Embed Size (px)
Citation preview

Ethology 101, 51-66 (1995) 0 1995 Blackwell Wissenschafts-Verlag, Berlin ISSN 0179-1613
Institut f u r Verhaltensbiologie, Freie Universitat Berlin, Berlin
Factors Affecting Acoustic Variation in Barbary-macaque (Macaca sylvanus) Disturbance Calls
JULIA FISCHER, KURT HAMMERSCHMIDT & DIETMAR TODT
FISCHER, J., HAMMERSCHMIDT, K. & TODT, D. 1995: Factors affecting acoustic variation in Barbary- macaque (Macucu sylvunns) disturbance calls. Ethology 101, 51-66.
Abstract
Semi-free-ranging Barbary macaques (Mucucu sylvunus) were observed to utter distinctive calls after disturbances in the surroundings (e.g. presence of a predator, occurrence of some unusual phenomenon). These calls differed from calls given in other contexts. Most of these calls were uttered in a serial manner, some of which lasted as long as the corresponding context. The aim of this study was to determine the object-related specificity of calls and variation of acoustic parameters within a call series. The analysis revealed that acoustic features of calls varied among contexts. Within one apparently homogeneous context, both temporal and frequency characteristics shifted gradually. Furthermore, the disturbance calls clearly differed between individuals. To examine the disturbance calls’ meaning, playback experiments were conducted in which alarm calls and disturbance calls were presented. After playback of an alarm call, subjects typically showed an escape response, whereas, in response to disturbance calls, they most often scanned the surroundings. Juvenile animals generally showed stronger responses than adults.
Corresponding author: Julia FISCHER, Institut fur Verhaltensbiologie, Freie Universitat Berlin, Haderslebener Strasse 9, D-12163 Berlin, Germany.
Introduction
An important aim in bio-acoustic research is to determine factors that govern acoustic variation in animal signals. Various studies have been conducted to identify the influence of numerous factors on signal variation, including devel- opmental state (ELOWSON et al. 1992; HULTSCH 1993), caller identity (SNOWDON & CLEVELAND 1980; CHAPMAN & WEARY 1990), social context (GREEN 1975; GOUZOULES et al. 1984; SCHRADER & TODT 1993), emotional state (SCHERER & KAPPAS 1988;JURGENS 1992), and articulation (HAUSER 1991; HAUSER et a]. 1993). Traditional analyses of animal communication have emphasised the relationship between signal morphology and the state of the sender (e.g. DARWIN 1872; MORTON 1982). Since STRUHSAKER’S (1967) report on vervet-monkey alarm calls, however, many studies have suggested that, in some cases, certain species are able
U.S. Copyright Clearance Center Code Statement: 0179-1613/95/~011-0051$11.00/0

52 J . FISCHER, K. HAMMERSCHMIDT & D. TODT
to encode and convey information about external events, for example, the type of predator (STRUHSAKER 1967; SEYFARTH et al. 1980; MACEDONIA 1990). In address- ing questions of signal meaning, primates and sciurid rodents have played a major role (OWINGS & VIRGINIA 1978; LEGER et a]. 1979; SEYFARTH et al. 1980; CHENEY & SEYFARTH 1990; PEREIRA & MACEDONIA 1991).
Studies addressing questions of signal meaning have dealt mainly with alarm calls because these have several properties which make them especially suitable for analysis. In general, the eliciting stimulus - the referent - is often readily identified. In our usage of the term ‘referent’, we are following the restricted definition given by SEYFARTH et al. (1980) and MARLER et al. (1992), reserving the term for objects or events external to the signaller. Another property is that animals typically show obvious responses to the appearance of a predator (review: MACEDONIA & EVANS 1993).
Most studies have so far focused on either signal variation with context or with internal state (but see HAUSER 1991). This study attempts to integrate the analysis of signal variation within one apparently homogenous context and com- pare it to the variation among different contexts. The third factor considered is the influence of individual characteristics, as these are reported to cause appreciable variation in signal patterns (HAMMERSCHMIDT & TODT 1995).
The set of calls used in this analysis are Barbary macaques’ vocalizations that were given to objects or events in their surroundings (presence of a predator or occurrence of some unusual phenomenon). Some of these calls actually elicited escape reactions in conspecifics, whereas the majority caused group members to scan the surroundings. However, to the human ear, there was no difference to be detected between calls that elicited flight reactions and those that did not. PETTER & CHARLES-DOMINIQUE (1979) termed all vocalizations that were given as a reaction to disturbances in the surroundings ‘alarm calls’. We suggested naming calls that did not elicit an escape reaction, but nevertheless seemed to call monkeys’ attention, ‘disturbance calls’. We reserved the term ‘alarm call’ for calls that clearly elicited escape responses.
Preceding observations had shown that the Barbary macaques in Rocamadour react to an observer’s sudden approach towards their sleeping trees at night with protracted call series. These vocalizations are referred to as ‘nocturnal disturbance calls’ (NDCs). Firstly, the nocturnal disturbance calls were compared to calls given to other objects in the external world to elucidate their potential referentiality. Secondly, the N D C s were examined for within-context variation. This extremely stable situation provided a good chance of investigating changes within the call series. O n e major point of interest in the analysis was the identification of acoustic variables that shift with time and, thus, may be correlated to a change in some internal variable (TODT 1995). As conditions were very similar for the different individuals, the data set also provided a good opportunity to compare the individual characteristics of the acoustic signals.
We conclude with a report from playback experiments that were designed to examine both the context independence of Barbary- macaque alarm calls and the proximate function of nocturnal disturbance calls. Context independence refers

Acoustic Variation in Barbary-macaque Disturbance Calls 53
to the hypothesis that referential signals alone should be sufficient to allow receivers to select an appropriate response (SEYFARTH et al. 1980; MACEDONIA & EVANS 1993), i. e. alarm calls should also elicit an escape reaction in group members in the absence of a predator. Another aim was to evaluate the nocturnal disturbance calls’ alarm-eliciting potential.
