Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Oct., 1966Copyright © 1966 American Society for Microbiology
Vol. 92, No. 4Printed in U.S.A.
Fractionation and Partial Characterization ofthe Products of Autolysis of Cell Walls
of Bacillus subtilisFRANK E. YOUNG'
Department of Pathology, Western Reserve University, Cleveland, Ohio, and Departments of Microbiology andExperimental Pathology, Scripps Clintic and Research Foundation, La Jolla, California
Received for publication 6 June 1966
ABsrRACTYOUNG, FRANK E. (Western Reserve University, Cleveland, Ohio). Fractionation
and partial characterization of the products of autolysis of cell walls of Bacillussubtilis. J. Bacteriol. 92:839-846. 1966.-Autolysis of the cell wall of Bacillus subtilisby an indigenous autolytic enzyme results in solubilization of 90% of the cell wall.The solubilized cell wall (supernatant fraction) was fractionated by the combina-tion of ion-exchange chromatography on diethylaminoethyl cellulose and gel filtra-tion on Sephadex G-25 into polysaccharides (composed of N-acyl glucosamine andN-acyl muramic acid), mucopeptides, peptides, and teichoic acid. The chemicalcomposition of the products of autolysis confirms the proposed mechanism of au-tolysis and establishes the autolytic enzyme as an N-acyl muramyl-L-alanine amidase.The heteropolymers in the cell wall are linked by peptide bridges. Two peptideswhich account for 70% of the peptides of the cell wall have a molar ratio of 1.0:0.9:1.3 for diaminopimelic acid, glutamic acid, and alanine, respectively. Other minorpeptides contain diaminopimelic acid, glutamic acid, and alanine in molar ratios of1.0:0.9:1.5, 1.0:0.5:1.0, and 1.0:1.5:1.7, respectively. The procedures employedin this study should be applicable to the fractionation of heteropolymers in cellwalls of other gram-positive organisms and thereby aid in the study of the struc-ture of antigenic determinants and endotoxins.
Lytic enzymes from animal tissue (13, 14, 17),bacteria (19), bacterial viruses (23), and fungi (6,20) have been utilized in the analysis of the struc-ture of the cell wall of microorganisms. Four as-pects of this approach require careful considera-tion. First, extreme care must be taken during thepreparation of cell walls to avoid partial degrada-tion by indigenous autolytic enzymes (16). Sec-ond, the enzymes which hydrolyze structuralmucopolysaccharides frequently have strict stere-ospecific requirements for substrate. For example,although a mucopeptide composed of N-acylglucosamine, N-acyl muramic acid, and aminoacids is common to all gram-positive .cell wallsstudied to date, there is a great variation in thesusceptibility of these cell walls to lysozyme (18).Furthermore, the copolymer of N-acyl muramicacid and N-acyl glucosamine, which theoretically
1 Faculty Research Associate, American CancerSociety. Present address: Department of Microbiology,Scripps Clinic and Research Foundation, La Jolla,Calif.
should be a good substrate for lysozyme, is notreadily hydrolyzed by lysozyme (27). A thirdproblem of this method is the degree of purity ofthe lytic enzyme. Not infrequently, extracts oforganisms and ifitrates of cultures contain a mix-ture of hexosaminidases and amidases (4). Incertain organisms, such as group D streptococci,the autolytic enzyme requires an initial activationby a proteolytic enzyme (2) which in itself mightalter the structure of the wall. Finally, the use oflytic enzymes to establish the structure of theheteropolymers and as reagents for the prepara-tion of antigens and endotoxin has been hamperedby the lack of procedures to isolate and purify theproducts of the hydrolytic reaction. Methods havebeen developed for the isolation of oligosac-charides, tetrasaccharides, disaccharides, andsome of the mucopeptides released during en-zymatic hydrolysis of cell walls, but these productsusually do not account for more than 25% of thetotal cell wall (13, 15, 28). Furthermore, Dowexion-exchange resins, which have been employed in
839
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
28
Dec
embe
r 20
21 b
y 24
.13.
92.1
89.

J. BACTERIOL.
some studies for fractionation of oligosaccharides,tetrasaccharides, and disaccharides (13), irre-versibly absorb some of the heteropolymers in thecell wall. Ghuysen, Tipper, and Strominger (7)recently isolated the products of hydrolysis of aglycopeptide-teichoic acid complex by the "32enzyme" of Streptomyces albus G; however, theirprocedure is restricted to small amounts of sub-strate and requires a combination of electro-phoresis and gel filtration.The cell walls of Bacillus subtilis 168 contain an
autolytic enzyme (N-acyl muramyl-L-alanineamidase) which rapidly solubilizes the cell wall byhydrolyzing the amide bond between the carbonylgroup of N-acyl muramic acid and L-alanine(24a, 27). The method of fractionation of theproducts of autolysis of the cell walls of B. subtilisdescribed in this communication provides a simpleprocedure for separating the heteropolymers ofthe cell wall. It is applicable to large quantities ofautolysate (100 to 300 mg), employs volatile buf-fers to preclude the problem of desalting, and isnot dependent on a specific spray or chemicalreaction for detection of the compounds. Themajor limitation of the method is the inability ofthese techniques to yield all of the fractions in achromatographically homogeneous form. Anal-ysis of the products verifies the proposed mech-anism of autolysis (24a, 27), demonstrates vari-ability in the subunits of the cell wall, reveals thepresence of three compounds which were notidentified in earlier studies of the chemical com-position of the cell wall of B. subtilis (26), andprovides an insight into the organization of thecell wall.
