Embed Size (px)
Citation preview

Cell Tissue Res (1981) 219:93-109Cell and Tissue
Research
@ Springer-Verlag 1981
Fine structure and metabolismof multiply innervated fast muscle fibres
in teleost fish
Ian A. Johnston and Thomas W. Moon *
Department of Physiology, University ofSt. Andrews, St. Andrews, Fife, Scotland, Great Britain
Summary. Both the fast and slow muscle fibres of advanced teleost fish aremultiply innervated. The fraction of slow-fibre volume occupied bymitochondria is 31.3%,25.5% and 24.6%, respectively, for the myotomalmuscles of brook trout (Salvelinusfontinalis), crucian carp ( Carassius carassius),and plaice (Pleuronectes platessa), respectively. The corresponding figures forthe fast muscles of these species are 9.3 %, 4.6% and 2.0 %, respectively.Cytochrome-oxidase and citrate-synthetase activities in the fast muscles of 9species of teleost range from 0.20-0.93 ~moles substrate utilised, 9 wet weightmuscle-1 min-1 (at 15° C) or around 4-17% of that of the corresponding slowfibres. Ultrastructural analyses reveal a marked heterogeneity within the fast-fibre population. For example, the fraction of fibres with < 1% or > 10%mitochondria is 0,4,42% and 36,12 and 0%, respectively, for trout, carp andplaice. In general, small fibres ( < 500 ~m2) have the highest and large fibres( > 1,500 ~m2) the lowest mitochondrial densities. The complexity ofmitochon-drial cristae is reduced in fast compared to slow fibres.
Hexokinase activities range from 0.4-2.5 in slow and from 0.08-0.7 ~moles,g wet weight- I min-1 in fast muscles, indicating a wide variation in their
capacity for aerobic glucose utilisation. Phosphofructokinase activities are 1.2to 3.6 times higher in fast than slow muscles indicating a greater glycolyticpotential. Lactate dehydrogenase activities are not correlated with either thepredicted anaerobic scopes for activity or the anoxic tolerances of the species
studied.The results indicate a considerable variation in the aerobic capacities and
principal fuels supporting activity among the fast muscles of different species.Brook trout and crucian carp are known to recruit fast fibres at low swimmingspeeds. For these species the aerobic potential of the fast muscle is probablysufficient to meet the energy requirements of slow swimming.
Send offprint requests to: Ian A. Johnston, Department of Physiology, University of St. Andrews, St.
Andrews, Fife, Scotland, KY169TS, Great Britain* Permanent address: Department of Biology, University of Ottawa, Ottawa, ' Ontario, Canada
K1N9B4
0302-766X/81/0219/0093/$03.40

94 .A. Johnston and T.W. Moon
Key words: Quantitative cytology -Fish muscle -Muscle metabolism -Muscleultrastructure -Teleosts
In all fishes fast fibres comprise between 70-100 % of the trunk musculature (seeGreer-Walker andPuI11975). The preponderance of fast fibre types is a reflection ofthe power requirements of aquatic locomotion [ ""' power cx velocity3] (Webb 1975).Thus small increments in performance at high speed require the recruitment ofincreasing numbers of fast motor units.
The dogfish (Scyilorhinus cannicula), in common with other elasmobranchs, hasfast fibres with single motor endplates (Bone 1964). In this species the fast motorsystem is reserved for butst locomotory activity (Bone 1966). Fast-muscle in sharksis pure white in colour, poorly vascularised and mitochondria usually occupy lessthan 1% of fibre volume (Kryvi 1977; Totland et al. 1980). It is likely that thesefibres are entirely dependent on anaerobic glycogenolysis for energy (Bone 1978a).Exhaustion in dogfish occurs after only 1-2min of vigorous swimming and co-incides with the depletion of glycogen in fast muscles (Bone 1966).
In contrast to the elasmobranchs, fast muscle fibres in most teleosts are multiplyinnervated as are the slow fibres of fish, amphibia, birds and some extra-occularmuscles in mammals (Barets 1961; Bone 1964; Hesse 1970). Typically each fibrereceives a dense network of innervation with numerous endplates, often derivedfrom more than one motor axon (Barets 1961 ;'Bone 1964,1970). For example, fastfibres in the scorpeaniform fish Myoxocephalus scorpius have up to 22 endplateseach ofwhich is probably derived from a separate axon (Hudson 1969). This type offast muscle innervation differs from that of all other vertebrates includingchondrosteans, elasmobranchs and certain primitive teleost groups (e.g., gon-orythichiformes, clupeiformes, anguilliformes) all of which have single motorendplates (Bone 1964, 1970).
Electromyographical studies have shown that fast fibres with multipleinnervation are recruited at both low-sustainable and burst-swimming speeds (seeBone 1978a; Johnston 1981a, b). For example, the threshold speed for recruitmentof fast fibres is only 0.5-2.1 bodylength S-l in carp (Johnston et al. 1977; Bone et al.1978),0.8-1.9 bodylength S-l in saithe (Johnston and Moon 1980a) and 0.5-1.3bodylength S-l in rainbow trout (Hudson 1973; Bone et al. 1978).
Little is known about the metabolism and energy supply of fast muscles inteleosts at low swimming speeds. However, it seems likely that the metabolism ofthese fibres may differ from the anaerobic pattern characteristic of elasmobranchs.The present study investigates both the fine structure and energy metabolism ofmuscles from teleost species with a range of different swimming behaviours andactivity levels.
Materials and methods
Fish
Marine fish were obtained from the Firth of Forth, Scotland during April 1980. They were used forexperiments the same day as capture. Freshwater fish were supplied by local fish farms and maintainedat 8-10° C for up to a week prior to sacrifice. Details of the species of fish used and their size ranges aregiven in Table 1. All fish were killed by a sharp blow to the head and transsection of the spinal cord.

Structure and metabolism of fish muscle
Table I. Lengths and weights of species used in th,ese investigations
Length (cm) Weight (g)Common name Scientific name Habitat
Freshwater rivers 18.2:t 0.3Freshwater lakes 17.9:t0.9Freshwater lakes 16.9:t 0.5Marine 32.9:t 1.3Marine 44.4:t 2.7Marine/Esturine 29.5:t 0.8Marine 27.2:t 1.3Marine 24.8:t 2.1Marine 24.2:t 0.7
59.2:95.8:54.7.
316.7.881.4.307.5:223.0.165.9.145.7.
Brook troutCrucian carpTench
WhitingCodFlounderLemon solePlaiceDab
Salvelinus fontinalis (Mitchil/)Carassius carassius L.Tinca tinca L.Merlangius merlangus L.Gadus morhua L.Platichthys flesus L.Microstomus kitt L.Pleuronectes platessa L.Limanda limanda L.
Ultrastructural studies
Muscle samples were initially fixed in situ for 1 h by injecting 3% glutaraldehyde, 0.15 M phosphatebuffer pH 7.4 into the posterior third of the dorsal myotomal muscle. During this time fish were kept onice. In the species studied different types of fibre are anatomically separated (see Bone 1978a). Smallbundles of fast and slow fibres can be easily dissected, free from contamination, using a binocularmicroscope. Samples of fast muscle were dissected at random from all depths within the body. Fibrebundles were held at their resting length by pinning to cork strips. Fixation was continued for a further2-24h at 4° C in 3% glutaraldehyde, 0.15 M phosphate buffer pH 7.4. Tissue samples were post-fixed in1% osmium tetroxide in 0.1 M phosphate pH 7.4, dehydrated in a series of alcohols up to 100% andembedded in araldite resin. Ultrathin sections were cut on a Reichart OM U2 ultramicrotome anddouble stained with uranyl acetate and lead citrate. Sections were examined with a Phillips 301 electronmicroscope. Orientation of muscle fibres in embedded material was ascertained from examination of1 ~m sections stained with either toluidine blue or p-phenylene diamine in 1:1 isopropanol:methanol
(Hollander and Vaaland 1968).