The design of playback experiments is generally restricted by the fact that primates tend to habituate quickly to the setup (BYRNE 1982). Therefore, only a limited number of trials can be conducted. We chose to use only very few calls and perform several trials per call exemplar. This design enabled us to estimate the variance of subjects’ responses. Additionally, it controlled for influences of other design variables such as, for instance, the direction of the loudspeaker or the playback site.
Methods
Study Animals and Study Site
Observations were conducted in Aug., Sep., and Dec. 1992, and in May, 1993, in a semi-free confinement (10 ha) in Rocamadour, France. The enclosure is a visitor park where monkeys range freely while visitors are restricted to a path. The monkeys are fed with monkey chow provided in feeder huts, and with apples, grain, and seeds which are spread throughout the park. During observations, the population consisted of approximately 240 individuals. The population is split into two subgroups of 50 and 190 animals, respectively. Individuals are tattooed with an individual code on the inside of the thigh. For details on park management and park size, see SMALL (1990), and DE TURCKHEIM & MERZ (1984). A review of studies conducted on Barbary macaques in Rocamadour is given in TODT et al. (1992). Additionally, wild Barbary macaques were observed in Morocco. Several groups were tracked in the ‘Gorges d’Ouzoud’ and in the ‘Gorges de l’Oued el Abid’ in the Southern region of the Middle Atlas as well as in the Cedar Forest near Ain Kahla in the Central region of the Middle Atlas (for a more detailed description of observation sites, see TODT et al. 1995).
Data Sampling
Vocalizations were recorded continuously with a Sony WM DC6 and a Sennheiser KN3/ME80 or ME88 microphone with a Sennheiser windscreen. Both focal-animal sampling techniques and ad libitum sampling were applied. When the focal animal vocalized, a detailed description of the situation before and after the calling, including activity of the caller and possible interactants, was spoken onto the tape recorder.
Vocal Activity after Nocturnal Disturbance
Observations at the beginning of our study had shown that monkeys that were already on their sleeping trees reacted to the appearance of an observer at night with protracted call series. Usually, the monkeys spent the night quietly (ANSORGE et al. 1992; HAMMERSCHMIDT et al. 1994 b). To provoke such serial calling, monkeys were repeatedly approached at night. Disturbances were conducted over 18 nights. Disturbances consisted of a noisy approach towards a sleeping tree followed by a quiet interval, and ended with either a further approach or retreat. Most often, a number of animals started to vocalize after they had noticed the observer. However, for analysis, only recordings were chosen during which only one animal was vocalizing at a time. Because of poor light conditions, it was not possible to individually identify the animals on the trees. For subsequent disturbances in the same night, different sleeping areas were chosen. Age was determined by size class, while sex could be identified from shape in adult animals. Only once could the caller’s identity be determined the following morning. Identification the next morning was impaired because monkeys tended to change their

54 J. FISCHER, K. HAMMERSCHMIDT & D. TODT
sleeping place after a disturbance. In addition, 2-3 sleeping clusters were frequently observed in the same tree (for details on the Barbary macaques’ sleeping habits, see ANSORGE et al. 1992).
Analysis of Call Sequences
For this analysis, a nocturnal call sequence is operationally defined as the succession of all calls occurring after an approach until: a. The subsequent approach; or b. The retreat. For a detailed analysis, 16 sequences from nine different subjects were selected. Six of the subjects were juveniles of different age classes, and three were adult females. Three subjects provided three sequences each (first approach and two following approaches), one subject provided two sequences, and the remaining five sequences were recorded from five different subjects. The mean number of calls in first-approach sequences was 140.1 (minimum 99, maximum 326), the mean number of calls in repeated approaches was 147.8 (minimum 64, maximum 244). Although it cannot be completely ruled out that different sequences were recorded from the same subject, this possibility was extremely unlikely considering the great number of animals in the park (n = 240).
Because sequences varied both in length and number of calls per sequence, it was necessary to find a measure which allowed a comparison between sequences. Therefore, for each sequence, 20 calls from each of the three positions ‘start’, ‘middle’ and ‘end’ were chosen for analysis (n = 540 calls). ‘Middle’ refers to the median of the number of calls of a given sequence. For a comparison among subjects (individual characteristics), only the first sequence per subject was chosen. Data from repeated approaches were used to check for intra-individual variation in animals’ responses.
Analysis of Vocal Patterns
For visual inspection, vocalisations were transferred to a spectrum analyser (NSC UA 500A), projected onto a 35mm positive film through an oscilloscope (Phillips PM 320), and examined using RTS (Engineering Design). Selected calls were digitized at a sample frequency of 37.5 kHz. Subsequently, a fast Fourier transformation (normal 1024-pt FFT) was conducted with Signal sound analysis system (time resolution: 5 ms; frequency range: 15 kHz; frequency resolution: 36.9 Hz). For evaluation of those parameters that characterize the structure of a vocal pattern, a statistic-threshold filter was applied to emphasise dominant frequency bands. Finally, on the basis of the filtered and unfiltered frequency-time spectra, 60 parameters were calculated to characterize the call in terms of its frequency and time dynamics. The term ‘dominant frequency band’ is applied to frequencies with an amplitude exceeding a statistical threshold in a consecutive number of cells. Dominant frequency bands do not necessarily correspond to the fundamental, second and third harmonic. The following parameters were calculated: 1. Temporal parameters calculated from filtered and unfiltered spectra: call duration, duration from start until maximum and minimum peak amplitude; 2. Frequency parameters calculated from filtered frequency-time spectra: start, end, maximum, minimum, and mean frequency of first, second, and third dominant-frequency band (dfb), mean width, modulation, and maximum modulation of first and second dfb; difference between first and 2nd dfb at start and end of call, maximum, minimum, and mean difference between first and second dfb; mean, maximum, and minimum number of dfbs; frequency of peak amplitude at start, end, and at maximum amplitude; minimum and mean amplitude, start, end, maximum and mean frequency range; 3. Frequency parameters calculated from power spectra: mean, start, and end of Ist, 2nd, and 3rd peak, mean relative amplitude, standard deviation of mean relative amplitude, maximum frequency, start, end and width of frequency range, density, percentage of high-frequency amplitudes, intensity, and maximum intensity. Filtering and parameter calculations were made using a software program (LMA 3.0) developed by K. HAM- MERSCHMIDT (HAMMERSCHMIDT & TODT 1995).
