Embed Size (px)
Citation preview

FORECLOSURES AND INVASIVE INSECT SPREAD:THE CASE
OF ASIAN CITRUS PSYLLID
TIMOTHY J. RICHARDS, DAVID W. SHANAFELT, AND ELI P. FENICHEL
Human economic activity is considered to be an important factor in exacerbating the speed ofinvasive species spread, but may also play an important role in preventing it. In this study, weinvestigate the role of home foreclosure in the spread of Asian Citrus Psyllid (ACP, Diaphorinacitri Kuwayama) throughout residential areas of Southern California. We find that foreclosures areindeed a significant factor in explaining ACP spread, even after controlling for other human andenvironmental effects. Our results suggest that human economic activity may be more importantin controlling the spread of invasive species than previously realized, and that the external costs ofthe foreclosure problem may also be underestimated.
Key words: Asian Citrus Psyllid, citrus greening disease, foreclosure, gravity model, invasivespecies, property rights, reaction-diffusion model.
JEL codes: C23, Q57.
Understanding the movement of invasivespecies in a host environment is critical todetection, management, and eradication.Statistical models of insect spread typicallyinclude measures of the time and distancefrom introduction, biologic suitability of thehost environment, and perhaps a controlactivity such as spraying or the introductionof natural predators.1 Only recently, however,has the role of human economic activity beenrecognized as exacerbating the speed of inva-sive species spread (Bossenbroek, Kraft, andNekola 2001; Sharov et al. 2002; Gilbert et al.2004; Muirhead et al. 2006). Most insectshave the ability to move only short distanceson their own over short periods of time, solong-distance dispersion is typically thoughtto result from human transport. Becausehuman-monitoring is an important tool fordetecting and eradicating invasive species
Richards is the Morrison Professor of Agribusiness in theMorrison School of Agribusiness, Arizona State University.Shanafelt is a PhD student in the School of Life Sciences, Ari-zona State University, and Fenichel is an Assistant Professor inthe School of Forestry and Environmental Economics at YaleUniversity. Correspondence may be sent to: [email protected] from the Specialty Crop Research Initiative (NIFA-USDA, grant no. 2010-01182) is gratefully acknowledged. Allconclusions remain the authors and do not represent officialpositions of the USDA.
1 See Liebhold and Tobin (2008), or Hastings et al. (2005) forrecent reviews of the modeling literature.
in urban areas, abandoned homes providea potentially critical pathway for uninter-rupted spread (USDA-APHIS 2012). In fact,home ownership provides an example ofhow property owners’ self interest can helpmitigate the spread of invasive species. In thisstudy, we investigate the role of home fore-closure in the spread of Asian Citrus Psyllid(ACP, Diaphorina citri Kuwayama) insectsthroughout residential areas of SouthernCalifornia.
The ACP is perhaps the pest most fearedby citrus growers. As a vector for the bacteriathat causes citrus greening disease, or huang-longbing (HLB), it can devastate citrus overan entire region. Huanglongbing causes citrusyields to fall, drastically reduces fruit quality,can ultimately kill the tree itself, and has noknown cure once a tree is infected (Halbertand Manjunath 2004). In 2009, Californiacitrus represented nearly $1.8 billion indirect cash receipts, $3.0 billion in total eco-nomic activity, and some 26,000 jobs in directand related activities (USDA-ERS 2012;Richards 2009). Therefore, the potential eco-nomic damage caused by widespread ACPmovement is indeed significant, but thehuman role in mediating ACP spread, andperhaps facilitating greater damage, is notwell understood.
Invasive insect species follow a three-stageprocess when invading a new environment:
Amer. J. Agr. Econ. 96(3): 615–630; doi: 10.1093/ajae/aat099Published online December 24, 2013
© The Author (2013). Published by Oxford University Press on behalf of the Agricultural and Applied EconomicsAssociation. All rights reserved. For permissions, please e-mail: [email protected]
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

616 April 2014 Amer. J. Agr. Econ.
introduction, establishment, and spread(Liebhold and Tobin 2008). Once an invasivepopulation is established, optimal manage-ment programs require a careful parameteri-zation of its spread (Epanchin-Niell andWilen 2012). There are essentially four typesof biological models that describe thespread of invasive species: (a) reaction-diffusion models (Skellam 1951); (b) integro-difference equation models (Kawasaki andShigesada 2007; Kot, Lewis, and Van denDrissche 1996; Neubert and Parker 2004);(c) gravity models (Bossenbroek, Kraft,and Nekola 2001); and (d) stratified dif-fusion models (Shigesada, Kawasaki, andTakeda 1995; Havel, Shurin, and Jones 2002;Gilbert et al. 2004; Muirhead et al. 2006).Reaction-diffusion models tend to explainlocalized spread well, but imply continuousrates of expansion, asymptotically constantspeeds of wave travel, and Gaussian disper-sion from the origin that is not necessarilydescriptive of most invasive insect popula-tions. Empirical observation tends to findthat spread is rather leptokurtic, exhibit-ing fat tails rather than Gaussian spread(Hastings et al. 2005; Liebhold and Tobin2008). Integro-difference equations are moreappropriate for explaining populations thattend to spread in non-continuous waves, butdo not explain seemingly random, long-termdispersal. Stratified diffusion and gravitymodels, on the other hand, are well-suitedto explain both local spread and long-termdiffusion while permitting empirical testsof mechanisms that facilitate both. A strati-fied diffusion model assumes that insects “...disperse by two independent dispersal pro-cesses occurring at different spatial scales...,”(Gilbert et al. 2004), one over short distanceswithin the range of natural movement, andone over longer distances, generally at rangesthat can only be explained by aided trans-port. In this paper, we estimate the spread ofACPs throughout Southern California using astratified diffusion model in which we intro-duce measures of home-foreclosure activityas an explicit argument in the rate of invasiveinsect spread.
Mass foreclosures impose a number ofexternalities, some expected and some not.The effect of foreclosures on neighboringhome prices is relatively well-understood, asis the effect on local tax receipts (Immergluckand Smith 2006a; Schuetz, Been, and Ellen2008; Lin, Rosenblatt, and Yao 2009). Lessobvious, but nonetheless apparent, are the
effects of foreclosures on the rate of violentcrime (Immergluck and Smith 2006b) andthe psychological and physiological healthof those going through a foreclosure (Pol-lack and Lynch 2009). When a homeownerabandons a foreclosure, not only does thecommunity lose a monitor with a vestedinterest in maintaining the health of his orher trees, but gains an owner with the indif-ference of scale (the bank) and often losesaccess to trap and (if required) spray to con-trol the invasive insect. Moreover, lowerhome values reduce existing homeowners’incentive to manage their own trees. To theextent that foreclosed homes are rented bybanks, renters are likely to have less of aninterest in monitoring and reporting insectson trees that they do not own.
Further, the potential economic damageof widespread movement of HLB-infectedACP has drawn the California Departmentof Agriculture (CDFA) into partnershipwith not only the USDA, but state-level cit-rus grower organizations (California CitrusMutual) and research organizations (Cali-fornia Citrus Research Board) in the hopesthat combining resources will make theirjoint efforts more effective. Detection andtreatment by the CDFA, however, is moredifficult, if not impossible, without access toforeclosed homes. One of the key strategiesused by the CDFA to control the spread ofACPs is a detection hotline. The existence ofunoccupied homes represents a weakness inthis strategy, as reporting requires monitor-ing. With a high concentration of foreclosuresin an urban neighborhood, the result is thusa virtually uninterrupted path by whichACPs can migrate through residential areason their way to other, perhaps commercial,orchards.
Controlling the spread of an invasivespecies is sufficiently complex that removingone pathway does not necessarily solve theproblem. In fact, some find that it is not theabsence but the presence of human economicactivity that facilitates spread (Gilbert et al.2004). Transporting infested citrus, failingto clean trucks or farm implements, or shar-ing implements are but three ways that ahigher population density can increase theprobability of infestation. This observationemphasizes both the need to control forother types of activity in identifying the fore-closure effect, as well as the more generalinsight that eliminating one externality may,in fact, create others.