Morphometric methods
Total cross-sectional areas of fibres were determined by tracing outlines from electron micrographs( x 4,940), using a summagraphics digitiser in conjunction with a mini-computer (Walesby and Johnston1980). Quantitative analyses of electron micrographs (magnification 7,000 to 15,000 x) were carried outusing a point-grid method (WeibeI1969). Good agreement was found between the stereological methodsof Weibel and direct estimates of cell component fractional volumes using the digitiser and mini-computer. The fraction of total fibre volume occupied by mitochondria ( %) (MF) was determined for 50
fast- and 50 slow-muscle fibres from brook trout, crucian carp and plaice.
Enzyme assays
Superficial red fibers were rapidly dissected from both sides of the entire length of the body. White-muscle samples of about 2 9 were dissected from the dorsal third of the trunk musculature. Muscle was
minced with scissors and homogenized at 00 C with an Ilado-X10 homogeniser (I.C.A. GmbH,Dottinger, W. Germany) for three periods of 25 s with cooling in 5-8 vols of 50 mM Tris-HCI 5 mM
EDTA, 2mM Mg C12, 1mM dithiothreitol pH7.5.Homogenates were centrifuged at 600 g for 20 min and filtered through glass wool. Enzyme
activities were determined in the supernatant and expressed in terms of ~oles substrate utilised per gdry weight muscle, min- 1. Measurements of enzyme activity were performed at 15° C with appropriatecontrols (usually substrate deletion). Concentrations of substrates and co-ions, and pH were establishedto give conditions for measurements of maximal enzyme activities on the basis of preliminaryexperiments with trout, carp and plaice. Assay procedures for the individual enzymes were as follows:
Citrate synthetase (CS)
Citrate synthetase was assayed in a medium of 100mM Tris-HCI, 0.5mM oxaloacetic acid, 0.3mMA""tvl r"A () 1 m M 5-51-rlit.hinhi,;-2-nit.robenzoic in 40 mM ohosohate. oH 8.0. The reaction was
: 3.1
: 10.7
: 7.5
: 54.3
: 147.0
: 45.0
: 25.5
: 17.2
: 14.1
96 A. Johnston and T.W. Moon
started by addition of oxaloacetic acid and the increase in extinction at 412 nm wavelength monitored.Enzyme activity was calculated using EmM = 13.6.
Cytochrome oxidase (CO )
Cytochrome oxidase activity was assayed by following the oxidation of reduced cytochrome C in 50 mMphosphate buffer pH7.6 at 550nm. Enzyme activity was calculated using EmM (red-ox) = 19.1.
Hexokinase ( H K)
Hexokinase activity was assayed using anA TP-regenerating system in a medium containing 85 mM Tris-HCI pH 7.5, 8mM MgCI2 0.8mM EDTA, lmM glucose, 2.5mM ATP, 0.4mM NADP, 10mMphosphoryl creatine, 100 I1g creatine phosphokinase, and 100 I1g glucose-6-phosphate dehydrogenase.Control assays contained the ~bove medium with glucose omitted.
Lactate dehydrogenase (LDH)
Lactate-dehydrogenase activity was assayed spectrophotometrically in a medium of 50 mM phosphatebuffer pH 7.5, 1 mM sodium pyruvate and 0.27 mM NADH.
Phosphofructokinase ( P F K)
Phosphofructokinase activity was assayed spectrophotometrically in a medium of 50 mM Tris-HClpH7.4, 4.5mM Fructose-6-phosphate, 3mMATP, 0.2mMAMP25mM KC!, 6mM MgCl2, 0.15mMNADH and excess alsolase, triose phosphate isomerase and glycerolpho~phate dehydrogenase.