A powerful tool for analysing differences between contexts is the discriminant analysis that compares multivariate patterns resulting from any interaction of variables. The discriminant-function analysis provides a classification procedure that assigns each call either to its appropriate group (correct assignment) or to another group (incorrect assignment; GOUZOULES & GOUZOULES 1989; HAUSER 1991). It also calculates the rank order in which parameters contribute to a correct assignment (identification of predictor variables). X*-analyses revealed whether or not the classification of calls was significantly greater than chance. A Kruskal-Wallis analysis of variance was used to check for univariate context-related differences. For each individual, one data point was entered in the analysis. A Friedman test was used to examine differences between positions in a sequence. After rejecting the
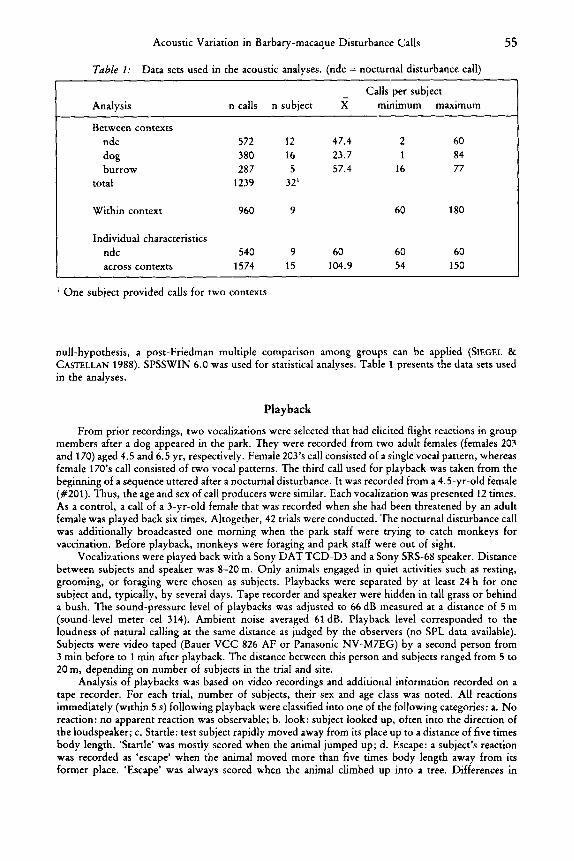
Acoustic Variation in Barbaymacaque Disturbance Calls 55
Table 1: Data sets used in the acoustic analyses. (ndc = nocturnal disturbance call)
- Calls per subject Analysis n calls n subject X minimum maximum
Between contexts ndc 572 12 47.4 2 60 dog 380 16 23.7 1 84 burrow 287 5 57.4 16 77
total 1239 32’
Within context 960 9 60 180
Individual characteristics ndc 540 9 60 60 60 across contexts 1574 15 104.9 54 150
~~~~~~~~ ~ ~
’ One subject provided calls for two contexts
null-hypothesis, a post-Friedman multiple comparison among groups can be applied (SIEGEL & CASTELLAN 1988). SPSSWIN 6.0 was used for statistical analyses. Table 1 presents the data sets used in the analyses.
Playback
From prior recordings, two vocalizations were selected that had elicited flight reactions in group members after a dog appeared in the park. They were recorded from two adult females (females 20.1 and 170) aged 4.5 and 6.5 yr, respectively. Female 203’s call consisted of a single vocal pattern, whereas female 170’s call consisted of two vocal patterns. The third call used for playback was taken from the beginning of a sequence uttered after a nocturnal disturbance. It was recorded from a 4.5-yr-old female (#201). Thus, the age and sex of call producers were similar. Each vocalization was presented 12 times. As a control, a call of a 3-yr-old female that was recorded when she had been threatened by an adult female was played back six times. Altogether, 42 trials were conducted. The nocturnal disturbance call was additionally broadcasted one morning when the park staff were trying to catch monkeys for vaccination. Before playback, monkeys were foraging and park staff were out of sight.
Vocalizations were played back with a Sony DAT TCD-D3 and a Sony SRS-68 speaker. Distance between subjects and speaker was 8-20 m. Only animals engaged in quiet activities such as resting, grooming, or foraging were chosen as subjects. Playbacks were separated by at least 24 h for one subject and, typically, by several days. Tape recorder and speaker were hidden in tall grass or behind a bush. The sound-pressure level of playbacks was adjusted to 66 dB measured at a distance of 5 m (sound-level meter cel 314). Ambient noise averaged 61 dB. Playback level corresponded to the loudness of natural calling at the same distance as judged by the observers (no SPL data available). Subjects were video taped (Bauer VCC 826 AF or Panasonic NV-M7EG) by a second person from 3 min before to 1 min after playback. The distance between this person and subjects ranged from 5 to 20 m, depending on number of subjects in the trial and site.