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

Richards, Shanafelt, and Fenichel Foreclosures and Invasive Insect Spread: The Case of Asian Citrus Psyllid 617
We contribute to the literature on inva-sive species management by documentingan important class of human-insect interac-tions that has previously been ignored: theeffect of home abandonment (or orchardabandonment of any type) on the rate atwhich invasives are allowed to spread. Ourfindings suggest a very simple implication,namely that homeowners with a vested eco-nomic interest in maintaining the health oftheir citrus can serve as an effective, decen-tralized means of controlling spread. Ourcase differs from others in the literaturebecause visitors to public recreation areasor campgrounds have no property rightsover the trees or lakes they unwittinglyinfect. Because homeowners have propertyrights over host trees, controlling spreadthrough residential areas is a fundamentallydifferent issue than spread through publiclands. We also contribute to the literature onspread modeling by introducing an econo-metric model of ACP diffusion that not onlyincludes gravity-type measures of populationdensity, but explicit measures of economicactivity that are thought to provide a catalyticeffect to insect movement.
The paper is structured as follows. In thefirst section we provide a brief narrativeon the entomological and policy back-ground underlying the ACP problem. Wealso describe our data collection process andsummarize the ACP sightings data in the firstsection. In the second section we develop anempirical model of ACP spread and diffusionthat represents a synthesis of existing gravityand stratified diffusion models. Our estima-tion results are presented in the third section,along with a discussion of their implicationsfor policy and future model efforts. The finalsection concludes.
Asian Citrus Psyllid: Background
Invasive insect species represent a significanteconomic risk to both the financial viabil-ity of specialty crop producers and to thesustainability of U.S. agriculture more gener-ally. With the rapid growth of internationaltrade in agricultural commodities of all types,agricultural systems in the United States areunder constant threat that new species willbe introduced, spread, and thrive in an envi-ronment lacking natural predators. Indeed,Pimentel, Zuniga, and Morrison (2005) claimthat insects alone are responsible for losses
amounting to 13% of total U.S. crop pro-duction, valued at $33.0 billion. Accordingto these authors, 40% of all insects can beregarded as invasive. Therefore, includingthe indirect costs associated with control, thetotal damage attributable to invasive insectspecies in the United States is $13.5 billionin 2001 dollars. The ACP problem is butone example of how invasive insect damagemanifests in one specific industry.
Asian Citrus Psyllids represent a catas-trophic threat if allowed to spread uncheckedwhen the pathogens that cause HLB arealso present (Halbert and Manjunath 2004).The ACP is an invasive insect species thathas been identified in Mexico, CentralAmerica, South America, sub-tropical Asia,Afghanistan, Mauritius, Saudi Arabia, andReunion (Grafton-Cardwell et al. 2006), andhas been present in Florida since 1998 (Hal-bert 1998). Now established throughoutFlorida, ACPs were accidentally introducedto the Rio Grande Valley in Texas in 2001,and were first found in San Diego County,California, in August 2008. Asian CitrusPsyllids have since been sighted in ImperialCounty in October 2008 and Orange and LosAngeles counties in August 2009. As of Octo-ber 2009, the California Cooperative AsianCitrus Psyllid Project (CCACPP) reportedpositive tests for ACPs at 614 sites out of atotal of 107,650 surveyed. Administrativeefforts to stop the spread of ACPs included awide-area quarantine placed on citrus move-ments from parts of San Diego, Riverside,and Imperial counties, and all of Los Ange-les and Orange counties in September 2009,while the USDA announced a $5.8 millionprogram to help prevent the spread of ACPsin California in October 2008. From the firstsighting in 2008 until early 2012, there hadnot been a positive test for HLB in SouthernCalifornia. However, in late March 2012,CDFA officials reported the first positivetest in a hybrid-lemon tree in a residentialneighborhood, confirming the importanceof orchards in residential landscapes to thelikelihood of devastating damage resultingfrom ACP spread (Nelson 2012).
Asian Citrus Psyllids damage citrus directlyby feeding on the leaf and depositing a largeamount of honeydew, leading to the forma-tion of molds (Manjunath 2004). Duringfeeding, ACPs also deposit a toxin that stuntsthe leaf and causes extreme malformation(Michaud 2004). Adults cluster to feed andmate on leaf bud sites, which impairs their
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

618 April 2014 Amer. J. Agr. Econ.
growth and further reduces fruit yield. Ontheir own, however, these issues are rela-tively minor. More significantly, ACPs vectorthe bacterium that causes HLB, which isoften referred to as “...one of the most dev-astating diseases to citrus in the world...”(Grafton-Cardwell et al. 2006). Huanglong-bing causes a number of physiological prob-lems to the tree and to the fruit itself. Treesinfected with HLB are smaller, have fewerleaves, bloom off-season, and will drop bothleaves and fruit (Halbert and Manjunath2004; Grafton-Cardwell et al. 2006). More-over, fruit from HLB-infected trees are small,lopsided, and hard with immature seeds and,perhaps not surprisingly, green. Most treesinfected with HLB survive only 5–8 yearsand typically fail to produce any valuablefruit over their lifetimes. Although CDFAofficials detected ACPs in San Diego andImperial counties, and found a positive testfor HLB in March of 2012, they have yet tofind widespread evidence of HLB in Califor-nia citrus. In fact, HLB was not detected inFlorida until 2005, despite the presence ofACPs for almost ten years (Meyer, Hoy, andSingh 2007). Because many species closelyrelated to citrus can serve as viable hosts,there are a number of avenues for the spreadof ACPs throughout California.2
Data Description
The primary insect-count data for this studywere gathered under the auspices of theCDFA between October 2008 and March2011. Both yellow panel traps and visualinspection were used to establish counts ofnymph, adult, and dead ACPs at site loca-tions determined by CDFA entomologists.Although visual inspection is not neces-sarily confined to the same location as thetraps, they are reported as such in the CDFA
2 Control methods include a combination of biological, chemi-cal, managerial, and integrated pest management (IPM) strategies.Because ACPs have a number of natural predators, providingan environment with spiders, pirate bugs, or beetles can bean effective control mechanism. In terms of chemical control,systemic uptake of imidacloprid (Admire) is generally used toprotect younger trees, as more mature stands are less affectedby damage due to direct feeding (Grafton-Cardwell et al. 2006).Neonicotinoids or pyrethroids are used if more immediate controlis necessary. Among managerial control methods, tree removalis the most common technique to remove the food source andto prevent further spread within a community. Tree removal,however, means the loss of an industry in the long run. Indeed,in parts of Asia in which CGD is particularly severe, citrus canno longer be grown.
sightings data. That is, each visual sighting isassociated with a specific latitude and longi-tude coordinate in a manner similar to thetrap sightings, so we pool all sightings dataand assume they are generated by the sameprocess. Only positive sightings are reported,so developing a probabilistic indicator ofpresence or absence of ACPs is not possible.In the empirical model below, we take thetruncated nature of the CDFA sightings datainto account. Latitude and longitude mea-sures for each trap are used to establish thelocation of each sighting. Because of the rel-atively long distances between sightings, andthe fact that the earth is not flat, Euclideandistance is not an acceptable method of cal-culating the distance from the first sightingin October 2008 and each subsequent sight-ing. For any appreciable distance there willbe a significant error associated with usingEuclidean distance to measure travel distancefor insects, so we use the great circle dis-tance between each sighting and the origin.3Table 1 provides a summary of the CDFAsightings data, as well as the hypothesizedmediating variables described below.
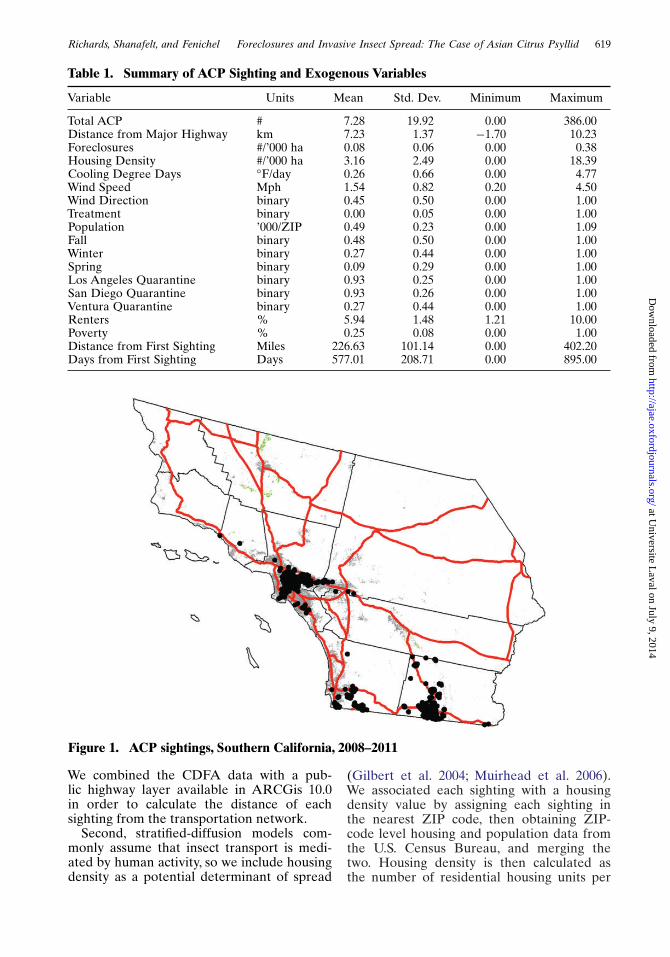
The geographic distribution of sightings isremarkable for the concentration of ACPs inurban areas (figure 1). While invasive speciesare typically regarded as an agricultural prob-lem, ACPs are clearly non-discriminatory inmoving through residential citrus. In fact, itis the stylized fact that ACPs move throughresidential citrus that provides the corehypothesis for this research, namely thathuman-mediated movement represents acritical driver in the spread of any invasivespecies.
