Embed Size (px)
Citation preview

FoxP2 - A gene under selectionSusan Ptak, Wolfgang Enard, Johannes Krause, Victor Wiebe,
Ines Hellmann, Michael Lachmann and Svante Paabo

Hurst et al. (1990) Dev Med Child Neurol 32, 352-355
male female
affected non-affected
A family with normal intelligence, buta speech-disorder:
http://www.youtube.com/watch?v=Fg2rLOkoL9Q

FOX
1 2 3 4 5 6 7 8 9 10 111213 141516 17
atg tga
Chromosomal breakpoint, patient CS R553H mutation in KE family
Lai et al (2001) Nature 413, 519–523
PolyQ
STOP mutation in unrelated patient
Mapping of SPCH1
- linkage map points to 7q31 to a 3.8 cM region (Fisher et al. 1998, Nat. Genet.) - cloning of the chromosomal breakpoint of an unrelated individual:

2 amino acid on the human lineage:
sequenced in a sample of 164 chromosomes, only one has a morenegative value (22.25). A second prediction for a selective sweep ata recombining locus is that more derived (that is, non-ancestral)alleles at high frequency are expected than under the standardneutral model, a feature reflected in a negativeH value17. To estimateH, we inferred the ancestral states of variable positions seen amongthe humans by using the chimpanzee and orang-utan DNAsequences. The H value of 212.24 deviates significantly from theneutral expectation of zero (P ! 0.042) andwould be even less likelyby chance under a model with population growth13. The stronglynegativeD andH reflect an extreme skew in the frequency spectrumof allelic variants at FOXP2 towards rare and high-frequency alleles.Because we considered a worldwide sample of humans, populationstructure might contribute to the negative D value. However, thistype of sampling scheme is highly unlikely to produce a significantlynegative H value. In contrast to demographic explanations, aselective sweep affecting the FOXP2 gene can account for bothaspects of the frequency spectrum. We do not observe a reduceddiversity at human FOXP2 relative to its divergence from thechimpanzee, as expected under a simple selective-sweep model.However, the magnitude of the reduction in variability expectedafter a selective sweep depends crucially on the rate of recombina-tion. Estimates of recombination between intronic polymorphismstaken from a study of FOXP2 (ref. 4) suggest that this region of thegene experiences rates of genetic exchange roughly five times thegenome-wide average. If we assume that a selective sweep at a linkedsite does account for the patterns of variability recovered at FOXP2,it is noteworthy that the next gene is located 286 kilobases (kb) awayfrom the sequenced segment. A selective sweep is not expected tolead to an excess of high-frequency derived alleles at sites that are286 kb distant from the target of selection13,17. Thus, the bestcandidates for the selected sites are the two amino-acid substi-tutions specific to humans in exon 7.
Individuals with disruption of FOXP2 have multiple difficultieswith both expressive and receptive aspects of language and gram-mar, and the nature of the core deficit remains a matter of debate18–20. Nevertheless, a predominant feature of the phenotype of affectedindividuals is an impairment of selection and sequencing of fineorofacial movements18, an ability that is typical of humans and notpresent in the great apes. We speculate that some human-specificfeature of FOXP2, perhaps one or both of the amino-acid substi-tutions in exon 7, affect a person’s ability to control orofacialmovements and thus to develop proficient spoken language. Ifthis speculation is true, then the time when such a FOXP2 variantbecame fixed in the human population may be pertinent withregard to the evolution of human language. We estimated thistime point using a likelihood approach. Under a model of arandomly mating population of constant size, the most likely datesince the fixation of the beneficial allele is 0, with approximate 95%confidence intervals of 0 and 120,000 years. Our point-estimate of 0
reflects the fact that high-frequency alleles rapidly drift to fixation,so an excess is most likely immediately after a selective sweep.However, if population growth soon succeeds the fixation of theadvantageous allele, the rate of drift will be decreased and high-frequency alleles may persist longer in the population. Thus, theinclusion of population growthmay push this time estimate back byat most the time since the onset of human population growth, some10,000–100,000 years ago21. In any case, our method suggests thatthe fixation occurred during the last 200,000 years of human history,that is, concomitant with or subsequent to the emergence ofanatomically modern humans22. This is compatible with a modelin which the expansion of modern humans was driven by theappearance of a more-proficient spoken language22. However, toestablish whether FOXP2 is indeed involved in basic aspects ofhuman culture, the normal functions of both the human and thechimpanzee FOXP2 proteins need to be clarified. A
