Embed Size (px)
Citation preview

Journal of Surgical Oncology 2008;98:377–383
Expression of Mismatch Repair Proteins, hMLH1/hMSH2, in Non-Small Cell
Lung Cancer Tissues and Its Clinical Significance
HIDENORI KOUSO, MD,* ICHIRO YOSHINO, MD, NAOKO MIURA, MD, TOMOYOSHI TAKENAKA, MD,TARO OHBA, MD, TOMOFUMI YOHENA, MD, ATSUSHI OSOEGAWA, MD, FUMIHIRO SHOJI, MD,
AND YOSHIHIKO MAEHARA, MD
Department of Surgery and Science, Graduate School of Medical Sciences, Kyushu University, Fukuoka, Japan
Background: hMLH1 and hMSH2 have been implicated to be involved in the DNA mismatch repair (MMR) system. The purpose of this study is
to investigate the expression of hMLH1 and hMSH2 DNA MMR proteins in non-small cell lung cancer (NSCLC) tissue and to elucidate their
clinical significance.
Methods: The hMLH1 and hMSH2 protein expression was evaluated by immunohistochemistry for a consecutive series of 113 NSCLC patients.
The expressions of each protein were examined for an association with the clinicopathological variables, including genetic alterations analyzed by
high resolution fluorescent microsatellite analysis.
Results: Regarding the hMLH1 expression, the MSI-positive patients showed significantly lower scores than the MSI-negative patients. For
hMSH2 expression, the patients with a 20 or higher pack-year index (PYI) showed significantly higher scores than the patients with a PYI less
than 20. The expression status of proteins did not affect both the disease free and overall survival of the patients. No significant correlation was
observed among the scores for the proteins.
Conclusions: The expressions of hMLH1 and hMSH2 are independently regulated and play different roles in NSCLC. The genetic instability is
possibly due to the reduced expression of hMLH1 protein, and hMSH2 expression is associated with smoking status.
J. Surg. Oncol. 2008;98:377–383. � 2008 Wiley-Liss, Inc.
KEY WORDS: hMLH1; hMSH2; microsatellite instability; mismatch repair; non-small cell lung cancer
INTRODUCTION
Lung cancer is one of the most prevalent cancers in the world, and
its mortality rate is significantly associated with the consumption of
cigarettes [1]. The main factor in the process of lung carcinogenesis is
the accumulation of genetic alterations caused by the toxic compounds
contained in cigarette smoke [2]. Cigarette smoke contains approxi-
mately 4,000 chemicals including around 40 carcinogens [3], some of
which generate DNA adducts in the respiratory cells. During DNA
replication, these lesions form DNA mismatches, which have to be
repaired by various systems, otherwise resulting in genetic mutations
[4–6]. Of the various DNA repair systems, the mismatch repair
(MMR) system is essential to maintain the stability of the genome
during repeated duplication. Its primary function is to eliminate any
single-base mismatches and insertion-deletion loops that may arise
during DNA replication [7]. In clinical lung cancer, A to T conversion
of p53 mutations and K-ras codon 12 mutation are closely related to
smoking status and microsatellite instability of the genome [8,9].
Therefore, the MMR system may play a crucial role in lung
carcinogenesis.
The MMR system is composed of a few well-conserved proteins.
The essential components of the MMR system, MutS, MutL, MutH and
Uvr, were identified in Escherichia coli [10,11]. In addition, all
eukaryotic organisms, including yeast, mice and humans, have MutS
homologs and MutL homologs [12]. Genetic and biochemical studies
have indicated that MSH2, one of the MutS homolog proteins, is
required for all mismatch corrections in nuclear DNA, whereas MSH3
and MSH6, which are also MutS homolog proteins, are required for the
repair of some distinct and overlapping types of mismatched DNA
during replication [13]. These three MutS homologs make two
heterodimers, MutSa (MSH2/MSH6) and MutSb (MSH2/ MSH3),
which recognize the mismatched DNA in eukaryotic MMR [7,14,15].
Although not as well studied as the MutS homologs, the eukaryotic
MutL homologs also appear to function as heterodimers. The best
characterization of these in humans has been MutLa, a MLH1/PMS2
heterodimer, which is capable of supporting the repair initiated by
MutSa or MutSb [15–18].
The loss of this system leads to carcinogenesis by accelerating the
accumulation of mutations in the oncogenes and tumor suppressor
genes [19,20], and, in general, such a disorder of the MMR system is
detected by gains or losses of short repeat units within the microsatel-
lite sequences, which is also known as microsatellite instability (MSI)
[9]. MSI is in fact detected in various clinical malignancies, including
colorectal, stomach, endometrial, and lung malignancies [21].
Regarding the mechanisms of the dysfunction of the MMR system,
germ line mutations of the human MMR genes, such as hMLH1 and
hMSH2, cause susceptibility to hereditary nonpolyposis colon cancer
(HNPCC) [7,22,23], whereas hypermethylation of the promoters of
these genes is considered to be another cause of the loss of the MMR
system in various sporadic cancers [24–27]. The inactivation of the
MMR genes is generally associated with the loss of immunohisto-
chemical expression of the corresponding protein [28].