MATERIALS AND METHODS
Cell walls. Cell walls were prepared from B. subtilis168 I-C+ as described previously (26).
Preparation of the autolysate. A 300-mg sample ofcell walls was suspended in 0.02 M (NH4)2CO3. After60 min of incubation at 38 C, there was a 90 to 95%decrease in the optical density (measured at 600 m,u)of the suspension of cell walls. The suspension wasincubated for an additional 40 min at 38 C, and thenwas centrifuged in a Lourdes (model LRA) refrig-erated centrifuge (rotor 9 RA) at 12,000 X g for 20min at 4 C. The supernatant fraction (SF) was lyo-philized twice to ensure removal of the (NH4)2CO3 andwas then weighed. The precipitate was washed threetimes with cold distilled water, lyophilized, weighed,hydrolyzed in 4 N HCI at 105 C for 11 hr, and ana-lyzed on a Spinco (model 120B) automatic aminoacid analyzer (26). The precipitate, which is primarilyproteinaceous material (25), was not studied further.
Ion-exchange chromatography. Diethylaminoethyl(DEAE) cellulose (high capacity) type 20 with anexchange capacity of 1.05 meq/g (Schleichler &Schuell Co., Keene, N.H.) was suspended in 30 times
its weight of water, stirred vigorously, and allowed tosettle for several hours. The supernatant fluid con-taining the fine particles was decanted, and the wash-ing was continued until the supernatant liquid wasclear after 30 min of settling. The DEAE cellulose wasthen washed on a coarse sintered-glass funnel with asolution containing 0.125 N NaOH and 0.125 M NaCl(regenerator solution). The DEAE cellulose wasfinally suspended in the regenerator solution andpacked into chromatographic tubes at room tempera-ture with constant mechanical stirring in the reservoir.The column was then washed with a solution of 1%HCI in 0.5 M NaCl and finally was equilibrated withthe starting buffer. The column was transferred to acold room (5 C), equilibrated with 1 liter of startingbuffer, and stored until used. The DEAE cellulose wasremoved from the chromatographic tube after eachfractionation, rewashed in regenerator solution, andtransferred again into chromatographic tubes.
Chromatographic separation of the total SF of theautolysate was accomplished by step elution. Thesample [usually 200 to 400 mg suspended in 10 ml of0.02 M (NH4)2CO31 was added to a bed of DEAEcellulose measuring 35 to 40 by 2.5 cm in diameterwhich had previously equilibrated with 0.02 M(NH4)2CO3. The column was then eluted stepwise with0.02, 0.05, 0.075, 0.10, 0.15, 0.20, 0.25, 0.30, and 0.40M (NH4)2CO3 at 5 C with a flow rate of 30 ml/hr. Theeffluent was collected in 5-ml fractions. The opticaldensity of the effluent was measured at 220 mu in aZeiss PM Qll spectrophotometer. When the opticaldensity of the effluent from each step of elution de-creased below 0.08, the next higher concentration of(NH4)2CO3 was added.
Gelfiltration. Sephadex G-25 (Pharmacia, Uppsala,Sweden) was suspended in 50 times its weight ofdistilled water and allowed to settle slowly. The super-natant liquid containing fine particles was decanted,and the procedure was repeated until the supernatantliquid was clear after 30 min of settling. A slurry ofSephadex G-25 in distilled water (1: 3, v/v) was addedto chromatographic tubes with constant stirring.Extreme care was taken to keep the column verticalduring packing with Sephadex.
Each of the fractions obtained from chromatog-raphy of the autolysate on DEAE cellulose was lyo-philized twice, weighed, and suspended in 1.5 ml ofdistilled water. A 1.0-ml sample of each fraction wasadded to a 150- by 1-cm column containing a 146- by1-cm bed of Sephadex G-25. The column was elutedwith distilled water at room temperature. After theeffluent had passed through a Vanguard spectropho-tometer (model 1056A), 2.2-ml fractions were col-lected.
Chemical analyses. Samples of the autolysate andfractions obtained after column chromatography weredried over P20. for 24 hr, weighed, suspended in 4 NHCl, hydrolyzed at 105 C for 11 hr, and analyzed onthe Spinco (model 120B) automatic amino acid ana-lyzer as described previously (26). The autolysate andthe chromatographic fractions were analyzed forphosphorus (27) and dipicolinic acid (12). Glucosewas measured by the Glucostat assay (WorthingtonBiochemical Co., Freehold, N.J.).