MethodsIsolation of cDNA sequencesFor all analysed species, we amplified by polymerase chain reaction (PCR) and sequencedoverlapping fragments of the FOXP2 coding region from first-strand cDNA. Details areavailable in Supplementary Information.
Genomic sequencingFull details are available in Supplementary Information. In brief, we designed primersfrom a human bacterial artificial chromosome (BAC) sequence (accession numberAC020606), PCR-amplified fragments of 6–14 kb, re-amplified 2.2-kb fragments fromthese products that were then sequenced with internal primers. For each individual, eachnucleotide position was read from both strands. Sequence traces were manually analysedfor polymorphic positions using the program Seqman of the DNAStar package (see alsoSupplementary Information).
Data analysisWe aligned sequences with the help of the program ClustalW23 and calculated moststatistics with DnaSP 3.51 (ref. 24). P values for D and H were obtained by coalescentsimulations implemented for a fixed number of segregating sites, and assuming norecombination. If we take into account recombination within the 14 kb, the P valuesdecrease (for example, P , 0.01 for H and P , 1024 for D if one assumes an effectivepopulation size of 104 and a recombination rate of 5 centimorgans (cM) per Mb). Becausethe chimpanzee and orang-utan do not differ at any polymorphic position compared withhumans, we assumed no backmutations when estimating the P value forH. The likelihoodratio tests for non-silent and silent substitutions were performed using the PAMLpackage12 as described25 (see Supplementary Information). We predicted the structure ofhuman, chimpanzee, mouse and orang-utan FOXP2 using the program PredictProtein(http://www.embl-heidelberg.de/predictprotein/predictprotein.html)6, which includesprediction of sites of protein kinase C phosphorylation by PROSITE26. The orang-utan-specific alanine-to-valine change at position 6 results in the prediction of a b-sheet atpositions 8–10 in the orang-utan, and the human-specific change at position 325 results inthe prediction of a b-sheet in positions 323–326. However, these are not reliable and maynot be relevant. We used the University of California at Santa Cruz Human GenomeProject Working Draft, 22 December 2001 assembly (http://genome.cse.ucsc.edu), toestimate distances to the closest genes. The middle of the sequenced region is 220 kb awayfrom the known 59 end and 54 kb away from the 39 end of FOXP2, respectively. The nextgene (supported by the cDNA sequence with GenBank accession number AF054589) islocated 286 kb distant in the 39 direction.
Modelling the selective sweepA summary likelihood method (compare with ref. 27) was used to estimate the time, T,since the fixation of the beneficial allele in the population. The polymorphism data wassummarized as vH (ref. 17) and p (ref. 28). We then ran coalescent simulations of aselective sweep with recombination as in ref. 13. These simulations assume that we havepolymorphism data for a neutral locus, at some distance from a selected site, and thatselection acted on a newly arising variant. The likelihood of T is estimated as theproportion of n simulated data sets, where jvHobs 2 vHsimj, 1 and jpobs 2psimj, 1(here, n ! 3 £ 106 and 1 ! 0.2). The likelihood of Twas evaluated over a grid of pointsspaced every 1,000 generations. We then chose the T value that maximizes the probabilityof obtaining the observed (vH, p) values. In addition to T, several additional parametersare in this selective sweep model: the distance to the selected site, the effective populationsize of humans, the strength of selection, the mutation rate and the recombination rate. Itis not computationally feasible to co-estimate all of these parameters, and we proceeded byassuming that the values of most nuisance parameters are known exactly.We hypothesizedthat one of the substitutions on the human lineage was the selected site and used a pointestimate of the population mutation rate (assuming 5 Myr to the common ancestor of ahuman and chimpanzee DNA sequence). We modelled uncertainty in the recombinationrate per megabase by choosing the rate for each simulation from a g distribution withparameters (5, 1); the mean was set to the recombination rate estimated from twopolymorphic markers in introns 2 and 16, respectively, of the FOXP2 gene4. The effective
0/7
1/2
0/5
1/131
0/5
0/2
0/2
2/0***
0/2
Human
C himp
G orilla
Orang-utan
Rhesus
Mouse
Figure 2 Silent and replacement nucleotide substitutions mapped on a phylogeny ofprimates. Bars represent nucleotide changes. Grey bars indicate amino-acid changes.
letters to nature
NATURE |VOL 418 | 22 AUGUST 2002 | www.nature.com/nature 871© 2002 Nature Publishing Group
FOX
1 2 3 4 5 6 7 8 9 10 111213 141516 17
atg tga
PolyQ
T->N N->S
Enard et al., Nature, 2002
Note: the amino acid changes are fixed --not polymorphic in ~1000 humans of the CEPH diversity panel

