Embed Size (px)
Citation preview

Scientia Horticulturae 121 (2009) 199–205
Flower greening in phytoplasma-infected Hydrangea macrophylla grown underdifferent shading conditions
Elly Kesumawati a,*, Munetaka Hosokawa a, Takushi Kimata a, Tatsuya Uemachi b, Susumu Yazawa a
a Laboratory of Vegetable and Ornamental Horticulture, Department of Agronomy and Horticultural Science, Graduate School of Agriculture, Kyoto University, Oiwake-cho,
Kitashirakawa, Sakyo-ku, Kyoto 606-8502, Japanb The School of Environmental Science, Department of Biological Resources Management, Shiga Prefecture University, Hassaka-cho 2500, Hikone, Shiga 522-8533, Japan
A R T I C L E I N F O
Article history:
Received 10 July 2006
Received in revised form 6 January 2009
Accepted 13 January 2009
Keywords:
Hydrangea
Saxifragaceae
Light intensity
Flower greening
JHP-phytoplasma
Polymerase chain reaction (PCR)
Green-area ratio
A B S T R A C T
To determine the effect of light intensity on flower greening, the Japanese hydrangea phyllody (JHP)
phytoplasma-infected hydrangea cultivars ‘Midori’, ‘Libelle’, ‘Rosea’ and ‘Madame E. Mouillere’ plants
were grown under different shade conditions. In the first-year experiment, the results indicate that the
flowers of the JHP-phytoplasma-infected hydrangea become green under shaded conditions (70% and
49% sunlight intensities). On the other hand, under full sunlight intensity (100% sunlight intensity), the
flowers of ‘Midori’, ‘Rosea’, and ‘Libelle’ plants were blue, pink or white. To calculate the percentage of
flower greening, inflorescences of these plants were separated and divided into individual flowers, and
classified into four types by green-area ratio, calculated using Adobe Photoshop. Under shading with one
sheet of cheesecloth (70% sunlight intensity), the inflorescences of ‘Midori’, ‘Libelle’ and ‘Madame E.
Mouillere’ plants were composed of more than 40% completely green flowers (0.8 2 green-area ratio),
whereas those of ‘Rosea’ plant had 0% completely green flowers. Under shading with two sheets of
cheesecloth (49% sunlight intensity), the inflorescences of ‘Midori’, ‘Libelle’ and ‘Madame E. Mouillere’
plants had more than 75% completely green flowers; ‘Rosea’ plants had 28%. In the second-year
experiment, under full sunlight intensity, ‘Midori’ plants had four types of flower depending on their
green-area ratio, namely, completely blue or pink, pink-green, greenish and completely green flowers.
Under shading with two sheets of cheesecloth, ‘Midori’ plants had more than 90% completely green
flowers. The JHP-phytoplasma could not be identified by PCR analysis in flowers with a green-area
ratio = 0 (completely blue/pink/white flowers). On the other hand, in flowers with a green-area ratio > 0,
the JHP-phytoplasma was detected by PCR analysis. Thus, we conclude that shading enhances flower
greening in hydrangea by increasing the JHP-phytoplasma concentration in the flowers.
� 2009 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Scientia Horticulturae
journa l homepage: www.e lsev ier .com/ locate /sc ihor t i
1. Introduction
Hydrangea macrophylla, a woody genus of Saxifragaceae, hasbeen a popular garden and greenhouse plant since 1789(McClintock, 1957). This plant is valued for its large brightlycolored flowers. Hydrangeas grow well at moderate temperatures(Bailey, 1914). During summer production at northern latitudes(428–598N), they grow vigorously under little or no shading(Littlere and Stromme, 1975); however, they also grow well undermoderate shading (20–50%) at southern latitudes (328–428N)(Shanks and Link, 1951). Hydrangeas prefer partial shading (30%)in areas and seasons with high light intensities (Weiler, 1980), anda definite light minimum is needed for adequate shoot develop-
* Corresponding author. Tel.: +81 75 753 6048; fax: +81 75 753 6068.
E-mail addresses: [email protected], [email protected]
(E. Kesumawati).
0304-4238/$ – see front matter � 2009 Elsevier B.V. All rights reserved.
doi:10.1016/j.scienta.2009.01.025
ment and flower bud differentiation (Ray, 1946; Struckmeyer,1950; Littlere and Stromme, 1975).
In 1996, Kanehira et al. reported on a Japanese hydrangeaphyllody (JHP) disease of hydrangeas that spreads whereverhydrangeas are grown in Japan, and determined that the diseaseis caused by the JHP-phytoplasma. In our previous study, weobserved that hydrangea flowers infected by the JHP-phyto-plasma become green but with variability (Kesumawati et al.,2006). We clarified that this variability in flower color is causedby the difference in the JHP-phytoplasma concentration in theplant.