The hMLH1 and hMSH2 DNA MMR proteins are known to be
implicated in human cancer, with colon cancer being the most well-
studied model. However, information on the status of these two
*Correspondence to: Hidenori Kouso, MD, Department of Surgery andScience, Graduate School of Medical Sciences, Kyushu University,Maidashi 3-1-1, Higashi-ku, Fukuoka 812-8582, Japan. Fax: 81-92-642-5482. E-mail: [email protected]
Received 15 November 2007; Accepted 20 May 2008
DOI 10.1002/jso.21108
Published online 21 July 2008 in Wiley InterScience(www.interscience.wiley.com).
� 2008 Wiley-Liss, Inc.

proteins in lung cancer is limited. In this study, we investigated the
expression of hMLH1 and hMSH2 DNA MMR proteins in non-small
cell lung cancer (NSCLC) tissues and their clinical significance.
MATERIALS AND METHODS
Patients
A consecutive series of 113 patients with NSCLC who underwent
surgical resections at Kyushu University Hospital between 2000 and
2003 were analyzed in this study. The histological diagnosis of the
tumors was based on criteria of the World Health Organization [29],
and the TNM stage was determined according to criteria revised in
1997 [30]. The age of the patients ranged from 36 to 84 years (mean of
68.8 years) and included 71 men and 42 women; 72 stage I, 15 stage II,
24 stage III and 2 stage IV; 46 with a pack-year index (PYI) of less than
20, and 67 with a PYI of 20 or more; 89 adenocarcinomas, 20
squamous cell carcinomas and 4 others (Table I). Written informed
consent was obtained from each patient for this study. The institutional
review board of our university gave approval for this study.
Follow-Up of Patients
In order to investigate the overall survival and the disease free
survival of the patients who underwent complete resections, a follow-
up examination was prospectively performed every 2 months for the
first 2 years and thereafter every 3–4 months. The mean follow-up
period of the patients was 37.8 months (median, 43.4 months; range,
0.4–75.2). The follow-up included a physical examination, complete
blood count, blood chemistry, and chest radiography. The majority of
the patients underwent CT and a radionuclide bone scan twice a year, at
the time when symptoms related to recurrence appeared. For recurrent
disease, feasible treatments such as systemic chemotherapy, radio-
therapy or surgery were performed.
High Resolution Fluorescent Microsatellite
Analysis (HRFMA)
Five dinucleotide microsatellites, D2S123, D5S107, D10S197,
D11S904 and D13S175, were used as markers for the analysis of MSI
and LOH [31]. Using genomic DNA derived from the tissue
specimens, the five microsatellite sequences were amplified by
polymerase chain reaction (PCR). Oligonucleotide primers corre-
sponding to the microsatellite sequences [31] were then synthesized
and purified using high performance liquid chromatography. 50 primers
were labeled with the fluorescent compound, ROX (6-carboxy-x-
rhodamine) or HEX (6-carboxy-20,40,70,4,7-hexachloro-fluorescein).
PCR reactions were performed using TAKARA Taq Reagent Kits
(TAKARA Co. Ltd., Tokyo, Japan) and analyzed using the Perkin–
Elmer GeneAmp PCR system 9600 or 2400 (Norwalk, USA). Each
50 ml reaction mixture contained 1� reaction buffer, 350 mM of each
dNTP, 10 pmol of each primer, 2.5 U of the polymerase, and 25 mg of
the genomic DNA. The thermal conditions of the system were as
follows: one cycle at 958C for 4 min, 35 cycles at 958C for 0.5 min,
558C for 0.5 min, 728C for 0.5 min, one cycle at 728C for 10 min. Next,
0.5 U of T4 DNA polymerase was added to the mixture, followed by
incubation at 378C for 10 min. Each 1.5 ml product was mixed with
0.5 ml loading buffer (blue dextran, 25 mM EDTA), 2.5 ml of
formamide, and 0.5 ml of dH2O. In order to compare the
electrophoretic profiles of the two samples, 1.2 ml of ROX-labeled
product and 0.3 ml of HEX-labeled product were mixed together. The
samples were denatured and loaded onto the ABI 373A sequencer
(Applied Biosystems, Foster City, CA). In each case, a size marker
labeled with TMRA (N,N,N0,N0-tetramethyl-6-carboxyrhodamine) was
always electrophoresed in each lane in order to standardize the
mobility of the sample. The running conditions were 1500 V, 20 mA,
and 30 W for 5.5 h. The data were processed using the ABI software
package from GeneScan (Applied Biosystems). The method of
Journal of Surgical Oncology
TABLE I. Demographics of the Patients in This Study
Factors Number of cases
Age
Median 66.8 years
Range 36–84
Sex
Male 71
Female 42
Pack-year indexa
Median 33.9
Range 0–156
20 or more 67
Under 20 46
Tumor histology
Adenocarcinoma 89
Squamous cell carcinoma 20
Others 4
Pathological stage
IA 49
IB 23
II 15
III 24
IV 2
MSI
Positive 9
Negative 103
LOH
Positive 61
Negative 52
aCalculated by the number of packs of cigarettes consumed a day� years.