Analysis of playbacks was based on video recordings and additional information recorded on a tape recorder. For each trial, number of subjects, their sex and age class was noted. All reactions immediately (within 5 s) following playback were classified into one of the following categories: a. No reaction: no apparent reaction was observable; b. look: subject looked up, often into the direction of the loudspeaker; c. Startle: test subject rapidly moved away from its place up to a distance of five times body length. ‘Startle’ was mostly scored when the animal jumped up; d. Escape: a subject’s reaction was recorded as ‘escape’ when the animal moved more than five times body length away from its former place. ‘Escape’ was always scored when the animal climbed up into a tree. Differences in

56 J . FISCHER, K. HAMMERSCHMIDT & D. TODT
responses between adults and juveniles and among calls were tested using the Craddock-Flood X’-test of contingency (BORTZ et al. 1990).
Results
Several events occurring outside the social group elicited vocalizations in the Rocamadour monkey population. Such situations were: 1 . A dog close to the monkeys; 2. Park staff trying to catch monkeys for vaccination; 3. A fox in the park; 4. Facing a new burrow; 5. Sudden appearance of a human in the park; and 6. The observer approaching the monkeys’ sleeping trees at night (see above). Some of these vocalizations elicited flight reactions in group members. However, the greatest number of vocalizations was recorded from serially calling individuals while other animals did not show escape responses. Those animals closest to the eliciting stimulus were calling repeatedly while others did not show any signs of moving away or up in a tree. One or few monkeys (most often juvenile or subadult males) typically sat in a tree, stared at the stimulus and called repeatedly while the other animals stayed on the ground and kept, for instance, foraging. During daytime, such situations were observed 32 times. Three of the contexts provided a sufficient number of vocalizations recorded from a sufficient number of animals to be included in the analysis. These were: different dogs in or outside the park (‘dog’), facing a burrow (‘burrow’) and after a nocturnal disturbance (‘NDC’). ‘Burrow’ describes an odd situation where several monkeys were sitting in trees outside the enclosure in front of a burrow’s opening at a very steep slope. They stared into the opening and repeatedly vocalized. As the observer was sitting on top of the opening, it was not possible to identify the actual cause of the monkeys’ reaction.
From their acoustic properties, vocalizations that were given after a dis- turbance in the surroundings resemble the Barbary macaques’ alarm calls but differ from calls given in other contexts. In terms of the actual sound, both disturbance and alarm calls may be described as barks. Most vocalizations start with a harmonic, more-or-less high-pitched segment followed by a harsh sound. An analysis of the entire vocal repertoire indicated significant differences between calls given to the objects or events listed above and vocalizations given during other contexts (HAMMERSCHMIDT et al. 1994 a). Generally, disturbance calls are given in sequences, the length of which seemed to be determined by the stimulus’ presence. Alarm calls, on the other hand, were uttered in short bouts containing only few vocal patterns. They either elicited flight reactions or cryptic behaviour in group members. These findings are supported by observations made in the field in Morocco. Approaches by the observers in Morocco most often led to calls causing immediate flight reactions. In two incidents, however, animals stayed at a relatively safe distance while one animal staring at the observers kept vocalizing.
Between-context Variation Fig. 1 presents spectrograms of calls which were given in the three contexts
selected for analysis. For each context, three calls recorded from three different

Acoustic Variation in Barbary-macaque Disturbance Calls 57
1 s time
Fig. I: Spectrograms of calls given in the three contexts examined: a. Nocturnal disturbance; b. Calls given to a dog; c. Calls given to the burrow. Three exemplars by three subjects per context
individuals are shown. A total of 1239 calls from 32 different individuals were submitted to analysis (see Table I). Of 1239 calls classified, the discriminant analysis’s classification procedure yielded a correct assignment of vocal patterns to the three contexts of 87 %. This is significantly more than what would be expected from random assignment (33.3 %; x2 = 258.8, df = 2, p < 0.001). The classification results for each context are presented in Table 2a. Calls given after a nocturnal disturbance and to ‘burrow’ show a relatively higher correct assignment than calls given to ‘dog’.
As noted earlier, the discriminant analysis also provides information on which variables contribute most to the discrimination of groups. The five most decisive variables are listed in Table 3a. Group means calculated from individual averages, standard errors, x2 and probability levels are also given in Table 3a. The univariate analyses show that not all parameters separate all groups equally well.

58
Table 2:
J. FISCHER, K. HAMMERSCHMIDT & D. TODT
Classification results of the discriminant function analysis for (a) between-context and (b) within-context variation (ndc = nocturnal disturbance call)
Predicted group membership a ndc (YO) Dog (YO) Burrow (YO) n
Actual group ndc 92.1 3.9 4.0 569 dog 7.1 77.1 15.8 380
burrow 3.1 6.6 90.2 287 (average correct assignment: 87 Yo ; random assignment: 33.3 Yo)
b Predicted group membership
Start (YO) Middle (YO) End (YO) n
Actual group start 58.0 19.4 22.6 320 middle 20.1 47.0 32.9 320
end 10.3 25.7 63.9 320 (average correct assignment: 56.32 YO; random assignment: 33.3 Yo)
Within-context Variation Changes within a call series were studied in those call series that were uttered
after animals were approached by an observer at night. An example is presented in Fig. 2. The average length of the 16 sequences analysed was about 6 min (246.1 s, SD 68.5 s). Mean call rate (measured across 20 calls after start and after 1 min, respectively) dropped from 1.23 (SD 0.56) calls/s after start to 0.66 (SD 0.27) calls/s after 1 min. Call length, interval length (time between onsets of successive calls), and call density (call lengthlinterval length) were chosen for measuring temporal variation along a call series. Overall mean call duration was 112 ms (SD 42 ms); overall mean interval length was 1.9 s (SD 2.2 s). Call duration decreased, interval length became longer, and call density decreased (Fig. 3). A Friedman test revealed that temporal parameters differed significantly (x2 for call duration 10.8 (p < 0.01); interval length 6.89 (p < 0.05), call density 14.89 (p < 0.001); n = 9; df = 2). A post-hoc multiple comparison showed that ‘start’ and ‘end’ differed in all cases significantly. Call density also exhibited significant differences between ‘start’ and ‘middle’ (p < 0.05).
