Embed Size (px)
Citation preview

The Prostate 28: I 53- I6 I ( I 996)
Expression of CD44 Isoforms in Human Prostate Tumor Cell Lines
Jeff W. Stevens, Patricia L. Palechek, Tomas L. Griebling, Ronald J. Midura, Oskar W. Rokhlin, and Michael B. Cohen
Departments of Orthopaedic Surgery 0. W.S., R.J.M.), Pathology (P. L P., 0. W. R.), and Urology (T.LG., M.B.C.), University of Iowa and Veterans Afairs Medical Center,
Iowa City
ABSTRACT: We have examined the expression of the transmembrane glycoproteins CD44 in four human prostate tumor cell lines. Expression was examined at the protein level by flow cytometric analysis and Western blot, and at the mRNA level by reverse transcription- polymerase chain reaction (RT-PCR). All four cell lines (DU145, LNCaP, PC3, and NDI) expressed the standard CD44 isoform (CD44s) at the mRNA level and all cell lines except LNCaP expressed CD44s at the protein level. All four cell lines contained one or more isoforms containing the v6 region (exon 10) at the mRNA level, which has been associated with metastatic potential. However, a subpopulation of LNCaP and NDl cells showed protein expression of v6. In addition, soluble CD44 isoforms were identified in cultured supernatants from all cell lines except LNCaP. These results show that CD44 isoforms are expressed on human prostate tumor cell lines, including the expression of variant isoforms containing the v6 region, and provide a rationale for the further study of this cellular adhesion molecule in prostate cancer. In addition, preliminary results indicate altered expression of CD44 in human prostatic adenocarcinomas examined immunohistochemically.
KEY WORDS:
0 1996 Wiley-Liss, Inc.
CD44, prostatic neoplasms, RT-PCR
INTRODUCTION
Cell surface receptors have long been recognized for the roles they play in the interactions between cells as well as between the cell and its surrounding extracellular matrix. These receptors have become in- creasingly important with respect to tumor cell be- havior in tissue matrices; namely tumor cell invasion and metastasis. CD44, a cell surface glycoprotein, is one of these cell surface receptors that has gained increasing attention with respect to tumor cell biology [l-71. It was originally identified on hematopoietic cells by antibody-binding studies and has since been identified on many different cell types from several animal species. Its protein sequence has been de- duced from many of these tissue sources [1,6]. Con- siderable variation in the size of the CD44 molecules has been identified, ranging from -85 to -280 kDa. Some of this variability has been attributed to carbo- hydrate modification and alteration in the presence of glycosylation recognition sites resulting from a differ-
ence in primary amino acid sequence found between the CD44 isoforms, generated by a mechanism of al- ternative mRNA splicing. Nineteen exons within the single CD44 gene copy located on chromosome llp13 have been identified that encode for the CD44 tran- scripts [B]. The most common transcript encodes for a 361 amino acid sequence species (CD44s). Several larger CD44 transcripts have been identified, result- ing in an increase in the CD44 molecule by as much as 425 amino acids. These transcripts are derived from alternative splicing of exon 6 through exon 14 by in- sertions at a single site in the extracellular encoded domain of CD44s [9,10]. The functional significance of these structural alterations in the CD44 molecule,
Received for publication November 11, 1994; accepted March 3, 1995. Address reprint requests to Michael B. Cohen, MD, Department of Pathology, University of Iowa, 200 Hawkins Drive, 5216 RCP, Iowa City, IA 52242-1009.
0 1996 Wiley-Liss, Inc.

154 Stevens et al.
Oiigomer: * + + Elq-) E12(-) E14(+)
Fig. 1. Schematic representation of the human CD44 gene. From top to bottom: primers (P), exons (boxed area representing size of individual exon), and oligomers (E) used as probes or for initiating DNA sequencing are shown.
resulting from expression of these variant transcripts, has yet to be identified. However, a series of ex- pressed transcripts that contain sequences derived from exon 10 have been correlated with carcinoma (e.g., breast, colon, stomach) [ll-151. Our interest in CD44 stemmed from a previous study of cellular ad- hesion molecules on human prostate tumor cell lines [16]. The following studies were designed to identify the CD44 isoforms found in four human prostate tu- mor cell lines.
MATERIALS AND METHODS
Cell Culture
The human prostate tumor cell lines studied were as follows: LNCaP.FGC (ATCC CRL1740), metastasis to lymph node; CU145 (ATCC HTBSl), metastasis to brain; PC3 (ATCC CRL1435), metastasis to bone; and a primary tumor cell line, ND1, kindly provided by Dr. P. Narayan (University of California, San Fran- cisco, CA) [17]. DU145 and PC3 cell lines were cul- tured in RPMI 1640 medium supplemented with 100 U/ml penicillin, 100 mg/ml streptomycin, 10% heat- inactivated fetal bovine serum (FBS; HyClone, Logan, UT), 0.1 mM 2-mercaptoethanol, and 2 mM L-glu- tamine. LNCaP and ND1 cell lines were cultured in the same media with the addition of insulin-transfer- rin-sodium selenite media supplement (Sigma, St. Louis, MO). Cells were seeded at 3 x lo5 cells in a 75 cm2 flask and incubated at 37°C in 7% CO, for 3-4 days and then subcultured at 1:5 to 1 : l O dilutions by trypsinization (0.025% trypsin-1 mM EDTA) for 2-3 min at room temperature or using a cell scraper. Cell counts were performed with a hemocytometer.