Fig. 1. Analysis of microsatellite instability and loss of hetero-zygosity by high resolution fluorescent system. (A) A case judgedpositive for microsatellite instability. (B) A case judged positive forloss of heterozygosity.
378 Kouso et al.
detecting MSI and LOH used here has been described previously [31].
Briefly, for the detection of MSI, the y-axis corresponds to length of
DNA fragment and fluorescence of y-axis is longer than that of the
normal control when the MSI occurred in the amplified region of the
genomic DNA derived from the tumor tissue. For the detection of
LOH, the fluorescence of a peak is decreased more than 30% of that of
the normal control when the LOH occurred (Fig. 1). MSI in at least one
locus was detected in 9 cases and LOH in at least 1 locus was detected
in 61 cases (Table I).
Immunohistochemistry
An immunohistochemical study was carried out on the formalin-
fixed, paraffin-embedded tissue sections. Four-micrometer sections
were deparaffinized with xylene and then rehydrated in a series of
ethanols. Heat-induced epitope retrieval was performed in 0.1 M of
NaOH-citrate buffer (pH 7.0) for hMLH1 and hMSH2 immunostain-
ing, and the samples were heated in an autoclave at 1218C for 15 min.
Endogenous peroxidase was blocked at room temperature using 3%
hydrogen peroxide in methanol for 30 min. After blocking with normal
goat serum, the slides were incubated with mouse monoclonal
antibody against hMLH1 (BD Bioscience, Franklin Lakes, NJ) and
hMSH2 (CALBIOCHEM, Germany) using a dilution of 1:50 and
1:200 at 48C overnight, respectively. After washing the sections, they
were treated for 60 min at room temperature with goat-antimouse
immunoglobulin. Staining for hMLH1 and hMSH2 was completed
using the streptavidin–biotin–peroxidase complex method (DAKO,
Denmark) with diaminobenzidine as a chromogen, and then the slides
were counterstained with hematoxylin. The normal staining pattern for
both hMLH1 and hMSH2 was localized at the nuclear portion. The
tumor cells that exhibited an absence of nuclear staining in the
presence of non-neoplastic cells with nuclear staining (with normal
bronchial epithelium as an internal control) were considered to have an
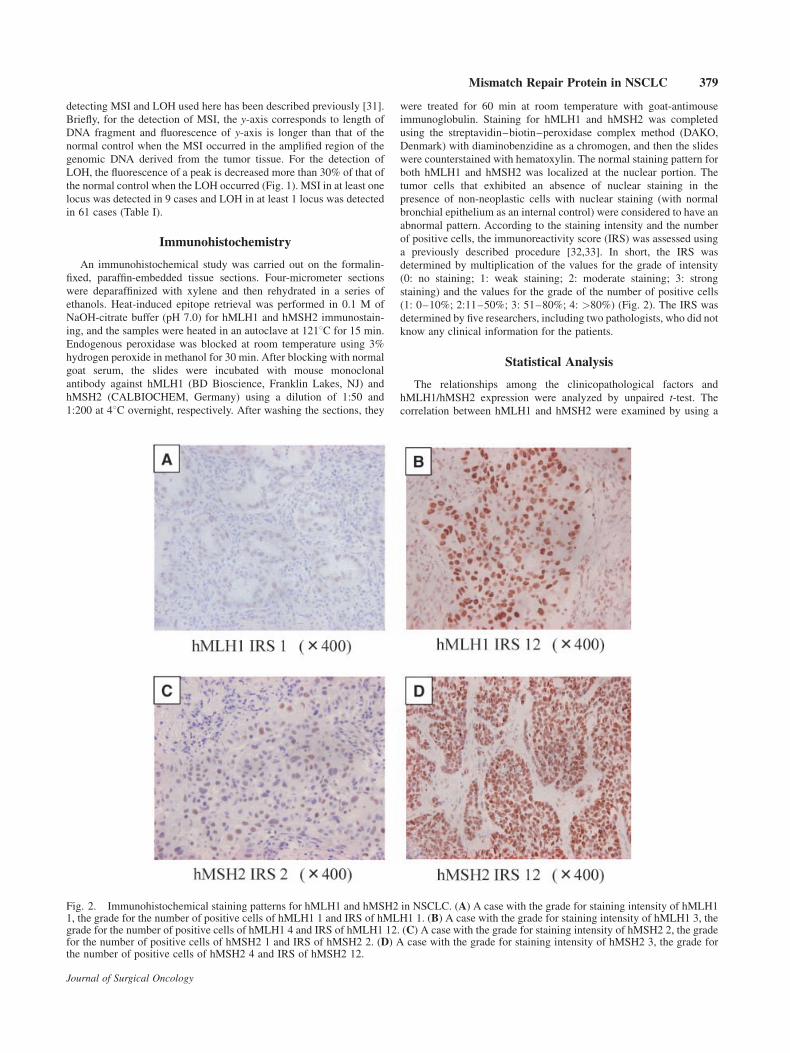
abnormal pattern. According to the staining intensity and the number
of positive cells, the immunoreactivity score (IRS) was assessed using
a previously described procedure [32,33]. In short, the IRS was
determined by multiplication of the values for the grade of intensity
(0: no staining; 1: weak staining; 2: moderate staining; 3: strong
staining) and the values for the grade of the number of positive cells
(1: 0–10%; 2:11–50%; 3: 51–80%; 4: >80%) (Fig. 2). The IRS was
determined by five researchers, including two pathologists, who did not
know any clinical information for the patients.