A discriminant analysis of the frequency-time spectra with regard to the different positions ‘start’, ‘middle’, and ‘end’ in a sequence yielded a correct assignment of 56.32 % (Table 2b). Mean values and SD, as well as x2 and probability levels for the five most decisive variables are given in Table 3b. A multiple comparison among groups revealed that all parameters but one (mean relative amplitude) differed significantly between ‘start’ and ‘end’. Finally, the repeated approaches were examined for effect of order by comparing mean values of the ‘start’ position among first, second, and third approach separately for each

Acoustic Variation in Barbary-macaque Disturbance Calls 59
Univariate statistics (X, SE) and xz for the five most decisive acoustic variables by (a) context -
Tubfe3: and (b) position
a Context Rank Variable ndc Dog Burrow xz p
1 mean 2nd dfb (Hz) 2183 1352 1766 18.2 <0.001
2 call duration (ms) 136 156 133 2.2 ns
3 mean frequency range (Hz) 2796 2043 1729 12.2 <0.01
4 diff. 1st to 2nd dfb (Hz) 601 383 314 13.0 cO.01
5 mean width 2nd dfb (Hz) 125 104 93 7.1 <0.05
SE 101 67 268
SE 10 9 20
SE 183 136 194
SE 13.6 8.8 7
SE 7 7 9 (Kruskal-Wallis test; n = 33; df = 2)
b Position Rank Variable Start Middle End x3 p
1 relative amplit. 58.7 55.15 44.41 2.7 ns
2 max. modul. 1st dfb (Hz) 442.9 415.1 328.6 8.7 C0.05
SE 9.35 5.43 4.4
SE 54.5 57.0 39.3
3 call duration (ms) 138.4 125.7 113.1 10.8 <0.01 SE 12.5 11.5 10.8
4 end f peak 3 (Hz) 4843.1 4045.1 3680.8 6.9 c0.05
5 mean 1st dfb (Hz) 1667.2 1605.6 1499.7 8.7 c0 .05 SE 658.3 343.4 202.3
SE 24.6 22.3 20.2 (Friedman test; n = 9; df = 2; dfb: dominant frequency band, f: fre-
quency)
20 30 S
Fig. 2: Spectrogram of a nocturnal call series (first 35 s)

60
- 2 . 5 . E c 2 8 154
0 5
0.5
J . FISCHER, K. HAMMERSCHM~DT & D. TODT
Call duration
153 1 3.5
31
O L
Interval length
i 5 4 : 5 O W
I
Fig. 3: Changes in temporal parameters (call duration, interval length, and call density) within nocturnal call sequences. X and first and third quartile for each of the three positions start, middle,
and end in a sequence are shown (n = 960 calls)
individual. There was no detectable effect of order, such as a constant decline or rise.
Individual Characteristics
Although calls varied within individuals, there was a pronounced inter- individual variation (Fig. 1). The classification procedure for the nine individuals vocalizing after a nocturnal disturbance yielded an average correct assignment by the discriminant analysis of 89.1 Yo; 11.1 % would be expected from random assignment. The most decisive parameters for discrimination between individuals were mean first dominant-frequency band, call duration, and standard deviation of mean relative amplitude. A second analysis determined the assignment to individuals across all contexts examined. A total of 15 animals who were indi- vidually identified were entered into the analysis. The average correct assignment was still as high as 86 Yo; 6.7 % would have been expected from random assign- ment.
Playback
The results of playbacks and statistical tests are presented in Table 4. Only the alarm calls elicited escape reactions, and only juveniles were observed to respond to the disturbance call with a startle response. The control stimulus elicited either 'no reaction' o r 'look'. Juveniles generally showed stronger responses than adults after playback of an alarm call or the disturbance call, but exhibited no difference in response to the two alarm calls. Adults, by contrast, were more prone to run away or up a tree after playback of 'double alarm' than after playback of 'single alarm'. Analysis of video recordings showed that adult animals typically looked up after the first call and ran away after the second. After playback of 'single alarm', adults most often scanned the surroundings.
Playback of 'nocturnal disturbance call' frequently elicited 'no apparent reaction' and no visual orientings in animals. There was no significant difference

Acoustic Variation in Barbary-macaque Disturbance Calls 61
Table 4: a. Total number of responses per call type and age-class; b. Age-related differences in responses to playback of four call types; c. Differences in subjects’ reactions to four call types separately
for both age-classes
a Responses Double alarm Single alarm Disturbance call Control
adults juveniles adults juveniles adults juveniles adults juveniles ~~ ~~
- - No reaction - 4 23 15 12 6 Look 12 4 25 9 19 9 4 4 Startle 3 4 6 10 5 - -
Escape 5 29 10 16 - - - -
-
b Call types Double alarm Single alarm Disturbance call Control
Juveniles x z = 17.5 xz = 12.8 x2 = 8.1 xz = .65 vs. d f = 2 d f = 3 df = 2 df = t adults p < 0.01 p < 0.01 p < 0.05 ns
C Test pairs Double alarm/ Single alarm / Single alarm/ Disturbance
single alarm disturbance call control call /control
x2 = 8.2 x2 = 32.4 x2 = 28.9 x 2 = 1.9
p < 0.05 p < 0.001 p < 0.001 ns x2 = 0.17 x2 = 30.1 x2 = 27.6 x 2 = 1.9
ns p < 0.001 p < 0.001 ns
Adults d f = 3 df = 3 d f = 3 df = 2
Juveniles df = 1 d f = 3 df = 3 d f = 1
between adults’ and juveniles’ responses after both a nocturnal disturbance call and the control stimulus.