We combine the insect-count data with fivemeasures of human and physical environ-ments that may be relevant to insect spread.First, highway transport is often cited as ameans by which invasives are able to movedistances much longer than the limits of self-transport (Godfrey and Grafton Cardwell2006; Burnett et al. 2008; Halbert et al. 2010).
3 Great circle distances are calculated from latitude/longitudecoordinates using the following algorithm. First, we convert thecoordinates to degree decimals by dividing the seconds measure by60. Second, we define latitude and longitude measures for stand-point (s, origin) and forepoint (f , destination) as: φs, λs, φf , andλf , respectively, and convert the result to radians. We then applythe law of cosines: cos �σ = sin φs sin φf + cos φs cos φf cos �λ,where �σ and �λ are the differences between the latitudeand longitude coordinates, respectively. Third, we solve for �σ bytaking the arccos of the result. This gives the geodesic distancein radians. Finally, we multiply by 6,372.8, which is the radius ofthe earth in miles, to arrive at distance measure.
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from
Richards, Shanafelt, and Fenichel Foreclosures and Invasive Insect Spread: The Case of Asian Citrus Psyllid 619
Table 1. Summary of ACP Sighting and Exogenous Variables
Variable Units Mean Std. Dev. Minimum Maximum
Total ACP # 7.28 19.92 0.00 386.00Distance from Major Highway km 7.23 1.37 −1.70 10.23Foreclosures #/’000 ha 0.08 0.06 0.00 0.38Housing Density #/’000 ha 3.16 2.49 0.00 18.39Cooling Degree Days ◦F/day 0.26 0.66 0.00 4.77Wind Speed Mph 1.54 0.82 0.20 4.50Wind Direction binary 0.45 0.50 0.00 1.00Treatment binary 0.00 0.05 0.00 1.00Population ’000/ZIP 0.49 0.23 0.00 1.09Fall binary 0.48 0.50 0.00 1.00Winter binary 0.27 0.44 0.00 1.00Spring binary 0.09 0.29 0.00 1.00Los Angeles Quarantine binary 0.93 0.25 0.00 1.00San Diego Quarantine binary 0.93 0.26 0.00 1.00Ventura Quarantine binary 0.27 0.44 0.00 1.00Renters % 5.94 1.48 1.21 10.00Poverty % 0.25 0.08 0.00 1.00Distance from First Sighting Miles 226.63 101.14 0.00 402.20Days from First Sighting Days 577.01 208.71 0.00 895.00
Figure 1. ACP sightings, Southern California, 2008–2011
We combined the CDFA data with a pub-lic highway layer available in ARCGis 10.0in order to calculate the distance of eachsighting from the transportation network.
Second, stratified-diffusion models com-monly assume that insect transport is medi-ated by human activity, so we include housingdensity as a potential determinant of spread
(Gilbert et al. 2004; Muirhead et al. 2006).We associated each sighting with a housingdensity value by assigning each sighting inthe nearest ZIP code, then obtaining ZIP-code level housing and population data fromthe U.S. Census Bureau, and merging thetwo. Housing density is then calculated asthe number of residential housing units per
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

620 April 2014 Amer. J. Agr. Econ.
thousand hectares. Ownership status is alsolikely to be an important measure of theoccupant’s incentive to maintain the valueof the property and, thereby, monitor andcontrol ACP spread. For this reason, we alsoinclude housing tenure (rentals as a pro-portion of total occupied housing) on a ZIPcode basis from the Census Bureau. Becausepopulation density may also be important inits own right (as a proxy measure of the levelof economic activity) regardless of housingdensity (Gilbert et al. 2004), we include ZIPcode level population as well. Low incomehouseholds may also lack either the ability tomonitor infested trees, or lack the resourcesto treat those that are identified as infected.Therefore, we include a measure of the pro-portion of households earning below thefederal poverty line in each ZIP code, as wellas from the Census Bureau.
Third, we merged ZIP code level fore-closure data obtained from RealtyTrac, Inc.RealtyTrac follows foreclosure activity on amonthly, ZIP code basis. Because foreclosureis a lengthy process, however, there are manydifferent ways to track foreclosure activity.Our data include five measures, each repre-sented as counts at a ZIP code geographylevel: (a) Notice of Default (NOD) is filed bya trustee after a homeowner has defaulted ona mortgage, and represents the initial step inthe pre-foreclosure process; (b) Lis Penden(LIS) is a notification of pending lawsuit andis the first judicial step in the pre-foreclosureprocess as it is filed by lawyers and not thetrustee; (c) Notice of Trustee’s Sale (NTS)is a notice announcing a public auction; (d)Notice (Judgment) of Foreclosure Sale (NFS)is a formal order, signed by a judge, thatinstructs the trustee to publish a “Noticeof Sale” and to sell the property at publicauction; and (e) Real Estate Owned (REO)indicates that the property has been con-veyed back to the lender and represents thefinal step in the foreclosure process. The totalof all five measures represents an indicator ofhow many homes in a given geographic areathere are at any stage of the pre-foreclosureand foreclosure processes.4 Table 1 provides asummary of the foreclosure data.
4 We included foreclosures to capture the fact that foreclosedhomes are likely to be unoccupied, but this variable may alsocapture how the property was managed prior to the foreclosuretaking place. To the extent that financial management and propertymanagement skills tend to be correlated, the foreclosure variablemay reflect the possibility that the property was neglected priorto foreclosure.
Fourth, insect growth and spread arecritically dependent upon weather condi-tions (Halbert and Manjunath 2004). AsianCitrus Psyllid reproduction has been shownto reach optimal levels at approximately65◦ F (Halbert et al. 2010), so we include ameasure of cooling degree days (CDD)—cumulative degrees above 65◦ F in theprevious 30 days—to capture the temper-ature effect on reproduction.5 Long-distanceACP spread is commonly thought to beeither wind-aided or human-aided, becauseACPs are notoriously poor flyers.6 Further,the first ACP sighting in California was inSan Diego County, so virtually all of the cit-rus lies further to the northeast, directly inthe path of the prevailing winds. Therefore,we also include average windspeed and winddirection to measure wind-aided spread. Wemeasure wind direction by including a binaryvariable for observations where the fastesttwo-minute wind (a metric provided by ourdata source) was from the southwest.7 Bothof these variables are from the National Cli-matic Data Center of the U.S. Department ofCommerce (NCDC - DOC).
Fifth, seasonal variation is important toboth ACP spread and detection becauseACPs feed on leaf flush (Grafton-Cardwell2012) that is more likely to appear in bothspring and fall. Although greater oppor-tunities for feeding would suggest moremovement and spread, insect traps of thetype used by CDFA are more effective whentrees are not flushing.8
CDFA control activities may effect theobserved rate of spread in a number of ways.
5 Asian Citrus Psyllid reproduce every 28 days, so 30-daytemperature and wind histories are appropriate.
6 Halbert and Manjunath (2004) cite a lack of reliable dataon ACP flight capabilities, but document flight-limits on similarinsects of less than 1.5 km.
