Embed Size (px)
Citation preview

DEVELOPMENTAL GENETICS 1442-50 (1993)
Expression of Endogenous and Microinjected hsp 30 Genes in Early Xenopus Zaeuis Embryos ADNAN ALI, PATRICK H. KRONE, AND JOHN J. HEIKKILA Department of Biology, University of Waterloo, Waterloo, Ontario, Canada
ABSTRACT In the present study, we have examined the regulation of expression of a newly isolated member of the hsp 30 gene family, hsp 30C. Using RT-PCR, we found that this gene was first heat-inducible at the tailbud stage of develop- ment. We also examined the expression of two mi- croinjected modified hsp 30C gene constructs in Xenopus embryos. One of the constructs had 404 bp of hsp 30C 5’-flanking region, whereas the other had 3.6 kb. Both gene constructs had 1 kb of 3’-flanking region. RT-PCR assays were employed to detect the expression of these microinjected genes. The presence of extensive 5‘- and 3’-flank- ing regions of the hsp 30C gene did not confer proper developmental regulation, since heat-in- ducible expression of both of the microinjected constructs was detectable at the midblastula stage. The premature expression of the microinjected hsp 30 gene was not a result of high plasmid copy number or the presence of plasmid DNA se- quences. These results suggest that the microin- jected genes contain all the cis-acting DNA sequences required for correct heat-inducible reg- ulation but do not contain the elements required for the proper regulation of hsp 30 gene expression during development. It is possible that regulatory elements controlling the developmental expression of the hsp30 genes may reside upstream or down- stream of the entire cluster. o 1993 WiIey-tiss, Inc.
Key words: Develop men t, transcription, heat shock protein, microinjection, polymerase chain re- action, Xenopus laevis, mRNA
INTRODUCTION Almost all organisms examined to date respond to
environmental stresses such as heat shock or exposure to heavy metal ions or sodium arsenite by rapidly in- creasing the expression of a set of heat shock protein genes [reviewed by Morimoto et al., 19901. In a variety of animal systems, synthesis of heat shock proteins (hsps) is developmentally regulated [Heikkila et al., 1986; Browder et al., 1989; Hightower and Nover, 19911. During early Xenopus embryogenesis, certain heat shock genes are not heat inducible until the mid-
blastula transition (MBT), a point in development that coincides with the activation of the zygotic genome [Bienz, 1984a; Heikkila et al., 19871. At this stage of development, heat shock-induced accumulation of hsp 70, hsp 87, and ubiquitin mRNAs has been observed [Bienz, 1984a; Heikkila et al., 1985,1987; Ovsenek and Heikkila, 19881. In contrast to the pattern of these hsp genes, heat shock-induced accumulation of hsp 30 mRNA was reported to first occur a t the tailbud stage of development [Krone and Heikkila, 19891.
We have previously used microinjection to examine potential cis-acting regulatory regions involved in the developmental regulation of hsp gene expression in Xe- nopus [Krone and Heikkila, 19891. It was demon- strated using a hsp 70lCAT fusion gene that all of the cis-acting sequences required for correct heat-inducible activation of hsp 70 gene expression at the midblastula stage were present within the first 750 bp of promoter region. However, a microinjected hsp 3OAiCAT fusion gene also exhibited heat-inducible expression at the midblastula stage, which was earlier in development than the expression of the endogenous hsp 30 genes was first detectable. Furthermore, the addition of se- quences within the coding and 3’-regions did not confer correct developmental regulation on the microinjected hsp 30A gene. Thus, it was possible that cis-acting se- quences required for correct developmental regulation of hsp 30 gene expression lay beyond those present in the hsp 30A genomic clone used for the microinjection studies.
In order to obtain hsp 30 genes containing more ex- tensive flanking regions, a Xenopus genomic library was screened and a 14.3-kb clone containing a tandem array of hsp 30 genes was isolated [Krone et al., 19921.
Received for publication October 26, 1992; accepted November 30, 1992.
Address reprint requests to Dr. John J. Heikkila, Department of Biology, University of Waterloo, Waterloo, Ontario N2L 3G1, Canada.
Dr. Krone is now at the Department of Anatomy, College of Medicine University of Saskatchewan, Saskatoon, Saskatchewan, S7N OWO, Canada.
0 1993 WILEY-LISS, INC.
HSP 30 GENE EXPRESSION IN XEIVOPUS EMBRYOS 43
This clone contained 2 complete hsp 30 genes (hsp 30C and hsp 30D) as well as a portion of a third gene (hsp 30E). Thus, extensive flanking regions of the hsp 30C gene, the central gene in the cluster, were available for the construction of microinjection vectors. In the present study, we have used the highly sensitive tech- nique of reverse transcription-polymerase chain reac- tion (RT-PCR) to compare the expression of these new microinjection vectors with the endogenous hsp 30C gene during early development. We demonstrate that the hsp 30C gene is regulated in the same manner as the hsp 30A gene in that heat-inducible expression is first detectable at the tailbud stage of development. Furthermore, the presence of extensive flanking re- gions does not confer correct developmental regulation on the microinjected hsp 30C gene. Taken together with our previous results obtained using the hsp 30A gene, it is possible that expression of the hsp 30 gene family is repressed prior to the tailbud stage and that the repression mechanism probably does not involve inhibitory promoter elements.