840 YOUNG
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
28
Dec
embe
r 20
21 b
y 24
.13.
92.1
89.

AUTOLYSIS OF CELL WALLS OF B. SUBTILIS
RESULTSIon-exchange chromatography. The SF obtained
after autolysis of the cell wall could be resolvedinto 13 sulbfractions by chromatography onDEAE cellulose (Fig. 1). Because the cell wallscontain only traces of aromatic amino acids, thecompounds eluted from the column could not beidentified by their optical density at 280 mju. In-stead, the optical density at 220 mjs was utilized;however, the high blanks introduced by the vola-tile salt and variation in optical density due toslight contamination of the samples with dustpreclude calculation of recovery of the startingmaterial by this method. For example, in ninechromatographic separations, 90 to 150% of theoptical density of the starting material was re-covered. Thus, the sum of the weights of eachfraction, despite losses encountered in lyophiliza-tion, provides the only accurate estimation ofrecovery. As shown in Table 1, 178.4 mg of a 210-mg sample of the SF was recovered followingcolumn chromatography. Fractions 10 and 11contain most of the phosphorus in the SF (Fig. 1).
Gelfiltration. Each of the 13 fractions obtainedfrom chromatography of the supernatant fractionof the autolysate of cell walls was chromato-graphed on Sephadex G-25, as described inMaterials and Methods. Figure 2 shows that mostof the fractions obtained from DEAE cellulosecould be resolved into more subfractions, many
3.0
1.0
o 0.8
z 0.6
2 0.4-
0.21
7 810
200 400 600TUBE NUMBER
0.02 0.05 0.075 0.10 0.15 020 0.25 0.30 0.40BUFFER CONCENTRATION (MOLES/I)
FIG. 1. Chromatography of the supernatant fractionof autolysates of cell walls on DEAE cellulose. A210-mg sample ofan autolysate ofcell walls ofBacillussubtilis was added to a 40 X 2.5-cm column ofDEAEcellulose and eluted stepwise with (NH4)2CO3 at a flowrate of30 ml/hr. The optical density at 220 m,u (solidline) and total phosphorus content (dashed line) ofeachsample was determined as described in Materials andMethods. The volumes of effluent pooled for analysis(Tables I and 2) are designated by brackets and arabicnumerals.
TABLE 1. Recovery and chemical aiialysischromatographic fractions of autolysate of
cell walls of Bacillus subtilis
Frac-tionno.
SFla2a2c3c4a4b5aSb6a6b7a7b8a8b9a9b101112d13d
Re-covery"
mg
210.020.011.0
2.314.0
1.9
18.0
1.8
4.0
3.5
40.058.91.71.3
of
Molar ratiosb
Glu-tamicacid
1.2tr00.4
0.51.50.90.50.90.91.00.80.90.90.90.91.00.3
Ala- Glucos- Muramicnine amine acid
2.3 1.0 0.7tr 11.8 12.8tr6.6
1.01.71.31.01.31.31.61.21.61.51.51.51.50.4
17.70
7.803.500.0800.900.100.103.02.8
17.70
8.203.700.0801.000.100.102.81.5
Galac-tos-
amine
0.50.62.30
1.703.10000.40000012.12.3
a Recovery of each of the fractions from chrom-atography on DEAE cellulose (Fig. 1) not the sub-fractions (letters) from chromatography onSephadex G-25 (Fig. 2), e.g., the 20 mg listed nextto la is the recovery for the entire Fraction 1.
bMolar ratios of the compounds in fractions ob-tained from chromatography on Sephadex G-25(Fig. 2). Diaminopimelic acid (DAP) is present in all ofthe fractions except 3, 12, and 13. Because DAP isequated to 1.0 and all other calculations made withreference to DAP, the values for DAP (1.0) wereomitted from the table. Therefore, the complete molarratios for F6b for example are 1.0:0.9:1.3 for DAP,glutamic acid, and alanine, respectively.
c Only trace amounts of amino acids and aminosugars are present in this fraction.dThese fractions contain various amounts of
18 amino acids without a unique distribution.
of which were still heterogeneous. To ascertainthe chemical composition of these fractions, thepeak areas of each (Fig. 2) were lyophilized andanalyzed as described in Materials and Methods.The analyses demonstrated that the adsorptivecapacity of DEAE cellulose for the variousheteropolymers in the cell wall is: polysaccharide< mucopeptide < peptides < teichoic acid.
Polysaccharide. Two copolymers of N-acylmuramic acid and N-acyl glucosamine were ob-tained (Fraction la and Fraction 2a). These co-
VOL. 92, 1966 841
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
28
Dec
embe
r 20
21 b
y 24
.13.
92.1
89.