Phytoplasmas are the smallest self-replicating life forms onearth and are characterized by the lack of a firm cell wall and by anextremely small size (Razin et al., 1998); their concentration ininfected plants is very low (Melamed et al., 2003). Phytoplasmaconcentration changes depending on seasonal environmentalconditions (Garcia-Chappa et al., 2003) and temperature(Kaminska et al., 2000).

E. Kesumawati et al. / Scientia Horticulturae 121 (2009) 199–205200
The presence of phytoplasmas during the phyllody (thedevelopment of floral parts into leafy structures) and virescence(the development of green flowers and the loss of normal flowerpigments) of hydrangeas has been reported in Europe (Muller, 1971;Welvaert et al., 1975), USA (Hearon et al., 1976), France (Cousin et al.,1986) and Japan (Kanehira et al., 1996; Sawayanagi et al., 1999).
In Japan, green hydrangea flowers have a potential market ascut flowers, dried flowers or potted plants and are sold at a highprice. An observation in the field showed that the inflorescences ofhydrangeas grown under the shade of neighboring plants arealways green. This is important information for inducing flowergreening. To date, no studies of the relationship between shadingand flower color in phytoplasma-infected hydrangeas have beenconducted. In this study, the effect of light intensity (varied byshading with cheesecloth) on flower greening in hydrangea plantsinfected by the JHP-phytoplasma was examined.
2. Materials and Methods
2.1. Plant materials and shade conditions
2.1.1. First-year experiment
The experiment was conducted at Kyoto University experi-mental farm, Japan (latitude, 358010N; longitude, 1358470E). Two‘Midori’ plants infected by the JHP-phytoplasma were used as stockplants for propagation. Thirty 10-cm-long shoot cuttings from eachof a single stock ‘Midori’ plant that was infected by the JHP-phytoplasma were propagated in August 2003. Twenty-fourcutting from each of three hydrangea cultivars ‘Libelle’, ‘Rosea’and ‘Madame E. Mouillere’ were individually grafted to a rootstockfrom another single ‘Midori’ plant that was also infected by theJHP-phytoplasma. All the shoots and grafted cuttings were plantedin trays containing vermiculite and placed under mist condition.
In September 2003, twenty-five ‘Midori’ plants and five plantseach of ‘Libelle’, ‘Rosea’ and ‘Madame E. Mouillere’ in the samegrowth stage were transferred and grown in 15-cm-diameter potsfilled with potting soil (Metro Mix 360 Scotts Co., Marysville, Ohio).Transplants were grown in a greenhouse under natural photo-period conditions at 35/17 8C (max/min). Each plant was fertilizedweekly with 1 L of a solution containing 1.25 g L�1 water-soluble5N–10P–5K fertilizer (Hanakoujou, Takeda, Tokyo, Japan). All theplants were irrigated with tap water on nonfertilization days.
In March 2004, the plants were divided and grown in the fieldand covered with black cheesecloth. Shading percentage wasexpressed as an increase in the number of sheets of cheeseclothused in shading the plants from sunlight. The shade conditions inthis experiment were full sunlight intensity (100% sunlightintensity), shading with one sheet of cheesecloth (70% sunlightintensity) and shading with two sheets of cheesecloth (49%sunlight intensity). Light intensity was measured by Luxmeter(UNITEST Digital Luxmeter 93408). There was no significantdifference in temperature among the shading conditions.
2.1.2. Second-year experiment
In March 2005, we repeated the treatment under full sunlightintensity and shading with two sheets of cheesecloth. ‘Midori’plants that were infected by JHP-phytoplasma from first yearexperiment were used in the second-year experiment. Six pottedplants each of the JHP-phytoplasma-infected ‘Midori’ plants weregrown under full sunlight intensity and shading with two sheets ofcheesecloth.
2.2. Preparation of the JHP phytoplasma non-infected plants
The infected ‘Midori’ plants were grown in a greenhouse. InApril 2002, to produce JHP-phytoplasma-free ‘Midori’ plants,
vegetative shoots were collected from these plants and sterilizedfor 15 min in 0.05% sodium hypochlorite and washed threetimes with sterile water. Uniform-size shoot tips with two leafprimordia were dissected. The excised shoot tips were culturedon a modified Knop medium containing Knop’s macroelementsat half strength (Knop, 1865) plus ferric-ethylenediaminete-traacetate (Fe-EDTA) (Murashige and Skoog, 1962), Ringe andNitsch’s microelements plus organic acids, and 20 g L�1 sucrose.This medium was solidified by adding 3 g L�1 gellan gum. Priorto autoclaving at 121 8C for 15 min, 0.5 ppm of plant pre-servative mixture (PPM) (Nacalai, Kyoto) was added to themedium to prevent bacterial proliferation; the medium’s pH wasadjusted to 5.8. After autoclaving, 5 mL of modified Knopmedium was solidified in a 6-cm-diameter petri dish. Shoot tipswere cultured at 20 � 3 8C under a 16-h photoperiod with a lightintensity of 70 mE m�2 s�1 provided by cool white fluorescent tubelights. After buds arose, they were subcultured on modifiedMurashige–Skoog (MS) medium containing MS macroelementsplus Fe-EDTA (Murashige and Skoog, 1962) and Ringe and Nitsch’smicroelements plus organic acid (Ringe and Nitsch, 1968) for shootelongation.