Monoclonal Antibodies
Murine anti-human monoclonal antibodies to CD44s and CD44v6 (Biosource International, Cama- rillo, CA) were used in flow cytometric and Western blot analysis for expression of CD44 isoforms. FITC-
conjugated murine anti-human CD44s (Becton and Dickinson Co., Mountain View, CA) was additionally used in flow cytometric analysis. Vectastain Elite ABC kit, mouse IgG (Vector Laboratories, Burlin- game, CA) was used for the detection of the anti- CD44 antibodies in the Western blot analysis. Control mouse immunoglobulin and affinity-purified unla- beled FITC and PE-labeled goat antibodies specific for mouse immunoglobulins were purchased from Southern Biotechnology Associates (Birmingham, AL) .
Synthetic Oligonucleotides
The following oligonucleotides were synthesized based on the previously published sequence of the human CD44 encoding for exon 1 through 19; see Figure 1 for the gene structure of CD44 and for exon identification from which the oligonucleotides were derived [9]. Three oligonucleotide Primer Sets were synthesized for polymerase chain reaction (PCR) am- plification of CD44 transcripts (5’ to 3’) as follows.
Primer Set I: P1 (+), GACACTATTGCTTCAAT- GCTTCAGC (exon 3/4, position 334-388 and P1 (-), GATGCCAAGATGATCAGCCATTCTGGAAT (exon 16/17, position 1939-1967).
Primer Set 11: P2 (+), CCTGAAGAAGATTGTA- CATCAGTCACAGAC (exon 4, position 373-402 and P2 (-), CCACTGTTCCTTCTGGGTAGCTGT (exon 10, position 1186-1209).
Primer Set 111: P3 (+), ACAGCTACCCAGAAG- GAACAGTGG (exon 10, position 1186-1209 and P3 (-), GATGCCAAGATGATCAGCCATTCTGGAAT (exon 16/17, position 1939-1967).
The E l0 (-) oligonucleotide, GTTGTACTACTAG- GAGTTGCCTG (exon 10, position 1156-1178), was synthesized for use as a probe to identify PCR prod- ucts containing the exon 10 sequence. Oligonucle- otide primers E l 2 (-), GATTTGGACAGGACAG-

CD44 lsoforms in Human CaP Cell Lines I55
GACCTC (exon 12, position 1480-1501 b) and El4 ( + ), GTCCTGCTTTCCTTCGTGTGTG (exon 14, po- sition 1694-1715) were synthesized as internal prim- ers for the sequencing of the 788 bp fragment, Primer Set 111-generated, PCR product.
PCR of CD44 Transcripts
The first-strand cDNA was made from cultured cells using the Micro-Fast Track mRNA isolation kit and the cDNA cycle kit (Invitrogen, Inc., San Diego, CA). Briefly, 1 x lo6 cells were lysed, polyA mRNA affinity purified, and cDNA synthesis generated with AMV reverse transcriptase using poly dT as an olig- onucleotide primer. Synthesized first-strand cDNA was dissolved in 20 pl of water. A 1 p1 aliquot of either a 10- or 100-fold dilution of the first-strand so- lution was used per 50 pl PCR. The PCR mix con- sisted of 1.5 mM MgCl,, 50 mM KCl, 10 pmol of each oligonucleotide primer, 200 mM dNTP, 10 mM Tris- C1, pH 8.3, with 1.5 U of Taq DNA polymerase (Boeh- ringer Mannheim Co., Indianapolis, IN), under min- eral oil. Prior to the addition of the Taq DNA polymerase, the reaction mixture was incubated at 99.9"C for 10 min. Following the addition of Taq poly- merase and oil, the reaction mix was heated to 94°C for 1 min, 62°C for 1 min, and 72°C for 2 min in a thermal cycler (Temp-Tronic, Thermolyne, Dubuque, IA) for 35 cycles. G3P-DH and p-actin housekeeping gene Primer Sets were used for monitoring relative synthesis of first-strand cDNA [18]. The G3P-DH Primer Set yielded a 309 bp fragment while the p-ac- tin yielded a 450 bp fragment.
Additional PCR amplification was performed on the Primer Set I-generated products in order to selec- tively amplify transcript fragments containing exon 10, thus reducing the overpowering presence of the CD44s transcript. Primer Set I1 was used to amplify transcript sequences between exons 4 and 10, while Primer Set I11 was used for sequences between exons 10 and 17.