As mentioned above, the nocturnal disturbance call was additionally broad- casted once while the park staff was trying to catch monkeys for vaccination. The staff was out of sight from the playback site. Immediately after playback, all monkeys ran away or up a tree.
Discussion
Between- and within-context variation
After disturbances, Barbary macaques uttered distinctive calls that, in their acoustic features, varied with context. It might be the case that these calls designated different objects, since the different calls were not clearly related to different behavioural responses, such as, for instance, specific escape strategies, as in vervet monkeys (CHENEY & SEYFARTH 1990). Therefore, it is not quite clear which

62 J . FISCHER, K. HAMMERSCHMIDT & D. TODT
information is conveyed in these calls and how they are perceived. So far, it can not be ruled out completely that Barbary macaques’ disturbance calls do not point to different external referents. Instead, they might primarily express different levels of arousal.
The comparison of alarm and disturbance calls given to a dog suggests that differences related to object specific characteristics are greater than differences between alarm and disturbance calls given to the same object. Due to the small number of calls available for analysis, the results need to be interpreted cautiously. Significant features of ‘alarming’ remain to be determined.
The nocturnal call series consisted of a succession of calls of the same call type (‘homo-type series’; TODT 1986). Along the course of a nocturnal call series, a gradual decline in both temporal and frequency characteristics could be observed. The results suggest that these changes reflect a change in some internal variable exhibiting the influence of a tonic component (TODT 1995). We assume that the animals’ assessment of the situation altered with increasing time from stimulus onset. Presumably, they become less distressed with time. Interestingly enough, the same parameters that decreased in the nocturnal call series increased in Barbary- macaque infant-crying sequences (TODT et al. 1995). Infants who are prevented from returning to their mother by a male caretaker often emit long lasting cry sequences. SCHRADER & TODT (1993) obtained similar results in a study investigating context-related variation in Common-marmoset phee calls (Callithrix jacchrrs). In this species, median frequency, call duration, and amplitude increased with increasing separation from the animals’ companion.
The Barbary-macaque disturbance calls might be viewed as an example of ‘intermediate signalling’ (MARLER et al. 1992), as disturbance calls were found to exhibit variation with context and with time from stimulus onset. Those acoustic features that characterize ‘nocturnal disturbance calls’ did not change within a series. Eventually, playback studies are needed to investigate the significance of these call variations for communication. A number of studies investigating salient features of primate vocal signals have been conducted (e.g. SNOWDON & POLA 1978; MASATAKA 1983). MAY et al. (1989) and HOW et al. (1992), for instance, have shown that the peak position along the F, contour is the most decisive feature in the discrimination of Japanese macaque ‘coo’ calls.
Individual Characteristics
Individual recognition has been demonstrated in a number of species, e.g. Squirrel monkeys (Sairniri sciureus; SYMMES & BIBEN 1986), Vervet monkeys (CHENEY & SEYFARTH 1980), and Japanese macaques (PEREIRA 1986). HAM- MERSCHMIDT (1990) showed that Barbary-macaque mothers are able to identify their infant offspring by voice alone. His analysis of acoustic features of infant crying suggested that individual characteristics are expressed by a specific con- figuration of several acoustic parameters. That author suggested that an animal’s ‘signature’ (BEECHER 1982) probably has to be learned by its group members. Individual characteristics were also detected in Barbary-macaque females’ mating

Acoustic Variation in Barbary-macaque Disturbance Calls 63
calls (TODT et al. 1995). The outcome of this study supports the view of individual cues in Barbary macaques’ vocal patterns.
Acoustic Variables With regard to the acoustic analysis, this study supports the view (HAM-
MERSCHMIDT & TODT 1995) that a detailed examination of several acoustic para- meters is a crucial prerequisite for discriminating among groups. A previous analysis (HAMMERSCHMIDT et al. 1994a) showed that the degree of correct assign- ment depends on the number of parameters available. The number of parameters required for a reliable discrimination of statistical groups increases with the number of groups to be separated. At the same time, however, we observed that groups do not need to differ in all parameters. For example, calls given after a nocturnal disturbance and to ‘burrow’ differed clearly in mean frequency range, but were similar in call duration. This is also true for calls given to ‘dog’ and ‘burrow’, which differed in call duration, but were similar in difference between first and second dominant-frequency band.
Playback Experiments Interpretation of these playback experiments is restricted by the fact that
only few call exemplars were used (for a thorough discussion of playback designs and the problem of pseudoreplication, see KROODSMA 1989; SEARCY 1989; MCGREGOR 1992). In spite of this limitation, playback of ‘single’ and ‘double alarm’ showed that these alarm calls were independent of context, in other words, playback was sufficient to elicit an appropriate response in conspecifics, even in the absence of a predator. The outcome of these trials corresponds to results of playback studies conducted in other species, e.g. Vervet monkeys (SEYFARTH et al. 1980; CHENEY & SEYFARTH 1990) or Ringtailed lemurs, Lemur cuttu (MACEDONIA 1990).
A replication of playback of the same call, however, allows estimation of the variability in animals’ responses. Observations suggested that the outcome of a playback experiment not only depended on signal meaning but also on the receivers’ motivational state and their assessment of the situation. This view was supported by monkeys’ escape responses after playback of the nocturnal disturbance call while park staff were trying to catch monkeys for vaccination. This was the only time that monkeys responded to playback of a disturbance call with an escape response.
Juveniles generally responded more intensely than adult animals. Unlike adults, juveniles sometimes startled in response to playback of a nocturnal dis- turbance call. Juveniles might generalize stimuli, whereas adults exhibited more refined perceptual abilities. It also appeared as if adults were more experienced in judging a situation, and as if they were occasionally waiting for further cues, such as further calling or sight of the caller, a behaviour similar to that reported by SEYFARTH et al. (1980). Age-related differences in alarm calling and responses to alarms may result from observational learning, reinforcement, or some other social process (SEYFARTH & CHENEY 1986). This view was supported by HAUSER (1989),

64 J. FISCHER, K. HAMMERSCHMIDT & D. TODT
who hypothesized that perception of vocalizations depends on the age of subject. In Barbary macaques, auditory and social experience may also influence responses.
