Embed Size (px)
Citation preview

JOURNALOF
Dermatological Science
ELSEVIER Journal of Dennatological Science 12 (1996) 104- 109
Fibrous-long spacing fiber formation by collagen and non-collagenous acidic components from calf skin
Shingo Tajima* Department of Dermatology, Keio University School of Medicine, 35 Shinanomachi, Shinjuku-ku, Tokyo 160, Japan
Received 9 February 1995; revision received 29 June 1995; accepted 26 July 1995
Abstract
Fibrous-long spacing (FLS) fibers were reconstituted in the supernatant fraction after disaggregation of calf insoluble collagen fibers. Macromolecular components of the fibers were biochemically analyzed. Amino acid analysis demonstrated that the major component of FLS was collagen consisting of types I and III collagen in the relative percentage of 92 and 8% respectively. Hyaluronic acid and dermatan sulfate were the major components of glycosaminoglycans present in the ratio of 1:2. Polyacrylamide gel electrophoresis (PAGE) revealed several minor non-collagenous proteins which were not derived from serum components. The results suggest that non-collagenous components including glycosaminoglycans, are involved in the formation of FLS.
Keywords: Fibrous long spacing fiber; Collagen; Glycosaminoglycan; Non-collagenous protein
1. Introduction
Collagen is a major component of dermal con- nective tissue and plays an essential role in the development of fibrotic diseases such as sclero- derma and keloid. The mechanism of collagen fibrogenesis is not fully understood yet and the factors regulating the collagen fibrogenesis are not known. Type I collagen fibril formation in the native fibrils is controlled to a large extent bq’ the amino acid sequence of the collagen and by the distribution of polar and hydrophobic residues
* Corresponding author.
[1,2]. However, the facts that collagen can be organized in the different macromolecular struc- tures in the different tissues and that type I collagen solution can form unusual aggregates such as fibrous-long spacing (FLS) or segment-long spac- ing (SLS) fibers under certain in vitro conditions indicate the involvement of additional regulatory elements other than the collagen molecule, such as interaction with non-collagenous constituents [1,2].
FLS aggregates have been reported to be recon- stituted in vitro when soluble collagen solution was incubated with serum @,-acid glycoprotein [3] or proteoglycan [4]. FLS fibers are thought to be formed by the bridged interaction between un- paired positively charged residues of collagen
0923-181 l/96/$15.00 0 1996 Elsevier Science Ireland Ltd. All rights reserved SSDI 0923-1811(95)00468-8

S. Tajima /Journal of Dermatological Science 12 (1996) 104-109 105
molecules. There are at least four different forms of FLS, FLS I-IV, with similar symmetric band- ing patterns but different intermolecular staggers [4]. FLS fibers have been observed in the various tissue in vivo under normal and diseased condi- tions [5]. The mechanism by which FLS fibers were formed in vivo remained unclear at present.
In the course of exploring the mechanism of collagen fibrogenesis using insoluble collagen fibrils, fiber and fiber-bundle isolated by disaggre- gating calf skin chips [6], we found that FLS fibers were formed in the supernatant fraction [7]. We attempted to analyze the macromolecular components involved in the formation of FLS fibers.
2. Materials and methods
2.1. Preparation of FLS fibers from acid-insoluble calf skin chips
Three-month-old calf skin removed 2-3 h after sacrifice was obtained from Toyo Chikusan Co. Ltd., Tokyo and stored at - 40°C until use. A piece of the frozen skin was thawed and hairs and subcutaneous tissues were removed with razor blades and scissors. The skin was washed with chilled water several times and cut into 11 cm chips with scissors. The chips were placed in 0.5 M acetic acid containing protease inhibitor cock- tail (1 mM of EDTA, NEM and PMSF) for 2 weeks at -40°C to make the collagen fibers swollen [7]. The chips were then thawed and minced with scissors and razor blades to obtain a slurry of insoluble collagen fibers [6], then cen- trifuged at 8000 x g for 30 min at 4°C. The precipitate was washed with cold distilled water several times until the pH of the solution becomes neutral. To about 50 g of the material 1 1 of 0.1 M Tris-HCl, pH 7.8, containing 0.5 M NaCl, 0.05 M EDTA and 0.2 M /?-mercaptoethanol was added and the suspension was stirred at 4°C for 3 days to disaggregate the insoluble collagen fibers, then centrifuged at 53 000 x g for 2 h at 4°C. The supernatant was exhaustively dialyzed against distilled water for 2 days at 4°C. The fibrous precipitate formed during dialysis was isolated with the centrifugation at 10 000 x g for 30 min at 4°C and washed with distilled water twice, then
subjected IO electron microscopic observation or biochemical analysis after lyophilization.
