Embed Size (px)
Citation preview

See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/24180853
Gago S, Elena SF, Flores R, Sanjuán R.. Extremely high mutation rate of a
hammerhead viroid. Science 323: 1308
Article in Science · April 2009
DOI: 10.1126/science.1169202 · Source: PubMed
CITATIONS
107
4 authors, including:
Some of the authors of this publication are also working on these related projects:
VIROID TAXONOMY View project
Evolution and Structure of Bipartite Networks View project
Selma Gago
Leibniz Institute for Plant Biochemistry
40 PUBLICATIONS 894 CITATIONS
SEE PROFILE
Santiago F Elena
Spanish National Research Council
472 PUBLICATIONS 10,778 CITATIONS
SEE PROFILE
Rafael Sanjuán
177 PUBLICATIONS 3,641 CITATIONS
SEE PROFILE
All content following this page was uploaded by Selma Gago on 21 May 2014.
The user has requested enhancement of the downloaded file.
Extremely High Mutation Rate of aHammerhead ViroidSelma Gago,1 Santiago F. Elena,1 Ricardo Flores,1 Rafael Sanjuán1,2*
Mutation rates vary by orders of magni-tude across species (1, 2), with the high-est rates measured so far corresponding
to RNAviruses (3), but little is known about otherRNA replicons. Viroids are plantpathogens with minimal non-protein-coding RNA genomesreplicated by host RNA poly-merases (4). We estimated themutation rate of Chrysanthemumchloroticmottle viroid (CChMVd),a 399-nucleotide chloroplasticviroid with hammerhead ribo-zymes. Hammerheads are RNAmotifs formed by three double-helix regions flanking a core of15 highly conserved nucleotidescritical for catalytic activity (5),which mediate self-cleavage ofreplicative intermediates and,hence, are essential for viroidreplication. Hammerhead viroidsshow elevated genetic variabil-ity (6), but this variability resultsfrom the combined action ofmutation and selection and there-fore cannot be used to directlyestimate mutation rates.
To achieve this goal, we in-oculated plants with an in vitrotranscript of CChMVd (7), andat the onset of symptoms wescreened for mutations at the 15 core nucleotidesplus the nucleotide preceding the self-cleavage sitein each of the two hammerheads (32 sites).Considering that these mutations are lethal forthe viroid, their population frequency must equalthe mutation rate because, despite multiplereplication rounds downstream from inoculation,they have necessarily been generated during thelast one. In three independent experiments, wefound three, seven, and five mutations in 63, 64,and 61 reverse transcription polymerase chainreaction (RT-PCR) clones, respectively (188 × 32= 6016 total target sites), yielding a mutation rateof 0.0025 T 0.0006 (SEM) per site and replicationcycle, that is, one mutation per 400 nucleotides(fig. S1).
In a control experiment, we sequenced RT-PCR clones from the in vitro transcript used forinoculations and found a single substitution in6525 sites. This result gives an error rate 17-foldlower than the estimated CChMVd mutation rateand discards any significant effect of RT-PCRartifacts. To confirm the lethality of the hammer-
head mutations sampled in vivo, we recreated themutations by site-directed mutagenesis and as-sayed for infectivity. Northern-blot hybridizationsindicated that plants inoculated with these mutants
had no detectable viroid RNA (fig. S2A). Further,self-cleavage analyses confirmed that all exceptone of the mutant hammerheads showed severelyreduced or null catalytic activity (fig. S2B).
To determine the strength of selection againstmutations elsewhere in the viroid genome, wecompeted 24 random-point mutants against thewild type. Sequencing of 138 RT-PCR clonesrevealed that 20/24 mutations had been purgedby selection at the onset of symptoms. In con-trast, 51 new polymorphisms appeared in thistime interval, showing that genetic variability israpidly regenerated because of highly error-pronereplication (fig. S3). We also inferred that ham-merheads are unlikely to constitute mutationalhotspots because polymorphisms did not mapmore frequently in hammerheads than in the restof the genome (Fisher exact test, P = 0.963)whereas the fraction of point mutations that wereselected against was also similar for these tworegions (7/8 and 13/16, respectively).