PCR Product and Southern Analyses
PCR products were identified by ethidium bro- mide following resolution of the products on a 2% Seakem agarose gel (FMC BioProducts, Rockland, ME) and further processed for Southern blotting [19]. Following capillary action transfer of the DNA onto nylon membrane (Nytran, 0.45 m, Schelicher & Schuell, Keene, NH), the DNA was crosslinked to the membrane using a crosslinker (Fisher Scientific, Itasca, IL). A nonisotopic method (Genius System, Boehringer Mannheim) was used for screening spe- cific exon sequences. The membrane was prehybrid- ized 1 hr at 68°C in 5 x SSC, 0.1% Sarkosyl, 0.02%
SDS, 1 % blocking reagent (Boehringer Mannheim). The hybridization solution was identical to the pre- hybridization solution, with a 21 oligonucleotide digoxigenin tailed oligomer and was hybridized at 54°C for 3 hr. Washes with 2 x SSC, 0.1% SDS at 54°C were used prior to detection of the digoxigenin with an anti-digoxigenin antibody conjugated with alka- line phosphatase. The membrane was washed with 100 mM Tris-C1, pH 7.6; 150 mM NaCl, followed by a wash with 100 mM Tris-C1, pH 9.5; 100 mM NaCl, 50 mM MgCl,. Lumi-Phos 530 was add to the membrane and exposed to X-ray film (Kodak XAR5) to record the chemiluminescent signal.
DNA Sequencing
The PCR Primer Set I generated a -500 bp oligo- nucleotide product, Primer Set I1 a -350 bp product, and Primer Set I11 a -800 bp product. Sequencing of these PCR products was performed following Centri- con-100 purification. The PCR products (-800 bp and -250 bp fragments) generated with Primer Set 111 in DU145 and ND1 were gel purified in low melt 2% agarose SeaPlaque (FMC BioProducts) prior to se- quencing. Sequencing of PCR products was per- formed in the DNA Core Facility in the College of Medicine, University of Iowa, using an Applied Bio- system automated sequencer system (model 373A; Foster, CA). CD44-specific oligonucleotides were used for initiation of sequencing of the purified PCR products.
Western Blot Analysis
Following washing of cells in a 30 cm dish (3 X lo6 cells) with PBS, 400 pl of protein extract solution was added, containing 20 mM Tris-C1, pH 7.5; 2 mM EDTA; 2 mM EGTA; aprotinin, 25 pg/ml; 5 pM pep- satin; leupeptide, 25 pg/ml; 1 mM phenylmethylsul- fonyl fluoride; and 1% SDS [20]. Total protein con- centration was determined using the Micro BCA protein assay method (Pierce Chemical Co., Rock- ford, IL). Protein extract (60 pg protein) was resolved on 7.5% SDS-PAGE. Proteins were electroblotted onto polyvinylidene difluoride membrane (Immo- bilon-P, Millipore Co., Bedford, MA) and immuno- detection for specific proteins was performed as fol- lows. Membranes were blocked with 5% nonfat skim milk, 1% whole goat serum, 150 mM NaCl, 100 mM Tris-C1, pH 7.5, for 2 hr at room temperature, then primary antibody in blocking buffer minus goat se- rum at 4"C, overnight. Washings were performed with Triskaline buffer (pH 7.5) containing 1% nonfat skim milk. Vectastain Elite ABC kit (Vector Laborato-

156 Stevens et al.
ries) and the ImmunoPure Metal Enhanced DAB Sub- strate Kit (Pierce Chemical Co.) were used according to manufacturer's instructions for the detection of the primary antibody. Molecular weight standards (Broad Range, BioRad, Hercules, CA) were detected with Amido Black staining following electrotransfer of the proteins co-blotted on to the membrane with the total cell protein extract.
Flow Cytometric Analysis
For cell surface staining, viable cells (5 x lo5) were incubated with monoclonal antibodies (50 &ml) in 50 ~1 of BPBS (0.15 M NaCl, 0.01 M sodium phos- phate, pH 7.2, 2% bovine serum albumin [BSA], and 0.1% sodium azide) for 30 min at 4"C, washed twice, and FITC- or PE-conjugated goat anti-mouse antibod- ies were used as a developing reagent. For cytoplas- mic staining, cells were first treated with 2.5% para- formaldehyde in PBS for 60 min at 4"C, washed once with PBS, and then incubated with 0.25% Tween-20 for 15 min at 37°C. Cells were then washed twice with PBS and incubated with antibodies, as for cell surface staining. Stained cells were analyzed using a FACS 440 instrument (Becton Dickinson Co.). A minimum of 10,000 events for each sample was collected in list mode.
Soluble CD44s Production
Soluble CD44s was performed by an ELISA assay (Bender Med Systems, Vienna, Austria) per the man- ufacturer's protocol. Briefly, prostate tumor cells were seeded at 3 x lo5 in a 75 cm2 flask in RPMI 1640 media supplemented with 10% FBS. Cells were incu- bated for 4 days, and when they reached about 3 x lo6 cells per flask, 10 ml of supernatant was removed and assayed. This kit recognizes all CD44 isoforms and does not distinguish between the standard form and variant isoforms.