Concerning the function of both nocturnal and day-time disturbance calls, they might announce the presence of a potential hazard which is still at a ‘safe’ distance. This hypothesis could be tested, for instance, by broadcasting a series of calls and examining the checking rate before and after playback. One would expect elevated checking rates after playback of disturbance calls. It might also be the case that disturbance calls are addressed to the predator in order to announce that it has been detected (KLUMP & SHALTER 1984).
Acknowledgements We would like to thank Ellen MEW and Gilbert de TURCKHEIM for permission to conduct this
study at ‘La For& des Singes’, and Lucien AMBIEHL and Emanuelle LE GRELLE for support in the field. Thanks go to Viveka ANSORGE for useful comments on an earlier version of this manuscript, and to Henrike HULTSCH, Jorg BOHNER, Marc NAGUIB, and Lars SCHRADER for fruitful discussions. Stefan GEIS and Roger MUNDRY provided companionship and support throughout the study.
Literature Cited ANSORGE, V., HAMMERSCHMIDT, K., & TODT, D. 1992: Communal roosting and formation of sleeping
dusters in Barbary macaques (Mucucu syIvunns). Am. J . Primatol. 28, 271-280. BEECHER, M. 1982: Signature systems and kin recognition. Amer. Zool. 22, 477-490. BORTZ, J., LIENERT, G. A. & BOEHNKE, K. 1990: Verteilungsfreie Methoden in der Biostatistik.
BYRNE, R. W. 1982: Primate vocalizations: structural and functional approaches to understanding.
CHAPMAN, C. A. & WEARY, D. M. 1990: Variability in spider monkeys’ vocalizations may provide
CHENEY, D. L. & SEYFARTH, R. M. 1980: Vocal recognition in free-ranging vervet monkeys. Anim.
-- & - - 1990: How Monkeys See the World. Univ. of Chicago Press, Chicago. DARWIN, C. 1872: The Expressions of the Emotions in Man and Animals. Murray, London. ELOWSON, A. M., SNOWDON, C. T., & SWEET, C. J. 1992: Ontogeny of trill and J-call vocalizations
in the pygmy marmoset, Cebuella pygmueu. Anim. Behav. 43, 703-705. GOUZOULES, H. & GOUZOULES, S. 1989: Design features and developmental modification of pigtail
macaque, Mucucu nemesrrinu, agonistic screams. Anim. Behav. 37, 383-401. -- & -- & MARLER, P. 1984: Rhesus monkey (Mucucu rnulattu) screams: representational
signalling in the recruitment of agonistic aid. Anim. Behav. 32, 182-193. GREEN, S. 1975: Variation of vocal pattern with social situation in the Japanese monkey (Mucucu
fuscutu): a field study. In: Primate Behavior, Vol. 4 (ROSENBLUM, L. A., ed.). Acid. Press, New York. pp. 1-102.
HAMMERSCHMIDT, K. & TODT, D. 1995: Individual differences in vocalisations of young Barbary macaques (Mucucu sylvanus): a multi-parametric analysis to identify critical cues in acoustic signalling. Behaviour, in press.
-_ , ANSORGE, V., & FISCHER, J . 1994 a: Age-related variations in the vocal repertoire of Barbary macaques. In: Current Primatology, Vol. 2 (ROEDER, J . J., THIERRY, B., ANDERSON, J. R. & HERRENSCHMIDT, N., eds). Univ. Louis Pasteur, Strasbourg. pp. 287-295.
for social shelter. Am. J. Primatol. 32, 277-289.
(Cercopitbectrs uerbiops) vocalizations. J . Comp. Psychol. 103, 149-158.
89, 2-46.
Springer Verl., Berlin, New York, Heidelberg.
Behavior SO, 241-257.
basis for individual recognition. Am. J. Primatol. 22, 279-284.
Behav. 28, 362-367.
-_ _ _ , _ _ & TODT, D. 1994 b: Dusk calling in Barbary macaques (Mucucu sylvanus): demand
HAUSER, M. D. 1989: Ontogenetic changes in the comprehension and production of vervet monkey
-- 1991 : Sources of acoustic variation in rhesus macaque (Macucu mwlatru) vocalizations. Ethology

Acoustic Variation in Barbary-macaque Disturbance Calls 65
-- , EVANS, C. S., & MARLER, P. 1993: The role of articulation in the production of rhesus monkey, Mucuca mulattu, vocalizations. Anim. Behav. 45, 423-433.
HOPP, S. L., SINNO?T, J. M., OWREN, M. J., & PETERSEN, M. R. 1992: Differential sensitivity of Japanese macaques (Mucacu fuscatu) and humans (Homo sapiens) to peak position along a synthetic coo call continuum. J. Comp. Psychol. 106, 128-1
HULTSCH, H. 1993: Tracing the memory mechanisms in the song acqu on of nightingales. Nether]. J . Zool. 43, 155-171.
JURGENS, U. 1992: O n the neurobiology of vocal communication. In: Nonverbal Vocal Com- munication (PAPOUSEK, H., JURGENS, U. & PAPOUSEK, M., eds). Cambridge Univ. Press, Cambridge. pp. 31-42.
KLUMP, G. M. & SHALTER, M. D. 1984: Acoustic behaviour of birds and mammals in the predator context. I. Factors affecting the structure of alarm signals. 11. The functional significance and evolution of alarm signals. Z. Tierpsychol. 66, 189-226.
KROODSMA, D. 1989: Suggested experimental designs for song playbacks. Anim. Behav. 37, 600- 609.