MATERIALS AND METHODS Embryo Manipulation and Microinjections
Xenopus eggs were obtained, fertilized and dejellied by the methods described in Heikkila et al. [19851. The developmental stages of the embryos were determined according to Nieuwkoop and Faber [19671. Embryos at the one to two-cell-stage were used in the microinjec- tion experiments. Unless otherwise stated, embryos were microinjected with gel purified supercoiled plas- mid DNA. Twenty nl of the DNA was microinjected into the animal pole of the embryos using the appara- tus described by Hitchcock and Friedman [19801. Fol- lowing microinjection, embryos were maintained a t 22°C in Steinberg’s solution containing 4% Ficoll until stage 7, at which point they were transferred to Stein- berg’s solution containing 2% Ficoll [Krieg and Melton, 19851. Following stage 8, the microinjected embryos were kept in Steinberg’s solution without Ficoll.
Heat shock experiments were performed with the embryos (injected or noninjected) a t selected stages of development. They were maintained at 22°C or incu- bated for 1 h in a petri dish submerged in a waterbath which had been equilibrated to 33°C. Only embryos which appeared normal were selected and used for RNA isolations.
RNA Isolation Total lithium chloride precipitable RNA was isolated
from embryos by the method of Auffrey and Rougeon [19801, as modified by Mohun et al. [1984]. Frozen em- bryos were homogenized on ice in 4 ml of RNA isolation buffer [3 M lithium chloride, 6 M urea, 0.5% (w/v) SDS, 70 mM 2-mercaptoethanol, 10 mM sodium acetate (pH 5.2)l per 40 embryos. Homogenates were transferred to 15-ml sterile glass Corex tubes and incubated over-
A . HSP30C
1 2 3 4 5 6
500 - 400 - 300-
200-
6 . ACTIN
1 2 3 4 5 6
500-
400-
300- c b c c
200-
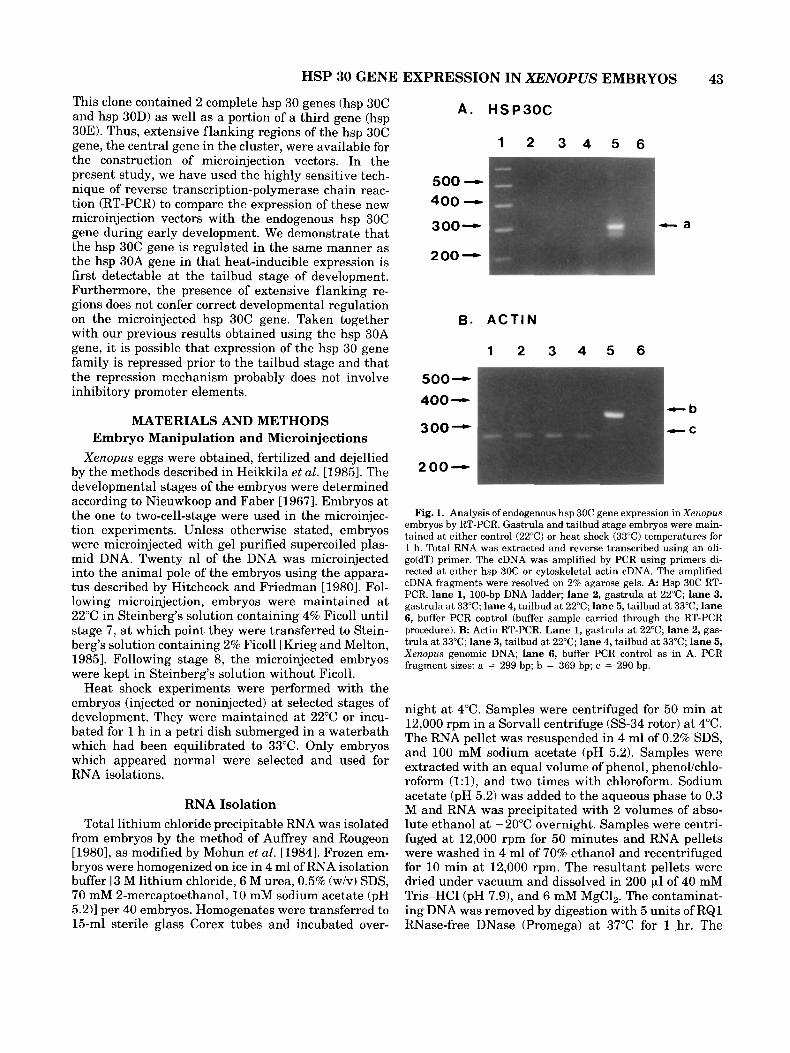
Fig. 1. Analysis of endogenous hsp 30C gene expression in Xenopus embryos by RT-PCR. Gastrula and tailbud stage embryos were main- tained at either control (22°C) or heat shock (33°C) temperatures for 1 h. Total RNA was extracted and reverse transcribed using an oli- go(dT) primer. The cDNA was amplified by PCR using primers di- rected at either hsp 30C or cytoskeletal actin cDNA. The amplified cDNA fragments were resolved on 2% agarose gels. A Hsp 30C RT- PCR. lane 1, 100-bp DNA ladder; lane 2, gastrula at 22°C; lane 3, gastrula at 33°C; lane 4, tailbud at 22°C; lane 5, tailbud at 33°C; lane 6, buffer PCR control (buffer sample carried through the RT-PCR procedure). B: Actin RT-PCR. Lane 1, gastrula a t 22°C; lane 2, gas- trula at 33°C; lane 3, tailbud at 22°C; lane 4, tailbud a t 33°C; lane 5, Xenopus genomic DNA; lane 6, buffer PCR control as in A. PCR fragment sizes: a = 299 bp; b = 369 bp; c = 290 bp.
