioside. The broad spectrum of sialogly- co-lipid structures showing this kind of activity suggests a mechanism involving general membrane perturbation, possibly of a physical-che- mica1 nature. This mechanism would be somewhat independent of car- bohydrate structure, except for an apparent requirement of sialic acid
Fig. 2. Structure of glycero-ganglioside.

Fig. 3. Phase contrast photomicrographs of fixed cultures of 9-day embryonic chick DRG explants after 4Xh incubation with medium alone (A) or medium supplemented with GM, (B), methyl ester of GM, (C) or GM,-AL (D). Photographed at 100 x . Reproduced from ref. 10 with permission.

or related moiety. The latter is inferred from the fact that asialo GM1 and various other neutral glycolipids were without activity toward neu- ro-2A cells (2) (although a few such substances along with other sur- factants were recently claimed to exert trophic effects toward certain primary neurons in culture - see ref. 29). The effect appears specific for lipids since sialo-proteins were without effect, at least toward neu- ro-2A cells (12).
Physical-chemical perturbations of the above postulated type would be expected to require at least moderately high concentrations of sia- lolipids, which may explain why these substances have generally been employed in the range 10-100 pM. The mechanism of this effect could differ fundamentally from that of the "N-type" phenomenon which requires an order-of-magnitude lower concentration (16). Thus far two neuroblastoma lines of the N type have been reported, GOT0 and NB-1 (27). In addition to the lower concentration requirement, these cells showed great selectivity in responding only to GQlb.
USE OF GM1 DERIVATIVES
The principal derivative we employed for comparing in vivo with in vitro trophic effects was the primary alcohol of GMl (termed GM1- AL) generated by NaBH4-reduction of the methyl ester of GMl (4). Application to neuro-2A neuroblastoma cells at the same concentration as GMl (0.12 mM) and to PC12 pheochromocytoma cells at 9 pM (to- gether with nerve growth factor at 5 BU/ml) caused significant in- crease in the percent of neurite bearing cells above controls: neuro-2A, 18O/o vs. 52OIo; PC12, 19OIo to 42OIo. Neurite length of both cell types was also significantly increased. Similar results were obtained with dorsal root ganglia explants from 9-day old chick embryos; at a concentration of 60 yM both GMl and GM1-AL increased the number of neurites exten- ding beyond the leading edge of the non-neuronal migration (Fig. 3).
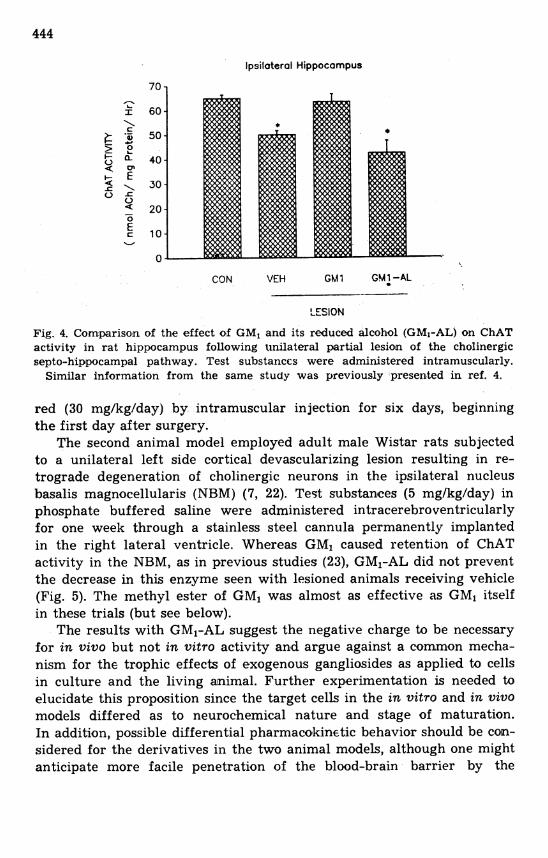
In contrast to the neuritogenic properties of GM1-AL in the above three cell culture systems, two in vivo animal models failed to reveal any trophic activity with this compound. One of these models employed unilateral partial lesion of the septo-hippocampal pathway in adult rats, leading to loss of choline acetyltransferase (ChAT) and acetylcholin- esterase (AChE) in the hippocampus (9, 31). The effect of GMl on hippocampal segments ipsilateral to the lesion was to attenuate reduction in these enzymes, as previously reported (9, 31). In contrast, GMl-AL produced virtually no effect, both enzymes remaining at virtually the same level in the hippocampus as vehicle injected controls (Fig. 4). The methyl ester of GM1 produced an increment in both enzyme activities that was not statistically significant. Test substances were administe-
lpsilateral Hippocampus
CON VEH Ghi7 GMi-AL
LESION
Fig. 4. Comparison of the effect of GM, and its reduced alcohol (GMI-AL) on ChAT activity in rat hippocampus following t~nilateral partial lesion of the cholinergic septo-hippocampal pathway. Test substances were administered intramuscularly.