7 More specifically,daily temperature highs and lows for the timeperiod of interest in southern California (latitude <=36.1388◦)were gathered from Historical Climatology Network stationsmaintained by National Oceanic and Atmospheric Administrationvia the NCDC website. Any missing data points were clearedfrom each set. To associate each trap with temperature data,a nearest-neighbor multivariate interpolation relied on latitude,longitude, elevation (meters), and day as independent variables.Open source mathematical computing libraries for Python, SciPy,and NumPy, were used to evaluate the interpolation. Values wereinterpolated for each trap collection event and 29 days previousfrom both the high and low data sets. To calculate CDD, the30 daily temperature values were respectively subtracted by orfrom 65◦ F (18◦ C). All subsequent positive values were summedto obtain CDD from highs and lows at each trap site. A similarprocedure was used to interpolate average daily wind speed anddirection of the fastest two-minute wind at each trap site. A30-day average was taken for each data set at each trap date.
8 We thank a reviewer for pointing this out.
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

Richards, Shanafelt, and Fenichel Foreclosures and Invasive Insect Spread: The Case of Asian Citrus Psyllid 621
Following a positive sighting at a trap site,the surrounding area is typically treated withinsecticide to ensure that any further spreadis controlled. Therefore, we include an indica-tor of whether a site was treated in the pastas an explanatory variable for the numberof ACPs on the current inspection. Similarly,California Department of Agriculture offi-cials enforced citrus quarantines on counties,or portions of counties, following positivesightings. During our sample period, quaran-tines were extended to Orange County onAugust 28, 2009, to Los Angeles County onSeptember 2, 2009, to Northern San DiegoCounty on November 18, 2009, and to Ven-tura, Southern Santa Barbara, and WestRiverside counties on December 31, 2010.Quarantines are likely to negatively impactthe probability of a positive sighting becauselong-distance spread is generally thought toresult from the physical movement of eitherfruit or nursery material from one area toanother, among the other factors describedhere (Grafton-Cardwell 2012).
Empirical Model of ACP Spread
We test for the effect of foreclosures on therate of ACP spread within the context ofa bio-economic model of invasive disper-sal. We specify a stratified diffusion model,which combines a reaction-diffusion model oflocal ACP spread and a long-range dispersalcomponent. In this sense, stratified diffusionmodels are a synthesis of theory-based andpurely empirical models, since they are meantto explain movement that derives from bio-logical factors, environmental factors, andhuman-mediated influences. Further, becausestratified diffusion models nest a reaction-diffusion model and an empirical model oflong-term dispersal, they are inherently moregeneral.
A stratified diffusion process assumes thatshort-range diffusion is determined by fac-tors inherent in the insect, and as such is afunction only of time and distance from theorigination point. Thus, long-range diffusionis driven more by environmental factors.At the core of the short-range portion ofthe spread model is the reaction-diffusionmodel of Skellam (1951). The reaction-diffusion model assumes a population growsat a constant exponential rate, and spreadsaccording to a process based on Fick’s Lawfrom particle physics (Fick 1855). In this
regard, the Skellam (1951) model impliesa continuous, asymptotically constant rateof spread from a well-defined origin andnormally distributed population density. Areaction-diffusion model of ACP spread iswritten as:
(1) b(s, t) = b(s0, t0)(
exp(rt − s2/4ψt)4πψt
),
where the initial population, b(s0, t0), isassigned a value of 1 in October, 2008 sincethat represents the first sighting at t = 0, ands = 0; b(s, t) is the number of ACPs sightedat location s and time t, where s and t aremeasured as the geographic and temporaldistances (in miles and days, respectively)from the first sighting in October, 2008.Moreover, r is the rate of exponential growth,and ψ is the diffusion coefficient. Althoughthe reaction-diffusion model has been shownto perform well in explaining local diffusion,it cannot explain long-distance dispersal(Liebhold and Tobin 2008). Therefore, we fol-low Havel, Shurin, and Jones (2002), Gilbertet al. (2004), and Muirhead et al. (2006) anddesign a stratified-diffusion model in whichlocal diffusion is explained with a reaction-diffusion model, while long-distance dispersalis explained using attributes unique to theenvironments surrounding each trap.
More formally, we test whether strat-ification is necessary by combining thereaction-diffusion model of Skellam (1951)with a term that explains long-range dis-persal. The stratified diffusion model for aninsect population b(s, t) at location s and timet is written as:
b(s, t) = b(s0, t0) exp( f (z))
×(
exp(rt − s2/4ψt)4πψt
),(2)
where f (z) is the linear dispersal kernel thatreflects the influence of economic, and demo-graphic and environmental factors (z) onlong-distance movement. A dispersal kernelrepresents the contribution to the probabilityof observing insects at a particular site thatare more distant than could be explained bynatural diffusion, and is independent of theshort-range or diffusion function (Gilbertet al. 2004). This kernel captures features ofmore distant locations that help explain the
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

622 April 2014 Amer. J. Agr. Econ.
probability of distant observations, and is akernel in the sense that it measures probabil-ities (through its substitution into equation(4) below). We use an exponential form ofthe dispersal kernel in order to facilitateestimation.
The elements of z include each of theenvironmental variables described above:distance to the nearest highway; cumula-tive CDD over the past 30 days; windspeed;wind direction (binary indicator for south-west wind); household density (number ofhouseholds per hectare); total populationin the ZIP code area; rental properties as apercentage of total housing; the proportion ofhouseholds earning below the federal povertylevel; a summary measure of the total rate offoreclosures (per hectare) in the surroundingZIP code; season indicator variables; binaryindicators for whether a particular site hadbeen treated with insecticide in the past; anddummy variables indicating whether or not acounty was under quarantine at the time ofthe sighting.9 Therefore, the dispersal kernelis written as: f (z) = ∑
βk zk where the zk areelements of z and βk is the effect of the kthelement on the contribution to long-termdispersal. Temporal and spatial distances toall other sightings are calculated based onthe initial date and latitude/longitude coordi-nate. Temporal distance is calculated as thenumber of days after t = 0, while distance iscalculated using the great-circle algorithmdescribed above.10
Our data capture only positive sightingsand not traps for which there were no ACPsobserved. Therefore, the dependent variableis truncated. Further, because CDFA over-samples in areas in which there has beena positive sighting, our sample is endoge-nously stratified. Englin and Shonkwiler
9 We included several measures together (NOD, LIS, NTS,NFS, and REO), but they proved highly collinear, so we use anaggregate, total value in the statistical model. Further, an idealdata set would include attributes of the previous owners wholost their homes, as there may be an association between howthe properties were managed previously and their current state.Without these data, we assume biologic and economic neglect tobe independent, and that our ZIP code data describe current aswell as previous residents, on average.
10 We recognize that some of the elements in z could alsoexplain local diffusion, but these elements are commonly usedto explain divergence from diffusion that would otherwise bepredicted by the Skellam (1951) model. In this regard, the Skellam(1951) model is intended to capture diffusion of an insect undernormal environmental conditions. Any deviation from this normal,intrinsic growth rate, which is manifest in unusually long-distancetravel, is captured by factors that may augment the normal processof biological movement.
(1995) develop a method for estimating trun-cated, endogenously-stratified (T-ES) countdata models that simplifies the maximumlikelihood approach of Shaw (1988). Ourmodel extends the Englin and Shonkwiler(1995) approach to allow for a non-linearindex function. Formally, we observe ACPsonly if the latent count value is positive,or: b(s, t) = b∗(s, t), if b∗(s, t) > 0. Assumingthat the conditional density of the numberof ACPs at location s and time t is givenas h(b∗(s, t) | z, s, t, θ), where θ is the set ofparameters in (2) above, then Shaw (1988)shows that the density function for observedcounts is given by:
g(b(s, t) | z, s, t, θ)
= b(s, t)h(b(s, t) | z, s, t, θ)∑∞t=1 t(h(t | z, s, t, θ))
.(3)
Further, if we assume the conditional densityis Poisson with location parameter λ(s, t)(interpreted as the mean number of ACPsper day), then we write the sample densityfunction from the observed sites as:11
g(b(s, t) | z, s, t, θ)
= exp(−λ(s, t))λb(s,t)−1
(b(s, t) − 1)! ,(4)
where b(s, t, ) are the observed ACP val-ues and λ(s, t) is modeled according to thestratified-diffusion model in (2). Englin andShonkwiler (1995) show that (4) is more eas-ily estimated by defining w(s, t) = b(s, t) − 1and estimating using conventional maximumlikelihood methods.
