Embed Size (px)
Citation preview

DEVELOPMENTAL DYNAMICS 204:421-431(1995)
Expression of the Integrin Subunit a9 in the Murine Embryo ANGELA WANG, LAURA PATRONE, JOHN A. MCDONALD, AND DEAN SHEPPARD Lung Biology Center, San Francisco General Hospital, University of California, San Francisco, California 94143 (A. W., D.S.); Samuel C. Johnson Medical Research Center, Mayo Clinic, Scottsdale, Arizona (JA.M. , L.P.)
ABSTRACT The a9 integrin subunit is ex- pressed in adult skeletal muscle, visceral smooth muscle, hepatocytes, squamous epithelium, and airway epithelium. The in vivo function of this protein is unknown. Thus far, only a single a9- containing integrin has been identified (a9pl) and only a single ligand (tenascin) has been found for this integrin. In order to gain insight into the po- tential function of a9 integrin(s), we examined the spatiotemporal distribution of the a9 subunit and tenascin during murine embryogenesis. In all tis- sues where a9 was expressed, its appearance was associated with other evidence of cell differentia- tion. In developing airway, visceral, and vascular smooth muscles, the onset of a9 expression either coincided with or immediately followed the ex- pression of a-SM actin. Expression of a9 in epithe- lia was restricted to the choroid plexus and the basal cell layer of squamous epithelia where its appearance coincided with the development of stratification. a9 immunostaining was first de- tected in developing skeletal musculature when skeletal rnyotubes formed. Tenascin expression was detected in many, but not all tissues found to express a9. For example, the hair germs of matur- ing hair follicles exhibited high levels of a9 stain- ing, but no tenascin immunoreactivity was de- tected either within the hair germ themselves or in the adjacent dermis. In some tissues where tenas- cin expression colocalized with a9, expression patterns were not synchronous. Although a9 ex- pression was associated with the onset of tissue differentiation, its expression was not limited to terminally differentiated cells. In fact, in the skin, a9 expression appeared restricted to cells known to retain the capacity to proliferate, i.e., basal cells and hair germs. Thus, a9 integrinb) are not likely to contribute to the early steps in organ formation, but probably play a role in the maturation and/or maintenance of a variety of differentiated tissues. The expression of a9 without its only known ligand, tenascin, suggests the existence of addi- tional ligands. o 1996 Wiley-Liss, Inc.
Key words: Integrin subunit a9, Murine embryo- genesis, Tenascin
INTRODUCTION The integrin family of cell adhesion receptors medi-
ates both cell-cell and cell-extracellular matrix inter- actions (Hynes, 1992). Integrins are heterodimers, composed of non-covalently associated alpha (a) and beta (PI subunits; 16 a subunits and 8 p subunits have been identified. Substrate specificity appears to be a function of not only the heterodimer itself, but also of the specific tissue or cells in which the integrin is ex- pressed. Some integrins bind only a single extracellu- lar matrix protein while others recognize multiple ex- tracellular proteins or even other integrins. An extracellular matrix protein may also be a ligand for more than one integrin; e.g., alpl, a2p1, a&, and a6pl all bind laminin in vitro. Whether all of these interactions are physiologically relevant and whether functional differences between integrins with the same substrate specificity exist in vivo is unknown. Cer- tainly, a given cell may express multiple integrins with overlapping substrate specificities (Hynes, 1992). a9 is a recently described integrin subunit that forms
a heterodimer with pl (Palmer et al., 1993). In contrast to some other integrin subunits, only one splice variant of a9 has been identified to date. Cell adhesion assays demonstrate that a9pl is a receptor for the extracellu- lar matrix protein tenascin-C (hereafter referred to as tenascin). In the adult mouse, a9 is selectively ex- pressed in a few highly specialized tissues: squamous epithelia, smooth and skeletal muscles, airway epithe- lium, and hepatocytes; however, the role a9 might be playing in these diverse tissues remains unknown (Palmer et al., 1993). Studies of the spatiotemporal dis- tribution of integrins and their ligands during embryo- genesis have provided clues to their biological func- tions. For example, the co-localization of the a4 integrin subunit and of the a4-integrin ligand VCAM-1 on developing myotubes provided the first evidence that these proteins played a central role in myotube fusion. Differences in the temporal pattern of expres- sion of the integrin subunits a1 and a6 during avian embryogenesis (Muschler and Horwitz, 1991; Bronner-
Received January 9, 1995; accepted August 9, 1995. Address reprint requests/correspondence to Angela Wang, Lung Bi-
ology Center, Box 0854, University of California, San Francisco, San Francisco, CA 94143.
0 1995 WILEY-LISS. INC.

422 WANG ET AL.
Fraser et al., 1992; Duband et al., 1992) have suggested that these integrins play distinct roles during skeletal myogenesis and neurogenesis. Therefore, in order to further elucidate the possible function(s) of a9 and its relationship with tenascin, we used immunohistochem- istry to examine and compare the expression pattern of these two proteins during murine embryogenesis. We found that the onset of a9 expression occurred rela- tively late during development and was associated with either biochemical and/or morphological evidence of tissue maturation. Although, in general, a9 expres- sion colocalized with that of tenascin, differences in both temporal and spatial localization of these two pro- teins suggest that another ligand(s) for a9 exists.