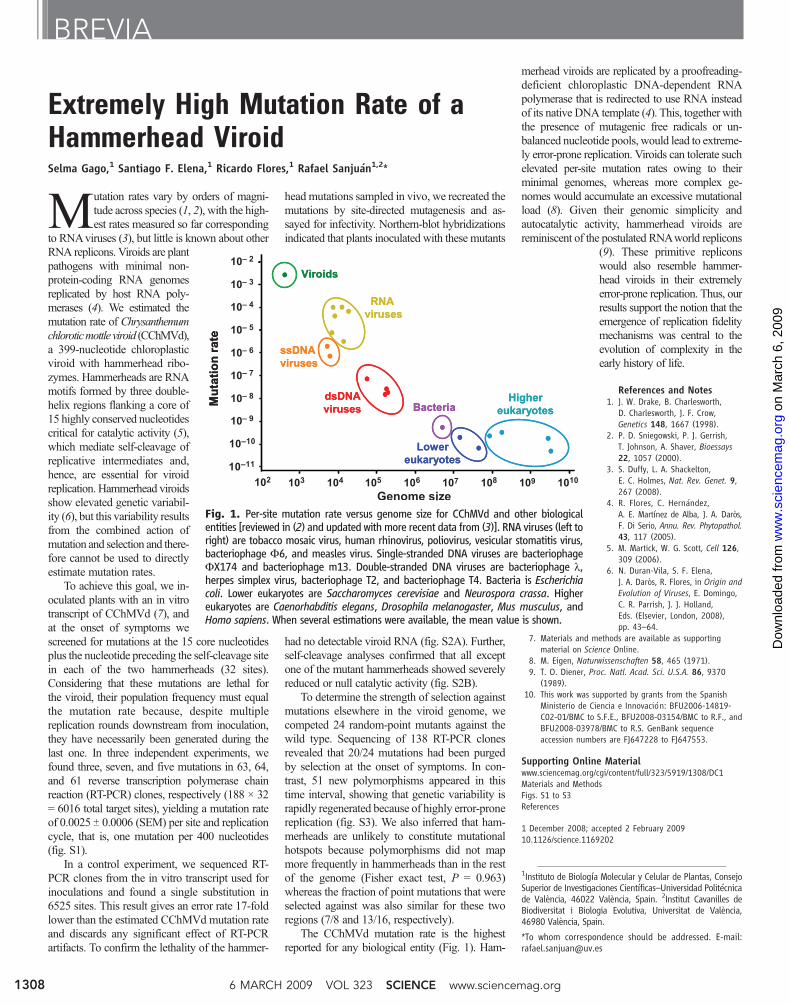
The CChMVd mutation rate is the highestreported for any biological entity (Fig. 1). Ham-
merhead viroids are replicated by a proofreading-deficient chloroplastic DNA-dependent RNApolymerase that is redirected to use RNA insteadof its native DNA template (4). This, together withthe presence of mutagenic free radicals or un-balanced nucleotide pools, would lead to extreme-ly error-prone replication. Viroids can tolerate suchelevated per-site mutation rates owing to theirminimal genomes, whereas more complex ge-nomes would accumulate an excessive mutationalload (8). Given their genomic simplicity andautocatalytic activity, hammerhead viroids arereminiscent of the postulated RNAworld replicons
(9). These primitive repliconswould also resemble hammer-head viroids in their extremelyerror-prone replication. Thus, ourresults support the notion that theemergence of replication fidelitymechanisms was central to theevolution of complexity in theearly history of life.