Statistical analyses
Results were compared using a Student's t-test.
Abbreviations; M F Fraction of total muscle fibre volume occupied by mitochondria ( %); CS Citratesynthetase; CO Cytochrome oxidase; H K Hexokinase; LDH Lactate dehydrogenase; p F K
Phosphofructokinase
Results
Enzyme activity profiles
Enzyme activities of slow and fast muscles are shown in Tables 2 and 3 respectively.Based on CS and CO activities the aerobic capacities of the slow muscles studiedwould appear to be broadly similar. The ratio of the activities of these two enzymesin fast: slow muscles therefore allow a crude assessment of the relative aerobic
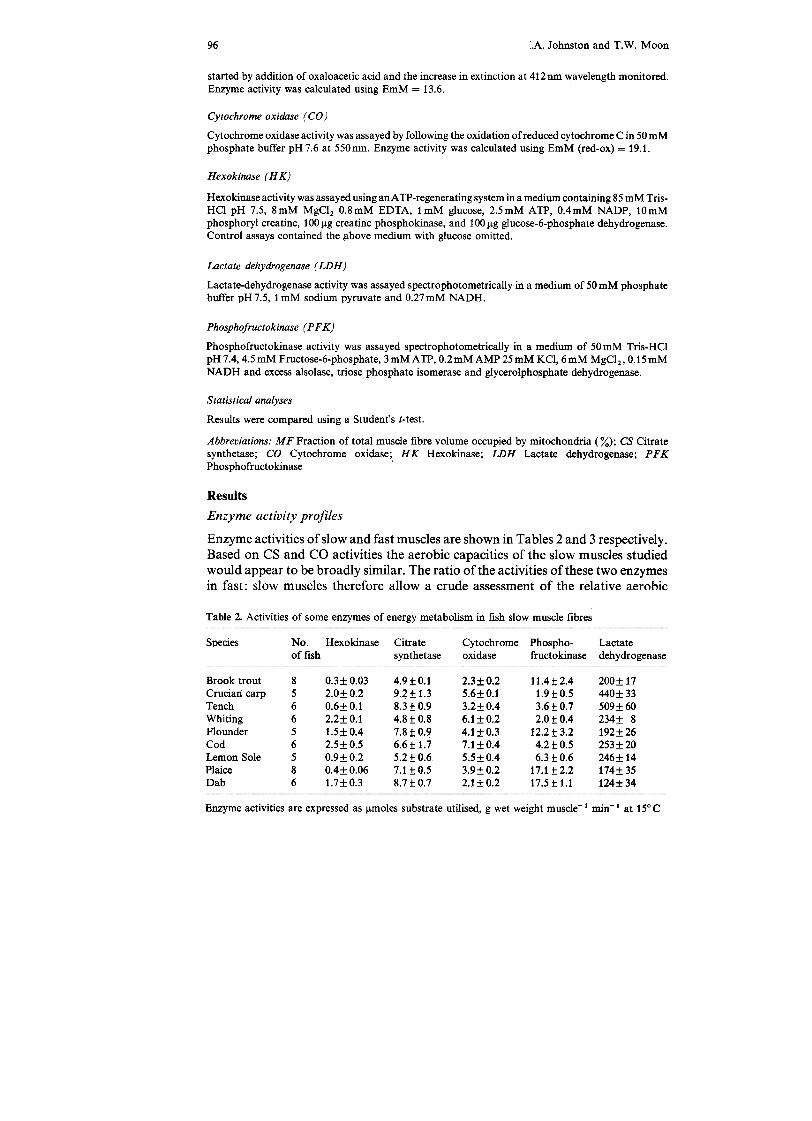
Table 2. Activities of some enzymes of energy metabolism in fish slow muscle fibres
Species No. Hexokinase
of fish
Citrate
synthetase
Cytochrome Phospho- Lactateoxidase fructokinase dehydrogenase
8
~6656586
0.3:t 0.032.0:t 0.20.6:t 0.12.2:t 0.11.5:t0.42.5:t 0.50.9:t 0.20.4:t 0.061.7:t0.3
4.99.28.34.87.86.65.27.18.7
2.3:t 0.2
S.6:t 0.1
3.2:t 0.4
6.1 :t 0.2
4.1:t0.3
7.1 :t 0.4
S.S:t 0.4
3.9:t 0.2
2.1::1: 0.2
11.41.93.62.0
12.24.26.3
17.117.5
200:t17440:t 33509:t 60234:t 8192:t 26253:t 20246:t 14174:t 35124::!: 34
Brook trout
CrucialicarpTench
WhitingFlounderCodLemon SolePlaiceDab
Enzyme activities are expressed as I.lmoles substrate utilised, 9 wet weight I1iUSCle-l min- 1 at 15° C
to.1t 1.3to.9to.8to.9t 1.7to.6to.5to.7
:!:2.4:!:0.5:!:0.7:!:0.4:!:3.2:!:0.5:!:0.6:!:2.2t 1.1
~tructure and metaboJi~m of fish muscle 97
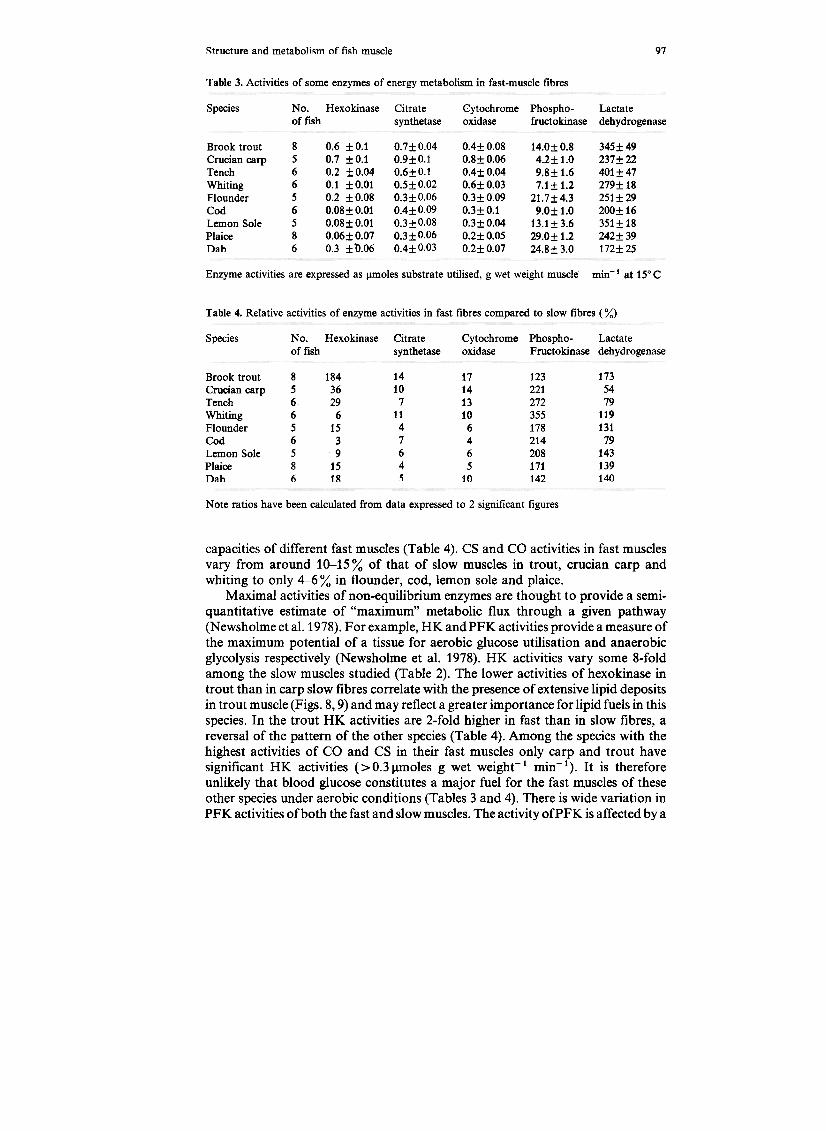
Table 3. Activities of some enzymes of energy metabolism in fast-muscle fibres
Species No. Hexokinase
of ti~h
Citratesynthetase
Cytochrome Phospho- Lactate
oxidase fructokinase dehydrogenase
856656586
0.6 :!:0.10.7 :!:0.10.2 :!: 0.040.1 :!: 0.010.2 :!: 0.080.08:!: 0.010.08:!: 0.010.06:!: 0.070.3 :!: "0.06
O.7:tO.9:tO.6:tO.5:tO.3:tO.4:tO.3:tO.3:tO.4:t
0.4:t 0.080.8:t 0.060.4:t 0.040.6:t 0.030.3:t 0.090.3:t 0.10.3:t 0.040.2:t 0.050.2:t 0.07
14.0:!: 0.84.2:!:1.09.8:!: 1.67.1:!: 1.2
21.7:!:4.39.0:!:1.0
13.1 :!: 3.629.0:!: 1.224.8:!: 3.0
345:!::49237:!::22401 :!:: 47279:!:: 18251:!:: 29200:!:: 16351:!::18242:!:: 39172:!::25
Brook .troutCrucian carpTenchWhitingFlounderCodLemon SolePlaicenab
Enzyme activities are expressed as ~moles substrate utilised, 9 wet weight muscle min-i at 15°C
Table 4. Relative activities of enzyme activities in fast fibres compared to slow fibres ( %)
Species No. Hexokinaseof fISh
Citratesynthetase
Cytochrome Phospho- Lactateoxidase Fructokinase dehydrogenase
856656586
184
36
29
6
15
3
9
15
18
14
10
7
11
4
7
6
4
5
17
14
13
10
6
4
6
5
10
123221272355178214208171142
1735479
119131
79143139140
Brook troutCrucian carpTench
WhitingFlounderCodLemon SolePlaicef)ah
Note ratios have been calculated from data expressed to 2 significant figures
capacities of different fast muscles (Table 4). CS and CO activities in fast musclesvary from around 10-15% of that of slow muscles in trout, crucian carp andwhiting to only 4-6% in flounder, cod, lemon sole and plaice.