night a t 4°C. Samples were centrifuged for 50 min at 12,000 rpm in a Sorvall centrifuge (SS-34 rotor) at 4°C. The RNA pellet was resuspended in 4 ml of 0.2% SDS, and 100 mM sodium acetate (pH 5.2). Samples were extracted with an equal volume of phenol, phenol/chlo- roform ( l : l ) , and two times with chloroform. Sodium acetate (pH 5.2) was added to the aqueous phase to 0.3 M and RNA was precipitated with 2 volumes of abso- lute ethanol at -20°C overnight. Samples were centri- fuged a t 12,000 rpm for 50 minutes and RNA pellets were washed in 4 ml of 70% ethanol and recentrifuged for 10 min at 12,000 rpm. The resultant pellets were dried under vacuum and dissolved in 200 pl of 40 mM Tr isHCl (pH 7.91, and 6 mM MgC1,. The contaminat- ing DNA was removed by digestion with 5 units of RQ1 RNase-free DNase (Promega) at 37°C for 1 hr. The

44 ALIETAL.
hsp30 gene cluster (1 4.3 kb)
I I T I I I I I I I I I I I I I + + +
hsp3OC hsp30D hsp30E
Fig. 2. Restriction enzyme map of the hsp 30 gene cluster. The coding regions are represented as thick solid bars. The direction of transcription is indicated by arrows below the respective genes.
RNA was then extracted once with phenol/chloroform (1:l) and once with chloroform and then precipitated as described above. The RNA pellet was resuspended in 100 pl of sterile distilled water that had been treated with 0.1% DEPC (diethylpyrocarbonate).
Construction of Modified hsp 30C Gene Vectors A 1.0-kb EcoRI/HindIII genomic fragment contain-
ing the 3'-end of the hsp 30C gene was filled in with the Klenow fragment of DNA polymerase I and blunt end ligated into the HincII site of the pUC19 plasmid. The resultant hsp 3OC/pUC19 construct was used to make two additional constructs both of which contained the hsp 30C coding region but differed in the length of the promoter region. A 1.1-kb BamHIIEcoRI genomic frag- ment, containing 404 bp of promoter region and 730 bp of hsp 30C coding region, was gel purified, filled in and blunt end ligated into the SmaI site of the hsp 30C/ pUC19 construct to get the plasmid p3OCplyl. This procedure left a 17-bp pUC19 polylinker separating the coding region and the 3'-flanking region of the hsp 30C gene. Similarly, another plasmid p3OCply2 was made by inserting a 4.3-kb EcoRI fragment into the EcoRI site of the hsp 3OC/pUC19 construct. In this case, the promoter region was 3.6 kb long, and a 30-bp pUC19 polylinker region separated the coding region and the 3'-flanking region of the gene.
Reverse Transcription Reverse transcription reactions were performed es-
sentially as described in Rappolee et al. [19891. One pg of total RNA was dissolved in 5 ~1 of sterile distilled water, treated with RNA Guard (40 U/pl; Pharmacia) a t a concentration of 80 U/ml. Five pl of Rt mix #1 [0.2 pl of oligo (dT) primer (1 mg/ml; Pharmacia), 1.0 pl of 10 mM dNTPs (100 mM stock diluted in RNA Guard- treated water; Pharmacia), 0.5 pl of 2 mg/ml BSA (10 mg/ml; Pharmacia), 0.1 pl of 1 M DTT, 2.0 p1 of 5 x Rt buffer [250 mM Tris-HC1 (pH 8.3 at 43"C), 300 mM KC1, 15 mM MgCl,, 50 mM DTTI, 0.125 p1 of RNA guard (40 U/pl), and 24 units of AMV reverse tran- scriptase (25 U/pl, Promega)] were added to each sam- ple of RNA for a total volume of 10 pl in a 0.5-ml
microcentrifuge tube. The contents were mixed and in- cubated a t 43°C for 1 hr. The tubes were then trans- ferred to 95°C for 3 min for heat denaturation of the complexes and then cooled on ice for few minutes. The samples were then mixed with 2.7 pl of Rt mix #2 (0.1 pl of 10 mM dNTPs, 0.5 p1 of BSA, 0.4 p1 of 5 x Rt buffer, 15 units of AMV reverse transcriptase and RNA Guard treated water to the final volume of 2.8 pl). Samples were then incubated at 43°C for 1 h: Seven- teen pl of sterile distilled water was added and the cDNA product was aliquoted and stored a t -20°C until needed.
Polymerase Chain Reaction Polymerase chain reaction (PCR) was performed as
described in Rappolee et al. [19891 and Rappolee 119901. The sequences of the primers were based on hsp 30C gene [Krone et al., 19921 and actin gene (type 5) se- quences [Mohun and Garrett, 19871. The downstream primer for the modified hsp 30C gene was the comple- mentary sequence of the polylinker region of the pUC19 plasmid. These primers were purchased from the Department of Genetics, University of Guelph, and are listed below:
5'-hsp 30C primer: 5'-CGACAGTCCAGAGAAAGT-
3'-hsp 30C primer: 5'-TCGAGGAGCCTGAATATG-
Expected length of the amplified hsp 30C fragment
5'-Actin primer: 5'-CATCCGTAAGGACCTGTAT-
3'-Actin primer: 5'-CAACGCATCTGCTAGCAG
Expected length of the amplified fragment from the
Expected length of the amplified fragment from actin
5'-modified hsp 30C primer: 5'-CATGAGCGGAAA-
GAGG-3'
CAGG-3'
= 299 bp
GCC-3'
TCCA-3'
actin gene (with intron) = 369 bp
cDNA = 290 bp
AGTGACACTGA-3'

HSP 30 GENE EXPRESSION IN XENOPUS EMBRYOS 45
A
730 bp Coding region
1 kb . ~ . . EcoRl , -. . .