EXPERIMENTAL PROCEDURES Antibodies and Embryos
Balb/c female mice were mated with Balb/c males and then checked every 8 hr for a vaginal plug, the appearance of which represented day 0. The female mice were killed at stated stages of gestation. Their uteri were removed via laparotomy and placed into Hanks’ buffered saline. The embryos were then dis- sected free and placed in 20% sucroselPBS for 15 min before being quick frozen in liquid nitrogen.
Purification of a polyclonal anti-peptide antibody di- rected against the predicted C-terminal domain of a9 has been previously described (Palmer et al., 1993). This antibody has been previously used for immuno- histochemistry (Palmer et al., 1993) and Figure 1 de- picts a Western blot using this antibody to detect a9 in adult mouse liver. The monoclonal antibodies against mouse tenascin and a synthetic peptide of human a-SM-actin (Skalli et al., 1986) were obtained from Sigma Chemical Co. (St. Louis, MO).
Immunoprecipitation and Western Blotting Freshly harvested adult mouse liver was minced and
then ultrasonicated in IP Buffer (150 mM NaC1, 1% Triton, 0.1% SDS, 1 mM CaC1). The homogenate was preabsorbed with protein A-Sepharose beads (Pharma- cia LKB Biotechnology, Piscataway, NJ) before being incubated with either the rabbit anti-a9 antibody or a nonspecific antibody overnight at 4°C. Samples were remixed with Protein A-sepharose and beads washed with IP Buffer containing 300 mM NaC1. Samples were eluted in Laemmli sample buffer containing DTT. The proteins were separated on a 10% polyacrylamide gel and then transferred onto Immobilon (Millipore, Bed- ford, MA) using a Hoefer transfer apparatus. The filter was blocked with 3% BSA overnight at 4°C and blotted with rabbit anti-a9 at 1200 dilution for 1 hr followed by a 45-min incubation with peroxidase conjugated goat anti-rabbit secondary antibody (Cappel, Malvern, PA; 1:2,000). The filter was then exposed to film after a brief incubation in Luminol (Amersham, Arlington Heights, IL).
Immunohistochemistry Embryos from gestation day 7,9, 10, 12.5,14.5, 16.5,
17.5, and 18.5 were studied. Sagittal sections (5 pm) of whole embryos were cut using a Jung-Reichart cry- ostat. Slides were air-dried for 5 min before being fixed in 2% paraformaldehyde at room temperature for 5 min. Sections were blocked with 0.5% casein/0.05% thimerosal/PBS, endogenous peroxidase activity blocked using Peroxoblock (Zymed, San Francisco, CAI and the sections incubated at 4°C overnight with either a 1:200 dilution of the polyclonal anti-a9 peptide anti- body (in 0.5% casein/0.05% thimerosal/PBS), anti-ag plus 0.5 pg/ml dilution of a9 peptide (peptide block), 1:1,500 dilution of the monoclonal anti-a-smooth mus- cle (a-SM) actin antibody, or 1:500 dilution of the monoclonal rat anti-mouse tenascin antibody. Sections were then rinsed, incubated with either a 1:250 dilu- tion of peroxidase-conjugated secondary goat anti-rab- bit (Vector, Burlingame, CAI or horse anti-mouse an- tibody (Vector) or 1:lOO dilution of the peroxidase conjugated sheep anti rat (Amersham) and color reac- tions developed using a DAB (3,3’-diaminobenzidine)- hydrogen peroxide kit (Zymed). Control slides were processed identically except that primary antibody was omitted.
RESULTS a9 Expression in Smooth Muscle
Pulmonary smooth muscle. The murine lung forms as an epithelial outpouching of the foregut into the surrounding mesenchyme at approximately days 8-9 gestation (embryonic stage). Between days 9-16 (E9-16), epithelial branching morphogenesis occurs, resulting in the formation of a primitive bronchial tree (glandular stage). Invasion by pulmonary vasculature begins at E l6 (canalicular stage) followed by the alve- olar stage which is characterized by the coalescing mesenchyme around the nascent airspaces and forma- tion of alveoli. Alveolar maturation is not completed until after birth (ten Have-Opbroek, 1991).
a9 immunoreactivity in the murine lung was not de- tected until E12.5. Moderate levels of staining were seen in the paralumenal cells surrounding the more- developed proximal, primary bronchioles (Fig. 2B ). On- set of a9 expression followed the rostra1 to caudal dif- ferentiation pattern of smooth muscle cells along the airways and coincided both temporally and spatially with that of a-SM actin (Fig. 2C) (Mitchell et al., 1990). The onset of tenascin expression in the lung also oc- curred in a proximal to distal sequence. Tenascin stain- ing was first detected on E12.5 when it was found sur- rounding only those airways already positive for a9 and a-SM actin (Fig. 2D).
The expression of a9 in airway smooth muscle rap- idly increased to adult levels and remained high throughout the rest of gestation. a9 expression in the lung was restricted to airway smooth muscle through- out all of the gestational ages examined (Fig. 3A,C). In

a9 IN MURINE EMERY 0 423
Fig. 1, Detection of murine a9 using polyclonal rabbit anti-a9 anti- body. Equal amounts of homogenized murine liver were immunoprecip- itated using either the anti-a9 antibody (lane A) or a nonspecific antibody (lane B). The resulting samples were separated on a 7.5% polyacryl- amide gel. lmmunoblotting was then performed using the polyclonal anti-a9 antibody. Molecular weight markers are shown to the left.
contrast, tenascin expression was initially restricted to the epithelial-mesenchymal interface and smooth mus- cle of the airways but was widely distributed through- out the mesenchyme of the nascent alveolar septa by E17.5 (Fig. 3B,D).