Maximal activities of non-equilibrium enzymes are thought to provide a semi-quantitative estimate of "maximum" metabolic flux through a given pathway(Newsholme et al. 1978). For example, HK andPFK activities provide a measure ofthe maximum potential of a tissue for aerobic glucose utilisation and anaerobicglycolysis respectively (Newsholme et al. 1978). HK activities vary some 8-foldamong the slow muscles studied (Table 2). The lower activities of hexokinase introut than in carp slow fibres correlate with the presence of extensive lipid depositsin trout muscle (Figs. 8,9) and may reflect a greater importance for lipid fuels in thisspecies. In the trout HK activities are 2-fold higher in fast than in slow fibres, areversal of the pattern of the other species (Table 4). Among the species with thehighest activities of CO and CS in their fast muscles only carp and trout havesignificant HK activities ( > 0.3 J.lmoles g wet weight-1 min-1 ). It is thereforeunlikely that blood glucose constitutes a major fuel for the fast muscles of theseother species under aerobic conditions (Tables 3 and 4). There is wide variation inPFK activities ofboth the fast and slow muscles. The activity ofPFK is affected by a
0.040.10.10.020.060.090.080.060.03

98 IA. .1 ohnston and T .W .Moon
Fig. I. Transverse section through myotomal slow muscle fibres of the brook trout. Note the largedeposits of lipid (L) between the muscle fibres; intracellular lipid droplets (L) and high mitochondrial
density (M7)
Fig. 2. Junction between two slow fibres from brook trout showing the subsarcolemmal mitochondrialzona (MS), and numerous lipid droplets (L)
99Structure and metabolism of fish muscle
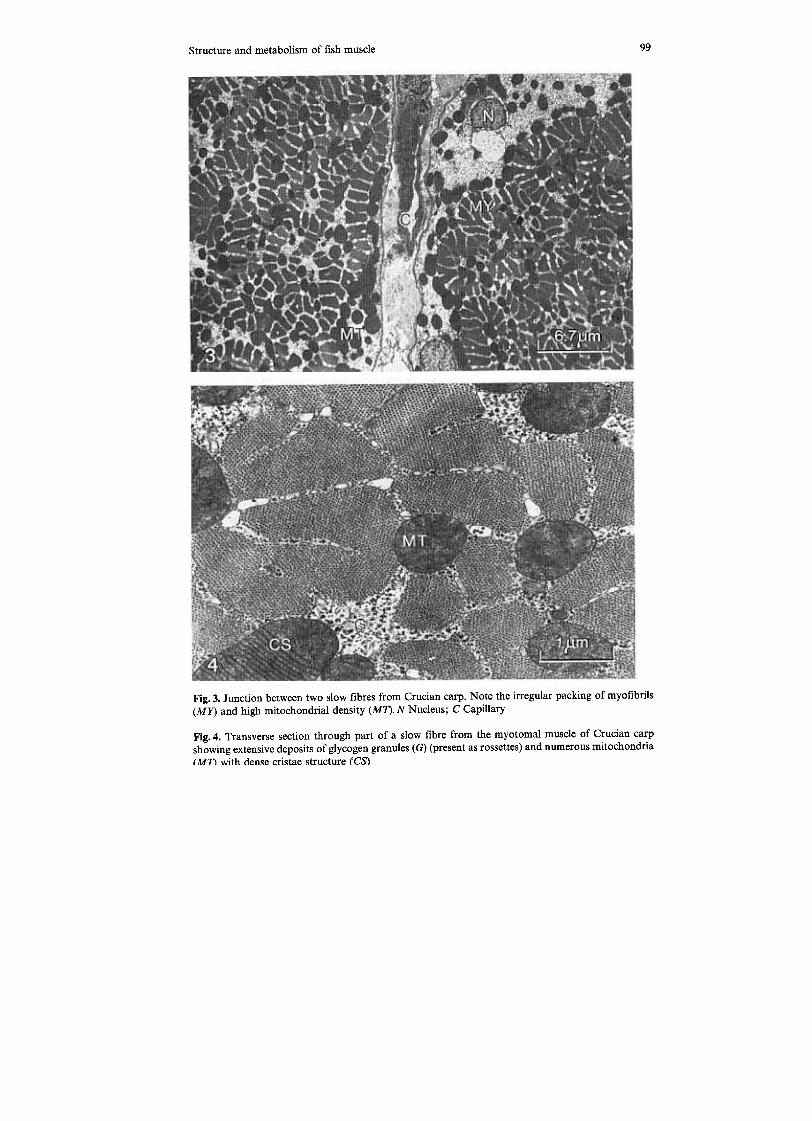
Fig. 3. Junction between two slow fibres from Crucian carp. Note the irregular packing of myofibrils
(MY) and high mitochondrial density (MI). N Nucleus; C Capillary
Fig. 4. Transverse section through part of a slow fibre from the myotomal muscle of Crucian carpshowing extensive deposits of glycogen granules (G) (present as rossettes) and numerous mitochondria
( MT\ with dense cristae structure (CS)

I.A. Johnston and T.W. Mool100
wide range of allosteric effectors including AMP , Pi and citrate. Thus some of thevariation in PFK activities may reflect differing degrees of activation of the enzymeamong the species since high-speed homogenates were employed. However, theoptimal conditions for fast and slow muscles were found to be similar for carp, troutand plaice, in which conditions for measurement of maximal activities weredetermined experimentally. It is likely, therefore, that the ratios ofPFK activitiesbetween fast and slow muscles proviGe an indication of the relative capacities ofthese fibre types for anaerobic glycolysis. PFKactivities vary from 1.2 (brook trout)to 3.6 (whiting) times higher in fast than slow muscles (Table 4).
LDH activities range from 124-5091!moles g wet weight-1 in slow fibres to 172-4011!moles g wet weight-:; 1 in fast muscles (Tables 2, 3). This enzyme functions to
maintain redox balance during anaerobic glycolysis by providing a continuoussupply of oxidized NAD. No obvious correlation is evident between muscle LDHactivity and either anoxic tolerance or known scope for anaerobic activity for thespecies. Unlike PFK and HK, LDH catalyses a reaction close to equilibrium andcannot be used as a quantitative index of capacity for anaerobic glycolysis (seeNewsholme and Start 1973).
Ultrastructural investigations
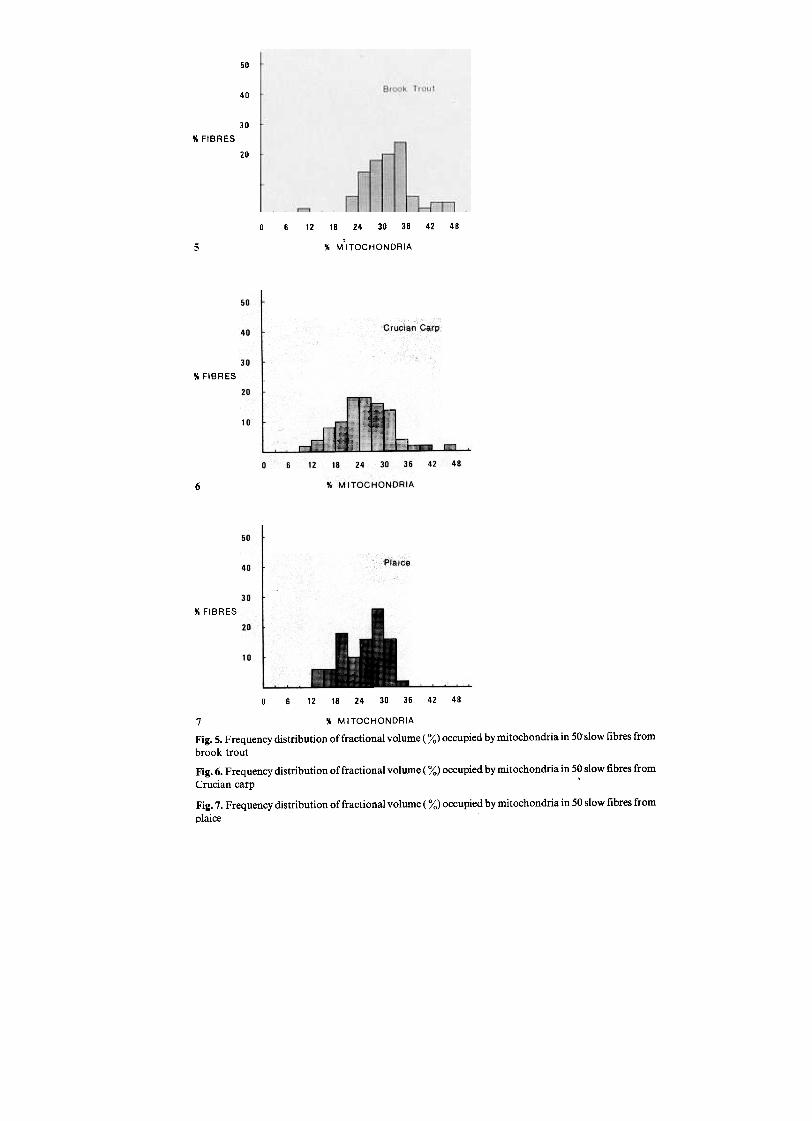
Typical structures of slow and fast fibres are shown in Figs. 1-4 and 8-10respectively. Measurements of activities of CS and CO only provide an averagemeasure of the aerobic capacities of different muscles. Morphometric analyses ofmitochondrial content of individual fibres reveal a considerable spread of aerobiccapacities within fibre populations, particularly for fast muscles (Figs. / , 15-17).The mean percentage of slow-fibre volume occupied by mitochondria (MF) issimilar among species and in the range 25-31% (Table 5). In contrast, the MF offast fibres varies from 8% of slow muscle values in plaice to 30% in trout (Table 5).