Statistical Analysis
The relationships among the clinicopathological factors and
hMLH1/hMSH2 expression were analyzed by unpaired t-test. The
correlation between hMLH1 and hMSH2 were examined by using a
Journal of Surgical Oncology
Fig. 2. Immunohistochemical staining patterns for hMLH1 and hMSH2 in NSCLC. (A) A case with the grade for staining intensity of hMLH11, the grade for the number of positive cells of hMLH1 1 and IRS of hMLH1 1. (B) A case with the grade for staining intensity of hMLH1 3, thegrade for the number of positive cells of hMLH1 4 and IRS of hMLH1 12. (C) A case with the grade for staining intensity of hMSH2 2, the gradefor the number of positive cells of hMSH2 1 and IRS of hMSH2 2. (D) A case with the grade for staining intensity of hMSH2 3, the grade forthe number of positive cells of hMSH2 4 and IRS of hMSH2 12.
Mismatch Repair Protein in NSCLC 379
single regression analysis. The survival curves were prepared by the
Kaplan–Meier method, and comparisons among the survival curves
were made using the log rank test. The data were considered to be
significant when the P value was less than 0.05. All of the analyses were
performed with using the Stat View software program, version 5.0.
RESULTS
Relationship Between hMLH1/hMSH2 Protein
Expression and Clinicopathological Factors
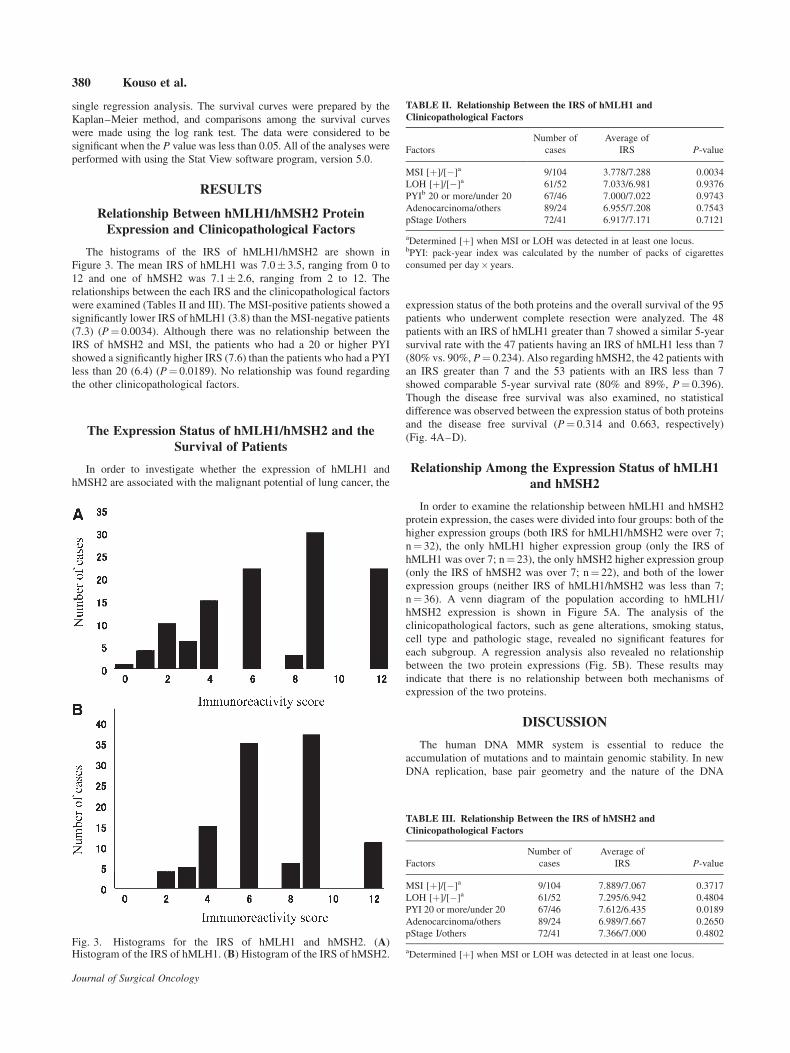
The histograms of the IRS of hMLH1/hMSH2 are shown in
Figure 3. The mean IRS of hMLH1 was 7.0� 3.5, ranging from 0 to
12 and one of hMSH2 was 7.1� 2.6, ranging from 2 to 12. The
relationships between the each IRS and the clinicopathological factors
were examined (Tables II and III). The MSI-positive patients showed a
significantly lower IRS of hMLH1 (3.8) than the MSI-negative patients
(7.3) (P¼ 0.0034). Although there was no relationship between the
IRS of hMSH2 and MSI, the patients who had a 20 or higher PYI
showed a significantly higher IRS (7.6) than the patients who had a PYI
less than 20 (6.4) (P¼ 0.0189). No relationship was found regarding
the other clinicopathological factors.