Gastrointestinal smooth muscle. Like the lung, the digestive tract matures in a rostra1 to caudal se- quence (Duband et al., 1992; Beaulieu et al., 1993). At E10.5, neither a-SM actin nor a9 was detectable any- where along the gastrointestinal tract. By E12.5, a9 and a-SM actin were strongly expressed by differenti- ating smooth muscle cells of the esophagus (Fig. 4A,B), while tenascin staining was present in the matrix sur- rounding the cells as well as at the epithelial-mesen- chymal interface (Fig. 40. Distally, the condensing mesenchyme surrounding the intestines was distin- guished by prominent staining for a-SM actin (Fig. 4E) as well as faint expression of a9 (Fig. 4D). Onset of tenascin expression lagged behind that of a9; tenascin staining was not apparent in the lower gastrointestinal tract at this stage (Fig. 4F). By E17.5, the muscular layers of the intestines could be readily distinguished. Both smooth muscle layers of the digestive tract showed moderate a9 (Fig. 5A,C) and tenascin staining (Fig. 5B,D). a9 was also present in the muscularis mu-
cosa while tenascin was expressed at the epithelial- mesenchymal interface.
Vascular smooth muscle. In contrast to its pattern of expression in airway and gastrointestinal smooth muscle, a9 expression in vascular smooth muscle changed with development. Vascular smooth muscle appeared to differentiate earlier than either airway or visceral smooth muscle as a-SM actin staining was very prominent surrounding vascular structures by E10.5 (not shown). a9 staining, however, did not ap- pear until E12.5, coinciding with the onset of tenascin expression. Although tenascin staining was seen to completely encircle small- and medium-sized arteries (Fig. 4D-F), its expression around the dorsal aorta was somewhat irregular a t this stage (Fig. 4G-I). By E17.5, a9 staining had disappeared from the aorta and other large vessels (Fig. 5A), while tenascin continued to be highly expressed by all vascular smooth muscle (Fig. 5B).
at9 Expression in Skeletal and Cardiac Muscle In the mouse, somite formation occurs between
E8-10 (Furst et al., 1989). a9 was not detected during this stage, nor at any point during subsequent myo- tome development and myoblast migration. Low levels of a9 immunoreactivity in skeletal muscle were ini- tially detected in the diaphragm and tongue (both of somitic origin) on E12.5, correlating with the onset of myotube formation. The skeletal musculature of the trunk and limbs mature somewhat later and a9 stain- ing was not seen until E14.5. As in smooth muscle, staining for a9 in skeletal musculature was initially faint, increased rapidly with myotube differentiation and then remained constant after cell maturation.
At E12.5, both the diaphragm and tongue already exhibited moderate to high levels of tenascin staining (not shown) as did the limb buds (Fig. 4E) although these latter structures were still negative for a9 (Fig. 4D). The tongue musculature continued to exhibit mod- erate levels of a9 and tenascin staining through late gestation (Fig. 6A,B), however, tenascin expression in limb muscles had greatly declined by late gestation, while remaining intense in tendons and myotendinous junctions (not shown).
No immunoreactivity for a9 was detected in cardiac muscle during any of the gestational ages examined.
a9 Expression in Epithelial Tissues Embryonic skin consists initially of two cell layers: a
temporary, protective upper layer known as the peri- derm and a basal layer that eventually becomes the germinative layer of the epidermis. Gradually, the periderm regresses as the cells of the stratum germi- nativum divide, become stratified and keratinized.
Expression of a9 was not detected in the skin until E16.5, coinciding with epidermal stratification and cornification. a9 immunoreactivity was confined to the basal cell layer and was very intense (Fig. 7A). Tena- scin immunoreactivity in the skin was also detectable

424 WANG ET AL.
Figs. 2 and 3.

a9 IN MURINE EMBRYO 425
by E16.5, and staining, although variable in intensity, was continuous along the dermo-epidermal junction (Fig. 7B).
Hair development begins after the onset of stratifi- cation. Initially, cells comprising the early pregerm in- vaginate into the dermis. These cells reacted strongly with the a9 antibody. As the follicle elongated, how- ever, a9 staining became restricted to the cells com- prising the hair germ (Fig. 7A,C). In contrast to the epidermis where keratinization is general and contin- uous, the process in hair follicles is restricted to the hair germ (Kaplan and Holbrook, 1994). The outer root sheath of more mature hair follicles exhibited no 019 immunoreactivity, even though this layer is continu- ous with the basal cell layer of the skin. Tenascin stain- ing varied with the stage of hair follicle development. With the onset of follicle growth, intense tenascin staining appeared in the mesenchymal cells immedi- ately underlying the hair germ as well as along the outer follicle sheath (Fig. 7B). As the follicles matured, a proximal to distal gradient developed with strongest tenascin staining seen along the follicle sheath closest to the epidermis and very little or none detected in association with the dermal condensation (Fig. 7B,D). No tenascin immunostaining was detected within the hair germ of mature hair follicles.