A good correlation was obtained for the activities of CS and CO in trout, carpand plaice fast muscles with the mean MF in these fibres (Tables 3, 4 and 5).
In general, small fast fibres of the carp have a higher MF than large ones(~1500I.Lm2) (Figs. 13, 14, 18). Fast fibres with areas >2500I.Lm2 invariablycontained very few mitochondria (Fig. 18). In plaice white muscle 42% of fibreshave a MF ofless than 1% compared to only 4% in carp (Table 5). The proportion
Table 5. Mitochondrial content of fast and slow muscles
Species Type of fibre Mitochondrial content (Volume %)
Mean:tS.E,
%
% Fibres~1%
% Fibres
?5%
% Fibres
~10%
10036
100
Slow
Fast
Slow
Fast
Slow
Fast
31.3:!: 0.99.3:!:0.7
25.5:!: 1.0
4.6:!:0.1
24.6:!: 0.82.0:!:0.3
oo
04
O42
O
76
O
32
O
12
Brook trout
Crucian carp
1000
Plaice
50
40
30
" FIBRES20
18 24 30 36 42
II MITOCHONDRIA
480 6 12
5
" FIBRES
6
" FIBRES
0 6 12 18 24 30 36 42 48
7 " MITOCHONDRIA
Fig. 5. Frequency distribution of fractional volume ( %) occupied by mitochondria in 50 slow fibres from
brook trout
Fig. 6. Frequency distribution of fractional volume ( %) occupied by mitochondria in 50 slow fibres fromCrucian carp .
Fig. 7. Frequency distribution of fractional volume ( %) occupied by mitochondria in 50 slow fibres from
plaice
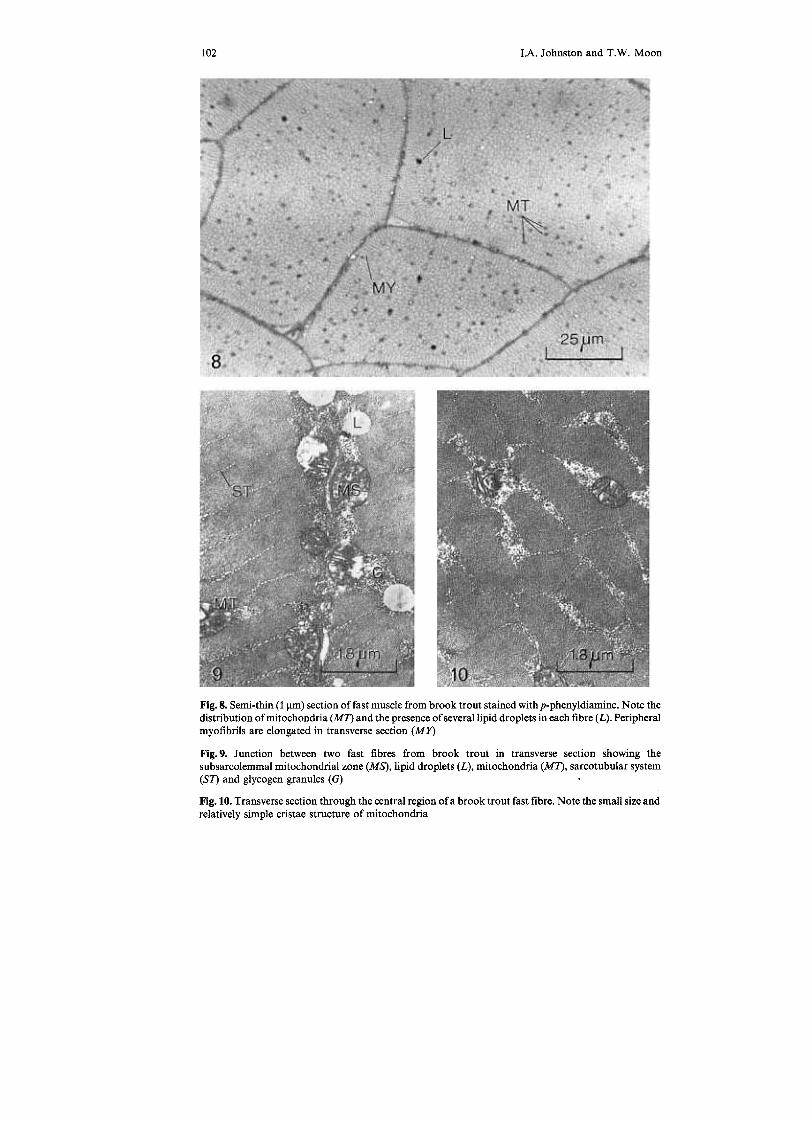
IA. Johnston and T.W. Moon102
Fig.8. Semi-thin (1 ~m) section of fast muscle from brook trout stained with p-phenyldiamine. Note thedistribution of mitochondria (M7) and the presence of several lipid droplets in each fibre (L). Peripheralmyofibrils are elongated in transverse section (MY)
Fig.9. Junction between two fast fibres from brook trout in transverse section showing thesubsarcolemmal mitochondrial zone (MS), lipid droplets (L), mitochondria (M7), sarcotubular system(S7) and glycogen granules (G)
Fig. 10. Transverse section through the central region ofa brook trout fast fibre. Note the small size andrelatively simple cristae structure of mitochondria
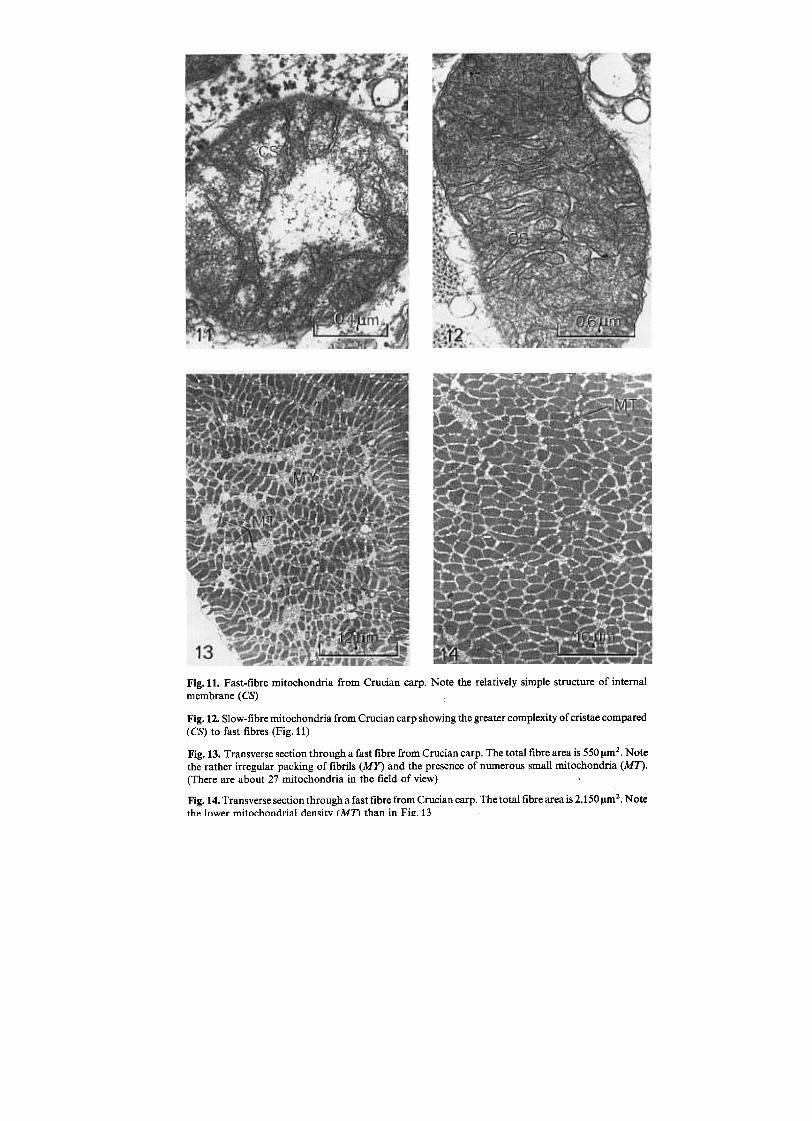
Fig. 11. Fast-fibre mitochondria from Crucian carp. Note the relatively simple structure of internal
membrane (CS)
Fig. 12. Slow-fibre mitochondria from Crucian carp showing the greater complexity of cristae compared