ANCESTRAL STATE A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A C C T G C A A G A A T G AOrang-Utan A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A C C T G C A A G A A T G AChimpanzee A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A Y C T G C A A G A A T G A
Human Consensus A A A T A G A A A C A C C A G A G A G A G A C G T C C G A T A C G T G C T A T A A T G A1 Australian Abor. "HAY-KJ" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - G - - - - - - - - - - - - -2 Warao South Amer Ind "SPACH"- - G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -3 Chinese "XLI-ND" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -4 Japanese "KT12" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -5 Thai "DC009" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- T - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -6 PNG (coastal) "100" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - -7 PNG (highlander) "35" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - -8 Nasioi (Melanesia) "966" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -9 Iranian "KHAGNI" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - T - - - - - - - - G - - -10 French "DUCAF" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - - - - - - - - - - G - - -12 German "EHM" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - - - - - - - - - - G - - -13 English "IHAF" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - G - - - - - - - - - - G - - - - - - - - - - - - - - - - - - - - - - - - - - - -14 Italian "WT24" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -15 Mbuti Pygmy "1039" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - T - - - - A - - - C - - - - - - - - - - - - - - - T T - C - - - C - -16 Biaka Pygmy "785" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -17 Biaka Pygmy "805" - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - G - - - A - A G C - - - - - - T A - - A - - - - T T - - - - - C - -18 Nigerian (Ibo) "044" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
C - - G - A G T - - - T T G A - - - - T A - - - - - - - C A - - C C - - A - G T - - - G19 Nigerian (Yoruba) "025" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - G - - - - - A - - - - - - G - - - - - - - - - - - - - T - - - - - - - -20 Nigerian (effik) "002" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - G - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -21 Nigerian (Hausa) "006" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - C - - A - - - - - - - - - - - - - - A -
FOX
1 2 3 4 5 6 7 8 9 10 111213 141516 17
atg tga
PolyQ
14 kb & 47 variable sites
Enard et al., Nature, 2002
# of Singletons = 31θw = 7.9 10−4
π = 3.0 10−4
θH = 11.7 10−4
Tajimas D = −2.20Fay & Wu H = −12.24