lmmunohistochemistry
An immunohistochemical reaction for CD44 was performed on six cases of primary prostate adenocar- cinoma using a labeled streptavidin-biotin method. Sections were deparaffinized in xylene, rehydrated in graded alcohols, and rinsed. Antigen retrieval was accomplished using citrate buffer for 10 min at high power in the microwave. Endogenous peroxidase ac- tivity was blocked using 0.3% H,O,/dH,O. Sections were covered with primary antisera, incubated over- night at 4"C, and rinsed. Sections were then covered with LSAB I1 Link Ab (Dako Corp., Carpinteria, CA) for 20 min at room temperature, rinsed, and then covered with LSAB I1 Label for 20 min at room temp
and rinsed. Sections were incubated with 0.05% DABlO.1 M TRIS with 0.015% H,O, to demonstrate signal of the primary antibody. A counterstain of 10% Harris Hematoxylin without acid for 1-2 min was used. Negative control slides were prepared by sub- stituting normal mouse serum for the primary anti- sera. Anti-CD44 antibody was purchased from Bec- ton Dickinson (San Jose, CA) and used at a dilution of 1:50. Anti-CD44 v6 antibody was purchased from Bender Med Systems and used at a dilution of 1 : l O O . Biotinylated link and label were purchased from Dako Corp. All rinses were performed using 1 Final Buffer (code-on) prepared from 10 x Automation Buffer (Biomeda Corp., Foster City, CA).
RESULTS
Variant Transcripts Are Present in All Cell Lines
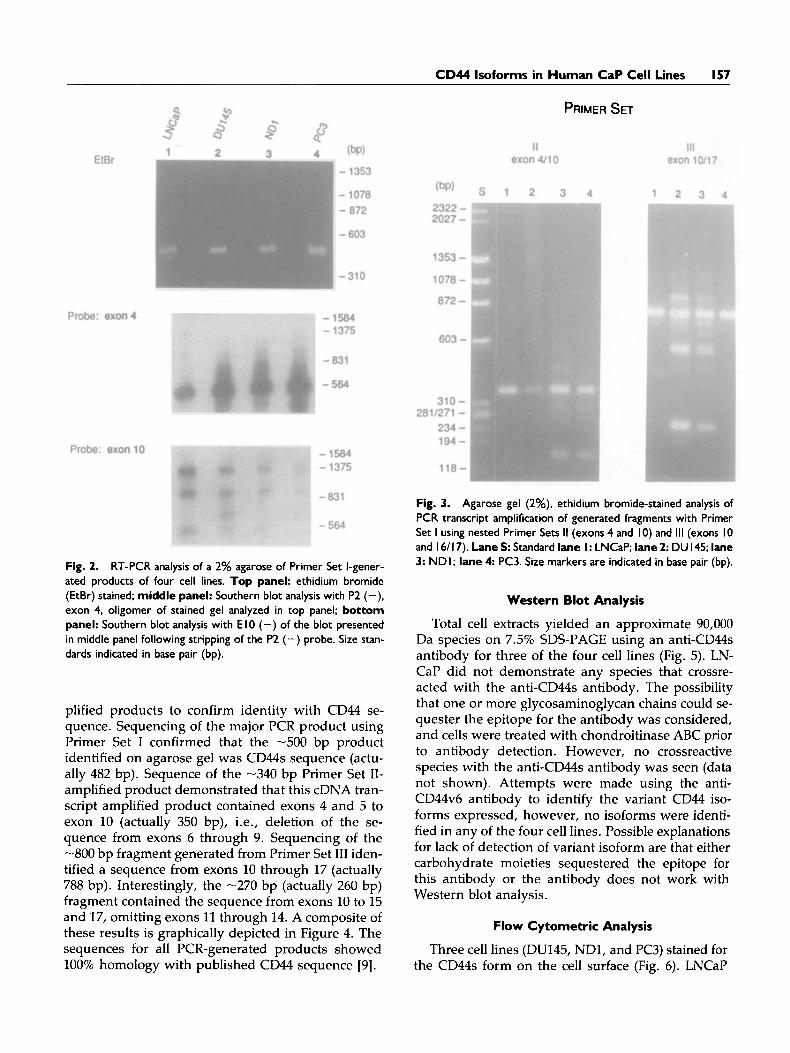
A -500 bp reverse transcription (RT)-PCR-ampli- fied product was identified by ethidium bromide staining from each cell line, when the CD44 Primer Set I containing oligonucleotide sequences from exon 3 and 16/17 was used (Fig. 2). (In some instances, multiple PCR products, -900 bp and -1200 bp, in particular, from LNCaP and DU145 were observed.) From all cell lines the -500 bp fragment hybridized with the exon 4 specific oligonucleotide P2 (+) (Fig. 2), suggesting that this fragment was derived from the CD44s transcript. Using the oligonucleotide probe, El0 (-), with a sequence encoded from exon 10, only bands -900 bp and -1,200 bp fragment were identified, suggesting the presence of alternatively spliced transcripts in all cell lines (Fig. 2). An addi- tional -600 bp band was observed in DU145, and to a lesser extent in ND1, that was not apparent in LN- CaP and PC3.
