Embed Size (px)
Citation preview

FACTORS AFFECTING THE METABOLIC CONJUGATION
OF ARYLACETIC ACIDS
by
PATRICK FRANK AYODELE DIXON
a thesis submitted for the degree of Doctor of Philosophy
in the
University of London
May 1976 Department of Biochemistry
St. Mary's Hospital Medical School,
London, W2 1PG

2
ABSTRACT
The metabolic conjugation of I -naphthylacetic, diphenylacetic, and
hydratropic acid has been studied in man, some selected sub-human primates
and non-primate species, by examining the nature of the conjugates excreted
in the urine after administration of the 14
C-labelled acids. I-Naphthylacetic
acid forms glycine, taurine,glutamine and glucuronic acid conjugates but the
pattern of conjugation varied with species. Hydratropic acid is excreted
mainly as a glucuronic acid conjugate except in the cat which excretes also
the glycine and taurine conjugates. Diphenylacetic acid is excreted mainly
as its glucuronic acid conjugate irrespective of species. At low doses
1-naphthylacetic acid is excreted in the rat conjugated mainly with glycine but
at higher doses, with glucuronic acid. Dose level did not affect the metabolic
pattern of diphenylacetic and hydratropic acids in the rat which are conjugated
solely with glucuronic acid.
The pharmacokinetic behaviour of these acids and phenylacetic acid
was examined in the rabbit; and 1 -naphthylacetic, diphenylacetic and hydratropic
acids which are conjugated mainly with glucuronic acid, were shown to have a
low blood clearance, a low biological half-life and a high elimination rate
constant while the reverse is the case for phenylacetic acid.
The affinities of these four acids for the sites of conjugation (mitochondria
for amino acid and microsomes for glucuronic acid conjugations) and the
conjugating enzymes associated with these structures were investigated. It
was found that the pattern of conjugation of an arylacetic acid in the rat is
influenced by its affinity for uptake (as measured by binding)by mitochondria and
endoplasmic reticulum and affinity for the conjugating enzyme systems associated
with these structures (as measured by their abilities to conjugate with glycine and
glucuronic acid).

3
ACKNOWLEDGEMENTS -
The work described in this thesis was carried out in the Department
of Biochemistry, St. Mary's Hospital Medical School.
I wish to thank Professor R. T. Williams, F. R. S. , for the great interest
he has shown in this project.
To Dr. J. Caldwell and Professor R. L. Smith, I am deeply grateful
for their encouragement and helpful advice over the last three years. I will
always appreciate the willingness of my fellow research workers, especially
Jeff, members of staff and the technical staff to assist practically whenever
necessary and in providing helpful information.
My sincere thanks to Miss Sally Turner for so capably typing this
thesis.
Finally, words can never adequately express my appreciation to members
of my family especially my brother Rowland, who financed this project and for
the understanding, patience and encouragement shown to me at all times.

4
INDEX
Page
Abstract 2
Acknowledgements 3
Chapter One Introduction 5
Two Materials and Methods 52
Three Metabolism of 1-Naphthylacetic Acid 78
Four Metabolism of Diphenylacetic Acid 106
Five Metabolism of Hydratropic Acid 118
Six Pharmacokinetics and Subcellular Aspects 179
of Arylacetic Acid Conjugation
Seven General Discussion and Conclusion 164
Appendix 175
References 179

5
CHAPTER ONE
Introduction
Contents
Page
Introduction 7
Biological Factors Affecting Drug Metabolism 8
Effect of Chemical Structure on Drug Metabolism 14 Cyclohexanecarboxylic Acids 17 Phenols 17 Polychlorinated Phenols 19 Benzoic Acids 19 Hydratropic Acid and Related Compounds 22 Aryl Alkyl Sulphones 23
Conjugation Mechanisms 25
Glucuronide Formation 25
Amino Acid Conjugation 27
Conjugation Patterns of Some Aromatic- and Arylalkyl- 30 carboxylic A cids
Aromatic Carboxylic Acids 31 Benzoic Acid 31 2 - (4 ' -Aminobenzoyloxy)benz oic Acid 33 Quin ()line -2-carboxylic Acid 34
Primary Arylacetic Acids 34 Phenylacetic Acid and its Simple Derivatives 34 1-Naphthylacetic Acid 38 Indole-3-acetic Acid 38 Indomethacin 39 Myalex 39 p-(Cyclopropylcarbonyl)phenylacetic Acid 40 (SQ 20, 650)
Metiaz inic Acid 41
Imidazole-4-acetic Acid 41
Secondary Arylacetic Acids 42 Ibuprofen (2-[4-isobutylpheny1]-propionic Acid) 42
Fenoprofen (dl-2-[3-phenoxyphenyl]7propionic Acid) 42 a - [4-(1 -Oxo-2-isoindoliny1)-phenyl]-propionic 43
Acid Diphenylacetic Acid 43

6
Tertiary Arylacetic Acids 43 a, a -Dimethylphenylacetic Acid 43 Triphenylacetic Acid 44 Benzilic Acid (Diphenylglycollic Acid) 44
Compounds Metabolised to Arylacetic Acids 44
4-(2:4:5-Trichlorophenoxyl)-butyric Acid 44 Diphenhydramine 45 Brompheniramine 46 Haloperidol 46 o, p -DDD, [1- (o-Chlorophenyl) -1 -(p' -chlorophenyl) -2,2- 47 dichloroethane] DDT [1,1 -Bis (p -chlorophenyl) -2,2,2, -trichloroethane] 48
X-ray Contrast Media 49
Some Endogenous Arylacetic Acid Derivatives Normally Occuring in the 49 Urine
Scope of the Present Investigation 50

7
Introduction
Most organic compounds which enter the body undergo metabolic
transformation prior to their excretion, although a few compounds are not
metabolised and are therefore excreted unchanged. These are usually
strongly polar compounds e.g. strong acids or bases. In addition a few non-
polar compounds such as the chlorinated biphenyl 2,4,5 , 2' , 4', 5'- hexachloro
biphenyl are excreted unchanged but this is a relatively unusual situation.
Some examples of these three types of compounds are shown in Table 1.1
Table 1.1
Compounds Eliminated without undergoing biochemical transformation
(Smith, 1974).
Strong acids
Strong bases
Non-polar
Methotrexate 5,5'7Methylenedisalicylic acid 2,4,5 -Trichlorophenoxyacetic acid
Hexamethonium salts Methylglyoxal-bisguanylhydrazone
2,4,5,2' , 4', 5' -Hexachlorobiphenyl Ether Barbitone
The metabolism of a foreign compound is generally a biphasic phenomenon
(Williams, 1959). In the first stage (Phase I) the compound undergoes a bio-
chemical reaction which may be oxidation, reduction or hydrolysis and which
usually introduces into the molecule a functional group such as hydroxyl (OH),
carboxyl (-COOH), amino (-NH2) or thiol (SH). In the second stage (Phase II)
the metabolite so formed is combined, usually through the newly introduced
functional group, with a molecule provided by the body which is derived from
carbohydrate, amino acid or other sources.

aromatic hydroxylation
conjugations OH
HO
Benzene Phenol pKa 10
Phenyl-p-glucuronide, pKa 3.4
OH
8
The conjugated product is typically an acid, largely ionised at body pH and
readily excreted in the urine and / or the bile. This pattern is well illustrated
by the metabolic fate of benzene. This compound undergoes a number of
metabolic reactions, the major pathways being oxidation (aromatic hydroxy-
lation) to phenol, followed by conjugation of the latter with glucuronic acid to
form phenyl-p-glucuronide, a strong water-soluble acid (pKa 3. 4) which is
readily excreted.
COOH
If the compound already possesses a suitable functional group conjugation may
occur directly. A number of Phase I and Phase II (conjugation) reactions are
shown in Tables 1. 2 and 1.3 .
The metabolic route or routes which a particular compound may undergo
will be determined by a wide variety of factors, such as the following:
i) biological factors e.g. species, strain, age and sex.
ii) chemical factors e. g. chemical structure (including molecular
size and geometry), ionization and lipid solubility.
iii) other factors, such as dose, route of administration, presence
of another drug, diseases, nutritional status, temperature
and altitude.
Biological factors affecting drug metabolism
These factors are principally, species, strain, age and sex. Species
variations in metabolism can occur both in respect to the speed at which

9
Table 1.2
Phase 1 - Reactions of Foreign Compounds
Reaction Class Example
Oxidations
Reduction
Hydroxylation N- and 0-dealkylation Deamination Replacement of S by 0 Ether cleavage Aromatisation Oxidation of thioethers
to sulphoxides
Reduction of nitro and keto groups
Reductive cleavages of azo groups
Reduction of C=O, C=C
Hydrolysis Hydrolysis of esters Amide hydrolysis

10
Table 1.3
The Principal Phase II (Conjugation) Reactions
Conjugation Reaction Conjugating agent Source
Glucuronide synthesis glucuronic acid
Glycine conjugation glycine
Glutamine conjugation glutamine
Methylation methionine
Mercapturic acid synthesis cysteine
carbohydrate
amino acids
Ethereal sulphate synthesis sulphate
Acetylation acetyl miscellaneous
Thiocya.nate formation thio (S-SO3H)

11
metabolism occurs and in the metabolic pathways employed, and these arise
mainly because of interspecies differences in the enzymic control of Phase
I and Phase II reactions. The enzymes occur mainly in the liver but are also
found, usually to a lesser extent, in several other tissues, including the
intestine, kidney and lung. Species differences in the pattern of metabolism
commonly arise because of one or more of the following reasons (Williams, 1967) :
i) competing reactions for example, the hydroxylation and ring
cleavage in the metabolism of coumarin (Table 1.4)
Table 1.4
Competing Reactions : Metabolism of Coumarin
(Kaighen and Williams, 1961 ; Gangolli et al. , 1974)
% of dose excreted as :
Hydrox-y -coumarins Ring open products
69 - 92
low
3
50
41
23
Species
Man
Rat
Rabbit
ii) the occurrence of metabolic defects in some species appear
to lack the capacity to carry out certain metabolic reactions
and some examples are shown in Table 1.5.
Although the rat is defective in the N-hydroxylation of
chlorphentermine, this reaction takes place extensively in man
rhesus monkey, rabbit and guinea pig (Caldwell et al. , 1975 a).
(Table 1.6).

Table 1.5
Species Defects in Common Metabolic Reactions
Defective Reaction Species References
N-Hydroxylation of aliphatic amines Rat, Marmoset Caldwell et al (1975a)
Glucuronide formation Cat, Gunn rat Robinson and Williams (1958)
Sulphate formation Pig, Opossum Stekol (1936) ; Combs and Hele (1927)
Arylamine acetylation Dog, Fox Marshall (1954)
Mercapturic acid formation Guinea pig Bray , et al. (1959) Bray and James (1960)
Glycine conjugation Fruit Bat Bababunmi et al (1973)

13
The cat forms little or none of the glucuronic acid
conjugate of phenol (Capel et al. 1972) but this defective
characteristic seems to be substrate dependent (Millburn, 1974)
(Table 1. 7).
Table 1. 6
Species Difference in the N-Hydroxylation of Chlorphentermine
Cl
CH3
CH C-NH Cl 21 2 CH3
CH3 -CH C-NHOH 21
CH3
Species % of Urinary 14C as
N-Hydroxy-p-chlorphentermine
Rat 0
Guinea Pig 50
Rabbit 42
Marmoset 0
Rhesus monkey 76
Man 44

14
Table 1. 7
Glucuronic Acid Conjugation in Cats : Variation with Substrate
Compound % of 24 h excretion conjugated with:
Glucuronide Sulphate
Phenol 1 95
1-Naphthol 1 98
2-Naphthol 3 97
Pax acet am ol 3 86
Phenolphthalein 60 40
iii) The occurrence of unusual metabolic reactions may be
restricted to certain species or a group of species and also
to certain compounds (Table 1. 8).
Some metabolic reactions appear to be restricted in
their species occurrence to man and other primate species
(Table. 1. 9).
Effect of Chemical Structure on Drug Metabolism
The metabolism of a compound depends among other things on its
chemical structure, which influences its physico-chemical properties such as
ionization and lipid solubility. A change in the chemical structure may vary the
physico-chemical properties and also affect the affinity of the compound for the
metabolising enzymes, and hence induce a possible change in its metabolic pattern.

15
Table 1. 8
Some Thcommon Metabolic Conjugation Reactions
Conjugating Agents Species
Ornithine Certain birds and reptiles
Taurine Pigeon, Ferret
Amino-acids Serine Rat, Rabbit
Glycyltaurine Cat
Arginine Ticks and Spiders
Glucose Insects
Carbo-hydrates Ribose Rat, Mouse
N-Acetylglucos- Rabbit amine
Phosphate Cat, Dog, Man
Acids Formate Rat, Dog
Succinate Rat, Dog

Table 1. 9
Metabolic Reactions Apparently Restricted to Primate Species
Metabolic reaction Species occurrence
N1-Glucuronide formation
Glutamine conjugation
Aromatisation of quinic acid
Man, New and Old World Monkeys and prosimiaas
Man, New and Old World Monkeys
Man and Old World Monkeys
0-Methylation of 4-hydroxy-3, Man, Monkeys 5-diiodo-benzoic acid
16

OSO3H 0. C6I1
90
6
17
Cyclohexanecarboxylic acids
Certain cyclohexanecarboxylic acid derivatives undergo aromatization
in the body, but the extent to which the process occurs has been found to depend
upon the nature of substituents attached to the cyclohexane ring and the animal
species studied, (Williams, 1959). Table 1. 10shows the metabolic pattern of
some cyclohexanecarboxylic acids and related derivatives.
Phenols
Phenols undergo metabolic conjugation with both glucuronic acid and
sulphate.
Although monosubstituted phenols are metabolised in a similar manner
to phenol, there are quantitative differences related to the chemical nature and
position of the substituent groups (Williams, 1938).
The more complex phenols such as stilboestol, hexoestrol and dienoestrol j
OH OH
OH
CHC2H5
CHC2H5
C=CHCH3
C=CHCH3
OX
OX
OX
Hexoestrol (X=H)
Stilboestrol (X=H) Dienoestrol (X=H)

Table 1.10
The Metabolism of Cyclohexanecarboxylic Acid and Related Acids
(Adapted from Williams, 1959)
18
R = C6H11 or
Derivative Structure Aromatic Metabolite
Cyclohexanecarboxylic R. C 00H acid
Cyclohexylacetic acid R. CH2COOH
g-cyclohexylpropionic R. CH2CH2COOH
y-cyclohexylbutyric acid R. (CH2)3COOH
N-Methylhexahydrobenz- R. CONHCH3 amide
Hexahydrobenzoylalanine R. CONHCH(CH3)COOH
benzoic acid
no aromatization; complete oxidation
benzoic acid
no aromatization ; (oxalic acid ).
benzoic acid
no aromatisation; original compound ex-creted
acid

19
are conjugated mainly with glucuronic acid in the rabbit (Dodgson et al. , 1948,
Mazur and Shorr, 1942).
Another phenol which is highly conjugated with glucuronic acid in the
rabbit but not at all with sulphuric acid is p-hydroxybenzophenone (Robinson
and Williams, 1957).
CO OH
High glucuronic acid conjugation and almost negligible sulphate conjugation
seems to be characteristic of phenols containing two isolated benzene rings.
Polychlorinated phenols
The main reaction of these phenols is again conjugation, except in the
case of pentachlorophenol and other chlorinated phenols with pKa values of less
than 7. 2:4-Dichloro- 2:4:5-trichloro-and a tetrachloro-phenol have been
reported as being excreted conjugated with glucuronic acid in the rabbit
(Deichmann and Thomas, 1943). As far as chlorinated phenols are concerned,
those with pKa less than 7 do not form ethereal sulphates in the rabbit as shown
in Table 1.11.
Benzoic Acids
The pattern of conjugation of the monosubstituted benzoic acids is
qualitatively similar to that of benzoic acid, but there are considerable
quantitative variations in the relative extent of the glycine and glucuronic acid
conjugations with this series of acids. Thus in the rabbit 2-toluic acid is
conjugated solely with glucuronic acid and not at all with glycine, 2-nitrobenzoic
acid is largely excreted unchanged, whilst anisic acid gives rise to more of its
glucuronic acid conjugate than its glycine conjugate when given in doses at which
benzoic acid is conjugated mainly with glycine. The quantitative data on the

20
Table 1.11
Ethereal Sulphate Formation and pKa of Chlorinated Phenols
(taken from Dodgson et al. , 1950)
(dose about 0.2 g/kg )
Phenols % of dose excreted
as ethereal sulphate pKa
4-Chlorophenol 22 9.2
2:4-Dichlorophenol 16 7.7
2:6-Dichlorophenol 0 6.8
2:4:5-Trichlorophenol 10 7.7
2:4:6-Trichlorophenol 0 6.4
Pentachlorophenol 0 5.3

Table 1.12
The Conjugation of Monosubstituted Benzoic Acids in Rabbits
Sub stituent pKa Approximate dose g/ kg
% Conjugation
Reference Glycine glucuronic acid
(ester type) Free
None 4. 2 0.4 83 15 1 Bray et al. (1955) 4-Nitro * 3.4 0.1 - 0.2 0 3? 80 - 90 Bray et al. (1949 a) 2-Chloro 2. 9 0. 3 5 19 60 Bray et al. (1952) 4-Chloro 4. 0 0.3 63 21 7 Bray et al. (1952) 2-Methyl 3. 9 0. 3 0 73 Bray et al. (1949 b) 4-Methyl 4.4 0. 3 46 14 73 Bray et al. (1949 b) 3-Hydroxy + 4.1 0. 5 ? 6 70 Bray et al. (1955) 4-Methoxy 4. 5 0.4 38 57 1 Bray et al. (1955) 4-Acetamido 4. 3 0.4 2 0 77 Bray et al. (1955) 4-Amino 4. 9 0. 5 + + 13 - 30 ++ Bray et al. (1948) '
* 11-21% of these acids is reduced to aminobenzoic acids + 5-14% of 2-, 2-19% of 3- and 0-18% of 4-hydroxybenzoic acids form ether glucuronides +÷ This acid is partly acetylated.

22
metabolism of a number of substituted benzoic acids in the rabbit are given in
Table 1.12.
Similar studies on the dog have been made by Quick (1932 a,b) who
found that substituents at 3- and 4-positions in the benzoic acid molecule had
little influence on the extent of glycine conjugation when compared with benzoic
acid, except when they were hydroxyl group which caused a depression of glycine
conjugation. Substitution in 2-position always reduced the extent of glycine
conjugation. With glucuronic acid conjugation, substitution in 3- position had
little effect, while acidic groups in the 2- and 4- positions reduced it and basic
groups increased it. Here again, acid strength and alternative metabolic
reactions are important factors.
Hydratropic acid and related compounds
Early experiments showed that tropic acid (a -hydroxymethylphenylacetic
acid, see below) is not metabolised and is excreted unchanged in the urine of cat
(Kay and Raper, 1922). Later work in the rat and the mouse showed that 95-98%
of a dose of tropic acid was recovered unchanged in the urine within two hours
of dosing (Gosselin et al. , 1955). Its isomer, atrolactic acid, behaves
similarly in dogs and cats (Kay & Raper, 1922), but its dehydrated analogue,
atropic acid is believed to be completely destroyed in dogs (Kay and Raper, 1922).
In the rabbit
CHCOOH
CH2OH
C. COOH CH. COOH
CH2 CH3
Hydratropic acid (pKa 4. 6)
Tropic acid
(pKa 4.1) Atropic acid
(pKa 3. 85)
—COH. COOH
CH3 Atrolacetic acid
(pKa 3. 53)

CHCONH CH 3 2 2-\ CH 3
23
the racemic, dextro- and laevo- forms of the hydratropic acid are highly con-
jugated with glucuronic acid and are excreted as hydratropoylglucuronide
(Robinson et al. , 1955) but Kay and Raper (1922) have also isolated hydra-
tropoylglycine in the dog.
The metabolic route of tropic, atropic, atrolatic and hydratropic acids
seems to depend on the type of substituents in the a-carbon.
Aryl alkyl suiphones
Aminomethylphenyl methyl sulphone (V 335) possesses antibacterial
properties, and when administered to rabbits is excreted mainly as 4-methyl-
sulphonylbenzoic acid (Hartles and Williams, 1947).
NH2 02C113 oxidative
HOO 0 CHI 3 deamination
The acetylated derivative of V 335, 4-acetamidomethylphenyl methyl sulpbone. ,
has no antibacterial activity and on administration to rabbits is excreted
unchanged (Hartles and Williams, 1949). Table 1.13 shows the metabolic fate of
some other derivatives in the rabbit. The size of the acyl substituent seems to
determine whether or not it is excreted unchanged or metabolised, or excreted
in the faeces.

24
Table 1.13
Metabolism of 4-Acylaminomethylphenyl Methyl Sulphones
R. CONHCH2 C6 H4 . SO4. CH3 dose 1 g / rabbit
(Adapted from Williams, 1959)
Derivatives
Urinary Excretion Faecal excret-ion in 5 days of unchanged drug %
1
R unchanged
%
l metabolised*
%
V335 - 0 87
Formyl H 48.5 15.8
Acetyl CH3 61.1 0
Propionyl C2H5 42.9 13.1
Butyryl C3H7 3.0 62.1 0
Hexoyl C51111 0 56.8 2.3
Decoyl C9H19 0 22.2 40
Tetradecoyl C13H27 0 0 50
* Excreted in the urine as 4-methylsulphonylbenzoic acid.

25
Conjugation Mechanisms
The biosynthetic reactions between a foreign compound and a substance
provided by the body (see Table1.3)are catalysed by specific enzymes which
require certain activated nucleotides for their enzymic action (Williams, 1969).
The intermediate nucleotide may contain either the conjugating agent or the
foreign compound. These two types of conjugation may be represented as : -
1) Activated ener source
gy Foreign Compound> Conjugated Conjugating > conjugating
agent agent transferase product
Activated energy conjugating ■
2) Foreign > Foreign / Conjugated source agent
Compound Co mpound product + transferase
The first class of detoxication mechanism includes the formation of
glucuronides, acetylation and methylation.
Peptide conjugations with glycine, glutamine and ornithine come under
the second class of detoxication mechanism (Williams, 1969). In these cases, the
appropriate transferase enzyme is associated with the mitochondria. Table 1.14
shows some examples of these two classes of conjugation reactions.
Glucuronide Formation
Conjugation with glucuronic acid is widespread among species, occurring
in mammals (Teague, 1954), marsupials (Hinks and Bolliger, 1957), birds
(Baldwin, Robinson and Williams, 1960), reptiles and amphibia (Smith, 1964).
In insects, the mechanism appears to be replaced by p-glucoside formation. This
is a characteristic insect detoxication and probably occurs in other invertebrates
as well (Smith, 1968). Glucuronide formation can occur with compounds
possessing -OH, -COOH, -NH2 and - SH groups. The mechanism of glucuronide

Table 1.14
Mechanistic Classification of Conjugation Reactions
(Adapted from Williams, 1974)
Reaction Intermediate Transferring enzyme Location
1. Intermediate containing conjugating agent
Glucuronide synthesis
Glucoside
Ether sulphate sunthesis
Methylation
Acetylation
Cyanide detaNication
UDP-glucuronic acid
UDP -glucose
Adenosine 3'-phosphate 5'-sulphatophosphate
Adenosylmethionine
Acetyl-CoA
Thiosulphate (S-SO3H)
Glucuronyltransferase
UDP-glucose glucosyl-transferase
Sulphotransferase
Methyltransferase
Transferase
Sulphurtransferase
Liver, kidney, lung, spleen, urinary bladder, gastro-intestinal tract
In insects - hepatic caecum
Liver, kidney, intestine
Liver ,kidney, spleen, brain, small7dnte stine , lung, heart, muscle .
Liver, kidney, brain, lung, pancrease, spleen, blood.
Wide distribution in the body but high concentration in the liver.
2. Intermediate containing foreign compound
Glycine conjugation Aroyl-CoA Glycine acyltransferase Liver and /or kidney only
Glutamine conjugation Arylacetyl-CoA Not defined (glutamine acyltransferase ?).
In man - kidney and liver
Ornithine conjugation Aroyl-CoA Not defined (ornithine acyltransferase ? ).
In chickens - kidney only
Mercapturic acid Epoxide * of foreign Glutathione S-opoxide- Liver kidney, pancreas synthesis compound transferase *.
* Some compounds are sufficiently reactive to combine with glutathione without further changes ; others such as hydrocarbons require
activation by oxidation to epoxides ; several glutathione-S-transferases occur(see Boyland & Chasseaud, 1969);

27 s3rnthesis can be described as follows: -
1) a-Glucose-1- phosphate + UTP
Uridyl- UDP glucose + pyrophosphate transferase
2) UDP-glucose
UDP -glucose dehydrogenase
DPN
UDP glucuronic acid
3) UDP glucuronic UDP - glucuronyl Aglycone glucuronide +
acid +Aglycone UDP transferase
The enzyme UDP-glucuronyltransferase is in, or associated with-the
microsomes (Dutton, 1961). Table 1.15 shows some carboxylic acids that form
ester glucuronides.
Amino Acid Conjugation
The mechanisms of amino acid conjugation involve a three-stage process,
the first two stages result in the activation of the carboxylic acid ; and the third
in the conjugation of the ,activated carboxylic acid with an amino acid, the reaction
1) R. COOH +ATP R. CO. AMP + pyrophosphate
2) R. CO. AMP + CoA -SH .> R. COS CoA + adenylic acid
3) R. COS CoA + H2 NCHCOOH > R. CONH. CH. COOH I I R' IV
CoA-SH
being catalysed by the respective amino acid-N-acylase. , This process takes
place in the mitochondria (Schachter and Taggart, 1953). Table 1.16 shows the
amino acids used by some species in conjugating some carboxylic adds.
Birds classed as Galliformes (hens, turkeys) and Anseriformes (ducks
and geese) excrete aromatic acids and arylacetic acids as ornithine conjugates.

Monkey Lan et al. 1975
Dog, monkey Harman et al. 1964
Pig-tailed and Foulkes (1970) talapoin monkeys
Robinson et al. 1955
Rubin et al. 1972
Robinson .& Williams (1955)
McChesney and Hoppe 1954.
Rabbit
Man
Rabbit
Dog, cat
28
Table 1.15
Some carboxylic acids which form ester glucuronides
Compound Species References
2-Ethylbutyric acid Trimethylacetic acid 3:3-Dimethylbutyric acid 2:4:4-Trimethylpentoic
acid 2 -Ethylhexoic acid
Benzoic acid
[p-(Cyclopropylcarbonyl) -phenyl ] acetic acid
Indomethacin
Myalex
Hydratropic acid
Fenoprofen
a, a -Dimethylphenylacetic acid
Iopanoic acid
Rabbit
Dziewiatkowski et al. (1949)
Indian fruit bat Bridges et al. 1970

Table 1.16
The amino acids used in conjugation reactions
Amino acids Species Substrate References
Glycine rat, rabbit, man cat, monkey
benzoic acid Bridges et al., 1970
Glutamine man, monkey phenylacetic acid James et al., 1972 a
Taurine dog [p-(Cyclopropylcarbony1)- phenyl]acetic acid.
Lan et al., 1975
Ornithine chicken phenylacetic acid. James et al., 1972 a
Aspartic acid rat bis (p -chlorophenyl) acetic acid
Pinto et al.,1965
Alanine mouse, hamster bis(p-chlorophenyl)acetic acid
Wallcane et al 1973
Serine rat o,p'-dichlorodiphenylacetic acid bis(p-chlorophenyl) acetic acid
Reif and Sinsheimer, 1975 Pinto et al 1965
Glutamic acid fruit bat benzoic acid Idle et al. , (1975)

30
Ornithine conjugation is not characteristic of all birds since the pigeon and dove
(Columbiformes) excrete aromatic acid as glycine conjugates (Baldwin, Robinson
and Williams, 1960).
Reptiles, which have a close phylogenetic relationship to birds, are
also capable of conjugation with ornithine, though, in the species examined,
glycine conjugation is also found. Thus urine from tortoises dosed with aromatic
acids was found to contain both ornithine and glycine conjugates ; similar
results were found with a grass snake, two species of lizard and an alligator
(Smith, 1964).
Extensive conjugation of aromatic acids is found in Arachnids. Although
the excreta may contain conjugates with arginine, glutamine, glutamic acid,
citrulline, ornithine and agmatine, it is thought that, with the exception of the
glutamine conjugate, all these are derived from the original arginine conjugate
(Hitchcock and Smith, 1966).
Kaihara and Price (1965) have also reported glycyltaurine and glycyl-
glycine conjugates of quinoline-2-carboxylic acid in the cat.
Conjugation Patterns of Some Aromatic - and Arylalkyl - Carboxylic Acids
Aromatic - and short-chain arylalkyl - carboxylic acids are metabolised
in general by conjugation either with an amino acid or with glucuronic acid.
Some which have low pKa may be excreted unchanged (e. g. 2-nitrobenzoic acid
pKa 2.2). The nature of the amino acid used varies with species, the two main
ones being glycine, in sub-primate mammals, and glutamine in primates.
Conjugation with a variety of other amino acids, including taurine, serine,
aspartic acid, glutamic acid and alanine have also been reported (see Table
1.16).