The Expression Status of hMLH1/hMSH2 and the
Survival of Patients
In order to investigate whether the expression of hMLH1 and
hMSH2 are associated with the malignant potential of lung cancer, the
expression status of the both proteins and the overall survival of the 95
patients who underwent complete resection were analyzed. The 48
patients with an IRS of hMLH1 greater than 7 showed a similar 5-year
survival rate with the 47 patients having an IRS of hMLH1 less than 7
(80% vs. 90%, P¼ 0.234). Also regarding hMSH2, the 42 patients with
an IRS greater than 7 and the 53 patients with an IRS less than 7
showed comparable 5-year survival rate (80% and 89%, P¼ 0.396).
Though the disease free survival was also examined, no statistical
difference was observed between the expression status of both proteins
and the disease free survival (P¼ 0.314 and 0.663, respectively)
(Fig. 4A–D).
Relationship Among the Expression Status of hMLH1
and hMSH2
In order to examine the relationship between hMLH1 and hMSH2
protein expression, the cases were divided into four groups: both of the
higher expression groups (both IRS for hMLH1/hMSH2 were over 7;
n¼ 32), the only hMLH1 higher expression group (only the IRS of
hMLH1 was over 7; n¼ 23), the only hMSH2 higher expression group
(only the IRS of hMSH2 was over 7; n¼ 22), and both of the lower
expression groups (neither IRS of hMLH1/hMSH2 was less than 7;
n¼ 36). A venn diagram of the population according to hMLH1/
hMSH2 expression is shown in Figure 5A. The analysis of the
clinicopathological factors, such as gene alterations, smoking status,
cell type and pathologic stage, revealed no significant features for
each subgroup. A regression analysis also revealed no relationship
between the two protein expressions (Fig. 5B). These results may
indicate that there is no relationship between both mechanisms of
expression of the two proteins.
DISCUSSION
The human DNA MMR system is essential to reduce the
accumulation of mutations and to maintain genomic stability. In new
DNA replication, base pair geometry and the nature of the DNA
Journal of Surgical Oncology
Fig. 3. Histograms for the IRS of hMLH1 and hMSH2. (A)Histogram of the IRS of hMLH1. (B) Histogram of the IRS of hMSH2.
TABLE II. Relationship Between the IRS of hMLH1 and
Clinicopathological Factors
Factors
Number of
cases
Average of
IRS P-value
MSI [þ]/[�]a 9/104 3.778/7.288 0.0034
LOH [þ]/[�]a 61/52 7.033/6.981 0.9376
PYIb 20 or more/under 20 67/46 7.000/7.022 0.9743
Adenocarcinoma/others 89/24 6.955/7.208 0.7543
pStage I/others 72/41 6.917/7.171 0.7121
aDetermined [þ] when MSI or LOH was detected in at least one locus.bPYI: pack-year index was calculated by the number of packs of cigarettes
consumed per day� years.
TABLE III. Relationship Between the IRS of hMSH2 and
Clinicopathological Factors
Factors
Number of
cases
Average of
IRS P-value
MSI [þ]/[�]a 9/104 7.889/7.067 0.3717
LOH [þ]/[�]a 61/52 7.295/6.942 0.4804
PYI 20 or more/under 20 67/46 7.612/6.435 0.0189
Adenocarcinoma/others 89/24 6.989/7.667 0.2650
pStage I/others 72/41 7.366/7.000 0.4802
aDetermined [þ] when MSI or LOH was detected in at least one locus.
380 Kouso et al.

polymerase involved result in an error rate of 10�4 to 10�5 at the
nucleotide insertion step of DNA synthesis [34], and the proofreading
exonuclease associated with some DNA polymerases edits this
mistake. Mistakes that escape these fidelity devices are corrected by
the MMR system, further elevating the fidelity by 50–1,000-fold [15].
These replication errors are particularly evident in the microsatellite
sequences, consisting of repeats of 1–4 base pairs. If the MMR system
is inactivated, the errors are fixed as a mutation, such as the addition or
deletion of one or more repeat units, after another next round of
replication. These altered microsatellite sequences are called micro-
satellite instability and are considered to reflect a cellular MMR
deficiency [35].
In the ubiquitous process of MMR, the hMLH1 and hMSH2
proteins have different roles but act serially. The hMSH2 protein
works to recognize mismatched DNA, whereas the hMLH1 protein
coordinates the interplay among the mismatch recognition complex
and other involved proteins [7]. However, in clinical cancer, these
protein expressions are not correlated well with each other. Xinarianos
et al. [36] reported that the comparative analysis of hMLH1 and
hMSH2 protein expression in the NSCLC tissue did not reveal any
associations between the expressions of these two genes, and that
the tumor specimens with a combined reduced expression of both
the hMLH1 and hMSH2 proteins did not correlate with any of the
clinicopathological parameters. In this study, we examined the
expression status of both the hMLH1 and hMSH2 in 113 surgical
specimens of NSCLC, and examined them for a relationship with
the clinicopathological parameters and microsatellite alterations of the
DNA. The expressions of the hMLH1 and hMSH2 proteins were
independent of an association with smoking status with hMSH2 but not
with hMLH1, which was determined by a regression analysis among
the IRSs of the two proteins. Moreover, only the hMLH1 expression
showed a significant relationship with the presence of MSI but not of
hMSH2. It is thus likely that the clinical significance of the hMLH1
and hMSH2 expressions is quite different in lung cancers.