404 bp ' 3-Flanking region 5'-Flanking region
Hindlll BsmHl -
- \ .' (17 bp Polylinkar region)
730 bp l k b Coding region ,@yA 3'-Flanking region
3.6 kb 5'-Flanking region
(30 bp Polylinkar region)
EcoRl
Fig. 3. Diagrams of the modified hsp 30C gene constructs used in the microinjection experiments. These constructs were made in a pUC19 vector. A: p30Cplyl plasmid. This construct contains a short 5'-flanking region (404 bp) upstream of the transcription initiation site. A small region of pUC19 polylinker (17 bp) separates the coding region and 3'-flanking region. B: p30Cply2 plasmid. This construct
carries 3.6 kb of 5'-flanking region of the hsp 30C gene and a 30-bp region of pUC19 polylinker that separates the coding region from 3'-flanking region of the gene. The coding region of the hsp 30C gene is shown as thick solid bar and the 3'-untranslated region as a shaded box.
3'-modified hsp 30C primer: 5'-TTAGAATTGACT
Expected length of the amplified fragment from the modified hsp 30C cDNA containing the long 5'-flank- ing region = 320 bp
Expected length of the amplified fragment from the modified hsp 30C cDNA containing the short 5'-flank- ing region = 307 bp
A dose of 1.5 p1 of each primer (5'- and 3'-) (0.35
CTAGAGGATCCC-3' pg/pl) was added to a 0.5-ml microcentrifuge tube con- taining 44 pl of PCR mix [4.8 pl of 10 x PCR buffer 100 mM Tr isHCl (pH 8.3 at 22"C), 500 mM KC1, 25 mM MgCl,, 1 mg/ml BSAI, 3.0 p1 of 10 mM dNTPs, 0.2 p1 of Taq DNA polymerase (5 U/p1; Pharmacia), and 36.1 pl of sterile distilled water. The contents were then com- bined with 3.0 p1 of cDNA, and two drops of mineral oil were added to each tube, to reduce evaporation and contamination. The samples were heat denatured a t

46 ALIETAL.
94°C for 4 min; PCR was then performed by incubating the tubes in a DNA thermal cycler (Perkin-Elmer-Ce- tus, Norwalk, PA) for 40 cycles. Each cycle had three temperature points, including denaturation at 94°C for 1 min, annealing at 55°C for 1 min, and polymerization at 72°C for 1.5 min. When 40 cycles were completed, the tubes were incubated at 72°C for 7 min on the same thermal cycler and then stored at 4°C. The amplified PCR fragments were resolved on a 2% a agarose gel containing 0.4 pg/ml ethidium bromide. Electrophore- sis was performed in l x TAE buffer and DNA size marker (100 bp DNA ladder; Pharmacia) was used to determine the size of amplified cDNA fragments.
A . MODIFIED HSP30C
1 2 3 4 5 6 7
369-
246-
123-
RESULTS Detection of Heat Shock-Induced
Endogenous hsp 30C Gene Expression by RT-PCR in Xenopus Embryos
In earlier studies employing Northern blot and RNase protection analysis, it was reported that heat shock-induced accumulation of hsp 30 mRNA occurred at the tailbud stage during Xenopus development [Krone and Heikkila, 1988, 19891. The cloning and se- quencing of the hsp 30 gene cluster has permitted the examination of the expression of the hsp 30C gene dur- ing Xenopus development by the more sensitive tech- nique of RT-PCR. In these experiments, total RNA was isolated from control (22°C for 1 hr) or heat shocked (33°C for 1 hr) embryos and subjected to reverse tran- scription using an oligo(dT) primer. The cDNA synthe- sized during this reaction was then amplified by PCR using specific primers. The endogenous expression of the hsp 30C gene was detected using primers directed at the 3'-coding region of hsp 30C gene. In Figure IA, RT-PCR yielded a 299-bp amplified fragment from heat-shocked tailbud stage RNA (lane 5) . This frag- ment was not detected in control tailbud (lane 4) or control or heat-shocked gastrula RNA (lanes 2 and 3). A second amplified fragment in lane 5 likely represents another hsp 30 type gene. The identity of the amplified hsp 30 cDNA fragments was confirmed by Southern blot analysis (data not shown). Control RT-PCR exper- iments were carried out with primers directed at the cytoskeletal actin gene coding region. Since these primers also flank the intron of the Xenopus cytoskel- eta1 actin gene it was possible to detect genomic DNA contamination in these samples. This control was im- portant since hsp 30 genes do not contain introns. As shown in Figure lB, a 290-bp RT-PCR fragment was detected in all gastrula and tailbud samples (lanes 1-4). None of these samples yielded the 369-bp frag- ment that would signify genomic DNA contamination. This observation was confirmed by Southern blot anal- ysis using the cytoskeletal actin gene as a probe (data not shown). These results support previous results demonstrating that hsp 30 gene expression is not heat- inducible until the tailbud stage of development.
B. ACTIN
1 2 3 4 5 6
369-
246-
- b C C
123-
Fig. 4. Detection of the expression of a microinjected modified hsp 30C gene containing a short 5'-flanking region. Fertilized eggs were microinjected with 400 pg of the modified hsp 30C gene (p30Cplyl). Expression of the gene was detected by means of RT-PCR assay, using specific primers. A: Modified hsp 30C RT-PCR. Lane 1,123 bp DNA ladder; lane 2, gastrula a t 22"C, lane 3, gastrula a t 33°C; lane 4, neurula at 22°C; lane 5, neurula a t 33°C; lane 6, positive control (p3OCplyl plasmid DNA); lane 7, buffer PCR control. B Actin RT- PCR. Lane 1, gastrula at 22°C; lane 2, gastrula at 33°C; lane 3, neurula at 22°C; lane 4, neurula at 33°C; lane 5, Xenopus genomic DNA; lane 6, buffer PCR control. PCR fragment sizes: a = 307 bp; b = 369 bp; c = 290 bp.