COON
Benzoic acid
Hippuric acid
31
Aromatic Carboxylic Acids
Benzoic Acid
Benzoic acid generally forms hippuric acid and benzoylglucuronide in
the animal body, and the relative amounts of these conjugates depend on the
dose and the species.
CONHCH2COOH
COOC6H906
Benzoylglucuronide
Bridges et al. (1970) have investigate the metabolic pattern of benzoic acid in
a range of species and some of their results are shown in Table 1:17. They have also
reported that chicken, turtle (side-necked) and gecko excrete benzoic acid mainly
as ornithuric acid and with small amount of hippuric acid. Indian fruit bat produced
mainly benzoylglucuronide (Bridges et al. , 1970) and this possibily indicates a
defect in hippuric acid formation. This possible defect has been confirmed by
Bababunmi et al . (1973).
Idle et al. (1975) have recently reported that benzoylglutamic acid is one
of the metabolites of benzoic acid in the fruit bat.

32
Table 1.17
Metabolites of Benzoic Acid in Some Species
(Adapted from Bridges et al. ; 1970)
Species dose
mg/kg
% 24 h excretion found as
Hippuric acid
Benzoyl glucuronide
Man 1 100 0
Rhesus monkey 20 100 0
Rabbit, cat and capuchin
50 100 0
Dog, squirrel monkey, ferret, hedgehog and pigeon
50 80 20
Ferret 200 47 44
Ferret 400 30 49

— 0 - C 11 0
unchanged (50%)
COOH
OC6H903
COOC6H
90
6
OH
CONHCH2COOH
OH
33
2-(4'-Aminobenzoyloxy)benzoic acid
The metabolism of 2-(4'-am_inobenzoyloxy)benzoic acid has been studied
in man (Smyth et al. , 1974). It is metabolised to salicylic acid which is
excreted as salicyluric acid and salicyloyl glucuronides, and 4-aminobenzoic
acid which is excreted as N-acetyl-4-aminobenzoic acid.
COOH
2-(4'-Aminobenzoyloxy)benzoic acid
COOH
OH COOH
Salicylic acid
NH2
4-Aminobenzoic acid
N-acetylation
Salicyluric acid
Salicyloyl glucuronides ( 15 % ) (35%)

34
Quinoline-2-carboxylic acid
The aromatic acid quinoline-2-carboxylic acid (see below) and certain
hydroxylated derivatives of this acid, give rise to unusual conjugation reactions
in certain species. In the rat, the acid is conjugated with glycine. In the cat,
however, a major metabolite of this acid is quinaldylglycyltaurine, and small
amounts of quinaldylglycylglycine are also formed (Kaihara and Price, 1965).
Quinoline-2-carboxylic acid
The cat is also able to form the glycyltaurine conjugate of 4-hydroxy-
quinoline-2- carboxylic acid. The dihydroxylated derivative, 4, 8-dihydroxy-
quinoline-2-carboxylic acid, is excreted in the rat as double conjugate, giving
two ether glucuronides (8-monoglucuronide and 4, 8-diglucuronide) each with a
serine residue attached to the 2-carboxyl group, whereas in the rabbit, the
hydroxyl groups are conjugated with sulphate but again, a serine residue is
attached to the 2-carboxyl group (Rothstein and Greenberg, 1957).
Primary Arylacetic Acids
Phenylacetic acid and its simple derivatives
Some earlier workers showed that phenylacetic acid is excreted conjugated
with glycine in rabbits, dogs, sheep, horses and rhesus monkey (Salkowski and
Salkowsi, 1879 ; Salkowski, 1884 ; Vasiliu, 1909 ; Sherwin, 1917) and with
ornithine in the hen (Totani, 1910). Thierfelder and Sherwin (1914, 1915) showed
that phenylacetic acid is excreted as the glutamine conjugate by man , and the
same conjugate has also been found in the urine of chimpanzee (Power, 1936).

35
CH2COOH
Phenylacetic acid
-CH2CONHCH2
COOH
CH 2
H CONHCH \CH 2
COOH CONH2
Phenacetylglycine Phenacetylglutamine
COOH
CH2CONH(CH2
)3CHNHCOCH2
Diphenacetylornithine
Sherwin also studied the metabolism of some simple substituted
phenylacetic acids in man, dog and rabbit. He found that in man 2-, 3-, and
4-hydroxyphenylacetic acids, 2-,3- and 4-nitrophenylacetic acids were excreted
in the urine unchanged, and that 4-aminophenylacetic acid was acetylated but not
conjugated at the -COOH group. He also reported that 2- and 4-chloro-, bromo-
and iodo-phenylacetic acids were excreted as glycine conjugates by man and dog
(Sherwin 1918, Sherwin 1923 ; Cerecedo and Sherwin 1924 ; Muenzen, Cerecedo
and Sherwin, 1926).
In 1958 the glutamine conjugate of 3,4-dihydroxy-5-methoxyphenylacetic
acid was found as a metabolite of mescaline in man (Harley-Mason et al. 1958).

36
3,4,5-Trimethoxyphenylacetic acid was also reported to be excreted unconjugated
as a metabolite of mescaline (Charalampous et al. , 1964). Another substituted
phenylacetic acid, 4-methoxyphenylacetic acid is excreted as 4-methoxy-phenacetyl-
glutamine and its 0-deme,thylate metabolite 4-hydroxyphenacetylglutamine in man
(Oakley and Seakins, 1971).
The metabolic studies of phenylacetic acid by James et al. (1972 a) in
a number of species have shown the pattern of conjugation represented in Table
1.18.
The same workers also showed that two avian species differed in the
conjugates excreted. Pigeons excrete phenacetylglycine and phenacetyltaurine
while chickens excrete mainly diphenacetylornithine. They also showed that
phenacetyltaurine was found in significant amounts in the urine of ferret, bush-
baby, slow loris, squirrel monkey, capuchin and pigeon but small amounts
were also found in most other species.
James et al (1972 b ) also reported that 4-nitrophenylacetic acid is excreted
unchanged by man and rhesus monkey, but conjugated with glycine by rats, and
that 4-chlorophenylacetic acid is similar to phenylacetic acid in its species
distribution of glycine and glutamine conjugation.
Recently Ette et al (1974) have reported that phenacetylglycine is the
major metabolite, while phenacetylglucuronide is a minor metabolite of phenylacetic
acid in the Indian fruit bat. This is an indication that glycine conjugation in this
species is substrate dependent, since it does not occur with benzoic acid
(Bababunmi et al. , 1973).

37
Table 1.18
Metabolic Conjugation of Phenylacetic Acid
Class
% of 24 h excretion conjugated with
Glutamine Glycine
Man 93 0
Old World Monkeys (8) 30 - 90 0.1 - 1
New World Monkeys (3) 64 - 80 1 - 10
Lemurs (2) 0 85
Sub-primates (10) 0 80 - 100
(number of species tested shown in parentheses)

38
1 -Naphthylacetic Acid
Lethco and Brouwer (1966) have shown that when 1-naphthylacetic
acid is administered orally to the rat, it is conjugated with glycine and
glucuronic acid. With small doses, the conjugation with glycine is the major
metabolic route, whereas the glucuronide formation becomes the major route
with larger doses. Bernhard and Caflisch-Weill (1949) found the glycine
conjugate in the urine of a dog which had been fed 1-naphthylacetic acid but
they did not detect this metabolite in the urine of rats and rabbits receiving this
acid.
Indole-3-acetic Acid
CH 2COOH
H
Indole-3-acetic acid is excreted as its glutamine conjugate in man and
old world monkeys, as glutamine and glycine conjugates in new world monkeys
and as glycine conjugate in prosimian species and lower animals (Evans 1972).
Further work by Bridges et aL,(1974) indicates that in man, the glycine conjugate
was not formed, the metabolites being the glucuronic acid and glutamine
conjugates of indole -3-acetic acid. Indolylacetyltaurine - is the only metabolite in
the pigeon, while it is a substantial metabolite in the green monkey, squirrel
monkey, the capuchin monkey and the ferret.
Cl Cl
C=0
CH3 0
39
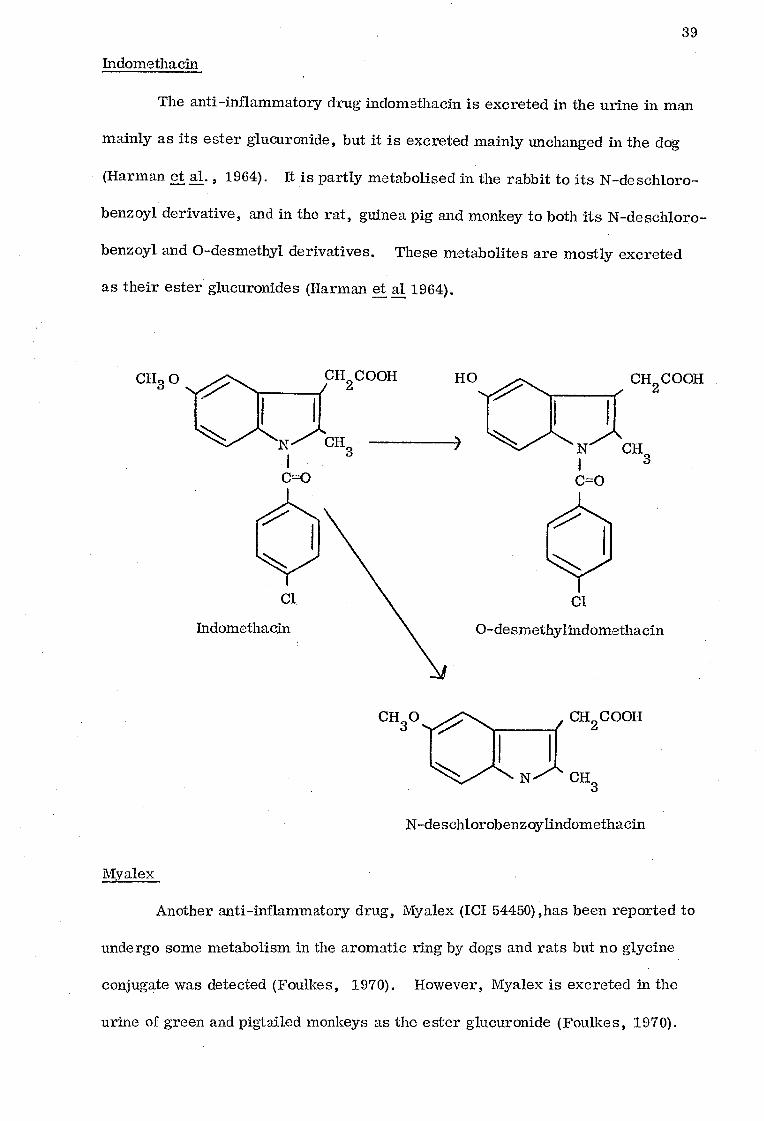
Indomethacin
The anti-inflammatory drug indomethacin is excreted in the urine in man
mainly as its ester glucuronide, but it is excreted mainly unchanged in the dog
(Harman et al. , 1964). It is partly metabolised in the rabbit to its N-deschloro-__
benzoyl derivative, and in the rat, guinea pig and monkey to both its N-deschloro-
benzoyl and 0-desmethyl derivatives. These metabolites are mostly excreted
as their ester glucuronides (Harman et al 1964).
Indomethacin 0-de smethylindomethacin
CH30 CH2COOH
CH3
N-deschlorobenzoylindomethacin
Mvalex
Another anti-inflammatory drug, Myalex (ICI 54450),has been reported to
undergo some metabolism in the aromatic ring by dogs and rats but no glycine
conjugate was detected (Foulkes, 1970). However, Myalex is excreted in the
urine of green and pigtailed monkeys as the ester glucuronide (Foulkes, 1970).

monkey (88%)
Glucuronic acid conjugate
Dog (27%)
Taurine conjugates
CH2COOH
Taurine CH2COOH
Dog og (30%) Glycine
Dog (31")> conjugate
CH2COOH
40
CH2COOC6H906
. Cl
Cl
Myalex Myalex ester glucuronide
p-(Cyclopropylcarbonyl)phenylacetic Acid (SQ 20, 650).
p-(Cyclopropylcarbonyl)phenylacetic acid a nonsteroidal anti-inflammatory
agent with analgesic activity, exhibits species variations in its metabolism
(Lan et al. 1975). In the rhesus monkey it is conjugated with glucuronic acid,
and in the dog with taurine. Its reduction product (a-cyclopropyl-a-hydroxy-p-
tolyl)acetic acid (SQ 21, 316) is conjugated with both taurine and glycine in the
dog.
( SQ 20, 650 )
Reduction Dog (53% ) Monkey (8-9%)
(SQ 21, 316)

41
Metiazinic Acid
Metiazinic acid is conjugated with both amino (-NH2) and methyl groups
in rabbit and man, and excreted as both amide and methylester (Populaire et al
1969). This is an unusual conjugation pattern since arylacetic acids are
usually conjugated with an amino acid or glucuronic acid.
Imidazole-4-acetic Acid
CH2 COOH
N CH
2COOH i 1 i
N j0
H V 0 CH-CHOH-CHOH-CHCH2OH
The metabolic conjugation of imidazole-4-acetic acid is unique when
compared with the other arylacetic acids already considered. The conjugation
does not involve the carboxylic acid group of the compound or the usual conjugating
agents utilised by arylacetic acids, but involves the use of ribose as the conjugating
agent. Schayer (1956) has reported that in rat and mice imidazole-4-acetic acid
is excreted as 1-ribosylimidazole-4-acetic acid. This riboside is also formed
as a metabolite of histamine which is degraded to imadazole-4-acetic acid. In
rats and mice the riboside is a major metabolite of small doses of histamine,
whereas in cats and dogs it is a very minor metabolite (Schayer 1956).

42
Secondary Arylacetic Acids
Ibuprofen (2-K-isobutylphenyll-propionic acid).
CH3
CH - CH
CH3
CH3
CH. C0011
Ibuprofen, an anti-inflammatory agent is metabolised by the oxidation of
the isobutyl side chain, but the carboxylic acid group is unaffected by its bio-
transformation in man (Adams et al. 1969) . This is unlike hydratropic acid
which is conjugated with glucuronic acid in the rabbit (Robinson et al. 1955) and
with glycine in the dog (Kay and Raper, 1922).
Fenoprofen (d1-2-[3-phenoxyphenyl]-propionic acid)
Fenoprofen, an anti-inflammatory analgesic agent is excreted mainly as
fenoprofen glucuroniceand 4'-hydroxyfenoprofen acyl-glucuronide in man (Rubin
et al. 1972). There are also very small amounts of unidentified acid -labile
conjugates of fenoprofen and 4'-hydroxyfenoprofen,
CH3 CH-COOH
fenoprofen

43
a - - (1 -Oxo-2-isoindoliny1)-phenyll-propionic Acid (K 4277)
CO\
N
CH2
CH3
CHCOOH
The metabolic fate of the anti-inflammatory agent K 4277 has been examined
in the rhesus monkey by Chasseaud et al. , (1974), who reported that it is excreted
mostly unchanged (78% of dose) with some 5% as the 5-hydroxy derivative and a
very small amount as glucuronide. K 4277 is excreted mainly as its ester
glucuronide conjugate by man and rabbit (Fuccella et al. , 1973 ; Goldaniga et al. ,
1973).
Diphenylacetic acid
Diphenylacetic acid (pKa 3. 94) is excreted as its ester glucuronide in man, dog
and rabbit (Miriam et al. , 1927a).
Tertiary Arylacetic Acids
a, a'-Dimethylphenylacetic Acid
a, a-Dimethylphenylacetic acid is metabolised exclusively to an ester
glucuronide in the rabbit (Robinson &Williams 1955).

44
Triphenylacetic Acid
Triphenylacetic acid , (pKa '3.96) has been shown by Miriam et al (1927b)
to be excreted totally unchanged by rabbit, dog and man.
Benzilic Acid (Diphenylglycollic acid)
Benzilic acid (pKa 3. 06) is excreted unchanged by rabbits (Sieberg and
Harloff, 1919).
Compounds Metabolised to Arylacetic Acids
4-(2:4:5-Trichlorophenoxy)-butyric Acid
Bohme and Grunow (1974) have examined the metabolism of 4-(2:4:5-
trichlorophenoxyl)-butyric acid, and 2:4:5-trichlorophenol was found as its
principal metabolite following oral administration to rats. 2:4:5-Trichlorophenoxy-
acetic acid, the final product of a-oxidation was also excreted in the urine. In a

45
different study, Grunow and Bohme (1974) have shown that 2:4:5-trichlorophenoxy-
acetic acid and 2:4-dichlorophenoxyacetic acid are excreted by rat and mice
as glycine and taurine conjugates.
Cl
Cl OCH2CH
2CH
2COOH
CI
OCH2COOH CI
4-(2:4:5 -Trichlorophenoxy) -butyric acid
Cl
Cl
V conjugated with glycine and
taurine
OH
Cl
2:4:5-Trichlorophenol
Diphenhydramine
Drach and Howell (1968) have shown that the antihistamine drug diphen-
hydramine (Benadryl: 2-diphenyl-methoxy-N,N-dimethylethylamine) is metabolised
to diphenylmethoxyacetic acid which is excreted as a glutamine conjugate by the
rhesus monkey.
CH 3 HC-OCH CH N
, oxidative 2 2 \CH3 deamination HC-OCH2COOH
Diphenhydramine Diphenylmethoxyacetic acid
conjugated with glutamine

CH 3 oxidative HC-CH CH N 2 2\ HC-CH
2C00:-I
deamination CH
3
46
Brompheniramine
Another antihistamine drug, brompheniramine which bears some structural
resemblance to diphenhydramine, is also metabolised to a substituted acetic acid
but this is reported to be excreted by man and dog partly as the glycine conjugate
(Bruce etal 1968).
Brompheniramine 47
conjugated with glycine
Haloperidol
Haloperidol, and other structurally related butyrophenone neuroleptic
drugs were reported to be metabolised in rats to p-fluorophenylacetic acid,
the glycine conjugate of which was the major urinary metabolite of the drug in
rats. (Braun, Saudijn and Poos, 1967).
CO-(CH2)„ -N/
>R CH2COOH
1
R' = -OH Cl
conjugated with
glycine
Haloperidol

Cl
(HO)2
Cl - - - - -> Cl
47
1 -( 0 -Chlorophenyl) -1 -(p'-chlorophenyl) -2,2 -dichloroethane (o, p' -DDD).
The metabolism of 0,pt-DDD has been investigated in the rat (Reif and
Sinsheimer, 1975). After 100 mg oral dose to rats, 7% is recovered in the
urine and 88% in faeces within 8 days. The urine was found to contain
o -p' -dichlorodiphenylacetic acid (0,p' -DDA) as well as 4-hydroxy-, 3-hydroxy-
and 3, 4-dihydroxy- substituted o -p'DDA. The serine and glycine conjugates
of 2,4' -DDA were also identified. In addition to the above metabolites the faeces
contained o-pi-DDD, 1-(2-chloropheny1)-1-(4-chloropheny1)- 2-chloroethylene,
and the aspartic acid conjugate of o,p'-DDA.
In the human studies, serine and glycine conjugates of o,p'-DDA have
been identified also (Reif and Sinsheimer, 1974).
Cl
3,4-Dihydroxy derivative 3-hydroxy- and 4-hydroxy
derivatives

48
DDT [1,1 -bis (p-chlorophenyl) -2,2,2 -trichloroethane I
DDT is partly metabolised to bis (p-chlorophenyl) acetic acid which is
excreted as serine and aspartic conjugates in rat (Pinto et al. 1965). The
mouse and hamster conjugate bis(p-chlorophenyl)- acetic acid with alanine and
glycine (Wallcane et al.1973). However, when bis(p-chlorophenyl) acetic acid
is administered intravenously to rat, it undergoes enterohepatic circulation
almost completely, and the only biliary metabolite is a glucuronide
(Gingell, 1975).
Cl -j Cl Cl
DDT
Cl H
COOH
bis(p-chtorophenyl)a cetic acid

49
X-ray Contrast Media
Compounds which are opaque to x-rays have been used as contrast media
after their injection to patients. Those which have high molecular weights
are excreted extensively in the bile, and produce gallbladder shadows which can
be used in diagnosis. Examples of such compounds are iopanoic acid, iophenoxic
acid and pheniodol.
Iopanoic acid is excreted in dog and cat as its ester glucuronide
(McChesney and Hoppe, 1958), and iophenoxic acid is excreted in dog as its
mono-ester-, mono-ether- and di-glucuronide (Wade et al., 1970) but pheniodol
seems to be excreted unchanged in rabbit and man (Junkman 1941).
C2H
5 1 CH
2CHCOOH
Iopanoic acid
C H 2 5
CH2CHCOOH
Iophenoxic acid
CH 2 —CHCOOH
Pheniodol
Some Endogenous Arylacetic Acid derivatives Normally Occuring in the Urine
A number of ring substituted arylacetic acids derived from the metabolism
of amino acids are found in normal urine with the carboxylic acid group unconjugated.
Included in this group are the phenolic acids which are normal constituents of

50
urine. Indeed the urinary levels of these acids can be important in diagnosis.
These include 3,4-dihydroxyphenylacetic acid (a metabolite of DOPA), and its
metabolites 3-methoxy-4-hydroxyphenylacetic acid and 3-hydroxyphenylacetic
acid, 2,5 -dihydroxyphenylacetic(homogenti dc acid) , 2 -hydroxyphenylacetic
acid, 4-hydroxyphenylacetic acid, 5-hydroxyindole-3-acetic acid (a metabolite
of tryptophol, and imidazoleacetic acid (a metabolite of histidine), (De Eds
et al. , 1955, Armstrong et al. , 1956, Neuberger et al. , 1947, Delvigs et al. ,
1965, Snyder et al. , 1964).
Scope of the present investigation
The literature survey indicates that there is enough evidence to suggest
that the structure of an aromatic - or arylalkyl - carboxylic acid greatly
influence its metabolic pattern of conjugation. The aromatic acids are con-
jugated with both amino acid and glucuronic acid, but the small primary
arylacetic acids are conjugated mainly with amino acids, however with increase
in complexity in the structure of the primary arylacetic acid there is a shift
from amino acid conjugation to glucuronic acid conjugation. Secondary arylacetic
acids and the small tertiary arylacetic acids are conjugated mainly with
glucuronic acid, but the large tertiary arylacetic acids are excreted unchanged.
The question arises as to what factors determine whether or not an amino
acid or glucuronic acid conjugation takes place. One or an interaction of two or
more of the following factors may be responsible : -
1) Ionization
2) Lipid solubility
3) Pharmacokinetic behaviour
4) Molecular size and geometry
5) Affinity for subcellular conjugation sites (mitochondria and
endoplasmic reticulum) and the associated enzymes.

51
Therefore this thesis describes investigation into the following: -
i) effect of chemical structure and dose on the metabolic
route of three arylacetic acids, namely 1-naphthylacetic,
diphenylacetic and hydratropic acids, in some selected
species.
ii) pharmacokinetic behaviour of these three acids and phenylacetic
acid in the rabbit.
iii) the affinities of these four acids to the conjugating sites
(mitochondria and microsomes) and the conjugating enzymes
associated with these structures.

52
CHAPTER TWO
Materials and Methods
Contents
Compounds
Radiochemical Synthesis
[Carboxyl-14C]diphenylacetic acid
[14 C -Methyl] - ( -Hydratropic acid
Synthesis of 1-Naphthylacetic Acid Conjugates
1 -Naphthylacetylglycine 1 -Naphthylacetyl-L -glutamine 1-Naphthylacetyltaurine 1-Naphthylacetylglucuronide
Synthesis of Diphenylacetic Acid Conjugates
Pages
54
54
54
55
56
56 56 57 58
58
Diphenylacetylglycine 58 Dipheny lacetyl -L -glutamine 59 Diphenylacetyltaurine 59 Diphenylacetylglucuronide 60
Synthesis of (±) Hydratropic Acid Conjugates 61
(±) -Hydratropoylglycine 61 (±) -Hydratropoyl-L-glutamine 62 (±) -Hydratropoyltaurine 62 (+) -Hydratropoylglucuronide 63
Benzylamine salt of (±)-Hydratropic acid 64
Metabolic Studies 65
Animals 65
Collection of carbon dioxide in the expired air 65 Paper and Thin-layer Chromatography 65 Location of Compounds on Chromatography 66
Ultra-violet (U. V.) light 66 Spray Reagents 66 Naphth ore sor cinol spray 66 4-Dimethylaminobenzaldehyde spray 66 Chlorine-starch/potassium iodide detection 67
reagent Ninhydrin spray 67

53
Chromatographic properties of taurine 67
Radiochemical Techniques 67 Spectra 71 Treatment of urine samples 71
Pharmacokinetic Studies 72
Animals 72
Cannulation of Marginal Vein 72
Administration of Compounds 72
Blood Analysis for 14C-content 73
In Vitro Studies 73
Preparation of Tissue Subfractions 73
Mitochondria 73 Microsomes 73
Binding Studies 74
Protein Determination 75
Enzyme Affinity 75
Glycine Conjugation 75 Glucuronic Acid Conjugation 75
Reverse Isotope Dilution Experiments 76

54
Materials and Methods
Compounds
[Carboxyl-14C]-1-naphthylacetic acid (specific activity, 44 mCi/mmol. ),
[Carboxyl u]-phenylacetic acid (specific activity, 59 mCi/mmol), sodium
[14
C]-cyanide (specific activity 55.5 mCi/mmol) and [14C]-methyl iodide (specific
activity 58 mCi/mmol) were purchased from the Radiochemical Centre,
Amersham, England. Phenylacetic, 1-naphthylacetic, diphenylacetic and
hydratropic acids were obtained from commercial sources and purified as
appropriate. Phenacetylglycine was a sample prepared by James et al (1972a).
Uridinediphosphoglucuronic acid (UDPGA) and adenosine triphosphate
(ATP) were purchased from Sigma Chemical Co, Surbiton, England, and co-
enzyme A (CoA) was obtained from Boehringer Corp. , Ealing, England.
Radiochemical Synthesis
[Carboxyl-14C]diphenylacetic acid
Chlorodiphenylmethane (1.06 g) was mixed with cuprous [14C]cyanide
(0.54 g ; 2 mei ; prepared from [14C] sodium cyanide, by the method of Reid et al. ,
(1951) ) and heated for 2 h in an oil-bath at 200-210°. The mixture was
allowed to cool and then extracted with acetone (30 ml). The acetone extract was
_14 filtered and evaporated to dryness. The residue of crude diphenylaceto- [ C]-
nitrile was hydrolysed by heating under reflux with stirring for 3 h with 48 % hydro-
bromic acid (50 ml) following which the hydrolysate was extracted with ether in a
continuous extractor for 3 h. The ether phase was extracted with N-NaOH
(25 ml), the latter separated and acidified with 2N-HCI. The mixture was
extracted once more into ether and then back again into N-NaOH (10 ml). The
latter was separated and acidified with 2N-HC1. The precipitate of [14C] —
diphenylacetic acid that separated was filtered and recrystallised from water to

55
give white crystals m. p. 147° specific activity 3jCi/mg (yield 0.28 g,
14
radio-
chemical yield from cuprous [ C]cyanide, 42%). It was shown by chromato-
graphy in solvent/D and F followed by radiochromatogram scanning to be
radiochemically pure as shown by the appearance of a single 14C peak at Rf
values 0.88 and 0.92 respectively corresponding to diphenylacetic acid.
Reverse isotope dilution analysis showed a radiochemical purity of 99. 5%.
14 r c-Methy1]-( 4 -hydratropic acid
To a suspension of NaH-mineral oil (51. 6%) in dry dimethyl sulphoxide
(25 ml) under nitrogen was added dropwise benzyl cyanide, (3 g) dissolved in
dimethyl sulphoxide (30 ml) and the reaction mixture was stirred at room
14 _ temperature for 4 h. [ C]Methyl iodide (3.6 g ; 1 mCi) was then added slowly,
the temperature being maintained at 10° with an ice/water bath. After stirring
for 2 h, a further portion of unlabelled methyl iodide (2.6 g) was added and the
solution stirred overnight. The reaction mixture was then treated with dilute
acetic acid and extracted twice with ether ( 2 x 50 ml). The ether extract was
washed with saturated sodium bicarbonate solution, evaporated to afford a residue
of crude [2-[14
C]-methyl]benzy1 cyanide.
The latter was dissolved in ethanol (200 ml) and refluxed for 7 h with a
solution of KOH (30 g) in water (70 ml). The reaction mixture was evaporated
to dryness and the residue acidified with 2N HC1 and extracted with ether (2 x 50 ml).
The latter was then extracted with saturated sodium bicarbonate solution (15 ml),
acidified with 2N-HC1 and reextracted with ether ( 2 x 50 m1). The ether was
evaporated leaving [14C]-hydratropic acid, specific activity 0.21 pCi/mg (yield
3.5 g, radiochemical yield, 73. 6%). It was shown by chromatography in solvents
D and F followed by radiochromatogram scanning to be radiochromatographically
pure as shown by the appearance of a single 14C peak at Rf values 0.75 and 0.87

56
respectively corresponding to hydratropic acid. Reverse isotope dilution
analysis showed a radiochemical purity of 96. 5%. However, as stated above,
radiochromatography of the product showed only one 14C peak, and since hydra-
tropic acid is an oil at room temperature, the apparently low value for the
radiochemical purity of this material determined by isotope dilution is probably
due to traces of solvent which remained after evaporation.
Synthesis of 1-naplithylacetic acid conjugates
1 -Naphthylacetylglycine was prepared from 1-naphthylacetylchloride
and glycine according to the method of Friedman and Masse(1910). It
was recrystallised from aqueous ethanol to give white crystals m. p.
148 - 149° (lit. 148 - 150°) and had an equivalent weight by titration
(0.1 N - NaOH) of 246 (requires 243) C14H1303 N requires C, 69.14 ; H, 5.35
and N, 5.76. Found C, 68.88 ; H, 5.44 and N, 5.77. The mass spectrum
of the methyl ester showed a molecular ion at m/e 257 (relative
intensity 22.6) with prominent peaks at 115 (24), 141 (76), 142 (95) and
168 (29).
1 -Naphthylacetyl-L-glutamine was prepared from 1-naphthylacetyl
chloride and L-glutamine according to the method of Thierfelder and
Sherwin (1914). It gave white needle crystals m. p. 185 - 186° when
recrystallised from methanol. Equivalent weight by titration (0.1 N - NaOH)
was 315 (requires 314). C17111804N2 requires C, 64. 97; H, 5.73 and N,
8.92. Found, C, 64. 81; H, 5. 91 and N, 9. 05.
The mass spectrum of 1 -naphthylacetyl-L-glutamine methyl ester pre-
pared by treatment of the free acid with ethereal diazomethane showed
a molecular ion at m/e of 328 (relative intensity 3.3) with prominent peaks
at 116 (29), 141 (78), 142 (56), 155 (17), 168 (78) and 187 (3.2). (cracking
pattern, see Appendix).