Other epithelia. a9 staining of the basal cell layer of oropharyngeal epithelium also appeared late in ges- tation, after the onset of stratification. Although the subepithelial mesenchyme of both the tongue, palate, and esophagus exhibited moderate to high levels of te- nascin immunoreactivity by E12.5, staining for a9 ap- peared gradually, progressing caudally from the lips towards the esophagus. By E17.5, a9 staining could be clearly seen in the basal cell layer of the tongue epi- thelium and had just begun to appear in that of the
~
Fig. 2. a9 and tenascin expression during pulmonary smooth muscle development. Sagittal sections through lung bud at E12.5 gestation. A: No primary antibody, staining with secondary anti-mouse antibody only; none of the secondary antibodies used exhibited nonspecific staining. Low power view of lung bud in which multiple developing airways can be seen. Thick arrow points to airway shown in high-power view in B, C, and D. B: Stained for as; (C) for a-SM actin; (D) for tenascin. At this stage, a9 is found only in the paralumenal cells (thin arrows) surrounding the more proximal primary bronchioles and posterior trachea, coinciding with the onset of a-smooth muscle (SM) actin immunoreactivity. Tenascin expres- sion is seen only in the condensing mesenchyme, including the epithelial- mesenchymal border, of airways positive for a9. A, x 10; B-D, x 40.
Fig. 3. Expression of a9 and tenascin in the trachea and lung of an E17.5 murine embryo. Sagittal sections through the trachea (A, B) and the lung (C, D) are shown. Sections were stained for a9 (A, C) or for tenascin (B, D). The smooth muscle of the trachea (found only along the posterior wall) and airways demonstrate moderate a9 staining. Note that the airway epithelium is negative for a9 (thin arrows). In addition to its presence in smooth muscle, tenascin expression can now be seen along the epithelial-mesenchymal interface (thick arrows) circumscribing the trachea, airways as well as the developing alveoli. An accompanying blood vessel in the lung parenchyma remains positive for tenascin while no expression of a9 is evident. tr, trachea; a, airway; bv, blood vessel; ep, airway epithelium. A-0, x 20.
proximal hard palate (Fig. 6A). No airway epithelial expression of a9 was detected at any of the gestational timepoints studied, and morphologically, the airway epithelium appeared immature. None of the three pri- mary cell types found in adult airway epithelium, in- cluding ciliated cells, was present (Fig. 3A). No stain- ing of intestinal, transitional, or dental epithelium for a9 was seen in the developing embryo. Tenascin stain- ing appeared at the epithelial-mesenchymal interface in all of these tissues (not shown).
a9 Expression in the Nervous System a9 was not detectable in the embryonic murine ner-
vous system except in the epithelium covering the' choroid plexus. The choroid plexus is actually a spe- cialized meninx which produces cerebrospinal fluid and also regulates the concentration of micronutrients in the CSF (Spector and Johanson, 1989). The choroid arises from the pia mater, the innermost meninx of the brain. The pia mater forms a vascular plexus over the roof of the ventricle. This plexus, which is covered by ependymal epithelium, then invaginates into the ven- tricle. By E12.5, the frond-like structure of the choroid plexus could clearly be seen, its epithelial covering staining intensely for a9 (Fig. 8A). Tenascin immuno- reactivity was also detected in the choroid plexus, just beneath the epithelium (Fig. 8B).
Other Tissues Although a9 is found in adult liver, embryonic and