(CS) to fast fibres (Fig.11)
Fig. 13. Transverse section through a fast fibre from Crucian carp. The total fibre area is 550 !1m2. Notethe rather irregular packing of fibrils (MY) and the presence of numerous small mitochondria (MI').(There are about 27 mitochondria in the field of view)
Fig. 14. Transverse section through a fast fibre from Crucian carp. The total fibre area is 2,150 !1m 2. Note
the lower mitochondrial densitv (MT\ than in Fil!.13
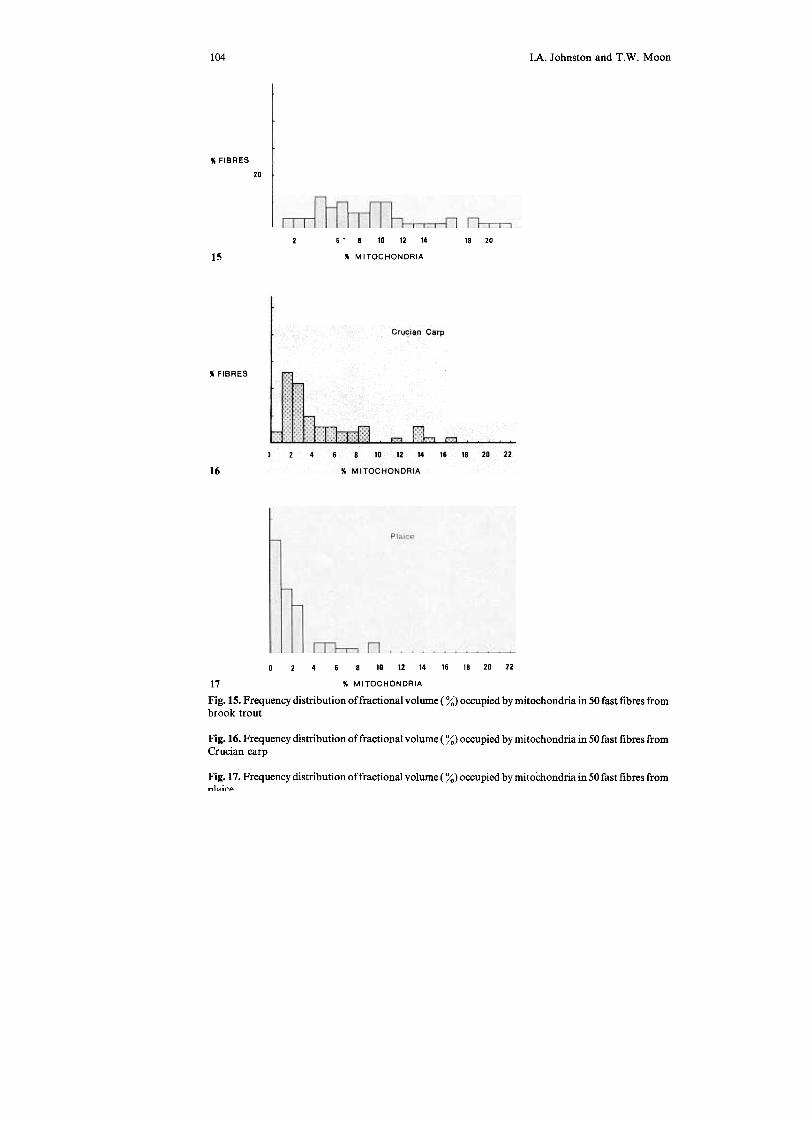
104 IA. Johnston and T.W. Moon
" FIBRES20
2 6 8 10 12 14
" MITOCHONDRIA
18 20
15
" FIBRES
16
0 2 4 6 8 10 12 14 16 18 20 22
17 " MITOCHONDRIA
Fig. 15. Frequency distribution of fractional volume ( %) occupied by mitochondria in 50 fast fibres frombrook trout
Fig. 16. Frequency distribution of fractional volume ( %) occupied by mitochondria in 50 fast fibres fromCrucian carp
Fig. 17. Frequency distribution of fractional volume ( %) occupied by mitochondria in 50 fast fibres from,,1,,;,..,.

105Stnlcture and metabolism of fish muscle
Fig. 18. A graph showing thefractional volume occupied bymitochondria ( %) fibres of Cruciancarp of different sizes. Units of fibrearea are I1m2, Note the data doesnot constitute a random sample(unlike that presented in Fig. 16) dueto the difficulty of measuring thearea of very large fibres at sufficientresolution to distinguish individualmitn"hnnciria ,
«~ozO:I:uOf-
~~
of fast fibres having MF of more than 10% varies from 0 in plaice to 36% in trout.However, in no case is there a significant overlap between the MF of fast and slowfibres (Figs. 5,--7, 15-17; Table 5). The cristae of mitochondria from fast muscles oftrout, carp and plaice have a relatively simple structure compared to those of slowmuscles (Figs. 11, 12). It therefore seems likely that the ratio of aerobic capacities offast/slow fibres is somewhat less than a simple measure of relative mitochondrialdensities would indicate. Nevertheless, the results clearly demonstrate that afraction of the fast fibre population has a significant aerobic capacity. For example,in the case of an active fish, such as brook trout, around 10% of fast-muscle fibre~have a MF which is more than twice that of the mean (Fig. 15).