2.2. Morphological observations A suspension of the precipitate was placed on a
grid (VEC’O 200 mesh, Oken Shoji Co. Ltd., Tokyo) and stained with 2% uranyl acetate for 3-4 s. Immediately after removal of the excess staining solution with a filter paper, the stained materials were examined with a Hitachi model HU-1 1A electron microscope.
2.3. Chemi’cal analyses Amino acid composition was determined as
described in our previous study [6,7]. Gly- cosaminoglycans were isolated from the fibrous
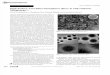
Fig. 1. Electron microscopic observation of the fibrous precip- Fig. 1. Electron microscopic observation of the fibrous precip- itate. A fibrous component formed during dialysis of the itate. A fibrous component formed during dialysis of the supematant fraction was stained with uranyl acetate. The scale supematant fraction was stained with uranyl acetate. The scale indicates 0.2 pm. indicates 0.2 pm.

106 S. Tajima / Journal of Dermatological Science 12 (1996) 104109
Table 1 Amino acid compositions of FLS fibers from dispersed calf dermis (Res/l 000 Res)
FLS Acid soluble collagen
HYP Asp Thr Ser Glu Pro @Y Ala l/2-cys Val Met Ileu Leu W Phe Trp His HY~ LYS A%
Total 1001 998
99 100 49 45 19 18 36 37 13 70
121 123 322 331 104 105
23 5
14 28 3
14 - 5
26 21 52 51
- 21 6
11 23 2
10 - 5
precipitate and quantified according to the method of Hata and Nagai [9,10]. Hexosamine content was determined on the amino acid auto- analyzer, model JEOL-6AS, after hydrolysis with 5 M HCl in an evacuated sealed tube at 110°C for 12 h [7,8].
2.4. Estimation of type I and type III collagens About 5 mg dry weight of sample material was
digested with 0.02% pepsin ( x 3 crystallized, Nutritional Biochemicals Corporation, Cleveland, OH) in 3 ml of 0.5 M acetic acid at 4°C for 24 h. Pepsin-solubilized collagen was precipitated by dialysis against 0.02 M Na,HP04 and were sub- mitted to 5% SDS-polyacrylamide gel elec- trophoresis (SDS-PAGE) in the presence of 3.6 M urea. The relative amounts of type I and type III collagen was estimated as described previously [l 11.
2.5. Electrophoretical analysis Samples were dissolved in 10
pH 6.8 containing 1% SDS and 100°C for 2 min and resolved
mM Tri-HCl, 1 mM DTT at on 7.5% SDS-
PAGE [12]. For native disc electrophoresis FLS- fibers were dissolved in 73 mM Tris, 0.35 M Glycine, pH 8.3 and PAGE was carried out ac- cording to Ornstein and Davis under the condi- tion of pH 8.9 [13] where collagen does not enter into the separating gel.
For the preparation of standard sample for electrophoresis and amino acid analysis, acid sol- uble collagen was extracted from calf skin with 0.5 M acetic acid for 2 days at 4°C and precipi- tated at both neutral and acidic pH, then lyophilized.
To know whether serum components like tlr- acidic glycoprotein are involved in FLS forma- tion, FLS fibers were dissolved in 0.05 M Verona1 buffer, pH 8.6 and fractionated on an agar plate, then reacted with anti-bovine serum antibody. The agar plate was stained with amide black.
3. Results
3.1. Ident@ation of FLS fibers on electron microscope
An electron microscopic observation demon- strated that the fibrous precipitate had a unique structure with repeating striations of 2000-3000 A and varying diameters (Fig. l), which is in agreement with the morphological structure of FLS fibers [5].
3.2. Amino acid composition of FLS fibers Amino acid composition of FLS fibers showed
no marked differences from that of acid soluble collagen, indicating that the major component of FLS is collagen. When compared with the amino acid composition of acid soluble collagen, FLS was relatively abundant in acidic (Asp and Glu) and neutral (Ileu and Leu) amino acids (Table 1).
