Embed Size (px)
Citation preview

Plant Molecular Biology 26: 1423-1437, 1994. © 1994 Kluwer Academic Publishers. Printed in Belgium. 1423
Fatty acid signalling in plants and their associated microorganisms
Edward E. Farmer Institut de Biologie et de Physiologie VOgdtales, B4timent de Biologie, 1015 Lausanne, Switzerland
Received 10 March 1994; accepted in revised form 26 April 1994
Key words: fatty acids, signal transduction, wounding, jasmonic acid, traumatin, plant-microorganism interaction
Introduction
Fatty acid signals in plants have a long history. A fatty acid derivative, traumatic acid, was iden- tiffed as a cell division promoter in wounded bean mesocarp and was among the first ever biologi- cally active molecules isolated directly from a plant tissue. A structure for this molecule was published in 1939 [20]. Indolyl-3-acetic acid (IAA), an auxin isolated from urine in 1934, was definitively isolated from plant tissues in 1942 by Haagen-Smit et aL [32]. Like the complex story of the discovery of the structures of auxins, the true nature of traumatic acid has been confusing due to the more recent discovery that this com- pound may have become modified during the ex- traction leading to its discovery [77]. With re- newed interest in fatty acid signalling, some of the old literature is being revised and many new fac- ets of fatty acid signalling are emerging. One com- pound in particular, jasmonic acid, has been the focus of much attention as a signal that stimulates the expression of an array of wound-inducible and defence-related genes as well as playing roles in various developmental responses. While much of the current interest in plant-derived fatty acid signals centres on jasmonic acid, it is apparent that other biologically active fatty acids exist in plants.
Known fatty acid-derived signals in plants can be divided into several categories. These include
the jasmonate family (octadecanoid-derived cy- clopentanones and cyclopentanols; e.g. A and B in Fig. 1), the traumatin family and related alk- enals resulting from the action of hydroperoxide lyases on fatty acid hydroperoxides (Fig. 1D, E, F), and other classes such as highly oxygenated fatty acid derivatives reminiscent of animal lipoxins (e.g. phaseolic acid, Fig. 1C). Within these presently known families are examples of volatile molecules (Fig. 1, compound E and the methyl ester of A), glycosides (Fig. 1B), conju- gates with other, unrelated molecules, and fatty acids that are potentially unstable and reactive (especially the alkenals, Fig. 1D, E, F). Some of these properties have caused difficulties in experi- mentation, but on the whole add greatly to the interest of these compounds.
Of the presently known plant fatty acid signals jasmonic acid has been the most studied. Numer- ous reviews concerning jasmonates have ap- peared in the past five years [e.g. 3, 24, 33, 41, 49, 50, 62, 66, 72]. This review aims at summarizing some of the very recent information concentrat- ing on jasmonate as a mediator of cellular re- sponses. Progress in the quantitation of jas- monate and inhibition of its synthesis is discussed. Other fatty acid signals of interest in plants are highlighted, and finally some examples from the rapidly growing area of fatty acid signalling be- tween plants and microorganisms are summa- rized.
[187]

1424
A
B O ~ O73-GIc
D
E
OH ~ ~ [ ' ~ CHO
C o . F ~ ~ ~ A ~'COOH
HOHzC - OT- H y v O~ H C I O C ~ , ~ ~ ~ CHO
Fig. 1. Diverse signal functions for plant fatty acids. A. Jasmonic acid, involved in wound signalling, induces the synthesis of a variety of low- and high-molecular-weight defence-related compounds. Jasmonic acid is involved in several developmental re- sponses including the stimulation of tendril coiling. Other closely related cyclopentanone fatty acids and their derivatives appear to play diverse roles in development. As a methyl ester jasmonate is a volatile floral scent. Methyl jasmonate might also act as an air-borne defence signal. B. Tuberonic acid-fl-glucoside, a powerful natural inducer of tuberization in potato. The aglycone is less potent. C. Phaseolic acid (2-oxo-5,8,12-trihydroxydodecanoic acid) stimulates elongation in pea stem segments and induces e-amylase activity in barley endosperm. D. (2E)-4-Hydroxy-2-nonenal, a potent cytotoxin long known in animals; its synthesis by bean extracts was recently discovered. E. (2E)-hexenal, a toxin highly active against pests and pathogens; highly volatile, also a floral scent. F. Traumatin, 12-oxo-(10E)-dodecenoic acid, a cell division promoter in wounded bean parenchyma.
Jasmonates as wound signals
The jasmonate family of C12 fatty acids includes members which play roles in defence and devel- opment. Amongst the best studied jasmonate- responsive genes are many that are wound- inducible ones. Two classes of wound-inducible genes that have been studied in most detail are wound-inducible proteinase inhibitors and veg- etative storage proteins. The potential roles of jasmonate in regulating the expression of these classes of genes has been the subject of much investigation which has been reviewed [24, 66].
A useful preliminary classification of wound- inducible genes and their response to jasmonate has been developed by Creelman etal. [14]. Wound-inducible proteins (in dicots) include rep- resentatives of the following classes: (1) phenyl- alanine ammonialyase and chalcone isomerase, (2) extensins, glycine-rich proteins and hydrox- yproline-rich glycoproteins, (3) proteinase inhibi- tors and vegetative storage proteins (including some lipoxygenases), and (4)chitinase and fl-glucanase. Jasmonate-inducible members of the
first three classes have all been characterized. The final class of wound-inducible genes contains members that are ethylene-inducible although some representatives that are jasmonate-induc- ible are likely to emerge.
The list of genes which are both wound- and jasmonate-inducible is growing but there is still a lack of detailed information concerning mono- cots. A monocot protein which is clearly both wound- and jasmonate-inducible is a 61 kDa pro- tein from barley leaves [4]. This protein is ho- mologous to ribosome-inactivating proteins (W. Becker and K. Apel, personal communication) and may thus play a fundamental role in trans- lational regulation. A better understanding of wound signalling in monocots is likely to emerge from work on wound-inducible Bowman-Birk trypsin inhibitor in maize seedlings [19]. In this system a wound signal is transmitted unidirec- tionally (upwards) to tissues distal to the wound site making this an attractive model for systemic signalling.
In summary, jasmonate does not stimulate the expression of all wound-inducible genes but may
[188]

be an important and wide-spread regulator of many such genes throughout the plant kingdom. It will be interesting to look at the possible in- volvement ofjasmonic acid (or other fatty acids) in the regulation of other classes of plant genes that could be classed as wound-inducible, for ex- ample the cell division control protein kinase p34 ca°2. While many wound-inducible genes are clearly jasmonate-inducible, the converse is not always true. Not all jasmonate-inducible genes are wound-inducible [5]. Further critical investi- gations of the relationship of jasmonate-induc- ibility to wound-inducibility are needed. In the meantime wound-inducible proteinase inhibitors provide an excellent system with which to study the role of jasmonate as a wound signal.
Systemically wound-inducible proteinase in- hibitors are defence proteins which accumulate in the aerial parts of tomato plants upon wounding, for example during insect attack [30]. The sys- temic signal stimulating proteinase inhibitor gene expression at sites distal to the wound is an oc- tadecapeptide, systemin [ 51 ]. A working model for the wound induction of proteinase inhibitor gene expression in tomato leaves has been pub- lished [25]. The model proposes that jasmonate is synthesized in response to systemin and pos- sibly to oligogalacturonides, which may be locally released during pathogen attack [25]. The model suggests that jasmonate arises from the action of a lipase on membrane-associated linolenic acid, i.e. jasmonate is synthesized de novo. Jasmonate itself would then act to regulate the expression of proteinase inhibitor genes. In order to test this model several experimental approaches are being used and some of the chief questions being ad- dressed are: are intracellular levels of jasmonic acid elevated on wounding leaves and do these levels correlate with gene expression? Is jas- monate synthesized de novo or released from cel- lular conjugates? Does jasmonate play a role in signalling downstream from other signals such as systemin or oligouronides, o r , what is the signal order? Is a lipase or a lipase-like enzyme(s) re- sponsible for linolenic acid release from cell mem- branes and how does jasmonate regulate gene expression? Answers to some of these questions
1425
are presented in the sections concerning cellular levels of jasmonate and inhibition of the octade- canoid pathway. In general, much of the work to date is consistent with an in vivo role for jasmonate as a regulator of wound-inducible genes but many details are lacking, especially considering the lat- ter question: How does jasmonate regulate gene expression? The whole area concerning jas- monate receptors is unexplored. These is no evi- dence for the location of the receptor(s); they may reside near or in the nucleus or at the cell surface.