Similar information from the same study was previously presented in ref. 4.
red (30 mg/kg/day) by intramuscular injection for six days, beginning the first day after surgery.
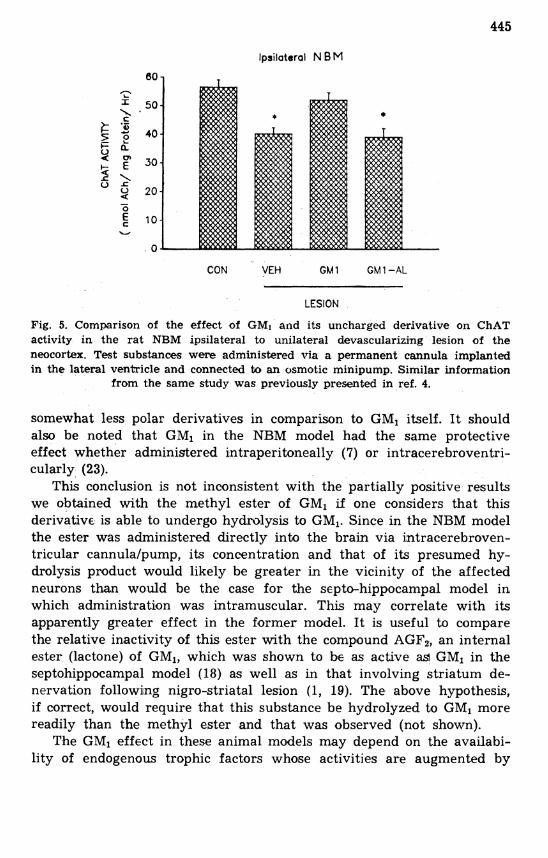
The second animal model employed adult male Wistar rats subjected to a unilateral left side cortical devascularizing lesion resulting in re- trograde degeneration of cholinergic neurons in the ipsilateral nucleus basalis magnocellularis (NBM) (7, 22). Test substances (5 mg/kg/day) in phosphate buffered saline were administered intracerebroventricularly for one week through a stainless steel cannula permanently implanted in the right lateral ventricle. Whereas GM1 caused retentian of ChAT activity in the NBM, as in previous studies (23), GM1-AL did not prevent the decrease in this enzyme seen with lesioned animals receiving vehicle (Fig. 5). The methyl ester of GM, was almost as effective as GMl itself in these trials (but see below).
The results with GMl-AL suggest the negative charge to be necessary for in vivo but not in vitro activity and argue against a common mecha- nism for the trophic effects of exogenous gangliosides as applied to cells in culture and the living animal. Further experimentation is needed to elucidate this proposition since the target cells in the in vitro and in vivo models differed as to neurochemical nature and stage of maturation. In addition, possible differential pharmacokinetic behavior should be con- sidered for the derivatives in the two animal models, although one might anticipate more facile penetration of the blood-brain barrier by the
lpsilatarol N B M
CON VEH GMl GMl-AL
LESION
Fig. 5. Comparison of the effect of GMI and its uncharged derivative on ChAT activity in the rat NBM ipsilateral to unilateral devascularizing lesion of the neocortex. Test substances were administered via a permanent cannula implanted in the lateral ventricle and connected to an osmotic minipump. Similar information
from the same study was previously presented in ref. 4.
somewhat less polar derivatives in comparison to GM, itself. I t should also be noted that GMl in the NBM model had the same protective effect whether administered intraperitoneally (7) or intracerebroventri- cularly (23).