We estimate the non-linear stratified diffu-sion model in (4) using maximum likelihood.The full non-linear stratified-diffusion model(model 3) is compared against alternativespecifications in order to examine the robust-ness of our findings. Namely, we test ourmaintained model (model 3) against a purereaction-diffusion model that accounts forlocal movement only, ignoring long-rangedispersal (model 1), and a linear stratified-diffusion version (model 2) that measuresspread and growth as direct functions ofdistance and time, respectively, without the
11 We choose a daily time interval, as there are cases wherethe same trap was inspected on successive days. We recognize,however, that not all traps will be sampled at the same interval,so the Poisson will only approximate the true arrival probability.
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

Richards, Shanafelt, and Fenichel Foreclosures and Invasive Insect Spread: The Case of Asian Citrus Psyllid 623
structure of the Skellam (1951) model. Wealso estimate a version of the model in whichwe assume, incorrectly, that the dependentvariable is not truncated (model 4). Wereport all estimation results, and the find-ings of our robustness tests, in the followingsection.
Results and Discussion
We begin our presentation of the resultswith a set of specification tests that com-pare a diffusion-only model of ACP spread(model 1) to a stratified-diffusion model thataccounts long-range dispersal, but is linearin time and distance (model 2), and to amodel that includes the non-linear Skellam(1951) diffusion term (model 3). All threeof these models account for the T-ES natureof our data. We then compare the parame-ter estimates from model 3 to one that doesnot account for the T-ES nature of our data,in order to demonstrate the extent of biasinvolved. Once a preferred model is selected,we then interpret the estimated spread andgrowth parameters both in terms of ourhypotheses regarding the likely mediators forACP spread, and the implications for currentmanagement and eradication plans.
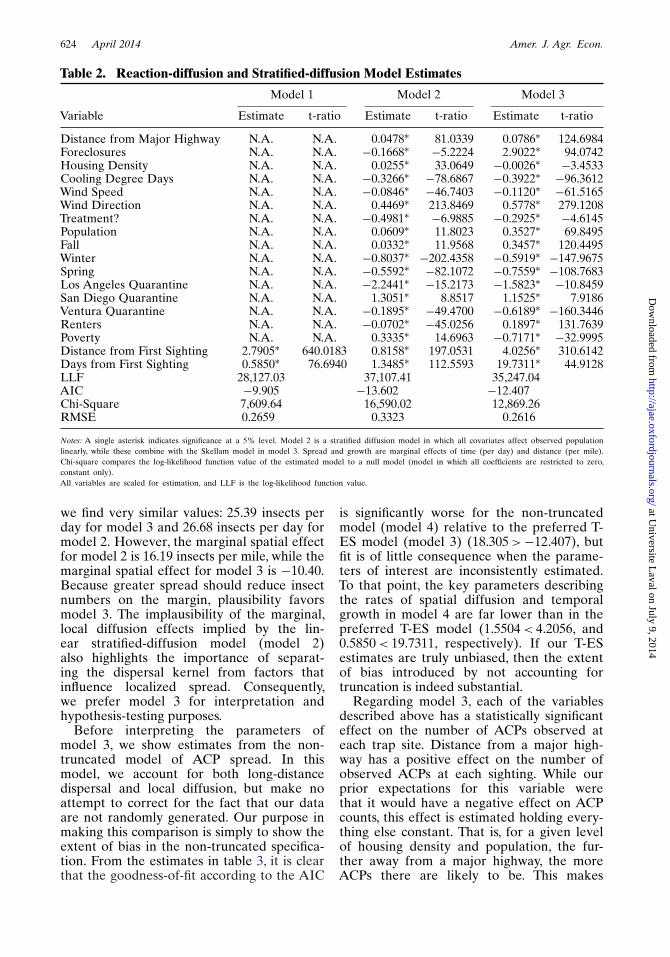
We present the estimates from models 1–3in table 2 below, and for model 4 in table 3.We compare the first three specificationsusing three criteria: (a) likelihood-ratio (LR)tests, (b) root-mean-square error (RMSE)in-sample, (c) Akaike information criterion(AIC), and (d) the significance and plau-sibility of individual parameters. In eachmodel, the dependent variable is the same,so LR tests are an appropriate model selec-tion method. We first compare model 1 tomodel 2 using the LR criteria. For this com-parison, the calculated LR chi-square teststatistic value is 17,960.76, and the criticalchi-square value with 18 degrees of freedomis 26.27, so we easily reject the null hypoth-esis that the two models are equivalent(p-value = 0.00) and prefer model 2. We alsocompare model 1 to model 3 using the LRcriteria, which yields a chi-square test statisticof 14,240.02, again favoring model 3 overmodel 1 (p-value = 0.00). Comparing the twostratified-diffusion models, the LR chi-squaretest statistic with two degrees of freedom(two parameter restrictions are required torender the non-linear and linear models thesame) is 5.99, and the chi-square statistic is
3,720.74 (p-value = 0.00), so we favor model2 over model 3. According to the AIC, weprefer model 2, as the information-loss isminimized in the linear stratified-diffusionmodel. If these were the only evaluation cri-terion, we would reject the theory of insectdiffusion developed by Skellam (1951) infavor of a linear stratified-diffusion model.
Predictive ability, however, yields a dif-ferent ranking. According to predictiveability (as measured by RMSE), model 3predicts substantially better than model2 (0.2596 < 0.3323), while model 3 pre-dicts only marginally better than model 1(0.2616 < 0.2659). Based on this comparison,model 3 appears to explain the data as wellas a model that does not account for long-range dispersal (model 1), and better than anatheoretic (model 2) alternative.
According to our third criteria, the signifi-cance and plausibility of individual parameterestimates, we prefer model 3. The first rea-son is because the parameters describinglong-distance dispersal in model 3 are all sta-tistically significant, the individual parameterestimates of model 1 are likely to be biaseddue to the likely omitted-variables bias inthe local-diffusion-only model. Therefore,we compare the estimates in model 2 withthose in model 3. The second reason for ourpreference is that, because of their primacyto the stratified-diffusion model, we focuson the temporal-growth and spatial-spreadeffects implied by each model. Both the spa-tial and temporal parameters estimated usingmodels 2 and 3 are statistically significant, sosignificance does not provide a useful basisfor comparison.12 Therefore, we calculatethe marginal effect (at the mean of the data)implied by each model and compare it to ourprior expectations based on the theory ofinsect diffusion developed in the entomol-ogy literature (Liebhold and Tobin 2008).Namely, if the population in any given areaexceeds a certain minimum threshold, thenwe expect the growth rate of the populationto be positive. Further, all else being constant,if the same population is spread over a widerdistance, we expect the marginal effect ofdistance from the origin to be negative, asinsects have more difficulty finding matesthe greater the dispersion. Calculating themarginal temporal effect in models 2 and 3,
12 Comparing the size of t-statistics to determine which is“more” statistically significant is not a valid basis for comparison,given that the models differ in structure.
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from
624 April 2014 Amer. J. Agr. Econ.
Table 2. Reaction-diffusion and Stratified-diffusion Model Estimates
Model 1 Model 2 Model 3
Variable Estimate t-ratio Estimate t-ratio Estimate t-ratio
Distance from Major Highway N.A. N.A. 0.0478∗ 81.0339 0.0786∗ 124.6984Foreclosures N.A. N.A. −0.1668∗ −5.2224 2.9022∗ 94.0742Housing Density N.A. N.A. 0.0255∗ 33.0649 −0.0026∗ −3.4533Cooling Degree Days N.A. N.A. −0.3266∗ −78.6867 −0.3922∗ −96.3612Wind Speed N.A. N.A. −0.0846∗ −46.7403 −0.1120∗ −61.5165Wind Direction N.A. N.A. 0.4469∗ 213.8469 0.5778∗ 279.1208Treatment? N.A. N.A. −0.4981∗ −6.9885 −0.2925∗ −4.6145Population N.A. N.A. 0.0609∗ 11.8023 0.3527∗ 69.8495Fall N.A. N.A. 0.0332∗ 11.9568 0.3457∗ 120.4495Winter N.A. N.A. −0.8037∗ −202.4358 −0.5919∗ −147.9675Spring N.A. N.A. −0.5592∗ −82.1072 −0.7559∗ −108.7683Los Angeles Quarantine N.A. N.A. −2.2441∗ −15.2173 −1.5823∗ −10.8459San Diego Quarantine N.A. N.A. 1.3051∗ 8.8517 1.1525∗ 7.9186Ventura Quarantine N.A. N.A. −0.1895∗ −49.4700 −0.6189∗ −160.3446Renters N.A. N.A. −0.0702∗ −45.0256 0.1897∗ 131.7639Poverty N.A. N.A. 0.3335∗ 14.6963 −0.7171∗ −32.9995Distance from First Sighting 2.7905∗ 640.0183 0.8158∗ 197.0531 4.0256∗ 310.6142Days from First Sighting 0.5850∗ 76.6940 1.3485∗ 112.5593 19.7311∗ 44.9128LLF 28,127.03 37,107.41 35,247.04AIC −9.905 −13.602 −12.407Chi-Square 7,609.64 16,590.02 12,869.26RMSE 0.2659 0.3323 0.2616
Notes: A single asterisk indicates significance at a 5% level. Model 2 is a stratified diffusion model in which all covariates affect observed populationlinearly, while these combine with the Skellam model in model 3. Spread and growth are marginal effects of time (per day) and distance (per mile).Chi-square compares the log-likelihood function value of the estimated model to a null model (model in which all coefficients are restricted to zero,constant only).All variables are scaled for estimation, and LLF is the log-likelihood function value.