Discussion
The MF in slow fibres of fish is around 25-31% in the species studied (Table 5).Even higher values have been obtained for the Atlantic mackeral (Scomberscomber) (35.5 %) (Bone 1978b) and the European anchovy (Engraulisencrasicolus)(45.5 %) (Johnston 1981, unpublished results). These are comparable to those of theventricles of the mouse (38 %) and finch (34 %) (Bossen et al. 1978) andconsiderably higher than that of slow twitch fibres from mammalian limb muscles.For example, soleus muscles ofrat and guinea pig have a MF of 4.9% (Eisenberg etal. 1974) and 5.7-7.4% (Stonnington and Engel 1973) respectively. Somescrombroid fishes (e.g. Katsuwonus) maintain elevated temperatures in brain andmuscle by means of a counter-current vascular heat exchange (Carey and Teal1969; Stevens and Neill 1978). Slow muscles of the skipjack tuna, Katsuwonuspelamis, have a MF which is only around half that of Scomber scomber which doesnot operate at temperatures much above ambient (Bone 1978b). It seems possiblethat the elevated body temperatures of homeotherms allow mitochondria tooperate somewhat more efficiently than those of ectotherms. Nevertheless, it isclear that slow fibres of fish have among the highest aerobic capacities of any
vertebrate skeletal muscles.The average MF of the teleost fast muscles studied, 2-9% ' (Table 5) are
somewhat higher than that reported for species with single motor endplates: sharks -
20
10
r o~ ° \ 00 .° 0. .. .(,
. \ ...
1000 2000 3000
FIBRE AREA

TA Jnhn.tnn and T .W .Moon10fi
Scyliorhinus 1% (Totland et al. 1980); Etmopterus spinax 1 %, Galeus melastomus0.5% (Kryvi 1977) and the sturgeon Acipenser ste//atus 0.7% (Kryvi et al. 1980).Such low mitochondrial densities are likely to be associated entirely with restingmetabolism, e.g., maintenance of ion gradients, protein turnover, etc.
There is now abundant evidence that, in contrast to elasmobranchs, someteleosts recruit fast motor units at low sustainable swimming speeds. For example,Bone and his co-workers ( 1978) found that white fibres in common carp ( Cyprinuscarpio) were active at all speeds above 0.5 body length S-I. The threshold speed forrecruitment of fast fibres in brook trout, of similar size to those in the present study,is around 1.8 bodylength S-I (Johnston and Moon 1980b). It has been generallyassumed that contractions of teleost fast fibres are largely supported by anaerobicglycogenolysis as is the case for elasmobranchs (see Biliilski 1974 and Bone 1978a,for reviews). However, anaerobic pathways constitute an inefficient means ofproducing A TP for sustained activity. The present study provides evidence thatthere is not a complete dichotomy in the metabolic characteristics of fast and slowmuscles in teleosts (Tables 2-5). For example, it can be seen from Table 5 that 36 %of trout fast muscle fibres contain more than 10% mitochondria. A brook trout of59 g contains around 2.5 g of slow fibres ( 4.2% body weight) and 32.9 g of fast fibres(55.8% body weight). Thus, fibres in which the MF is more than 10% constitute anactive mass of muscle at least comparable to that of the slow fibres: Although theaerobic capacity of fast fibres is far less than that of slow fibres on the basis of theabundance and structure of mitochondria, this clearly represents a significant
,<\:."t.q,~~ ~t...~~t...t."'t.t..The question arises as to what fraction of the energy requirements of white
fibres can be met entirely aerobically. This is likely to be critically dependent onswimming speed. Studies of oxygen consumption (Smit et al. 1971) and wholeanimal metabolite balance (Wokoma and Johnston 1980) have provided evidencethat at high sustainable speeds both goldfish (3.4 bodylength s- 1) and rainbowtrout (3.5-5 bodylength S-I ) have a significant anaerobic component. Under theseconditions it is necessary to control both redox balance and intracellular pH withinthe fast muscle if anaerobic contractions are to be supported for any length of time.Small trout can swim at 51engths s- I for more than 4 h during which time total body
lactate rises from 16 to 29 ~moles g, body weight (Wokoma and Johnston 1980). IIhas been suggested that lactate produced in the white muscle is transferred to moreaerobic tissues such as red muscle, gills, liver and kidney for subsequent oxidatioIJto pyruvate (Bilinski and Jonas 1972; Bone 1975). However, the quantitativeimportance of these pathways in unknown for any fish species.
The fraction of total energy needs supplied by anaerobic glycogenolysis at lo~swimming speed is more difficult to assess experimentally. However, it woul(appear that for at least trout and carp (Table 3) contractions of fast fibres at the ver)low speeds might be largely supported by aerobic pathways. Principal fuels artlikely to be either blood glucose (probably only important in trout and carp amon~the species studied) or local glycogen stores (see Tables 3 and 4). Interestingly, fasjfibres in brook trout contain intracellular lipid droplets (Fig.9). In this specie!activities of 3-0H acyl CoA dehydrogenase (fatty acid catabolis~ enzyme) arearound 18% of slow fibres, indicating some ability to use lipid fuels (Johnston an(Mnnn 1QROh\

107St.rllct.ure and metabolism of fish muscle
To utilise glycogen aerobically, it is necessary to have a mechanism for shuttling
hydrogen between cytoplasm and mitochondria. The two most importantpathways in vertebrate muscle are thought to be the glycerophosphate cycle and the
malate-aspartate shuttle (Newsholme and Start 1973; Dawson 1979). The former
appears to be the most important in the white muscle of skipjack tuna on the basis
of enzyme activity measurements (Guppy and Hochachka 1978; Hochachka et al.1978). There are differences betwee.n species with respect to the threshold speed for
recruitment of fast fibres (see Bone 1978a and Johnston 1981a, b). For example,
electromyograms can be recorded at speeds as low as 0.8 body length-1 in small
Pollachius virens (Johnston and Moon 1980a) but not until 3.2 and 4.5 body
lengths-1, respectively in striped bass (Morone saxatilis) and bluefish (Pomatomussultatnix)(Freadman 1979). Tables 2 and 5 indicate that the aerobic capacities of
multiply innervated fast muscles also vary widely between species as does their
degree of vascularization (Mosse 1978, 1979). It seems probable that some of this
species variation in aerobic capacity reflects different degrees of involvement qf the
fast muscles in sustained activity.In all species studied there is a wide distribution of fibre size within the fast
muscle. There is a trend for the MF to be' greatest in small fibres. For example, incrucian carp, fast fibres larger than 1000 Jlm 2 in cross-sectional;u:ea contain < 2 %
mitochondria whereas very small fibres ( < 100 Jlm 2) contain 12-17 % mitochondria(Fig. 18). The fast fibres of these species are histochemically homogenous on the
basis of staining for myofibrillar ATPase activity (Johnston 1981b). Unlike
mammals, fibre number increases throughout life in many fish by cell division and
proliferation ofmyosatellite cells (Greer-Walker 1970; Kryvi 1975). Recently, wehave shown that each fast motor axon in cod (Gadus morhua) innervates fibres with
a wide range of different diameters (Altringham and Johnston, unpublishedresults). It is thus likely that the range of fibre size represents states of fibre growth
and maturation rather than distinct fibre types (see also Bone 1978a; Johnston
1981b). An interesting possibility is that fast fibres with a higher than average
mitochondria content have a role in the aerobic catabolism of lactate produced by
the recruitment fast motor units at sustained swimming speeds.
Acknowledgements. Supported by a grant from Science Research Council to I.A. Johnston. T. W. Moonwas supported in part by an operating grant from the NSERC of Canada.