Expression of a Microinjected Modified hsp 30C Gene in Xenopus Embryos
In previous studies, we found that a microinjected Xenopus hsp 30AlCAT fusion gene containing 402 bp (previously the size was incorrectly reported as 350 bp) of promoter sequences was heat inducible after the midblastula stage unlike the endogenous hsp 30 gene, which was not heat inducible until the early tailbud stage [Krone and Heikkila, 19891. Given these results, it was possible that the cis-acting DNA elements re- quired for correct developmental regulation may not reside within the hsp 30A genomic clone. Therefore, a Xenopus genomic library was screened and an hsp 30 gene cluster was isolated [Krone et al., 19921. One of the 3 hsp 30 genes present within the cluster, namely hsp 30C, contained complete 5'- and 3'-flanking re- gions (Fig. 2). In the present study, we have repeated the earlier microinjection experiments using modified hsp 30C constructs containing 1 kb of 3'-flanking re-

492 - 369 - 246 -
HSP 30 GENE EXPRESSION IN XENOPUS EMBRYOS 47
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3
123 - Fig. 5. Detection of the expression of a microinjected modified hsp
30C gene containing extensive 5’-flanking sequences. Fertilized eggs were microinjected with 400 pg of the modified hsp 30C gene (p3OCply2). Expression of this gene was detected by means of RT-PCR using specific primers. Lane 1, 123-bp DNA ladder; lane 2, cleavage at 22°C; lane 3, cleavage at 33°C; lane 4, midblastula at 22°C; lane 5,
midblastula a t 33°C; lane 6, gastrula at 22°C; lane 7, gastrula a t 33°C; lane 8, neurula at 22°C; lane 9, neurula a t 33°C; lane 10, tailbud at 22°C; lane 11, tailbud at 33°C; lane 12, positive control (p30Cply2 plasmid DNA); lane 13, buffer PCR control. PCR fragment sizes: a = 320 bp.
gion and either 404 bp (p3OCplyl; Fig. 3A) or 3.6 kb of 5’-flanking region (p3OCply2; Fig. 3B). The p3OCplyl vector was constructed such that a 17-bp region of pUC19 polylinker separated the coding region and 3’- flanking region of the hsp 30C gene. The presence of this tagged polylinker region was required to discrim- inate between the transcripts of endogenous and mi- croinjected genes. This modified construct was micro- injected into fertilized eggs and total RNA was isolated from control or heat-shocked embryos at gastrula and neurula stages. RT-PCR assay was employed to detect expression of the modified hsp 30C gene. The down- stream primer was directed at the pUC19 polylinker region used to tag the hsp 30C gene while the upstream primer was directed at the coding region. The expres- sion of the injected gene was detectable in heat-shocked gastrula stage as well as in heat-shocked neurula stage embryos (Fig. 4A, lanes 3 and 5) as a 307-bp PCR- amplified fragment. The expression of the microin- jected gene was also detected in heat shock blastula stage embryos (data not shown). No detectable expres- sion of the injected gene was observed in any of the embryos maintained under control conditions (Fig. 4A, lanes 2 and 4), whereas, in Figure 4B, lanes 1-4 dem- onstrate the presence of actin cDNA in each sample. These results agree with the previous data showing that the injected hsp 30A gene was heat inducible dur- ing pretailbud stages but was not developmentally reg- ulated in a similar manner to the endogenous hsp 30 genes.
The second modified hsp 30C gene construct (p3OCply2) was similar to the p3OCplyl construct ex- cept that it contained 3.6 kb of 5‘-flanking region as well as a 30 bp pUC19 polylinker tag (Fig. 3B). This clone was microinjected into fertilized eggs and at key developmental stages the embryos were either incu- bated at 22°C or 33°C for 1 h, followed by RNA isolation and RT-PCR. As shown in Figure 5, the expression of
the microinjected gene was first detectable by RT-PCR using specific primers in heat-shocked midblastula stage embryos (lane 5), as well as in heat-shocked gas- trula, neurula, and tailbud embryos (lanes 7, 9, and 11). Expression of this injected gene was not detected in control or heat-shocked cleavage stage embryos or in any later stage embryos maintained under control con- ditions (lanes 2 ,3 ,4 ,6,8, and 10). Control experiments utilizing the actin primers indicated that genomic DNA was not present as a contaminant in these RNA samples. As well, primers used to detect the expression of the modified injected hsp 30C genes did not detect endogenous hsp 30C gene expression (data not shown). In each of the PCR assays, the specificity of the ampli- fied cDNA fragments was confirmed by Southern blot analyses (data not shown). This experiment reveals that the injected gene is up-regulated by heat shock but is not regulated during development in a similar man- ner to the endogenous hsp 30 genes.