fetal liver exhibited only faint staining that was diffi- cult to distinguish from background. In contrast, ten- ascin expression in developing liver was quite high. Neither a9 nor tenascin immunoreactivity was de- tected in the pancreas and thyroid at any of the gesta- tional timepoints examined. Thymus, cartilage, and bone remained negative for a9, but stained strongly for tenascin during development (not shown).
DISCUSSION We have investigated the spatioternporal expression
pattern of the a9 integrin subunit and its proposed ligand tenascin during murine embryogenesis. Expres- sion of a9 was detected in visceral and vascular smooth muscles, squamous epithelia, and epithelium of the choroid plexus, and skeletal muscle. In all of these tis- sues, onset of a9 immunoreactivity occured relatively late during gestation. No evidence of a9 expression could be detected prior to gestational day 12.5. Rather than being associated with the early stages of organ morphogenesis, the appearance of a9 expression corre- lated with the development of the mature tissue phe- notype. Tenascin expression colocalized to most murine fetal tissues found to express a9. In some cases, e.g., airway smooth muscle, the patterns of expression of both proteins were quite similar: tenascin expression, like that ofa9, was persistent and constant throughout late stage of gestation. In other sites, expression pat- terns of 019 and tenascin were either dyssynchronous
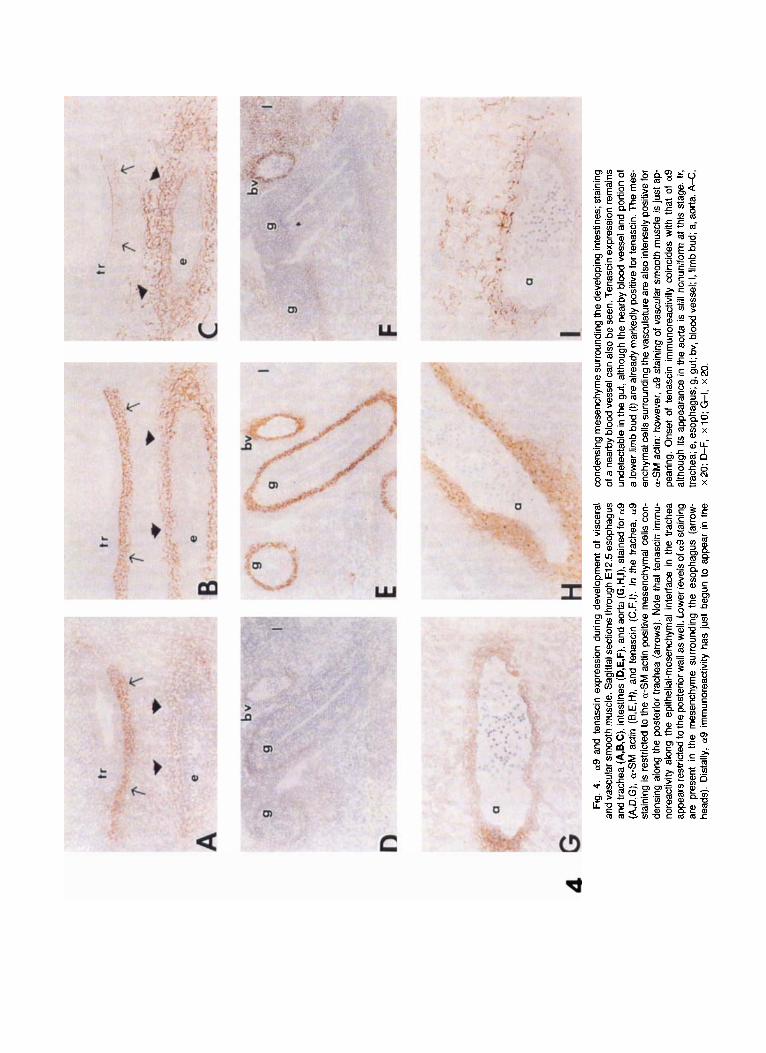
4 Fi
g. 4
. a9 a
nd t
enas
cin
expr
essi
on d
urin
g de
velo
pmen
t of
vis
cera
l an
d va
scul
ar s
moo
th m
uscl
e. S
agitt
al se
ctio
ns th
roug
h E
l 2.5
eso
phag
us
and
trach
ea (A
,B,C
), in
test
ines
(D,E
,F),
and
aorta
(G,H
,I),
stai
ned
for a9
(A,D
,G),
a-S
M a
ctin
(B,
E,H
), an
d te
nasc
in (
C,F
,I). I
n th
e tra
chea
, a9
stai
ning
is r
estri
cted
to th
e a-
SM
act
in p
ositi
ve m
esen
chyr
nal c
ells
con
- de
nsin
g al
ong
the
post
erio
r tra
chea
(ar
row
s). N
ote
that
tena
scin
imm
u-
nore
activ
ity a
long
the
epith
elia
l-rne
senc
hyrn
al in
terfa
ce i
n th
e tra
chea
ap
pear
s res
trict
ed to
the
post
erio
r wal
l as
wel
l. Lo
wer
leve
ls o
f a9
stai
ning
ar
e pr
esen
t in
the
mes
ench
yme
surro
undi
ng t
he e
soph
agus
(ar
row
- he
ads)
. Dis
tally
, a9 i
rnm
unor
eact
ivity
has
just
beg
un to
app
ear
in th
e
cond
ensi
ng m
esen
chym
e su
rroun
ding
the
deve
lopi
ng in
test
ines
; sta
inin
g of
a n
earb
y bl
ood
vess
el c
an a
lso
be s
een.
Ten
asci
n ex
pres
sion
rem
ains
un
dete
ctab
le in
the
gut,
alth
ough
the
near
by b
lood
ves
sel a
nd p
ortio
n of
a
low
er li
mb
bud
(1) ar
e al
read
y m
arke
dly
posi
tive
for t
enas
cin.
The
mes
- en
chym
al c
ells
sur
roun
ding
the
vasc
ulat
ure
are
alSo
inte
nsel
y po
sitiv
e fo
r a-
SM ac
tin; h
owev
er, a9
stai
ning
of v
ascu
lar s
moo
th m
uscl
e is
just
ap-
pe
arin
g. O
nset
of
tena
scin
im
mun
orea
ctiv
ity c
oinc
ides
with
tha
t of
a9
alth
ough
its
appe
aran
ce in
the
aorta
is s
till n
onun
iform
at t
his
stag
e. lr
, tra
chea
; e, e
soph
agus
; g, g
ut; b
v, b
lood
ves
sel;
I, lim
b bu
d; a
, aor
ta. A
-C,
x 20
; D
-F,
x 10
; GI,
x 20
.

a9 IN MURINE EMBRYO 427
5
6
Fig. 5. Expression of a9 and tenascin in the lower esophagus and aorta of an E17.5 murine embryo. Sagittal sections through esophagus (e) and aorta (a) (A,B), and transverse sections through esophagus (C,D) stained for a9 (A,C) or for tenascin (B,D) are shown. a9 and tenascin are present in the muscle layers of the esophagus (thin arrows), more easily seen in the transverse views. In contrast, expression of a9 by aortic smooth muscle (thick arrows) has decreased significantly while that of tenascin remains high. The esophageal epithelium remains negative for a9. e, esophagus; a, aorta. A& x 20; C,D, x 40.