This conclusion is not inconsistent with the partially positive results we obtained with the methyl ester of GM1 if one considers that this derivative is able to undergo hydrolysis to GM,. Since in the NBM model the ester was administered directly into the brain via intracerebroven- tricular cannulalpump, its concentration and that of its presumed hy- drolysis product would likely be greater in the vicinity of the affected neurons than would be the case for the septo-hippocampal model in which administration was intramuscular. This may correlate with its apparently greater effect in the former model. It is useful to compare the relative inactivity of this ester with the compound AGF,, an internal ester (lactone) of GM,, which was shown to be as active aa GMI in the septohippocampal model (18) as well as in that involving striatum de- nervation following nigro-striatal lesion (1, 19). The above hypothesis, if correct, would require that this substance be hydrolyzed to GM, more readily than the methyl ester and that was observed (not shown).
The GM1 effect in these animal models may depend on the availabi- lity of endogenous trophic factors whose activities are augmented by

GM1. I t has been noted that certain conditions are required for gan- gliosides to show a trophic effect, referred to as "permissive conditions" for in vivo models (23) or a "window of opportunity" in the c e of in vitro effects (30). Endogenous trophic factors which could provide the appropriate conditions were shown to be produced in brain immediately after injury (14, 17). The ability of GM1 to potentiate trophic factors has been well demonstrated with primary neurons in vitro (11). If potentia- tion of neurotrophic factors (13) is the operative mechanism, it remains to be determined why the GMl structure, effective both in vitro and in vivo, loses its modulatory activity in the latter model but not the former with removal of the negative charge.
CONCLUSIONS
In attempting to unravel the mystery of ganglioside mechanisms in the nervous system it has become useful to keep certain distinctions clearly in mind. First is that between endogenous and exogenous gan- gliosides, some evidence indicate that these do not function in the same way. This is the conclusion from in vitro studies which, in contrast to specific appearance of the gangliotetraose series during normal diffe- rentiation, showed a wide variety of natural and synthetic sialoglycoli- pids inducing neuritogenesis as exogenous agents. Possibly this nonspe- cific behavior originates in physical-chemical perturbation of the mem- brane triggering internal changes. In addition, in considering the phar- macological activity of exogenous gangliosides a distinction must be drawn between in vivo and in vitro behavior. This was seen most strikingly with an uncharged derivative of GMl which, although quite neuritogenic in three diverse cell culture systems, failed to exhibit trophic effects in two animal models for which GM1 was active. There is also an important distinction to be made between promotion of neurite outgrowth and facili- tation of survival involving protection against mechanical, toxicological and ischemic modes of damage. Finally, although the data at present are less concrete, it appears likely that differences exist between respanse to gangliosides of a mature neuron as opposed to one that is still in its developmental stage. Recognition of the diversity of phenomena now generally categorized as "ganglioside mechanism of action" may assist in clarifying the nature of specific aspects that can be fruitfully addres- sed in future studies.
Drs' L. Garofalo, M. Grqdkowska, M. Skup and M. Zaremba participation in experiments is acknowledged. This study was supported by USPHS grant NS 04834 (R.W.L.), a grant from the MRC (Canada) (A.C.C.), and project CPBP 04.01 of the Polish Academy of Sciences (B.0.-N.) M.S.C. received assistance from N.I.H. training grant NS 07098.

ABBREVIATIONS
GMl-AL, NaBH4 reduction product of GMl methyl ester, in which the carboxyl group of GM1 is converted to a primary alcohol. ChAT, choline acetyltransferase; AChE, acetylcholinesterase; NBM, nucleus ba- salis magnocellularis. Ganglioside nomenclature is that of Svenner- holm (25).
REFERENCES
1. ALDINIO, C., VALENTI, G., SAVOINI, G. E., KIRSCHNER, G., AGNATI, L.F. and TOFFANO, G. 1984. Monosialoganglioside internal ester stimulates the dopaminergic reinnervation of the striatum after unilateral hemitransection in rat. Int. Dev. Neurosci. 2: 267-275.
2. BYRNE, M. C., LEDEEN, R. W., ROISEN, F. J., YORKE, G. and SCLAFANI, J. R. 1983. Ganglioside-induced neuritogenesis: verification that ganglio- sides are the active agents, and comparison of molecular species. J. Neuro- chem. 41: 1214-1222.