Jasmonates as developmental signals
Jasmonates are well known inducers of a wide variety of physiological responses in plants (re- viewed in [41, 62, 74] and are likely to play roles in plant development. A number of interesting developmental phenomena that are jasmonate- inducible are currently under investigation. A concrete developmental role for ajasmonate fam- ily member is emerging from the search for a tuber-inducing factor in potatoes [41]. The dis- covery of the tuber-inducing stimulus as tuberonic acid/3-glucoside [76] was a major breakthrough involving a great deal of demanding analytical work. The low levels of this substance and its high activity suggest that it may be a major regulator of tuberisation in potato, where it is synthesized in the leaves and transported downwards to the stolons. The fact that the fl-glucoside of tuberonic acid is more active than the free acid indicates the potential importance of conjugated fatty acid metabolites as natural regulators.
The induction of low-molecular-weight meta- bolites by exogenous jasmonic acid has been in- vestigated in a large number of plant suspension cultures [31 ]. In nearly every culture investigated (representing over 36 species)jasmonate has been found to induce the production of low-molecular- weight metabolites many of which have proposed roles in plant defence. The same compounds were in many cases induced by a yeast-derived elicitor which was shown to stimulate the accumulation of endogenous 3R,7S-jasmonic acid in suspen- sion-cultured cells of a variety of dicot, monocot and gymnosperm species [47]. In all cases exam-
[1891

1426
ined the increased synthesis of low-molecular- weight compounds correlated with the accumu- lation of endogenous 3R,7S-jasmonic acid. These quantitative data (some of which are presented in Table 1) have added to our knowledge of jas- monate as a signal, particularly since the kinetics of formation of a characterized stereoisomer was followed. By taking advantage of a suspension culture system, Mueller et al. [47] were also able to show that linolenic acid was released in re- sponse to elicitor treatment of cells, providing further evidence consistent with de novo syn- thesis ofjasmonate prior to the induction of low- molecular-weight compounds. A recent parallel study of alkaloid synthesis in Catharanthus (peri- winkle, Apocynaceae) and Cinchona (quinine, Rubiaceae) extends these studies to the whole plant level [ 1 ]. The seedlings of these two plants synthesize alkaloids at a precise developmental period. The plants are responsive to methyl jas- monate vapour, which increases the synthesis of alkaloids, only within this time frame. This work suggests that not all plant tissues appear to be jasmonate-responsive at all times.
One of the experimental systems of great inter- est in jasmonate research is the touch-induced coiling of the Bryonia tendril. Bryonia dioica is a dioecious climbing plant in the Cucurbitaceae. Both sexes produce tendrils which are mecha-
nosensory organs specialized to localize solid ob- jects, such as the stems of other plants, and then coil around them to provide support. Touching the ventral side of the tendril promotes a coiling reaction whereas touching the dorsal side of the tendril inhibits initiation of the coiling reaction. A series of developmental changes take place dur- ing touch-induced coiling. These include the lig- nification of a field of sclerenchyma cells (known as the Bianconi plate) as well as the proliferation of a ventral layer of collenchyma extending from the subepidermal cell layer to the lignified Bian- coni plate [36]. Linolenic acid, 12-oxo-phytodi- enoic acid and jasmonic acid can all induce the coiling response as well as some of the develop- mental changes associated with coiling [22]. En- dogenous levels of jasmonic acid increase dra- matically on coiling ([2]; see Table 1) strongly suggesting an in planta role for this molecule. It has been suggested that jasmonate, or one of its precursors, may couple the primary response, mechanostimulation, to the coiling response [75]. The fact that jasmonate stimulates tendril coiling as well as cell-type-specific developmental changes makes this a system of great interest since the bryony tendril is a relatively simple organ with a well defined cellular architecture. Another fea- ture of interest in the Bryonia tendril is the pres- ence of numerous longitudinally extended air
Table 1. Stimulus-induced changes in jasmonic acid levels in plants.
Plant Tissue Resting JA level Stimulus Maximum JA level Profile Reference
Arabidopsis thaliana L 40 + 1.1 ng per g fwt Wound 1360 + 708 ng per g fwt step Lycopersicon L 14 + 3.8 ng per g fwt Wound 52 + 13.8 ng per g fwt intermed.
esculentum Glycine m a x H ca. 80 ng per g fwt Wound ca. 500 ng per g fwt step Phaseolus vulgaris CC 2 ng per g dwt Elicitor 190 ng per g dwt intermed. Rauvolfia canescens CC 25 ng per g dwt Elicitor 1370 ng per g dwt spike Eschscholtzia CC 1.9 ng per g dwt Elicitor 74 ng per g dwt intermed.
californica Taxus baccata CC 3 ng per g dwt Elicitor 40 ng per g dwt step Agrostis tenuis CC 10 ng per g dwt Elicitor 1000 ng per g dwt spike Avena sativa L 14 ng per g fwt Wound 49 ng per g fwt intermed. Bryoniadioica L 15 + 1.1 ng per g fwt Wound 54+ 5 ng per g fwt intermed.
Bryonia dioica T 10 + 3 ng per g fwt Touch 52 + 18.7 ng per g fwt NYA
W.E. Weiler, pers. comm. W.E. Weiler, pers. comm. Albrecht et al. [2] Creelman et al. [ 14] Mueller et al. [47] Gundlach et al. 1992 Mueller et al. [47]
Mueller et al. [47] Mueller et al. [47] Albrecht et al. [2] W.E. Weiler, pers. comm. Albrecht et al. [2] Weiler et aL [75]
~Abbreviations: L, leaf; H, hypocotyl; CC, cell culture; T, tendril; fwt, fresh weight; dwt, dry weight; NYA, not yet available.
[190]

spaces, particularly those in two bands of meso- phyll-like cells which subtend the edges of the dorsal epidermis. It has been suggested that these air spaces play a role in the coiling response by allowing the air-borne intercellular transport of methyl jasmonate [22].
High levels of exogenous j asmonates frequently cause dramatic responses such as senescence or growth inhibition in plant tissues [62, 70]. Thus special care must be taken to provide physiologi- cally relevant jasmonate concentrations in the study of the developmental role of jasmonate in plants. Perhaps related to the effects of jas- monates on growth inhibition and senescence in plants are the recent studies of Reinbothe et al. [56, 57, 58] who explored the effects of exogen- ous jasmonate on gene expression in barley leaf segments. In addition to the stimulation of ex- pression of a number of genes involved in defence and development exogenous jasmonates have in- teresting inhibitory effects on the translation of a number of nuclear and plastid-encoded RNAs. The translation of a specific subset of cytoplasmic mRNAs (amongst which is the nuclear-encoded ribulose- 1,5-bisphosphate carboxylase/oxygen- ase small subunit, S SU) is inhibited in jasmonate- treated barley leaf segments [56]. This raises a question about the functions these mRNAs have in common, and about the special features of jasmonate-inducible mRNAs that may enable their sustained high translation in jasmonate- treated leaf segments. A detailed study of the translation of SSU mRNA, normally associated with large polysomes in control leaf segments, showed that after exposure of leaf segments to methyl jasmonate the SSU transcripts became associated with smaller polysomes, probably due to decreased chain elongation during translation [57].