Developmental Regulation of Expression of the Microinjected Modified hsp 30C Gene Is Not
Affected by Copy Number It was possible that a high copy number of the mi-
croinjected modified hsp 30C plasmid DNA may over- load the regulatory mechanism(s1 controlling hsp 30C gene expression. This could allow for premature tran- scription of the construct in pretailbud stage embryos. To address this question, fertilized eggs were microin- jected with 10 and 50 pg of the p3OCply2 plasmid. Heat shock-induced expression of the injected gene in gas- trula embryos was detectable when as little as 10 pg of DNA was injected (Fig. 6, lane 5). This amount of DNA is less than 1 copy per haploid genome equivalent at the gastrula stage of development. Figure 6 panel B shows the presence of actin cDNA in all of these sam- ples. Constitutive expression of the injected modified hsp 30C gene in control gastrula stage embryos was
48 ALIETAL.
A. MODIFIED HSP30C 1 2 3 4 5 6 7
1 2 3 4 5 6 7 8 9
5 0 0 - 400- 300- 200-
500- 400- 300-
- a 200-
- a -b
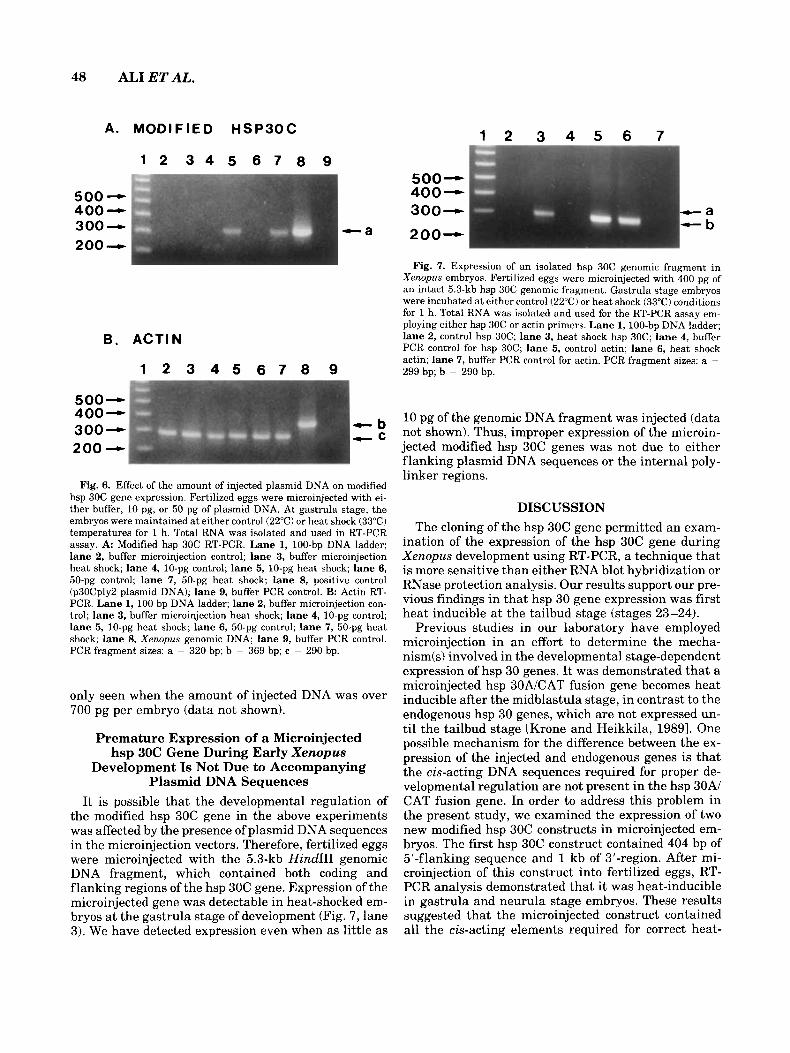
Fig. 7. Expression of an isolated hsp 30C genomic fragment in Xenopus embryos. Fertilized eggs were microinjected with 400 pg of an intact 5.3-kb hsp 30C genomic fragment. Gastrula stage embryos were incubated at either control (22°C) or heat shock (33°C) conditions for 1 h. Total RNA was isolated and used for the RT-PCR assay em- ploying either hsp 30C or actin primers. Lane 1, 100-bp DNA ladder; lane 2, control hsp 30C; lane 3, heat shock hsp 30C; lane 4, buffer PCR control for hsp 30C; lane 5, control actin; lane 6, heat shock actin; lane 7, buffer PCR control for actin. PCR fragment sizes: a =
299 bp; b = 290 bp.
6. ACTIN
1 2 3 4 5 6 7 8 9
500- 400- 300 - 200 - - b
c c
Fig. 6. Effect of the amount of injected plasmid DNA on modified hsp 30C gene expression. Fertilized eggs were microinjected with ei- ther buffer, 10 pg, or 50 pg of plasmid DNA. At gastrula stage, the embryos were maintained at either control (22°C) or heat shock (33°C) temperatures for 1 h. Total RNA was isolated and used in RT-PCR assay. A Modified hsp 30C RT-PCR. Lane 1, 100-bp DNA ladder; lane 2, buffer microinjection control; lane 3, buffer microinjection heat shock lane 4, 10-pg control; lane 5, 10-pg heat shock lane 6, 50-pg control; lane 7, 50-pg heat shock; lane 8, positive control (p3OCply2 plasmid DNA); lane 9, buffer PCR control. B: Actin RT- PCR. Lane 1, 100 bp DNA ladder; lane 2, buffer microinjection con- trol; lane 3, buffer microinjection heat shock; lane 4, 10-pg control; lane 5, 10-pg heat shock; lane 6, 50-pg control; lane 7, 50-pg heat shock; lane 8, Xenopus genomic DNA; lane 9, buffer PCR control. PCR fragment sizes: a = 320 bp; b = 369 bp; c = 290 bp.
only seen when the amount of injected DNA was over 700 pg per embryo (data not shown).
Premature Expression of a Microinjected hsp 30C Gene During Early Xenopus
Development Is Not Due to Accompanying Plasmid DNA Sequences
It is possible that the developmental regulation of the modified hsp 30C gene in the above experiments was affected by the presence of plasmid DNA sequences in the microinjection vectors. Therefore, fertilized eggs were microinjected with the 5.3-kb Hind111 genomic DNA fragment, which contained both coding and flanking regions of the hsp 30C gene. Expression of the microinjected gene was detectable in heat-shocked em- bryos at the gastrula stage of development (Fig. 7, lane 3). We have detected expression even when as little as
10 pg of the genomic DNA fragment was injected (data not shown). Thus, improper expression of the microin- jected modified hsp 30C genes was not due to either flanking plasmid DNA sequences or the internal poly- linker regions.