To further characterize the variant transcripts, we amplified transcript fragments using nested primers (Primer Sets I1 and 111) containing an oligonucleotide primer with a sequence from exon 10 and a second oligonucleotide primer sequence from either exon 3 or 16/17. Using Primer Set 11, which contained one oligonucleotide sequence 5' of the splice site, results indicated amplification of a single -340 bp fragment for all cell lines (Fig. 3). Primer Set 111, which con- tained one primer with a sequence from exon 16/17, yielded a -800 bp fragment in all cell lines following PCR amplification (Fig. 3). In the cell lines DU145 and ND1, two additional products (-270 bp) were iden- tified using Primer Set 111. Identification of several PCR products using Primer Set I11 and not Primer Set I1 suggests that alternative splice variants exist from splicing out exons between 10 and 16.
DNA sequencing was performed on all PCR-am-
CD44 lsoforms in Human CaP Cell Lines 157
PRIMER SET
Fig. 2. RT-PCR analysis of a 2% agarose of Primer Set I-gener- ated products of four cell lines. Top panel: ethidium bromide (EtBr) stained; middle panel: Southern blot analysis with P2 (-), exon 4, oligomer of stained gel analyzed in top panel; bottom panel: Southern blot analysis with E I0 (-) of the blot presented in middle panel following stripping of the P2 (-) probe. Size stan- dards indicated in base pair (bp).
plified products to confirm identity with CD44 se- quence. Sequencing of the major PCR product using Primer Set I confirmed that the -500 bp product identified on agarose gel was CD44s sequence (actu- ally 482 bp). Sequence of the -340 bp Primer Set II- amplified product demonstrated that this cDNA tran- script amplified product contained exons 4 and 5 to exon 10 (actually 350 bp), i.e., deletion of the se- quence from exons 6 through 9. Sequencing of the -800 bp fragment generated from Primer Set I11 iden- tified a sequence from exons 10 through 17 (actually 788 bp). Interestingly, the -270 bp (actually 260 bp) fragment contained the sequence from exons 10 to 15 and 17, omitting exons 11 through 14. A composite of these results is graphically depicted in Figure 4. The sequences for all PCR-generated products showed 100% homology with published CD44 sequence [9].
Fig. 3. Agarose gel (2%), ethidium bromide-stained analysis of PCR transcript amplification of generated fragments with Primer Set I using nested Primer Sets I1 (exons 4 and 10) and 111 (exons I0 and I 6/ 17). Lane S: Standard lane I : LNCaP; lane 2: DU 145; lane 3: NDI; lane 4 PC3. Size markers are indicated in base pair (bp).
Western Blot Analysis
Total cell extracts yielded an approximate 90,000 Da species on 7.5% SDS-PAGE using an anti-CD44s antibody for three of the four cell lines (Fig. 5). LN- CaP did not demonstrate any species that crossre- acted with the anti-CD44s antibody. The possibility that one or more glycosaminoglycan chains could se- quester the epitope for the antibody was considered, and cells were treated with chondroitinase ABC prior to antibody detection. However, no crossreactive species with the anti-CD44s antibody was seen (data not shown). Attempts were made using the anti- CD44v6 antibody to identify the variant CD44 iso- forms expressed, however, no isoforms were identi- fied in any of the four cell lines. Possible explanations for lack of detection of variant isoform are that either carbohydrate moieties sequestered the epitope for this antibody or the antibody does not work with Western blot analysis.
Flow Cytometric Analysis
Three cell lines (DU145, NDI, and PC3) stained for the CD44s form on the cell surface (Fig. 6). LNCaP

158 Stevens et al.
B
Fig. 4. Schematic representation of human CD44 gene and transcripts identified in prostate tumor cell lines. A Exon numbering with relative size of individual exons indicated by box size of the human CD44 gene. B Proposed transcripts present, based on sequencing of PCR-generated CD44 frag- ments.
the cells were positive (data now shown). DU145 and PC3, however, did not react with anti-CD44v6 on the cell surface or within the cytoplasm.
Soluble CD44
ELISA assay for soluble CD44 detected 1 ng/ml of specific protein in DU145 and ND1 supernatants, 0.4 ng/ml in PC3, and was less than the limit of detection in LNCaP.
Fig. 5. Western blot analysis for CD44 isoforms using anti-hu- man CD44s monoclonal antibody following 7.5% SDS-PAGE res- olution of total protein extraction from four prostate tumor cell lines. Molecular weight standard as indicated by size in kDa. As- terisk indicates nonspecific staining.