An in vivo and in vitro inhibition of translation of the plastid-encoded mRNA for ribulose-l,5- bisphosphate carboxylase/oxygenase large sub- unit (LSU) was also observed in jasmonate- treated leaf segments. In this case primer extension analysis showed that transcripts syn- thesized in the presence ofjasmonate had under- gone alternative processing of the 5' end result-
1427
ing in larger transcripts [58]. These alternative transcripts contain a 35 base sequence between - 94 and - 59 which is highly complementary to the extreme 3' end of 16S rRNA. Formation of an intermolecular complex between the LSU RNA and ribosomal RNA would be a possible hin- drance to translation of this message [ 58 ]. While the physiological importance of these effects is still unclear, these studies have the potential to yield much new information on translational con- trol in a eukaryotic system. They have also em- phasized the potential diversity of the effects of jasmonate on gene expression in plants.
Experiments to investigate the early stages of jasmonate interaction with the plant cell are needed to answer questions concerning the nature ofjasmonate receptors. Of the many approaches available, mutagenesis to produce jasmonate- insensitive plants holds promise. Staswick et aL
[ 67 ] have described an Arabidopsis mutant which fails to respond to jasmonate-inhibition of root growth or jasmonate-induction of proteins simi- lar to vegetative storage proteins. It is hoped that the generation of mutants such as these will pro- vide the necessary backgrounds from which to isolate genes involved in jasmonate signal trans- duction.
At this stage it should be noted that jasmonates are not the only plant-derived fatty acid signals known to regulate developmental phenomena in plants. Traumatins will be discussed below, al- though it is far from clear how widespread their roles are. Another fatty acid derivative, phaseolic acid (Fig. 1C) was characterized in the 1960s [55]. This trihydroxydodecanoic acid, purified from bean seeds, was shown to stimulate the growth of pea stem segments as well as induce a-amylase synthesis in barley endosperm [55]. Exogenous phaseolic acid also retarded senes- cence in barley leaf segments. This compound thus deserves further investigation, particularly in light of the fact that its regulation of c~-amylase genes provides a sensitive assay for the barley compound. Several other reports of the synthesis of prostaglandin-like compounds in plants have appeared [ e.g. 48 ] although their activity in plants, if any, was not reported.
[191]
1428
Cellular levels of jasmonate in relation to gene expression
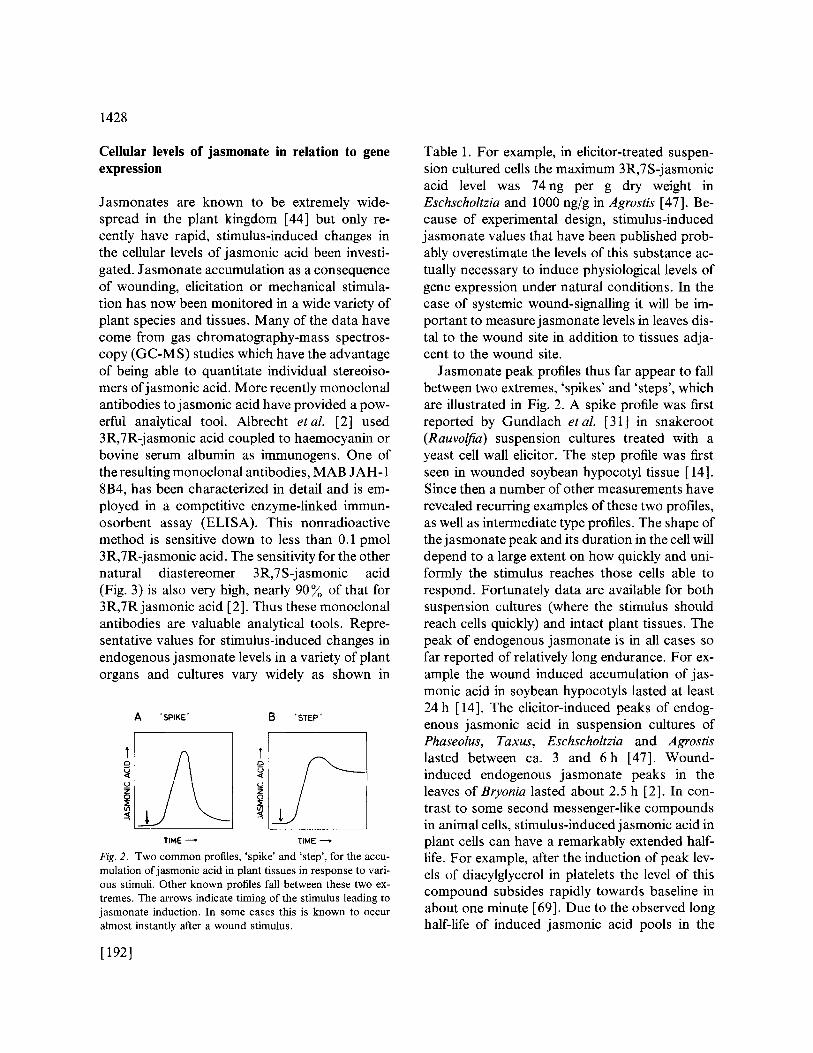
Jasmonates are known to be extremely wide- spread in the plant kingdom [44] but only re- cently have rapid, stimulus-induced changes in the cellular levels of jasmonic acid been investi- gated. Jasmonate accumulation as a consequence of wounding, elicitation or mechanical stimula- tion has now been monitored in a wide variety of plant species and tissues. Many of the data have come from gas chromatography-mass spectros- copy (GC-MS) studies which have the advantage of being able to quantitate individual stereoiso- mers ofjasmonic acid. More recently monoclonal antibodies to j asmonic acid have provided a pow- erful analytical tool. Albrecht etal. [2] used 3R,7R-jasmonic acid coupled to haemocyanin or bovine serum albumin as immunogens. One of the resulting monoclonal antibodies, MAB JAH- 1 8B4, has been characterized in detail and is em- ployed in a competitive enzyme-linked immun- osorbent assay (ELISA). This nonradioactive method is sensitive down to less than 0.1 pmol 3R,7R-jasmonic acid. The sensitivity for the other natural diastereomer 3R,7S-jasmonic acid (Fig. 3) is also very high, nearly 90 ~o of that for 3R,7R j asmonic acid [2]. Thus these monoclonal antibodies are valuable analytical tools. Repre- sentative values for stimulus-induced changes in endogenous jasmonate levels in a variety of plant organs and cultures vary widely as shown in
A 'SPIKE' B 'STEP'
o
z 0 u')
T
(J
TIME ----" TIME -'-,"
Fig. 2. Two c o m m o n profiles, ' spike ' and 's tep ' , for the accu- mulat ion o f j a s m o n i c acid in plant t i ssues in response to vari- ous stimuli. Other known profiles fall between these two ex- t remes. The arrows indicate t iming of the s t imulus leading to j a s m o n a t e induction. In some cases this is known to occur a lmost instant ly after a wound st imulus.
[192]
Table 1. For example, in elicitor-treated suspen- sion cultured cells the maximum 3R,7S-jasmonic acid level was 74ng per g dry weight in Eschscholtzia and 1000 ng/g in Agrostis [47]. Be- cause of experimental design, stimulus-induced jasmonate values that have been published prob- ably overestimate the levels of this substance ac- tually necessary to induce physiological levels of gene expression under natural conditions. In the case of systemic wound-signalling it will be im- portant to measure jasmonate levels in leaves dis- tal to the wound site in addition to tissues adja- cent to the wound site.