In March 2004, two-year-old plants from the JHP-phytoplasma-free ‘Midori’ buds that arose from the shoot tips were transplantedto 15-cm-diameter pots filled with potting soil (Metro Mix 360)and grown in a greenhouse under the same conditions as those forthe first experiment.
The JHP-phytoplasma detection test was performed on the budsthat arose from the shoot tips using PCR analysis.
On the other hand, twelve shoot cutting from each of threehydrangea cultivars ‘Libelle’, ‘Rosea’ and ‘Madame E. Mouillere’were taken from stock plants that were free from JHP-phytoplama.All of the shoots were planted in trays containing vermiculite,placed under mist conditions and used for comparison with graftedplants that were infected by JHP-phytoplasma.
2.3. Measurement of green-area ratio of individual flowers in
inflorescence
The green-area ratio of individual flowers in an inflorescencewas measured according to Kesumawati et al. (2006). A colordetector with fluorescent light (Taketombo Co., Ltd., Japan) wasused for taking a photograph of each flower separated from aninflorescence in a dark room. The photographs of individualflowers were taken using a digital camera at an Focus (F) of 5,6 anda shutter speed (S) of 1/25.
Green-area ratio of each flower from an inflorescence wasmeasured from a photograph using Adobe Photoshop with thefuzziness at 135. The green-area ratio of each flower was measuredby determining the number of green pixels divided by the totalnumber of pixels of the total flower area. From the green-area ratioof the flowers, we classified the flowers into four types: green-arearatio = 0, completely blue/pink/white flowers; 0 < green-arearatio < 0.4, blue/pink/white-green flowers; 0.4 2 green-arearatio < 0.8, greenish flower; 0.8 2 green-area ratio, completelygreen flower (Fig. 1).
2.4. Data collection and calculation
Flowers were collected from inflorescences of all ‘Midori’ plantsand grafted plants under different conditions. In the first-yearexperiment, six potted ‘Midori’ plants were used for data collectionand in this experiment used randomized block design. In thesecond-year experiment, all individual plants were used again fordata collection.
The data for the grafted plants ‘Libelle’, ‘Rosea’ and ‘Madame E.Mouillere’ were collected from one potted plant for each under

Fig. 1. Colors of individual flowers classified into four types by green-area ratio, determined by Adobe Photoshop (‘Midori’). (A) Green-area ratio = 0, completely blue/pink/
white flower, (B) 0 < green-area ratio < 0.4, blue/pink/white-green flower, (C) 0.4 2 green-area ratio < 0.8, greenish flower, and (D) 0.8 2 green-area ratio, completely green
flower.
E. Kesumawati et al. / Scientia Horticulturae 121 (2009) 199–205 201
different shade conditions. The JHP-phytoplasma-free ‘Midori’plants and the non-grafted ‘Libelle’, ‘Rosea’ and ‘Madame E.Mouillere’ plants were used for the comparison of flower color bythe JHP-phytoplasma-infected ‘Midori’ plants and JHP-phyto-plasma-grafted plants grown under full sunlight intensity.
2.5. DNA extractions and PCR conditions
Total DNA was extracted from the flowers of the JHP-phytoplasma-infected hydrangea ‘Midori’, Libelle’, ‘Rosea’ and‘Madame E. Mouillere’ plants under three conditions depending onthe types of flower; each type has three replicates.
Fresh flowers were frozen in liquid nitrogen, and ground intofine powder with a mortar and a pestle. The fine powder (0.07 g)was transferred to a 1.5-mL microtube to which 300 mL of DNAextraction buffer of the Mag Extractor kit (Toyobo, Osaka, Japan)was added. The extracted DNA was dried on a clean benchaccording to the manufacturer’s instructions of the Mag Extractorkit, and then resuspended in 100 mL of TE buffer (10 mM Tris–HCland 1 mM EDTA (pH 8.0)).
Fig. 2. Relationship between light intensity and flower color in the JHP-phytoplasma-in
‘Madame E. Mouillere’. (A, E, I, and M) Non-infected hydrangea cultivars grown under fu
infected hydrangea grown under full sunlight intensity (100% sunlight intensity). (C, G, K,
cheesecloth (49% sunlight intensity). (D, H, L, and P) The JHP-phytoplasma-infected h
intensity). The pictures were taken 2 weeks before harvest.