57
1-Naphthylacetyltaurine was synthesized as follows: -
1-naphthylacetyl chloride (17. 4 g) was added dropwise with stirring
over a period of 4 h to an ice-cold solution of taurine (9.2 g) dissolved
in N-NaOH (82 ml). The reaction was continued overnight after which
the mixture was adjusted to pH 2 with 2N-HC1 and extracted with ether
(3 x 30 ml) to remove 1-naphthylacetic acid. The aqueous layer was
separated and reduced.to dryness in a rotary evaporator and the residue
extracted with methanol (400 ml). The methanol extract was filtered and
reduced to 70 ml on a rotary evaporator. On addition of acetone to the
concentrate white crystals separated which were filtered and recrystallised
from the mixture of methanol and acetone (2:1 by vol.) to give 7 g of small
white crystals m. p. 228-229° of the sodium salt of 1-naphthylacetyltaurine.
The crystals gave a strong positive sodium flame test and did not react
with sodium bicarbonate. C14H14NO
4SNa requires C, 53. 33 ; H, 4.44 ;
N, 4.44 ; S, 10.16 ; Na, 7.30. Found C, 53.35; H, 4. 53 ; N, 4. 39 ;
S, 9.96 ; Na, 7.25.
The compound was shown to afford 1-naphthylacetic acid and taurine on
acid hydrolysis as follows : 10 mg of the compound was heated with 5N-HCI
(1 ml) in a sealed tube at 120° for 18 h. The hydrolysate was reduced to
dryness and the residue dissolved in water (0.25 ml). Portions (50 pd.)
of the latter were chromatographed on Whatman No. 4 paper using solvent
systems B and C. Spraying the chromatogram with ninhydrin revealed a
purple spot of Rf value 0.12 and 0.41 in solvents B and C respectively
which corresponded with taurine. Further portions of the hydrolysate
were chromatographed on thin-layer silica gel plates using solvent system
F. When viewed beneath u. v. light there appeared a dark purple spot Rf
0. 88 which corresponded to 1-naphthylacetic acid.

58
The infra-red spectrum (Nujol showed prominent absorption bands at
3260 cm-1 (N - H stretch), 1640 (Amide I band C = 0 stretch) 1555 (Amide
II band, C - N stretch), 1220 - 1165 (broad) and 1065 - 1020 (broad) (both
due to S = 0 stretch).
The mass spectrum of the methyl ester showed prominent peaks at m/e
185 (relative intensity 44%), 167 (12), 166 (9), 142 (74), 141 (100), 139
(23), 115 (46), 69 (20), 63 (11) and 44 (95) (cracking pattern shown in
Appendix).
1-Naphthylacetylglucuronide A total of 9 g of 1-naphthylacetic acid was
fed to three rabbits. The glucuronide gum was prepared from the
pooled 24 h urine, using the basic lead salt procedure of Kamil, Smith and
Williams (1952). The gum gave an intensely positive reaction with
naphthoresorcinol and was strongly reducing towards Fehlings and
Benedict's solution. When a small portion of the gum was incubated
over-night in pH 5 0. 5M-acetate buffer at 37° with 13 -glucuronidase or was
warmed with 2N-NaOH for 5 min it was shown chromatographically to
afford 1-naphthylacetic acid. The crude gum was used as standard
for chromatography. A small portion (about 1 g) was treated with
ethereal diazomethane and acetylated by method of Kamil et al. (1952)
but no crystals were obtained . No further characterisation was
undertaken.
Synthesis of diphenylacetic acid conjugates
Diphenylacetylglycine was prepared by reacting diphenylacetylchl.oride
(50 g) for 2 h with glycine (13 g) dissolved in N-NaOH (24 ml). The
reaction mixture was acidified with 2N-HC1 and the product that separated,
filtered washed with chloroform and recrystallised from aqueous ethanol to give

59
white needle crystals of diphenylacetylglycine (m. p. 141 - 142° ;
Miriam et al. ,1927a quote m. p. 157°). C16H1503N requires C, 71. 40;
H, 5. 62 ; N, 5. 20. Found C, 71. 16 ; H, 5. 71 ; N, 5. 12. Equivalent
weight by titration (0.1 N NaOH), 270 (required 269). The mass spectrum
of the methyl ester showed a molecular ion at m/e 283 (relative intensity 3)
with prominent peaks at 77 (22), 91 (49), 105 (68), 106 (100), 116 (8),
166 (11) and 167 (8) (for cracking pattern, see Appendix).
Diphenylacetyl-L-glutamine. Diphenylacetyl chloride (23 g), was added
gradually over a period of 3 h to a well stirred solution of L-glutamine
(13 g) dissolved in 100 ml of water containing sodium bicarbonate (23 g).
The reaction mixture after leaving overnight was filtered and acidifed with
2N-HCI. The precipitate was filtered, washed with 2N-HC1 and ether
and crystallized from water to give white crystals of diphenylacetyl-L-
glutamine (yield 12 g ; m. p. 149°). C19H2004N2 requires C, 67. 04 ;
H, 5. 92 ; N, 8.22. Found C, 67.1 ; H, 6. 07 ; N, 8. 31. Equivalent
weight by titration (0.1 N-NaOH) 342 (requires 342).
The mass spectrum of the methyl ester showed a molecular ion at rale of
355 (M +1; relative intensity, 0.2%) with prominent peaks at 155 (98),
165 (100), 166 (62), 167 (100), 168 (100), 169 (47) and 187 (62) (cracking
pattern, see Appendix).
Diphenylacetyltaurine . Diphenylacetyl chloride (25 g) was gradually added
with stirring to an ice-cold solution of taurine (11 g) dissolved in N-NaOH
(120 ml) over a period of 5 h and left to react overnight. The reaction
mixture was filtered and the filtrate acidified with 2N-HC1 and extracted
with ether ( 3 x 50 ml). The aqueous phase was separated and reduced to

60
dryness and extracted with hot methanol (250 ml). On cooling white needle
crystals of the sodium salt of diphenylacetyltaurine separated (24 g) m. p.
200 205 . The crystals gave a positive flame test for sodium and did not
give CO2
when treated with sodium bicarbonate. C16
H16
NO4
SNa requires,
C, 56.29 ; H, 4.73 ; N, 4.10 ; Na, 6.73 and S, 9.39. Found C, 56. 30 ; H,
4.74 ; N, 4. 04 ; Na, 6. 74 and S, 9. 38. The compound was shown to afford
diphenylacetic acid and taurine on acid hydrolysis as follows : the compound
(10 mg) was heated with 5N-HC1 (1 ml) in a sealed tube at 120 for 18 h. The
tube was broken, the hydrolysate removed and evaporated to dryness. The
residue was dissolved in water (0.25 ml) and portions (501/1) chromatographed
on Whatman No. 4 paper using solvent systems B and C. The dried chromato-
grams showed on treatment with ninhydrin a purple spot at Rf values 0. 11 and
0.42 in solvents B and C respectively which corresponded with taurine. Further
portions of the hydrolysate were subjected to t. 1. c. using solvent system F.
The developed chromatograms showed a dark purple spot Rf 0. 92 when
viewed beneath ultra violet light which corresponded to diphenylacetic acid.
Its infra red spectrum (Nujol) showed prominent absorption bands at 3380
N H stretch), 1650 (amide I band C = 0 stretch), 1510 (amide II band C - N
stretch), 1230 - 1170 (broad), and 1070 - 1050 (broad) (both due S = 0 stretch).
The mass spectrum of the methyl ester of diphenylacetyltaurine showed
prominent peaks at m/e 169 (relative intensity 81), 168 (53), 167 (94), 166 (17),
165 (48), 153 (8), 152 (29), 91 (6), 77 (8), 69 (17), 63 (9) and 44 (100)
(cracking pattern, see Appendix).
Dinhenylacetylglucuronide was isolated from the urine of rabbits dosed
with diphenylacetic acid. The glucuronide gum was prepared by the d
method of Kamil, Smith and Williams (1952) from the combine/24 11 urine
samples collected from three rabbits each fed 3 g of diphenylacetic acid.

61
The gum gave an intensely positive reaction with naphthoresorcinol and was
strongly reducing towards Fehling's and Benedict's solution. When
small portion of the gum was incubated overnight in pH 5 0. 5M-acetate
buffer at 37° with p-glucuronidase or was warmed with 2N-NaOH for 2 min
it was shown chromatographically to afford diphenylacetic acid. The
glucuronide gum was used as standard for chromatography.
A small portion of the gum was characterised as follows: about 1 g was
treated with ethereal diazomethane. Following removal of the solvent
the residue was recrystallised from water to give white crystalline needles
(200 mg) of diphenylacetylglucuronide methyl ester m. p. 175°. It gave a
positive reaction with -naphthoresorcinol and was strongly reducing
towards Fhlings and Benedicts reagents. C21H2208 requires C, 62. 68 ;
H, 5.51. Found C, 62.60 ; H, 5.44.
The mass spectrum of the diphenylacetylglucuronide methyl ester did not
show a molecular ion and gave peaks at m/e 207 (2), 173 (13), 168 (18),
167 (100), 166 (14), 165 (37), 91 (3), 90 (8) and 43 (100) (cracking pattern,
see Appendix).
Synthesis of (±) hydratropic acid conjugates
(±) - Hydratropoylglycine was prepared by treating a solution of glycine in
aqueous sodium bicarbonate with hydratropic acid chloride (Kay and Raper,
1922). It was recrystallised from aqueous ethanol to give white crystalline
needles m.p. 102 - 103° (lit. 103°). C11
l-11303N requires C, 63. 76 ; H, 6.32,
N, 6. 75. Found C, 63. 72 ; H, 6. 38 ; N, 6. 73. Equivalent weight by
titration (0. 1 N NaOH) 208 (requires 207).
The mass spectrum of the methyl ester showed a molecular ion at m/e
221 (relative intensity, 84%) with prominent peaks at m/e 116 (94) , 105 (96)

62
and 91 (100).(cracking pattern, see Appendix).
(±)-Hydratropoyl-L-glutamine Hydratropic acid chloride ( 34 g) was added
dropwise to a well-stirred solution of L-glutamine (24 g) in water (240
ml) containing sodium bicarbonate (50 g). After 3h the reaction mixture
was transferred to a separating funnel and the aqueous layer was
separated. The latter was acidified with 2N-HCI and reduced to dryness
using a rotary evaporator. The residue was extracted with hot methanol
(200 ml) and the extract reduced to dryness. The residue was
recrystallised from water to give white needle crystals (yield 30 g ; m. p.
139 -140 °).C141118 04N2 requires C, 60.42 ; H, 6. 52 ; N, 10. 06.
Found C, 60.50 ; H, 6.36 ; N, 10.43.
Equivalent weight by titration (0.1 N-NaOH) 280 (requires 278).
The mass spectrum of the methyl ester showed a molecular ion at m/e
of 292 (relative intensity 4%) with prominent peaks at 187 (54), 155 (98),
116 (96) and 91 (94) (cracking pattern, see Appendix).
(d)-Hydratropoyltaurine Hydratropic acid chloride (27 g) was added drop-
wise to an ice-cooled and well-stirred solution of taurine (15 g) in N-
NaOH (120 ml) and the stirred mixture allowed to react overnight. The
aqueous layer was separated from the oil that separated and acidified with
2N-HCI and evaporated to dryness on a rotary evaporator. The residue
was extracted with hot methanol (200 ml.) and the methanol extract
separated and reduced to dryness. The residue was recrystallised from
methyl ethyl ketone to give white crystals m. p. 88 - 89° (yield 20 g, 50%).
C11
H15
NO4S requires C, 51.35 ; H, 5.87 ; N, 5.44 ; S, 12.46. Found
C, 51.25; H, 5.89; N, 5.35 ; S, 12.42.
The compound was shown to afford hydrotropic acid and taurine on acid

63
hydrolysis as follows: 10 mg of the compound was heated with 5N-HC1
(1 ml) in a sealed tube at 120° for 18 h. The hydrolysate was reduced
to dryness and the residue dissolved in water (0.25 ml). Portions
(50 aul) of the latter were chromatographed on Whatman No. 4. paper
using solvent systems E and F. Spraying the chromatograms with
ninhydrin revealed a purple spot of Rf values 0.12 and 0.41 in solvents
B and C respectively which corresponded with taurine. Further
portions of the hydrolysate were chromatographed on thin-layer silica"
gel plates using solvent F. When viewed beneath ultra-violet light there
appeared a dark purple spot Rf 0. 87 which corresponded to hydratropic
acid.
Its infra-red spectrum (Nujol) showed prominent absorption bands at
3440 ( N - H stretch), 1650 (amide I band C = 0 stretch), 1570 (amide
II band C - N stretch), 1200 - 1140 (broad) and 1060 - 1010 (broad)
(both due to S = 0 stretch).
The mass spectrum of the methyl ester of hydratropoyltaurine gave a
molecular ion at m/e 271 (relative intensity 76%, with prominent peaks
at m/e 166 (90), 134 (80), 105 (96) and 91 (100) (cracking pattern, see
Appendix).
(±)-Hydratropoylglucuronide A total of 6 g. of hydratropic acid was fed
to three rabbits and the urine collected for 24 h. The glucuronide gum
was prepared from the pooled urine using the basic lead salt procedure
of Kamil, Smith and Williams (1952). The gum was dissolved in methanol
(5 ml) and ether (250 ml) followed by petroleum ether (b. p. 40-60°). After
standing for six months a white solid separated which was recrystallised
from water to give white needle crystals of (±)-hydratropoylglucuronide
and this was used as standard for chromatography m.p. 163° (lit.163-164°).

64
It reduced Benedict's and Fehling's solution readily and gave a
strongly positive naphthoresorcinol test. On warming with N-NaOH for 1 h
or incubating with f3 - glucur onidase in pH 5,0.5M - acetate buffer over-
night at 37° the conjugate was shown by chromatography to afford
hydratropic acid and glucuronic acid.
The conjugate was further characterised as its methyl ester following the
treatment of the remaining filtrate (see above) with diazomethane. The
solvents were removed by evaporation and the residue recrystallised
from water to give white needle crystals of (±)-hydratropoylglucuronide
methyl ester m.p. 164° (lit. 165 - 166°). It reduced both Fehlings and Benedicts
solution on warming and gave an intensely positive reaction with naphthore-
sorcinol. C16H2008 . H2O requires C, 53. 67 ; H, 6.18. Found C, 53. 89;
H, 5.76. Loss on drying at 110°, 5.08% (requires 5. 03%).
The mass spectrum showed a molecular ion at m/e of 356 (relative
intensity 2%) with major peaks at 191 (18), 173 (64), 105 (100) and 91 (16)
(cracking pattern, see Appendix).
Benzylamine salt of (±)-hydratropic acid was prepared for reverse
isotope dilution experiments with hydratropic acid. Hydratropic acid
(1 g) dissolved in ethyl acetate (3 ml) was added to benzylamine (1.5 g),
dissolved in hot ethanol. The mixture was heated for 5 min and the
solvent then removed using a rotary evaporator. The residue was
recrystallised from petroleum ether (b. p. 100-102°) to give white needle
crystals of the benzylamine salt of (±)-hydratropic acid m.p. 88° (yield 1. 8 g).
C16 H19 02 N requires C = 74. 68 ; H, 7.44 ; N, 5.44. Found C, 74.80 ;
H, 7. 37 ; N, 5. 24.

65
Metabolic Studies
Animals
All the species were obtained from dealers in the London area.
Equimolar amounts of _14 _14 C]-1-naphthylacetic acid (100 mg/kg) and C]-
diphenylacetic acid (114 mg-/kg) were given as aqueous solution dissolved in
the calculated amount of N-NaOH. [14Crilydratropic acid (81 mg/kg) was given
in a solution of /3-propylene glycol/water (2:1 v/v). For subhuman primates
the dose solution was sterilised by ultrafiltration prior to use. In the case
of human subjects the dose was 5 mg administered orally. The 0-24 and
24 - 48h urines were collected and adjusted to pH 5 with glacial acetic acid prior
to storage at 0°. Animals were kept in metabolism cages, which allowed the
separate collection of urine, and faeces, and maintained on an appropriate diet
with free access to water.
Bile-duct cannulated female rats were prepared as described by
Abou-El-Makarem et al. , (1967) and were injected intraperitoneally with the
dose solution (5-500 mg/kg). The bile was collected for 3 h and urine was
collected by bladder puncture.
Collection of carbon dioxide in the expired air
14 _ Rats dosed with either L C]-1-naphthylacetic acid or [14
Cj-diphenyl-
acetic acid were placed in Metabowl cages (Jencons) which allowed the collection
of expired air as well as urine and faeces. The expired air was drawn through a
drying trap of anhydrous CaC12 and then through two traps each containing 100 ml
of a 1:2 by vol. solution of ethanolamine (redistilled) in 2-methoxyethanol, (Jeffay
and Alvarez, 1961), at a rate which just prevented condensation inside the
Metabowls.
Paper and. Thin-Layer Chromatography
The solvent systems used were as follows : -

66
A. Butan-1-ol saturated with water
B. Butan-1-ol : acetic acid : water (4:1:1, by vol).
C. Propan-1-ol : ammonia (sp. gr. 0. 88) (7:3 v/v)
D. Benzene : acetone : acetic acid (2:2:1, by vol. )
E. Benzene : acetone : acetic acid (6:2:1, by vol).
F. Chloroform : methanol : acetic acid (24:8:1, by vol.)
Whatman No. 1 paper chromatograms were developed in solvent A
using descending technique , for identifying 1-naphthylacetic acid and its
conjugate. Whatman. No. 4 paper chromatograms were developed in solvent
systems B and C using the ascending technique for identifying taurine. Thin -
layer chromatograms, (aluminium backed silica gel 60 F254 plates, E. Merck
A. G. Darmstadt, Germany ; 0. 2 mm thick) were developed in solvent systems
D, E and F for identifying the arylacetic acids and their respective conjugates.
Location of Compounds on Chromatograms
Ultra-violet (U. V.) light
All the arylacetic acids and their conjugates considered were seen as
dark purple spots under U. V. lights (254 nm ; Hanovia Chromatolite, Slough,
Bucks. , U. K. ).
Spray Reagents
Naphthoresorcinol spray (Bridges, Kibby and Williams, 1965)
Chromatograms were sprayed with naphthoresorcinol (4% w/v) in acetone
to which phosphoric acid (10%) was added (4:1 , v/v) just before use. Glucuronides
showed up as blue spots on heating at 105° for 5 mins.
4-Dimethylaminobenzaldehyde spray (DMAB)
Chromatograms were sprayed with 4-dimethylaminobenzladehyde dissolved
in acetic anhydride (4% w/v) containing a little sodium acetate. After spraying

67
the chromatograms were gently heated with a hot-air blower, and glycine
conjugates showed up as orange spots.
Chlorine-starch/potassium iodide detection reagent
The chromatograms were exposed to chlorine generated from conc. HC1
and sodium hypochlorite for 30 min, aired and then sprayed with 1% (w/v)
aqueous solution of potassium iodide containing 1% (w/v) starch ; and amino acid
conjugates appeared as purplish brown spots.
Ninhydrin spray
Chromatograms were sprayed with 0. 3% ninhydrin in acetone. Amino
acids showed up as purple spots.
Chromatographic properties of taurine
Taurine has Rf values of 0.12 and 0.41 on Whatman No. 4 paper
chromatograms developed in solvent systems B and C respectively.
Radiochemical Techniques
The 14
C in the samples was determined using Packard Tri-Carb
Scintillation Spectrometers (models 3214 and 3320) and dioxan scintillator prepared
as described by Bridges et al. (1967). Urine (0.01 - 1 ml), bile (0. 01 - 0. 05 ml)
and cage washing (0. 05 - 1. 0 ml) were counted in triplicate. The radioactivity
in the expired carbon dioxide was estimated by counting 2 ml of the absorbent in
15 ml of a toluene : 2-methoxyethanol ( 2 : 1 v/v), scintillation medium containing
2, 5-diphenyloxazole (PPO, 5.5 g/l).
For the radiochromatogram scarmingthe urine (0. 01 - 0.2 ml) or bile
(0. 01 ml) containing about 1 x 104 d. p. m. was streaked on the chromatogram.
After development of the chromatograms they were scanned in a Packard
Radiochromatogram Scanner (Model, 7200). In some experiments the chromato-
grams were cut into sections (1 cm) and each counted in the scintillation counter
to enable a quantitation of the metabolites. Histograms were then plotted of the

Table 2. 1
Chromatographic Properties of 1 -Naphthylacetic Acid and its Conjugates
Technique: Compound
solvent:
Paper Thin-layer
A E F
1 -Naphthylacetic acid 0. 84 0. 86 0. 64
1 -Naphthylacetylglycine 0.46 0. 63 0. 37
1 -Naphthyl a cetyl glutamine 0. 24 0.40 0. 13
1 -Naphthylacetyltaurine 0.15 0.18 0. 0
1 -Naphthylacetylglucuronide 0.09 0. 0 0. 0
Rf value1
Colour reactions 1 i
Appearance Naphthoresorcinol Chlorine-
beneath U. V. spray DIVIAB starch/K1
reagent
dark purple
dark purple
dark purple
dark purple
dark purple
orange
blue
purplish brown
purplish brown
purplish brown

Table 2.2
Thin-Layer Chromatographic Properties of Diphenylacetic Acid and its Conjugates
Rf value Colour reactions
Compound Appearance
beneath U. V. solvent : D E F
Diphenylacetic acid 0.88 0.61 0.92
Diphenylacetylglycine 0.73 0.42 0.73
Diphenylacetylglutamine 0.54 0.21 0.35
Diphenylacetyltaurine 0.28 0.00 0.43
Diphenylacetylglucuronide 0.10 0.00 0.17
Naphthoresorcinol
spray DMAB
Chlorine - star ch/K1 reagent
dark purple
dark purple
dark purple
dark purple
dark purple blue
orange purplish brown
purplish brown
purplish brown

Appearance
beneath U. V.
Naphthore sorcinol
spray DMAB
Chlorine -starch/Kt reagent
dark purple
dark purple
dark purple
dark purple
dark purple
orange
blue
purplish brown
purplish brown
purplish brown
Table 2.3
Thin-Layer Chromatographic Properties of (±)Hydratropic Acid and its Conjugates
Rf value
Compound Solvent : D E F
(Li) -Hydratropie acid 0.75 0.75 0.87
(±)-Hydratropoylglycine 0.65 0.42 0.79
(±) -Hydratropoylglutamine 0. 53 0.18 0. 59
(1) -Hydratropoyltaurine 0. 30 0. 00 0. 33
(+)-Hydratropoylglucuronide 0.10 0. 00 0.13
Colour reactions

71
amount of C associated with each position on the original chromatogram. In
all the cases, quench correction was by channels ratio method.
Spectra
Infra-red (i. r.) spectra were recorded as liquid paraffin mulls on a
Perkin-Elmer Infracord 137 Spectrophotometer.
Mass spectra of compounds using direct insertion technique were
recorded on a Varian MAT CH5 Mass spectrometer : probe temperature
100 - 125°C ; chamber temperature, 160°C ; ionization energy, 70 eV ; ionization
current, 300 p.A.
Treatment of urine samples
Urine samples were chromatographed as obtained and following alkaline
hydrolysis or treatment with p-glucuronidase as follows:
(a) mild alkaline hydrolysis ; urine ( 1 ml ) was heated on a boiling water
bath with N-NaOH ( lml ) for 15 min. The solution was neutralised with 2N-
HCl and a portion chromatographed. This treatment was shown to
hydrolyse completely the glucuronic acid conjugates of the arylacetic
acids to their respective aglycones.
(b) j3-glucuronidase treatment ; urine ( 2 ml) was incubated with ketodase
( 1 ml ) and pH 5 0. 5 M-acetate buffer at 37° for 18 h. As a control
on the activity of the enzyme a tube was included containing phenolphthalein
glucuronide ( 1 mg ) dissolved in water ( 1 ml ) instead of urine.
At the end of the experiment the incubate was made alkaline with N-NaOH.
Appearances of a deep red-purple colour indicated that enzyme preparation
was active. Under these conditions arylacetylglucuronides were hydrolysed
to the free acids.

72
Pharmacokinetic Studies
Animals
Female Dutch rabbits weighing 2. 0 - 2. 5 kg ; (ay. wt. 2. 2 kg) were
used.
Cannulation of marginal vein
For intravenous administration of compounds and collection of blood the
marginal ear vein of the rabbit was cannulated as follows: The animal was
first restrained by being wrapped in a thick cotton cloth (100 x 100 cm) so that only
the head and the ears were exposed. The hairs around the vein were shaved
and xylene was used to dilate the blood vessel. A puncture was made in the vein
using a Gillette Scimitar disposable hypodermic needle ( 19 g x 5 cm ) and the
cannula inserted 2 - 3 cm into the vessel. The cannula was held in position
with an adhesive plaster.
The cannula was made from a 20 cm length of polythene tubing (Portex
Pp-60) which has been heated in an oven at 100°C and drawn out at one end. The
narrow end was cut at an angle to provide a sharp point. The wider end was
attached to a hypodermic needle (21 g x 3. 8 cm) and syringe containing heparinized
saline (500 I. U. /ml of heparin) (Paine s and Byrne Ltd. , Greenford, U.K.).
This enabled the cannula to be kept filled with saline medium without clotting.
After cannulation, the cannula was clamped with small (12 cm ) pair of artery
forceps to prevent the outflow of blood. The injection needle and the syringe
were disconnected from the cannula and replaced by a 2 cm length of a suitable
wire to act as a plug for the cannula. The artery forceps were then released.
Administration of compounds
After withdrawal of a blank blood sample, the [14q-arylacetic acids
69 ptmol/kg ; 6 teCi/kg ) were injected through the cannula.

73
Blood Analysis for 14C content
After administration of the compounds, blood samples (2 ml) were then
withdrawn at different time intervals up to 3 h. The cannula was kept full of
heparinized saline throughout the experiment. Blood samples (0.1 ml) were
counted in a dioxan based scintillation fluid for the 14C content. Blood samples
were also examined for the presence of metabolites as follows: blood (1 ml) was
mixed with acetone (1 ml) and centrifuged at 200 x g for 5 min. The supernatant
(100 pl) was then subjected to thin layer chromatography using solvents D and F.
In Vitro Studies
Preparation of tissue subfractions
Female Wistar albino rats , weighing 180-220 g, were killed with a blow
on the head, their livers excised and placed on ice. After weighing quickly, the
livers were homogenised in 3 volumes of ice-cold 0.25 M sucrose (for isolation
of mitochondria) or in ice-cold 1.15% w/v KCI (for isolation of microsomes)
using an Ultra-Turrax homogeniser. This homogenate was centrifuged for 10
min at 700 x g at 4°, using an MSE High -Speed 25 centrifuge, to remove nuclei
and cell debris.
Mitochondria
A 700 x g supernatant of liver in 0.25 M sucrose, was layered over an
equal volume of ice-cold 0.34M sucrose, and this centrifuged at 5000 x g for 10
min at 4°, using an MSE High Speed 25 centrifuge. The crude mitochondrial pellet
so obtained was redispersed in 1.15% KCI and centrifuged again using the above
conditions. This washed mitochondrial pellet was resuspended in 1.15% KC1 so that
1 ml contained mitochondria from 250 mg of liver.
Microsomes
A 700 x g supernatant, in 1.15% KCI was centrifuged at 5000 x g for 15
min at 4°. The supernatant from this was then centrifuged at 100,000 x g for

74 60 min at 4° in an MSE Superspeed 40 centrifuge, the pellet so obtained redispersed
in 1.15% KCl and centrifuged again under the same conditions as before. The
washed microsomal pellet so obtained was resuspended in 1.15% KCl so that 1 ml
contained microsomes from 250 mg of liver.
Binding studies
In order to distinguish between real binding of drugs to subcellular
fractions and non-specific extrapment, the approach of Goldstein, Lowney and
Pal (1971) was used. Two sets of tubes were set up, one of which (A) contained
14 only the . Cl-arylacetic acid and the other of which (B) was incubated with a
large excess of unlabelled acid prior to addition of the _14. Cl-arylacetic acid. In
the first set of tubes, A, the [14C]-arylacetic acid will participate in all possible
14 kinds of interaction with the organelles. In the second set, B, the _ Cl-arylacetic
acid will be blocked from entering saturable sites, but, their non-specific inter-
actions (trapped and dissolved) will be unaffected. The difference, A - B, there
fore measures the saturable binding of the arylacetic acid to the organelles.
In this experiment, the incubation, A, contained mitochondria or
microsomes (equivalent of 1 g liver), 0.2M Tris-HC1 buffer, pH 7.4 (2. ml)
together with the _14C]-arylacetic acid (10 nmol ; 50,000 d. p. m.) in final volume
of 4 ml and were incubated for 15 min at 37° with shaking.
The incubation B contained mitochondria or microsomes as above with
unlabelled arylacetic acid (1 p.mol) in a volume of 4 ml . After incubation at 37°
_ for 15 min with shaking, the 14C]-arylacetic acid (10 nmol) was added and then
incubated for a further 15 min at 37°.
After incubation the organelles were isolated by centrifugation as pre-
viously described and the supernatants removed and their [14C] content determined.
The incubation tubes were allowed to drain on tissue paper prior to resuspending
the pellet in 1.15% KCl (2 ml) for measurement of [14C] content.