LEGER, D. W., OWINGS, D. H., & BOAL, L. M. 1979: Contextual information and differential responses to alarm whistles in California ground squirrels. Z. Tierpsychol. 49, 142-1 55.
MACEDONIA, J. M. 1990: What is communicated in the antipredator calls of lemurs: evidence from playback experiments with ringtailed and ruffed lemurs. Ethology 86, 177-190.
-- & EVANS, C. S. 1993: Variation among mammalian alarm call systems and the problem of meaning in animal signals. Ethology 93, 177-197.
MARLFR, P., EVANS, C. S. & HAUSER, M. D. 1992: Animal signals: motivational, referential, or both? In: Nonverbal Vocal Communication (PAPOUSEK, H., JURGENS, U. & PAPOUSEK, M., eds). Cambridge Univ. Press, Cambridge. pp. 6 6 8 6 .
MASATAKA, N. 1983: Categorical responses to natural and synthesized alarm calls in Goeldis’s monkeys (Cullimico goeldiz). Primates 24, 40-51.
MAY, B., MOODY, D. B., & STEBBINS, W. C. 1989: Categorical perception of conspecific com- munication sounds by Japanese macaques, Mucucafuscatu. J. Acoust. SOC. Am. 85, 837-847.
MCGREGOR, P. K. 1992: Playback and Studies of Animal Communication. Plenum Press, New York. MORTON, E. S. 1982: Grading, discreteness, redundancy, and motivational-structural rules. In:
Acoustic Communication in Birds, Vol. 1 (KROODSMA, D. E. & MILLER, E. H., eds). Acad. Press, New York. pp. 183-212.
OWINGS, D. H. & VIRGINIA, R. A. 1978: Alarm calls of California ground squirrels (Spermophilus beecheyi). Z. Tierpsychol. 46, 58-70.
PEREIRA, M. E. 1986: Maternal recognition of juvenile offspring vocalizations in Japanese macaques. Anim. Behav. 34, 935-937.
-- & MACEDONIA, J . M. 1991: Ringtailed lemur anti-predator calls denote predator class, not response urgency. Anim. Behav. 41, 543-544.
PETTER, J. J . & CHARLES-DOMINIQUE, P. 1979: Vocal communication in prosimians. In: The Study of Prosimian Behavior (DOYLE, G. A. & MARTIN, R. D., eds). Acad. Press, New York. pp.
SCHERER, K. R. & KAPPAS, A. 1988: Primate vocal expression of affective states. In: Primate Vocal Communication (TODT, D., GOEDEKING, P. & SYMMES, D., eds). Springer Verl., Berlin, New York, Heidelberg. pp. 171-194.
SCHRADER, L. & TODT, D. 1993: Contact call parameters vary with social context in common marmosets, Callithrin j . jacchus. Anim. Behav. 46, 1 0 2 6 1 0 2 8 .
SEARCY, W. A. 1989: Pseudoreplication, external validity and the design of playback experiments. Anim. Behav. 38, 715-717.
SEYFARTH, R. M. & CHENEY, D. L. 1986: Vocal development in vervet monkeys. Anim. Behav. 34,
-- , CHENEY, D. L. & MARLER, P. 1980: Monkey responses to three different alarm calls: Evidence
SIEGEL, S. & CASTELLAN, N. J . 1988: Nonparametric Statistics for the Behavioral Sciences. McGraw-
SMALL, M. F. 1990: Alloparental behaviour in Barbary macaques, Mucacu sylvanus. Anim. Behav. 39,
247-305.
1640-1 658.
of predator classification and semantic communication. Science 210, 801-803.
Hill, New York.
297-306.

66 FISCHER et al., Acoustic Variation in Barbary-macaque Disturbance Calls
SNOWDON, C. T. & CLEVELAND, J. 1980: Individual recognition of contact calls by pygmy marmosets. Anim. Behav. 28, 717-727.
-_ & POLA, Y. V. 1978: Interspecific and intraspecific responses to synthesized pygmy marmoset vocalizations. Anim. Behav. 26, 192-206.
STRUHSAKER, T. T. 1967: Auditory communication among vervet monkeys (Cercopithecus uethiops). In: Social Communication among Primates (ALTMANN, S . A., ed.). Univ. of Chicago Press, Chicago. pp. 281-324.
SYMMES, D. & BIBEN, M. 1986: Maternal recognition of individual infant squirrel monkeys from isolation call playbacks. Am. J. Primatol. 9, 39-46.
TODT, D. 1986: Hinweis-Charakter und Mittler-Funktion von Verhalten. 2. Semiotik 8, 183-232. _ _ 1993: The dynamic of crying and its interactional role: phasic versus tonic components. In:
Social and Biological Aspects of Infant Crying (LESTER, B., NEWMAN, J. D. & PEDERSEN, F., eds). Plenum Press, New York, in press.
- _ , HAMMERSCHMIDT, K., ANSORGE, V. & FISCHER, J. 1995: The vocal behaviour of Barbary macaques. In: Current Topics in Primate Vocal Communication (ZIMMERMANN, E., NEWMAN, J . D., & JURGENS, U., eds). Plenum Press, New York, in press.
Perspective and projects of a long-term study. Primate Rep. 32, 19-30. DE TURCKHEIM, G. & MERZ, E. 1984: Breeding Barbary macaques in outdoor open enclosures. In:
The Barbary Macaque: a Case Study in Conservation (FA, J. F., ed.). Plenum Press, New York. pp. 241-261.
_ _ , _ _ & HULTSCH, H. 1992: The behaviour of Barbary macaques (Mucucu sylvanus L. 1758):
Received: August 25, 1994
Accepted: March 23, 1991 (W. Wickler)





