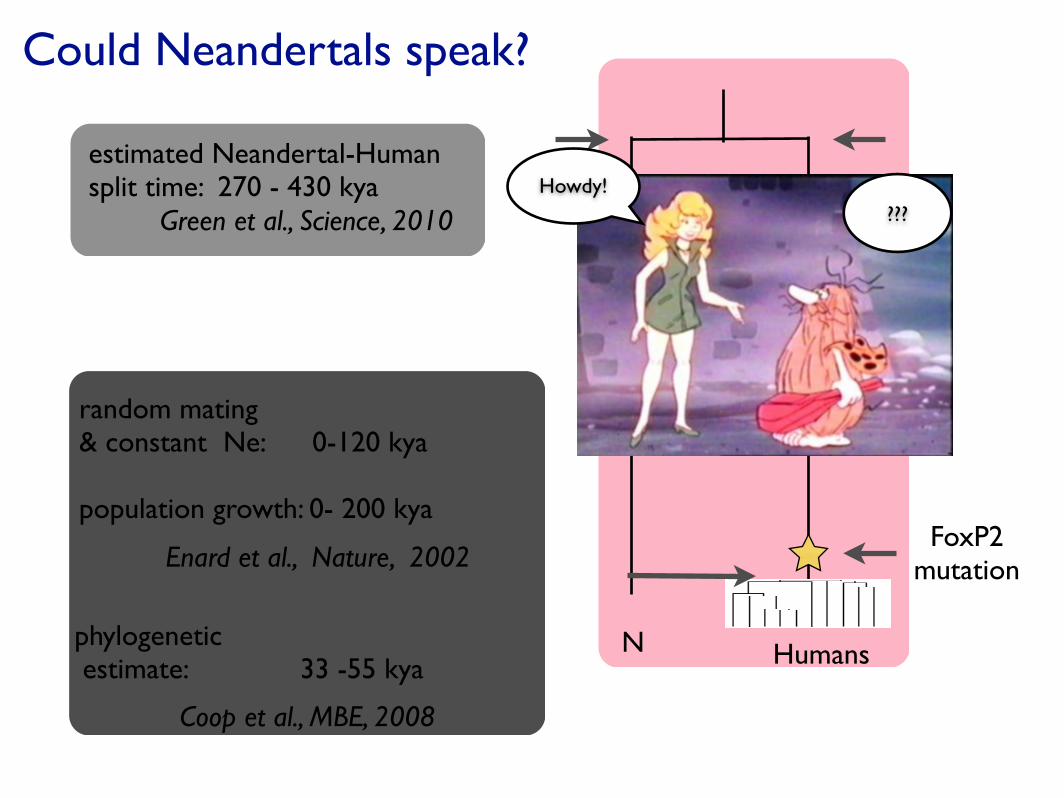
Could Neandertals speak?
random mating & constant Ne: 0-120 kya population growth: 0- 200 kya
Enard et al., Nature, 2002
Coop et al., MBE, 2008
phylogenetic estimate: 33 -55 kya
estimated Neandertal-Humansplit time: 270 - 430 kya
Green et al., Science, 2010
N Humans
FoxP2 mutation
random mating & constant Ne: 0-120 kya population growth: 0- 200 kya
Enard et al., Nature, 2002
Coop et al., MBE, 2008
phylogenetic estimate: 33 -55 kya
estimated Neandertal-Humansplit time: 270 - 430 kya
Green et al., Science, 2010
N Humans
FoxP2 mutation
Howdy!???
Could Neandertals speak?