75
Protein determination
The protein content of subcellular fractions were determined by the
Biuret method.
Enzyme Affinity
Glycine conjugation
The conjugation of the arylacetic acids with glycine was measured in
the rat liver homogenates and in an isolated mitochondria by an adaption of the
method of Caldwell et al (1976) for hippuric acid formation in human liver.
A typical incubation consisted of 14C-labelled acid ( 10nmol, 50,000 d. p. m. ),
glycine (60 mmol), MgC12 (3 pmol), glutathione ( 20 pmol) and mitochondria or
a whole liver homogenate (equivalent to 40m; of liver) in 0.2 M-Tris/HCl
buffer, pH 8.4 (1 ml ). After incubation for 30 min at 37° the reaction was
stopped by the addition of acetone ( 1 ml ) containing the appropriate carrier acid
( 5 mg) and its glycine conjugate (5 mg ). After centrifuging to remove the
protein (0. 05 ml) was chromatographed on thin-layer plates, and developed in
solvents E and F. The developed plate was viewed beneath U. V. light, and the
dark-quenching areas corresponding to the free acid and its glycine conjugate
were scraped from the plate, and the associated 14C was determined by liquid-
scintillation counting. For each compound optimal substrate and cofactor
conditions were determined.
Glucuronic acid conjugation
The liver microsomal conjugation with glucuronic acid of the arylacetic
acids was determined by the method of Dingell et al (1974). Typical incubation
mixture (3 ml) contained the [14q-arylacetic acid (10-500 nmol), UDPGA (1pmol)
and microsomes in 1. 15% KCI (corresponding to 100 mg liver ; 1. 5 mg protein
per incubation) in 0.2M phosphate buffer (pH as shown in results). These

76
mixtures were incubated in 50 ml ground-glass stoppered tubes at 37° for
various time intervals with shaking, and the reaction then stopped by adjusting
pH to 1.5 with 2M-HC1. The conjugation of the acids was then assayed by
solvent extraction of the free acid, leaving the glucuronide conjugates in the
aqueous phase.
With hydratropic and phenylacetic acids, extraction was with water-
saturated ether (2 x 15 ml) while with 1-naphthyl- and diphenyl-acetic acids,
water-saturated toluene ( 1x15 ml) was used. After centrifuging to separate
the phases, aliquots (2 ml) of each were counted for 14C as described earlier.
Reverse isotope dilution experiments
1 -Naphthylacetic acid, 1-naphthylacetyl-glycine and -glutamine,
diphenylacetic acid, diphenylacetyl-glycine and -glutamine, hydratropoyl-
glycine and -glutamine were all processed in the same way. 1 g of the appro-
priate acid, dissolved in aqueous NaHCO3 was added to urine samples containing
about 0. 5 - 1 /Xi of 14C. The urine was adjusted to pH 1 with 2N-HCl and the
precipitate that formed, filtered and recrystallised from water in the case of
1 --naphthylacetic acid m. p. 131; 1 -naphthylacetylglutamine m. p. 186°, hydra -
tropoyl-glycine m. p. 103° and -glutamine in. p. 139°, diphenylacetic acid m. p.
147° and diphenylacetylglutamine m. p. 149°, and from aqueous ethanol in the case
of 1 -naphthylacetylglycine m. p. 149°, and diphenylacetylglycine m. p. 142° until
constant specific activity was achieved.
In the case of 1-naphthylacetyltaurine, diphenylacetyltaurine and hydra-
tropoyltaurine, ig of the appropriate compound was added to urine samples and
the whole reduced to dryness on a rotary evaporator. The residue was treated
with hot methanol (5m1) and filtered. For diphenylacetyltaurine, the m3thanolic
extract was allowed to cool and the crystals that separated were recrystallised

77
from methanol to constant specific activity, m. p. 200-205°. In the case of
1-naphthylacetyltaurine the methanolic extract was treated with acetone (3 ml)
and the white crystals that separated were recrystallised from methanol/acetone
(2 : 1 v/v) to constant specific activity, m. p. 229°. The methanolic extract
for hydratropoyltaurine, was further reduced to dryness and the residue was
recrystallised from methyl ethyl ketone to constant specific activity m. p. 89°.
Hydratropic acid was treated differently. 1 g of the acid was added
to urine samples and the whole reduced to dryness. The residue was taken up
in ethylacetate (5 ml) and filtered. The filtrate was added to benzylamine (1.5g)
in boiling ethanol (25 ml) and allowed to boil for 5 min. Solvents were removed
using a rotary evaporator. The crystals of benzylamine salt of hydratropic acid
that formed were triturated with petroleum ether (b. p. 40-60°) filtered and
recrystallised from petroleum ether (b. p. 100-120°) to constant specific
activity, m. p. 88°.

78
CHAPTER THREE
Metabolism of 1 - Naphthylacetic Acid
Contents
Pages
Identification of Urinary Metabolites of 1-Naphthylacetic acid
79
Man 79 Rhesus and cynomolgus monkey 81 Capuchin, marmoset and squirrel monkey 84 Bushbaby 84 Cat 89 Rabbit and rat 91 Fruit bat 91
Influence of Dose on the Pattern of Metabolism and Excretion of 1-Naphthylacetic Acid in the Rat 94
Urine 94 Bile 94
Results
94
Man 95 Rhesus monkey 95 Cynomolgus monkey 95 Squirrel monkey 95 Capuchin 96 Marmoset 96 Bushbaby 96 Cat 96 Rabbit 96 Rat 97 Fruit bat 97
Influence of dose on the pattern of metabolism and excretion of 1-Naphthylacetic acid in the rat 97
Urinary excretion products 97 Biliary excretion products 100
Discus sion 100

79
Tne Metabolism of 1-Naphthylacetic Acid
The metabolic fate of 1-naphthylacetic acid has been studied in man,
6 sub-human primates and 4 non-primates . Additionally the influence of dose
on the pattern of metabolism and excretion of this acid has been studied in the
rat.
Identification of urinary metabolites of 1-naphthylacetic acid
The urines of animals dosed with 1-naphthylacetic acid were subjected to
paper and thin-layer chromatography as described in Chapter 2.
Man
Paper chromatography followed by radiochromatogram scanning of
human urine samples using solvent A revealed the presence of a large 14C peak
at Rf 0. 09 (Fig. 3:1). This gave a positive reaction with naphthoresorcinol
and disappeared when the urine was treated with p-glucuronidase or N-NaOH, to
be replaced by a new 14C peak at R
f 0. 84, corresponding to 1-naphthylacetic
acid. A portion of the glucuronide gum obtained from rabbits fed with 1-naphthyl-
acetic acid also showed on chromatography as above a naphthoresorcinol positive
spot at Rf 0. 09 which on treatment with p-glucuronidase or N-NaOH afforded
1-naphthylacetic acid (see chapter 2). The radioactive component found in the
urine with Rf 0. 09 was therefore identified as 1-naphthylacetylglucuronide.
There was also a small 14C peak at Rf 0.14 corresponding to 1-naphthylacetyl-
taurine. Similarly, thin-layer chromatography in solvent D showed the presence
on scanning of a single large 14C peak at the origin which gave a positive
reaction with naphthoresorcinol and was labile to p-glucuronidase or N-NaOH
treatment to afford 1-naphthylacetic acid (R1 0. 82). Reverse isotope dilution also
confirmed the presence of 1-naphthylacetyltaurine, and the presence of 1 -naphthyl - acid
acetic/was confirmed after the treatment of urine samples with p-glucuronidase or
N-Na0H.

0 S F.
2 3
after mild alkaline hydrolysis
S. F.
80
Fig. 3.1 Radiochromatogram scan of urine from man dosed orally
[14C]-1-naphthylacetic acid (5 mg ; 5 tiCi) and chromatographed
on Whatman No. 1 paper in solvent system A.
1. 1-Naphthylacetylglucuronide; 2. 1-naphthylacetyltaurine;
3. 1-naphthylacetic acid; 0 = origin; S. F. = solvent front

81
Rhesus and cynomolgus monkeys
Scans of chromatograms of rhesus monkey urine developed on Whatman
No. 1 paper in solvent A showed a single broad peak at Rf 0.09 and a minor peak
at Rf 0. 84 (Fig. 3.2). The major peak at Rf
0.09 gave a positive reaction with
naphthoresorcinol and disappeared when the urine was treated with g glucuronidase
or N-NaOH giving a 14C peak at Rf
0.85 which corresponded to 1-naphthylacetic
acid. The 14
C peak at Rf 0.09 was therefore identified as 1-naphthylacetyl-
glucuronide. There was a small 14C peak at R 0.14 corresponding to 1-
naphthylacetyltaurine, and the small peak in untreated urine at Rf
0. 84 was
identified as 1-naphthylacetic acid. Chromatography on thin-layer using solvent
D similarly revealed the main 14
C compound present to be 1-naphthylacetyl-
glucuronide together with small amounts of 1-naphthylacetyltaurine and 1-naphthyl-
acetic acid. The presence of these compounds was also confirmed by reverse
isotope dilution.
Chromatograms of cynomologus monkey urine developed on Whatman No.1
paper in solvent A showed on radiochromatogram scanning two major 14C peaks
(Fig. 3:3) ; at Rf values 0.09 and 0.84. The peak at Rf 0.09 gave positive
reaction with naphthoresorcinol and disappeared on treatment of urine samples
with j3-glucuronidase or N-NaOH and this was accompanied by an increase in
the size of the peak at Rf 0.84 due to 1-naphthylacetic acid. The peak at Rf
0.09 was therefore identified as 1-naphthylacetylglucuronide. There were also
two small peaks at Rf 0.14 and 0.26 corresponding to 1-naphthylacetyltaurine
and 1-naphthylacetylglutamine respectively.
14C scans on thin-layer chromatograms of cynomolgus monkey urine
developed in solvent D showed a large 14
C peak at the origin due to 1-naphthyl-
acetylglucuronide, and a second large 14C peak due to the unchanged acid at

after mild alkaline hydrolysis
3 S. F.
S. F.
82
Fig. 3.2 Radiochromatogram scan of urine from rhesus monkey
dosed intramuscularly with [14 C]-1-naphthylacetic acid
(100 mg /kg ; 17.2 pCi) and chromatographed on Whatman
NO. 1 paper in solvent system A.
1. 1 -Naphthylacetylglucuronide; 2, 1 -naphthylacetyltaurine;
3. 1-naphthylacetie acid; 0 = origin, S. F. = solvent front

0 S. F.
0 S. F.
after mild alkaline
hydrolysis 2 3 4
83
Fig. 3. 3 Radiochromatogram scan of urine from cynomolgus monkey
dosed intramuscularly with [14 C]-1-naphthylacetic acid
(100 mg/kg ; 21.2 pCi) and chromatographed on Whatman
No. 1paper in solvent system A.
1. 1 -Naphthy I acetylglucuronide ; 2, 1 -naphthylacetyltaurine ;
3. 1-naphthylacetylglutamine; 4. 1-naphthylacetic acid; 0 = origin ;
S. F. = solvent front

84
Rf 0. 86. Two minor
14C peaks appeared at R
f values 0.18 and 0. 38
corresponding to 1-naphthylacetyltaurine and-glutamine respectively. Reverse
isotope dilution also confirmed the presence of 1 -naphthylacetic acid, 1-
naphthylacetyl-taurine and - glutamine.
Capuchin, marmoset and squirrel monkey
Chromatography of urine samples on Whatman No. 1 paper using solvent
A revealed on radiochromatogram scanning the presence of five 14C peaks of Rf
values O. 09, 0.16, 0.25, 0.48 and 0. 86 (Figs. 3:4, 3:5 and 3:6). The 14
C
peak at 0. 09 gave a positive reaction with naphthoresorcinol and disappeared
when urine was treated with P-glucuronidase or N-NaOH to be replaced by a
larger 14
C peak at Rf 0.86, which corresponded to 1-naphthylacetic acid. This
peak at Rf 0.09 was therefore identified as 1-naphthylacetylglucuronide. The
14C peak at Rf 0.48 gave a positive reaction with the 4-dimethylaminobenzaldehyde
reagent and was identified as 1-naphthylacetylglycine. The 14C peaks of Rf
values, 0.16, and 0.25 corresponded to 1 -naphthylacetyl-taurine and-glutamine
respectively.
Similarly, chromatography on thin-layers using solvent D showed on
radiochromatogram scanning the presence of five 14C peaks, one at the origin
and others at Rf 0.18' 0. 40, 0.63 and 0.86 which corresponded to 1-naphthyl-
acetyl- glucuronide, taurine, glutamine, glycine and 1-naphthylacetic acid
respectively. The 14C peak at the origin disappeared from urine samples
treated with [3-glucuronidase or N-NaOH as above and it gave a positive
naphthoresorcinol reaction. The presence of 1 -naphthylacetyl- taurine, glutamine,
glycine and 1-naphthylacetic acid was also confirmed by reverse isotope dilution.
Bushbaby
14C scans of paper chromatograms of bushbaby urine developed with
solvent A showed three major radioactive peaks of R 0.08, 0.14 and 0. 44

0 S F.
0 S. F.
after mild alkaline hydrolysis
2 3 4 5
85
Fig. 3.4 Radiochromatogram scan of urine from capuchin monkey
dosed intramuscularly with [14C]-naphthylacetic acid
(100 mg/kg ; 8. 5 pCi) and chromatographed on Whatman
No.1 in solvent system A.
1. 1 -Naphthylacetylglucuronide; 2. 1-naphthylacetyltaurine
3. 1 -naphthylacetylglutamine; 4. 1-naphthylacetylglycine
5. 1-naphthylacetia acid . 0 = origin; S. F. = solvent front

0 S. F.
after mild alkaline hydrolysis
86
Fig. 3. 5
Radiochromatogram scan of urine from squirrel monkey
dosed intramuscularly with [14C]-1 -naphthylacetic acid
(100 mg/kg ; 5. 9 12 Ci) and chromatographed on Whatman
No.1 paper in solvent system A.
1. 1 -Naphthylacetylglucuronide ; 2. 1 -naphthylacetyltaurine ;
3. 1 -naphthylacetylglutamine; 4. 1 -naphthylacetylglycine
5. 1 -naphthylacetic acid. 0 = origin ; S. F. = solvent front

0 S. F.
after mild alkaline
hydrolysis
87
Fig. 3.6 Radiochromatogram scan of urine from marmoset
dosed intramuscularly with [14 C]-1-naphthylacetic
acid (100 mg/kg ; 5.9 iCi) and chromatographed on Whatman
No. 1 paper in solvent system A.
1. 1 -Naphthylacetylglucuronice; 2. 1-n.aphthylacetyltaurine
3. 1 -naphthylacetylglutaraine ;. 4. 1-naohthylacetylglycine
5. 1-naphthylacetic acid. 0 = origin ; S. F. solvent front

88
0
S. F.
0 Fig. 3. 7
10 cm 20 S F. Radiochromatogram scan and histogram of urine from bushbaby dosed intramuscularly with [14C]-1-naphthylacetic acid (100 mg/kg; 23. 6 p.Ci) and chromatographed on Whatman No. 1 paper in solvent system A.
1. 1- Naphthylacetylglucuronide ; 2. 1 -naphthylacetyltaurine
3. 1-naphthylacetylglycine ; 4. 1-naphthylacetic acid. 0 = origin ;
S. F. = solvent front

89
(Fig. 3:7) corresponding to 1-naphthylacetyl-glucuronide, -taurine and
-glycine respectively. The peak at Rf 0. 08 gave a positive naphthoresorcinol
reaction and disappeared from urine treated with 13-glucuronidase or N-NaOH
and was replaced by an enlarged peak at Rf 0. 84 corresponding to 1-naphthyl-
acetic acid. The peak at Rf 0. 08 was therefore identified as 1-naphthylacetyl-
glucuronide. The peak at Rf 0. 44 gave a positive reaction for a glycine
conjugate with 4-dimethylaminobenzaldehyde reagent and the peak at Rf 0.14
corresponded to 1-naphthylacetyltaurine. In addition to these three conjugates
urine also contained a small amount of 1-naphthylacetic acid as revealed by
the appearance of a minor 14
C peak at Rf 0. 84. Scans of thin-layer chromato-
grams developed in solvent D also revealed three major 14C peaks, Rf
0.0,
0.16, 0. 62 and a minor peak at 0. 84 corresponding to 1-naphthylacetyl-
glucuronide, -taurine, and-glycine and 1-naphthylacetic acid respectively.
Reverse isotope dilution also confirmed the presence of 1-naphthylacetyl-taurine
and - glycine and 1-naphthylacetic acid.
Cat
Radiochromatogram scanning of paper chromatograms of cat urine
developed in solvent A showed two major radioactive peaks of Rf
0.14 and 0.44
(Fig. 3:8) which corresponded to 1 -naphthylacetyl-taurine and -glycine respectively.
There also appeared a very minor peak at Rf 0. 84 corresponding to 1 -naphthylacetic
acid. Similarly, scans of thin-layer chromatograms developed with solvent D
revealed two major peaks of Rf
0. 15 and 0.60 corresponding 1-naphthylacetyl-
taurine and -glycine respectively and a minor peak at Rf 0. 85 due to unchanged
compound. Reverse isotope dilution also confirmed the presence of 1 -naphthyl-
acetyl-taurine and -glycine and 1-naphthylacetic acid.

90
0 S. F.
10 cm 20
S. F.
Fig. 3.8 Radiochromatogram scan and histogram of urine from cat dosed intraperitoneally with [14C]-1-naphthylacetic acid (100 mg/kg ; 14.8 pCi) and chromatographed on Whatman No. 1 paper in solvent system A.
1. 1 -Naphthylacetyltaurine; 2. 1 -naphthylacetylglycine
3. 1-naphthylacetic acid ; 0 = origin; S. F. = solvent front

91
Rabbit and rat
Radiochromatogram scans of rabbit and rat urine samples chromato-
graphed on paper and developed in solvent A revealed the presence of a large
14C peak at Rf 0. 09 and two smaller 14C peaks at Rf 0.46 and 0. 84 respectively
(Figs. 3:9, 3:10). The peak at 0. 09 gave a positive reaction with naphthoresorcinol
and disappeared when urine samples were treated with 13-glucuronidase or N-NaOH
Its disappearance was accompanied by an increase in the size of the peak at Rf
0. 84 which corresponded to 1-naphthylacetic acid. The peak at Rf 0.46 gave an
orange colour with the 4-dimethylaminobenzaldehyde reagent and was identified
as 1-naphthylacetylglycine. Similarly, scans of chromatograms developed in
solvent D showed a single large 14C peak at the origin and two smaller peaks at
Rf 0. 60 and 0. 86. As above the large peak at the origin disappeared when the
urine was treated with g-glucuronidase or N-NaOH to be replaced by an enlarged
peak at Rf 0. 86 corresponding to 1-naphthylacetic acid. The peaks at Rf 0.60
and 0. 86 were identified as 1-naphthylacetylglycine and 1-naphthylacetic acid and
there presence was also confirmed by reverse isotope dilution.
Fruit Bat
Paper chromatography of fruit bat urine followed by radiochromatogram
scanning showed the presence of a large 14C peak at R
f 0. 09 corresponding to
1-naphthylacetylglucuronide and a small peak at 111 0. 84 due to the unchanged
acid (Fig. 3:10). The peak at Rf 0. 09 gave a positive reaction with naphthore-
sorcinol and disappeared when urine samples were treated with (3-glucuronidase or
N-NaOH and this was accompanied by a large increase in the size of the peak at
Rf 0. 84 due to 1 -naphthylacetic acid. Chromatography on thin-layer in solvent D
showed the presence of 1-naphthylacetylglucuronide together with a small amount
of the unchanged acid.

92
Fig. 3. 9 Radiochromatogram scan and histogram of urine from
rat dosed intraperitoneally with [14C]-1-naphthylacetic
acid (100 mg/kg ; 5. 9 MCi) and chromatographed on Whatman
No. 1 paper in solvent system A.
1. 1-Naphtylacetylglucuronide; 2. 1-naphthylacetylglycine
3. 1-naphthylacetic acid 0 = origin; S.F. = solvent front

Fruit bat (dosed intraperitoneally, 10 ACi)
1
S. F. 0
93
Rabbit (dosed orally, 22. 5 ;lei)
2
S. F.
Fig. 3. 10 Radiochromatogram of urine from rabbit and fruit bat 14 _
dosed with [ C]-1-naplithylacetic acid (100 mg/kg )
and chromatographed on Whatman No. 1 paper in
solvent system A.
1. 1-Naphthylacetylglucuronide; 2. 1-naphthylacetylglycine;
3. 1-naphthylacetic acid; 0 = origin; S. F. = solvent front

94
Influence of dose on the pattern of metabolism and excretion of 1-naphthylacetic
acid in the rat
Bile and urine sample were collected over the first 3 h after dosing from
bile-duct cannulated rats dosed intraperitoneally with [14C1-1-naphthylacetic
acid (5 - 500 mg/kg) and were analysed for 14C and metabolites as described
in Chapter 2.
Urine - The thin-layer chromatograms of the urine samples developed in
solvent D revealed the presence of two major 14C peaks at Rf 0. 0 and 0. 62 and
a minor peak at Rf 0. 86. The peak at the origin gave positive reaction with
naphthoresorcinol reagent and disappeared when the urine samples were treated
with /3-glucuronidase or N-NaOH and this was accompanied by an increase in
the size of the peak at Rf 0. 86 corresponding to 1-naphthylacetic acid. The peak
at the origin was therefore identified as 1-naphthylacetylglucuronide. The 14C
peak at 0.62 gave an orange colour with 4-dimethylaminobenzaldehyde and was
identified as 1-naphthylacetylglycine. Thin-layer chromatograms developed
in solvent E confirmed the presence of 1-naphthylacetyl-glucuronide and -glycine
with very small amounts of unchanged acid.
Bile - The thin-layer chromatograms of bile samples developed in solvent D
revealed the presence of one major peak at the origin and a minor peak at Rf 0. 86.
The peak at the origin gave a positive reaction with naphthoresorcinol reagent
and disappeared on treatment of the bile samples with 13-glucuronidase or N-NaOH
and this was accompanied by an increase in the size of the peak at Rf 0. 86 due
to 1-naphthylacetic acid. The peak at the origin was therefore identified as
1 -naphthylacetylglucuronide.
Results
The quantitative and qualitative aspects of the excretion of 1-naphthylacetic

95
acid and its metabolites by the 11 species studied are shown in Table 3:1 and
those in rats at various dose levels are shown in Table 3:2.
Man - The two human subjects excreted in the urine about 97% of the radio-
activity in 24 h after an oral dose of 5 mg of [14
CJ-1-naphthylacetic acid. The
major excretion product was 1-naphthylacetylglucuronide (88 and 94% of the
dose respectively for the two subjects) with small amounts (7.8 and 3.4%) of
1-naphthylacetyltaurine. The glycine and glutamine conjugates of the acid
were not detected.
Rhesus monkey - The two rhesus monkeys excreted almost all the radioactivity
(99 and 100%) in 24 h after an intramuscular dose of 100 mg/kg of [140]-1-naphthyl-
acetic acid. The major excretion product was 1-naphthylacetylglucuronide
(83 and 94% of the dose respectively) with small amounts of 1-naphthylacetic acid
(12 and 6. 0%) and 1-naphthylacetyltaurine (3. 5 and 1%) but there were no
glycine or glutamine conjugate detected.
Cynomolgus monkey - The single cynomolgus monkey examined excreted 70% of
an injected dose (100 mg/kg, intramuscularly) of [14C]-1-naphthylacetic acid in
the urine within 24 h. The major excretion products were unchanged 1-naphthyl-
acetic acid (35% of the dose) and 1-naphthylacetylglucuronide (29%) with small
amounts of glutamine (2.6%) and taurine (3.5%) conjugates of the acid but the
glycine conjugate was not detected.
Squirrel monkey - The two squirrel monkeys examined excreted 39 and 22% of the
radioactivity in the urine within 24 h, after an intramuscular dose of 100 mg /kg
.14 f C1-1-naphthylacetic acid. The main radioactive compounds in the urine
were unchanged drug (13 and 7. 3% of the dose respectively), and glycine (7. 5 and
5.0), tauririe (7.5 and 5. 0), glucuronic acid (9.4 and 3.7%) and glutamine (2. 0 and
0.9%) conjugates of 1-naphthylacetic acid.