The PCR mixture was prepared as follows: 1 mL of templateDNA, 0.1 mL of each primer, 0.1 mL of KOD dash polymerase(TOYOBO, OSAKA, Japan), 1 mL of KOD dash buffer, 1 mL each ofdNTPs (2.0 mM), 0.1 mL each of primers (20 mM) for JHP-phytoplasma detection, and 6.7 mL of distilled water.
A pair of primers (JHPF1 (forward): 50-GTGTAGCCGGGCTGA-GAGGTCA-30 and JHPR1 (reverse): 50-TCCAACTCTAGCTAAA-CAGTTTCTG-30) was used to amplify the 16S rDNA of the JHPphytoplasma (Sawayanagi et al., 1999). The reaction mixture wasoverlaid with one drop of mineral oil, and PCR amplification wasperformed using a DNA thermal cycler (PerkinElmer, USA). Theprogram for PCR amplification was as follows: denaturation for3 min at 98 8C followed by 30 cycles (each cycle consisting of 30 sof denaturation at 98 8C, 5 s of annealing at 62 8C, and 30 s ofextension at 74 8C) and a final extension step at 74 8C for 5 min. A5-mL aliquot from each PCR analysis sample was separated on a1.5% agarose gel and visualized with ethidium bromide under anultraviolet transilluminator. The agarose gel contained TBE buffer(0.27 g of Tris, 0.14 g of boric acids and 0.1% 0.5 M EDTA (pH 8.0) in50 mL of distillated water).
fected hydrangea cultivars. (A–D) ‘Midori’; (E–H) ‘Libelle’; (I–L) ‘Rosea’; and (M–P)
ll sunlight intensity (100% sunlight intensity). (B, F, J, and N) The JHP-phytoplasma-
and O) JHP-phytoplasma-infected hydrangea grown under shading with one sheet of
ydrangea grown under shading with two sheets of cheesecloth (70% full sunlight
E. Kesumawati et al. / Scientia Horticulturae 121 (2009) 199–205202
3. Results
3.1. Effect of shading condition on flower color of the JHP-
phytoplasma-infected hydrangea cultivar ‘Midori’
By a visual inspection of the field, a marked difference in flowercolor was observed in ‘Midori’ plants grown at different lightintensities. In the first-year experiment, the JHP-phytoplasma-free‘Midori’ plants propagated by shoot tip culture had showed blueflowers under full sunlight intensity (100% light intensity) (Fig. 2A).Under the same conditions, when ‘Midori’ plants were infectedwith the JHP phytoplasma, all the flowers in an inflorescence werepink (Fig. 2B). Under shading with one sheet of cheesecloth, theflower was mixed pink and green (Fig. 2C); under shading with twosheets of cheesecloth, the flowers were completely green (Fig. 2D).
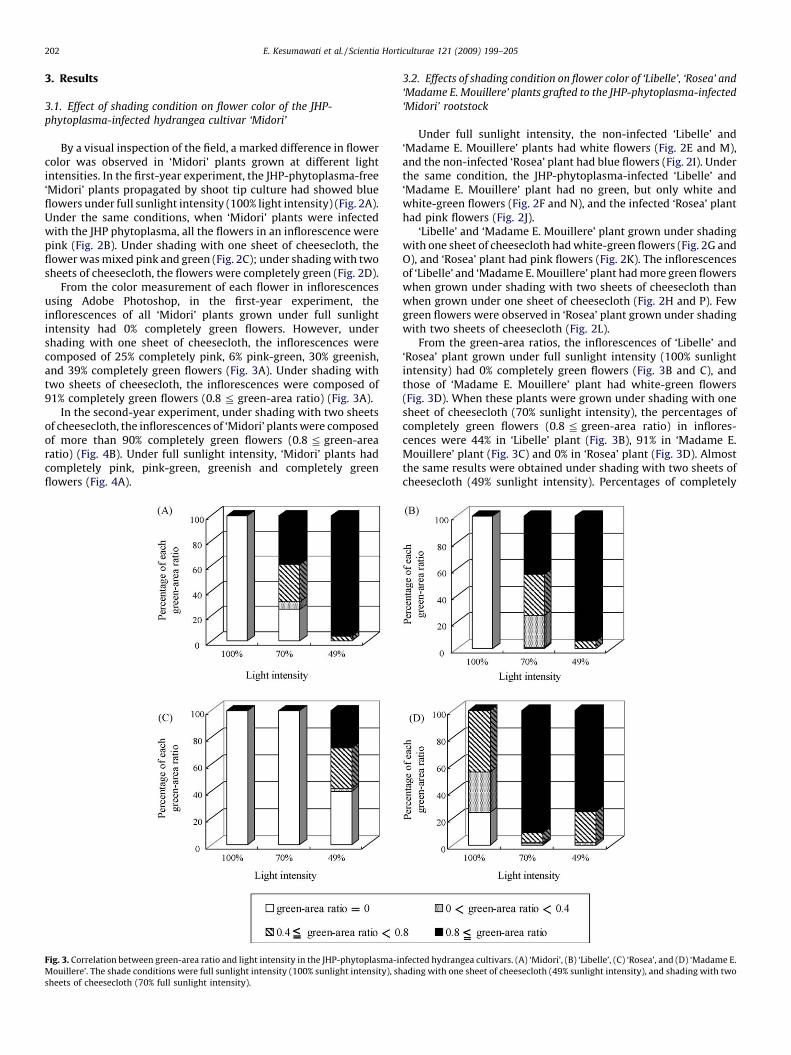
From the color measurement of each flower in inflorescencesusing Adobe Photoshop, in the first-year experiment, theinflorescences of all ‘Midori’ plants grown under full sunlightintensity had 0% completely green flowers. However, undershading with one sheet of cheesecloth, the inflorescences werecomposed of 25% completely pink, 6% pink-green, 30% greenish,and 39% completely green flowers (Fig. 3A). Under shading withtwo sheets of cheesecloth, the inflorescences were composed of91% completely green flowers (0.8 2 green-area ratio) (Fig. 3A).