FOX
1 2 3 4 5 6 7 8 9 10 111213 141516 17
atg tga
PolyQ
T->N N->S
Krause et al., Current Biol., 2007
But Neandertal has both human specific amino acids:
where Neandertals and contemporary humans can bedistinguished on the basis of several substitutions [8]fixed in each group. Among 46 DNA extracts preparedfrom 22 Neandertal bones, we identified two boneswhere approximately 98%–99% of the hominid mtDNAin the extracts are of the Neandertal type andwhere DNAis abundant enough for PCR. Both bones originate fromEl Sidron Cave in Asturias (north of Spain) [9]. Thesebones (numbers 1253 and 1351c) were removed understerile conditions directly from the excavation in 2006,immediately frozen, and transported to our clean roomfacility where extractions were performed. Becauseadditional DNA sequences from the mtDNA hypervari-able region show that they stem from different individ-uals (data not shown), they are referred to as the firstand the second Neandertal, respectively, below.Second, we designed a number of additional controls
for detecting modern human nuclear DNA contamina-tion. To this end, we used genomic sequence data pro-duced from a 38,000-year-old Neandertal from VindijaCave, Croatia [8] to identify seven sequence positionson autosomes and the X chromosome that are ancestral(i.e., identical to the chimpanzee sequence) in the VindijaNeandertal but derived (i.e., different from the chimpan-zee sequence) and not known to vary among current hu-mans. We avoided C to T or G to A differences becausethese are common artifacts of ancient DNA sequences[10–14]. We chose several control positions based onthe Vindija Neandertal because it is not certain thatany particular one would be shared with the El SidronNeandertals. Furthermore, given thatmost extracts con-tain very low amounts of Neandertal DNA, we expectonly a fraction of amplifications of nuclear target se-quences in any one experiment to yield products.Third, we took advantage of the detailed modern hu-
man Y chromosome phylogeny [15] and the fact thatthe time to the most recent common ancestor for mod-ern human Y chromosomes, approximately 90,000 yearsago [16], is more recent than the estimated populationsplit time with Neandertals [17]. A priori we therefore ex-pect Neandertal Y chromosomes to fall outside the var-iation of modern human Y chromosomes unless therewas male gene flow frommodern humans into Neander-tals. This allowed us to design primers that amplify fivepositions that define the deepest nodes in the tree ofcurrent Y chromosomes as well as subgroups includingalmost all European and Asian and most African Y chro-mosomes (Figure S1 in the Supplemental Data availableonline).We combined all control primer pairs with different
primer sets for the FOXP2 gene in four different primer
mixes and performed two-step multiplex PCRs [18]. Allprimers were amplified for 30 cycles in a first PCR fromwhich aliquots were removed and then used to amplifyeach specific target individually in a second PCR. Foreach primermix and individual, we also performedmockamplifications containing no template DNA. None of 108secondary PCRs from such negative controls yieldedany specific product. The results for the twoNeandertalsare summarized in Figure 2.For the autosomal controls, nine out of 20 secondary
PCRs yielded the relevant products. The fact that notall primer pairs yield products shows that the extractscontain such small amounts of nuclear DNA that ampli-fication success is only sporadic and likely to often startfrom single-templatemolecules [10]. For two of the threeautosomal positions analyzed, both individuals carry theancestral allele. Out of the five products retrieved for thethird autosomal position (A1, Table S1), both the ances-tral and derived alleles are found in both El Sidron Nean-dertals, suggesting that they were polymorphic at thisposition. The second individual in addition carried an al-lele not seen in modern humans or chimpanzees. Giventhat this sequence variant is a G to T substitution, nota substitution commonly associated with ancient DNAdamage [10], it is likely to represent a genuine allelepresent in the Neandertals. However, further workwouldbe necessary to verify this. Out of four X chromosomalpositions, only one yielded products in one individual.It carried the ancestral state. Both Neandertals yieldedproducts for Y chromosomal primer pairs, indicatingthat they were males. Strikingly, all 15 Y chromosomalproducts for the five assayed positions show the ances-tral allele. This includes two polymorphisms that definethe deepest split among current human Y chromosomes(Y2 and Y4, Figure S1) as well as two polymorphismsthat cover less common African Y chromosomes (Y3and Y5, Figure S1). These Y chromosome results mustderive, then, either from Y chromosomes that fall out-side the variation of modern humans or from the veryrare African lineages not covered by the assay (Fig-ure S1). For our purposes, this result shows that neitherthe maternally inherited mtDNA nor the paternally in-herited Y chromosome shows evidence of gene flowfrommodern humans into Neandertals or of subsequentcontamination of their mortal remains. From these re-sults, we infer that the multiplex-PCR products derivefrom Neandertal nuclear-DNA templates.Of the two FOXP2 substitutions in exon 7, position 911
was retrieved four times from the first individual andtwice from the second. In all cases, the recovered allelewas the derived allele seen in modern humans. Position
Figure 1. Sequence Alignment of Nucleotide Positions 880–1020 from the FOXP2 Gene
The two nonsynonymous nucleotide substitutions on the human lineage are indicated by arrows. Identical positions in the alignment are given asdots. The three primer pairs used to retrieve the two substitutions from the El Sidron Neandertals are indicated by arrows.
Current Biology Vol 17 No 212
Please cite this article in press as: Krause et al., The Derived FOXP2 Variant of Modern Humans Was Shared with Neander-tals, Current Biology (2007), doi:10.1016/j.cub.2007.10.008