we find very similar values: 25.39 insects perday for model 3 and 26.68 insects per day formodel 2. However, the marginal spatial effectfor model 2 is 16.19 insects per mile, while themarginal spatial effect for model 3 is −10.40.Because greater spread should reduce insectnumbers on the margin, plausibility favorsmodel 3. The implausibility of the marginal,local diffusion effects implied by the lin-ear stratified-diffusion model (model 2)also highlights the importance of separat-ing the dispersal kernel from factors thatinfluence localized spread. Consequently,we prefer model 3 for interpretation andhypothesis-testing purposes.
Before interpreting the parameters ofmodel 3, we show estimates from the non-truncated model of ACP spread. In thismodel, we account for both long-distancedispersal and local diffusion, but make noattempt to correct for the fact that our dataare not randomly generated. Our purpose inmaking this comparison is simply to show theextent of bias in the non-truncated specifica-tion. From the estimates in table 3, it is clearthat the goodness-of-fit according to the AIC
is significantly worse for the non-truncatedmodel (model 4) relative to the preferred T-ES model (model 3) (18.305 > −12.407), butfit is of little consequence when the parame-ters of interest are inconsistently estimated.To that point, the key parameters describingthe rates of spatial diffusion and temporalgrowth in model 4 are far lower than in thepreferred T-ES model (1.5504 < 4.2056, and0.5850 < 19.7311, respectively). If our T-ESestimates are truly unbiased, then the extentof bias introduced by not accounting fortruncation is indeed substantial.
Regarding model 3, each of the variablesdescribed above has a statistically significanteffect on the number of ACPs observed ateach trap site. Distance from a major high-way has a positive effect on the number ofobserved ACPs at each sighting. While ourprior expectations for this variable werethat it would have a negative effect on ACPcounts, this effect is estimated holding every-thing else constant. That is, for a given levelof housing density and population, the fur-ther away from a major highway, the moreACPs there are likely to be. This makes
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

Richards, Shanafelt, and Fenichel Foreclosures and Invasive Insect Spread: The Case of Asian Citrus Psyllid 625
Table 3. Non-truncated Model Estimates
Variable Estimate t-ratio
Distance from MajorHighway
0.0617∗ 19.6433
Foreclosures −0.1000 −0.7014Housing Density 0.0158∗ 5.0000Cooling Degree Days −0.1948∗ −14.0165Wind Speed −0.0598∗ −7.6314Wind Direction 0.4076∗ 26.1643Treatment? −0.3176∗ −2.2122Population 0.1085∗ 3.3992Fall 0.1186∗ 6.4510Winter −0.5984∗ −27.8858Spring −0.3867∗ −14.3823Los Angeles Quarantine −1.2924∗ −7.8402San Diego Quarantine 0.5669∗ 3.4907Ventura Quarantine −0.2585∗ −11.1953Renters −0.0333∗ −4.6601Poverty 0.2926∗ 3.1553Distance from First
Sighting1.5504∗ 21.5241
Days from First Sighting 0.5850∗ 45.6706LLF −51,193.75AIC 18.053Chi-Square 11,845.39
Note: A single asterisk indicates significance at a 5% level. Chi-squarecompares the log-likelihood function value of the estimated model toa null model (model in which all coefficients are restricted to zero,constant only).
sense, as these other measures capture the“transportation corridor” effect, while theremaining influence likely stems from the factthat most large orchards are far from majorhighways.
Among environment-related variables,CDD (higher values indicating more warmdays) and wind speed have negative effectson ACP counts, while wind direction has apositive effect. Because every increment inCDD is one degree above optimal breed-ing temperature, this negative effect is tobe expected (Halbert et al. 2004). The winddirection effect is also consistent with ourpriors, as it assumes a value of 1 when thewind blows from the southwest. Given thatACPs do not fly very far during their life-times, wind-aided transport is likely to bean important factor in long-term dispersal:if the prevailing winds in the previous 30days were from the southwest, the numberof ACPs rise significantly. However, the neg-ative wind speed effect means that strongerwinds are associated with fewer ACPs at eachtrap site, perhaps suggesting that traps (andvisual inspections) are less effective whenthe wind has been strong enough to blowACPs out of the area (Catling et al. 1972).
Finally, the literature suggests that ACPsfeed more actively during spring and fall(Grafton-Cardwell 2012). This effect could goeither way, as more active feeding may meanthat insects are moving around more, aremore abundant and are therefore easier totrap, or that they are focused on feeding andare less susceptible to being trapped. We findthat ACP counts fall in the spring and rise inautumn, so we cannot conclude definitivelyone way or the other.
Due to the potential damage inflictedby the uncontrolled spread of ACPs, bothfederal and state officials have developeda number of strategies aimed at not onlycontrolling spread, but eradicating the ACPpopulation. One strategy is that each site istreated with insecticide following a positiveACP sighting. As expected, the treatmentbinary variable has a significant negativeeffect on subsequent ACP counts. Anotherstrategy was that the USDA mandated aseries of quarantines on citrus movement ona county or sub-county basis in response toevidence that ACPs had become establishedin a particular region. Our findings show thatthe quarantines imposed in Los Angeles andVentura counties (including south Santa Bar-bara and west Riverside) had the expectednegative effects on ACP movement, while theSan Diego quarantine had a positive effect.Either this quarantine was singularly inef-fective, or this binary variable is associatedwith a period of rapid ACP spread that is notcaptured by any of our other variables.
We now examine our primary hypothesis,that home foreclosures increase the num-ber of ACPs observed at each trap site. Asexpected, foreclosures were found to be a sig-nificant factor in the spread of ACPs acrossSouthern California. Controlling for housingdensity, population, and the proportion ofrenters in the area surrounding a trap, wefind that the number of foreclosures in a ZIPcode is associated with a relatively large (2.9)and significant (t-ratio = 97.074) rise in theexpected number of ACPs. This finding isrobust to various measures of foreclosure,or whether we define foreclosures on a perhectare or per household basis. Acceleratingthe spread of invasive species is an importantexternality not commonly associated withthe foreclosure crisis. It is easily explained,however, as backyards that are not moni-tored for ACPs, either because the owner isnot attentive, or has lost his or her home tobankruptcy and the home is not occupied, are
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

626 April 2014 Amer. J. Agr. Econ.
more likely to serve as ACP hosts. Housingdensity has a negative influence when wecontrol for total population, the proportionof renters, and the number of foreclosuresin the surrounding area. While housing den-sity, as a measure of the level of economicactivity, may be expected to have a positiveeffect in a traditional gravity model, we con-trol for this effect by including populationin the NLSD model. What remains is thepossibility that more multi-family or small-yard housing units are simply associatedwith fewer opportunities to grow citrus.13 Asexpected, population and the proportion ofrenters have strong, positive effects on ACPcounts.
Controlling for long-distance dispersal,we find plausible estimates for the rates ofgeographic spread, and of temporal growth.Localized spread, in the absence of human-or weather-aided transport, occurs at a rate ofabout four-tenths of a mile per year. BecauseACPs do not fly long distances, this estimateis reasonable. Population growth occurs ata rate of 0.025 insects per day, which mayseem to be low but is reasonable given thatthe sightings data used here are actually den-sity estimates, as there is no way to obtainan accurate measure of total populationsthrough random-trapping.