DISCUSSION The cloning of the hsp 30C gene permitted an exam-
ination of the expression of the hsp 30C gene during Xenopus development using RT-PCR, a technique that is more sensitive than either RNA blot hybridization or RNase protection analysis. Our results support our pre- vious findings in that hsp 30 gene expression was first heat inducible at the tailbud stage (stages 23-24).
Previous studies in our laboratory have employed microinjection in an effort to determine the mecha- nism(s) involved in the developmental stage-dependent expression of hsp 30 genes. It was demonstrated that a microinjected hsp 30AlCAT fusion gene becomes heat inducible after the midblastula stage, in contrast to the endogenous hsp 30 genes, which are not expressed un- til the tailbud stage [Krone and Heikkila, 19891. One possible mechanism for the difference between the ex- pression of the injected and endogenous genes is that the cis-acting DNA sequences required for proper de- velopmental regulation are not present in the hsp 30Al CAT fusion gene. In order to address this problem in the present study, we examined the expression of two new modified hsp 30C constructs in microinjected em- bryos. The first hsp 30C construct contained 404 bp of 5’-flanking sequence and 1 kb of 3’-region. After mi- croinjection of this construct into fertilized eggs, RT- PCR analysis demonstrated that it was heat-inducible in gastrula and neurula stage embryos. These results suggested that the microinjected construct contained all the cis-acting elements required for correct heat-

HSP 30 GENE EXPRESSION IN XENOPUS EMBRYOS 49
inducible regulation but putative DNA elements re- quired for correct developmental regulation did not re- side within the short 5‘-flanking region. Therefore, it was important to examine the expression of a microin- jected hsp 30 gene with more extensive 5’-flanking re- gion. The second modified hsp 30C construct, which contained 3.6 kb of 5’- and 1 kb of 3’-flanking region, was microinjected into fertilized Xenopus eggs and RT- PCR analysis was employed to detect its expression during development. The results indicated that this microinjected modified hsp 30C gene was also heat in- ducible at the midblastula stage. Thus, the presence of extensive 5’- and 3’-flanking regions did not repress hsp 30 gene expression at the midblastula stage. It was determined that the expression of the injected modified hsp 30C gene after midblastula was not due to a high copy number of the injected construct which might have overridden any inhibitory system. However, we cannot exclude the possibility that some DNA se- quences may be sequestered and concentrated within the embryos resulting in a localized titration of puta- tive hsp 30 inhibitory factors. Furthermore, it was shown that the expression of the modified hsp 30C gene after the midblastula stage was not due to the presence of plasmid DNA sequences associated with the micro- injected gene, since an isolated DNA fragment contain- ing the hsp 30C gene was also expressed at the early gastrula stage.
While some microinjected genes (e.g., GS 17) have been expressed correctly during Xenopus development [Krieg and Melton, 19851, some microinjected genes have not. For example, a microinjected Xenopus a- globin gene was expressed in gastrula stage embryos in contrast to the endogenous genes, which were not de- tectable until metamorphosis [Bendig and Williams, 19831. A common feature between the Xenopus globin and hsp 30 genes is that neither is normally activated during early frog development. It has been suggested that genes activated later than gastrulation may not be regulated properly during development following their microinjection into fertilized eggs [Sargent and Mathers, 19911.
Another possible common feature between Xenopus globin and hsp 30 genes is that they occur in clusters. In chicken, regulation of specific globin genes is con- trolled at the level of the gene cluster [Choi and Engel, 19881. Recently, it has been determined that clustered human globin genes are regulated at the level of chro- matin conformation by a globin locus element posi- tioned 10-20 kb upstream [Lowrey et al., 19921. Thus, it is possible that an entire hsp 30 gene cluster could be involved in developmental regulation. For example, if all the hsp 30 genes are present on a single loop of chromatin, they could assume a transcriptionally ac- tive form which would allow their expression to be reg- ulated coordinately.
Alternatively, the expression of hsp 30 genes could be regulated by localized alterations in chromatin
structure associated with each gene. Several studies have indicated that chromatin does indeed have a role in gene regulation [reviewed by Workman et al., 19911. It has been suggested that the precise placement of the core nucleosome particles in chromatin is related to particular structural properties of DNA and that such positioning has important implications for the regula- tion of eukaryotic genes [Workman et al., 19911. Sev- eral cases have been described where nucleosomes are positioned to overlap transcriptional control regions. Specific activation of such genes may require the re- moval of these positioned nucleosomes. One such ex- ample is the activation of the yeast PH05 gene which is associated with the removal of four nucleosomes up- stream of the transcriptional start site and the conse- quent unmasking of an additional regulatory sequence [Almer et al., 19861. In Drosophila, stage-dependent regulation of small heat shock proteins has been ob- served [Dura, 19811. It was also shown that alterations in chromatin structure were associated with the acti- vation of Drosophila small hsp genes [Thomas et al., 19851. In Xenopus, hsp 30 gene expression may be con- trolled at the level of chromatin conformation. Perhaps the microinjected hsp 30 genes are not packaged into an endogenous hsp 30-like chromatin conformation that prevents their expression until the tailbud stage.
ACKNOWLEDGMENTS This research has been supported by a Natural Sci-
ences and Engineering Research Council of Canada grant to J.J.H.