References and Notes1. J. W. Drake, B. Charlesworth,
D. Charlesworth, J. F. Crow,Genetics 148, 1667 (1998).
2. P. D. Sniegowski, P. J. Gerrish,T. Johnson, A. Shaver, Bioessays22, 1057 (2000).
3. S. Duffy, L. A. Shackelton,E. C. Holmes, Nat. Rev. Genet. 9,267 (2008).
4. R. Flores, C. Hernández,A. E. Martínez de Alba, J. A. Daròs,F. Di Serio, Annu. Rev. Phytopathol.43, 117 (2005).
5. M. Martick, W. G. Scott, Cell 126,309 (2006).
6. N. Duran-Vila, S. F. Elena,J. A. Daròs, R. Flores, in Origin andEvolution of Viruses, E. Domingo,C. R. Parrish, J. J. Holland,Eds. (Elsevier, London, 2008),pp. 43–64.
7. Materials and methods are available as supportingmaterial on Science Online.
8. M. Eigen, Naturwissenschaften 58, 465 (1971).9. T. O. Diener, Proc. Natl. Acad. Sci. U.S.A. 86, 9370
(1989).10. This work was supported by grants from the Spanish
Ministerio de Ciencia e Innovación: BFU2006-14819-C02-01/BMC to S.F.E., BFU2008-03154/BMC to R.F., andBFU2008-03978/BMC to R.S. GenBank sequenceaccession numbers are FJ647228 to FJ647553.
Supporting Online Materialwww.sciencemag.org/cgi/content/full/323/5919/1308/DC1Materials and MethodsFigs. S1 to S3References
1 December 2008; accepted 2 February 200910.1126/science.1169202
BREVIA
1Instituto de Biología Molecular y Celular de Plantas, ConsejoSuperior de Investigaciones Científicas–Universidad Politécnicade València, 46022 València, Spain. 2Institut Cavanilles deBiodiversitat i Biologia Evolutiva, Universitat de València,46980 València, Spain.
*To whom correspondence should be addressed. E-mail:[email protected]
Fig. 1. Per-site mutation rate versus genome size for CChMVd and other biologicalentities [reviewed in (2) and updated with more recent data from (3)]. RNA viruses (left toright) are tobacco mosaic virus, human rhinovirus, poliovirus, vesicular stomatitis virus,bacteriophage F6, and measles virus. Single-stranded DNA viruses are bacteriophageFX174 and bacteriophage m13. Double-stranded DNA viruses are bacteriophage l,herpes simplex virus, bacteriophage T2, and bacteriophage T4. Bacteria is Escherichiacoli. Lower eukaryotes are Saccharomyces cerevisiae and Neurospora crassa. Highereukaryotes are Caenorhabditis elegans, Drosophila melanogaster, Mus musculus, andHomo sapiens. When several estimations were available, the mean value is shown.
6 MARCH 2009 VOL 323 SCIENCE www.sciencemag.org1308
on
Mar
ch 6
, 200
9 w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

www.sciencemag.org/cgi/content/full/323/5919/1308/DC1
Supporting Online Material for
Extremely High Mutation Rate of a Hammerhead Viroid Selma Gago, Santiago F. Elena, Ricardo Flores, Rafael Sanjuán*
*To whom correspondence should be addressed. E-mail: [email protected]
Published 6 March 2009, Science 323, 1308 (2009)
DOI: 10.1126/science.1169202
This PDF file includes:
Materials and Methods Figs. S1 to S3 References

Material and Methods
Site-directed mutagenesis
A pUC18 derivative carrying a dimeric head-to-tail CChMVd wild-type sequence (CM20) (EMBL accession AJ878085) (S1), was used for site-directed mutagenesis as previously described (S2, S3).