FOX
1 2 3 4 5 6 7 8 9 10 111213 141516 17
atg tga
PolyQ
T->N N->S
Krause et al., Current Biol., 2007
But Neandertal has both human specific amino acids:
where Neandertals and contemporary humans can bedistinguished on the basis of several substitutions [8]fixed in each group. Among 46 DNA extracts preparedfrom 22 Neandertal bones, we identified two boneswhere approximately 98%–99% of the hominid mtDNAin the extracts are of the Neandertal type andwhere DNAis abundant enough for PCR. Both bones originate fromEl Sidron Cave in Asturias (north of Spain) [9]. Thesebones (numbers 1253 and 1351c) were removed understerile conditions directly from the excavation in 2006,immediately frozen, and transported to our clean roomfacility where extractions were performed. Becauseadditional DNA sequences from the mtDNA hypervari-able region show that they stem from different individ-uals (data not shown), they are referred to as the firstand the second Neandertal, respectively, below.Second, we designed a number of additional controls
for detecting modern human nuclear DNA contamina-tion. To this end, we used genomic sequence data pro-duced from a 38,000-year-old Neandertal from VindijaCave, Croatia [8] to identify seven sequence positionson autosomes and the X chromosome that are ancestral(i.e., identical to the chimpanzee sequence) in the VindijaNeandertal but derived (i.e., different from the chimpan-zee sequence) and not known to vary among current hu-mans. We avoided C to T or G to A differences becausethese are common artifacts of ancient DNA sequences[10–14]. We chose several control positions based onthe Vindija Neandertal because it is not certain thatany particular one would be shared with the El SidronNeandertals. Furthermore, given thatmost extracts con-tain very low amounts of Neandertal DNA, we expectonly a fraction of amplifications of nuclear target se-quences in any one experiment to yield products.Third, we took advantage of the detailed modern hu-
man Y chromosome phylogeny [15] and the fact thatthe time to the most recent common ancestor for mod-ern human Y chromosomes, approximately 90,000 yearsago [16], is more recent than the estimated populationsplit time with Neandertals [17]. A priori we therefore ex-pect Neandertal Y chromosomes to fall outside the var-iation of modern human Y chromosomes unless therewas male gene flow frommodern humans into Neander-tals. This allowed us to design primers that amplify fivepositions that define the deepest nodes in the tree ofcurrent Y chromosomes as well as subgroups includingalmost all European and Asian and most African Y chro-mosomes (Figure S1 in the Supplemental Data availableonline).We combined all control primer pairs with different
primer sets for the FOXP2 gene in four different primer
mixes and performed two-step multiplex PCRs [18]. Allprimers were amplified for 30 cycles in a first PCR fromwhich aliquots were removed and then used to amplifyeach specific target individually in a second PCR. Foreach primermix and individual, we also performedmockamplifications containing no template DNA. None of 108secondary PCRs from such negative controls yieldedany specific product. The results for the twoNeandertalsare summarized in Figure 2.For the autosomal controls, nine out of 20 secondary
PCRs yielded the relevant products. The fact that notall primer pairs yield products shows that the extractscontain such small amounts of nuclear DNA that ampli-fication success is only sporadic and likely to often startfrom single-templatemolecules [10]. For two of the threeautosomal positions analyzed, both individuals carry theancestral allele. Out of the five products retrieved for thethird autosomal position (A1, Table S1), both the ances-tral and derived alleles are found in both El Sidron Nean-dertals, suggesting that they were polymorphic at thisposition. The second individual in addition carried an al-lele not seen in modern humans or chimpanzees. Giventhat this sequence variant is a G to T substitution, nota substitution commonly associated with ancient DNAdamage [10], it is likely to represent a genuine allelepresent in the Neandertals. However, further workwouldbe necessary to verify this. Out of four X chromosomalpositions, only one yielded products in one individual.It carried the ancestral state. Both Neandertals yieldedproducts for Y chromosomal primer pairs, indicatingthat they were males. Strikingly, all 15 Y chromosomalproducts for the five assayed positions show the ances-tral allele. This includes two polymorphisms that definethe deepest split among current human Y chromosomes(Y2 and Y4, Figure S1) as well as two polymorphismsthat cover less common African Y chromosomes (Y3and Y5, Figure S1). These Y chromosome results mustderive, then, either from Y chromosomes that fall out-side the variation of modern humans or from the veryrare African lineages not covered by the assay (Fig-ure S1). For our purposes, this result shows that neitherthe maternally inherited mtDNA nor the paternally in-herited Y chromosome shows evidence of gene flowfrommodern humans into Neandertals or of subsequentcontamination of their mortal remains. From these re-sults, we infer that the multiplex-PCR products derivefrom Neandertal nuclear-DNA templates.Of the two FOXP2 substitutions in exon 7, position 911
was retrieved four times from the first individual andtwice from the second. In all cases, the recovered allelewas the derived allele seen in modern humans. Position
Figure 1. Sequence Alignment of Nucleotide Positions 880–1020 from the FOXP2 Gene
The two nonsynonymous nucleotide substitutions on the human lineage are indicated by arrows. Identical positions in the alignment are given asdots. The three primer pairs used to retrieve the two substitutions from the El Sidron Neandertals are indicated by arrows.
Current Biology Vol 17 No 212
Please cite this article in press as: Krause et al., The Derived FOXP2 Variant of Modern Humans Was Shared with Neander-tals, Current Biology (2007), doi:10.1016/j.cub.2007.10.008
Howdy! And how do you do