3. CANNELLA, M. S., ACHER, A. J. and LEDEEN, R. W. 1988. Stimulation of neurite outgrowth in vitro by a glycero-ganglioside. Int. J. Dev. Neurosci. 6: 319-326.
4. CANNELLA, M. S., ODERFELDJNOWAK, B., GRADKOWSKA, M., SKUP, M., GAROFALO, L., CUELLO, A. C. and LEDEEN, R. W. 1990. Derivatives of ganglioside GM, as neuronotrophic agents: comparison of in vivo and in vitro effects. Brain Res. 513: 286-294.
5. CANNELLA, M. S., ROISEN, F. J., OGAWA, T., SUGIMOTO, M. and LEDEEN, R. W. 1988. Comparison of EPI-GM3 with GM3 and GMI a s stimulators of ceurite outgrowth. Dev. Brain Res. 39: 137-143.
6. CANNELLA, M. S., WU, G., VASWANI, K. K. and LEDEEN, R. W. 1988. Neu- ritogenic effects of exogenous gangliosides and synthetic sialoglycolipids: comparison to endogenous ganglioside requirements. In R. W. Ledeen, E. L. Hogan, G. Tettarnanti, A. J. Yates and R. K. Yu (ed.), New trends in ganglioside research: neurochemical and neuroregenerative aspects. Fidia Research Series. Vol. 14. Liviana Press, Padova, p. 379-390.
7. CUELLO, A. C., STEPHENS, P. H., TAGARI, P. C., SOFRONIEW, M. V. and PEARSON, R. C. A. 1986. Rotrograde changes in the nucleus basalis of the rat, caused by cortical damage, are prevented by exogenous ganglio- side GMl. Brain Res. 376: 373-377.
8. FERRARI, G., FABRIS, M. and GORIO, A. 1983. Gangliosides enhance neurite outgrowth in PC12 cells. Dev. Brain Res. 8: 215-222.
9. GRADKOWSKA, M., SKUP, M., KIEDROWSKI, L., CALZOLARI, S. and ODER- FELD-NOWAK, B. 1986. The effect of GMl ganglioside on cholinergic and serotonergic systems in the rat hippocampus following partial denervation is dependent on the degree of fiber degeneration. Brain Res. 375: 417-422.
10. LEDEEN, R. W. 1984. Biology of gangliosides: Neuritogenic and neuronotrophic properties. J. Neurosci. Res. 12: 147-159.
11. LEDEEN, R. W. 1989. Biosynthesis, metabolism, and biological effects of gan- gliosides. In R. U. Margolis and R. K. Margolis (ed.), Neurobiology of glycoconjugates. Plenum Publishing Corporation, New York, p. 43-83.
23 - Acta Neurobiol. Exp. 4-5,90

12. LEDEEN, R. W. and CANNELLA, M. S. 1987. The neuritogenic effect of gan- gliosides i n cell cultures. In H. R a h m a m (ed.), Gangliosides and modula- tion of neuronal functions. Springer-Verlag, Berlin, p. 491-500.
13. LEON, A., DAL TOSO, R., PRESTI, D., BENVENGNU, D., FACCI, L., KIR- SCHNER, G., TETTAMANTI, G. and TOFFANO, G. 1988. Development and survival of neurons in dissociated fetal mesencephalic serum-free cell cultures: 11. Modulatory effects of gangliasides. J . Neurosci. 8: 746-753.
14. MANTHORPE, M. A., NIETO-SAMPEDRO, M., SKAPER, S. D., LEWIS, E. G., BARBIN, G., LONGO, F. M., COTMAN, C. W. and VARON, S. 1983, Neuronotropic activity in brain wounds of the developing rat. Correlation with implant survival in the wound cavity. Brain Res. 267: 47-56.
15. MORGAN, J. I. and SEIFERT, W. 1979. Growth factors and gangliosides: A possible new perspective in neuronal growth control. J . Supramol. Struct. 10: 111-124.
16. NAGAI, Y. and TSUJI, S. 1988. Cell biological significance of gangliosides in neural differentiation and development: critique and proposals. In R. W. Ledeen, E. L. Hogan, G. Tettamanti, A. J. Yates and R. K. Yu (ed.), New trends in ganglioside research: neurochemical and neuroregenerative aspects. Liviana Press, Padova, p. 329-350.