In vitro transcription
Plasmid DNA was linearized and transcribed at 37ºC with T7 or T3 RNA polymerase to obtain dimeric CChMVd RNA of (+) or (−) polarity, respectively. For in vitro self-cleavage analysis, transcriptions were incubated 20 min in 2 mM MgCl2, quenched with 3 volumes of denaturing solution (8 M urea, 0.25% bromophenol blue and xylene cyanol), heated at 95ºC for 1 min, and the resulting RNAs were separated by PAGE in 5% gels containing 8 M urea that were stained with silver. For plant inoculation the DNA template was digested with DNaseI following transcription and the RNA was recovered by phenol extraction and ethanol precipitation. For competition assays plasmid DNA of the 24 mutants and of the wild-type were quantified spectrophotometrically by triplicate, pooled at equal amounts, and transcribed, whereas for the infectivity assays of the hammerhead mutants, each mutant clone was treated separately.
Plant inoculation
Transcripts were diluted in borate buffer and mechanically inoculated to one-month old chrysanthemum plants (Dendranthema grandiflora Tzvelev, cv. ‘Bonnie Jean’) (S1). In a preliminary titration assay, we determined that 10 ng per plant of the wild-type transcript elicited symptoms in all plants. For competition assays plants were inoculated with 200 ng of RNA to avoid random sampling of sequences, and for infectivity assays of hammerhead mutants, plants were inoculated with 25 ng per plant to minimize the probability of mutant reversion. Plants were maintained in a greenhouse at 20-22ºC with thermal oscillation between 25ºC and 15ºC and a photoperiod of 16 h.
RNA extraction
Apices and the first two expanding leaves were collected on ice and stored at −80ºC, ground in liquid nitrogen, and homogenized in extraction buffer (0.1 M Tris-HCl pH 9.0, 0.1 M NaCl, 10 mM EDTA, 0.1 M 2-mercaptoethanol, 5 M urea). Following centrifugation the supernatant was phenol-extracted, and the RNAs were recovered by isopropanol precipitation and resuspended in 0.5 mL of nuclease-free water.
1

RT-PCR, cloning and sequencing
RNA preparations were mixed with a specific complementary primer, heated for 2 min at 95ºC, ice-cooled and used for reverse transcription (RT) with MMLV reverse transcriptase and PCR amplification with the high-fidelity PhusionTM DNA polymerase. Reverse/forward primers annealing at positions 377-399/1-25, respectively, were used for RT-PCR amplification of the full-length CChMVd sequence. For amplification of the hammerhead region, reverse/forward primers annealed at positions 149-170/346-366, respectively. PCR products were extracted from agarose gels, ligated into pUC19/SmaI, column-purified, and used for electroporation. Plasmid DNA was purified from individual colonies and molecular clones were sequenced using vector-based primers.
We performed control experiments in which we applied the same RT-PCR protocol to the in vitro transcript used for inoculations. We found a single C → A substitution in 45 clones of a 145-nt region encompassing the two hammerheads, yielding an experimental error of 1.5 × 10−4.
Northern-blot analysis
Aliquots corresponding to 150 mg of fresh tissue per plant were mixed with 3 volumes of denaturing solution, heated at 95ºC for 1 min and separated by PAGE in 5% gels containing 8 M urea. Gels were stained with ethidium bromide and RNAs were electrotransferred to nylon membranes and fixed by UV-irradiation. Membranes were hybridized overnight at 70ºC in the presence of 50% formamide with a 32P-labeled riboprobe for detecting the CChMVd (+) strand, washed, and autoradiographed or quantitatively scanned with a bioimage analyzer.