In the second-year experiment, under shading with two sheetsof cheesecloth, the inflorescences of ‘Midori’ plants were composedof more than 90% completely green flowers (0.8 2 green-arearatio) (Fig. 4B). Under full sunlight intensity, ‘Midori’ plants hadcompletely pink, pink-green, greenish and completely greenflowers (Fig. 4A).
Fig. 3. Correlation between green-area ratio and light intensity in the JHP-phytoplasma-i
Mouillere’. The shade conditions were full sunlight intensity (100% sunlight intensity), sh
sheets of cheesecloth (70% full sunlight intensity).
3.2. Effects of shading condition on flower color of ‘Libelle’, ‘Rosea’ and
‘Madame E. Mouillere’ plants grafted to the JHP-phytoplasma-infected
‘Midori’ rootstock
Under full sunlight intensity, the non-infected ‘Libelle’ and‘Madame E. Mouillere’ plants had white flowers (Fig. 2E and M),and the non-infected ‘Rosea’ plant had blue flowers (Fig. 2I). Underthe same condition, the JHP-phytoplasma-infected ‘Libelle’ and‘Madame E. Mouillere’ plant had no green, but only white andwhite-green flowers (Fig. 2F and N), and the infected ‘Rosea’ planthad pink flowers (Fig. 2J).
‘Libelle’ and ‘Madame E. Mouillere’ plant grown under shadingwith one sheet of cheesecloth had white-green flowers (Fig. 2G andO), and ‘Rosea’ plant had pink flowers (Fig. 2K). The inflorescencesof ‘Libelle’ and ‘Madame E. Mouillere’ plant had more green flowerswhen grown under shading with two sheets of cheesecloth thanwhen grown under one sheet of cheesecloth (Fig. 2H and P). Fewgreen flowers were observed in ‘Rosea’ plant grown under shadingwith two sheets of cheesecloth (Fig. 2L).
From the green-area ratios, the inflorescences of ‘Libelle’ and‘Rosea’ plant grown under full sunlight intensity (100% sunlightintensity) had 0% completely green flowers (Fig. 3B and C), andthose of ‘Madame E. Mouillere’ plant had white-green flowers(Fig. 3D). When these plants were grown under shading with onesheet of cheesecloth (70% sunlight intensity), the percentages ofcompletely green flowers (0.8 2 green-area ratio) in inflores-cences were 44% in ‘Libelle’ plant (Fig. 3B), 91% in ‘Madame E.Mouillere’ plant (Fig. 3C) and 0% in ‘Rosea’ plant (Fig. 3D). Almostthe same results were obtained under shading with two sheets ofcheesecloth (49% sunlight intensity). Percentages of completely
nfected hydrangea cultivars. (A) ‘Midori’, (B) ‘Libelle’, (C) ‘Rosea’, and (D) ‘Madame E.
ading with one sheet of cheesecloth (49% sunlight intensity), and shading with two

Fig. 4. Effects of light intensity and green-area ratio on green-flower stability in the
JHP-phytoplasma-infected ‘Midori’ plants. (A) Under full sunlight intensity (100%
sunlight intensity). (B) Under shading with two sheets of cheesecloth (49% sunlight
intensity).
E. Kesumawati et al. / Scientia Horticulturae 121 (2009) 199–205 203
green flowers (0.8 2 green-area ratio) in inflorescences were 94%in ‘Libelle’ plant (Fig. 3B), 75% in ‘Madame E. Mouillere’ plant(Fig. 3C), and 28% in ‘Rosea’ plant (Fig. 3D).