17. NIETO-SAMPEDRO, M., MANTHORPE, M., BARBIN, G., VARON, S. and COTMAN, C. W. 1983. Injury-induced neuronotrophic activity in adult ra t brain: correlation with survival of delayed implants in the wound cavity. J. Neurosci. 3: 2219-2229.
18. ODERFELD-NOWAK, B., GRADKOWSKA, M., ZAREMBA, M. and KIEDROW- SKI, L. 1988. Protection against secondary degradation of nerve terminals h target structure - a possible mechanism to account for early beneficial effects of ganglioside treatment in injured brain. In R. W. Ledeen, E. L. Hogan, G. Tettamanti, A. J. Yates and R. K. Yu (ed.), New trends i n ganglioside research: neurochemical and neuroregenerative aspects. Fidia Research Series. Vol. 14. Liviana Press, Padova, p. 567-577.
19. RAITERI, M., VERSACE, P , and MARCHI, M. 1985. GM1 monosialoganglioside inner ester induces early recovery of striatal dopamine uptake in rats with unilateral nigrostriatal lesion. Eur. J. Pharmacol. 118: 347-350.
20. ROISEN, F. J., BARTFELD, H., NAGELE, R. and YORKE, G. 1981. Ganglio- side stimulation of axonal sprouting i n vitro. Science 214: 577-578.
21. SPOERRI, P. E. 1983. Effects of gangliosides on the i n vitro development of neuroblastoma cells: a n ultrastructural study. Int. J. Dev. Neurosci. 1: 383-391.
22. STEPHENS, P. H., CUELLO, A. C., SOFRONIEW, M. V., PEARSON, R. C. A. and TAGARI, P. 1985. Effect of unilateral decortication on choline acetyl- transferase activity in the nucleus basalis and other areas of the ra t brain. J. Neurochem. 45: 1021-1026.
23. STEPHENS, P. H., TAGARI, P. C., GARAFOLO, L., MAYSINGER, D., PIOT- TE, M. and CUELLO, A. C. 1987. Neural plasticity of basal forebrain cho- linergic neurons: effect of gangliosides. Neurosci Lett. 80: 80-84.
24. SUZUKI, K. 1965. The pattern of mammalian brain gangliosides. 111. Regional and developmental differences. J. Neurochem. 12: 969-979.
25. SVENNERHOLM, L. 1963. Chromatographic separation of human brain gan- gliosides. J. Neurochem. 10: 613-623.

26. SVENNERHOLM, L., BROSTROM, K., FREDMAN, P., MANSSON, J.-E., RO- SENGREN, B. and RYNMARK, B.-M. 1989. Human brain gangliosides: developmental changes from early fetal stage to advanced age. Biochim. Biophys. Acta 1055: 109-117.
27. TSUJI, S., ARITA, M. and NAGAI, Y. 1983. GQlb, a bioactive ganglioside that exhibits novel nerve growth factor (NGF)-like activities in the two neuroblastoma cell lines. J. aiochem. 94: 303-306.
28. TSUJI, S., YAMASHITA, T., TANAKA, M. and NAGAI, Y. 1988. Synthetic sialyl compounds as well as natural gangliosides induce neuritogenesis in a mouse neuroblastorna cell line (Neuro 2A). J. Neurochem. 50: 414-423.
29. UNSICKER, K. and WIEGANDT, H. 1988. Promotion of survival and neurite outgrowth of cultured peripheral neurons by exogenous lipids and de- tergents. Exp. Cell Res. 178: 377-389.
30. VARON, S., SKAPER, S. D. and KATOH-SEMBA, R. 1985. Neuritic responses to GM1 in several in vitro systems. Satelite symposium on neuronal pla- sticity and gangliosides. Int. Soc. Neurochem., Mantova, Abstract 13, p. 19.
31. WOJCIK, M., ULAS, J. and ODERFELD-NOWAK, B. 1982. The stimulating effect of ganglioside injections on the recovery of choline acetyltransferase and acetylcholinesterase activities in the hippcampus of the r a t after septa1 lesions. Neuroscience 7: 495-499.





