Fig. 6. Expression of a9 and tenascin in the tongue and orophatynx of an E17.5 embryo. Sagittal section through the oral cavity stained for a9 (A) or tenascin (6). Moderate expression of both proteins is evident in the skeletal muscle comprising the tongue. Tenascin expression can also be seen in the subepithelial mesenchyme of both the palate and tongue. The epithelium lining the tongue expresses a9 at this stage, while the palatal epithelium is still negative. t, tongue; p, palate; ep, epithelium. A,B, x 20.
(intestinal smooth muscle, skeletal muscle), or spa- tially discordant (hair follicle). These data suggest that although tenascin may be a biologically relevant ligand for a9-containing integrin(s), other potential ligands exist.
Onset of a9 expression in skin, smooth and skeletal
muscle tissues closely correlated with the appearance of biochemical and/or morphological evidence of tissue differentiation. Its appearance in pulmonary and vas- cular smooth muscle precursors either coincided with or soon followed the expression of a-SM actin (Skalli et al., 1987). The onset of a9 immunoreactivity in the

428 WANG ET AL
7
Fig. 7. Expression of a9 and tenascin in the skin of an E17.5 embryo. Sagittal section through the upper lip (A,B) and cross-sections through a hair germ (C,D). Sections were stained for 019 (A,C) or for tenascin (B,D). By E17.5, the follicles of the upper lip vibrissae are almost fully differen- tiated and expression of a9 has become restricted to the hair germ (thin arrows). Tenascin staining follows the dermo-epidermal junction com- pletely around nascent hair follicles (thick arrow): the hair germs and adjacent dermal papillae of the more mature vibrissae express little or no tenascin. A,B, x 10; C,D, x 40.
Fig. 8. Expression of a9 and tenascin in the choroid plexus. Sagittal sections through the choroid plexus of the fourth ventricle of an E12.5 embryonic murine brain, stained with 019 (A) or tenascin (6). The choroid plexus exhibits intense 019 immunoreactivity. No other evidence of 0.9 expression was detected in the developing murine nervous system. Weak tenascin staining is restricted to the mesenchymal-epithelial interface of the choroid (arrows).
incipient smooth muscle layers of the gastrointestinal tract also paralleled that of a-smooth muscle actin. Al- though a-smooth muscle actin is not a precise marker of visceral smooth muscle differentiation, both the tim- ing and rostral-to-caudal pattern of onset of a9 immu- noreactivity suggest that the appearance of a9 in em-
bryonic visceral smooth muscle is also associated with the onset of differentiation. In skeletal muscle, a9 was not detected until the completion of myoblast migra- tion and the beginning of myotube formation. Onset of epidermal expression of a9 coincided with the develop- ment of mature, stratified tissue. Expression of a9 at

a9 IN MURINE EMBRYO 429
TABLE 1. Distribution of a9 in Developing and Adult Murine Tissues
Tissue Embryo Adult Smooth muscle
Airway Onset E 1 2 ~ 5 * * ~ ** GI Onset E12.5** ** Vascular E12.5-16.5 Large veins only"
Skin Onset E16.5*** Basal cells only*** Choroid plexus Onset E12.5*** Unknown Airway Negative **x
Tongue Onset day 17.5** ** Esophagus Negative ***
Epithelium
*** Pharyngeal Onset day 18.5*
Cornea Negative Principally in limbus** Skeletal muscle Onset E12.5-14.5*** ** Thymus Negative Negative Thyroid Negative Negative Liver Negative Hepatocytes* Pancreas Negative Negative Bone Negative Negative "Asterisks refer to intensity of expression: *=weak; ** =moderate; *** =strong.
each of these sites in the adult mouse has been previ- ously reported (Palmer et al., 1993). This suggests that a9 expression is not only a marker but also a persistent feature of the differentiated phenotype.
There were a few tissues, all epithelial, in which a9 staining was seen in the adult mouse but not during the embryonic stages studied (Table 1). For instance, the squamous epithelium lining the adult murine esopha- gus exhibited high levels of a9 immunoreactivity, al- though it remained negative for a9 through E18.5. The progressively more caudal appearance of a9 in oral and then pharyngeal epithelium during development, how- ever, paralleled the earlier pattern of smooth muscle development. We did not detect a9 immunostaining on hepatocytes or basal airway epithelial cells during ges- tation although these cells exhibited moderate to high levels of expression in the adult mouse. Murine airway epithelium does not mature until after birth (C. Bas- baum, personal communication). Similarly, maturation and stratification of the cornea occur postnatally (Chung et al., 1992); no a9 immunostaining was de- tected (at these sites) during gestation. Hence, the lack of a9 immunoreactivity in these epithelial tissues dur- ing gestation appears to reflect their continued devel- opment during very late stages of embryogenesis and postnatally, as well. Finally, the distribution of a9 in the adult murine central nervous system is unknown. During embryogenesis, a9 immunostaining was seen only in the specialized epithelium of the choroid plexus. Here again, the appearance of a9 coincided with mor- phologic evidence of maturation and subsequently per- sisted through late stages of embryogenesis.