Jasmonate peak profiles thus far appear to fall between two extremes, 'spikes' and 'steps', which are illustrated in Fig. 2. A spike profile was first reported by Gundlach et al. [31 ] in snakeroot (Rauvolfia) suspension cultures treated with a yeast cell wall elicitor. The step profile was first seen in wounded soybean hypocotyl tissue [14]. Since then a number of other measurements have revealed recurring examples of these two profiles, as well as intermediate type profiles. The shape of the j asmonate peak and its duration in the cell will depend to a large extent on how quickly and uni- formly the stimulus reaches those cells able to respond. Fortunately data are available for both suspension cultures (where the stimulus should reach cells quickly) and intact plant tissues. The peak of endogenous jasmonate is in all cases so far reported of relatively long endurance. For ex- ample the wound induced accumulation of jas- monic acid in soybean hypocotyls lasted at least 24 h [14]. The elicitor-induced peaks of endog- enous jasmonic acid in suspension cultures of Phaseolus, Taxus, Eschscholtzia and Agrostis lasted between ca. 3 and 6h [47]. Wound- induced endogenous jasmonate peaks in the leaves of Bryonia lasted about 2.5 h [2]. In con- trast to some second messenger-like compounds in animal cells, stimulus-induced jasmonic acid in plant cells can have a remarkably extended half- life. For example, after the induction of peak lev- els of diacylglycerol in platelets the level of this compound subsides rapidly towards baseline in about one minute [69]. Due to the observed long half-life of induced jasmonic acid pools in the

plant cell, as well as other features of jasmonate signalling, this molecule should not be regarded as a classical second messenger. Jasmonates seem to have more in common with other forms of biological mediators such as prostaglandins, and may share features in common with paracrine signals [25]. Bearing in mind that prolonged ex- posure to elevated jasmonate levels can cause growth inhibition, it would be interesting to know if the jasmonate step profiles are associated with decreased or inhibited cell growth. In other words, are particular jasmonate profile shapes associ- ated with particular responses?
A similar question concerning the biological activities of various jasmonic acid stereoisomers has been addressed. Newly synthesized jasmonic acid epimerises to form two diastereomers at or near thermodynamic equilibrium (Fig. 3). This epimerization takes place in vivo converting 3R,7S-jasmonic acid to the 3R,7R-diastereomer [47]. Different physiological responses are in- duced to different levels by the naturally occurring jasmonate isomers. Koda eta l . [40] examined the effects of the four jasmonate stereoisomers (see review by Sembdner and Parthier [62]) on four different physiological responses and found that each response differed in its sensitivity to a given stereoisomer. An extrapolation of these re- suits might mean that individual genes are differ- entially sensitive to jasmonate stereoisomers. Epimerisation of jasmonic acid within the cell (Fig. 3) may thus alter its biological activity or affects its turnover or conjugation within the cell.
Other ways that the cell could remove free jas- monate from the cellular pool include the forma- tion of conjugates either with the carboxyl group
A B
0 0
Fig. 3. The epimerization of biosynthetic 3R,7S-jasmonic acid (A) to 3R,7R-jasmonic acid (B) which occurs within plant cells and which could, in part, regulate the life-time of the active signal within the cell.
1429
by esterification to amino acids or by the addition of glucose to hydroxyl groups introduced at car- bons 11 or 12 [45,46]. Conjugates of dihydro- jasmonate with a variety of other molecules, es- pecially the hydrophobic amino acids leucine, isoleucine and valine, have been characterized [45]. The possibility that simular conjugates arise from the proteolysis of polypeptides linked to jas- monic acid should be considered, and could per- haps be addressed using the monoclonal anti- bodies to jasmonic acid [2] which would be expected to have a high affinity for jasmonate- protein conjugates.
Inhibition of the octadecanoid signal pathway
Biochemical experiments to investigate the need for jasmonate biosynthesis in signal transduction have been conducted and cumulative evidence suggests that induced jasmonate is, at least in part, synthesized de novo in response to a stimu- lus and not simply released from 'storage' conju- gates or pools. Ideally, disruption of the jas- monate biosynthetic pathway could be achieved with a genetic method, for example by expressing the mRNA for a jasmonate synthesizing enzyme in antisense. Until this is reported the available tools include a variety of inhibitors of fatty acid metabolism in plants, some of which have been borrowed from animal research. Experiments in- volving the use oflipoxygenase inhibitors are often difficult and it is possible that plant lipoxygenases involved in signal generation in plants differ somewhat from the animal enzymes for which the inhibitors were originally developed. Several such compounds have been employed to block the syn- thesis of wound-inducible proteinase inhibitors and vegetative storage proteins in leaves. These inhibitors include aspirin [ 18], and ibuprofen, N-propyl gallate, salicylhydroxamic acid and phe- nylbutazone [65]. A recently introduced inhibi- tor, ursolic acid, was shown to reduce endog- enous levels of jasmonic acid in barley leaf segments as well as to inhibit the expression of a number of genes encoding jasmonate-inducible proteins [73]. This compound may prove to
[193]

1430
be a broadly useful inhibitor. Results strongly supporting the role ofjasmonic acid in the wound induction of proteinase inhibitor gene expression were recently published. Pel"la-Cortes et al. [52] showed that aspirin blocked the accumulation of endogenous jasmonic acid in wounded leaves of tomato. Concomitant with reduced levels ofjas- monate was the reduced accumulation of the messenger RNA for proteinase inhibitor II as well as the mRNAs for threonine dehydrase and cathepsin D-inhibitor. The proposed site of ac- tion of aspirin was on hydroperoxide dehydrase.
Recently a different type of inhibitor has emerged: sodium diethyldithiocarbamic acid (DIECA) was found to partially block the wound and systemin induction of proteinase inhibitors I and II in tomato leaves. A series of attempts to identify the site of action of DIECA led to the observations that this compound is a weak in- hibitor of tomato leaf lipoxygenase activity but can react directly with 13(S)-hydroperoxylino- lenic acid in vitro to form a compound with prop- erties expected of 13-hydroxylinolenic acid (Farmer, Caldelari, Pearce, Walker-Simmons and Ryan, Plant Physiol, in press). Thus this inhibi- tor has at least two potential sites of action and this compound, or a similar less toxic derative, might prove useful since its action at the level of the lipoxygenase products (fatty acid hydroper- oxides) would bypass the need to inhibit lipoxy- genase directly. It must be noted that the appar- ent toxicity of DIECA might limit its utility as an inhibitor but some tentative conclusions have been drawn from its use in the tomato/proteinase inhibitor system. The most important conclusion from this study was that, since DIECA blocked the systemin-induced expression of proteinase in- hibitor genes by interruption of fatty acid meta- bolism, the signal order systemin --, jasmonate gene is likely. Further experiments are necessary to test this proposed signal order.
Another way to inhibit j asmonate synthesis and thus to test various models for the regulation of wound-inducible gene expression would be to generate mutants incapable of producing wild- type levels of the jasmonate. These tests will be possible using Arabidopsis mutants deficient in the
[1941
jasmonate precursor linolenic acid (M. McConn and J. Browse, 1993, Abstract B3, National Plant Lipid Cooperative Symposium, Minneapolis; J. Browse, personal communication) and it will be interesting to study j asmonate levels as well as the expression of wound-inducible genes in these plants.
The traumatin family
Fatty acid hydroperoxides (including the 13-(S)- hydroperoxide of linolenic acid, which is a pre- cursor to jasmonic acid) can be metabolized by lytic reactions to yield two aldehydes (reviewed in [28]) which are candidates for further enzymic or nonenzymic oxidation to carboxylic acids. One of these molecules, dicarboxylic acid called trau- matic acid, was isolated directly from the meso- carp of wounded beans and was shown to en- hance cell proliferation at the wound site [20]. Traumatic acid has not received the attention it deserves because its effects appear to be limited to one or a few bean cultivars, and because it is likely that the traumatic acid originally identified by English et al. [20] arose through the oxidation of an unstable alkenal, traumatin [77]. It is this dicarboxylic acid that is commercially available and is often used in place of the less stable oxoacid which may have a higher biological activity [77]. For example, traumatic acid causes abscission of cotyledon petioles in cotton explants, and this effect is reduced by auxin treatment [68]. Several other studies reporting the effects of traumatin on plants or pathogens have employed traumatic acid and in some cases it is not clear if the authors were aware of the difference between these mol- ecules. To further complicate the matter, trauma- tin itself arises through the rearrangements of an isomeric compound (9Z)12-oxo-dodecenoic acid (A9-ODA) [34]. This is often the primary me- tabolite produced from 18:2 or 18:3 fatty acid hydroperoxides by plant leaf extracts. For clari- fication the probable relationship between mem- bers of the traumatin family is outlined in Fig. 4. Further studies are needed to clarify the biosyn- thesis and biological activities of traumatin, a 9-

18:2, 18:3
~L tipoxygenose
A. I hydroperoxide lyase O H C ~ C O O H
I isornerisation
B.
autooxidation
C, HOOC ~ c O O H
Fig. 4. Traumatin and its dodecanoid relatives. A. 12-oxo- 9(Z)-dodecenoic acid (A9-ODA). B. Traumatin, 12-oxo- 10(E)-dodecenoic acid (A 10-ODA). C. Traumatic acid, 10(E) dodeca-1,12-dicarboxylic acid. Product A originates from the action of hydroperoxide lyase on 13-hydroperoxyoctade- canoids. This compound isomerizes in the presence of some plant extracts to B, 'traumatin'. Compound C, traumatic acid, is an oxidized derivative of B and was the structure originally characterized by Bonner 's group in the 1930s as a wound hormone. The possibility that product B ' traumatin' is pro- duced directly in some tissues is not excluded.