2

�
��������� �������������� ��������� �������������� ������� ��������
���������� �������� �� ��� ����������� ���� ���� ��� ���������� ������
���������� ����� �� ��� ���� �� ���� ����� ��� ����� ����� ��� ��� ��������
���������� ���������������������� ����� ������� ����������� �������
���!������ "���������������� �������������� ��� ��������!��� ����������
#$���� ����� ������������������ ������%� ���#&���� �� �������
����� ��� ��!�� � ������ �� �� �!�� ������ ��� ����� �� ������� ���������
����������� ������� '������������ ���(��� ����� �����������������������
���� ��������������������� �������� ��������� ������ ���������� ��#��$�
������ ��)��#*����������� ����������+������ ��������!��� �� ��&��
����� � ��� ���������� ����� � ,�� ������ �� ������� ��� �� �� ���
�������������� ���� ��� -� �� �%�������� �χ+ �� � �. /��+0� �����������
������� ����� ������������������� ����+ �� �
CChMVd (+)
Helix IHelix II
Helix III
G AGU
U AG CU G
CG
G CG CA UA UU AU A
C GC GU AU A
C
3´ 5´
Loop 2
U AU A
GAGA
C
G
AA
AGU
G CG C
A GUC
C GC G
G U
CU
GA
C
UC
U
G CG CAA
U A
A10
3´ 5´
CGC
GAGA
C
G
AA
AGU G U
CU
GA
C
3´ 5´
GA
C
G
AA
AGU
C
G
C
A
UG
17
16.115.1
3
4
5678
14
16.215.2
9
10.111.11213
Loop 1A
GC
UGA
UUA
3´ 5´
AG
UC
M1: U16.1→→→→�C17→→→→�
CG�
GAGA
C
G
AA
AG� G U
CU
GA
C AGC
UGA
UUA
AG
UC
C�
�
GAGA
C
G
AA
AGG G U
CU
GA
C AGC
UGA
UUA
AG
UC
CG
G
GAGA
C
G
AA
AGU G U
CU
GA
C AGC
UGA
UU
A
AG
UC
M2: U16.1→→→→GC17→→→→�G1.1 →→→→�
M3: C17→→→→G
CGC
GAGA
C
G
AA
AGU G U
UU
GA
C AGC
UGA
UUA
AG
UC
CGC
GAGA
C
G
AA
AGU G U
CU
AA
C AGC
UGA
UUA
AG
UC
CGC
AA
C
G
AA
AGU G U
CU
GA
C AGC
UGA
UUA
AG
UC
CGC
GAGA
U
G
AA
AGU G
U
CU
GA
C AGC
UGA
UU
A
AG
UC
M4: C3→→→→U M5: G5→→→→A M6: G10.1→→→→A M7: C15.2→→→→U
10.211.2
2.1
1.22.2
1.1
3´ 5´
3´ 5´
3´ 5´ 3´ 5´
3´ 5´ 3´ 5´3´ 5´ 3´ 5´ 3´ 5´
3´ 5´ 3´ 5´ 3´ 5´ 3´ 5´ 3´ 5´3´ 5´ 3´ 5´ 3´ 5´
3´ 5´
3´ 5´ 3´ 5´ 3´ 5´
Wild-type
CChMVd (-)
Helix I
Helix II
Helix III
G AGU
U AU GU A
CG
A UA UG CG CC GC G
G CG CG CG C
C
3´ 5´
Loop 2
A UA U
G CG C
A A
U AU A
U
A
U AU A
GA
17
16.115.1
34
5678
14
16.215.2
9
10.111.11213
Loop 1
10.211.2
2.1
1.22.2
1.1
UGG
U
C GC GC GC GU AU AC GC G
A
U GU GU AU AC GC GG CG C
AC
AC
CG
A UA UG CG C
1.31.41.51.6
GA
C
G
AA
AGU
C
G
C
A
UG
U AU GU A
CG
A UA UG CG CC GC G
G CG C
C
G CG C
U AU A
U
UG3´ 5´
3´ 5´
3´ 5´
GA
C
G
AA
AGU
C
G
C
A
UA
U AU GU A
CG
A UA UG CG CC GC G
G CG C
C
G CG C
U AU A
U
UG3´ 5´
3´ 5´
3´ 5´
GA
C
G
AA
AGU
C
G
C
A
UG
U AU GU A
CG
A UA UG CG CC GC G
G CG C
U
G CG C
U AU A
U
UG3´ 5´
3´ 5´
3´ 5´
GA
C
G
AA
AGU
C
A
C
A
UG
U AU GU A
CG
A UA UG CG CC GC G
G CG C
C
G CG C
U AU A
U
UG3´ 5´
3´ 5´
3´ 5´
GA
C
G
AA
AGU
C
G
C
AU
G
UUU
C
A UA UG CG CC GC G
G CG C