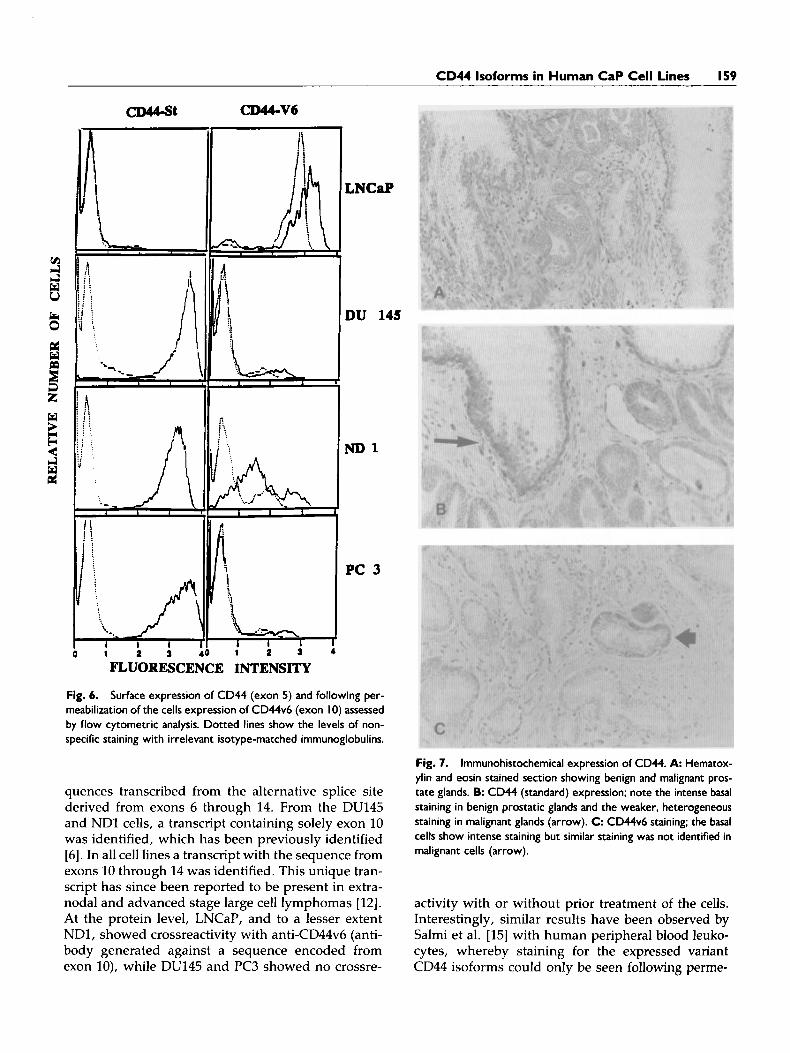
did not stain for CD44s by surface staining; however, -30% of the LNCaP cells stained with anti-CD44v6, which is specific for the expressed amino acid se- quence encoded from exon 10. Anti-CD44v6 staining was identified both on the cell surface (data not shown) and, much brighter, within the cytoplasm (Fig. 6). ND1 showed minimal reactivity with anti- CD44v6 on the cell surface (data not shown), but was clearly positive when cytoplasmic staining was per- formed (Fig. 6). Immunocytochemical staining of LN- CaP with anti-CD44v6 showed that the majority of
lmmunohistochemistry
Immunohistochemical staining of nonneoplastic prostate revealed intense membrane staining of basal epithelial cells with both anti-CD44 and anti-CD44v6 antibodies. In contrast, primary prostatic adenocarci- noma stained heterogeneously for CD44 with both a decrease in the intensity as well as the percentage of cells (Fig. 7). No staining was identified with anti- CD44v6 in prostatic adenocarcinoma (Fig. 7).
DISCUSSION
We have examined CD44 isoform expression in four human prostate tumor cell lines (LNCaP, DU145, ND1, and PC3) at both the transcriptional and protein levels. CD44s transcript was identified at variable levels in all cell lines. Interestingly, at the protein level, CD44s was seen in three of the four cell lines. CD44s was not detected by immunofluoresence staining using flow cytometric analysis, either before or following permeabilization of the LNCaP cells, by immunocytochemical analysis on fixed cells, or by Western blot analysis of total cell protein. All four cell lines contained one or more transcripts with se-
CD44 lsoforms in Human CaP Cell Lines 159
i i
\.I -. .., I 1 I
LNCap
DU 145
ND1
PC 3
FLUORESCENCE INTENSITY
Fig. 6. Surface expression of CD44 (exon 5) and following per- meabilization of the cells expression of CD44v6 (exon 10) assessed by flow cytometric analysis. Dotted lines show the levels of non- specific staining with irrelevant isotype-matched immunoglobulins.
quences transcribed from the alternative splice site derived from exons 6 through 14. From the DU145 and NDl cells, a transcript containing solely exon 10 was identified, which has been previously identified [6]. In all cell lines a transcript with the sequence from exons 10 through 14 was identified. This unique tran- script has since been reported to be present in extra- nodal and advanced stage large cell lymphomas [12]. At the protein level, LNCaP, and to a lesser extent NDI, showed crossreactivity with anti-CD44v6 (anti- body generated against a sequence encoded from exon lo), while DU145 and PC3 showed no crossre-
Fig. 7. lmmunohistochemical expression of CD44. A Hematox- ylin and eosin stained section showing benign and malignant pros- tate glands. B: CD44 (standard) expression; note the intense basal staining in benign prostatic glands and the weaker, heterogeneous staining in malignant glands (arrow). C: CD44v6 staining; the basal cells show intense staining but similar staining was not identified in malignant cells (arrow).