3.3. Components of glycosaminoglycans of FLS fibers
Glycosaminoglycan components of FLS were hyaluronic acid and dermatan sulfate (Table 2). No other glycosaminoglycans were detectable by

Table 2
S. Tajima / Journal of Dermatological Science 12 (1996) 104-109 107
Amounts of collagen, glycosaminoglycans and non-collagenous proteins in FLS fibers formed from disaggregation of insoluble collagen fibers
Components Amounts (pg/mg dry weight)
Proteins
Glycosaminoglycans
Collagen 787 Type 1 92% Type III 8%
Non-collagenous protein 160 HA 3 DS 6
The amount of collagen was calculated from the hydroxyprohne value obtained by amino acid analysis. Non-collagenous protein content was calculated by subtracting the collagen content from total protein content estimated by the data of amino acid analysis. The collagen content was calculated considering the hydroxyprohne values of q (I), a2 (I) and GI, (III) chains as 97, 83 (16) and 121 residues (17) per 1000 residues, respectively.
cellulose acetate electrophoresis. The ratio of hyaluronic acid to dermatan sulfate was 0.5.
+top
-a,(m)
+w( 1)
+a2
-bottom
Fig. 2. Separation of type I and III collagens by SDS-PAGE in the presence of 3.6 M urea. Pepsinized samples were resolved on 5% SDS-PAGE without (lane 1) or with (lane 2) 1 mM dithiothreitol.
3.4. Content of type I and III collagen in FLS fibers
The amount of pepsin-insoluble material was < 1% of the starting material. Densitometrical scanning of the polyacrylamide gel revealed that the relative percent of type III collagen was 8%. No other collagenous bands indicative of type VI collagens were detected by SDS-PAGE (Fig. 2, Table 2).
3.5. Characterization of non-collagenous proteins involved in FLS fibers
SDS-PAGE showed three minor bands with apparent molecular weight of 60, 12 and 10 kDa were associated with FLS fibers which were not found in acid-soluble collagen (Fig. 3a). PAGE performed under the condition of Davis demon- strated that the doublet polypeptides distinct from serum q-acid glycoprotein were involved in FLS fibers but not in acid-soluble collagen (Fig. 3b).
Immunoelectrophoresis showed that the proteins involved in FLS fiber formation are not derived from serum (Fig. 4).
4. Discussion
It is now recognized that collagens are orga- nized into classical long, unbranched banded fibrillar structure (native form) with a characteris- tic periodicity of 60-70 nm [1,2]. FLS fibers ob- served in this experiment may be formed in association with the components extracted from the insoluble collagen fibers since purified type I collagen precipitated at both neutral and acidic

108 S. Tajima / Journal of Dermatological Science 12 (1996) 104109
a> b)
I =5% -66k -43k
I 2 3 4 I 2 3
Fig. 3. Separation of the proteins by PAGE. (a): The proteins of FLS fibers (lane 1), bovine serum albumin (lane 2) ovalbu- mm (lane 3) and acid soluble calf skin collagen (lane 4) were dissolved in 0.1 M Tris-HCl; pH 6.8, containing 1% SDS and resolved on 7.5% SDS-PAGE under reducing condition. (b): FLS (lane I), acid-soluble calf skin collagen (lane 2) and a,-acid glycoprotein (lane 3) were dissolved in Tris-Glycine buffer, pH 8.3 and resolved on native PAGE at pH 8.9. Both upper and lower gels were shown in the figure.
Fig. 4. Immunoelectrophoresis of FLS and the supernatant
Anti-BS
Anti-BS
fraction of FLS. Calf serum, FLS and the supernatant fraction (designated as 53 000s sup) of FLS were fractionated on an agar plate and reacted with anti-bovine serum antibody at 50 x dilution.
pH has been previously demonstrated to form native type fibers [I ,2].
Similar cross-banded structures have been re- ported in vivo and in vitro [14]. One is tissue microfibrils with a diameter of 5- 10 nm and a periodicity of about 100 nm which was identified as type VI collagen by immunoelectronmicro- scopic study [15], or by isolating a similar fibril with monoclonal antibodies [15]. FLS fibers ob-
tained in our experiment are distinct from type VI collagen microfibrils because the FLS had longer periodicity than type VI fibers and consisted of type I and III collagens not of type VI collagen based on the SDS-PAGE of pepsin digestion of FLS.