ODA and traumatic acid. It is already known that the levels of traumatin increase in wounded bean pod tissue [77] and it is interesting to note that the common precursor for jasmonate and traumatin, 13(S)-hydroperoxylinolenic acid, gives rise to two wound-inducible signals, one of which (jasmonate) is a defence signal and the other (traumatin) a wound-healing signal.
Traumatin is only one of a potentially larger class of alkenals that can be generated by the lytic action of hydroperoxide lyase on various fatty acid hydroperoxides. Since a potentially larger number of fatty acid hydroperoxides could be produced by the action of lipoxygenase on 1,4- Z,Z-pentadiene fatty acids, an even larger family of aldehydes could be generated in planta. The aldehyde function is labile and the double bond proximal to the aldehyde group is subject to cis- trans isomerization (e.g. Fig. 4). Many of these molecules are volatile and might rapidly diffuse through tissues and into the air. Examples of
1431
volatile and toxic alkenals are discussed in the next section.
Volatile and cytotoxic fatty acid oxidation prod- ucts
Amongst the most toxic natural products of fatty acid oxidation are the unsaturated aldehydes (alkenals) and their more highly oxidized deriva- tives. Plants have long been known to produce large quantities of alkenals. One of the earliest papers on alkenal production highlighted both the wide-spread occurrence of this phenomenon in the plant kingdom and the potential toxicity of these molecules [60]. The antimicrobial proper- ties of several unsaturated aldehydes, in particu- lar (2E)-hexenal, are likely to play important roles in plant defence against fungi [42, 71], bacteria [15, 17] and arthropods [35,42]. This volatile compound is transiently released into the air upon wounding leaves [60] and during the hypersensi- tive response of Phaseolus leaves to infection with Pseudomonas syringae pv. phaseolicola [ 15 ]. The volatility of (2E)-hexenal may contribute to its rapid diffusion through damaged or infected tis- sues. In addition to their toxicity, volatile alkenals such as (2E)-hexenal are often floral scent com- ponents [39].
A recent discovery of interest in plants is the production of (2E)-4-hydroxynonenal from (3Z)- nonenal by broad bean Vicia faba seed extracts [ 29]. (2E)-4-hydroxynonenal is a highly cytotoxic and genotoxic fatty acid derivative that can be produced in animal cells [21 ]. The possibility that this compound could act in defence against fungi has been investigated. (2E)-4-Hydroxynonenal was found to be an inhibitor of the growth of Colletotrichun truncatum and Scherotium rolfsii when added to the fungal growth medium at lev- els as low as 0.3 #mol per ml medium [71]. The effects of (2E)-4-hydroxynonenal on plant cells were not examined but judging by its toxicity to animal cells [21], this compound could poten- tially destroy plant cells at very low concentra- tions. Could unsaturated aldehydes or their de- rivatives be the cytotoxins or signals that kill plant
[1951

1432
cells during the hypersensitive response? If so, how is the generation of these toxic molecules regulated?
Methyl jasmonate vapour is a powerful signal for the induction of proteinase inhibitor gene ex- pression in the leaves of several species of plants in the Solanaceae (e.g. tomato, potato, tobacco [23, 26]) and Fabaceae (e.g. alfalfa [26]). The low levels of this vapour necessary to trigger pro- teinase inhibitor gene expression, as well as to cause certain physiological responses like tendril coiling in Bryonia [22], suggest that volatile me- thyl jasmonate may play physiological roles. This compound was shown to be present on the leaf surfaces of sagebrush (Artemisia tridentata) in suf- ficient quantity to induce proteinase inhibitor ac- cumulation in the leaves of neighbouring tomato plants under laboratory conditions [23]. The pos- sible consequences of plants releasing this highly active signal into the air have not been investi- gated adequately, and the case of A. tridentata remains intriguing. This plant, North America's most abundant shrub (National Geographic, January 1989), could potentially affect neighbour- ing plants in a variety of ways. For example, it could induce defence responses or affect other aspects of physiology including growth inhibition or senescence. One possibility is that methyl jas- monate could inhibit the growth of competing plants, particularly climbing plants which are rarely observed on Great Basin A. tridentata populations (E. Farmer, unpublished; E.D. McArthur, personal communication).
Although a number of volatile fatty acid de- rivatives are known to be released by leaves there are still few examples of their role or utilization in nature. In contrast, a great many fatty acid- derived floral scents are known [39]. It is con- ceivable that the great number of fatty acid- derived floral scents may be paralleled in the production of volatile fatty acid derivatives by healthy or wounded leaves. Bearing in mind the volatile nature of many fatty acid derivatives, it will be interesting to reappraise the distribution and function of intercellular air spaces in plant tissues in connection with plant development and pathogenesis.
[196]
Fatty acid signalling between plants and micro- organisms
The use of jasmonate as a spray for the protec- tion of plants from pests and pathogens has been investigated in a number of systems. The idea behind these experiments was that jasmonate would induce native plant defences thus protect- ing the crop. Cohen et al. [ 13] observed that the application of jasmonic acid to potato plants caused local and systemic protection from the late blight fungus Phytophthora infestans. Although jasmonic acid was found to have a direct effect on the mode of germination ofP. infestans sporangia, it is likely that the protective effect of jasmonic acid was due to the induction of antifungal de- fences since leaves distal to those sprayed with jasmonic acid were protected from late blight. The nature of the induced systemic defences was not established, but the authors speculated that an oxidative defence involving lipoxygenase acti- vation may have been responsible for the protec- tive effect ofjasmonate.
A recent surprise was the observation that jas- monic acid, apart from its signalling effect in plants, may also act as a signal in fungi. Barley (Hordeum) plants sprayed with jasmonic acid so- lutions were found to be protected from infection by the pathogenic fungus Erysiphe graminis f.sp. hordei [61 ]. Several jasmonate-inducible proteins accumulated when barley was sprayed with jas- monic acid, however these proteins were found to have no antifungal activity against Erysiphe. Fur- thermore, pretreating plants with cordycepin (a transcription inhibitor) had no effect on the ability of jasmonic acid to protect barley against E. graminis. These first results suggested an un- expected and interesting result, that inhibition of pathogenesis was due, at least in part, to the di- rect effect of jasmonate on the life cycle of Ery- siphe. Jasmonic acid was found to block the differentiation of appressoria (thus inhibiting pen- etration of the fungus) at concentrations which were not inhibitory to mycelial growth. These re- sults highlight the potential complexity of the role of jasmonate in the plant-fungal interaction. However Schweizer etal. [61] caution that the

levels ofj asmonic acid which accumulate in planta are probably too low to have a direct effect on fungal differentiation.
The mycelial membranes of Phytophthora in- festans contain C20 fatty acids (including arachi- donic acid; Fig. 5B) which are powerful inducers of phytoalexin accumulation and browning in po- tato tuber tissue [6, 7]. Experiments suggest that lipoxygenase is necessary for defence gene induc- tion by arachidonic acid [54]. However, the structures of the biologically active arachidonate metabolites are not yet known and the possibil- ity remains that arachidonate can affect defence
A. D. 0 % ^ ^ . ,
I 13-GIcN, [~ -.GIcNAc]n - B -GIc NAc
I Ac
3. OH
E. OH
C.