References
Barets A (1961) Contribution a retude des systemes moteur lent et rapide des muscle lateral des
teleosteens. Arch Anat Microsc Morphol Exp 50 Suppl:91-187Bilinski E, Jonas REE (1972) Oxidation of lactate to carbon dioxide by rainbow trout (Salmo gairdnerl)
tissues. J Fish Res Board Can 29: 1467-1471Bilinski E (1974) Biochemical aspects offish swimming. In: Malins DC, SargentJR (eds) Biochemical
perspectives in Marine Biology: Academic Press, New York, Vo11, pp 239-288Bone Q (1964) Patterns of muscular innervation in the lower chordates. Int Rev Neurobio16:99-147Bone Q (1966) On the function of the two types ofmyotomal muscle fibres in elasmobranch fish. J Mar
Biol Ass UK 46:321-349Bone Q (1970) Muscular innervation and fish classification. Simp Int Zoofi11st Univ Salamanca, pp
369-377Bone Q (1975) Muscular and energetic aspects offish swimming. In: Wu TY -T , Brok~w CJ, Brennen C
(.,ti.) Swimming and flving in nature. Plenum. New York. Vol 2. pp 493-528

108 I.A. Johnston and T.W. Moon
Bone Q (1978a) Locomotor muscle. In: Hoar WS, Randall DJ (eds) Fish Physiology. Academic Press,New York San Francisco London, Vol 7, pp 361-424
Bone Q (1978b) Myotomal muscle fibres types in Scomber and Katsuwonus. In: Sharp GD, Dizon AE(eds) The physiological ecology of tunas. Academic Press, New York, London, San Francisco, pp183-205
Bone Q, Kicenuik J, Jones DR (1978) On the role of the different fibre types in fish myotomes atintermediate swimming speeds. Fish Bull 76:691-699
Bossen EH, Sommer JR, Waugh RA ( 1978) Comparative stereology of the mouse and finch left ventricle.Tissue and CeI110:(4) 773-784
Carey FG, Teal JM (1969) Regulation of body temperature by the blue fin tuna. Comp Biochem Physiol28:205-213
Dawson AG (1979) Oxidation of cytosolic NADH formed during aerobic metabolism in mammaliancells. TIBS August 171-1""{6
Eisenberg BR, KudaAM, Peter JB (1974) Stereological analysis ofmammalian skeletal muscle. I. Soleusmuscle of the adult guinea pig. J Cell Bioi 60: 732-754
Freadman MA (1979) Role of partitioning of swimming musculature of striped Bass, Morone saxatilisWalbaum and Bluefish, Pomatomus saltatrix L. J Fish Bioi 15:417-423
Greer-Walker MG (1970) Growth and development of the skeletal muscle fibres of the cod (Gadusmorhua L.). J Cons Perm Int Explor Mer 33:228-244
Greer-Walker M, Pull GA (1975) A survey ofred and white muscle in marine fish. J Fish Bioi 7:295-300Guppy M, Hochachka PW (1978) II Skipjack tuna white muscle: A blueprint for the integration of
aerobic and anaerobic carbohydrate metabolism. In: Sharp GD, Dizon AE (eds) The Physiologicalecology of tunas: Academic Press, New York London San Francisco, pp 175-181
Hesse A (1970) Vertebrate slow muscle fibres. Physiol Res 50:40-62Hochachka PW, Hulbert WC, Guppy M (1978) The Tuna power plant and furnace. In: Sharp GD,
Dizon AE (eds) The physiological ecology of tunas: Academic Press, New York London SanFrancisco, pp 153-174
Hollander H, Vaaland J (1968) A reliable staining method for semi-thin sections in experimentalneuroanatomy. Brain Res 10: 120-126
Hudson RCL (1969) Polyneuronal innervation of the fast muscles of the marine teleost Cottus scorpiusL. J Exp Bioi 50: 47-67
Hudson RCL (1973) On the function of the white muscles in teleosts at intermediate swimming speeds. JExp Bioi 58:509-522
Johnston lA (1981a) Specializations of fish muscle. In: Goldspink DF (ed) Development andspecializations of muscle. Society Exp Bioi Seminar Series: Cambridge University Press 7: 123-148
Johnston lA (1981 b) Structure and function offish muscles. In: Day MH (ed) Verterbrate Locomotion.Zool Soc Symp, Ac~demic Press, London 48:72-113
J ohnston lA, Moon TW ( 1980a ) Endurance exercise training in the fast and slow muscles of a teleost fish(Pollachius virens). J Comp Physiol B 135:147-156
Johnston lA, Moon TW (1980b) Exercise training in skeletal muscle of brook trout (Salvelinusfontinalis). J Exp Bioi 87:177-194
Johnston lA, Patterson S, Ward PS, Goldspink G (1974) The histochemical demonstration ofmyofibrillar adenosine triphosphatase activity in fish muscle. Can J Zool 52:871-877
J ohnston lA, Ward PS, Goldspink G ( 1975) Studies on the swimming musculature of the rainbow trout.I: Fibre types. J Fish Bioi 7:451-458
Johnston lA, Davison W, Goldspink G (1977) Energy metabolism of carp swimming muscles. J Comp
PhysioI114:203-216Kryvi H (1975) The structure of myosatellite cells in axial muscles of the shark (Galeus melastomus).
Anat Embryol.147:35-44Kryvi H (1977) Ultrastructure of the different fibre types in axial muscles of the sharks Etmopterus
spinax and Galeus melastomus. Cell Tissue Res 184:287-300Kryvi H, Flood P, Guljaei D (1980) The ultrastructure and vascular supply of the fibre types in the
sturgeon (Acipenser stellatus). Cell Tissue Res 212:114-126Mosse PRL (1978) The Distribution of capillaries in the somatic musculature of two vertebrate types
with particular reference to teleost fish. Cell Tissue Res 187:281-303 .Mosse PRL (1979) Capillary distribution and metabolic histochemistry of the lateral propulsive
musculature of pelagic teleost fish. Cell Tissue Res 203: 141-660

Structure and metabolism of fish muscle 109
Newsholme EA, Start C (1973) Regulation in metabolism. Wiley, New York, ppNewsholme EA, Zammit V A, Crabtree B (1978) The role of glucose and glycogen as fuels for muscle.
Biochem Soc Trans 6:512-520Smit H, Amelink-Koutstall IM, Vijverberg I, von Vaupel-Klein IC (1971) Oxygen consumption and
efficiency of swimming goldfish. Comp Biochem Physiol A39: 1-28Stevens ED, Neill WH (1978) Body temperature relations of tunas, especially skipjack. In: Hoar WS,
Randall DI (eds) Fish Physiology: Academic Press, New York, Vol 7, pp 315-359Stonnington HH, Engel AG (!973) Normal and denervated muscle. Neurology 23:714-725Totland GK, Kryvi H, Bone Q, Flood PR (1980) Vascularization of the lateral muscle of some
elasmobranchiomorph fishes. I Fish BioI (In Press)Walesby NI, Johnston lA (1980) Fibre types in the locomotory muscles of an Antarctic teleost
Notothenia rossii: A histochemical, ultrastructural and biochemical study. Cell Tissue Res 208: 143-164 ,
WebbPW (1975) Hydrodynamics and energetics offish propulsion. Bull Fish Res Board Can No 190.158 pp
Weibel EC (1969) Stereological principles for morphometry in electron microscopy catology. Int RevCyt 26:235-299
WokomaA, Johnston lA (1980) Lactate production at high sustltinable cruising speeds in rainbow trout(Salmo gairdneri Richardson). I Exp BioI 90:361-364
Accepted February 17, 1981





