In several types of malignancies, the protein expressions of hMLH1
and/or hMSH2 have been demonstrated to be significantly associated
with a DNA mismatch repair deficiency and MSI in the stomach,
endometrial and lung [37–42]. However, which of the proteins is
essential to the presence of MSI remains to be clarified. In this study,
the IRS of hMLH1 was expectedly lower in the MSI-positive cases
(P¼ 0.0034), whereas there was no statistical difference in the
relationship between hMSH2 and MSI. Chang et al. [42] also reported
a relationship to exist between MSI and a reduced hMLH1 protein
expression, but not with hMSH2 protein expression in NSCLC. In
an experiment of cancer cell lines with MSI, the reversal of
methylation by a demethylating agent, 5-azacytidine, not only induced
Journal of Surgical Oncology
Fig. 4. Correlation of the IRS of hMLH1 and hMSH2 with the prognosis of patients. No statistical difference was observed between theexpression status of hMLH1/hMSH2 and both the disease free and overall survival.
Mismatch Repair Protein in NSCLC 381

the reexpression of the hMLH1 protein, but also restored the MMR
capacity in the MMR-deficient cell line [24,25]. These results suggest
that isolated hMLH1 deficiency leads to MSI.
Although germ line mutations in the MMR genes causes MSI in
HNPCC and sporadic colorectal cancer with MSI, there are a subset of
sporadic colorectal cancers with MSI which have been identified as
having no mutations of the MMR genes [43]. The hypermethylation of
the promoter of hMLH1 also causes the inactivation of the hMLH1
protein expression, and consequently, MSI is caused in most sporadic
cancers including colorectal, stomach, and ovarian carcinoma [24–26].
In NSCLC, hypermethylation of the promoter of the hMLH1 gene was
reported to be significantly associated with the reduction in protein
expression [27]. This report also indicated a significant relationship
between MSI and hypermethylation of the promoter of hMLH1.
Hence, in sporadic cancers, hypermethylation of the promoter of
hMLH1 causes alteration of its protein expression and MSI. Therefore,
hypermethylation of the promoter of hMLH1 may be a meaningful
factor for carcinogenesis. In NSCLC, the regulation of hMLH1 protein
expression, possibly through gene silencing, may have a valuable role
in lung carcinogenesis.
It should be noted that the IRS of hMSH2 was higher in the over-20
PYI cases than in the less-than-20 PYI cases (P¼ 0.0189). The
chemical compounds included in tobacco smoke, such as aromatic
hydrocarbons, frequently cause genetic alterations through the
formation of DNA adducts [44]. On the other hand, inactivation of
the MMR genes promotes carcinogenesis by accelerating the
accumulation of mutations in many oncogenes and tumor suppressor
genes, including p53, K-ras, APC, TGFbIIR, and BAX [20,45–48].
In clinical lung cancer, A to T conversion of p53 mutations and K-ras
codon 12 mutation are closely related to smoking status and an
unfavorable prognosis [8,9]. The MMR system may play a significant
role in overcoming the crisis of the respiratory epithelial cells;
otherwise, the reduced expression of the MMR proteins may result in a
susceptibility to lung carcinogenesis. The present results showed the
close relationship between the hMSH2 expression in lung cancer
tissues and the smoking status of the corresponding patients, and
supported the above hypothesis.
Scartozzi et al. [49] demonstrated that a lower expression of
hMLH1 in the tumor cells was associated with a favorable survival in
patients with NSCLC. However, Skarda et al. [50] reported no
relationships to exist between the altered protein expression of
hMLH1/hMSH2 and both the disease free and overall survival in
NSCLC patients. In this study, the expression status of both hMLH1
and hMSH2 showed no significant impact on the disease free survival
or overall survival of patients. In the present cohort, the mean follow-
up period of the patients was 37.8 months and 72 patients (64%) were
stage I; therefore, it may be difficult to detect survival differences.
CONCLUSION
The genetic instability of NSCLC is due to the reduced expression
of the hMLH1 protein. The hMSH2 expression is increased by
smoking stimulation, and it is associated with the repair of gene
damage due to smoking-related chemical substances. Further investi-
gation is required to elucidate the complete pathways of activation and
inactivation of the expression of these two proteins in NSCLC.
REFERENCES
1. World Health Organization. World Health Statistics Annual.Geneva: WHO; 1991.
2. Phillips DH: Smoking-related DNA and protein adducts in humantissues. Carcinogenesis 2002;23:1979–2004.
3. Hoffmann D, Hecht SS: Advances in tobacco carcinogenesis. In:Cooper CS, Grover PL, editors. Chemical Carcinogenesis andMutagenesis I, Springer-Verlag Berlin: 1990; 63–102.
4. Doherty KM, Sharma S, Uzdilla LA, et al.: RECQ1 helicaseinteracts with human mismatch repair factors that regulate geneticrecombination. J Biol Chem 2005;280:28085–28094.