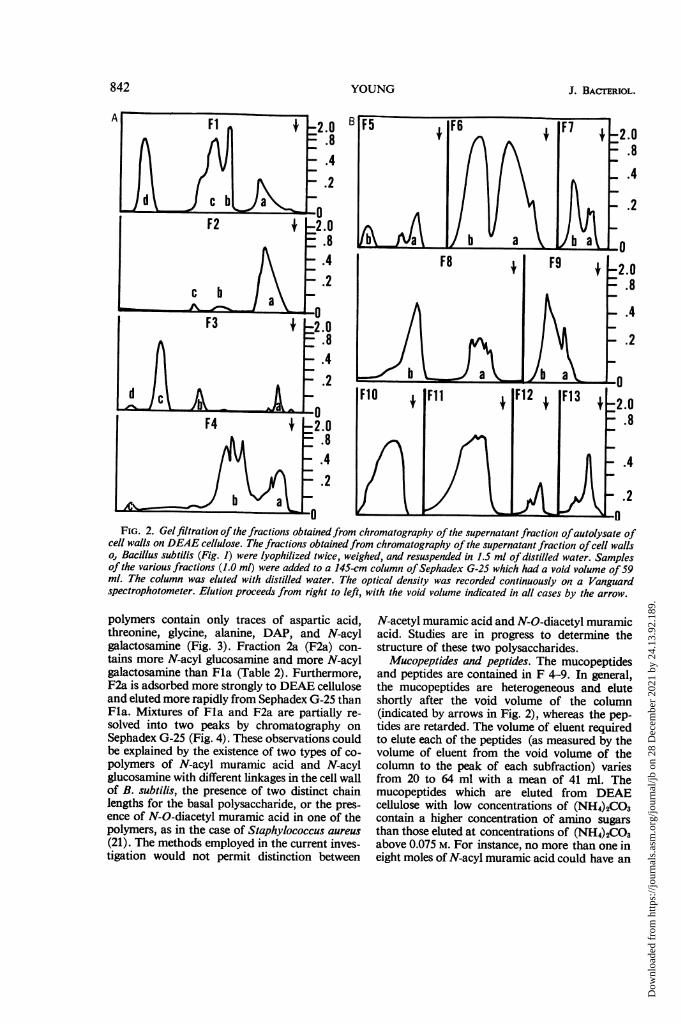
J. BACrERIOL.
FIG. 2. Gelfiltration of the fractions obtainedfrom chromatography of the supernatant fractionl ofautolysate ofcell walls on DEAE cellulose. The fractions obtainedfrom chromatography of the supernatant fraction ofcell wallso, Bacillus subtilis (Fig. 1) were lyophilized twice, weighed, and resuspended in 1.5 ml of distilled water. Samplesof the various fractions (1.0 ml) were added to a 145-cm column ofSephadex G-25 which had a void volume of59ml. The column was eluted with distilled water. The optical density was recorded continuously on a Vanguardspectrophotometer. Elution proceeds from right to left, with the void volume indicated in all cases by the arrow.
polymers contain only traces of aspartic acid,threonine, glycine, alanine, DAP, and N-acylgalactosamine (Fig. 3). Fraction 2a (F2a) con-tains more N-acyl glucosamine and more N-acylgalactosamine than Fla (Table 2). Furthermore,F2a is adsorbed more strongly to DEAE celluloseand eluted more rapidly from Sephadex G-25 thanFla. Mixtures of Fla and F2a are partially re-solved into two peaks by chromatography onSephadex G-25 (Fig. 4). These observations couldbe explained by the existence of two types of co-polymers of N-acyl muramic acid and N-acylglucosamine with different linkages in the cell wallof B. subtilis, the presence of two distinct chainlengths for the basal polysaccharide, or the pres-ence of N-O-diacetyl muramic acid in one of thepolymers, as in the case of Staphylococcus aureus(21). The methods employed in the current inves-tigation would not permit distinction between
N-acetyl muramic acid and N-O-diacetyl muramicacid. Studies are in progress to determine thestructure of these two polysaccharides.
Mucopeptides and peptides. The mucopeptidesand peptides are contained in F 4-9. In general,the mucopeptides are heterogeneous and eluteshortly after the void volume of the column(indicated by arrows in Fig. 2), whereas the pep-tides are retarded. The volume of eluent requiredto elute each of the peptides (as measured by thevolume of eluent from the void volume of thecolumn to the peak of each subfraction) variesfrom 20 to 64 ml with a mean of 41 ml. Themucopeptides which are eluted from DEAEcellulose with low concentrations of (NH4)2CO3contain a higher concentration of amino sugarsthan those eluted at concentrations of (NH4)2CO3above 0.075 M. For instance, no more than one ineight moles of N-acyl muramic acid could have an
842 YOUNG
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
28
Dec
embe
r 20
21 b
y 24
.13.
92.1
89.
AUTOLYSIS OF CELL WALLS OF B. SUBTILIS
.5.
.4'
1 2 34536 7 8K.0.