REFERENCES Almer A, Rudolph H, Hinnen A, Horz W (1986): Removal of positioned
nucleosomes from the yeast PH05 promoter upon PH05 induction releases additional upstream activating DNA elements. EMBO J 5:2689-2696.
Auffrey C, Rougeon F (1980): Purification of mouse immunoglobulin heavy chain messenger RNAs from total myeloma tumour RNA. Eur J Biochem 107:303-314.
Bendig MM, Williams J G (1983): Replication and expression of Xe- nopus Zueuis globin genes injected into fertilized Xenopus eggs. Proc Natl Acad Sci USA 80:6197-6201.
Bienz M. (1984a): Developmental control of the heat shock response in Xenopus. Proc Natl Acad Sci USA 81:3138-3142.
Bienz M. (198413): Xenopus hsp 70 genes are constitutively expressed in injected oocytes. EMBO J 3:2477-2483.
Browder LW, Pollock M, Nickells RW, Heikkila J J , Winning RS (1989): Developmental regulation of the heat shock response. In Di Beradino MA, Etkin LD (eds): “Developmental Biology. A Compre- hensive Synthesis.” New York: Plenum, pp 97-147.
Choi OR, Engel JD (1988): Developmental regulation of p-globin gene switching. Cell 55:17-26.
Dura JM (1981): Stage-dependent synthesis of heat shock induced proteins in early embryos of Drosophilu melunoguster. Mol Gen Genet 184:381-385.
Elgin SCR, Cartwright IL, Fleischmann G, Gilmour DS, Thomas GH (1989): Alterations in chromatin structure associated with the gene activation. In “DNA-Protein Interactions in Transcription.” New York: Alan R. Liss, Inc., pp 287-296.
Heikkila JJ, Browder LW, Gedamu L, Nickells RW, Schultz GA (1986): Heat shock gene expression in animal embryonic systems. Can J Gen Cytol 28:1093-1105.

50 ALIETAL.
Heikkila J J , Darasch SP, Mosser DD, Bols NC (1987): Heat and so- dium arsenite act synergistically on the induction of heat shock gene expression in Xenopus laeuis A6 cells. Biochem Cell Biol 65: 310-316.
Heikkila J J , Kloc M, Bury J , Schultz GA, Browder LW (1985): Ac- quisition of the heat shock response and thermotolerance during early development of Xenopus laeuis. Dev Biol 107:483-489.
Heikkila J J , Ovsenek N, Krone PH (1987): Examination ofheat shock protein mRNA accumulation in early Xenopus laeuis embryos. Bio- chem. Cell Biol 65237-94.
Hightower L, Nover L (1991): “Heat Shock and Development.” Springer-Verlag, Heidelberg.
Hitchcock MJM, Friedman RM (1980): Microinjection of Xenopus oocytes. An automated device for volume control in the nanolitre range. Anal Biochem 109:338-344.
Krieg PA, Melton DA (1985): Developmental regulation of a gastrula- specific gene injected into fertilized Xenopus eggs. EMBO J 4:3463- 3471.
Krone PH, Heikkila JJ (1988): Analysis of hsp 30, hsp 70, and ubi- quitin gene expression in Xenopus laeuis tadpoles. Development 103:59-67.
Krone PH, Heikkila JJ (1989): Expression of microinjected hsp 701 CAT and hsp 30ICAT chimeric genes in developing Xenopus laeuis embryos. Development 106:271-281.
Krone PH, Snow A, Ali A, Pasternak J J , Heikkila JJ (1992): Com- parison of the regulatory and structural regions of the Xenopus laeuis small heat-shock protein encoding gene family. Gene 110: 159-166.
Lowrey CH, Bodine DM, Nienhuis AW (1992): Mechanism of DNase I hypersensitive site formation within the human globin locus con- trol region. Proc Natl Acad Sci USA 89:1143-1147.
Mohun TJ, Brennan S, Dathan N, Fairman S, Gurdon J B (1984): Cell type-specific activation of actin genes in the early amphibian em- bryo. Nature 311:716-721.
Mohun TJ, Garrett N (1987): An amphibian cytoskeletal-type actin gene is expressed exclusively in muscle tissue. Development 101: 393-402.
Morimoto RI, Tissieres A, Georgopoulos C (1990): “Stress Proteins in Biology and Medicine.” Cold Spring Harbor, New York: Cold Spring Harbor Laboratory.
Nieuwkoop PD, Faber J (1967): “Normal Table of Xenopus laeuis (Daudin).” 2nd Ed. Amsterdam: North-Holland.
Ovsenek N, Heikkila JJ (1988): Heat shock-induced accumulation of ubiquitin mRNA in Xenopus laeuis is developmentally regulated. Dev Biol 129:582-585.
Rappolee DA (1990): Optimizing the sensitivity of RT-PCR. Amplifi- cations 45-7.
Rappolee DA, Wang A, Mark D, Werb Z (1989): A novel method for studying mRNA phenotypes in single or small number of cells. J Cell Biochem 39:l-11.
Sargent TD, Mathers PH (1991): Analysis of the class I1 gene regu- lation. In Kay BK and Peng HB (eds): “Methods in Cell Biology.” San Diego: Academic Press, Inc., pp 347-365.
Thomas GH, Siegfried E, Elgin SCR (1985): DNase I hypersensitive sites: a structural feature of chromatin associated with gene expres- sion. In Reeck G, Goodwin G, Puigdomenech P (eds): “Chromosomal Proteins and Gene Expression.” New York Plenum Press, p 77.
Workman JL, Taylor ICA, Kingston RE (1991): Activation domain of stably bound GAL 4 derivatives alleviate repression of promoters by nucleosomes. Cell 64533-544.