While the close association of a9 onset with other markers of differentiation implicates a role in regula- tion of the differentiation process, our results suggest that expression of a9 is not linked to terminal differ- entiation. Differentiation refers to expression of a rel- atively invariant phenotype associated with the spe-
cialized function of cells or tissues in contrast to terminal differentiation which is accompanied by the cessation of proliferation or cell division (Hedin et al., 1990). Visceral smooth muscle and skeletal muscle cells can retain or regain the capacity to proliferate after differentiation (Hedin et al., 1990; Hurme and Kalimo, 1992). In the skin, the restriction of a9 expres- sion to the basal cell layer and to the hair germ in hair follicles suggests that a9 expression might be limited to the basal keratinocytes that are still capable of di- viding. Finally, in the mature cornea, a9 is detected principally in the limbus and not the central cornea itself (M. Stepp, personal communication). Strikingly, the cells of the central cornea can no longer divide; the stem cells are found in the limbus. These data suggest a possible role for a9 integrin(s) in the regeneration of squamous epithelia.
Since expression of a9 occurred in conjunction with the appearance of the differentiated phenotype, it is not likely that signals initiated via a9-containing inte- grin(s) contribute to early tissue morphogenesis. The persistence of a9 expression following its initial ap- pearance in most tissues examined further suggests that or9-mediated interactions are involved in the maintenance rather than formation of organized struc- tures. One notable exception to this pattern was vas- cular smooth muscle. As in other smooth muscle, onset of a9 expression in aortic smooth muscle was associ- ated with the appearance of a-SM actin expression but by E17.5, a9 immunostaining in the aorta had declined from moderate to near background levels. No a9 ex- pression is seen in aortic smooth muscle of adult mice. The dynamic expression pattern of a9 in vascular smooth muscle suggests a possible role for a9 inte- grinb) during vasculogenesis.
In some tissues where expression of a9 and tenascin overlapped, distinct differences in expression and local- ization were found. For example, in skeletal muscle,

430 WANG ET AL.
onset of tenascin expression appeared to precede the appearance of a9 while in intestinal smooth muscle, the reverse was true. Furthermore, tenascin staining in skeletal muscle had clearly diminished by later stages of gestation whereas levels of a9 expression re- mained fairly constant. In the skin, a9 was detected on the lateral and apical surfaces of basal cells although tenascin expression appeared to be restricted to the basal surface. Furthermore, we find intense expression of a9 in distal adult human airway epithelium, but no tenascin immunoreactivity (Weinacker et al., 1995).
Thus, while our results support the hypothesis that tenascin is a biologically relevant ligand for a9, they suggest that other a9 binding proteins remain to be identified. Our in vitro studies have so far identified only p l as a partner for a9. Other a integrin subunits are capable of forming heterodimers with more than one p subunit. Since we examined a9-integrin expres- sion using an antibody directed against the a subunit itself, we cannot be certain whether a9pl is the only heterodimer expressed at all of the sites where a9 im- munoreactivity was detected. It is possible that at sites where a9 is expressed but tenascin is not, another a9 containing integrin heterodimer whose ligand specific- ities differ from a961 is present.
As expected, tenascin immunoreactivity was also de- tected in a number of developing murine tissues, e.g., brain and bone, that were devoid of a9 staining (Chi- quet-Ehrismann et al., 1986; Erickson, 1989; Chiquet, 1992). Detection of tenascin at sites distant for a9 ex- pression is not surprising, since it is clear that there are at least three other integrins, and at least one cell surface protein that is not an integrin, which have been identified as receptors for tenascin (Prieto et al., 1993; Sriramarao et al., 1993; Chung and Erickson, 1994). We have recently shown that transfected cells heterol- ogously expressing a9pl bind to both intact tenascin-C and to a recombinant fragment composed of its third fibronectin type 111 repeat (Yokosaki et al., 1994). At least two other av integrin-containing receptors (uvp3 and probably, avp6) (Prieto et al., 1993; Sriramarao et al., 1993) mediate cell binding and spreading via the same region. Both a2pl and a chondroitin sulfate con- taining proteoglycan have been reported to bind tena- scin via the terminal fibrinogen-like domain (Srirama- rao et al., 19931, while a cell surface form of the protein annexin I1 appears to be a receptor for the alterna- tively spliced segment of tenascin (Chung and Erick- son, 1994). Thus, tenascin expressed at sites where a9 is absent could still be detected by cells expressing these other tenascin receptors.
The gene encoding mouse tenascin-C has recently been inactivated and a strain of mice developed that is completely devoid of tenascin-C expression. These mice appear to develop and age normally, a finding that has called into question, the importance of tenascin-C in development (Saga et al., 1.992). In the present study, the onset of a9 expression occurred relatively late dur- ing development, after the process of organ formation
had begun, implying that a9-integrins do not mediate early steps in tissue morphogenesis. Thus, even at sites where ag-integrin(s) might be principally binding ten- ascin, it would not be surprising that organogenesis would be unaffected by the loss of any a9-integrinlte- nascin interaction. However, if, as we suggest, a9 in- tegrin(s) contribute to the later steps in organ matura- tion, a9 integrin(s) must be recognizing ligands other than tenascin-C in tenascin knockout mice. More de- finitive characterization of the role(s) that a9-inte- grin(s) play in development will ultimately require in- activation of the a9 gene, itself.
ACKNOWLEDGMENTS The authors thank Anita Jennings for the, prepara-
tion of the murine embryos. This work was supported in part by National Institutes of Health grants HLI A133259 and HL47412 (to D.S.), HL02765-02 (to A.W.), and GM-38276 and funds from the Mayo Foundation for Education and Research (to J.A.M.).
REFERENCES Beaulieu, J.-F., Jutras, S., Durand, J., Vachon, P.H., and Perreault, N.