96
Capuchin The two Capuchin xamined excreted in the urine, 35 and 44% of
the injected dose (100 mg/kg, intramuscularly) of [14C]-1-naphthylacetic acid.
The major excretion products were the unchanged compound (16 and 4. 5% of
the dose respectively) taurine (14 and 15%) and glucuronic acid (2.2 and 13%)
conjugates with small amounts of glycine (2.2 and 8.8%) and glutamine (0.9 and
3.0%) conjugates of 1-naphthylacetic acid.
Marmoset - The single marmoset examined excreted 70% of an injected dose
(100 mg/kg, intramuscularly) of [14C]-1-naphthylacetic acid in the urine within
24 h. The major excretion products were 1-naphthylacetylglucuronide (49% of
the dose) 1-naphthylacetyltaurine (9.0%) and the unchanged compound (5.4%) the
with small amounts of/glutamine (1.6%) and glycine (1. 8%) conjugates of
1-naphthylacetic acid.
Bushbaby - The two bushbabies examined excreted 41 and 26% of the dose of
[14 C]-1-naphthylacetic acid (100 mg/kg, intramuscularly) respectively in the
urine in 48 h. The major excretion products were 1-naphthylacetylglycine
(21 and 4. 0% of the dose respectively), 1 -naphthylacetylglucuronide (11 and 3. 9%),
1 -naphthylacetyltaurine (4. 6 and 7. 5%) and some unchanged 1-naphthylacetic acid
(3.9 and 10.5%) but no glutamine conjugate of 1-naphthylacetic acid was detected.
Cat - Cats injected intraperitoneally with [14 C]-1-naphthylacetic acid (100 mg/kg)
excreted about 62% of the dose of radioactivity in the urine in 24 h. The major
excretion products were glycine (37% of dose) and taurine (25%) conjugates with
a small amount of the unchanged compound (1. 8%). 1- Naphthylacetylglucuronide
was not detected.
Rabbit - Rabbits dosed orally with [14
C]-1-naphthylacetic acid (100 mg/kg)
excreted about 81% of the dose of radioactivity in the urine in 24 h. The major
excretion product was 1-naphthylacetylglucuronide (71% of the dose) with small

97
amounts of 1 -naphthylacetylglycine (4. 5%) and 1-naphthylacetic acid (6.2%),
but no taurine conjugate of 1-naphthylacetic acid was detected.
14 . Rat - Rats injected intraperitoneally with [ C1-1- naphthylacetic acid (100 mg/kg)
excreted about 64% of the dose of radioactivity in the urine in 24 h. The major
excretion products were 1-naphthylacetylglucuronide (33% of the dose), 1-naphthyl-
acetylglycine (15%) and 1--naphthylacetic acid (17%). There was no taurine
conjugate detected.
In the experiment for the collection of expired carbon dioxide, there was
no radioactivity detected in the entrapped carbon dioxide indicating therefore,
that there was no metabolism by decarboxylation.
Fruit bat - The two fruit bats examined excreted 69 and 58% of the dose of
[14C]-1-naphthyIacetic acid (100 mg/kg, intraperitoneally) respectively in the
urine in 24 h. The major excretion product was 1-naphthylacetylglucuronide (62
and 56% of the dose respectively) with small amounts of the unchanged compound
(6.6 and 2. 0%). Neither the glycine nor the taurine conjugate was detected.
Influence of dose on the pattern of metabolism and excretion of 1-naplathylacetic
acid in the rat
Bile duct °annulated rats were injected intraperitoneally with [14C]-1-
naphthylacetic acid (5 to 500 mg/kg). From the dose level of 5 to 500 mg/kg,
the total amount of the radioactivity excreted in the urine in 3 h fell from 32 to 0.6%,
while the amount excreted in the bile fell from 37 to 10% of the dose respectively.
Urinary excretion products 1-Naphthylacetylglycine formed 88% of the excretion
products at 5 mg/kg dose level, but its proportion of the products fell to 11% at
the 500 mg/kg dose level. 1-Naphthylacetylglucuronide on the other hand formed
the minor metabolite at the lowest dose level (11%) but was the major metabolite
(88%) at the highest dose level. The unchanged acid formed a very small fraction

Species (NO. & Sex)
Primates
Man* (2M)
Rhesus monkey (1M,1F)
Cynomolgus monkey (1F)
Squirrel monkey (2F)
Capuchin (2F)
Marmoset (1M)
Busbaby (1M,1F)
Non-Primates
Cat (2F,1M)
Rabbit (3F)
Rat (3F)
Fruit bat (11', 1M)
Genus and Species Route of administration
Homo sapiens p. o.
Macaca mulatta i. m.
Macaca fascicularis (irus) i.m.
Saimiri sciureus i. m.
Cebus albifrons i. m.
Sanguinus oedipus i. m.
Galago crassicaudatus . i. m.
Fells cattus i. p.
Lepus caniculus p. o.
Rattus norvegicus i. p.
Pteropus giganteus i. p.
Dose of 14C /Xi/animal
14C excreted % of dose
5. 0 17.2
21.2
96, 97 99, 100
70
5. 9 39, 22
8. 5 35, 44
5.9 70
23.6 41, 26
14.8 62(57-69)
22. 5 81(76-87)
5. 9 64(59-70)
10.0 69, 58
* Dose 5mg + 48h excretion
Table 3 : 1
Conjugates of [14C1-1-naphthylacetic acid in various species
14 [ C]-1-naphthylacetic acid (100 mg/kg) dissolved in the appropriate amount of N-NaOH was administered as described in chapter two. The urine was collected for 2 days and the 0-24 h urine analysed for metabolites. Results are means for three animals with ranges in parentheses. Where only one or two animals were used individual values are given.
Amount of [14C] -1 -naphthylacetic acid excreted conjugated with various compounds in 24 h as % dose
Unconjugated Glutamine Glycine Taurine Glucuronic Acid
- - 7.8, 3.4 88, 94 12, 6 - - 3.5, 1 83, 94
35 2.6 - 3.5 29 13, 7.3 2.0, 0.9 7.5, 5.0 7.5, 5.0 9.4, 3.7 16, 4.5 0.9, 3.0 2.2, 8.8 14, 15 2. 2, 13
5.4 1.6 1.8 9.0 49 3.9, 10.5 - 21, 4 4.6, 7.5 11, 3.9
1. 8(1. 0-2. 2) - 37(29-48) 25(17-39) 6. 2(3. 6-10. 3) - - 4. 5(4. 0-5. 2) - 71(68-73) 17(15-18) - 15(10-21) - 33(31-34) 6.6, 2.0 - = - 62, 56
co

Table 3 :2
Influence of dose on the pattern of metabolism and excretion of 1-naphthylacetic acid it the rat
Bile-duct cannulated female rats were injected intraperit one ally with [14C I -1 -naphthylacetic acid (dissolved in the calculated amount of N-NaOH and diluted with water) and the bile collected for 3 h and urine removed from the bladder. Bile and urine were analysed for 14C and metabolites-as described in the text.
Urine Bile 14 I
% C excreted conjugated with: Dose (mg/kg) % dose excreted Unchanged Glycine Glucuronic
Acid
5 32 (31-33) 1.0 (0.8-1.2) 88 (84-91) 11 (8.2-15)
25 30 (27-37) 1.0 (0.9-1.1) 55 (49-60) 44 (38-49)
50 14 (11-16) 2.0 (1.8-2.1) 48 (43-52) 52 (47-56)
250 12 (8.2-13 5.1 (2.3-6.2) 12 (7.4-20) 85 (78-90)
500 0.6 (0.5-1.0) 2.4 (1.5-3.1) 11 (8.1-13) 88 (84-91)
r I
% dose
% 14C excreted conjugate
Unchanged Glycine
t
Glucuronic excreted Acid
37 (32-42) 5.3 (3.1-7.0) 4.1 (3.2-5.6) 91 (87-94)
34 (29-39) 5.9 (5.0-8.2) 1.0 (1. 0-1. 0) 92 (90-94)
44 (40-46) 5.1 (5.0-5.3) 1.0 (0.7-1.2) 94 (93-95)
15 (7.5-21) 8.4 (7.0-9.2) 1.0 (0.9-1.1) 91 (90-92)
10 (8.2-13) 16 (15-17) 1.1 (0.8-1.4) 84 (83-85)

100
of the urinary excretion products.
Binary excretion products. Irrespective of the dose level 1-naphthylacetyl-
glucuride was the major excretion product with only traces of 1-naphthylacetyl-
glycine. The unchanged acid rose from 5. 3% at the lowest dose level to 16%
at the highest level.
Discus sion
The metabolic fate of 1-naphthylacetic acid has been studied in eleven
species and the results show that 1-naphthylacetic acid can be conjugated at the
carboxyl group with amino acids and/or glucuronic acid, the pattern of which
varies with species. Table 3:3 summarises the conjugation pattern of 1-naphthyl-
acetic acid in the species studied and it suggests that man and the Old World
monkeys conjugate it mainly with glucuronic acid, the New World monkeys
(except the marmoset) and the bushbaby mainly with amino acids and to a small
extent with glucuronic acid, the cat with amino acids extensively, the rat and
rabbit principally with glucuronic acid and the fruit bat entirely with glucuronic
acid. The amino acids used were mainly glycine and taurine while the very
small amount of glutamine conjugation is restricted to the New World monkeys
and thecynomolgus monkey, a conjugation reaction which James et al (1972 a)
have reported to be restricted to man, New and Old World monkeys.
The taurine conjugation is predominant in the cat and the New World
monkeys and glycine conjugation also in the cat and bushbaby. James et al ,
(1972 a) have also reported that the taurine conjugate of phenylacetic acid is
a major metabolite in the New World monkeys, (except the marmoset), the
prosimians and in the ferrets. The quantitative distribution of the taurine
conjugate of 1-naphthylacetic acid in the species studied although haphazard,
was not detected in the fruit bat, rat and rabbit. Taurine conjugation has
previously been known to occur only with bile acids (see Sobotka 1937; Haselwood 1967),

Table 3:3
Species Variations in the pattern of conjugation of 1-Naphthylacetic acid.
1-naphthylacetic acid conjugates as % of the 14C excreted in the urine
Species Unchanged Glutamine Glycine Taurine Total conjugated with Glucuronic acid
Primates
amino acid
Man - - - 5.8 5.8 95
Rhesus monkey 8. 8 - - 2. 3 2. 3 89
Cynomolgus monkey 50 3. 7 - 5.0 8. 7 41
Squirrel monkey 33 4. 6 21 21 47 21
Capuchin 2. 9 4. 5 4. 2 36 45 17
Marmoset 7.7 2. 3 2.6 13 18 70
Bushbaby 25 - 53, 16 11, 29 64, 45 27, 15
Non-Primates
Cat 1.8 - 59 40 99 0
Rabbit 7. 6 - 5. 5 - 5. 5 87
Rat 26 - 23 <1 24 51
Fruit Bat 6.5 - - - 0 94

102
although a conjugate containing taurine, quinaldylglycyltaurine has been reported
as a metabolite of quinoline-2-carboxylic acid (quinaldic acid) and of 4-hydroxy-
quinoline-2-carboxylic acid (kynurenic acid) in the cat (Kaihara & Price 1961).
The cat formed glycine and taurine conjugates of 1-naphthylacetic acid but did
not form a glucuronic acid conjugate which may indicate a possible defect in
glucuronide formation with this compound, a characteristic which is highly
substrate dependent in the cat (Capel et al. , 1972 ; Millburn, 1974). Some
Carnivores such as the lion, civet and genet form high amounts of glycine and/or
taurine conjugates of 1-naphthylacetic acid but are defective in the glucuronide
formation (French etall, 1974) whereas others such as dog, ferret and hyaena
form glycine, taurine and glucuronic acid conjugates of the acid (Caldwell et al. ,
1975b; Idle et al, 1975). Table 3:4 shows conjugation of three compounds in
some carnivores and cat, and it shows the ability to form Nt-glucuronide of
sulphadimethoxine in the dog but not in the others. It also shows that the dog
and ferret form glucuronic acid conjugate with phenol and not in the rest but all
do form the sulphate conjugate. This finding suggests that the defect in the glucur-
onide formation do occur in some other carnivores other than the cat. The
fruit bat does not form a glycine conjugate with 1-naphthylacetic acid, and the
defect in glycine conjugation with benzoic acid has been reported by Bridges et al,
(1970) and confirmed by Bababunmi et al, (1973) but Ette et al, (1974) have
shown that glycine conjugation with phenylacetic acid does take place in this
species. They therefore suggest that glycine conjugation in the fruit bat is
highly substrate dependent.
The metabolic conjugation pattern discussed so far has been on one dose
level (100 mg/kg, except in the human subjects which received 5 mg). The
effect of dose on the pattern of conjugates in the urine and bile of bile duct-cannulated
rats receiving lrnaphthylacetic acid was shown in Table 3:2. It is clear from the

Table 3:4
Conjugation reactions in some Carnivores (Adapted from Caldwell et al, 1975 b)
Compound Conjugate found % of 24 h excretion in :
in urine DOG
50 18
56 7
25
0 19
FERRET
58 40
6 26 63
27 0
HYAENA
90 tr
46 40 11
0 4
CAT
90 tr
59 0
37
18 0
CIVET
97 0
74 tr 6
66 0
GENET
99 0
70 tr 18
50 0
Phenol
1 -Naphthylacetic acid
Sulphadimethoxine
Sulphate Glucuronide
Glycine Glucuronic acid Taurine
N4-Acetyl
1 N -Glucuronide
1

104
figures for urine that glucuronic acid conjugation is taking over from glycine
conjugation as the dose rises, but for the bile the pattern is practically
unchanged for all the doses. The bile pattern is explained by the fact that
1-naphthylacetylglucuronide (M. W. 362) is extensively excreted in the bile in
the rat whereas 1-naphthylacetylglycine (M. W. 231) is not. At saturation doses
the glucuronidation capacity in 3 h as derived from the data on Table 3:2 is
70nmol/kgThe molecular weight of the glucuronide is within the range 325+ 50
given by Hirom et al (1972) for extensive biliary excretion in the rat whereas
that of the glycine conjugate is below this range and this conjugate is unlikely
to be excreted in the bile in large quantities. This result is consistent
with examples in the literature which show that the size of the dose of a compound
could alter the pattern of its metabolism in so far as a large dose could exhaust the
mechanism by which a small dose is metabolised (Williams, 1959). The
compound could then be partly excreted unchanged or undergo another metabolic
reaction.
- For example it has been known for a considerable time that in man and
the pig, small doses of benzoic acid are metabolised entirely by conjugation
with glycine, but with large doses glucuronic acid conjugation become predominant
(see Williams, 1959). The cat, as already mentioned has a defective glucuronic
acid conjugation for certain compounds including benzoic acid. Consequently
benzoic acid is more toxic to the cat than most common species of animals
(Bedford & Clarke, 1971, 1972), for the cat conjugates benzoic acid entirely with
glycine which is limited in supply and glucuronic acid conjugation is not available
to take over when large doses of benzoic acid are administered. Another example
is 4-acetamidophenol which produces liver necrosis in high doses. This drug
forms a sulphate, a glucuronide and a mercapturic acid. The toxic effect of high

105
doses appears to be related to the depletion of the liver of glutathione which is
used to form the mercapturic acid. Studies in the hamster, a species most
sensitive to the necrotic effects of 4-acetamidophenol, have shown that at low
doses more of the drug is excreted as sulphur conjugates - sulphate and
mercapturic acid - than glucuronide, but at high doses the reverse is true
(Jollow et al. , 1973).
It is clear from the study of the effect of dose on glycine, sulphate
and glutathione conjugations that these mechanisms have a limited capacity
unless supplemented from outside sources and that when they are exhausted
the glucuronic acid mechanism takes over. It follows from this that with any
compound which is conjugated with glucuronic acid only the pattern of metabolites
will not change very much with dose unless this is excessive.
In general, it seems probable that the extent of metabolic pathways will
change with increase in dose, since it is to be expected that different mechanisms
have different capacities, and furthermore, some compounds can induce their
own metabolism and possibly one pathway more than another. These changes,
however, may also vary with species as in the case of the cat with its defective
glucuronic acid mechanism.

106
CHAPTER FOUR
Metabolism of Diphenylacetic Acid
Contents Page s
Chromatographic identification of urinary metabolites 107
Chromatographic identification of urinary and binary metabolites in the rat. 108
Results
108
Influence of dose on the pattern of metabolism and excretion of diphenylacetic acid in the rat
113
Discussion 113

107
The Metabolism of Diphenylacetic Acid
The metabolic fate of diphenylacetic acid has been examined in man,
6 sub-human primates and 4 non-primate species. Additionally the influence
of dose level in its pattern of metabolism and excretion in the rat has also
been studied.
Chromatographic identification of urinary metabolites
14 _ Urine samples from the 11 species dosed with [ Cidiphenylacetic
acid were subjected to thin-layer chromatography (see Chapter 2, Table 2:2).
Thin-layer chromatography in solvent F of urine samples from man,
rhesus,cynomolgus, squirrel and capuchin monkeys, marmoset, bushbaby, cat,
14 _ rabbit, rat and fruit bat dosed with [ C]diphenylacetic acid revealed on radio-
chromatogram scanning the, presence of two 14
C peaks at R f 0.17 and 0.90
respectively (see Figs. 4:1, 4:2 and 4:3). The peak at R1 0.17 gave a positive
naphthoresorcinol reaction and disappeared from urine samples treated with p
glucuronidase or N-NaOH and this was accompanied by an enlarged peak at Rf
0.92 which corresponded with diphenylacetic acid. A portion of the glucuronide
gum obtained from rabbits fed with diphenylacetic (shown to contain diphenyl -
acetylglucuronide following its conversion to its methyl ester) also showed on
chromatography as above a naphthoresorcinol positive spot at Rf 0.17 which on
treatment with fl-glucuronidase or N NaOH afforded diphenylacetic acid ( see
chapter 2). The radioactive component found in the urine of the species dosed
with [14C]diphenylacetic acid and showing an Rf 0.17 was therefore identified as
the ester glucuronide conjugate of the acid. Similarly, thin-layer chromatography
of urine samples in solvent D followed by radiochromatogram scanning showed
two 14C peaks at Rf 0.10 and 0.88 which corresponded to diphenylacetylglucuronide
and diphenylacetic acid respectively.

108
There were marked species differences in the relative sizes of the
two 14C peaks found on radiochromatogram scanning. Thus, thin-layer
chromatogram of urine samples obtained from the fruit bat, rabbit and rat
and developed in solvent F showed a large peak at Rf
0.17 corresponding to
diphenylacetylglucuronide and only a minor peak at Rf 0. 90 due to the free
acid. In the case of the cynomolgus and squirrel monkeys the size of the two
peaks were about the same whereas for the capuchin and rhesus monkeys the
cat and man the peak at Rf 0.17 was about twice the size of that at Rf 0.90.
In the case of the bushbaby, however, there occurred a major peak at Rf 0. 90
with minor one at Rf 0.17.
Chromatographic identification of urinary and biliary metabolites in the rat
3 h urine and bile samples from bile-duct cannulated rats dose intra-
peritoneally with [14C]diphenylacetic acid (5-500 mg/kg) were subjected to
thin-layer chromatography and developed in solvent F followed by radio-
chromatogram scanning. This revealed a major 14C peak at R
f 0.16, and
a very small peak at Rf 0. 92. The peak at 0.16 gave a positive naphthoresorcinol
reaction and disappeared from urine and bile samples treated with fl-glueuronidase
or g-Na0}1 and this was accompanied by an enlarged peak at Rf 0. 92 which
corresponded with diphenylacetic acid. The peak at Rf 0.16 was therefore
identified as diphenylacetylglucuronide.
Similarly chromatograms developed in solvent D revealed the presence
of diphenylacetylglucuronide and diphenylacetic acid.
Results
The quantitative and qualitative aspects of the excretion of diphenylacetic
acid and its metabolites by the 11 species studied are shown in Table 4:1 and those
in rats at various dose levels are shown in Table 4:2.
Table 4:1 shows a low urinary excretion of the radioactivity of the administered

0 S. F.
Man (5 orally)
S. F.
t 1
• t Rhesus Monkey 414 rng/ikgi ! (intramilsc-utarly
• • 2 ; ; 2
•
109
S. F.
Fig. 4:1 Radiochromatogram scans of urine after a dose of [
14C]
diphenylacetic acid. 1. Diphenylacetylglucuronide.
2. Diphenylacetic acid
Solvent system F. 0 = origin. S. F. = solvent front

0 S. F.
• Squirrel monkey
110
Fig 4:2 Radiochrornatogram scans of urine after a dose of [1 C] diphenylacetic acid. 1. Diphenylacetylglucuronide
2. Diphenylacetic acid
Solvent system F. 0 = origin. S. F. = solvent front
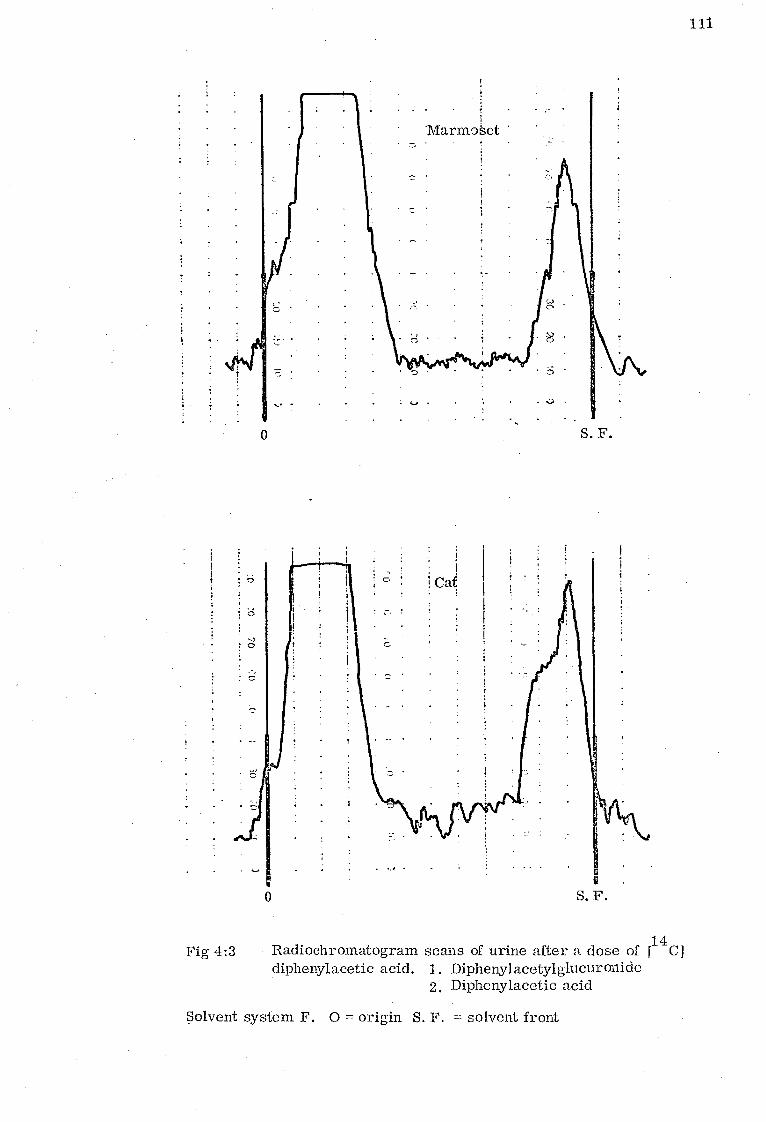
111
0
S. F.
Solvent system F. 0 = origin S. F. solvent front
Fig 4:3
Radiochromatogram scans of urine after a dose of [14
C] diphenylacetic acid. 3 . Diphenylacetylglucuronide
2. Diphenylacetic acid

0 S. F.
Fruit bat
(intraperitoneally)
S. F.
Rat . (intraperitoneally)
112
0 S. F.
Rabbit (Orally)
2. Diphenylacetic acid Solvent system F. 0 = origin S. F. = solvent front
Fig 4:4 Radiochromatogram scans of urine after a dose of [
14c]
diphenylacetic acid 1. Diphenylacetylglucuronicle

113
dose of [14C]diphenylacetic acid in 48 h in squirrel monkey (14% of the dose),
capuchin (15 and 21%), bushbaby (23 and 34%) fruit bat (29 and 35%), cat (40%),
rhesus monkey (45 and 49%) and rat (48%) but relatively higher values for
cynomolgus (57%) and marmoset (66%) monkeys. The rabbit excretes about 84%
whereas all the radioactivity is excreted by man. The major excretion products
are diphenylacetylglucuronide and diphenylacetic acid but the quantities varied
with species. Diphenylacetylglucuronide is the major excretion product in
most of the species except in the cynomolgus monkey and bushbaby which have
the unchanged acid as the major excretion product. There are traces of the glycine
conjugate in the marmoset, bushbaby, cat and rat, and also of the taurine conjugate
in the cat as shown by reverse isotope dilution.
Influence of dose on the pattern of metabolism and excretion of diphenyl-
acetic acid in the rat.
At 5 mg/kg dose of [14C]diphenylacetic acid (intraperitoneally) about 86%
of the radioactivity was excreted in the bile in 3 h while about 11% was in the
urine. The percentage excreted decreased with increasing dose and at 500 mg/kg
dose level, 20% is excreted in the bile , with some 8 % in the urine. The major
excretion product was diphenylacetylglucuronide with only small amounts of the un-
changed compound irrespective of the dose level.
Discussion
The metabolic fate of diphenylacetic acid has been studied in eleven species
and the results show that diphenylacetic acid is only conjugated with glucuronic
acid irrespective of species but the quantities varied with species. Table 4:3
summarises the conjugation pattern of diphenylacetic acid in the species studied
and it suggests among other things that bushbaby has a low capacity in forming a
glucuronic acid conjugate with diphenylacetic acid and bearing in mind the low
glucuronic acid conjugation with 1-naphthylacetic acid (see Chapter 3, Table 3:1)

Amount of [14C]-diphenylacetic acid excreted conjugated with various compounds in 48 h as % dose
Unconjugated Glutamine Glycine Taurine Glucuronic Acid
30, 34
12, 17
29
n.d.
n. d.
n. d.
n. d.
n. d.
n. d.
5. 7 n. d. n. d.
2.6, 6.7 n.d. n. d.
17 n. d. trace
29, 17 n. d. trace
9.6 (7.4-13) n.d. trace
6. 9 (5. 6-9.2) n. d. n. d.
2.6 (2: 2-2.9) n. d. trace
2.8, 1.5 n. d. n. d.
n. d. 70, 65
n. d. 30, 36 n. d. 27 n. d. 8.4
n. d. 18, 10
n. d. 49
n. d. 3.2, 8.7
trace 30 (26-32)
n. d. 77 (75-79)
n. d. 45 (45-45)
n. d. 26, 34
Table 4:1
Conjugates of [14C]-diphenylacetic acid in various species
14 [ C]-diphenylacetic acid (114 mg/kg) dissolved in the appropriate amount of N-NaOH was administered as described in the text. The urine was collected for 48 h and analysed for metabolites. Results are means for three animals with ranges in parentheses. animals were used individual values are given.
Species (No. & Sex) Genus and Species Dose of 14C Route of administration
Where only one or two
14C excreted %
Primates pCi/animal of dose
Man * (2M) Homo sapiens P. 0. 5. 0 100, 100 Rhesus monkey (1M,1F) Macaca mulatta i. m. 13. 0 45, 49 Cynomolgus monkey (1F) Macaca fascicularis (Iris) i. m. 11.4 57 Squirrel monkey (1F) Saimiri sciureus i.m. 14. 0 14 Capuchin (2F) Cebus albifrons i.m. 11.4 15, 21 Marmoset (1M) Sanguinus oedipus i. m. 14,0 66 Bushbaby(1M, 1F) Galago crassicaudatus i. m. 6. 0 23, 34
Non-Primates
Cat (2F, 1M) Felis cattus i. m. 11.4 40 (36-42) Rabbit (3F) Lepus canaliculus P.O. 21. 0 84 (88-91) Rat (3F) Rattus norvegicus i.p. 1.4 48 (42-53) Fruit bat (1F,1M) Pteropus giganteus 1. p. 11. 3 29, 35
* Dose 5 mg as shown by reverse isotope dilution n. d. not detected

Table 4:2
Influence of dose on the pattern of metabolism and excretion of diphenylacetic acid in the rat
Bile-duct cannulated female rats were injected intraperitoneally with diphenylacetic acid (dissolved in the calculated amount of N-NaOH and diluted with water) and the bile collected for 3h and urine removed from the bladder. Bile and urine were analysed for 14C and metabolites as described in the text.
Bile Urine I I
14C excreted conjugated with: % 14 % C excreted conjugated with:
Dose (mg/kg) % dose excreted Unchanged Glucuronic % dose excreted Unchanged Glucuronic Acid Acid
5 86 (74-95) 7.1 (6.2-8.4) 93 (92-94) 11 (4.4-19) 13 (2.1-34) 87 (66-98)
25 73 (72-74) 16.0 (12-19) 84 (88-89) 12 (7.3-17) 2.3 (2.1-3.3) 98 (97-98)
50 30 (27-36) 9.0 (8. 9-9. 2) 91 (91-92) 9.3 (4. 5-15) 2.9 (1.1-4.2) 98 (96-99)
250 39 (16-53) 7.2 (6. 8-10) 93 (91-93) 19 (13-22) 1.9 (1.7-3.0) 98 (97-98)
500 20 (15-44) 9.1 (8.3-10) 91 (90-92) 8.2 (7.3-9.4) 4.9 (4.0-6.2) 95 (94-96)

Table 4:3
Species Variation in the pattern of conjugation of Diphenylacetic acid
Diphenylacetic abid conjugates as % of the 14C excreted in the
urine
Species
Primates
Unchanged Glycine Glucuronic acid
Man 32 - 68
Rhesus monkey 31 - 69
Cynomolgus monkey 52 - 49
Squirrel monkey 40 - 60
Capuchin 17, 31 - 83, 69
Marmoset 25 < 1 75
Bushb aby 87, 74 <1 14, 26
Non Primates
Cat * 24 <1 76
Rabbit 8.2 - 92
Rat 5.4 <1 95
Fruit Bat 9.7 - 93
* 1% conjugated with taurine

117
one wonders if this is not characteristic with this species. The ability of
the cat to form glucuronic acid conjugateAurther substantiates the fact that
the glucuronide formation in this species is highly substrate dependent. (see
Chapter 3, Capel et al, 1972 ; Millburn 1974). This report is in agreemnt with
the finding of Miriam et al (1927a) which has shown that dog, man and rabbit
conjugate this acid with glucuronic acid.
The conjugation of diphenylacetic acid at different dose levels in the bile
duct-cannulated rats was also investigated (see Table 4:2). The figures show
that higher amounts were excreted in the bile than in the urine irrespective of
the dose and that diphenylacetylglucuronide was the only conjugated excretion
product in both urine and bile. This is understandable since this is the only
metabolic conjugation route employed by the rat, unlike in the case of
1-naphthylacetic acid (see Chapter 3) where there was a shift from glycine
conjugation at low doses to glucuronide conjugation at higher doses. At saturation
doses the glucuronidation capacity in 3h as derived from the data on Table 4:2 is
140nmol/kg.Finally the molecular weight of the diphenylacetyl glucuronide is 388
and this value is a prequisite for extensive biliary excretion in the rat. A
structurally related compotmd,bis (p-chlorophenyl)acetic acid has been reported
to undergo extensive enterohepatic circulation when administered intravenously
to rat (Gingell, 1975). Comparing the amount of diphenylacetic acid excreted in the
intact rat in 48 h and the high biliary excretion in the bile duct-cannulated rat in 3h.
It is suggested that the metabolite excreted in the bile is reabsorbed and excreted
in the urine slowly. The only metabolite is diphenylacetylglucuronide which
could be reabsorbed as such or after hydrolysis to the free acid by the gut flora and/
or secretions (see Smith and Williams, 1966). p-Glucuronidase activity is widely
distributed among the intestinal bacteria (Hawksworth et al. , 1971).

118
CHAPTER FIVE
Metabolism of Hydratropic Acid
Contents Pages
Chromatographic Identification of Urinary Metabolites 119
Man 119 Rabbit, Rat and Rhesus Monkey 119 Cat 122
Chromatographic Identification of Urinary and Biliary Excretion 122 in the Rat
Results 123
Man 123 Rhesus Monkey, Rabbit and Rat 123 Cat 123
Influence of Dose on the Pattern of Metabolism and Excretion of Hydratropic Acid in the Rat 126
Discussion 126

119
The Metabolism of Ilydratropic Acid
The metabolic fate of hydratropic acid has been studied in man, rhesus
monkey, cat, rabbit and rat. Additionally the influence of dose level on its
pattern of metabolism and excretion in the rat has been examined.
Chromatographic identification of urinary metabolites
Urine samples from the 5 species dosed with [14C]hydratropic acid were
subjected to thin-layer chromatography using the systems described in Chapter 2.
(see Table 2 : 3).
Man Chromatography of urine samples on thin-layers using solvent F showed on radio-
chromatogram scanning the presence of one major 14C peak of R
f 0.13 (Fig. 5:1).
This peak gave a positive reaction with naphthoresorcinol and disappeared when
urine samples were incubated with fl-glucuronidase or warmed with N-Na0H. Its
disappearance was accompanied by an appearance of a new peak at Rf 0. 88.
corresponding to hydratropic acid. The peak at Rf 0.13 therefore was identified
as hydratropoylglucuronide. Chromatograms developed in solvent D showed a
single large peak at the origin which corresponded with hydrotropoylglucuronide.
As described above this large peak at the origin disappeared when the urine was
treated with j3-glucuronidase or N-NaOH to be replaced by a new peak at R1 0. 76
corresponding to hydratropic acid.
Rabbit, rat and rhesus monkey . Thin-layer chromatography of urine samples
using solvent F, followed by radiochromatogram scanning showed the presence
of a large 14C peak at R
f 0.14 corresponding to hydratropoylglucuronide and a
small 14C peak at Rf 0.86 due to the unchanged compound (Fig. 5:2). The peak
at Rf 0.14 gave a positive reaction with naphthoresorcinol and disappeared when
urine samples were treated with f3 - g lu c u rani dase or N-NaOH and this was
accompanied by a large increase in the size of the peak at Rf 0. 86 due to hydra--

120
0
S. F.
S. F.
Fig 5:1 Hydratropic acid conjugates in urine chromatograms were developed in solvent F. 1. Hydratropoylgrucuronide. 9, . Hydratropoyltaurine 3. Hydratropoylglycine, 4. Hydratropic acid.