�
GG
U AU A
U
UG3´ 5´
3´ 5´
3´ 5´
GA
C
A
AA
AGU
C
G
C
AU
G
U AU GU A
CG
A UA UG CG CC GC G
G CG C
C
G CG C
U AU A
U
UG3´ 5´
3´ 5´
3´ 5´
GA
C
G
AA
AGU
C
G
C
AA
G
U AU GU A
CG
A UA UG CG CC GC G
G CG C
C
G CG C
U AU A
U
UG3´ 5´
3´ 5´
3´ 5´
GA
A
G
AA
AGU
C
G
C
AU
G
U AU GU A
CG
A UA UG CG CC GC G
G CG C
C
G CG C
U AU A
U
UG3´ 5´
3´ 5´
M9: C17→→→→�G1.1 to C1.6→→→→�
M8: C17→→→→UWild-type
�
�
�
�
��
M10: U4→→→→A
M11: G5→→→→A M12: G8→→→→A M13: G12→→→→A M14: C15.2→→→→A
3´ 5´
a
b

b
899 nt
177 nt722 nt
576 nt
399 nt
227 nt655 nt
626 nt
399 nt
256 nt
882 nt
CChMVd (-) hammerhead mutants
M8 M9 M10 M11 M12 M13 M14 WT882655626
399
256
227
M1 M2 M3 M4 M5 M6 M7 WT899722576
399
323
177
CChMVd (+) hammerhead mutants
a
P1 P2 P3 P4 P5 P6 P7 P8 (+)P10P9 (-)
M1 M2 M3 M4 M5 M6 M7 H (+)(-) M8 M9 M10 M11 M12 M13 M14 H (+)(-)
P1 P2 P3 P4 P5 P6 P7 P8 (+)P10P9 (-)
CChMVd (+) hammerhead mutants
Wild-type Wild-type
4S
4S
4S
4S
CChMVd (-) hammerhead mutants
323 nt
�
��������� ����������������� ��������������������������� ������� ������� ��
��� ������� � �� ��� ��� ��� ����������� �� �� �� ���� ��� ��� �!�� ""�#$�%� � ��
������ �����!���� �������� ���� �� �&'(�� �����%��)���������� �� �&�!!���!�����(*�
� ���+���!��!��� ��������� ����������� ���!��� �� ����!�����*� ����������� ��� ���
�!��&�� ���!�����(*������������������!��� � ����!�����������!��� ���%��,�������� ���
� ���+����������!������ ��-�.�����/�.������������� ���������������� �� ��������� ���
��� �!������� *����!�� ����%��+��������� ���!������� ��������0������� ���!��� ��
&1(*���������������! ��� ��&'(����������������� ���&−(���� ���� ����������������%���2�+����� ����� ��� ����������� ���������������������� ����%��3���!������ �
������ ��� ������ �������� �� ��� �� ��������!���� �� ������ ��� �� ��� ������
""�#$��&'(�+��*� ������������ ��� ��������������� ���� ���������������%�
�2������������������� ��������� ������! ������ ������ ������������� �� ������ ��
��� �!�%� � 4!!��� !�����5� ������ �� ��!����� � ��� ��� ��� ������ ����� �� ���
������! ������ �����6!�� �����������������!����� �%��$�� ��������������7�����������
���� ��������0����� ����� ���*����!�� ����*� ������������������ ����������0�����
���� ������ �����6!�� ��� ������������� ����� ������� ��� ������ ���� ��� ���%��
,� ���!�����5��������������� ��������������������� ����� ����������� �����
������������� �� ������ ��� ��� �!�%������������ ������ ����� �� ���������� ���
!������ ������! ��������� ���!����� ������� �������������������������� ����������
���������������������������� ����������������������� ��%�

�
GCUGA
120
CU
U CAG G UC
U CG ACU
G
. . . . . . . . . . . . . . . . . . . . . . . .