1 23 45 61i 8 9
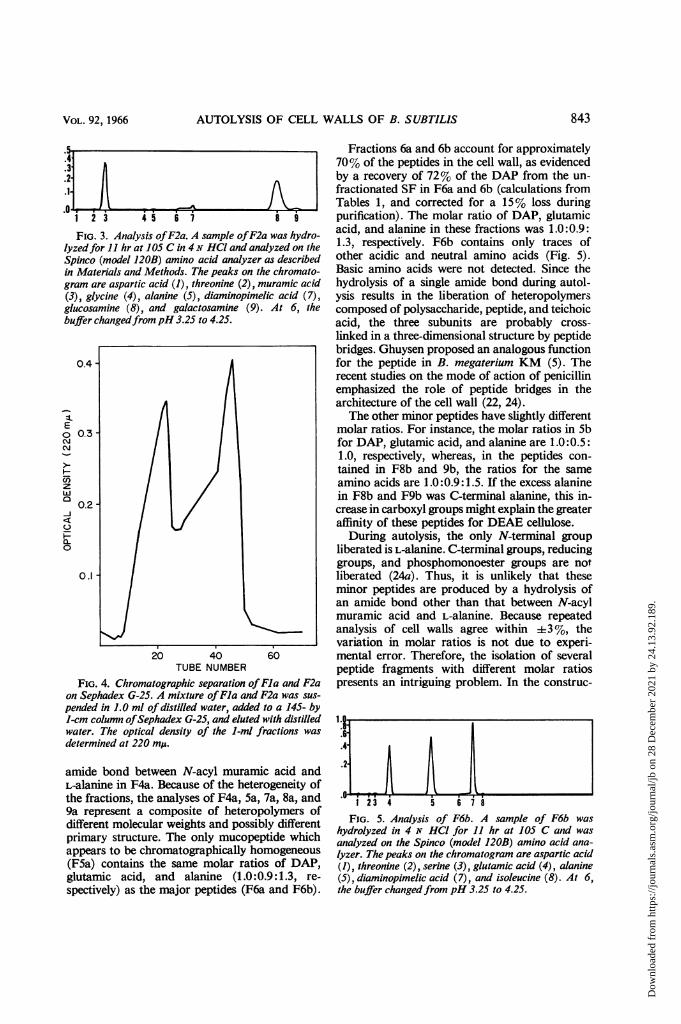
FIG. 3. Analysis ofF2a. A sample ofF2a was hydro-lyzedfor 11 hr at 105 C in 4N HCl and analyzed on theSpinco (model 120B) amino acid analyzer as describedin Materials and Methods. The peaks on the chromato-gram are aspartic acid (1), threonine (2), muramic acid(3), glycine (4), alanine (5), diaminopimelic acid (7),glucosamine (8), and galactosamine (9). At 6, thebuffer changedfrompH 3.25 to 4.25.
0.4
E
o 0.3
z
w
0.2-
Q-0
0.1
20 40 60TUBE NUMBER
FIG. 4. Chromatographic separation ofFla and F2aon Sephadex G-25. A mixture ofFla and F2a was sus-
pended in 1.0 ml of distilled water, added to a 145- by1-cm columtn ofSephadex G-25, and eluted with distilledwater. The optical density of the 1-ml fractions wasdetermined at 220 mp.
amide bond between N-acyl muramic acid andL-alanine in F4a. Because of the heterogeneity ofthe fractions, the analyses of F4a, 5a, 7a, 8a, and9a represent a composite of heteropolymers ofdifferent molecular weights and possibly differentprimary structure. The only mucopeptide whichappears to be chromatographically homogeneous(F5a) contains the same molar ratios of DAP,glutamic acid, and alanine (1.0:0.9:1.3, re-
spectively) as the major peptides (F6a and F6b).
Fractions 6a and 6b account for approximately70% of the peptides in the cell wall, as evidencedby a recovery of 72% of the DAP from the un-fractionated SF in F6a and 6b (calculations fromTables 1, and corrected for a 15% loss duringpurification). The molar ratio of DAP, glutamicacid, and alanine in these fractions was 1.0:0.9:1.3, respectively. F6b contains only traces ofother acidic and neutral amino acids (Fig. 5).Basic amino acids were not detected. Since thehydrolysis of a single amide bond during autol-ysis results in the liberation of heteropolymerscomposed of polysaccharide, peptide, and teichoicacid, the three subunits are probably cross-
linked in a three-dimensional structure by peptidebridges. Ghuysen proposed an analogous functionfor the peptide in B. megaterium KM (5). Therecent studies on the mode of action of penicillinemphasized the role of peptide bridges in thearchitecture of the cell wall (22, 24).The other minor peptides have slightly different
molar ratios. For instance, the molar ratios in Sbfor DAP, glutamic acid, and alanine are 1.0:0.5:1.0, respectively, whereas, in the peptides con-
tained in F8b and 9b, the ratios for the sameamino acids are 1.0:0.9:1.5. If the excess alaninein F8b and F9b was C-terminal alanine, this in-crease in carboxyl groups might explain the greateraffinity of these peptides for DEAE cellulose.During autolysis, the only N-terminal group
liberated is L-alanine. C-terminal groups, reducinggroups, and phosphomonoester groups are notliberated (24a). Thus, it is unlikely that theseminor peptides are produced by a hydrolysis ofan amide bond other than that between N-acylmuramic acid and L-alanine. Because repeatedanalysis of cell walls agree within +3%, thevariation in molar ratios is not due to experi-mental error. Therefore, the isolation of severalpeptide fragments with different molar ratiospresents an intriguing problem. In the construc-
II..65.4-
1 23 4 5 687FIG. 5. Analysis of F6b. A sample of F6b was
hydrolyzed in 4 N HCI for 11 hr at 105 C and wasanalyzed on the Spinco (model 120B) amino acid ana-lyzer. The peaks on the chromatogram are aspartic acid(1), threonine (2), serine (3), glutamic acid (4), alanine(5), diaminopimelic acid (7), and isoleucine (8). At 6,the buffer changedfrom pH 3.25 to 4.25.