activity with or without prior treatment of the cells. Interestingly, similar results have been observed by Salmi et al. [15] with human peripheral blood leuko- cytes, whereby staining for the expressed variant CD44 isoforms could only be seen following perme-

160 Stevens et al.
abilization of the cells. The discordance between CD44s expression at the transcript vs. protein levels in LNCaP is most likely due to the sensitive detection using PCR, or possibly because the epitope may be masked by posttranslational modification. This dis- cordance is particularly intriguing because LNCaP is the only cell line studied for which growth is andro- gen dependent. Our findings are similar to those reported by Liu [21]. The results of our immunohis- tochemical findings of primary prostate adenocarci- noma, i.e., heterogeneous and decreased CD44 and absent CD44v6 staining, indicate decreased or lack of expression of this CAM. This finding is particularly intriguing when taken together with the protein ex- pression data of LNCaP. LNCaP is the only andro- gen-dependent cell line studied and raises questions about hormonal regulation of CD44 expression. In addition, we have extended these studies to the ex- amination of human prosatic adenocarcinomas. Our findings suggest altered expression of CD44 in pros- tatic adenocarcinomas in a manner similar to CD44 expression in LNCaP, i.e., decreased or absent pro- tein expression. The significance and etiology of these findings await future studies.
It has become increasingly apparent that CD44 has broad tissue distribution and recent studies have at- tempted to correlate its expression with tumor biol- ogy [6]. Protein and mRNA studies have identified certain CD44 isoforms that are expressed selectively on epithelial cells, as well as carcinomas [ll-151. Fur- ther, the expression of certain CD44 variants has been linked with metastasis and tumor progression [2,5,7,11,15]. The most convincing evidence for this was the report by Gunthert et al. [22] who transfected a rat pancreatic carcinoma cell line with a clone en- coding a variant exon (v6; exon 10) of CD44. The new isoform that was expressed was only found in metas- tasizing cells and was not expressed in nonmetasta- sizing cells, nor in most normal rat tissues. In other studies, CD44 expression has been examined in a va- riety of human tumors, including carcinomas of the breast, colon, and stomach [ll-151. Correlations have been identified between protein expression and bio- logc behavior. For example, Mayer et al. [14] found a significant positive correlation between immunohis- tochemical detection of CD44 in primary gastric can- cers with distant metastases, and with recurrence and mortality in patients resected with curative intent.
Hyaluronic acid (HA) binding has been identified with CD44 and HA is thought to be the major ligand for CD44s [23,24]. HA is a large polysaccharide found in specific distributions within most connective tissue matrices and pericellular layers [23]. It is thought that tumor cells bind to the HA that surround them via specific cell surface receptors and this binding event
transduces a prolifeative and/or migratory behavior in these cells. Consequently, the presence of HA re- ceptors on the surface of certain tumor cells has been implicated in the proliferation and metastasis of these cells [4,23,24]. The function of the variant isoforms of CD44 is not known and HA in fact may not be a ligand for these isoforms, e.g., v6 isoform.
Consensus sequences indicate that CD44 also has homology with ras and that CD44 may be a guanine nucleotide-binding protein with GTPase activity [25]. This would suggest that CD44 may have unique sig- nal transduction pathway(s). In addition, activated H-ras can induce CD44 promoter activity in cloned rat embryo fibroblast (CREF) cells and lead to a low level expression of CD44 isoforms, including those con- taining v6, and therefore may be able to modulate metastatic behavior [26]. Recently, Mackay et al. [27] have shown that CD44 variant isoform expression may be modulated by certain cytokines, including tu- mor necrosis factor-a and y-interferon. Taken to- gether, these results suggest that CD44 expression is a highly regulated process that may be altered during tumor transformation and progression.
CONCLUSIONS
Our findings add to the growing litrerature of the ubiquitous nature of CD44 expression and provide a rationale for further study of this particular cellular adhesion molecule in human prostate cancer.
ACKNOWLEDGMENTS
The authors wish to thank Dr. D. Moser of the DNA Core Facility in the University of Iowa College of Medicine for performing the DNA sequencing, and Kristina Gaunt for her editorial assistance in pre- paring this manuscript. Supported by the Roy Carver Trust Fund of the University of Iowa and an Ameri- can Cancer Society Institutional Research Seed grant to J.W.S. and R.J.M.; and a VA Merit Award and an American Cancer Society Clinical Oncology Career Development Award (92-12) to M.B.C.
REFERENCES
1. Haynes BF, Liao HX, Patton KL: The transmembrane hyaluronate receptor (CD44): Multiple functions, mul- tiple forms. Cancer Cells 337-350, 1991.
2. Hart IR, Birch M, Marshall JF: Cell adhesion receptor expression during melanoma progression and me- tastases. Cancer Metastasis Rev 10:115-128, 1991.
3. Sy MS, Guo YJ, Stamenkovic I: Distinct effects of two CD44 isoforms on tumor growth in vivo. J Exp Med
4. Thomas L, Byers HR, Vink J, Stamenkovic I: CD44H regulates tumor cell migration on hyaluronate-coated substrate. J Cell Biol 118:971-977, 1992.
174~859-866, 1991.