121
Rhesus monkey
ri
0
0
0
S. F.
Fig 5:2 Hydratropic acid conjugates in urine Chromatograms were developed in solvent F. 1. Hydratropoylglucuronide 2. Hydratropic acid

122
tropic acid. Chromatography on thin-layers in solvent D showed the presence
of hydratropoylglucuronide together with a small amount of the unchanged acid.
The presence of hydratropic acid before and after P-glucuronidase or N-NaOH
treatment was confirmed by reverse isotope dilution.
Cat 14C-scans of thin-layer chromatogram of cat urine developed in solvent F
showed four radioactive peaks of Rf values 0.12, 0. 33, 0.72 and 0. 87
respectively (Fig. 5:1). The peak at Rf 0.12 gave a positive reaction with
naphthoresorcinol and disappeared when urine samples were treated with
p-glucuronidase or N-NaOH and was accompanied by a large increase in the
size of the peak at Rf 0. 87 corresponding to hydratropic acid. The peaks at R1
0.33 and 0.72 gave a purple brown colour reaction with chlorine-potassium
iodide/starch reagent, and the peak at Rf 0. 72 gave also positive reaction with
the 4-dimethylaminobenzaldehyde reagent. Therefore, these peaks at Rf
0.12, 0.33, 0.72 and 0.87 were identified as hydratropoylglucuronide, -taurine
and -glycine and hydratropic acid respectively. Similarly chromatograms
developed in solvent D revealed four 14C peaks of Rf values 0.1, 0. 30, 0.63
and 0. 75 corresponding to hydratropoyl- glucuronide, -taurine and -glycine
and hydratropic acid respectively. The presence of hydratropoyl- taurine and
-glycine and hydratropic acid was also confirmed by reverse isotope dilution.
Chromatographic identification of urinary and biliary excretion in the rat
Urine and bile samples were collected over the first 3 h from bile-duct
cannulated rats dosed intraperitoneally with [14 C]+hydratropi c acid (5-500 mg/kg)
were streaked on thin-layers and developed in solvent F. Radiochromatogram
scanning revealed a major 14C peak at Rf 0.13 and a minor peak at Rf 0. 87. The

123
peak at Rf 0.13 gave a positive reaction with naphthoresorcinol and disappeared
when urine and bile samples were treated with p-glucuronidase or N-NaOH and
this was accompanied by a large increase in the size of the peak at R 0.87
due to hydratropic acid. The peak at R 0.13 therefore was identified as hydra-
tropoylglucuronide. Similarly chromatograms developed in solvent D reveals
the presence of hydratropoylglucuronide and hydratropic acid.
Results
The quantitative and qualitative data on the excretion of hydratropic acid
and its metabolites in the five species studied are shown in Table 5:1 and those
in rats at various dose levels are shown in Table 5:2.
Man. The two human subjects excreted 97 and 100% of the radioactivity in the
urine in 24 h after an oral dose of 5 mg [ CJ±hydratropic acid. The major
excretion product was hydratropoylglucuronide (95 and 100% of the dose respect-
ively) with 2.1% of the unchanged hydratropic acid in one of the subjects.
Rhesus monkey, rabbit and rat
The urinary excretion of the radioactivity of the administered dose of [14
C}-1-
hydratropic acid (81 mg/kg) in 24 h is about 97% in the rhesus monkey, 82% in the
rabbit and 81% in the rat. The major excretion product was hydratropoylglucuronide,
the quantities varied in the species, the rhesus monkey excreted 82 and 71.1% ( of
the dose in the two examined), rabbit 73.1% and rat 64.1%. There were also
small amounts of the unchanged compound.
Cat. Cats injected intraperitoneally with [14C1-thydratropic acid (81 mg/kg)
ex_creted about 58% in the urine within 48 h. The major excretion products were
hydratropoylglucuronide (24.1% of the dose), hydratropoyltaurine (13.1%),
hydratropoylglycine (8. 7%) and some unchanged hydratropic acid (12. 1%).

Table 5:1
Conjugates of [14C]-hydratropic acid in various species
[14C]-hydratropic acid (81 mg/kg) in solution of propylene glycol/water (2:1 by vol.) was administered as described in the text. The urine was collected for 24 h and analysed for metabolites. Results are means for three animals with ranges in parentheses. Where only one or two animals were used individual values are given.
Species (No. & Sex) Dose of 14C 14C excreted %
Genus and Species Route of administration tCi/animal of dose
Amount of [14C]-hydratropic acid excreted conjugated with various compounds in 24 h as % dose
Unconjugated Glycine Taurine Glucuronic Acid
Primates *
Man (2M) Homo sapiens p. o. 2. 5 97, 100 2.1, 0 95.0, 100
9.6
8. 5
9. 6
5. 2
Rhesus monkey (1M ,1F) Macaca mulatta i. m.
Non-Primates
Cat (2F, 1M) Fells cattus i. p.
Rabbit (3F) Lepus caniculus p. o.
Rat (3F) Rattus norvegicus i.p.
97, 97.5
58 (53-64)
82 (72-90)
81 (63-90)
15.0, 26.9 82.0, 70.6,
11.9 (8.3-13.9) 8.7(7.9-9.9) 13.3 (8. 1-17. 6) 24.1 (22. 3-26. 1)
9.3 (0. 0-16.6) 72.7 (65.4-82. 0)
16.6 (15.6-17.9) <1 63.9 (63.1-65.4)
* dose was 5 mg/man - = not detected + 48 h excretion

Table 5:2
Influence of dose on the pattern of metabolism and excretion of hydratropic acid in the rat
Bile-duct cannulated rats were injected intraperitoneally with [14C1-hydratropic acid (in solution of propyleneglycol/water, 2:1 v/v) and the bile collected for 3 h and urine removed from the bladder. Bile and urine were analysed for 14C and metabolites as described in the text
Urine Bile i
% 14C excreted conjugated with:
Dose (mg/kg) % dose excreted Unchanged Glucuronic % dose excreted Unchanged Glucuronic Acid Acid
5 26 (16-39) 7.2 (6.3-8.2) 93 (92-94) 32 (21-46) 28 (18-38) 73 (62-81)
25 23 (20-25) 8.6 (5. 8-19) 90.1 (81-94) 28 (20-37) 25 (22-28) 74.2 (72-78)
50 21 (19-23) 14 (3.2-26) 85.7 (74-97) 23 (14-32) 24 (17-25) 76.1 (75-78)
250 22 (7.3-31) 4.2 (2.1-7.0) 95.6 (94-98) 29 (5.5-36) 23 (15-32) 77.8 (68-83)
500 2.6 (2.2-3.0) 15.3 (8.6-25) 84.4 (73-91) 23 (7.9-41) 19 (8.9-25) 80.9 (75-91)
14 % excreted conjugated with:
*.hydratropoylglycine not detected in urine or bile

126
Influence of dose on the pattern of metabolism and excretion of hydratropic
acid in the rat
Bile-duct cannulated rats injected intraperitoneally with [ C]iydra-
tropic acid (5-500 mg/kg) excreted at 5 mg/kg dose level about 32% of the
radioactivity in the bile and 28% in the urine. These values fell with increasing
dose, and at the dose level of 500 mg/kg while 23% of the dose was excreted
in the bile only 2.6% was excreted in the urine. The major excretion product
was hydratropoylglucuronide in both urine and bile irrespective of the dose
level, and small amounts of the free acids were also excreted.
Discussion
The metabolic fate of hydratropic acid has been studied in the five species.
Table 5:2 shows the conjugation pattern of this acid in the species studied and it
suggests that hydratropic acid can be conjugated with amino acids and / or
glucuronic acids the pattern depending on the species. Man, rhesus monkey,
rabbit and rat form glucuronic acid conjugate of hydratropic acid, and earlier
Robinson et al (1955) have found this conjugate in the rabbit. The cat on the
other hand forms glucuronic acid, glycine and taurine conjugates of this acid and
Kay and Raper, (1922) have reported that another carnivore, the dog, conjugates
this acid with glycine. The ferret also conjugates the compound mainly with glycine
and taurine (unpublished data of J. R. Idle). The ability to form glucuronides in the
cat is substrate dependent (Millburn, 1974) and hydratropic acids seems a very
important substrate in the shift from amino acid to glucuronic acid conjugation
since this species forms only amino acid conjugates with phenylacetic (James et al
1972 a) and 1-naphthylacetic (see Chapter 3) acids but glucuronic acid conjugate
with diphenylacetic acid (see Chapter 4. )
The effect of dose on the metabolism and excretion of hydratropic acid in
bile duct cannulated rats has also been examined and the results shown in Table
5:2, suggests a fairly even excretion in the bile and urine except at the dose level

Table 5:3
Species Variation in the pattern of Conjugation of Hydratropic acid
Hydratropic acid conjugates as % of the 14C excreted in the urine
Species Unchanged Glycine Taurine Total conjugated with Glucuronic acid
Primates
amino acids
Man 1 - - 0 99
Rhesus monkey 16, 28 - - 0 85, 72
Non-Primates
Cat 21 15 23 38 42
Rabbit 11 - - 0 89
Rat 21 L. 1 - 1 79

128
of 500 mg/kg where the biliary excretion was higher. Hydratropoylglucuronide
was the only conjugated excretion product in bile and urine. This is to be
expected since this is the only metabolic conjugation route for hydratropic acid
in the rat. At saturation doses the glucuronidation capacity in 3 h as derived from
the data on Table 5:2 is 145nmol/kg. Hydratropoylglucuronide (M. W. 326) has a
molecular weight with the range 325 ± 50 given by Hirom et al. (1972) for
extensive biliary excretion in the rat. The excretion in intact rats and the bile
duct-cannulated rats suggest a reabsorption of the glucuronic acid conjugate as
stated in Chapter 4.

129
CHAPTER SIX
Pharmacokinetic and Subcellular Aspects of Arylacetic
Acid Conjugation
Contents Pages
Introduction 130
Drug Distribution 130
Membranes 131
Results
Discussion
Mitochondria 131 Microsomes 132
Drug Binding 134
Lipid Solubility and Metabolism 135
Affinities of Conjugation Sites for Drugs and 136 their Polar Metabolites
Pharmac °kinetics 137
Enzyme Affinity 140
Scope of the Present Investigation 141
141
PharmacokMetics 141
1-Naphthylacetic Acid 141 Diphenylacetic Acid 141 Hydratropic Acid 149 Phenylacetic Acid 149
Binding Studies 149
Enzyme Affinity 152
Glycine C onjugation 152
Glucuronic Acid Conjugation 152
152
Summary and Conclusion 162

130
Pharmacokinetics and subcellular Aspects of Arylacetic Acid Conjugation
Introduction The results of the in vivo studies (see Chapter 3, 4 and 5) and the work
of others reported elsewhere in the literature strongly suggest that the structure
of an arylacetic acid is the most important factor influencing its pattern of metabolic
conjugation. The pharmacokinetic behaviour of a compound may give information
as to its manner of distribution in the body , binding to body components and
more especially its access to the metabolic conjugation sites, in this case the
mitochondria (site of amino acid conjugation) Schachter and Taggart 1954) and the
endoplasmic reticulum (microsomes, site of glucuronic acid conjugation, Dutton,
1966). The overall metabolism would be a result of the affinity of the arylacetic
acid for the conjugating enzymes associated with these structures. The differential
sites for conjugation may be related to the ability of the mitochondria to absorb
ionic forms, whereas metabolism by microsomes requires in the main lipid
soluble nonpolar molecules.
Drug distribution
After a drug is absorbed or injected into the blood stream, it can enter or
pass through the various body fluid compartments — plasma, interstitial fluid,
transcellular fluids anc cellular fluids. Some drugs cannot pass cell membranes
and therefore, are restricted in their distribution and in their sites of action,
whereas others penetrate into cells and thereby distribute throughout all fluid
compartments. The important factors that influence distribution include the extent
of binding of drug to plasma and tissue proteins and other components of cells
such as mitochondria and endoplasmic reticulum (Burns et al. , 1953 ; Dingell
et al. 1964) ; regional differences in pH ; the permeability characteristics of — —
special membranes to drug ; and the lipid content of the cells (Melmon and Morrelli,
1972). Tissue uptake of a drug is dependent upon the mass of the tissue and the

131
rate of blood flow to it, as well as the partition coefficient of the drug between
blood and tissue (Gillette, 1973).
Membranes
For a drug introduced into the body to getto its metabolic site it has to
penetrate some membranes. The degree of penetration depends on the
constitution of the membrane, type of transport mechanism employed and the
partition coefficient of the drug between the membrane and its environment.
It is a widely accepted view that biological membranes possess both
physical and functional plasticity and this had led to the hypothesis of a membrane
structure in which mobile arrays of proteins and lipids are bound together by
non-covalent hydrophobic bonds (Singer, 1971 ; Gilter, 1972). In this context,
it has been shown by paramagnetic resonance and x-ray diffraction that fatty
acid chains in a lipid bilayer and in some biological membranes exhibit a high
degree of molecular motion (Luzzati, 1968 ; Hubbell and McConnell, 1971).
Furthermore, Branton et al. (1972) have shown with freeze-etched preparations
that the organisational changes associated with the fluidity of the membrane are
caused both by intrinsic modifications of the membrane and by changes in its
surrounding environment. More recent work on freeze-etched tetrahymena
membranes (Hubbell and Wunderlich, 1973) has further suggested that membrane
intercalated particles may be capable of transme,mbranal mobility.
Mitochondria
The penetration of drugs into subcellular particles such as mitochondria
appears to be an important aspect of drug distribution and action ; and the
penetration of drugs follows the same principles as for cell membranes. In
contrast to indications of membrane plasticity, the results of biochemical
studies on mitochondria have given a rather static picture of enzyme localization

132
in mitochondrial membranes (Waksman and Rendon, 1974). It has been shown
that some macromolecular components (enzymes and proteins) of mitochondrial
inner membrane are capable of large amplitude movements allowing them specific
and reversible escape from the field of membrane interactions (Waksman and
Rendon, 1971 ; Rendon and Waksman, 1971).
Waksman and Rendon (1974) have also reported that an integrated mito -
chondrial membrane system is endowed with the potential capacity to "read and
recognize" defined variations in the extramitochondrial environment and to
transduce this information by specific and reversible, large amplitude inter-
membranal protein movements. Since the behaviour of a protein is defined by
a set of interactions with its environment, the mobile protein in its membrane-
bound state should be related to its membrane partners by a characteristic set
of interactions. Once severed from these connections, it will become available
for new interactions in a new environment. At least two extreme possibilities ,
both of which depend on specific ligand-protein interactions are offered to the
released proteins. A first possibility is the environmentally induced metabolic
expression of the released enzymatic activities (Estrada-0 et al 1970) Thus some
mitochondrial enzymes may be modulated, by their shuttling to and away from
their substrates and ligands. A second possibility is that the protein shuttling
may be associated with the transport of small molecules, with some advantage
to cell economy. If these proteins are able to bind ligands, without necessarily
metabolizing them, the bound ligand might then be transported from one compart-
ment to another (Waksman and Rendon, 1974).
Microsomes
Many drugs are bound to microsomes, for example imipramine (Gillette,
1973) and halothane (Uehleke et al, 1973 ; Van Dyke and Wood, 1975). The
semipermeability of microsomal membrane/has certain important functional

133
implications, mainly since the accessibility of a number of substrates to the
membrane-bound enzymes is shown to be a limiting factor (Hers and deDuve, 1950 ;
Ernster and Jones, 1962 ; Mulder, 1970) . Changes in membrane permeability
by external or internal agents may be decisive for regulating the activity of
various enzymic systems (Blether, 1966 ; Rodbell, 1966).
Since the enzyme activity depends on the integrity of the membrane
structure, it follows that :
1) a membrane bound enzyme may as a result of interaction with
other membrane components have a different conformation ( and hence
different catalytic activity) from that which, it would possess in free
solution (Laidler and Bunting, 1973).
ii) depending on the location of the enzyme in the membrane its inter-
action with substrates may occur in an environment which is different
(e. g. in pH) from that outside the microsomal vesicles (Katchalski et al .
1972)
iii) if the enzyme is located in the microsomal membrane so that
a permeability barrier is interposed between substrates and active site,
partition of substrates between the aqueous and membrane phases,
permeability of the microsomal membrane and the nature of substrate
transport mechanisms may affect the overall rate of the enzyme reaction.
The size and the surface charge of the microsomal vesicles may be important.
Lysophosphatides, unsaturated fatty acids, phospholipase A and detergents
increase the permeability of microsomal membranes (Fiehn and Hasselbach,
1970 ; Kreibich et al. , 1973) and as a result might effect the apparent
enzyme activity e. g. rat liver UDP-glucuronyltransferase.
Nilsson et al (1971) studied the permeability of microsome,s

134
to different compounds and found that unchanged molecules freely
penetrated the microsomal membrane while smaller charged species did
not. They also demonstrated a high partition by a lipophilic substrate
like naphthalene into the microsomal phase by comparing its concentration
in the pellet and supernatant fraction after separation of the two phases.
Furthermore, Stier and Sackmann (1973) examined the interaction of
microsomal lipid and substrates with spin-label techniques. They have
concluded that the microsomal system consists of a heterogeneous lipid
matrix in which the enzymes are imbedded. This lipid matrix influences
the binding of substrate as well as the interaction of the enzymes since
the substrate is incorporated irto the membrane.
Drug binding
A drug can bind with tissue and subcellular components in a variety of
ways. Goldstein et al. (1971) have distinguished between non-saturable and
saturable interactions.
Non-saturable interactions are of two kinds. First, particles surrounded
by an osmotic membrane (Qg. synaptosomes) or having a spongy matrix can
contain entrapped drug in aqueous solution. Second, membranes will contain
dissolved drug, in amounts determined by the lipid/water partition coefficient
and the ambient aqueous concentration.
Non-specific saturable interaction arises through ionic bonds, hydrogen
bonds, and hydrophobic forces, but specific saturable interaction arises at the
receptor sites, where drug binding triggers the chain of events that leads to
the characteristic pharmacological effect. A measure of specific saturable
binding of an arylacetic acid to mitochondria and microsomes could be used as
an index of correlation as to the degree of amino acid (in the mitochondria) or
glucuronic acid (in the microsomes) conjugation. It has also been shown that

135
the parent drug and its nonpolar metabolite have more affinity for its metabolic
site than its polar metabolite (Bickel and Borner, 1974 ; Glauman et al. 1970 ;
Nilsson et al. 1971).
Lipid solubility and metabolism
The lipid solubility of a compound seems not only to determine its
penetration into cells and metabolic sites but also influences its metabolism.
The oxidative dealkylation of foreign N-alkylamines by rabbit liver microsomes
appears to be limited to compounds which are lipid soluble ; and since the micro-
somal hydroxylation of aromatic compounds also appears to be limited to lipid
soluble substances as well, Gaudette and Brodie (1959) suggested that an intra-
cellular fat-like boundary separates normally occuring polar substances from
the highly non-specific microsomal enzymes.
The microsomal system dealkylates a variety of highly lipid soluble N-
alkylamines,such as methylamphetamine, pethidine, ephedrine and codeine
(Gaudette and Brodie, 1959 ; Axelrod, 1956), but normally occurring N -alkyl-
amines such as sarcosine, dimethylglycine and epinephrine, which have very
low lipid solubility , are not demethylated by liver microsomes. The dimethyl-
ation of sarcosine and dimethylglycine, however, is accomplished by highly specific
systems in the mitochondria (Mackenzie and Frisell, 1958 ; Gaudette and Brodie,
1959) . A similar situation exists for the hydroxylation of aromatic compounds.
Despite the non-specificity of the microsomal hydroxylation system with respect
to foreign compounds, it does not promote the hydroxylation of naturally occuring
compounds such as L-phenylalanine, L--tryptophane, kynurenine and anthranilic acid,
which are hydroxylated by quite specific systems localised in other parts of the
liver cell (Mitoma et al. , 1956).
Other examples of the importance of lipid solubility are found in substrates

136
of the deamination enzyme present in rabbit microsomes, but absent in a number
of other mammalian species (Axelrod, 1954) e This enzyme deaminates the lipid
soluble substances amphetamine, methamphetamine and ephedrine, but does not act
on substances like tyramine or serotonin (Axelrod, 1954) , which are substrates
of monoamine oxidase.
The ability of certain drugs to increase the fluidity of lipid region in a
membrane may also affect the metabolism of that drug. Stier and Sackmann,
(1973) have suggested that the cytochrome P450 system, which is the drug
metabolising system situated in the microsomal fraction of the liver, may be
enclosed in a' gather rigid phospholipid halo'! A critical factor in the metabolism
of a substrate by the cytochrome P450 system would seem to be its lipid solubility.
However, the rate of metabolism of a drug could be determined not by its lipo-
philic nature alone but also by its ability to fluidise the phospholipid halo
surrounding the cytochrome P450 complex (see Cater et al. , 1974).
This lipophilic characteristic could be one of the vital factors among other things
operating in the metabolic conjugation of the arylacetic acids already examined, the
ones with high lipid solubility, or able to fluidise the lipid region of the micro-
somes would be, expected to be conjugated there.
Affinities of conjugation sites for drugs and their polar metabolites
Little is known on the uptake of drugs into cells, their binding to cellular
structures, as well as the nature of intracellular and intercompartmental trans-
location of drugs and their polar metabolites from binding and metabolic sites
to sites of incretion or excretion.
Bickel and Borner (1974) studied pharmacokinetic processes using
recirculating rat liver perfusion system, and determined the drug imipramine
and its major metabolites at various times in perfus ate, liver, bile and subcellular

137
liver fractions. They found that imipramine undergoes a rapid hepatic uptake,
the initial extraction being close to 100% ; and that most of the unchanged drug
is then localized in the microsomal fraction. Metabolism of imipramine is
not limited by uptake but by enzymatic factors. Like imipramine, its lipophilic
metabolite, desmethylimipramine is bound to microsomes, but the polar
glucuronides are easily released from endoplasmic reticulum, their site of
formation, into the cytosol and presumably from there excreted into the bile.
This would agree with the direct demonstration of this pathway for the polar
metabolites and conjugates of naphthalene (Glaumann et al. 1970) and morphine
(Nilsson et al. , 1971). The same general pattern of intra-hepatocytic binding,
and distribution has been observed by Bahr and Borga (1971) (desmethylimipramine --->
2-hydrov-desmethyl -imipramine ---)2-hydroxy-desmethylimipramine-
glucuronide) and by Levine and Singer (1972) with other compounds, where
glucuronides in contrast to the parent drugs, were mainly localized in the cytosol
fraction. Bickel and Steele (1974) also have demonstrated high affinities of liver
microsomes for imipramine and desmethylimipramine, but not their glucuronidated
derivatives. This finding could also apply to parent drugs and their polar
conjugates in the mitochondria.
Pharmacokinetics
A drug which when administered intravenously rapidly equilibrates between
the plasma and other body fluids and tissues, and when administered orally is not
subject to metabolism prior to reaching the systemic circulation, can usually be
adequately described by a one compartment model with first-order elimination
as depicted in scheme 1.
K input V
Scheme 1
output

138
In this scheme, K is the first-order rate constant for hepatic elimination and
V is the apparent volume of distribution of the drug. The apparent volume of
distribution is dependent on the size of drug molecule, lipid solubility and its
binding to plasma and tissue proteins ; and can be defined as that volume of
fluid into which the drug appears to distribute with concentration equal to that
in the blood.
Amount of drug in the body
Apparent Vol, of distribution - (1) Concentration of drug in the blood
The biologic half-life (T1/2) and clearance (Q) of a drug which confers
single compartment characteristics to the body are directly related to K as
follows:
T1/2 = 0.693 / K (2)
and
KV (3)
Since the liver is considered an integral part of this "homogeneous" one-
compartment system, the direct relationship between half-life and clearance, and
hepatic elimination as expressed in equations 2 and 3, would be anticipated.
Consequently, half-life and clearance should reflect the elimination characteristics
of this class of drugs (Perrier and Gibaldi, 1974).
A large number of drugs, however confer on the body the characteristics
of a multicompartment system rather than a simple one-compartment system.
For this class of drugs the body does not behave as a single kinetically "homogeneous"
compartment, but rather as two or more such compartments. The simplest
multicompartment system, a two-compartment system is illustrated in the scheme
2 where K12 and K21 are inte])..compartmental transfer rate constants and Ke is the

Central Compartment
V1
output \toe Ke
input
139
elimination rate constant.
K12
K21
Peripheral Compartment
V2
Scheme 2
Unless there is evidence to the contrary, the liver is usually assumed to be an
integral part of thehomogeneous central compartment. Hence, the rate
constant, Ke, is analogous to K in a one-compartment system in that it is also
the rate constant for hepatic elimination. The biologic half-life of a drug which confers
multicompartment characteristics on the body is not only a function of hepatic
elimination (Ke) but also of drug distribution (K12
and K21), and for a two
compartment model,
T1 = 0.693/ 2 [ (K12+ K21 Ke)- (K12 +K21 +Ke) 2 - 4 K2iKei (4)
-2-
The result is that the biologic half-life of this class of drugs is not
linearly related to the reciprocal of Ke, the hepatic elimination rate constant
(Gibaldi and Perrier, 1972). Clearance can be determined by employing the
relationship
Q = Dose AUC
where 'Dose' is the administered dose and AUC is the total area (from time zero to
infinity) under the plasma concentration-time curve. Clearance as calculated
previously for one-compartment model (equation 3) can also be determined employing
this relationship. For a drug which can be characterized by scheme 2
(5)

140
AUC = Dose = Dose Ke
pvd
. Q = KeV1 = ord (6)
where V1 is the apparent volume of the central compartment, Vd is the apparent
volume of distribution of the drug and 13 equals 0. 693/T1 where T1, the biologic
half -life is defined by equation 4. Equation 6 illustrates the direct relationship
between clearance and hepatic elimination. Therefore, clearance measurements
in multi-compartment systems should yield insight into the intrinsic elimination
characteristics of a drug, and hence serve as an index of hepatic elimination.
There exists a third group of drugs whose pharmacokinetics cannot be
explained adequately by the conventional single or multicompartment systems
discussed above. These are drugs which are subject to metabolism on their
initial pass through the liver after oral administration. This group includes,
among other drugs, propranolol (Shand et al. 1970) and propoxyphene (Perrier
and Gibaldi, 1972).
Enzyme affinity
The kinetic parameters of an enzymic reaction are defined by two
quantities namely Vmax and K . While K (Michaelis rate constant) is the m m
equilibrium constant of reaction, and represent the reciprocal of the affinity
of the enzyme for the substrate, Vmax is the measure of the velocity constant
of breakdown of the enzymes-substrate complex. These two quantities are bound
to influence the pharmacokinetic behaviour of a compound introduced into the
body.