843VOL. 92, 1966
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
28
Dec
embe
r 20
21 b
y 24
.13.
92.1
89.

J. BACTERIOL.
tion of the cell wall, there must be a mechanismfor limiting the length of the cross-linked peptidein the basal mucopeptide of the cell wall. Thedeletion of one of the diamino or dicarboxylicamino acids in the mucopeptide would of neces-sity prevent further cross-links. Peptides with a
preponderance of glutamic acid or DAP wouldproduce such an effect. Alternatively, the differ-ences in the ratio of the various amino acids inthe peptides may be due to geographic differencesin synthesis of cell wall within each organism or
may reflect the differences in differentiation in thenonsynchronous cultures during presporulationevents. It is also possible that the minor peptidesmay represent unique cross-links between differ-ent heteropolymers. Wise and Park (24) andTipper and Strominger (22) presented evidencethat penicillin inhibits transpeptidization inS. aureus. If this finding can be extended to B. sub-tilis, the peptides in the cell wall of organismsinhibited with penicillin should contain muco-peptides with different ratios of amino acids thanin the cross-linked peptides.
Teichoic acid. The cell walls of B. subtilis con-tain teichoic acid, a polymer of polyribitol phos-phate (1) or polyglycerol phosphate (3). Thepolyol may be substituted with amino acids suchas D-alanine (1) and carbohydrate such as glucose(1). In B. subtilis 168, hydrolysis of the amidebond between N-acyl muramic acid and L-alanineduring autolysis (24a), results in the liberation oftwo heteropolymers (FIO and Fll) which arecomposed of a polyol (because acid hydrolysatesof these fractions serve as a substrate for glyceroldehydrogenase, but not for ribitol dehydrogen-ase, the polyol in B. subtilis 168 is probablyglycerol; however, more sensitive and quantita-tive assays are required to establish this), glucose,phosphorus, and different amounts of aminosugars. The molar ratios of N-acyl muramic acid,N-acyl glucosamine, N-acyl galactosamine, glu-
cose, and phosphorus are 1.0:1.0:4.1:11:10 forF10, whereas the molar ratios of these com-
pounds in Fit are 1.0:1.8:1.7:22:20 [molarratios derived from analyses in Table 2 and glu-cose determinations (Young, unpublished data);muramic acid = 1.0]. Fractions 10 and 11,which constitute 55% of the mass of the cell wall,are occasionally eluted together in the 0.25 M step.The amino sugars in the teichoic acid fractionsprobably serve an important function in thelinkage of teichoic acid to the basal mucopeptide.For example, a peptide bridge between N-acylmuramic acid in teichoic acid and the N-acylmuramic acid in the basal mucopeptide couldestablish a covalent linkage between these twoheteropolymers. Hydrolysis of the amide bondsbetween N-acyl muramic acid and L-alanine in thecell wall during autolysis would result in the lib-eration of the major polymers isolated in thisstudy: (i) copolymers of N-acyl glucosamine andN-acyl muramic acid from the basal mucopeptide,(ii) cross-linked peptides, and (iii) teichoic acidcontaining amino sugars.
Teichuronic acid, a copolymer of N-acetylgalactosamine and glucuronic acid, was originallydescribed by Janczura et al. (11) in an organismdesignated B. subtilis NCTC 6346. This organismhas been reclassified as B. licheniformis NCTC6346 (10). Three observations demonstrate thatthe N-acyl galactosamine in FIO and Fll in B.subtilis 168 is covalently attached to the polyoland is not present in teichuronic acid. First,glucuronic acid is not present in cell walls of B.subtilis 168 (27), FIO, or Fll. Second, the aminosugars precipitate to the same extent as the polyolphosphate with 1-hexadecylpyridinium chloride(Young, unpublished data). Third, there is lessthan 1 phosphomonoester group per 500 residuesof organic phosphorus in the autolysate, whichdemonstrates that most of the organic phosphorusin the cell wall is covalently linked to another
TABLE 2. Chemical composition offractions of cell walls of Bacillus subtilis 168 I-C+a
Sample Alanine DAP Glutamic Glucosamine Muramic Galactosamine Phosphorusacid acid
Cell walls........ 0.68 0.30 0.35 0.30 0.22 0.15 1.28SFb... 0.64 0.30 0.34 0.30 0.27 0.17 1.27Fla............<0.03 0.11 <0.03 1.30 1.41 0.07 <0.05F2a............ 0.03 0.08 <0.03 1.42 1.42 0.18 <0.05F6a.. . . .. . . . .... 2.05 1.53 1.41 0.12 0.12 <0.01 <0.05F6b............ 2.65 2.10 1.73 0 0 0 <0.05F10............ 0.09 0.06 0.06 0.18 0.17 0.73 1.79Fil ............ <0.03 0.05 <0.03 0.16 0.09 0.15 1.80
a Expressed as micromoles per milligram.b This supernatant fraction, obtained by centrifugation of the autolysate at 12,000 X g for 20 min,
was the starting material employed in column chromatography.