H OH OH
Fig. 5. Fatty acid derivatives involved in signalling between microorganisms and plants. A. Jasmonic acid: not only a sig- nal molecule in plants, this fatty acid can inhibit appressorium formation in the fungus Erysiphe without inhibiting mycelial growth. B. Arachidonic acid, a component of the mycelial membranes of Phytophthora infestans, regulates a number of defence-related genes in potato tuber. C. 9,10,18-trihydroxy- octadecanoic acid, a component of plant cutin, is a powerful inducer of cutinase in the germinating spores of several patho- genic fungi including Fusarium solani pisi (Nectria hematococca). D. Nod Rlv-V(Ac,18:4) a decorated fatty acid produced by Rhizobium leguminosarum bv. viciae. This molecule causes root hair curling and cortical root cell division in pea. Modification of the fatty acid changes the ability of the signal to cause these effects in a given plant species. E. Syringolide 1, produced by bacteria (e.g. Pseudomonas syringae pv. tomato) expressing the avirulence gene avrD. Syringolide causes cell death only in plants expressing a complementary resistance gene, Rpg4, and is thus a specificity determinant in pathogenesis.
1433
signal transduction by other mechanisms. Ricker and Bostock [59] identified several hydroperox- yeicosanoic acids generated by the application of arachidonic acid to potato tubers. None of these compounds including 5-hydroperoxyeicosatet- raenoic acid (5-HPETE) had the ability to induce sesquiterpenoid phytoalexins. In contrast, Casto- ria et al. [10] reported that 5-HPETE could in- duce sesquiterpenoid phytoalexin accumulation in potato tuber tissue. If arachidonic acid hydro- peroxides are able to signal phytoalexin accumu- lation, it will be interesting to see if the biologi- cally active compound(s) is the fatty acid hydroperoxide or a downstream metabolite(s).
Recent studies on the regulation of a small fam- ily of HMG-CoA reductase (hmg) genes in potato tissues have revealed interesting patterns of dif- ferential gene regulation by fatty acids [ 11, 12]. The gene hmgl is wound-inducible and implicated in the synthesis of steroid glycoalkaloids which may be necessary for the restoration of membrane function during wound-healing. The wound- inducible hmgl gene is powerfully upregulated by jasmonic acid, but expression of this gene is sup- pressed by arachidonic acid [12]. This provides an example of a gene that appears to be differ- entially regulated by two different fatty acid sig- nals of plant and fungal origin respectively. The genes hmg2 and hmg3 are involved in the biosyn- thesis of sesquiterpenoid phytoalexins and are, in contrast to hmgl, powerfully induced by exogen- ous arachidonic acid.
Fatty acid-derived signals involved in the in- teraction between plants and pathogens do not always derive from the latter. Plant cutin compo- nents including 10,16-dihydroxyhexadecanoic acid and 9,10,18-trihydroxyoctadecanoic acid are powerful upregulators of cutinase transcription in nuclei from the phytopathogenic fungus Fusarium solanipisi (Nectria hematococca [ 53 ]). Further evi- dence for a biological role for fatty acid polyhy- droxides came from the observation that the mycelia of several phytopathogenic fungi lyse in the presence of 8-hydroxylinoleic acid. This com- pound is made by the soil fungus Laetisaria ar- valis, which is a disease suppresser in some crops [9]. More recently a detailed study oflinoleic acid
[1971

1434
metabolism by the wheat take-all fungus Gaeum- annomyces graminis has revealed the production of an interesting array of oxygenated fatty acids including (8R)-hydroxylinoleic acid and 7,8- dihydroxylinoleic acid [8]. (8R)-Hydroxylinoleic acid was shown to be a sporulation regulator in Aspergillus nidulans [43]. Unsaturated monohy- droxy fatty acids are also known to be antifungal [37]. It may only be a matter of time before these interesting molecules are found to have signalling roles in the plant.
The recent discoveries that several fatty acid- containing signal molecules can play governing roles in specificity determination and recognition phenomena has considerably broadened our un- derstanding of plant-microorganism interactions. Nodulation (Nod) factors secreted by Rhizobium species can be regarded as fatty acids which are decorated through N-acetyl linkage to glu- cosamine oligomers. These complex signal mol- ecules induce organogenesis leading to nodule formation in compatible legume species. The fatty acids found in Nod factors often contain trans (E) double bonds (e.g. Fig. 5D), and when con- jugated could be potentially reactive. The fatty acid substituents vary between species and strains of Rhizobium and help to determine host speci- ficity [27, 64]. This was probably the first indica- tion that fatty acids could help control specificity in a recognition response. Increasing evidence suggests that Nod-like signals are made by plants themselves and may play a natural role in plant development [ 16].
Other fatty acid derivatives involved in recog- nition phenomena are syringolides, a small fam- ily of compounds thought to result from the con- jugation of/~-keto fatty acid and xylulose [38]. Syringolides are produced by gram negative bac- teria carrying the avirulence gene avrD originally isolated from Pseudomonas syringae pv. tomato. Syringolides (Fig. 5E) cause cell death only in plants containing the cognate resistance gene Rpg4 [38]. It is not yet known why some bacte- ria make syringolides, and the possibility that they are signals for the bacterium itself cannot be ig- nored.
[1981
Conclusion
The number of biologically active fatty acids known in plants is increasing, although detailed information exists only for some members of the jasmonate family, chiefly jasmonic acid. This compound is a regulator of wound-inducible genes and its intracellular levels are transiently elevated in response to wounding. It can also regulate other defence-related genes. Members of the jasmonate family appear to play roles in a number of developmental responses including tu- berization in potato, and tendril coiling, and in inducing the synthesis of secondary plant com- pounds. Jasmonate is known to occur in plants, fungi and insects; perhaps it is only a matter of time until it is found in higher animals. Increas- ingly the literature suggests that new fatty acid signals in plants will be discovered, although much remains to be learnt of biologically active fatty acids discovered many years ago, for ex- ample, traumatin and phaseolic acid. In animal cells fatty acid-derived signals are numerous with tens of biologically active fatty acid-derived mol- ecules having been characterized and more await- ing discovery. Even considering the greater cel- lular complexity of higher animals, it is likely that more fatty acid signals exist in plants. The pos- sibilities become more exciting when one consid- ers the relationship between peptides and fatty acids. A large number of peptide signals in ani- mals influence cellular lipid metabolism. If there are more peptide hormones in plants, are there more fatty acid signals? The recent discoveries of interkingdom fatty acid signalling have illumi- nated a wide variety of roles for these molecules, including roles in recognition phenomena. In the next few years more fatty acid derivatives involved in the interaction of plants and other organisms will probably be found and attention will turn increasingly towards receptors for these mol- ecules.
Acknowledgements
The following people made data available prior to publication or otherwise assisted in providing

useful information: K. Apel, R. Bostock, I.D. Brodowsky, D. Caldelari, D. Hildebrand, Y. Koda, J. Mullet, B. Parthier, U. Rtlegg, P. Schweizer, W. Smith, B. Vick, C. Wasternack, E. Weiler, M.H. Zenk. I thank H. Berttoud for preparation of figures.
References
1. Aerts RJ, Gisi D, De Carolis E, De Luca V, Baumann TW: Methyl jasmonate vapor increases the developmen- tally controlled synthesis of alkaloids in Catharanthus and Cinchona seedlings. Plant J 5:635-645 (1994).
2. Albrecht T, Kehlen A, Stahl K, KnOfel H-D, Sembdner G, Weiler EW: Quantification of rapid, transient increases in jasmonic acid in wounded plants using a monoclonal antibody. Planta 191:86-94 (1993).
3. Anderson JM: Membrane-derived fatty acids as precur- sors to second messengers. In: Boss WF, Morre DJ (eds) Second Messengers in Plant Growth and Development, pp. 181-182. Liss, New York (1989).
4. Becker W, Apel K: Differences in gene expression be- tween natural and artificially induced leaf senescence. Planta 189:74-79 (1993).