3.3. Determination of the JH-phytoplasma by PCR analysis using
flowers with different green-area ratios
The JHP-phytoplasma infection in the hydrangea flowers withvarious green-area ratios as a result of the graft inoculation of theJHP-phytoplasma to ‘Midori’, ‘Libelle’, ‘Rosea’, and ‘Madame E.Mouillere’ plant were determined. The JHP-phytoplasma was notdetected by PCR analysis in completely blue/pink/white flowerswith a green-area ratio = 0 (Fig. 5). On the other hand, the JHP-phytoplasma was detected in the other flowers with differentgreen-area ratios (Fig. 5). The same result was obtained from thesecond-year experiment using ‘Midori’ plants (data not shown).
4. Discussion
The JHP-phytoplasma affects of the color change of hydrangeaflowers to green (Sawayanagi et al., 1999), but with variability(Kesumawati et al., 2006). To clarify the mechanism underlyingthis variability, JHP-phytoplasma concentration in the green, blue/pink and blue/pink-green flowers of JHP-phytoplasma-infected‘Midori’ hydrangea plants was measured by semiquantitative PCRanalysis (Kesumawati et al., 2006). Kesumawati et al. (2006) foundthat the JHP-phytoplasma concentration in green flowers was 16-fold that in blue/pink-green flowers and concluded that main-
taining a high JHP-phytoplasma concentration in flowers isimportant for flower greening in hydrangea.
This study showed that shading conditions (70% and 49%sunlight intensities) enhance the rate of greening of inflorescencesin JHP-phytoplasma-infected hydrangea cultivars. However, plantsgrown under full sunlight intensity (100% sunlight intensity) hadpink or white flowers. PCR analysis showed that the JHP-phytoplasma could not be detected in any of the hydrangeacultivars with completely pink or white flowers (green-arearatio = 0), but was identified in those with flowers of various green-area ratios (Fig. 5). This result suggests that the JHP-phytoplasmacould not proliferate well in hydrangea plants at a high lightintensity. It also indicates that shading affects the JHP-phyto-plasma concentration in an inflorescence and enhances flowergreening. Hearon et al. (1976) reported that, in infected hydrangeaplants, flowers vary from completely green to mixed color, withpink and green flowers, with and without foliar symptoms.Kesumawati et al. (2006) reported that PCR analysis cannot be usedin determining the JHP-phytoplasma concentration in blue/pinkflowers of infected hydrangea plants and that the JHP-phyto-plasma concentrations in the flowers are very low.
Under 100% sunlight intensity, the JHP-phytoplasma-free‘Midori’ plants that regenerated by the shoot tip culture had blueflowers; PCR analysis shown that the plants were free of the JHP-phytoplasma (data not shown). These results indicate that tissueculture can be successfully applied to the elimination of the JHP-phytoplasma from hydrangea plants. By shoot tip (Sear andKlomparens, 1989) or callus culture (Moellers and Sarkar, 1989),symptom-free plants have been obtained and such plants havebeen proved to be phytoplasma-free. On the other hand, under thesame conditions, ‘Midori’ plants infected with the JHP-phyto-plasma had pink flower in the first year. This suggests that the JHP-phytoplasma cannot proliferate when hydrangeas are grown underhigh light intensity. Bailey and Hammer (1989) reported thatenvironmental conditions have an affect on hydrangea leaf andinflorescence malformations, for example phytoplasmas can beeliminated from plants by high-temperature exposure (Gong,1990). High-temperature exposure creates disadvantageous con-ditions for pathogen replication, non-replacement of degradedpathogens, and an altered balance between synthesis anddegradation causing the elimination of pathogens (Kassanis, 1954).
In the second-year experiment, the results were similar to thosein the first-year experiment. However, under full sunlightintensity, ‘Midori’ plants showed variation in flower color, bearingcompletely pink, pink-green, greenish and completely greenflowers (Fig. 4A). These results might be correlated with theacclimatization of the plants to high light intensity. From theresults in the second-year experiment, ‘Midori’ plants wereassumed to be well adapted to full sunlight intensity and in thesewell adapted plants, the JHP-phytoplasma could proliferate well,resulting in flower greening (Fig. 4A).