844 YOUNG
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
28
Dec
embe
r 20
21 b
y 24
.13.
92.1
89.

AUTOLYSIS OF CELL WALLS OF B. SUBTILIS
compound (27). Treatment of the autolysate orpartially purified teichoic acid with trichloroaceticacid rapidly hydrolyzes the amino sugar link (27).Although many investigators have suggestedthat the amino sugars are linked to the teichoicacid through phosphodiester bonds (8; Tipper,Strominger, and Ghuysen, Federation Proc. 23:379, 1964) or by a phosphoramide bond (9),direct quantitative evidence for such a linkage isnot available. The presence of two subtypes ofteichoic acid in B. subtilis which can be obtainedin large quantities by column chromatographymay be helpful in elucidating the linkage betweenteichoic acid and polysaccharide.
Other compounds present within the autolysatesofcell walls. Autolysis at alkalinepH results in theliberation of alkaline labile amino acids fromteichoic acid which comprise Flb. Alanine con-stitutes 95% of the alkaline labile amino acids(Table 3).Three fractions have not been successfully
identified: Flc, Fld, and F3c. These fractions donot contain reducing groups, phosphorus, dipi-colinic acid, ribose, or amino acids. Their ultra-violet absorption spectrum is shown in Fig. 6.
Fractions 12 and 13 are proteins. These frac-
2.612.0
1.6
1.4-
i1.2
01.00
200 230 260 290 320
Wavelength (mei)
FIG. 6. Ultraviolet absorption spectrum of the un-known compounds. The ultraviolet absorption spectrumof Flc (solid line), Fid (dotted line), and F3c (dashedline) dissolved in distilled water was determined with aZeiss spectrophotometer.
TABLE 3. Molar ratios ofamino acids in Fractioni lb
Component Ratioa
Alanine ................. 158.0Aspartic acid ....................... <0.5Threonine. ..1 .2Serine ............................. 1.0Glycine ............................. 1.0Valine. ..1.7Isoleucine ............................. 1.4Leucine ............................ 1.6
a Serine = 1.0.
tions do not contain active autolytic enzyme.Together they constitute less than 1.5% of the SF.
ACKNOWLEDGMENTSThis investigation was supported by grant E 288 b
and grant PR 8 from the American Cancer Society,and by Public Health Service grant GM 12956-01from the Division of General Medical Sciences.The technical assistance of Andrew Jackson is
gratefully acknowledged.
LITERATURE CITED1. ARMSTRONG, J. J., J. BADDILEY, AND J. G. Bu-
CHANAN. 1961. Further studies on the teichoicacid from Bacillus subtilis walls. Biochem. J.81:254-261.
2. BLEISEIS, A. S., AND R. M. KRAUSE. 1965. The cellwalls of group D streptococci. I. The immuno-chemistry of type I carbohydrate. J. Exptl. Med.122:237-249.
3. BURGER, M. M., AND L. GLASER. 1964. The syn-thesis of teichoic acids. I. Polyglycerophosphate.J. Biol. Chem. 239:3168-3177.
4. GHUYSEN, J-M. 1960. Acetylhexosamine com-pounds enzymatically released from Micro-coccus lysodeikticus cell walls. Biochim. Bio-phys. Acta 40:473-480.
5. GHUYSEN, J-M., M. LEYH-BOvILLE, AND L.DIERICKX. 1962. Structure des parois de BacillusMegaterum. K. M. II. Etude des complexesmucopeptidique et phospho - mucopolysac-charidique. Biochim. Biophys. Acta 63:297-307.
6. GHUYSEN, J-M., AND J. L. STROMINGER. 1963.Structure of the cell wall of Staphylococcusaureus, strain Copenhagen. I. Preparation offragments by enzymatic hydrolysis. Biochemis-try 2:1110-1119.
7. GHUYSEN, J-M., D. J. TIPPER, AND J. L. STROMIN-GER. 1965. Structure of the cell wall of Staphylo-coccus aureus, strain Copenhagen. IV. Theteichoic acid-glycopeptide complex. Biochem-istry 4:474-485.
8. HALL, E. A., AND K. W. KNOX. 1965. Properties ofthe polysaccharide and mucopeptide compo-nents of the cell wall of Lactobacillus casei.Biochem. J. 96:310-318.