The FLS precipitate produced by the addition of serum q-acid glycoprotein in type I collagen solution in vitro has 220-240 nm periodicity [3]. Our results demonstrated that type I and III collagen complex is able to form FLS fibers which have the similar periodicity to type I collagen FLS. It is noted that the ratio of type I/type III collagen of FLS fibers (11.5) was greater than those of insoluble collagen fibrils fibers and fiber- bundles isolated by disaggregating the insoluble calf skin chips (3.6-4.5) [see Ref. 61. This suggests that the amount of type III collagen in the colla- gen fibers may be an essential factor for con- trolling the morphology of collagen fibers. In fact, it has been reported that a high proportion of type III collagen produce thinner collagen fibers in vitro [16]. Our results also demonstrated that FLS fibers are reconstituted under the interaction with the acidic proteins other than serum proteins, possibly with the proteins derived from connective tissue components of skin. Recent ad- vance in extracellular macromolecules in skin identified several novel non-collagenous proteins, including fibrillins, decorin tenescin, throm- bospondin, vitronectin [ 171 and dermatopontin [18]. Considering their molecular sizes, these non- collagenous proteins, however, are not likely to be involved in the FLS formation.
References
[l] Piez KA: Primary structure, in Biochemistry of collagen. Edited by GN Ramachandran, AN Reddi AH. Plenum Press, New York, 1976, pp. l-44.
[2] Kielty CM, Hopkinson I, Grant ME: The collagen family structure, assembly and organization in the extracellular matrix, in Connective Tissue and Its Heritable Disorders. Edited by PM Royce, B Steinmann. Wiley-Liss, New York, 1993, pp. 1033147.
[3] Highberger JH, Gross J, Schmitt FO: The interaction of mucoprotein with soluble collagen: an electron micro- scope study. Proc Nat1 Acad Sci USA 37: 2866291, 1951.
[4] Doyle BB, Hukins DWL, Hulmes DJS, Miller A Wood- head-Galloway J: Collagen polymorphism: its origins in the amino acid sequence. J Mol Biol 91: 79-99, 1975.

S. Tajima / Journal of Dermatological Science 12 (1996) Ill-109 109
[5] Edwards RP: Long-spacing collagen in skin biopsies from gels and related procedures, in The Proteins. Edited by H patients with lepromatous leprosy. Brit J Dermatol 93: Neurath, RL Hill. Volume 1, Academic Press, New York, 175-182, 1975. 1975, pp, 179-223.
[6] Matsumura T, Shinmei M, Nagai Y: Disaggregation of connective tissue: preparation of fibrous components from sea cucumber body wall and calf skin. J Biochem 73: 155-162, 1973.
[13] Davis B: Disc electrophoresis-II: method and application to human serum proteins. Ann NY Acad Sci 121: 404- 427, 1964.
[7] Tajima S, Nagai Y: Isolation and partial characterization of collagen fibers and fiber-bundles from insoluble calf dermis. Conn Tiss Res 7: 157- 163, 1980.
[8] Tajima S, Nagai Y: Distribution of macromolecular com- ponents in calf derrnal connective tissue. Conn Tiss Res 7: 65-71, 1980.
[14] Timpl R, Engel J: Type VI collagen, in Structure and Function of Collagen Types. Edited by R Mayne, RE Burgesoc. Academic Press, New York, 1987, pp. 105- 143.
[9] Hata R, Nagai Y: A rapid and micro method for separa- tion acidic glycosaminoglycans by two-dimensional elec- trophoresis. Anal Biochem 45: 462-468, 1972.
[lo] Hata R, Nagai Y: A micro calorimetric determination of acidic glycosaminoglycans by two-dimensional elec- trophoresis on a cellulose acetate strip. Anal Biochem 52: 652-656, 1973.
[1 I] Hayashi T, Nagai Y: Separation of the cc-chains of type I and III collagens by SDS-polyacrylamide gel elec- trophoresis. J Biochem 86: 453-459, 1979.
[12] Weber K, Osbom M: Proteins and sodium dodecyl sul- fate: molecular weight determination on polyacrylamide
[15] Bums RR, Press W, Engvall E, Timpl R, Gross J: Type VI collagen in extracellular, 100 nm periodic filaments and fibrils: identification by immunoelectron microscopy. J Cell Biol 103: 393-404, 1986.
[16] Lapiere CM, Nusgens B, Pierard GE: Interaction between collagen type I and type III in conditioning bundles organization. Conn Tiss Res 5: 21-29, 1977.
[17] von der Mark K, Goodman S: Adhesive glycoproteins, in Connective Tissue and Its Heritable Disorders. Edited by PM Royce, B Steinmann. Wiley-Liss, New York, 1993, pp. 21 l-236.
[18] Superti-F’urga A, Rocchi M, Schafer BW, Gitzelmann R: Complementory DNA sequence and chromosomal map- ping of a human proteoglycan-binding cell adhesion protein (dermatopontin). Genomics 17: 463-467, 1993.