CD44 lsoforrns in Human CaP Cell Lines 161
5. Arch R, Wirth K, Hofmann M, Ponta H, Matzku S, Herrlich P, Zoller M: Participation in normal immune responses of a metastasis-inducing splice variant of CD44. Science 257682-685, 1992.
6. Gunthert U: CD44: A multitude of isoforms with di- verse functions. Curr Top Microbiol Immunol 184:47- 63, 1993.
7. East JA, Hart IR: CD44 and its role in tumour progres- sion and metastasis. Eur J Cancer 29A:1921-1922, 1993.
8. Screaton GR, Bell MV, Bell JI, Jackson DG: The identi- fication of a new alternative exon with highly restricted tissue expression in transcripts encoding the mouse Pgp-1 (CD44) homing receptor. J Biol Chem 268:12235- 12238, 1993.
9. Screaton GR, Bell MV, Jackson DG, Cornelius FB, Gerth U, Bell JI: Genomic structure of DNA encoding the lymphocyte homing receptor CD44 reveals at least 12 alternatively spliced exons. Proc Natl Acad Sci USA
10. Jackson DG, Buckley J, Bell JI: Multiple variants of the human lymphocyte homing receptor CD44 generated by insertions at a single site in the extracellular domain. J Biol Chem 3674732-4739, 1992.
11. Matsumura Y, Tarin D: Significance of CD44 gene prod- ucts for cancer diagnosis and disease evaluation. Lan- cet 340:1053-1058, 1992.
12. Salles G, Zain M, Wei-meng J, Vassiliki AB, Shipp MA: Alternatively spliced CD44 transcripts in diffuse large- cell lymphomas: Characterization and comparison with normal activated B cells and epithelial malignancies. Blood 82:3539-3547, 1993.
13. Wielenga JM, Heider K-H, Johan G, Offerhaus JA, Gunther RA, van den Berg FM, Ponta H, Herrlich P, Pals ST: Expression of CD44 variant proteins in human colorectal cancer is related to tumor progression. Can- cer Res 53:4754-4756, 1993.
14. Mayer B, Jauch KW, Gunthert U, Figdor CG, Schild- berg FW, Funke I, Johnson JP: Denovo expression of CD44 and survival in gastric cancer. Lancet 342:1019- 1022, 1993.
15. Salmi M, Gron-Virta K, Sointu P, Grenman R, Kalimo H, Jalkanen S: Regulated expression of exon v6 con- taining isoforms of CD44 in man: Down-regulation dur- ing malignant transformation of tumors of squamocel- lular origin. J Cell Biol 122:431-442, 1993.
89:12160-12164, 1992.
16. Rokhlin OW, Cohen MB: Expression of cellular adhe- sion molecules on human prostate tumor cell lines. Prostate (in press).
17. Narayan P, Dahiya R: Establishment and characteriza- tion of a human primary prostate adenocarcinoma cell line (ND-I). J Urol 148:lOOO-1004, 1992.
18. Ujiie K: mRNA expression and synthesis of endothe- lin-1 along rat nephron segment. J Clin Invest 90:1043- 1048, 1992.
19. Sambrook J, Fritsch EF, Maniatis T: “Molecular Clon- ing. A Laboratory Manual.” Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 1989.
20. Borner C, Guadagno SN, Fabbro D, Weinstein IB: Ex- pression of four protein kinase C isoforms in rat fibro- blasts. J Biol Chem 26712892-12899, 1992.
21. Liu AY: Expression of CD44 in prostate cancer cells. Cancer Lett 76:63-69, 1992.
22. Gunthert U, Hofmann M, Wolfgang R, Reber S, Zoller M, Haubmann I, Matzku S, Wenzel A, Ponta H, Herr- lich P: A new variant of glycoprotein CD44 confers met- astatic potential to rat carcinoma cells. Cell 65:13-24, 1991.
23. Knudson W, Biswas C, Li XQ, Nemec RE, Toole BP: The role and regulations of tumour-associated hyaluro- nan. In Evered D, Whelan J (eds): ”The Biology of Hy- aluronan.” New York: Wiley, Chichester, 1989, pp 150- 169.
24. Underhill C: CD44: The hyaluronan receptor. J Cell Sci 103:293-298, 1992.
25. Lokeshwar VB, Bourguignon L: The lymphoma trans- membrane glycoprotein GP85 (CD44) is a novel gua- nine nucleotide-binding protein which regulates GP85 (CD44) ankyrin interaction. J Biol Chem 267:22073- 22078, 1992.
26. Hofmann M, Rudy W, Gunthert U, Zimmer SG, Zawadzki V, Zoller M, Lichtner RB, Herrlich P, Ponta H: A link between rus and metastatic behavior of tu- mor cells: rus induces CD44 promoter activity and leads to low-level expression of metastasis-specific variants of CD44 in CREF cells. Cancer Res 53:1516-1521, 1993.
27. Mackay CR, Terpe HJ, Stauder R, Marston W, Stark H, Gunthert U: Expression and modulation of CD44 vari- ant isoforms in humans. J Cell Biol 124:71-82, 1994.
- - ~





