For policy purposes, it is necessary to trans-late our estimates into measures of economicbenefit and cost. Our counterfactual is toassume that policymakers have the abilityto stop foreclosures entirely. If they could,the benefit to doing so, at least in terms ofthe problem at hand, would be removing asignificant number of ACPs from the envi-ronment. The marginal value of reducingthe number of ACPs through non-chemicalmethods is assumed to be the avoided costof additional treatments. The pesticides thathave proven most effective are standardneocontinoids and pyrethroids, at a cost peracre of some $746.99 (in 2013 dollars) basedon the application of pyrethroids, Movento,Delegate and imidacloprid in the fall andspring.14 Based on the estimates reportedabove, each treatment is responsible forremoving 0.29 ACPs, all else being con-stant, per trap site. Removing foreclosures,on the other hand, has a marginal effect of
13 We attempted to create a map of citrus cover during thistime period, but reliable data were not available at the ZIP codelevel of resolution.
14 Grafton-Cardwell, personal communication.
removing approximately 2.9 ACPs, or theequivalent of 10 treatments, or $7,469.90,assuming each treatment covers one acresurrounding the trap site. Aggregating overall sites provides the total value of eliminat-ing the foreclosure effect of $19.84 million,based on 2,656 unique trap sites in our data.Although this estimate is based on a numberof assumptions, it is based on reasonable andconservative estimates of the total savings byeliminating this source of externality. Froma policy perspective, this is the benefit thatshould be added to the other, numerous ben-efits of resolving the foreclosure crisis, andcompared to the cost of doing so. Accountingfor these other benefits, and the ultimatecost of helping homeowners, is beyond thescope of this paper, but would seem to be areasonable exercise for an agency with moremacro-policy responsibilities.
The implications of our findings areimportant on a number of levels. The firstimplication is that we identify and quantifyan important external effect of the fore-closure crisis that has been previouslyoverlooked. While it is well-understood thaturban blight imposes significant managementcosts on municipalities, our findings highlightadditional costs to the environment and toagricultural production. Once regarded asan urban problem, foreclosures represent aclear pathway through which the built envi-ronment imposes an unintended cost on foodproduction. More generally, if foreclosuresaid the spread of ACPs, then they are likelyto have the same effect on the spread ofother invasive species, controlling for othermeasures of economic activity as we do.When state officials were negotiating withthe major mortgage lenders in settlementof the lawsuit regarding unfair lending andforeclosure practices, we would hope thatexternalities like the one we identify weretaken into account (Dennis and Horwitz2012). Because it is more than likely theywere not, we would hope that any part of thesettlement that goes to public coffers wouldbe directed at remedying externalities such asthe one found here.
Another implication of our findings is thatif humans mediate spread, then humans mustalso serve as important agents of control andprevention. Because they have a vested inter-est in preventing spread, homeowners areperhaps best situated to monitor, report,and manage ACP dispersal throughoutdensely-populated urban areas.
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

Richards, Shanafelt, and Fenichel Foreclosures and Invasive Insect Spread: The Case of Asian Citrus Psyllid 627
Our findings are relevant to industry-sponsored information and monitoring acti-vities, as well as government-led controland eradication programs. Although theCalifornia Citrus Mutual (CCM), the CitrusResearch Board (CRB), and the CaliforniaDepartment of Food and Agriculture(CDFA) have initiated aggressive infor-mation campaigns targeting non-commercialcitrus growers, our findings suggest that theymay need to expand the range of potentialgrove-monitors to include mortgage lenders,city managers, and even commercial realestate management companies that may havecitrus on unoccupied properties. Designingincentives that encourage these groups toparticipate may be problematic.15 Fines fornon-resident owners would be effective, butwould certainly be resisted. One would hopethat local governments—whether munic-ipalities, counties, or even homeowners’associations—have the same broader, socialobjectives as state officials, but that is notlikely the case. Witholding services wouldbe an option. Detection and eradicationare inherently capital- and labor-intensiveprocesses, but our findings provide justifica-tion for intensifying trapping activities on ahousehold-level scale in order to be effec-tive. Deciding on the amount of resourcesto allocate to monitoring invasive speciesin an economically efficient way requirescareful consideration of the marginal costsand benefits involved. Although we quan-tify the benefit to removing a few ACPs byreducing the foreclosure effect, ultimatelythe public benefit from solving the problementirely may include preventing the loss ofthe California citrus industry. If this is thecase, then the cost of micro-level monitoringand eradication may be justified.
Suggesting that these other agents be heldresponsible for monitoring and controllingthe problem, however, begs the question ofhow precisely they would be incentivizedto comply. Many municipalities adopted asimple solution during the foreclosure cri-sis. Namely, in many of the most blightedareas, municipal governments enacted bylawsand regulations intended to make lenderslegally responsible for a full range of main-tenance issues on bank-owned properties.We envision a program similar to that usedin Oakland, CA, in which banks are fined for
15 We thank a reviewer for bringing this point to our attention.
not complying with maintenance ordinances(Drummond 2012). We would recommendthat such ordinances include a clause thatcovers invasive species, as well as trash andother more visible signs of neglect.
Conclusion
Human economic activity is an importantmediating factor in the spread of invasivespecies. International trade, recreationalactivities, commercial transport, or smug-gling are commonly cited as the primarymeans by which invasive plants or animalsmove from an infested area to one that isnot. Our understanding of the range andimportance of these activities, however, is notwell-developed.
In this study, we estimate a stratified-diffusion model of ACP spread throughoutSouthern California from 2008-2011. Local-ized spread is modeled as a Skellam (1951)reaction-diffusion model in which spatial-temporal growth depends on a non-linearfunction of time and distance from the firstsighting, while long-distance ACP spreadis modeled as depending on a host of envi-ronmental factors (temperature, windspeed,and wind direction) and economic factors(distance from a highway, population, treat-ment, quarantine, proportion of renters, andhousing density). Because citrus trees inabandoned backyards represent an ideal,unmonitored host environment for ACPs, weinclude measures of foreclosure activity inthe dispersal kernel of the stratified-diffusionmodel. We find that all factors hypothesizedto be important in long-distance dispersal arestatistically significant.
We also find that the number of foreclosedhomes is an important mediating factor forthe spread of an invasive species. Our find-ing is a unique and important outcome ona number of levels. While the difficultiesforeclosed homes create for citizen-basedmonitoring and eradication programs aresomewhat obvious, our finding adds onemore negative externality associated with themortgage crisis that began in 2007. Invasivespecies spread like crime and blight throughneglected urban environments, creating onemore reason to return foreclosed proper-ties to some form of responsible ownership.Perhaps more important to ongoing controlefforts, if humans mediate spread, they canalso help manage it. Because homeowners
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

628 April 2014 Amer. J. Agr. Econ.
have an incentive to maintain the valueof their homes, our findings suggest thatthey serve as an effective, if decentralized,means of managing the spread of invasivesthroughout residential areas.
Nevertheless, human-aided transport isa well-understood means by which insectsmove over long distances, so removing oneexternality may in fact also restore another.More generally, we show that establishingproperty rights over host environmentscan be an effective means of incentivizingindividual agents so that market control ofan invasive species can be part of a viablesolution, but not necessarily the entiresolution.
Our findings also allow us to parameterizelocal and long-distance ACP spread. Futureresearch, therefore, may use the estimatesshown here in the design of spatial-temporaloptimal control programs. Only by explic-itly calculating the marginal benefits andcosts of ACP eradication and control will itbe possible to determine whether invasivespecies control is an important factor in therestoration of property rights to foreclosedproperties.
One weakness of this study is the nature ofthe CDFA sightings data. Obtaining trap dataon invasive species is always problematic, asresearchers necessarily select trap sites usingheuristics and personal judgement. Datafrom a controlled experiment may capturethe underlying biology more accurately, butwould not be able to measure the underlyinghuman-environmental interactions that wefocus on here.
References
Bossenbroek, J.M., C.E. Kraft, and J.C.Nekola. 2001. Prediction of Long-distance Dispersal Using Gravity Models:Zebra Mussel Invasion of Inland Lakes.Ecological Applications 11: 1178–1788.
Burnett, K., S. D’Evelyn, B.A. Kaiser, P. Nan-tamanasikarn, and J.A. Roumasset. 2008.Beyond the Lamppost: Optimal Pre-vention and Control of the Brown TreeSnake in Hawaii. Ecological Economics67: 66–74.