141
Scope of the present investigation
Phenylacetic acid is conjugated mainly with amino acid (James et al. 1972 a)
1-na.phthyla.cetic acid with both amino acid and glucuronic acid (see Chapter 3),
hydratropic acid (except in the cat, see Chapter 5) and diphenylacetic acid (see
Chapter 4) are mainly conjugated with glucuronic acid. In this study an attempt
is made to correlate the metabolic patterns of these four arylacetic acids with
their pharmacokinetic behaviour, after intravenous administration, in the rabbit
and also their affinity for the conjugation sites (mitochondria and microsomes)
and the conjugating enzymes associated with these structures.
Results
Pharmacokinetics. The mean blood concentrations of 14C-labelled 1-naphthylacetic,
diphenylacetic and hydratropic acids at different time intervals in three rabbits
after an intravenous administration of these acids (69 i/mol/kg) are shown in
Tables 6:1, 6:2, 6:3 and 6:4 respectively, and after plotting the results on semi-
log paper, calculation of some of the pharmacokinetic parameters of these acids
were undertaken, the values of which are shown in Table 6:5. The blood samples
revealed on analysis the absence of any conjugates of these acids.
1 -Naphthylacetic acid. The semi-log plot of the mean blood concentration of
1 -naphthylacetic acid against time is shown in Fig. 6:1. This shows a mono-
phasic profiles and thus representative of one compartment system. Table 6:5
shows low values for blood clearance (Q, 0.72 ml. min 1), apparent volume of
distribution (VD, 0.137 1/kg) and elimination rate constant (Ke 9 0. 316 h-1
).
Diphenylacetic acid . The semi-log plot of the mean blood concentration of
diphenylacetic acid against time is shown in Fig. 6:2. This shows a biphasic pro -
file and thus representation of a multicompartment model (in this case taken
essentially as a two compartment system). Diphenylacetic acid has low clearance

142
Table 6:1
Mean blood concentrations of 1-naphthylacetic acid (pg/m1) in three rabbits
following intravenous administration of 28. 3 mg.
Time (min. )
5
10
15
Mean blood concentrations of 1 -naphthylacetic acid (µg/ml)
80.6 (59.9 - 102. 5)
90. 8 (75. 5 - 105. 9)
73.1 (68.7 - 81.7)
20 67. 4 (56. 7 - 80. 6)
30 55. 5 (45. 3 - 68. 0)
45 48. 3 (38. 9 - 57. 7)
60 34. 0 (28. 3 - 39. 7)
90 22.0 (17.3 -25.5)
120 13. 5 (9. 3 - 16. 0)
180 4. 0 ( 2. 6 - 5. 4 )
Table 6:2
Mean blood concentrations of diphenylacetic acid (µg/ml) in three rabbits
following intravenous administration of 32. 2 mg
Time Mean blood concentrations of diphenylacetic (min. ) acid (µg/m1)
5
10
44. 7
30.2
(42. 1 - 47. 2)
(30.1 - 30.4)
15 23. 5 (21. 8 - 26. 8)
20 15. 5 (12.1 - 18. 8)
30 9. 5 ( 8.1 - 10.1)
45 6. 1 ( 5. 8 - 6. 3 )
60 2.6
90 1.0
120 0. 3

143
Table 6:3
Mean blood concentrations of hydratropic acid (tteml) in three rabbits
following intravenous administration of 22. 7 mg
Time (min. )
5
Mean blood concnetrations of hydratropic acid (µg/m1)
59.4 (50.3 68. 5)
10 53. 0 (47. 0 - 59. 0)
15 38. 8 (30. 3 - 49. 5)
30 34. 5 (25. 0- 50. 5)
45 22. 2 (21. 0 - 23. 9)
60 19.9 (17. 8 - 22. 8)
90 14. 0 (12. 6 -16. 0)
120 9. 7 ( 9. 2 - 10.1)
180 4.5 ( 4.2 - 4.8)
Table 6:4
Mean blood concentration of phenylacetic acid (µg/m1) in three rabbits
following intravenous administration of 20. 5 mg.
Time (min. )
Mean blood concentrations of phenylacetic acid (pg/m1).
5 26. 2 (22. 4 - 29. 4)
10 19. 8 (14. 8 - 25. 2)
15 13.1 ( 7.4 - 20. 5)
20 11. 0 ( 8. 8 - 15. 0)
30 8.4 ( 7.0 - 9.7)
45 4. 2 ( 3. 5 - 5.3)
60 2. 2 ( 1. 8 - 2. 6)
90 O. 6 ( O. 5 - O. 7)
120 0. 4 ( O. 3 - 0.5)
180 0. 2

144
100 M
ean
bloo
d co
ncen
trat
ion
of 1
-nap
hthy
lace
tic
acid
(pg/
m1)
10
1.0
1 3
Time (h)
Fig. 6:1 Pharmacokinetic analysis of mean blood concentrations in three rabbits (ay. wt. 2. 2 kg) following intravenous administration of 28. 3 mg of [14C1-1-naphthylacetic acid.

145
Time ( h )
1
2
Fig 6:2 Pharmacokinetic analysis of mean blood concentrations in three rabbits (ay. wt. 2.2 kg) following intravenous administration of 32.2 mg of [14C]diphenylacetic acid.

Mea
n bl
ood
conc
entr
atio
n of
hydr
atro
pic
acid
(µg/
m1)
20
1.0
80 60
40
beta phase
alpha phase
0.1 -
1
146
Total (h) 1 2 3
Fig 6:3 Pharmacokinetic analysis of mean blood concentrations in three rabbits (ay. wt. 2.2 kg ) following intravenous administration of 22.7 mg of [14C]hydratropic acid.

Fig 6:4 Pharmacokinetic analysis of mean blood concentrations in three rabbits (ay. wt. 2.2 kg) following intravenous administration of 20.5 mg of 44 Cjphenylacetic acid.
blo
o d c
once
ntra
tion
of
phen
ylac
e tic
aci
d (d
ig/m
1)
alpha phase \
beta phase
Time (h )
147

Table 6:5
Pharmacokinetic parameters of arylacetic acids in three rabbits following intravenous administration of 69 pmolikg
Compound Compart - meat Model
Vc 1/kg
VD 1/kg
Ke h-1
K12
h-1 K21 h-1
Alpha (a)
h-1 aTi
2- h
Beta (g) h-1
gi- 1 2 h
Q
ml/min
1 -Naphthylacetic acid 1 - 0.137 0.316 - - - - O. 316 2.19 0.72
Diphenylacetic acid 2 0. 393 0.485 0.47 0. 053 1.57 1.492 0.469 0.495 1.40 4. 0
Hydratropic acid 2 0.320 0.256 0.21 0.604 2.44 1.75 0.396 0.296 3.3 1.33
Phenylacetic acid 2 0.301 5.481 0.086 2.0 3.07 0.835 0.83 0.314 8.66 28.7
VC = volume of central compartment ; VD = apparent volume of distribution ; Q = blood clearance
K12 = (a + 13) - (K21 + Ke) ; Ke = c /3/K21 ; K21 = (Aa(0)/3 + Ba(o)a) / (Aa(0) + Ba(o)) where Aa(o) = The intercept obtained
after extrapolating the alpha phase back to time zero. Ba(o) = The intercept obtained by extrapolating the beta phase back to time
zero.

149
(4.0 ml min-1) low apparent volume of distribution (0.485 1/kg) and low
elimination rate constant (0.47 h-1) although these values are higher than
in the case of 1-naphthylacetic acid (Table 6:5). The rate constant of distribution
represented by alpha is 1.492 h-1, and alpha half life ( aTi) is 0.469 h. The 2
ratio K12
: K21 is 1:30.
Hydratropic acid The semi-log plot of the mean blood concentration of hydra-
tropic acid against time is shown in Fig. 6:3. This shows a biphasic profile
and thus representative of a two-compartment model as stated above. Table 6:5
shows that hydratropic has a low clearance (1.33 ml. min-1), a low apparent
volume of distribution (0.256 1/kg) and low elimination rate constant (0.21 11-1).
The rate constant of distribution represented by alpha is 1.75 h-1 and the alpha
half life ( aTi) is 0.396 h-1. The ratio K12
: K21
is 1:4.
Phenylacetic acid The semi-log plot of the mean blood concentration of phenyl-
acetic acid against time is shown in Fig. 6.4. This shows a biphasic profile and
thus representative of a two-compartment model. Table 6:5 shows that phenyl-
acetic acid has a high clearance (28.5 ml. min-1), a high apparent volume of
distribution (5.481 1/kg) and a low elimination rate constant (0. 086 11-1). The
rate constant of distribution represented by alpha is 0.835 h-1 and the alpha
half life (aT 1) is 0.83 h. The ratio K12
: K21
is 2:3.. 2
Binding studies The binding of 1-naphthylacetic, diphenylacetic, hydratropic
and phenylacetic acids to initochondrial and microsomal fractions in 0.21VI Tris-HCI
buffer was investigated as described in Chapter 2, and the results are shown in
Table 6:5 and 6:7. The binding of these acids to mitochondria was in the proportion
phenylacetic acid, 40 ; 1-naphthylacetic acid, 10 ; diphenylacetic acid, 2 ; hydra-
tropic acid 1 (Table 6:6) and their binding to microsomes was in the proportion
hydratropic acid, 30 ; diphenylacetic acid, 27 ; 1-naphthylacetic acid, 20; phenyl-
acetic acid, 10 (Table 6:7).

Table 6:6
The binding of some arylacetic acids to mitochondria
Substrate concentration = 10 nmol
Binding in pmol / 10 mg protein
A B A -B
Compound Total Non-saturable Saturable (trapped and dissolved)
1 -Naphthylacetic acid 525 (521 - 527) 515 (509 - 519) 10 ( 8 - 12)
Diphenylacetic acid 330 (328 - 331) 328 (325 - 331) 2( 0- 3)
Hydratropic acid 599 (598 - 600) 598 (596 - 600) 1( 0- 2)
Phenylacetic acid 256 (251 - 264) 216 (203 - 229) 40 (35 - 48)

Table 6:7
The binding of some arylacetic acids to microsomes
Substrate concentration = 10nmol
Binding in pmol / 10 mg protein
A B A-B
Compound Total Novi saturable Saturable (trapped and dissolved)
1-Naphthylacetic acid 652 (648 - 654) 552 (546 - 558) 100 (96 - 102)
Diphenylacetic acid 400 (395 - 403) 265 (259 - 273) 135 (130 - 138)
Hydratropic acid 690 (681 - 696) 540 (525 - 555) 150 (141 - 156)
Phenylacetic acid 110 (101 - 115) 60 ( 46 - 74) 50 ( 41 - 55)

152
Enzyme affinity The conjugation of 1-naphthylacetic, diphenylacetic, hydra-
tropic and phenylacetic acids with glycine and glucuronic acid in vitro has been
examined as described in Chapter 2.
Glycine conjugation The experiment was performed in 0. 2M Tris-HC1 buffer pH 8.4
using both liver and kidney homogenates respectively as the enzyme sources. The
results are shown in Table 6:8. With kidney homogenate/there was 19% glycine
conjugate formed with phenylacetic acid as substrate (10 nmol), and none for the
other acids, but with liver homogenate it was 89% for phenylacetic acid, 1% for
1-naphthylacetic acid and none for both diphenylacetic and hydratropic acids.
Table 6:9 shows the kinetics of phenacetylglycine formation, while Fig. 6. 5 shows
the Lineweaver Burk plot of the results. The KM for the phenacetylglycine
formation was found to be 1.3 x 10-6M and the Vmax 5 nmol/ 40 mg liver / min.
Glucuronic acid conjugation The glucuronic acid conjugation of 1-naphthylacetic,
diphenylacetic, hydratropic and phenylacetic acids was studied using liver s the
microsomal fraction/as/enzyme source as described in Chapter 2. There was no
glucuronide formation detected with phenylacetic acid as substrate. The
glucuronidation kinetics for 1-naphthylacetic acid is shown in Table 6:10, diphenyl-
acetic acid, Table 6:11 and hydratropic acid Table 6:12 and the Lineweaver Burk
plots of these results are shown in Figs. 6:6, 6:7 and 6:8 respectively. The Km
for these acids were 1-naphthylacetic acid, 6.1 x 10-8
M ; diphenylacetic acid
2.3 x 10-8
M and hydratropic acid 2. 6 x 10-8
M. The V were 1-naphthylacetic max
acid, 11.1 nmol/mg protein/ min, diphenylacetic acid 1. 7 nmol/mg protein/ min and
hydratropic acid 1.4 nmol/mg protein/min.
Discussion
and The clearance / half-life of a drug can be used as indices of the intrinsic
capacity of the liver to metabolise drugs (Perrier and Gibaldi, 1974). The use

Table 6:8
Glycine conjugation
Substrate concentration = 10 nmol ; 0. 2M Tris-HC1 buffer pH 8.4
Glycine conjugation as % substrate/ 30min
Compound Kidney Liver
1 -Naphthylacetic acid 0 1.0
Diphenylacetic acid 0 0
Hydratropic acid 0 0
Phenylacetic acid 19 (16-23) 89 (80-93)
Table 6:9
Kinetics of phenacetylglycine formation
Buffer = 0.2 Tris-HC1 buffer pH 8.4.
S (nM) V 1/V 1 S x 0-5M nmo1/40mg liver / min
10 0.344 2.9 10
12.5 0.42 2.4 8
50.0 0.63 1.6 5
100.0 2.0 0.5 1
500.0 4.0 0.3 0.2
1000.0 5.0 0.2 0.1
153

154
Table 6:10
Kinetics of 1 -naphthylacetylglucuronide formation
Buffer = 0.2M
Phosphate buffer pH 5. 9
Substrate Initial
1/v 1/s I0-8M velocity (V)
Concentration (nmol/mg protein/ s (nM) minim
3. 3 0.59 1.7 3.0
4.2 0.75 1.3 2.4
8. 3 1.54 0. 7 1.2
16. 7 2.56 0.4 0. 6
33. 3 3.33 0.3 0. 3

Table 6:11
Kinetics of diphenylacetyl glucuronide formation
Buffer = 0.2M phosphate buffer pH 6. 2
S (nM)
V (nmol/mg protein/min)
1/V I/S x 10-8M
3.3 0.22 4.5 3.0
4.2 0.27 3.8 2.4
8.3 0.50 2.0 1.2
16.7 0.72 1.4 0.6
33.3 1.11 0.9 0.3
Table 6:12
Kinetics of hydratropoylglucuronide formation
Buffer = 0.2M phosphate buffer pH 5. 7
S (nM)
V (nmol/mg protein/min)
I/V I/S x 10-8M
3. 3 0.172 5. 8 3. 0
6. 7 0.308 3. 3 1.5
16. 7 0.60 1.7 0. 6
33. 3 0. 82 1.2 0. 3
50. 0 0.92 1.1 0. 2
66.7 1.06 0.9 0.15
155

156
Q
O
0
x 10 -5M
Fig. 6.5 Lineweaver-Burk plot of phenacetylglycine formation concentrations of phenylacetic acid were from 0. 01 - (50, 000 d. p. m. ) in the presence of 60 pmol of glycine. Reaction mixtures contained 3pmol MgCli , 20pmol giutathione, 0. 2M Tris/HCI buffer, pH 8.4 and 40 mg of rat liver homo - genate.
Km =1.3 x 10-6M V = 5nmo1/40 mg liver/mm

• 2-7
71) 0
0
-0.17 0
x 10-8M
Fig 6:6 Lineweaver-Burk plot of 1-naphthylacetylglucuronide formation. Concentrations of 1-naphthylacetic acid 3. 3 - 33.3 nM Incubation mixtures contained ltanol TJDPGA, 1. 5 mg microsomal protein and 0.2 M phosphate buffer pH 5. 9. Km = 6.1 x 10-8M, Vm = 11.1 nmol / mg protein/min.
157

158
6-
Fig. 6:7 Lineweaver -Burk-plot of diphenylacetyl.glucuronide formation. Concentrations of diphenylacetic acid were from 3. 3 to 33. 3 nM. The incubation mixtures contained Tµmol UDPGA, 1. 5
microsomal protein and 0.2 M phosphate buffer , pH 6.2 Km = 2. 3 x 10-8 M V = 1. 7 nmol/mg protein/min.

6- !•
0 -039
i/v
[nm
ol/m
g pr
otei
n/m
in].
6
2-
2 3
1 x 10 M
159
Fig 6:8 Lineweaver-Burk-plot of hydratropoylglucuronide formation. Concentrations of hydratropic acid were from 3. 3 - 66.7 nM. The incubation, mixtures contained linno1 UDPGA, 1.5 mg , mic.rosomal protein and Q. 2 M phosphate buffer, pH 5.7. K 2.6 x 10-8M V =1.4 nmol/mg protein/min. m

160
of these parameters as indices of hepatic elimination is highly dependent on the
pharmacokinetic characteristics of a particular drug. For drugs which confer
the pharmacokinetic characteristics of a single compartment model on the body,
both half-life and clearance serve a meaningful measures of hepatic elimination.
However, for a drug which confers multicompartment characteristics on the
body, biologic half-life does not reflect properly the intrinsic metabolic activity
of the liver. Clearance on the other hand, is a direct measure of this activity
regardless of the number of compartments conferred upon the body by a drug
provided the liver is an integral part of the central compartment (Perrier and
Gibaldi, 1974).
1-Naphthylacetic acid confers a one compartment model, while diphenyl-
acetic, hydratropic and phenylacetic acids confer a two compartment model on
the rabbit, but the rate of distribution (alpha) of diphenylacetic and hydratropic
acids is almost twice that of phenylacetic acid a factor which is reflected on the
alpha half life (aT1). The total body water of 2.2 kg rabbit is about 1. 7 litres 2
(extrapolated from the data of West et al. (1948) on 3. 5 kg infant with 2. 7 litres
body water). The apparent volume of distribution for phenylacetic acid is more
than the total body water of the rabbit and indicates that this acid is highly bound
to extravascular tissues. This acid has also a relatively higher blood clearance
compared with the other acids. 1-Naphthylacetic and hydratropic acids are
distributed into 17 and 32% respectively of body water and have very low blood
clearance and thus indicating a high vascular tissue binding for these acids.
Diphenylacetic acid on the other hand is 62% distributed into the body water and
has a fairly moderate blood clearance. The elimination rate constant is very
low for phenylacetic and this is also reflected on its very high biologic half-life
(flT1) when compared with the other acids. These pharmacokinetic parameters

161
in general may be influenced by the degree of binding of these acids to the
metabolic sites (in this case the conjugation sites, mitochondria and endoplasmic
reticulum), their affinity for the conjugating enzymes associated with these
structures and also their rate of conjugation with amino acid and / or glucuronic
acid.
The binding of 1-naphthylacetic, diphenylacetic, hydratropic and phenylacetic
acids to mitochondria and microsomes suggests a different binding capacities.
This differential binding capacity may arise as a result of a possible difference
in the mitochondrial and microsomal partition coefficient of these acidsdegree
of ionization, size and molecular geometry. These may also be the factors
influencing the glycine conjugation and the glucuronidation affinity of these acids.
Diphenylacetic and hydratropic acids have low binding capacity for mitochondria
and these did not form glycine conjugates while phenylacetic and 1-naphthylacetic
acids have a higher binding capacity and formed glycine conjugates although much
more in the case of phenylacetic acid. On the other hand phenylacetic acid has
the least binding capacity for the microsomes and it does not form a glucuronie
acid conjugate, whereas diphenylacetic, hydratropie and 1-naphthylacetic acids
with high binding capacity formed glucuronic acid conjugates although at different
rates and have different affinities for the conjugating enzymes as well. The
lipid solubility of a compound is known to determine not only its penetration into
cells and metabolic sites but also influences its metabolism (Gaudette and Brodie,
1959). Highly lipid soluble N-alkylamines are dealkylated in the microsomes
while the low lipid soluble ones in the mitochondria (Gaudette and Brodie, 1959 ;
Axelrod, 1956 ; Mackenzie and Frisell, 1958). The arylacetic acids seem to fall
into this discrimination pattern as regards conjugation sites and type of conjugation.
1-Naphthylacetic and diphenylacetic acids which have high binding capacity for

162
the microsomal fractions and form glucuronides have high lipid solubilities
while phenylacetic which has low lipid solubility has low binding capacity for
microsomes but high binding capacity for mitochondria does notform a glucuronic
acid but an amino acid conjugate. Although the difference in the lipid solubilities
of phenylacetic and hydratropic acids is small, it may be suggested that hydra-
tropic acid is able to fluidise the lipid region of the microsomes, a character -
istic Cater et al. (1974) have indicated may qualify a compound for micro-
somal metabolism.
The binding of the arylacetic acids to the subcellular fractions as reported
here, is rather a static reaction but correlating the binding to the conjugation
reaction (the kinetics) which is a dynamic reaction it may be possible to have
an insight to what happens in vivo when these compounds are administered to
an animal. Since non-polar compounds and their lipophilic metabolites have
more affinities for the metabolic sites than their polar conjugates (see Bickel
and Bonier, 1974 ; Glaumann et al. 1970 ; Nilsson et al, 1971) it follows
therefore that amino acid and glucuronic acid conjugates of these arylacetic acids
do have lower affinities for the conjugation sites than the parent compounds.
The conjugates therefore are excreted as soon as they are formed in the body
setting up a dynamic process which may be representative of the pharmaco-
kinetic study in the rabbit. It may be argued therefore that the pharmacokinetic
parameters calculated represent a combination of the binding to subcellular
fractions, metabolic conjugation and the influence the physico-chemical properties
of these arylacetic acids have on their metabolism in the rabbit.
Summary and Conclusion
1-Naphthylacetic acid confers a one-compartment model, while phenylacetic,
diphenylacetic and hydratropic acids confer a two-compartment model on the
rabbit. While phenylacetic acid has a high apparent volume of distribution, a high blood

163
clearance (Q) a high biologic half-life and a very low elimination rate constant,
1-naphthylacetic, diphenylacetic and hydratropic acids have low apparent volume
of distribution, relatively lower blood clearance, a low biologic half- life and a
high elimination rate constant. It may be suggested that the pharmacokinetic
parameters of phenylacetic acid may represent the index for arylacetic acids
which are conjugated mainly with an amino acid whereas those of the other three
acids for arylacetic acids which are conjugated mainly with glucuronic acid.
On the basis of the in vitro work it is also suggested that the pattern
of conjugation of an arylacetic acid in the rat is influenced by its affinity for
uptake (as measured by binding) by mitochondria and the endoplasmic reticulum
and affinity for the conjugating enzyme systems associated with these -structures.

164
CHAPTER SEVEN
Page
General Discussion and Conclusion 165

165
General Discussion and Conclusion
The previous chapters constitute an examination of the metabolic
conjugation pattern of 1-naphthylacetic, diphenylacetic and hydratropic acids
in some selected species and the effect of dose level on the pattern of
conjugation in the rat of these acids. Additionally the pharmacokinetic behaviour
of these acids and phenylacetic acid in the rabbit, and their relative affinity
for rat mitochondrial and microsomal fractions and the conjugating enzymes
associated with these structures have been studied.
1 -Naphthylacetic acid showed a distinct species variation in its metabolic
conjugation pattern in all the species studied, man and the Old World monkeys
(except the marmoset) and bushbaby mainly with amino acids and to a small
extent with glucuronic acid,the eat with amino acids and to a small extent with
glucuronic acid the cat with amino acids extensively, the rat and rabbit
principally with glucuronic acid and the fruit bat entirely with glucuronic acid.
Diphenylacetic acid on the other hand is conjugated with glucuronic acid irrespective
of the species, but hydratropic acid is conjugated mainly with glucuronic acid in
man, rhesus monkey, rat and rabbit and with both amino acids and glucuronic
acid in the cat. This study and similar work reported elsewhere in the
literature suggest that there is a shift from amino acid to glucuronic acid
conjugation with increase in complexity of the chemical structure of arylacetic
acids in most species. This concept applies to a limited degree to cat and some
other carnivores that are defective in glucuronic acid conjugation (French et al, 1974 ;
Caldwell et al, 1975b). Thus, arylacetic acids of relatively simple chemical
structure such as phenylacetic acid and its simple derivatives form mainly amino
acid conjugates (James et al, 1972 a,b) where as other more complex arylacetic
acids such as diphenylacetic acid (see Chapter 4), indomethacin (Harman et al, 1964)

166
and iopanoic acid (McChesney and Hoppe, 1954) form glucuronic acid
conjugates.
Arylcarboxylic acids such as benzoic and salicylic acids are conjugated
in man, and sub-human primates and non primate species with amino acid and
glucuronic acid the quantities differing with species. The simple primary
arylacetic acid such phenylacetic acid is conjugated mainly with amino acid
irrespective of species, but there is discrimination in the type of amino acid utilised.
The glutamine conjugation is restricted to man, Old and New World monkeys
while glycine conjugation is utilised by the rest. Taurine conjugation seems to be
predominate in the New World monkeys (except the marmoset) and the bushbaby
although their is a haphazard quantitative distribution in the other species (James
et al , 1972 a). In the more complex primary arylacetic acids such as
1-naphthylacetic acid there is a shift from amino acid to glucuronic acid
conjugation in most species except the cat, some other carnivores (see Chapter 3 ;
Caldwell et al, 1975b)the New World monkeys (except the marmoset) and the
bushbaby which largely conjugate this acid with glycine and taurine. For indolyl-
acetic acid the shift towards glucuronic acid conjugation was observed in man and not
in the other species, but there was glutamine conjugation in man, New and Old
World monkeys, a conjugation reaction which seems to be restricted to these species
(Bridges et al, 1974). With further increase in the complexity of the chemical
structure of primary arylacetic acids, there appears a distinctive discrimination
in the metabolism and conjugation pattern between the carnivores and the rest
of the species. Man conjugates indomethacin with glucuronic acid but the dog ex-
cretes it unchanged while, rat, guinea pig and the monkey conjugate its metabolised
derivative (N-deschlorobenzoyl and 0-desmethyl) with glucuronic acid(Harman et al
1964). Another example is p-(cyclopropylcarbonyl) phenylacetic acid (SQ 20, 650)

167
which is conjugated with glucuronic acid by the monkey but with taurine
by the dog (Lan et al. , 1975).
There has been little information in the metabolic conjugation pattern
of secondary arylacetic acids inmany species, but the available information
for hydratropic acid suggest a total shift from amino acid to glucuronic acid
conjugation in the species examined except in the carnivores. Although
hydratropic acid is conjugated with glucuronic acid by man, rhesus monkey,
rabbit, rat, the cat still conjugates this acid with glycine and taurine (38% of
the dose) and glucuronic acid acid (40%) (see Chapter 5). The dog is also
reported to conjugate this acid with glycine (Kay and Raper, 1922). It is
interesting to note that with the more complex secondary arylacetic acid,
diphenylacetic acid there is a total shift from amino acid to glucuronic acid
conjugation irrespective of species (see Chapter 4) a result which is in agree-
ment with the finding of Miriam et al , (1927a)in man, dog and rabbit.
The only information available on the metabolic conjugation of simple
tertiary arylacetic acid is the work of Robinson & Williams (1955) which showed
that a, a -dimethylphenylacetic acid is conjugated with glucuronic acid by the
rabbit. Miriam et al (1927 b) have also reported that the more complex chemical
structure, triphenylacetic is totally excreted uncharged by rabbit, dog and man.
This shift in the conjugation pattern with increase in the complexity of
the chemical structure of arylacetic acids has confirmed the highly substrate-
dependency of some conjugation reactions which are known to be deficient in
some species. For example, the fruit bat does not form glycine conjugate
with benzoic, 1-naphthylacetic and diphenylacetic acids but does with phenylacetic
acid (see Bababunmi et al, 1973 ; Ette et al , 1974 ; Chapters 3 and 4). The
cat is known to be deficient in glucuronide formation, and does not form glucuronic

168
acid conjugates with benzoic, phenylacetic and 1-naphthylacetic acids but
does with hydratropic and diphenylacetic acids (Bridges et al, 1970 ; James et al. ,
1972 a; Chapters 3, 4 and 5).
The effect of dose on the metabolic conjugation pattern of 1-naphthylacetic,
diphenylacetic and hydratropic acids has also been studied in bile duct- cannulated
rats (Chapters 3,4 and 5). At low doses the glycine conjugate is the major
urinary metabolite of 1-naphthylacetic acid but with increase in dose glucuronic acid
conjugation takes over, but at both low and high doses diphenylacetic and hydra-
tropic acids are conjugated extensively with glucuronic acid. This finding suggests the
that/glycine conjugation mechanism has limited capacity and that when it is exhausted
the glucuronic acid mechanism takes over, but with any compound which is con -
jugated with glucuronic acid only the pattern of metabolites will not change very
much with dose unless this is excessive. The cat as already mentioned has a
defective glucuronic acid conjugation for certain compounds including benzoic
acid. Consequently benzoic acid is more toxic to the cat than most common species
of animals (Bedford & Clarke, 1971, 1972), for the cat conjugates benzoic acid
entirely with glycine which is limited in supply, and glucuronic acid conjugation is
not available to take over when large doses of benzoic acid are administered. The
effect of dose on the shift from sulphate and glutathione conjugations to
glucuronic acid conjugation was discussed in Chapter 3.
These differences in the metabolic conjugation pattern of the aryl -
carboxylic and --acetic acids may be due to differences in their physico- chemical
properties which may influence their pharmaco-kinetic behaviour, when
administered to an animal, degree of penetration to the conjugation sites
(mitochondria and microsomes) and the affinity for the conjugating enzymes
associated with these structures. Some physico-chemical properties of some

169
selected acids are shown in Table 7:1. The acids are 99.9% ionized at pH 7.4
and thus the ionization does not play a significant role in there different metabolic
conjugation pattern, but Table 1:11 shows the influence of pKavalues on the pattern
of metabolism of chlorophenols. The data in Table 7:1 suggests that log P (P is
the molar partition coefficients between octanol and water) and the substitution
at the a-carbon and molecular geometry are the most influencing factors in
determining the metabolic conjugation pattern of arylacetic acids. Phenylacetic
acid with the least log P value (1.43) has planar molecular geometry and no a-carbon
substitution and is conjugated mainly with amino acids in all the species, but
the smaller molecule, benzoic acid with slightly higher log P (1.87), though
with planar molecular geometry has its a-carbon totally substituted and this
acid is conjugated with amino acid and glucuronic acid except in the carnivores
which conjugate mainly with amino acid. 1-Mphthylacetic acid on the other hand
has a high log P value (3.14), and is conjugated with both amino acid and
glucuronic acid (except in some carnivores) but like phenylacetic acid it has a
planar molecular geometry. Hydratropic and diphenylacetic acids have log P
values of 1.93 and 3.09 respectively, and have non-planar geometry with an
a-carbon substitution (one proton at a-C) are conjugated mainly with glucuronic
acid except in the case of hydratropic acid in the cat, ferret and dog. a, a-
Dimethylphenylacetic and triphenylacetic acids, although with high log P values
(2.46 and 5.08 respectively), with non-planar geometry and have their a-carbons
totally substituted, only a, a-dimethylphenylacetic acid has been reported to
form a glucuronic acid conjugate (Robinson & Williams, 1955) whereas triphenylacetic
acid is excreted unconjugated (Miriam et al. , 1972 b). This suggests that the
very high log P value is not a favourable factor for glucuronic acid conjugation,
and one wonders if the relationship between glucuronic acid conjugation and log P