(1993) Relationship between tenascin and or-smooth actin expres- sion in the developing human small intestinal mucosa. Anat. Em- bryol. 188: 149-158.
Bronner-Fraser, M., Artinger, M., Muschler, J., and Horwitz, A.F. (1992) Developmentally regulated expression of a5 integrin in avian embryos. Development 115:197-211.
Chiquet, M. (1992) Tenascin: An extracellular matrix protein in- volved in morphogenesis of epithelial organs. Kidney Int. 41:629- 31.
Chiquet-Ehrismann, R., Mackie, E.J., Pearson, C.A., and Sakakura, T. (1986) Tenascin: An extracellular matrix protein involved in tis- sue interactions during fetal development and oncogenesis. Cell 47:131-139.
Chung, C.Y., and Erickson, H.P. (1994) Cell surface annexin I1 is a high affinity receptor for the alternatively spliced segment often- ascin-C. J. Cell Biol. 126:539-548.
Chung, E.-H., Bukusoglu, G., and Zieske, J.D. (1992) Localization of corneal epithelial stem cells in the developing rat. Invest. Opthal- mol. Vis. Sci. 33:2199-2206.
Duband, J.-L., Belkin, A.M., Syfrig, J., Thiery, J.P., and Koteliansky, V.E. (1992) Expression of a1 integrin, a laminin-collagen receptor, during myogenesis and neurogenesis in the avian embryo. Devel- opment 116:585-600.
Erickson, H.P. (1989) Tenascin: An extracellular matrix protein prominent in specialized embryonic tissues and tumors. Annu. Rev. Cell. Biol. 5:71-92.
Furst, D.O., Osborn, M., and Weber, K. (1989) Myogenesis in the mouse embryo: differential onset of expression of myogeneic pro- teins and the involvement of titin in myofibril assembly. J. Cell. Biol. 109:517-527.
Hedin, U., Sjolund, M., Hultgardh-Nilsson, N., and Thyberg, J. (1990) Changes in expression and organization of smooth-muscle-specific a-actin during fibronectin mediated modulation of arterial smooth muscle cell phenotype. Differentiation 44(222-231):
Hurme, T., and Kalimo, H. (1992) Adhesion in skeletal muscle during regeneration. Musc. Nerve 15:482-489.
Hynes, R.O. (1992) Integrins: Versatility, modulation, and signaling in cell adhesion. Cell 69:ll-25.
Kaplan, E.D., and Holbrook, K.A. (1994) Dynamic expression pat- terns of tenascin, proteoglycans, and cell adhesion molecules during human hair follicle morphogenesis. Dev. Dyn. 199:141-55.
Mitchell, J., Reynolds, S., Leslie, K.O., Low, R.B., and Woodcock- Mitchell, J. (1990) Smooth muscle cell markers in developing rat lung. Am. J. Respir. Cell. Mol. Biol. 3515-523.

a9 IN MURINE EMBRYO 43 1
Muschler, J.L., and Horwitz, A.F. (1991) Down-regulation of the chicken a5pl integrin fibronectin receptor during development. De- velopment 113:327-337.
Palmer, E.L., Ruegg, C., Ferrando, R., Pytela, R., and Sheppard, D. (1993) Sequence and tissue distribution of the integrin a9 subunit, a novel partner of 81 that is widely distributed in epithelia and muscle. J. Cell Biol. 123:1289-1297.
Prieto, A.L., Edelman, G.M., and Crossin, K.L. (1993) Multiple inte- grins mediate cell attachment to cytotactin/tenascin. Proc. Natl. Acad. Sci. U.S.A. 90:10154-10158.
Saga, Y., Yagi, T., Ikawa, Y., Sakakura, T, and Aizawa, S. (1992) Mice develop normally without tenascin. Genes Dev. 6:1821-31.
Skalli, O., Ropraz, P., Trzeciak, A., Benzonang, G., Gillessen, D., and Gabbiani, G. (1986) A monoclonal antibody against a-smooth mus- cle actin: A new probe for smooth muscle differentiation. J. Cell Biol. 103:2787.
Skalli, O., Vandekerckhove, J., and Gabbiani, G. (1987) Actin-isoform
pattern as a marker of normal or pathological smooth-muscle and fibroblastic tissues. Differentiation 33:232.
Spector, R., and Johanson, C.E. (1989) The mammalian choroid plexus. Sci. Am. 264:68-74.
Sriramarao, P., Mendler, M., and Bourdon, M.A. (1993) Endothelial cell attachment and spreading on human tenascin is mediated by a2pl and avp3 integrins. J. Cell. Sci. 105:lOOl-1012.
ten Have-Opbroek, A.A. (1991) Lung development in the mouse em- bryo. Exp. Lung Res. 17:lll-30.
Weinacker, A., Ferrando, R., Elliott, M., Hogg, J., Balmes, J. and Sheppard, D. (1995) Distribution of integrins a$, and aspI and their known ligands, fibronectin and tenascin, in human airways. Am. J. Respir. Cell Mol. Biol. 12:547-56,
Yokosaki, Y., Palmer, E.L., Prieto, A.L., CrosBin, K.L., Bourdon, M.A., Pytela, R., and Sheppard, D. (1994) The integrin a9pl rnedi- ates cell attachment to a non-RGD site in the third fibronectin type I11 repeat of tenascin. J . Biol. Chem. 269:26691-26696.





