Catling, H.D. 1972. The Bionomics of theSouth Africa Citrus Psylla, Triozaerytreae (Del Guercio) (Homoptera:Psyllidae). Journal of the EntomologicalSociety of South Africa 35: 235–251.
Drummond, T. 2012. Bank Foreclosuresan Urban Blight. Contra Costa Times.http://www.contracostatimes.com/news/ci_20399645/tammerlin-drummond-bank-foreclosures-an-urban-blight.
Dennis, B., and S. Horwitz. 2012. $26B BankForeclosure Fraud Settlement to beAnnounced Thursday. Washington Post.http://articles.washingtonpost.com/2012-02-08/business/35443965_1_fraudulent-fore-closure-practices-attorney-general-eric-schneiderman-banks.
Englin, J., and S. Shonkwiler. 1995. Estimat-ing Social Welfare Using Count DataModels: An Application to Long-runRecreation Demand Under Conditionsof Endogenous Stratification and Trunca-tion. Review of Economics and Statistics77: 104–112.
Epanchin-Niell, R.S., and J.E. Wilen. 2012.Optimal Spatial Control of BiologicalInvasions. Journal of Environmen-tal Economics and Management 63:260–270.
Fick, A. 1855. Ueber Diffusion. Annalen derPhysik 170: 59–86.
Gilbert, M., J.-C. Gregoire, J.F. Freise, and W.Heitland. 2004. Long-distance Dispersaland Human Population Density Allowthe Prediction of Invasive Patterns in theHorse Chestnut Leafminer (Camerariaohridella). Journal of Animal Ecology 73:459–468.
Godfrey, K., and E. Grafton-Cardwell. 2006.Diaprepes Root Weevil and Asian CitrusPsyllid: Newcomers to California? Pro-ceedings of the California Conference onBiological Control V. Riverside, CA.
Grafton-Cardwell, E.E. 2012. Asian CitrusPsyllid and Huanglongbing Disease.http://www.ipm.ucdavis.edu/PMG/PESTNOTES/pn74155.html.
Grafton-Cardwell, E.E., K.E. Godfrey, M.E.Rogers, C.C. Childers, and P.A. Stansly.2006. Asian Citrus Psyllid. Universityof California, Division of Agricultureand Natural Resources, Publication No.8205.
Halbert, S., and K. Manjunath. 2004. AsianCitrus Psyllids (Sternorrhyncha: Psyll-idae) and Greening Disease of Citrus:A Literature Review and Assess-ment of Risk in Florida. The FloridaEntomologist 87: 330–353.
Halbert, S., K. Manjunath, C. Ramadugu, M.Brodie, S. Webb, and R. Lee. 2010. Trail-ers Transporting Oranges to Processing
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

Richards, Shanafelt, and Fenichel Foreclosures and Invasive Insect Spread: The Case of Asian Citrus Psyllid 629
Plants Move Asian Citrus Psyllids. TheFlorida Entomologist 93: 33–38.
Hastings, A., K. Cuddington, K.F. Davies, C.J.Dugaw, S. Elmendorf, A. Freestone, S.Harrison, et al. 2005. The Spatial Spreadof Invasions: New Developments in The-ory and Evidence. Ecology Letters 8:91–101.
Havel, J.E., J.B. Shurin, and J.R. Jones. 2002.Estimating Dispersal from Patternsof Spread: Spatial and Local Controlof Lake Invasions. Ecology 83: 3306–3318.
Immergluck, D., and G. Smith. 2006a. TheExternal Costs of Foreclosure: TheImpact of Single-family Mortgage Fore-closures on Property Values. HousingPolicy Debate 17: 57–79.
———. 2006b. The Impact of Single-familyMortgage Foreclosures on NeighborhoodCrime. Housing Studies 21: 851–866.
Kawasaki, K., and N. Shigesada. 2007. AnIntegrodifference Model for BiologicalInvasions in a Periodically FragmentedEnvironment. Japan Journal of Industrialand Applied Mathematics 24: 3–15.
Kot, M., M.A. Lewis, and T. van den Driess-che. 1996. Dispersal Data and the Spreadof Invading Organisms. Ecology 77:2027–2042.
Liebhold, A.M., and P.C. Tobin. 2006. Growthof Newly-established Alien Popula-tions: Comparison of North AmericanGypsy Moth Colonies with Inva-sion Theory. Population Ecology 48:253–262.
Liebhold, A.M., and P.C. Tobin. 2008. Popu-lation Ecology of Insect Invasions andTheir Management. Annual Review ofEntomology 53: 387–408.
Lin, Z., E. Rosenblatt, and V.W. Yao. 2009.Spillover Effects of Foreclosures onNeighborhood Property Values. Journalof Real Estate Finance and Economics 38:387–407.
Meyer, J.M., M.A. Hoy, and R. Singh. 2007.Low Incidence of Candidatus Liberib-acter Asiaticus in Diaphorina Citri(Hemiptera: Psyllidae) PopulationsBetween Nov 2005 and Jan 2006: Rele-vance to Management of Citrus GreeningDisease in Florida. Florida Entomologist90: 394–397.
Michaud, J.P. 2004. Natural Mortality ofAsian Citrus Psyllid (Homoptera: Psyl-lidae) in Central Florida. BiologicalControl 29: 260–269.
Muirhead J.R., B. Leung, C. van Overdijk,D.W. Kelly, K. Nandakumar, K. R.Marchant, and H.J. MacIsaac. 2006. Mod-elling Local and Long-distance Dispersalof Invasive Emerald Ash Borer Agrilusplanipennis (Coleoptera) in NorthAmerica. Diversity and Distributions12: 71–79.
Nelson, A. 2012. HLB Confirmed in Cali-fornia. The Packer http://www.thepacker.com/fruit-vegetable-news/HLB-confirmed-in-California-145762445.html.
Neubert, M.G., and I.M. Parker. 2004. Pro-jecting Rates of Spread for InvasiveSpecies. Risk Analysis 24: 817–831.
Pimentel, D., R. Zuniga, and D. Morrison.2005. Update on the Environmen-tal and Economic Costs Associatedwith Alien-invasive Species in theUnited States. Ecological Economics 52:273–288.
Pollack, C.E., and J. Lynch. 2009. Health Sta-tus of People Undergoing Foreclosurein the Philadelphia Region. AmericanJournal of Public Health 99: 1833–1839.
Richards, T.J. 2009. California Citrus in 2009:Impact Analysis and Policy Simulation.Report prepared for the California CitrusMutual, Exeter, CA.
Schuetz, J., V. Been, and I.G. Ellen. 2008.Neighborhood Effects of Concentra-ted Mortgage Foreclosures. Journal ofHousing Economics 17: 306–319.
Sharov, A.A., D. Leonard, A.M. Liebhold,E.A. Roberts, and W. Dickerson. 2002.‘Slow the Spread:’ A National Programto Contain the Gypsy Moth. Journal ofForestry 100: 30–35.
Shaw, D. 1988. On-site Samples’ Regres-sion: Problems of Non-negative Integers,Truncation, and Endogenous Strati-fication. Journal of Econometrics 37:211–223.
Shigesada, N., K.Y. Kawasaki, and Y. Takeda.1995. Modeling Stratified Diffusion inBiological Invasions. American Naturalist146: 229–251.
Skellam, J.G. 1951. Random Dispersal inTheoretical Populations. Biometrika 38:196–218.
U.S. Department of Agriculture. 2012. Ani-mal and Plant Health Inspection Service.http://www.hungrypests.com/the-threat/asian-citrus-psyllid.php.
U.S. Department of Agriculture. 2012.Economic Research Service. Fruit andTree Nut Yearbook. http://usda.mannlib.
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from

630 April 2014 Amer. J. Agr. Econ.
cornell.edu/MannUsda/viewDocumentInfo.do?documentID=1377.
U.S. Department of Commerce, National Cli-matic Data Center. 2012. Washington DC.http://www.ncdc.noaa.gov/oa/ncdc.html.
U.S. Department of Commerce, U.S. CensusBureau. 2012. American Commu-nity Survey. Washington DC. http://factfinder2.census.gov/faces/nav/jsf/pages/searchresults.xhtml?refresh=t.
at Universite L
aval on July 9, 2014http://ajae.oxfordjournals.org/
Dow
nloaded from





