5. Bolter CJ: Methyl jasmonate induces papain inhibitor(s) in tomato leaves. Plant Physiol 103:1347-1352 (1993).
6. Bostock RM, Kuc JA, Laine RA: Eicosapentaenoic and arachidonic acids from Phytophthora infestans elicit fun- gitoxic sesquiterpenes in the potato. Science 212:67-69 (1981).
7. Bostock RM, Yamamoto H, Choi D, Ricker KE, Ward BL: Rapid stimulation of 5-1ipoxygenase activity in potato by the fungal elicitor arachidonic acid. Plant Physiol 100: 1448-1456 (1992).
8. Brodowsky IR, Hamberg M, Oliw EH: A linoleic acid 8(R)-dioxygenase and hydroperoxide isomerase of the fungus Gaeurnannomyces graminis: Biosynthesis of 8(R)- hydroxylinoleic acid and 7(S),8(S)-dihydroxylinoleic acid from 8(R)-hydroperoxylinoleic acid. J Biol Chem 267: 14738-14745 (1992).
9. Bowers WS, Hoch HC, Evans PH, Katayama M: Thal- lophytic allelopathy: isolation and identification of laeti- saric acid. Science 232:105-106 (1986).
10. Castoria R, Fanelli C, Fabbri AA, Pasi S: Metabolism of arachidonic acid involved in its eliciting activity in potato tuber. Physiol Mol Plant Path 41:127-137 (1992).
11. Choi D, Ward BL, Bostock RM: Differential induction and suppression of potato 3-hydroxy-3-methylglutaryl co- enzyme A reductase genes in response to Phytophthora infestans and to its elicitor arachidonic acid. Plant Cell 4: 1333-1344 (1992).
12. Choi D, Bostock RM, Avdiushko S, Hildebrand DF: Lipid-derived signals that discriminate wound- and
1435
pathogen-responsive isoprenoid pathways in plants: methyl-jasmonate and the fungal elicitor arachidonic acid induce different HMG-CoA reductase genes and antimi- crobial isoprenoids in Solanum tuberosum L. Proc Natl Acad Sci USA 91:2329-2333 (1994).
13. Cohen Y, Gisi U, Mosinger E: Local and systemic pro- tection against Phytophthora infestans induced in potato and tomato plants byjasmonic acid and jasmonic-methyl- ester. Phytopath 83:1054-1062 (1993).
14. Creelman RA, Tierney ML, Mullet JE: Jasmonic acid/ methyl jasmonate accumulate in wounded soybean hy- pocotyls and modulate wound gene expression. Proc Natl Acad Sci USA 89:4938-4941 (1992).
15. Croft KPC, J~ittner F, Slusarenko AJ: Volatile products of the lipoxygenase pathway evolved from Phaseolus vul- garis (L.) leaves inoculated with Pseudomonas syringae pv. phaseolicola. Plant Physiol 101:13-24 (1993).
16. de Jong AJ, Heidstra R, Spaink HP, Hartog MV, Meijer EA, Hendriks T, Lo Schiavo F, Terzi M, Bisseling T, van Kammen A, de Vries SC: Rhizobium lipooligosaccharides rescue a carrot somatic embryo mutant. Plant Cell 6: 615-620 (1993)
17. Deng W, Hamilton-Kemp TR, Nelson MT, Andersen RA, Collins GB, Hildebrand DF: Effects of six-carbon aldehydes and alcohols on bacterial proliferation. J Agric Food Chem 41:506-510 (1993).
18. Doherty HM, Selvendran RR, Bowles DJ: The wound response of tomato plants can be inhibited by aspirin and related hydroxybenzoic acids. Physiol Mol Plant Path 32: 377-384 (1988).
19. Eckelkamp K, Ehmann B, Schopfer P: Wound-induced systemic accumulation of a transcript coding for a Bowman-Birk trypsin inhibitor-related protein in maize (Zea rnays L.) seedlings. FEBS Lettt 323:73-76 (1993).
20. English J, Bonner J, Haagen-Smit AJ: Structure and synthesis of a plant wound hormone. Science 90:329 (1939).
21. Esterbauer H, Schaur RJ, Zollner H: Chemistry and bio- chemistry of 4-hydroxynonenai, malonaldehyde and re- lated aldehydes. Free Rad Biol Med 11:81-128 (1988).
22. Falkenstein E, Groth B, Mith/ffer A, Weiler EW: Methyl jasmonate and ~-linolenic acid are potent inducers of tendril coiling. Planta 185:316-322 (1991).
23. Farmer EE, Ryan CA: Interplant communication: air- borne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc Natl Acad Sci USA 87: 7713-7716 (1990).
24. Farmer EE, Ryan CA: Octadecanoid-derived signals in plants. Trends Cell Biol 2:236-241 (1992).
25. Farmer EE, Ryan CA: Octadecanoid precursors ofjas- monic acid activate the synthesis of wound-inducible pro- teinase inhibitors. Plant Cell 4:129-134 (1992).
26. Farmer EE, Johnson RR, Ryan CA: Regulation of ex- pression of proteinase inhibitor genes by methyl jas- monate and jasmonic acid. Plant Physiol 98:995-1002 (1992).
[1991

1436
27. Fisher RF, Long SR: Rhizobium-plant signal exchange. Nature 357:655-659 (1992).
28. Gardner HW: Recent investigations into the lipoxygenase pathway of plants. Biochim Biophys Acta 1084:221-239 (1991).
29. Gardner HW, Hamberg M: Oxygenation of (3Z)-nonenal to (2E)-4-hydroxy-2-nonenal in the broad bean (Viciafaba L.). J Biol Chem 268:6971-6977 (1993).
30. Green TR, Ryan CA: Wound-induced proteinase inhibi- tor in plant leaves: a possible defence mechanism against insects. Science 175:776-77 (1972).
31. Gundlach H, MOiler MJ, Kutchan TM, Zenk MH: Jas- monic acid is a signal transducer in elicitor-induced plant cell cultures. Proc Natl Acad Sci USA 89:2389-2393 (1992).
32. Haagen-Smit AJ, Leach WD, Bergren WR: The estima- tion, isolation and identification of auxins in plant mate- rials. Am J Bot 29:500-506 (1942).
33. Hamburg M, Gardner HW: Oxylipin pathway to jas- monates: biochemistry and biological significance. Bio- chim Biophys Acta 1165:1-18 (1992).
34. Hatanaka A, Kajiwara T, Sekija J: Biosynthetic pathway of C6-aldehyde formation from linolenic acid in green leaves. Chem Phys Lipids 44:341-361 (1987).
35. Hildebrand DF, Brown GC, Jackson DM, Hamilton- Kemp TR: Effects of some leaf-emitted volatile com- pounds on aphid population increase. J Chem Ecol 19: 1875-1887 (1993).
36. Kaiser I, Engelberth J, Groth B, Weiler EW: Touch and methyl j asmonate induced lignification in tendrils of Bryo- nia dioica Jacy. Bot Acta 107:24-29 (1994).
37. Kato T, Yamaguchi Y, Hirano T, Yokohama T, Uyehara T, Yamanaka S, Harada T: Unsaturated hydroxyfatty acids, the self defence substances in rice plant against rice blast disease. Chem Lett: 409-412 (1984).
38. Keen NT, Sims J J, Midland S, Yoder M, Jurnak F, Shen H, Boyd C, Yucul I, Lorang J, Murillo J: Determinants of specificity in the interaction of plants with bacterial pathogens. In: Nester EW, Verma PS (eds) Advances in Molecular Genetics and Plant-Microbe Interactions, pp. 211-220. Kluwer Academic Publishers, Dordrecht (1993).
39. Knudsen JT, Tollsten L, Bergstrom G: Floral scents - a checklist of volatile compounds isolated by headspace techniques. Phytochemistry 33:253-280 (1993).
40. Koda Y, Kikuta Y, Kitahara T, Nishi T, Mori K: Com- parisons of various biological activities of stereoisomers of methyl jasmonate. Phytochemistry 31:1111-1114 (1992).