The green-area ratios of ‘Midori’ plants grown under shadingwith two sheets of cheesecloth (49% light intensity) showed thatthe inflorescences were composed of more than 90% completelygreen flower (0.8 2 green-area ratio) in the first and second-yearexperiment (Figs. 3A and 4B). This suggests that shading conditionsfavorable for hydrangea growth are also favorable for JHP-phytoplasma proliferation. These results are similar to thoseobtained by Nishigawa et al. (2002) who observed that becausephytoplasmas are so dependent on their hosts, any mechanism forincreasing their biological diversity could be of great significanceto their pathogenicity. Oshima et al. (2004) reported that becausephytoplasmas lack most of the genes necessary to synthesizenucleotides, they absorb them from their environment. Razin et al.(1998) reported that phytoplasmas are phloem-restricted bacteria.They are the first living organisms to be discovered to lack the eight

Fig. 5. Determination of the JHP-phytoplasma concentrations from hydrangea flowers with various green-area ratios by PCR analysis as 370 bp band. Line 1: M. Molecular
marker (wX 174/HincII digest, Toyobo), (1–4) ‘Midori’ and (5–8) ‘Libelle’. Line 2: M. Molecular marker (wX 174/HincII digest, Toyobo), (1–4) ‘Rosea’ and (5–8) ‘Madame E.
Mouillere’. Four types of flowers depending on green-area ratio are shown: (1, 5) green-area ratio = 0; (2, 6) 0 < green-area ratio < 0.4; (3, 7) 0.4 2 green-area ratio < 0.8; (4,
8) 0.8 2 green-area ratio. The flowers of the hydrangea cultivars used for PCR analysis are shown: Line 1: (1–4) ‘Midori’ and (5-8) ‘Libelle’. Line 2: (1–4) ‘Rosea’ and (5–8)
‘Madame E. Mouillere’.
E. Kesumawati et al. / Scientia Horticulturae 121 (2009) 199–205204
adenosine triphosphate (ATP)-synthase subunits (Mushegian andKoonin, 1996). Note that ATP synthesis is dependent on theglycolysis pathway (Santana et al., 1994). Recently, Oshima et al.(2004) have clarified that phytoplasmas lack the pentosephosphate cycle and ATP-synthase subunits, which are consideredto be essential to life. They explained this to be a result ofphytoplasmas living as the intracellular parasites in nutrient-richenvironments.
In this experiment, ‘Midori’ plants that arose from shoot tipshad blue flowers under 100% sunlight intensity, and the sameplants but infected by the JHP-phytoplasma had pink flowers(Fig. 2A and B). Under the same conditions, the non-infected‘Rosea’ plant had blue flowers; after the infection by the JHP-phytoplasma, it had pink flowers (Fig. 2I and J). The mechanismsunderlying this phenomenon are as yet unknown, but could be thevery low concentrations of the JHP-phytoplasma in the hydrangeaplants or of other bacteria or viruses infecting the plants.
In conclusion, this is to the best of our knowledge the first reporton the relationship between light intensity (varied by shading withcheesecloth) and the growth of the JHP-phytoplasma-infectedhydrangea plants. Here we showed that the JHP-phytoplasmacould proliferate better under shaded condition, than under fullsunlight intensity (100% sunlight intensity) and that is enhancesflower greening in hydrangea. The effect of light intensity on flowergreening in hydrangeas depends on cultivars.
References
Bailey, L.H., 1914. The Standard Cyclopedia of Horticulture, 2. Macmillan, New York,1619–1623.
Bailey, D.A., Hammer, P.A., 1989. Stimulation of hydrangea distortion throughenvironmental manipulation. J. Am. Soc. Horticult. Sci. 114, 411–416.
Cousin, M.T., Sharma, A.K., Rousseau, J., Poitevin, J.P., Savore, A., 1986. Hydrangeavirescence. I. Description of the disease and its transmission to the differentialhost plant Catharanthus roseous by Cuscuta subinclusa. Agronomie 6, 3–28.
Garcia-Chappa, M., Medina, V., Viruel, M.A., Lavina, A., Batle, A., 2003. Seasonaldetection of pear decline phytoplasma by nested-PCR in different pear cultivars.Plant Pathol. 52, 513–520.
Gong, Z.X., 1990. Prevention and cure of mycoplasma-like organism diseases inplants. In: Gong, Z.X., Chen, J.Y., Shen, J.Y. (Eds.), A Collection of IllustrativePlates of Plant Mycoplasma-like Organisms in China. Science Press, Beijing, pp.38–48.
Hearon, S.S., Lawson, R.H., Smith, F.F., McKenzie, J.T., Rosen, J., 1976. Morphology offilamentous forms of a mycoplasma-like organism associated with Hydrangeavirescence. Phytopathology 66, 608–616.
Kanehira, T., Horikoshi, N., Yamakita, Y., Shinohara, M., 1996. Occurrence ofhydrangea phyllody in Japan and detection of the causal phytoplasma. Ann.Phytopathol. Soc. Jpn. 62, 26–28.
Kaminska, M., Gabryszewska, E., Korbin, M., 2000. Phytoplasma detection in tissueculture Gladioulus plants grown under various conditions. Acta Soc. Bot. Pol. 69,197–200.
Kassanis, B., 1954. Heat therapy of virus infected plants. Ann. Appl. Biol. 41,470–474.