Alternative Scenarios:
N Humans
Old Model:

Alternative Scenarios:
N Humans
Old Model:
N Humans
1. Sweep in ancestral population or on standing variation?

Alternative Scenarios:
N Humans
Old Model:
N Humans
1. Sweep in ancestral population or on standing variation?
N Humans
2. Admixture of humans and Neandertals

Alternative Scenarios:
N Humans
Old Model:
N Humans
1. Sweep in ancestral population or on standing variation?
N Humans
2. Admixture of humans and Neandertals
N Humans
?
3. Neither of the amino acid changes caused the sweep

After a sweep, high frequency ancestral alleles do rarely occur on the same haplotype:

FOX
1 2 3 4 5 6 7 8 9 10 111213 141516 17
atg tga
PolyQ14kb
4kb3kbSamples:14 Nigerians,2 Melanesians1 Russian
-1
0
1
2
3
4
5
6
7
8
9
10
11
12
-1 0 1 2 3 4 5 6 7 8 9 10 11 12
10.8 - 1.00.6 - 0.80.4 - 0.60.2 - 0.40.0 - 0.2significant LDmarginal LD
R2/D'
acrossexon 7
acrossexon 7
downstreamexon 7
upstreamexon 7
37 40 54 55 56 58 59 60 61 62 63
63
62
61
60
59
58
56
55
54
40
37

Assuming we don’t know the Neandertal: what are the chances for the LD-pattern?
Selected amino acid site got on the ancestral background dueto gene conversion during the sweep.
The ancestral haplotype is a double recombinant.
P (DR ∧ no SR) < 0.001
P (GC | frl = 4 10−6) = 0.02P (GC | frl = 2 10−5) = 0.06P (GC | frl = 2 10−4) = 0.40

Howdy! And how do you do my dear?
Maybe Neandertals and Modern Humans
could speak with one another and lived happily ever after.
(or until the Neandertals disappeared)
Conclusions