41. Koda Y: The role of jasmonic acid and related com- pounds in the regulation of plant development. Int Rev Cytol 135:155-199 (1992).
42. Lyr H, Banasiak L: Alkenals, volatile defence substances in plants, their properties and activities. Acta Phytopath Acad Sci Hung 18:3-12 (1983).
43. Mazur P, Nakanishi K, E1-Zayat AAE, Champe SP:
[2001
Structure and synthesis of sporogenic psi factors from Aspergillus nidulans. J Chem Soc Chem Commun 20: 1486-1487 (1991).
44. Meyer A, Miersch O, BtRtner C, Dathe W, Sembdner G: Occurrence of the plant growth regulator jasmonic acid in plants. J Growth Regul 3 :1 -8 (1984).
45. Meyer A, Schmidt J, Gross D, Jensen E, Rudolph A, Vorkefeld S, Sembdner G: Amino acid conjugates as metabolites of the plant growth regulator dihydrojasmonic acid in barley (Hordeum vulgate). J Plant Growth Regul 10:17-25 (1991).
46. Meyer A, Gross D, Vorkfeld S, Kummer M, Schmidt J, Sembdner G, Schreiber K: Metabolism of the plant growth regulator dihydrojasmonic acid in barley shoots. Phytochemistry 28:1007-1011 (1989).
47. Mueller MJ, Brodschelm W, Spannagl E, Zenk MH: Sig- nalling in the elicitation process is mediated through the octadecanoid pathway leading to jasmonic acid. Proc Natl Acad Sci USA 90:7490-7494 (1993).
48. Panossian AG, Avetissian GM, Mantsakanian VA, Batrakov SG, Vartanian SA, Gabrielian ES, Amroyan EA: Unsaturated polyhydroxy acids having prostagladin- like activity from Bryonia alba. II. Major components. Planta Med 47:17-25 (1983).
49. Parthier B: Jasmonates: hormonal regulators or stress factors in leaf senescence? J Plant Growth Regul 9 :1 -7 (1990).
50. Parthier B: Jasmonates, new regulators of plant growth and development: many facts and few hypotheses on their actions. Bot Acta 104:446-454 (1991).
51. Pearce G, Strydom D, Johnson S, Ryan CA: A polypep- tide from tomato leaves activates the expression of pro- teinase inhibitor genes. Science 253:895-897 (1991).
52. Pefia-Cort6s H, Albrecht T, Prat S, Weiler EW, Will- mitzer L: Aspirin prevents wound-induced gene expres- sion in tomato leaves by blocking jasmonic acid biosyn- thesis. Planta 191:123-128 (1993).
53. Podila GK, Dickman MB, Kolattukudy PE: Transcrip- tional activation of a cutinase gene in isolated fungal nu- clei by plant cutin monomers. Science 242:922-925 (1988).
54. Preisig CL, Ku6 JA: Inhibition by salicylhydroxamic acid, BW755C, eicosatetraenoic acid, and disulfiram of hypersensitive resistance elicited by arachidonic acid or poly-L-lysine in potato tuber. Plant Physiol 84:891-894 (1987).
55. Redemann CT, Rappaport L, Thompson RH: Phaseolic acid: a new plant growth regulator from bean seeds. In: Wightman F, Setterfield G (eds) Biochemistry and Physi- ology of Plant Growth Regulator Substances, pp. 109- 124. Runge Press, Ottawa (1968).
56. Reinbothe S, Reinbothe C, Parthier B: Methyljasmonate represses translation initiation of a specific set of mRNAs in barley. Plant J 4:459-467 (1993).
57. Reinbothe S, Reinbothe C, Parthier B: Methyljasmonate- regulated translation of nuclear-encoded chloroplast pro-

1437
teins in barley (Hordeum vulgare L. cv. Salome). J Biol Chem 268:10606-10611 (1993).
58. Reinbothe S, Reinbothe C, Heintzen C, Seidenbecher C, Parthier B: A methyl j as monate-induced shift in the length of the 5' untranslated region impairs translation of the plastid rbcL transcript in barley. EMBO J 12:1505- 1512 (1993).
59. Ricker KE, Bostock RM: Eicosanoids in the Phytoph- thora infestans-potato interaction: fipoxygenase metabo- lism of arachidonic acid and biological activities of se- lected lipoxygenase products. Physiol Mol Plant Path 44: 65-80 (1994).
60. Schildknechkt VH, Rauch G: Die chemische Natur der Luftphytoncide von Blattpflanzen insbesondere von Ro- binia pseudoacacia Z Naturforsch 166:422-429 (1961).
61. Schweizer P, Gees R, M~Ssinger E: Effect of jasmonic acid on the interaction of barley (Hordeum vulgate L.) with powdery mildew Erysiphe graminis f.sp. hordei. Plant Physiol 102:503-511 (1993).
62. Sembdner G, Parthier B: The biochemistry and the physi- ological and molecular actions ofjasmonates. Annu Rev Plant Physiol Plant Mol Biol 44:569-589 (1993).
63. Spaink HP: Rhizobial lipo-oligosaccharides: answers and questions. Plant Mol Biol 20:977-986 (1992).
64. Spaink HP, Sheeley DM, van Brussel AAN, Glushka J, York WS, Tak T, Geiger O, Kennedy EP, Reinhold VN, Lugtenberg B J J: A novel highly unsaturated fatty acid moiety of lipo-oligosaccharide signals determines host specificity of Rhizobium. Nature 354:125-130 (1991).
65. Staswick PE, Huang J-F, Yoon R: Nitrogen and methyl jasmonate induction of soybean vegetative storage pro- tein genes. Plant Physiol 96:130-136 (1991).
66. Staswick PE: Jasmonate, genes, and frageant signals. Plant Physiol 99:804-807 (1992).
67. Staswick PE, Su W, Howell SH: Methyl jasmonate in- hibition of root growth and induction of a leaf protein are decreased in an Arabidopsis thaliana mutant. Proc Natl Acad Sci USA 89:6837-6840 (1992).
68. Strong FE, Kruitwagen E: Traumatic acid: an accelera- tor of abscission in cotton explants. Nature 215: 1380- 1381 (1967).
69. Takai Y, Kikkawa U, Kaibuchi K, Nishizuka Y: Mem- brane phospholipid metabolism and signal transduction for protein phosphorylation. Adv Cyclic Nucl Prot Phosph Res 18:119-149 (1984).
70. Ueda J, Kato J: Isolation and identification of a senescence-promoting substance from wormwood (Arte- misia absinthium L.) Plant Physiol 66:246-249 (1980).
71. Vaughn SF, Gardner HW: Lipoxygenase-derived alde- hydes inhibit fungi pathogenic on soybean. J Chem Ecol 19:2337-2345 (1993).
72. Vick BA: Oxygenated fatty acids of the lipoxygenase pathway. In: Moore TS (ed) Lipid Metabolism in Plants, pp. 167-191. CRC Press, London (1993).
73. Wasternack C, Atzorn R, Blume B, Leopold J, Parthier B: Ursolic acid inhibits synthesis of jasmonate-induced proteins in barley leaves. Phytochemistry 3 5 : 4 9 - 5 4 (1994).
74. Weiler WE: Octadecanoid-derived signalling molecules involved in touch perception in a higher plant. Bot Acta 106:2-4 (1993).
75. Weiler WE, Albrecht T, Groth B, Xia Z-Q, Luxem M, Liss H, Andert L, Spengler P: Evidence for the involve- ment ofjasmonates and their octadecanoid precursors in the tendril coiling response of Bryonia dioica. Phytochem- istry 32:591-600 (1993).
76. Yoshikara T, Omer E-SA, Koshino H, Sakamura S, Kikuta Y, Koda Y: Structure of a tuber-inducing stimu- lus from potato leaves. Agric Biol Chem 53:2835-2837 (1989).
77. Zimmerman DC, Coudron CA: Identification of trauma- tin, a wound hormone, as 12-oxo-trans-10-dodecenoic acid. Plant Physiol 63:536-541 (1979).
[201]




