GA
160
180
200
240
380
GGC
ACCUGAU
G
C
C
U A G
CGUC CA A CC C C
G G GAG
GAAA
GGGGUUGGGA
C
G
GGCC
AG
GGCC
AG
UCCCAGU
UCCCAGU
UGGUU
GGUUC
GCU
GCUC
UCGUAGUC
AC
AGCCACUGGGGA
ACC
UAG
GCAG
AU
GGCUGGAC GGCUGGACGG
A UGUC U AG U C C
CCG
AGGUC . . . . . . . . . . . . . . . . . .
.. .
. .
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
. . . . . . . . . . . . . . . . . . . . . . . .
1
UCCUUUGGAGU
UCCUUUGGAGUA
CUCCAGAGGA .
.
.
.
.
.
.
.
.
.
.
CCGGAA C
GUUCCG. . . . .
. . U
UG
GGU
UUGA
AAC C C
CC
AA
AG
GUA A
AUA
CC
UC
. .
. .
. CUCU
CUCU
AA G
GGAG
GGAG
.
.
.
.
.
.
.
. AGGU
CG
U A AAA
CU
UC
C. .
. . .
140
60 100
UC C A
U U UC UC A U CA G GAA
A C CC
A. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
U
GGCUG
GGCUGUG
CG .
20
UC
GGUG
UCUGAA
GAAACCUC
U
40
GAUCCAU
GACA
GGAUC .
.
.
.
.
80
UAA A
GGAG
GC
GGAG
G
GGAG
GC
UU U
C
C
C
CC
UC
CA
AG
GCU
UC
C
CC
UC
CA
AG
GCU
UC
CC
UC
CA
AG
GCU
UC
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
320320
340
360360
280
C C AU UU CGAGAUGG. . . . . . . . . . . . . . . .
300300
U C U
GAA
GG
. .
U
A G
CA
A. C
GUUGGA A U
UCU
CCUCGG
260
.
CU
A 220
G
CC
A
AA
UU
AA
UU
AA
A
U
UU
UA
A
UA
A
UA
U
A
G
G
C
U
U
U
C
A
UUAA
CC U
A
GG
UU
U
AA
AAA
GG GG
U
U
UA
U
U
U
U
UU
AUUAAUU
UU
A
GGGG
U
A
G
U
AAU
C
CC
A
U
U
�
���������������������� ������������������������������������������������������
������������������������������������������������� ���������������������� � ������
��������������������� ����������� !�"�����#���������� �������������$��"��������
�� ��� �!�������������%������������� �&� ���������������� ������������ ������
������ ���������"� ��������� ��� � '������ ����� �������� � ���� �(��!� %�����!� ����
����������� ������ � )���*+��������������������������� ��������������������!������"�
��� ��� ��!������ ����� ���� �� ������� �� � ����������� ��� #,*�� ���"� ��� ����� ����
����� ������ � - ������ ���� ������� ������ ��� ��������� ��������� ������ � ��� ��� ��
����� ��������������������� ������������ ��� �"���������� �"�������������������� ����
.������������!���������������������� ���������� ������������������� �&��������
����������������� ��������������������������������������� ���������� ��������

Supplementary References S1. B. Navarro and R. Flores, Proc. Natl. Acad. Sci. USA 94, 11262-11267 (1997). S2. R. Sanjuán and J.A. Daròs, J. Virol. Methods 145, 71-75 (2007). S3. S. Gago, M. de la Peña, R. Flores, RNA 11, 1073-1083 (2005). S4. M. Martick and W. G. Scott, Cell 126, 309-320 (2006). S5. R. Flores, C. Hernández, M. de la Peña, A. Vera, J. A. Daròs, Methods Enzymol. 341,
540-552 (2001).
6
View publication statsView publication stats