ANCESTRAL STATE A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A C C T G C A A G A A T G AOrang-Utan A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A C C T G C A A G A A T G AChimpanzee A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A Y C T G C A A G A A T G A
Human Consensus A A A T A G A A A C A C C A G A G A G A G A C G T C C G A T A C G T G C T A T A A T G A1 Australian Abor. "HAY-KJ" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - G - - - - - - - - - - - - -2 Warao South Amer Ind "SPACH"- - G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -3 Chinese "XLI-ND" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -4 Japanese "KT12" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -5 Thai "DC009" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- T - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -6 PNG (coastal) "100" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - -7 PNG (highlander) "35" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - -8 Nasioi (Melanesia) "966" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -9 Iranian "KHAGNI" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - T - - - - - - - - G - - -10 French "DUCAF" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - - - - - - - - - - G - - -12 German "EHM" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - - - - - - - - - - G - - -13 English "IHAF" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - G - - - - - - - - - - G - - - - - - - - - - - - - - - - - - - - - - - - - - - -14 Italian "WT24" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -15 Mbuti Pygmy "1039" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - T - - - - A - - - C - - - - - - - - - - - - - - - T T - C - - - C - -16 Biaka Pygmy "785" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -17 Biaka Pygmy "805" - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - G - - - A - A G C - - - - - - T A - - A - - - - T T - - - - - C - -18 Nigerian (Ibo) "044" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
C - - G - A G T - - - T T G A - - - - T A - - - - - - - C A - - C C - - A - G T - - - G19 Nigerian (Yoruba) "025" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - G - - - - - A - - - - - - G - - - - - - - - - - - - - T - - - - - - - -20 Nigerian (effik) "002" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - G - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -21 Nigerian (Hausa) "006" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - C - - A - - - - - - - - - - - - - - A -
Neandertal A C T G T
A
D
TC/T
... and not only that, the Neandertal also has the derived haplotype.

ANCESTRAL STATE A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A C C T G C A A G A A T G AOrang-Utan A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A C C T G C A A G A A T G AChimpanzee A A A T A G A T A C A C T G A A G A G A A A C G T C C G A A A Y C T G C A A G A A T G A
Human Consensus A A A T A G A A A C A C C A G A G A G A G A C G T C C G A T A C G T G C T A T A A T G A1 Australian Abor. "HAY-KJ" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - G - - - - - - - - - - - - -2 Warao South Amer Ind "SPACH"- - G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -3 Chinese "XLI-ND" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -4 Japanese "KT12" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -5 Thai "DC009" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- T - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -6 PNG (coastal) "100" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - -7 PNG (highlander) "35" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - -8 Nasioi (Melanesia) "966" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -9 Iranian "KHAGNI" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - T - - - - - - - - G - - -10 French "DUCAF" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - - - - - - - - - - G - - -12 German "EHM" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - - - - - A - - - - - - - T - - T A - - A - - - - - - - - - - G - - -13 English "IHAF" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - G - - - - - - - - - - G - - - - - - - - - - - - - - - - - - - - - - - - - - - -14 Italian "WT24" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -15 Mbuti Pygmy "1039" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - T - - - - A - - - C - - - - - - - - - - - - - - - T T - C - - - C - -16 Biaka Pygmy "785" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -17 Biaka Pygmy "805" - - - - - - - - - - - - - - A - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - G - - - - - - G - - - A - A G C - - - - - - T A - - A - - - - T T - - - - - C - -18 Nigerian (Ibo) "044" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
C - - G - A G T - - - T T G A - - - - T A - - - - - - - C A - - C C - - A - G T - - - G19 Nigerian (Yoruba) "025" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - G - - - - - A - - - - - - G - - - - - - - - - - - - - T - - - - - - - -20 Nigerian (effik) "002" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - G - - - - - A - - - - - - - - - - - - - - - - - - - - T - - - - - - - -21 Nigerian (Hausa) "006" - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - A - - - - - - - - - C - - A - - - - - - - - - - - - - - A -
Neandertal A C T G T
A
D
TC/T
... and not only that, the Neandertal also has the derived haplotype.
Howdy! And how do you do

1. & 2. Haplotype D was present in the ancestral populationor selection in the ancestral population
HFD-SNPs
Neandertal
N(1/1) H1
(39/40)H2
(1/40)
sweep starts
The chances of observing this tree are low!
0.000
0.005
0.010
0.015
0.020
0.025
0.030
0.035
0.040
0.045
0 0.0001 0.0002 0.0003 0.0004
population recombination rate
low, old high, old
low, recent high, recent
low, old high, old
low, recent high, recent
prob
abili
ty





