Kesumawati, E., Kimata, T., Uemachi, T., Hosokawa, M., Yazawa, S., 2006. Correlationof phytoplasma concentration in Hydrangea macrophylla with green-floweringstability. Sci. Horticult. 108, 74–78.
Knop, W., 1865. Quantitative untersuchungen uber die ernahrungsprozesse derpflanzen. Landwirtsch Vesrs Stn. 7, 93–107.
Littlere, B., Stromme, E., 1975. The influence of temperature, daylength and lightintensity on flowering in Hydrangea macrophylla (Thunb.) Serr. Acta Horticult.51, 285–298.
Melamed, S., Tanne, E., Ben-Haim, R., Edelbaum, O., Yogev, D., Sela, I., 2003.Identification and characterization of phytoplasmal genes, employing a novelmethod of isolating phytoplasmal genomic DNA. Am. Soc. Microbiol. 185,6513–6521.
Moellers, C., Sarkar, S., 1989. Regeneration of healthy plants from Catharantus roseusinfected with mycoplasma-like organism through culture. Plant Sci. 60, 83–90.
Murashige, T., Skoog, F., 1962. A revised medium for rapid growth and bioassay withtobacco tissue culture. Physiol. Plant. 15, 473–497.

E. Kesumawati et al. / Scientia Horticulturae 121 (2009) 199–205 205
Muller, H.M., 1971. Elektronenmikroskopischer nachweis von mycoplasmaahnli-chen organismen inphloem von Hydrangea macrophylla. Zentralkl. Bacteriol.Parasitenkd. Infections-krankh. Hyg. Naturwiss 126, 564–565.
Mushegian, A.R., Koonin, E.V., 1996. A minimal gene set for cellular life derived bycomparison of complete bacterial genomes. Proc. Natl. Acad. Sci. U.S.A. 93,10268–10273.
McClintock, E., 1957. A monograph of the genus hydrangea. Proc. Calif. Acad. Sci. 29,147–256.
Nishigawa, H., Oshima, K., Kakizawa, S., Jung, H., Kuboyama, T., Miyata, S., Ugaki, M.,Namba, S., 2002. Evident of intermolecular recombination between extrachro-mosomal DNAs in phytoplasmas: a trigger for the biological diversity ofphytoplasma? Microbiology 148, 1389–1396.
Oshima, K., Kakizawa, S., Nishigawa, H., Jung, H.Y., Wei, W., Suzuki, S., Arashida, R.,Nakata, D., Miyata, S., Ugaki, M., Namba, S., 2004. Reductive evolution suggestedfrom the complete genome sequence of a plant-pathogenic phytoplasma. Nat.Genet. 36, 27–29.
Razin, S., Yogev, D., Naot, Y., 1998. Molecular biology and pathogenicity of myco-plasmas. Microbiol. Mol. Biol. Rev. 62, 1094–1156.
Ray, S., 1946. Reduction of blindness in hydrangeas. Proc. Am. Soc. Horticult. Sci. 47,501–502.
Ringe, F., Nitsch, J.P., 1968. Conditions leading to flower formation of excisedBegonia fragments cultured in vitro. Plant Cell Physiol. 9, 639–652.
Santana, M., Ionescu, M.S., Vestes, A., Longin, R., Kunst, F., Danchin, A., Glaser, P.,1994. Bacillus subtilis F0F1 ATPase: DNA sequence of the atp operon andcharacterization of atp mutants. J. Bacteriol. 176, 6802–6811.
Sawayanagi, T., Horikoshi, N., Kanehira, T., Shinohara, M., Bertaccini, A., Cousin, M.T.,Hiruki, C., Namba, S., 1999. ‘Candidatus phytoplasma japonicum’, a new phy-toplasma taxon associated with Japanese hydrangea phyllody. Int. J. Syst.Bacteriol. 49, 1275–1285.
Sear,B.B., Klomparens, K.L.,1989. Leaf tip culturesof the eveningprimrose allow stableaseptic culture of mycoplasma-like organism. Can. J. Plant Pathol. 11, 343–348.
Shanks, J.B., Link, C.B., 1951. Some studies on the effects of temperature andphotoperiod on growth and flower formation in hydrangea. Proc. Am. Soc.Hortic. Sci. 58, 357–366.
Struckmeyer, B.E., 1950. Blossom bud induction and differentiation in hydrangea.Proc. Am. Soc. Hortic. Sci. 56, 410–414.
Welvaert, W., Samyn, G., Lagasse, A., 1975. Research on the virescence symptoms ofHydrangea macrophylla (Thunb.). Phytopathology 83, 152–158.
Weiler, T.C., 1980. Hydrangeas. In: Larson, R.A. (Ed.), Introduction to Floriculture.Academic Press, New York, pp. 354–375.





