5. Elliott B, Jasin M: Repair of double-strand breaks by homologousrecombination in mismatch repair-defective mammalian cells.Mol Cell Biol 2001;21:2671–2682.
6. Mu D, Tursun M, Duckett DR, et al.: Recognition and repair ofcompound DNA lesions (base damage and mismatch) by humanmismatch repair and excision repair systems. Mol Cell Biol1997;17:760–769.
7. Peltomaki P: Role of DNA mismatch repair defects in thepathogenesis of human cancer. J Clin Oncol 2003;21:1174–1179.
8. Levine AJ, Momand J, Finlay CA: The p53 tumour suppressorgene. Nature 1991;351:453–456.
9. Slebos RJ, Hruban RH, Dalesio O, et al.: Relationship between K-ras oncogene activation and smoking in adenocarcinoma of thehuman lung. J Natl Cancer Inst 1991;83:1024–1027.
10. Modrich P, Lahue R: Mismatch repair in replication fidelity,genetic recombination, and cancer biology. Annu Rev Biochem1996;65:101–133.
11. Wang H, Yang Y, Schofield MJ, et al.: DNA bending andunbending by MutS govern mismatch recognition and specificity.Proc Natl Acad Sci U S A 2003;100:14822–14827.
12. Jun SH, Kim TG, Ban C: DNA mismatch repair system. Classicaland fresh roles. FEBS J 2006;273:1609–1619.
13. Marsischky GT, Filosi N, Kane MF, et al.: Redundancy ofSaccharomyces cerevisiae MSH3 and MSH6 in MSH2-dependentmismatch repair. Genes Dev 1996;10:407–420.
Journal of Surgical Oncology
Fig. 5. Relationship between hMLH1 and hMSH2 protein expression(A) A venn figure of the population according to hMLH1/hMSH2expression. (B) A dot plot analysis for hMLH1/hMSH2. There was norelationship between the expression of two proteins.
382 Kouso et al.

14. Jascur T, Boland CR: Structure and function of the components ofthe human DNA mismatch repair system. Int J Cancer 2006;119:2030–2035.
15. Iyer RR, Pluciennik A, Burdett V, et al.: DNA mismatch repair:functions and mechanisms. Chem Rev 2006;106:302–323.
16. Li GM, Modrich P: Restoration of mismatch repair to nuclearextracts of H6 colorectal tumor cells by a heterodimer of humanMutL homologs. Proc Natl Acad Sci U S A 1995;92:1950–1954.
17. Fishel R, Lescoe MK, Rao MR, et al.: The human mutator genehomolog MSH2 and its association with hereditary nonpolyposiscolon cancer. Cell 1993;75:1027–1038.
18. Bronner CE, Baker SM, Morrison PT, et al.: Mutation in the DNAmismatch repair gene homologue hMLH1 is associated withhereditary non-polyposis colon cancer. Nature 1994;368:258–261.
19. Eshleman JR, Markowitz SD: Mismatch repair defects in humancarcinogenesis. Hum Mol Genet 1996;5:1489–1494.
20. Kinzler KW, Vogelstein B: Lessons from hereditary colorectalcancer. Cell 1996;87:159–170.
21. Lawes DA, SenGupta S, Boulos PB: The clinical importance andprognostic implications of microsatellite instability in sporadiccancer. Eur J Surg Oncol 2003;29:201–212.
22. Han HJ, Maruyama M, Baba S, et al.: Genomic structure ofhuman mismatch repair gene, hMLH1, and its mutation analysisin patients with hereditary non-polyposis colorectal cancer(HNPCC). Hum Mol Genet 1995;4:237–242.
23. Liu B, Parsons RE, Hamilton SR, et al.: hMSH2 mutations inhereditary nonpolyposis colorectal cancer kindreds. Cancer Res1994;54:4590–4594.
24. Herman JG, Umar A, Polyak K, et al.: Incidence and functionalconsequences of hMLH1 promoter hypermethylation in color-ectal carcinoma. Proc Natl Acad Sci U S A 1998;95:6870–6875.
25. Leung SY, Yuen ST, Chung LP, et al.: hMLH1 promotermethylation and lack of hMLH1 expression in sporadic gastriccarcinomas with high-frequency microsatellite instability. CancerRes 1999;59:159–164.
26. Geisler JP, Goodheart MJ, Sood AK, et al.: Mismatch repair geneexpression defects contribute to microsatellite instability inovarian carcinoma. Cancer 2003;98:2199–2206.
27. Wang YC, Lu YP, Tseng RC, et al.: Inactivation of hMLH1 andhMSH2 by promoter methylation in primary non-small cell lungtumors and matched sputum samples. J Clin Invest 2003;111:887–895.
28. Marcus VA, Madlensky L, Gryfe R, et al.: Immunohistochemistryfor hMLH1 and hMH2: A practical test for DNA mismatch repair-deficient tumors. Am J Surg Pathol 1999;23:1248–1255.