9. HAY, J. B., A. R. ARCHIBALD, AND J. BADDILEY.1965. The molecular structure of bacterial walls.The size of ribitol teichoic acids and the nature
VOL. 92, 1966 845
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
28
Dec
embe
r 20
21 b
y 24
.13.
92.1
89.

J. BACTERIOL.
of their linkage to glycosaminopeptides. Bio-chem. J. 97:723-730.
10. HUGUES, R. C. 1965. The isolation of structuralcomponents present in the cell wall of Bacilluslicheniformis NCTC 6346. Biochem. J. 96:700-709.
11. JANCZURA, E., H. R. PERKINS, AND H. J. ROGERS.1961. Teichuronic acid: a mucopolysaccharidepresent in wall preparations from vegetativecells of Bacillus subtilis. Biochem. J. 80:82-93.
12. JANSSEN, F. W., A. J. LuND, AND L. E. ANDERSON.1958. Colorimetric assay for dipicolinic acid inbacterial spores. Science 127:26-27.
13. JEANLOZ, R. W., N. SHARON, AND H. M. FLOWERS.1964. Synthetic muramic acid derivatives andthe chemical structure of Micrococcus lyso-deikticus cell walls, p. 49-62. In M. Landy andW. Braun [ed.], Bacterial endotoxins. RutgersUniv. Press, New Brunswick, N.J.
14. SALTON, M. R. J. 1952. Cell wall of Micrococcuslysodeikticus as the substrate for lysozyme.Nature 170:746-747.
15. SALTON, M. R. J. 1956. Studies of the bacterialcell wall. V. The action of lysozyme on cellwalls of some lysozyme-sensitive-bacteria. Bio-chim. Biophys. Acta 22:495-506.
16. SALTON, M. R. J. 1960. Surface layers of the bac-terial cell, p. 97-151. In I. C. Gunsalus andR. Y. Stanier [ed.], The bacteria, vol. 1. Aca-demic Press, Inc., New York.
17. SALTON, M. R. J., AND J-M. GHUYSEN. 1960. Ace-tylhexosamine compounds enzymatically re-leased from Micrococcus lysodeikticus cell walls.III. The structure of di- and tetrasaccharidesreleased from cell walls by lysozyme andStreptomyces F. enzyme. Biochim. Biophys.Acta 45:355-363.
18. SALTON, M. R. J., AND J. M. PAVLIK. 1960. Studiesof the bacterial cell wall. VI. Wall compositionand sensitivity to lysozyme. Biochim. Biophys.Acta 39:398-407.
19. STRANGE, R. E. 1959. Cell wall lysis and the re-lease of peptides in Bacillus species. Bacteriol.Rev. 23:1-7.
20. TIPPER, D. J., J-M. GHUYSEN, AND J. L. STROMIN-GER. 1965. Structure of the cell wall of Staphy-loccus aureus, strain Copenhagen. III. Furtherstudies of disaccharides. Biochemistry 4:468-473.
21. TIPPER, D. J., J. L. STROMINGER, AND J-M. GHUY-SEN. 1964. Staphylytic enzyme from Chalaropsis:Mechanism of action. Science 146:781-782.
22. TIPPER, D. J., AND J. L. STROMINGER. 1965.Mechanism of action of penicillins: Proposalbased on their structural similarity to acyl-D-alanyl-D alanine. Proc. Natl. Acad. Sci. U.S.54:1133-1141.
23. WEIDEL, W., AND J. PRIMOSIGH. 1958. Biochemicalparallels between lysis by virulent phage andlysis by penicillin. J. Gen. Microbiol. 18:513-517.
24. WISE, E. M., JR., AND J. T. PARK. 1965. Penicillin:Its basic site of action as an inhibitor of a pep-tide cross-linking reaction in cell wall muco-peptide synthesis. Proc. NatI. Acad. Sci. U.S.54:75-81.
24a. YOUNG, F. E. 1966. Autolytic enzyme associatedwith cell walls of Bacillus subtilis. J. Biol. Chem.241:3462-3467.
25. YOUNG, F. E., AND J. SPIZIZEN. 1963. Biochemicalaspects of competence in the Bacillus subtilistransformation system. II. Autolytic enzymeactivity of cell walls. J. Biol. Chem. 238:3126-3130.
26. YOUNG, F. E., J. SPIZIZEN, AND I. P. CRAWFORD.1963. Biochemical aspects of competence in theBacillus subtilis transformation system. I. Chem-ical composition of cell walls. J. Biol. Chem.238:3119-3125.
27. YOUNG, F. E., D. J. TIPPER, AND J. L. STROMIN-GER. 1964. Autolysis of cell walls of Bacillussubtilis: Mechanism and possible relationshipto competence. J. Biol. Chem. 239:PC3600-3602.
28. ZILLIKEN, F. 1959. Chemistry of bacterial cellwalls. Federation Proc. 18:966-973.
846 YOUNG
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
28
Dec
embe
r 20
21 b
y 24
.13.
92.1
89.





