Table 7:1
Characteristics of some Arylacetic Acids
Compound Log P ++
p-Ka
Molecular geometry
a -Carbon Molecular
Conjugation with :
Planar Non-planar Amino Glucuronic substitution(No. weight acid acid
Benzoic acid
Phenylacetic acid
1 -Naphthylacetic acid
Hydratropic acid
Diphenylacetic acid
a, a-Dime.thylphenylacetic acid
Triphenylacetic acid
1.87
1.43
3.14
1.93
3.09
2.46
5.08
4.21
4.32
4.21
4.6
3.94
3.96
+
+
+
-
-
of protons on a C)
122
136
186
150
212
164
288
+
+ **
+ *
-
-
-
+
-
+
+
+
+
+ (0)
- (2)
- (2)
+ (1)
+ (1)
+ (0)
+ (0)
except in the carnivores 4+ data obtained or calculated from Fujita et al . (1964) and Nys & Rekker (1973)
** except in the fruit bat

171
values is parabolic since at low and very high log P values there is no
glucuronic acid conjugation but at moderately high log P values glucuronic acid
conjugation seems to be favoured, or in the case of triphenylacetic acid whether
a steric effect is the inhibitory factor in this acid not being conjugated. The
glucuronyl acceptor substrates are lipid soluble and the affinity of enzymes
increase with their lipophilicity (Hanninen and Alanen, 1966), but from the
relationship between glucuronic acid conjugation and log P values shown above,
it is suggested that the compounds which are too lipophilic become trapped in
the external lipids of the cell walls and do not enter the cell and tim'efore have
no access to the metabolic conjugation sites (endoplasmic reticulum). A
similar concept was used to explain the parabolic relationship between lipo-
philic character and isoeffective doses of ethers and alkanes (Jeppsson, 1975)
and partition coefficient and narcosis (Hansch et al. , 1968). The conjugation
pattern shown by arylacetic acids is also seen in other compounds, for example
phenols of small molecular size are mainly conjugated with sulphate (Williams 1938 ;
Capel et al. , 1972) while those with large molecular size are conjugated mainly
with glucuronic acid (see Dodgson et al. , 1948 ; Mazur and Shorr, 1942). The
physico-cheinical properties shown in Table 7:1 may also be the factors directing
the pharmacokinetic behaviour of these acids when administered to an animal.
The pharmacokinetic behaviour of 1-naphthylacetic, diphenylacetic,
hydratropic and phenylacetic acids has been studied in the rabbit. The first
three acids are conjugated mainly with glucuronic acid, while phenylacetic acid
is conjugated mainly with glycine in this species. The result (see Table 6:5)
shows that phenylacetic acid has a high apparent volume of distribution, a
relatively high blood clearance, a low elimination rate constant and a high
biological half life (beta half life) while the reverse is true for 1-naphthylacetic,

172
hydratropic and diphenylacetic acids. This indicates that phenylacetic acid
is more quickly cleared from the blood and is heavily bound to extravasctlar
tissues and also has a relatively poor metabolic elimination mechanism as
compared with the other acids which seem also to be restricted in their binding
to either the vascular tissues or a preferential target site (es. endoplasmic
reticulum). From the binding studies (see Chapter 6) 1-naphthylacetic,
diphenylacetic and hydratropic acids have a higher affinity for the rat m5.crosomal
fraction than phenylacetic acid and the reverse is the case for the rat mIto-
chondrial fraction. The lipid solubility of a compound is known to determine
not only its penetration into cells and metabolic sites but also influences in. its
metabolism ( Gaudette and Brodie, 1959). Highly lipid soluble N-alkylamines are
dealkylated in the microsomes while the low lipid soluble ones in the mitochondria
(see, Gaudette and Brodie, 1959 ; Axelrod, 1956 ; Mackenzieand Frisell, 1958).
The in vitro conjugation study of 1-naphthylacetic, diphenylaceticlhydratropic
and phenylacetic acids with glucuronic acid and glycine (see Chapter 6) is in
agreement with the in vivo glucuronidation and amino acid conjugation capabilities
of these acids and has a direct relationship to their affinities for the metabolic
conjugation sites (as measured by binding to mitochondria and microsomal
fractions). 1-NaphthylaceticIdiphenylacetic and hydratropic acids are more lipid
soluble than phenylacetic acid thus the result of this study is in agreement with the
concept of Gaudette and Brodie, (1959). But non-polar compounds and their
lipophilic metabolites have more affinities for their metabolic sites than their
polar conjugates (see Bickel and Borner, 1974 ; Glaumann et al. , 1970) ; it follows
therefore that amino acid and glucuronic acid conjugates of these arylacetic acids
do have lower affinities for the conjugation sites than the parent compounds. The
conjugates therefore are excreted as soon as they are formed in the body setting

173
up a dynamic process which may be representative of the pharmacokinetic
study in the rabbit.
Amino acids and glucuronic acid have different conjugation mechanisms
as described in Chapter 1. For amino acid conjugation the substrate has to be
activated first (R. COSCoA) before reacting with the conjugating agent, whereas
for glucuronic acid conjugation the substrate reacts directly with the activated
(UDPGA) conjugating agent. The question arises as to whether or not the
arylacetic acids which do not form amino acid conjugates do form the activated
intermediate (R. COSCoA). These intermediates have been reported for benzoic,
phenylacetic , salicylic and.bile acids (Schacter and Taggart, 1953 ; Moldave and
Meister, 1957 ; Elliott, 1956 ; Tishler and Goldman, 1970) which are subsequently
converted to the respective amino acid conjugates by the specific amino acid N-
acylase . It may be interesting therefore to investigate at what stage in the
amino acid conjugation mechanism compounds like diphenylacetic acid are
defective. For example if the activated intermediates diphenylacetyl-AMP and
-CoA are formed then the deficiency is at the amino acid N-acylase enzyme stage
which may be explained in the terms that the activated intermediate -enzyme binding
forces may be too small to force the enzyme into the catalytically active conformation
and hence no conjugation takes place. On the other hand if these activated inter -
mediates are not formed it may be suggested that this reaction is not thermodyna-
mically feasible which may be a result of the chemical structure and the intrinsic
energy of the compound) and therefore prefer a different metabolic conjugation
pathway which is thermodynamically feasible.
In conclusion the metabolic conjugation pattern of arylacetic acids is
greatly influenced by the physico-chemical properties of the acid (such as lipid
solubility, chemical structure and molecular size and geometry) and the affinity

174
for the conjugation sites (mitochondria and endoplasmic reticulum) and the
conjugating enzymes associated with these structures.

APPENDIX.
175
Contents
Pages
176
Mass Spectral Fragmentation Pattern
1-Naphthylacetylglycine Methyl Ester 176
1 -Naphthy lac ety I -L -glutamine Methyl Ester 176
1-Naphthylacetyltaurine Methyl Ester 176
Diphenylacetyiglycine Methyl Ester 176
Diphenylacetyl -L -glutamine Methyl Ester 177
Diphenylacetyltaurine Methyl Ester 177
Diphenylacetylglucuronide Methyl Ester 177
Hydratropoylglycine Methyl Ester 178
Hydratropoyl-L-glutamine Methyl Ester 178
Hydratropoyltaurine Methyl Ester 178
Hydratropoylglucuronide Methyl Ester 178

105 cHpONFICH
2COOCH
3.
91\1
176 Mass Spectral Fragmentation Pattern
1 -Naphthylacetylglycine methyl ester
142 11681 1
CH IC*11CH2 COOCH3
1-Naphthylacetyl-L-glutamine methyl ester
142I164 I 1
CH 14THCHCOOCH 2I,_ ,_ L _ _ _ 3 — —116
0 - - --1 T H2
I CH 1 2 I CONH2 I
187
1 -Naphthylacetyltaurine methyl ester
1421 166
I 115 I I
CH21CONH1CH2CH2S03ICH3
185
Diphenylacetylglycine methyl ester
167

167i
I 1107
I
I ; CHpOp
COCH3
43
OH
177
Diphenylacetyl-L-glutamine methyl ester _
167
10 I CHICONHCHCOOCH3
1\ 1 CH 2
I CH, 1187 1 - CONH2
Diphenylacetyltaurine methyl ester
167 I 166
152;
CHFONHCH2CH2S031CH3
Diphenylacetylglucuronide methyl ester

178
Hydratropoylglycine methyl ester
I 116
1
- I-1- I CH4ONHCH2COOCH3
I 91
1051
Hydratropoyl-L-gliitamine methyl ester
1 187 CH
i 3 CH;
I
C ONHCHCOOCH3 1
H2 —
C — 1 I
I CH2
bONH2
— 116
Hydrotropoyltaurine methyl ester
1166 CH I
r „3-1, CHCONHCH CH SO CH
I I 2 2 3 3 I
911 1051 1131
Hydratropoyleucuronide methyl ester
j 191
I COOCH3
CH / I e -p xpo° I I I OH
91 OBI
101 173

179 REFERENCES
Abou-El-Makarem, M. M. , Millburn, P. , Smith, R. L. , Williams, R. T.
(1967) Biochem. J. 105, 1289-1293.
Adams, S. S. , Bough, R. G. , Cliffe, E. E. , Lessel, B. & Mills, R. F. N.
(1969) Toxicol. Appl. Pharmacol. 15, 310.
Armstrong, M.D., Shaw, K. N. F. & Wall, P.E. (1956) J. Biol. Chem. 218,
293 - 303.
Axelrod, J. (1954) J. Pharmacol. 110, 315.
Axelrod, J. (1956) J. Pharmacol. 117, 322.
Bababunmi, E. A. , Smith, R. L. & Williams, R. T. (1973) Life. Sci. 12,
317 - 326.
Bahr, C. Von, Borga, 0. (1971) Acta pharmacol. (Kbh) 29, 359 - 374.
Baldwin, B.C., Robinson, D. & Williams, R. T. (1960). Biochem. J. 76,
595.
Bedford, P. G. C. & Clarke, E. G. C. (1971) Vet. Record pp. 599 - 601.
Bedford, P. G. C. & Clarke, E . G. C. (1972) Vet. Record. pp. 53 - 58.
Bernhard, K., Caflisch-Weill, H. (1949) Hely. Chim.Acta. 32, 625.
Bickel, M. H. & Bonier (1974) Naunyn. Schmiedeberg's Arch. Pharmacol.
284, 339 - 352.
Bickel, M. H, , Steele, J.W. (1974) Chemico-biol. Interactions. 8, 151 - 162.
Blether, M. (1966) Biochem. Biophys. Res. Commun. 23, 293.
Bohme, C. & Grunow, W. (1974) Arch. Toxicol. 32, 227 - 231.
Boyland, E. & Chasseaud, L. F. (1969) Adv. Enzymol. Relat. Areas Mol.
Biol. 32, 173 - 219.
Branton, D., Elgsaeter, A. & James, R. (1972), FEBS 8th Meeting, p. 165,
North Holland / American Elsevier, Amsterdam.
Braun, G.A., Poos,G.I. & Soudijn, W. (1965) Eur. J. Pharmacol. 1, 58 - 62.

180
Bray, H. G. & James, S.P. (1960) Biochem. J. 74, 394.
Bray, H. G. , Clowes, R. C. , Thorpe, W.V. Wnite, K. & Wood, P.B. (1952)
Biochem. J. 50, 583.
Bray, H. G. , Franklin T. J. & James, S. P. (1959) Biochem. J. 73, 465.
Bray, H. G, Humphris, B.G., Thorpe, W.V. , White, K. &Wood, P.B.
(1955) Biochem. J. 59, 162.
Bray, H. G. , Lake, H. J. , Neale, F. C. , Thorpe, W. V. & Wood, P.B.
(1948) Biochem. J. 42 434.
Bray, H. G. , Thorpe, W. V. & Wood, P. B. (1949 a) Biochem. J. 44, 39.
Bray, H. G. , Thorpe, W. V. & Wood, P. B. (1949 b) Biochem. J. 45, 45.
Bridges, J.W., Davies, D. S. & Williams, R. T. (1967) Biochem. J. 105',
1261 - 1267.
Bridges, J.W., Kibby, M.R. & Williams, R. T. (1965) Biochem. J. 96,
829 - 836.
Bridges, J.W., Evans, M. E. , Idle, J. R. , Millburn, P., Osiyemi, F. 0.,
Smith, R. L. & Williams, R.T. (1974) Xenobiotica, 4, 645 - 652.
Bridges, J.W., French, M.R. , Smith, R. L. & Williams, R. T. (1970)
Biochem. J. 118, 47 - 51.
Bruce, R. B. , Turnbull, C. B. , Newman, J. H. & Pitts, J. C. (1968) J.
Medicin. Chem. 11, 1031 - 1034.
Burns, J. J. , Rose, R.K. , Chenkin, T. , Goldman, A., Schulert , A. &
Brodie, B.B. (1953) J. Pharmacol. Expt. Then 109, 346 - 356.
Caldwell, J., French, M.R. , Idle, J.R., Renwick, A.G., Bassir, 0. &
Williams, R. T. (1975 b) FEBS Lett. 60, 391 - 395.
Caldwell, J., Koster, U., Smith, R. L. & Williams, R.T. (1975 a) Biochem
Pharmacol. 24 2225 - 2232.

181
Caldwell, J, Moffatt, J.R. , Smith, R. L. (1976) Xenobiotica "in the press".
Capel, I.D., French, M.R. , Millburn, P. , Smith, R.L. & Williams, R. T.
(1972) Xenobiotica 2, 25.
Cater, B.R. , Chapman, D. , Hawes, S. M. & Saville, J. (1974) Biochim.
Biophys. Acta. 363, 54 - 69.
Cerecedo, L. R. & Sherwin, C. P. (1923) J. Biol. Chem. 58, 215 - 224.
Cerecedo, L . R. & Sherwin, C. P. (1924) J. Biol. Chem. 62, 217 - 230.
Charalampous, K.D., Orengo, A. , Walker, K.E. & Kinross-Wright, J. (1964)
J. Pharmacol. Exp. Ther. 145, 242 - 2460
Chasseaud, L. F. , Fry, B.J. , Valzelli, G. & Tosolini, G. O. (1974)
Arzneim-Forsch (Drug Research) 24, 1606 - 1609.
Combs, H.I. & Hele, T. S. (1927) Biochem. J. 21, 606.
De Eds, F., Booth, A.N. & Jones, F. T. (1955) Fed. Proc. 14, 332.
Deichmann, W.B. & Thomas, G. (1943) J. Industr. Hyg. , 25, 286.
Delvigs, P., Mclsaac, W. M. & Taborsky, R. G. (1965) J. Biol. Chem. 240,
348 - 350.
Dingell, J. V. , Sulser, F. & Gillette, J.R. (1964) J. Pharmacol. Exp. Ther.
143, 14 - 22.
Dodgson, K. S. , Garton, G.A., Stubbs, A. L. & Williams, R. T. (1948) Biochem.
J. 42, 357.
Dodgson, K. S. , Smith, J. N. & Williams, R. T. (1950) Biochem. J. , 46, 124.
Drach, J. C. & Howell, J. P. (1968) Biochem. Pharmacol. 17, 2125 - 2136.
Dutton, G.J. (1961) Proc. 1st Interntl. Pharmac. Meeting, Stockholm, 1961,
6, 65. , Pergamon Press.
Dutton, G.J. (1966) Glucuronic Acid Free and Combined, Academic Press,
N.Y.

182
Dziewiattowski, D.D. , Venkataraman, A. & Lewis, H.B. (1949) J. Biol.
Chem. 178, 169.
Elliott, W.H. (1956) Biochem. J. 62, 433 - 436.
Ernster, L. & Jones, L. C. (1962) J. Cell. Biol. 15, 563.
Estrado-O, S. , Montal, M. , Celiz, H. & Carabez, A. (1970) Eur. J. Biochem. ,
12, 227 - 235.
Ette, S.I. French, M.R. , Smith, R. L. & Williams, R. T. (1974) FEBS Lett.
46, 134 - 136.
Evans, 1VZ (1972) Ph.D. Thesis, Univ. of London.
Fiehn, W. & Hasselbach, W. (1970) Eur. J. Biochem. 13, 510 - 518.
Foulkes, D. M. (1970) J. Pharmacol. Exp. Ther. 172, 115 - 121.
French, M. R. , Bababunmi, E.A. , Golding, R.R. & Bassir, 0., Caldwell, J.,
Smith, R. L. & Williams, R. T. (1974) FEBS Lett. 46, 134 - 136.
Friedmann, E. & Masse, C. (1910) Biochem. J. 27, 97.
Fuccella, L. M. , Goldaniga, G. C. , Moro, E. , Tamassia, V. , Tosolini, G. P.
& Valzelli, G. (1973) Eur. J. Clin. Pharmacol. 6, 256.
Fujita, T. , Iwasa, J. & Hansch, C. (1964) J. Am. Chem. Soc., 86 , 5175 - 5180.
Gangolli, S.D. , Shilling, W. H. , Grasso, P. & Grant, I. F. (1974) Biochem.
Trans. , 2, 310.
Gaudette, L.E. & Brodie, B.B. (1959) Biochem. Pharmacol. 2, 89 - 96.
Gibaldi, M. & Perrier, D. (1972) J. Pharm. Sci. 61, 952 - 954.
Gillette, J. R. (1973) J. Pharmacokinetics and Biopharmaceutics 1, 497 - 520.
Gingell, R. (1975) Drug Metab. and Disposition 3, 42 - 46.
Gitler, C. (1972) Ann. Rev. Biophys. Bioengineer. 1, 1 - 92.
Glaumann, H. , Nilsson, R. Danner, G. (1970) FEBS Lett. 10, 306 - 308.
Goldaniga, G. C. , Pianezzola, E. & Valzelli, G. (1973) Carolo Erba Internal
Report.

183
Goldstein, A., Lowney, L. I. & Pal, B.K. (1971) Proc. Natl. Acad. Sci.
U. S. A. , 68, 1742 - 1747.
Gosselin, R. E. , Gabourel, J.D., Kaiser, S.C., & Wills, J.H. (1955)
J. Pharmacol. 115, 217.
Grunow, W. & Bohme, C. (1974) Arch. Toxicol. 32, 217 - 225.
Hanninen, 0. & Alanen, K. (1966) Biochem. Pharmacol. 15, 1465 - 1467
Hansch, C., Steward, A.R., Anderson, S. M. & Bentley, D. (1968) J. Med.
Chem., 11, 1 - 11.
Harley-Mason, J. , Laird, A. H. & Smithies, J. R. (1958) Confinia. Neurol (Basel)
18 152 - 155.
Harman R. E. , Meisinger, M. A. P. , Davis, G. E. & Kuehl, F. A. Jr. (1964)
J. Pharmacol. Exp. Ther. 143, 215 - 220.
Hartles, R. L. & Williams, R. T. (1947) Biochem. J. 41, 206.
Hartles, R. L. '& Williams, R. T. (1949) Biochem. J. 44, 335.
Hazelwood, G.A.D. (1967) J. Lipid. Res. 8, 535 - 550.
Hawksworth, G. M. , Drasar, B. S. & Hill, M. J. (1971) J. Med. Microbiol.
4, 451 - 459.
Hers, H. G. & de Duve. C. (1950) Bull. Soc. Chim. Biol. 32, 20.
Hinks, N. T. , Bolliger, A. (1957) Aust. J. exp. Biol. med. Sci. 3 37.
Hirom, P.C., Millburn, P., Smith, R. L. & Williams, R.T. (1972) Biochem.
J. 129, 1071 - 1077.
Hitchcock, M., Smith, J. N. (1966) Biochem. J. , 98, 736.
Hubbell, W. L. & McConnell, H. M. (1971) J. Am. Chem. Soc. 93 , 314 - 326.
Hubbell, W.L. & Wunderlich, F. (1973) Biochim, Biophys. Acta, 29, 621 - 628.
Idle, J.R. , Millburn, P. & Williams, R. T. (1975) FEBS. Lett. 59, 234 - 236.
James, M. O. , Smith, R. L. , Williams, R. T. & Reidenberg. M. (1972 a) Proc.
R. Soc. Lond. B. 182, 25 - 35.

184
James, M. 0. , Smith, R. L. & Williams, R. T. (1972 b) Xenobiotica. 2
499 - 506.
Jeffay, H. & Alvarez, J. (1961) Anal. Chem. 33, 612 - 615.
Jeppsson, R. (1975) Acta Pharmacol et toxicol. , 37, 56 - 64.
Jollow, D.J. , Thorgeirsson, S. S. , Patter, W. Z. , Mitchell, J.R. , Gillette,
J.R. &Brodie, B.B. (1973) Fed. Proc. „ 305
Junkman, W. (1941) Klin. Wschr. , 20, 125.
Kaighen, M. & Williams, R. T. (1961) J. Med. l'harm. Chem. 3, 25
Kaihara, M. & Price, J. M. (1965) J. Biol. Chem. , 240, 454.
• Kamil, I.A. , Smith, J. N. & Williams, R. T. (1952) Biochem. J. 50, 235.
Katchalsi, E. , Silman, I. & Goldman, R. (1972) Advan. Enzymol. 34, 445 - 536.
Kay, H.D. &Raper, H.S. (1922) Biochem. J. 16, 465.
Kreibich, G. , Debey, P. & Sabatini, D. D. (1973) J. Cell Biol. 58, 436 - 462.
Laidler, K. J. & Bunting, P. S. (1973) The Chemical Kinetics of Enzyme
Action pp. 382 - 412, Clarendon Press, Oxford.
Lan, S. J. , El-Hawey, A.M. , Dean, A. V. & Schreiber, E. C. (1975)
Drug Metab. and Disposition 3, 171 - 179.
Lethco, E. J. & Brouwer, E.A. (1966) J. Agr. Food. Chem. 14, 532 - 535.
Levine, W. G. , Singer, R.W. (1972) J. Pharmacol. Exp. Ther. 183, 411 - 419.
Luzzati, V. (1968) Biological Membranes, Physical Fact and Function.
(Chapman , D. , ed.), p 71, Academic Press, N.Y.
Mackenzie, C. G. & Frisell, W.R. (1958) J. Biol. Chem. 232, 417.
Marshall, E. K. Jr. (1954) J. Biol. Chem. 211, 499.
Mazur, A. & Shorr, E. (1942) J. Biol. Chem. 144, 283.
McChesney, E.W. & Hoppe, J. 0. (1954) Arch. int. pharmacology, 99, 127.
Melmon, K. L. & Morrelli, H. F. (1972) Clinical Pharmacology, Basic Principles
in Therapeutics, p. 41. Macmillan, New York.

185
Millburn, P. (1974) Biochem. Soc. Trans. 2, 126 - 130.
Miriam, S. R. , Wolf, J. T. & Sherwin, C. P. (1927 a) J. Biol. Chem.
71, 249 - 253.
Miriam, S. R. , Wolf, J. T. & Sherwin, C. P. (1927 b) J. Biol. Chem.
71, 695 - 698.
Mitoma, C., Posner, H. S. , Reitz, A. C. & Udenf riend, S. (1956) Arch.
Biochem. and Biophys. 61 431.
Moldave, K. & Meister, A. (1957) J. Biol. Chem. 229, 463 - 476.
Muenzen, J. B. , Cerecedo, L. R. & Sherwin, C. P. (1926) J. Biol. Chem.
67, 469.
Mulder, G.J. (1970) Biochem. J. 117, 319.
Neuberger, A. , Rimington, C. & Wilson,J. M. G. (1947) Biochem. J. 41,
438 - 449.
Nilsson, R., Pettersson, E., Danner, G. (1971) FEBS Lett. 15, 85 - 87.
Nys, G. G. & Rekker, R. F. (1973) Chimie Therapeutique No. 5. p. 521 - 535.
Oakley, S.E. & Seakins, J.W. T. (1971) Biochem. J. 121, 81 - 82.
Perrier, D. & Gibaldi, M. (1974) J. Pharmacol. Exp. Ther. 191, 17 -24.
Perrier, D. & Gibaldi, M. (1972) J. Clin. Pharmacol. 12, 449 - 451.
Pinto, J.D., Camien, M. N. & Dun, M.S. (1965) J. Biol. Chem. 240,
2148.
Populaire, P., Terlain, B. , Pascal, S., Lebreton, G. & Deconvelaere, B. (1969)
Arzneim-Forsch, 19, 1214 - 1221.
Power, F. W. (1936) Proc. Soc. Exp. Biol. Med. 33, 598 - 600.
Quick, A.J. (1932 a) J. Biol. Chem. 96, 73.
Quick, A. J. (1932 b) J. Biol. Chem. 96, 83 .
Reid, J. C. & Weaver, J. C. (1951) Cancer Res. 11, 188.

186
Reif, V. D. & Sinsheimer, J. E. (1975) Drug. Meta. & Disposition, 3, 15 - 25.
Reif, V. D. , Sinsheimer, J. E. , Ward, J. C. & Schteingart, D. E. (1974)
J. Pharmaceutical Sci. 63 1730 - 1736.
Rendon, A. & Waksman, A. (1971) Biochem. Biophys. Res. Commun, 42,
745 - 749.
Robinson, D. Smith, J. N. & Williams, R. T. (1955) Biochem. J. 59, 153.
Robinson, D. & Williams R.T. (1955) Biochem. J. 59, 159.
Robinson, D. & Williams, R. T. (1957) Biochem. J. 66, 46 p.
Robinson, D. & Williams, R.T. (1958) Biochem. J. 68, 23 p.
Rodbell, M. (1966) J. Biol. Chem. 241, 130.
Rothstein, M. & Greenberg, D. M. (1957) Archs. Biochem. Biophys. 68, 206.
Rubin, A. , Warrick, P. , Wolen, R. L. , Chernish, S. M. , Ridolfo, A. S. &
Gruber, C. M. Jr. (1972) J. Pharmacol. Exp. Ther. 183, 449 - 457.
Sabotka, H. (1937) Physiological Chemistry of the Bile pp. 22 - 23, Balliere,
Tindall & Cox, London.
Salkowski, E. (1884) Chem. Ber. 17, 3010 - 3012.
Salkowski, E. & Salkowski, H. (1879) Chem. Ber. 12, 653.
Schachter, D. & Taggart, J. V. (1953) J. Biol. Chem. 203, 925.
Schachter, D. & Taggart, J.V. (1954) J. Biol. Chem. 208, 263 - 275.
Schayer, R.W. (1956) Brit. J. Pharmacol. 11, 472.
Shand, D. G. , Nuckolls, E. M. , Oates, J. A. (1970) Clin. Pharmacol. Ther.
11, 112 - 120.
Sherwin, C.P. (1917) J. Biol. Chem. 31, 307 - 310.
Sherwin, C.P. (1918) J. Biol. Chem. 36, 309 - 318.
Sieberg, E. & Harloff, E. (1919) Z. Physiol. Chem. 108, 195. p. 58.
Singer, S. J. (1971) Structure and Function of Biological Membranes (Rothfield,
L. 1. ed. ) p. 145, Academic Press, N. Y.

187
Smith, J. N. (1964) In : Comparative Biochemistry, 6. Eds. Florkin, M. and
Mason, H.S., Academic Press, N.Y. and Lond.
Smith,J. N. (1968) In : Advances in Comparative Physiology and Biochemistry
3, Ed. Lowenstein ; Academic Press, N. Y. and Lond.
Smith, R. L. (1974) Ann. Nutr. Alim. 28, 335 - 349.
Smith, R. L. & Williams, R. T. (1966) in "Glucuronic Acid", (Dutton, G. J. , Ed).
pp. 457 - 491 Academic Press, New York.
Smyth, R. D. , Polk, A. , Diamond, J. , Burns, B. J. & Herczeg, T. (1974)
Xenobiotic 4, 383 - 391.
Snyder, S. H. , Axelrod, J. & Bauer, H. J. (1964) J. Pharmacol. Exp. Ther. 144,
373 - 379.
Stekol, J.A. (1936) J. Biol. Chem. 113, 675.
Stier, A. & Sackman, E. (1973) Biochim. Biophys, Acta. 311, 400 - 408.
Teague, R. S. (1954) Adv. Carbohyd. Chem. , 9, 185.
Thierfelder, H. & Sherwin, C. P. (1914) Chem. Ber. 47, 2630 - 2634.
Thierfelder, H. & Sherwin, C. P. (1915) Hoppe-Seyler s Z. Physiol. Chem. 94,
1 - 9.
Tishler, S. L. & Goldman, P. (1970) Biochem. Pharmacol. 19, 143 - 150.
Uehleke, H., Helimer, K. H. & Tabarelli-Poplawski. S. , (1973) Naunyn-
Schmiedeberg's Arch. Pharmacol. 279, 39 - 52.
Van Dyke, R.A. & Wood, C. L. (1975) Drug. Metab. and Disposition. 3, 51 - 57.
Vasilin, (1909) Mitt. Landw. Inst. Univ. Breslau. 4, 703.
Wade, D. N. , Desbiens, N. , Strewler, G. J. Jr. , Berndt, W. O. & Mudge, G. H.
(1971) J. Pharmacol. Exp. Ther. 178, 173.
Waksman, A. & Rendon, A. (1971) Biochem. Biophys, Res. Commun. 42,
1214 - 1219.

188
Waksman, A. & Rendon, A. (1974) Biochimie, 56, 907 - 924.
Wallcane, L. , Gingell, R. & Bronczyk, S. (1973) Toxicol Appl. Pharmacol.
25, 472.
West, J. R. , Smith, H.W. & Chasis, H. (1948) J. Pediat. 32, 10.
Williams, R T. (1938) Biochem. J. 32, 878, pp. 69, 72, 82, 94, 104, 105,
136, 149, 250.
Williams, R. T. (1959) Detoxication Mechanisms, 2nd Ed. Chapman & Hall Ltd.
Williams, R. T. (1967) Advancement of Sci. 23, 531 - 533.
Williams, R. T. (1969) Pure Appl. Chem, 18, 129.
Williams, R. T. (1974) Biochem. Soc. Trans. 2, 13 - 377.





