29. World Health Organization. Histological Typing of Lung andPleural Tumors. (3 ed). Springer-Verlag Geneva: 1999.
30. Mountain CF: Revisions in the International System for StagingLung Cancer. Chest 1997;111:1710–1717.
31. Oda S, Oki E, Maehara Y, et al.: Precise assessment ofmicrosatellite instability using high resolution fluorescent micro-satellite analysis. Nucleic Acids Res 1997;25:3415–3420.
32. Friedrich M, Villena-Heinsen C, Meyberg R, et al.: Immunohis-tochemical analysis of DNA ’mismatch-repair’ enzyme humanMut-S-Homologon-2 in ovarian carcinomas. Histochem J 1999;31:717–722.
33. Friedrich M, Meyberg R, Villena-Heinsen C, et al.: Immunohis-tochemical analysis of DNA mismatch-repair enzyme hMSH-2
and Ki-67 in breast carcinomas. Anticancer Res 1999;19:3349–3353.
34. Kunkel TA: DNA replication fidelity. J. Biol Chem 2004;279:16895–16898.
35. Boland CR, Thibodeau SN, Hamilton SR, et al.: A NationalCancer Institute Workshop on Microsatellite Instability for cancerdetection and familial predisposition: development of interna-tional criteria for the determination of microsatellite instability incolorectal cancer. Cancer Res 1998;58:5248–5257.
36. Xinarianos G, Liloglou T, Prime W, et al.: hMLH1 and hMSH2expression correlates with allelic imbalance on chromosome 3p innon-small cell lung carcinomas. Cancer Res 2000;60:4216–4221.
37. Lanza G, Gafa R, Maestri I, et al.: Immunohistochemical patternof MLH1/MSH2 expression is related to clinical and pathologicalfeatures in colorectal adenocarcinomas with microsatelliteinstability. Mod Pathol 2002;15:741–749.
38. Staebler A, Lax SF, Ellenson LH: Altered expression of hMLH1and hMSH2 protein in endometrial carcinomas with micro-satellite instability. Hum Pathol 2000;31:354–358.
39. Rosen DG, Cai KQ, Luthra R, et al.: Immunohistochemicalstaining of hMLH1 and hMSH2 reflects microsatellite instabilitystatus in ovarian carcinoma. Mod Pathol 2006;19:1414–1420.
40. Wilentz RE, Goggins M, Redston M, et al.: Genetic, immuno-histochemical, and clinical features of medullary carcinoma of thepancreas: A newly described and characterized entity. Am JPathol 2000;156:1641–1651.
41. Kawaguchi K, Oda Y, Takahira T, et al.: Microsatellite instabilityand hMLH1 and hMSH2 expression analysis in soft tissuesarcomas. Oncol Rep 2005;13:241–246.
42. Chang JW, Chen YC, Chen CY, et al.: Correlation of geneticinstability with mismatch repair protein expression and p53mutations in non-small cell lung cancer. Clin Cancer Res 2000;6:1639–1646.
43. Liu B, Nicolaides NC, Markowitz S, et al.: Mismatch repair genedefects in sporadic colorectal cancers with microsatelliteinstability. Nat Genet 1995;9:48–55.
44. Polynuclear aromatic compounds. General remarks on thesubstances considered. IARC Monogr Eval Carcinog Risk ChemHum 1983;32:33–91.
45. Konishi M, Kikuchi-Yanoshita R, Tanaka K, et al.: Molecularnature of colon tumors in hereditary nonpolyposis colon cancer,familial polyposis, and sporadic colon cancer. Gastroenterology1996;111:307–317.
46. Losi L, Ponz de Leon M, Jiricny J, et al.: K-ras and p53 mutationsin hereditary non-polyposis colorectal cancers. Int J Cancer1997;74:94–96.
47. Furukawa T, Konishi F, Shitoh K, et al.: Evaluation of screeningstrategy for detecting hereditary nonpolyposis colorectal carci-noma. Cancer 2002;94:911–920.
48. Fujiwara T, Stolker JM, Watanabe T, et al.: Accumulated clonalgenetic alterations in familial and sporadic colorectal carcinomaswith widespread instability in microsatellite sequences. Am JPathol 1998;153:1063–1078.
49. Scartozzi M, Franciosi V, Campanini N, et al.: Mismatch repairsystem (MMR) status correlates with response and survival innon-small cell lung cancer (NSCLC) patients. Lung Cancer 2006;53:103–109.
50. Skarda J, Fridman E, Plevova P, et al.: Prognostic value of hMLH1and hMSH2 immunohistochemical expression in non-small celllung cancer. A. tissue microarray study. Biomed Pap Med FacUniv Palacky Olomouc Czech Repub 2006;150:255–259.
Journal of Surgical Oncology
Mismatch Repair Protein in NSCLC 383

![hMLH1 and hMSH2 Expression Correlates with Allelic ... › content › canres › ... · [CANCER RESEARCH 60, 4216–4221, August 1, 2000] hMLH1 and hMSH2 Expression Correlates with](https://img.dokumen.tips/doc/110x75/5f191ad1de8c4842a60e1d57/hmlh1-and-hmsh2-expression-correlates-with-allelic-a-content-a-canres-a.jpg)



























