Embed Size (px)
Citation preview

Int. J. Cancer (Pred. Oncol.): 64,274-279 (1995) 0 1995 Wiley-Liss, Inc.
Publication of the International Union Against Cancer J Publication de I’Union Internationale Contre le Cancer
EXPRESSION OF 2 VARIANT FORMS OF FIBROBLAST GROWTH FACTOR RECEPTOR 1 IN HUMAN BREAST Yunus A. LUQMANI~J, Caroline MORTIMER’, Constantinos YIANGOU1~3, Caroline L. Dudley SINNETT3, Matthew LAW5 and R. Charles COOMBES~ Departments of ‘Medical Oncology and 3Surgely, Charing Cross Hospital, London, and jICR, Royal Marsden Hospital, Sutton, UK.
Gurpal S. BANSAL~,
The expression of variant mRNAs encoding isoforms of fibroblast growth factor receptor (FGFR)I with either 2 or 3 Ig-like loops in the extracellular domain was investigated in human breast tissues and cell lines using a polymerase chain reaction amplification method. Almost all tissues contained both forms of FGFR I, but cancers (n = 137) had a significantly lower proportion of the transcript that encoded the full 3-loop form compared with non-malignant biopsies (n = 34). This was confirmed using microdissected populations of normal and cancerous cells from frozen tissue sections. A high ratio of the 2- to 3-loop form was found to be predictive of reduced relapse- free survival. In both groups, however, the predominant form of FGFRl was that encoding the 2-loop receptor. Cell lines derived from a variety of tissues, including breast, also co-expressed both variants of FGFRI, suggesting their presence within the same cell type. Again, there was a similar preponderance of the shorter isoform. Our results were confirmed at the protein level, where out of 5 cancers analysed 4 expressed more of the 2-loop form than the 3-loop form. Our findings suggest that cells may normally simultaneously express several splice variants of FGFR I, and aberrant expression or a change in their relative amounts (Le., in malignancy) could contribute to modified responses to either autocrine or paracrine factors. o 1995 Wiley-Liss, Inc.
The family of fibroblast growth factors (FGFs) are a group of 9 structurally related heparin binding polypeptides which have been implicated in proliferation and differentiation of a wide variety of cells of mesenchymal, epithelial and neuroecto- dermal origins and play important and diverse roles in embryo- genesis, angiogenesis, wound healing, tissue regeneration, development and tumorigenesis (Baird and Walicke, 1989; Burgess and Maciag, 1989). The 2 prototypes, FGF 1 and FGF 2, are present in almost every human tissue examined (Luq- mani et al., 1992). They are also moderately mitogenic for breast cells in vitro (Takahashi et al., 1989), and FGF 2 can synergise with oestradiol and TGF-a to increase proliferation of MCF-7 cells (Stewart et al., 1992).
These factors mediate their cellular response through the type IV class of tyrosine kinase receptors, which are structur- ally similar to the type I11 PDGF receptor. Distinct gene products encode 4 highly homologous and well-conserved receptors which share 56-92% amino acid sequence identity and are composed of an external ligand binding domain in the form of 3 Ig-like structures and an intracellular portion which contains a split tyrosine kinase domain (Jaye et al., 1992). The FGF receptors (FGFRs) have open reading frames of 800-822 amino acids predicting proteins of about 90 kDa, which increase to 110-150 kDa after glycosylation.
FGFRl and -2 are both expressed in normal and malignant human breast tissues as well as almost all other normal tissues examined (Luqmani et al., 1992). An immunohistochemical evaluation (Hughes and Hall, 1993) also showed a wide tissue distribution of low levels of FGFRl protein with most of the positive reaction confined to the microvasculature, and epithe- lial cells in some tissues.
For FGFRl and -2, a large number of variant isoforms predicted from cDNA clones and PCR products have been described by several laboratories (Hou et at., 1991; Xu et al., 1992; Jaye et al., 1992). At the transcriptional level, these are considered to arise from alternate usage of splice sites (Johnson
et al., 1991) in the pre-mRNA, differential exon splicing, alternate polyadenylation signals, as well as small deletions, and affect all parts of the receptor. The main implications are for ligand binding in the extracellular region and signal transduction through the tyrosine kinase domain with poten- tial for heterologous transphosphorylation through heterodime- risation (Bellot et al., 1991).
The most commonly described variant mRNAs are those affecting the extracellular domain and particularly one in which the 267 bp exon encoding the 1st Ig-like loop has been deleted. Surprisingly this does not abolish the binding of either FGF 1 or FGF 2 (Jaye et al., 1992), which appears to be more dependent on the presence of the 2nd and 3rd loops (Xu et at., 1992); indeed, 2-loop transfectants appear to have increased binding affinity (Shi et al., 1993; Cheong et al., 1994).
In this study we have looked for the presence of FGFR transcripts encoding 2- and 3-loop forms in normal and cancerous breast tissues and for the expression of the corre- sponding FGFRl proteins. Their clinical significance was also determined.
MATERIAL Chemicals
Random hexamers and dNTPs were from Pharmacia (Uppsala, Sweden), MMLV reverse transcriptase and tissue culture media and serum were from GIBCO (Paisley, UK) and Taq polymerase from Penninsula (St Helens, UK). [32P]dCTP (3,000 Ci mmol-I) was obtained from Amersham (Aylesbury, UK). All other reagents were from Sigma (Poole, UK) unless otherwise indicated.
Tissues Cancer tissues were obtained from surgical operations on
143 patients (details in Table I) attending breast clinics at the Royal Marsden, St George’s and Charing Cross Hospitals. These patients had not undergone therapy prior to surgery. Diagnosis was confirmed by histological examination and samples snap frozen and stored in liquid nitrogen. We also collected 34 cases of non-malignant breast tissue which were composed of 30 samples from histologically uninvolved tissue adjacent to cancer, 3 samples of tissue from fibroadenomas and 1 reduction mammoplasty specimen.
Cell lines The origin and maintenance of the cell lines used in this
study were as described previously (Luqmani et al., 1992). Monolayer cultures of cells were generally harvested at about 80% confluency and stored as frozen pellets at -70°C.
*Present address: Faculty of Allied Health, Kuwait University, P.O. Box 31470 Sulaibikhat, 90805, Kuwait.
4To whom correspondence and reprint requests should be sent at: Department of Medical Oncology, Charing Cross Hospital, Fulham Palace Road, London W6 8RF, UK.
Received: April 10, 1995.

FGFR 1 ISOFORMS IN HUMAN BREAST 275
TABLE I ~ CLINICAL DETAILS OF PATIENTS IN THIS STUDY
Characteristic Number %
Total' 119 100 Age
55.5 Ranee 29-89 Median
Menopausal status Pre- Peri-
35 8
Post- 76
Negative 63 Positive 41 Not known 15
Nodal status
29 7
64
61 39
Clinical stage TlIT2 86 85 T3lT4 Not known
Pathological stage TlIT2 T3lT4
16 17
71 31
Not known 17
Invasive ductal 104 Invasive mixed 9 In situ ductal 1 Not known 5
Positive 30 Negative 21
Histological type
Oestrogen receptor
15
70 30
91 8 1
59 41
No; known 68
'Only patients with follow-up data are included.
RNA isolation and reverse transcription Total cellular RNA was extracted from pulverised frozen
tissue and frozen cell pellets as described before (Luqmani et al., 1992). Yield and integrity were determined by spectrom- etry and agarose gel electrophoresis, respectively. RNA resus- pended in water was stored at -70°C in small aliquots.
For cDNA synthesis, 2 pg RNA were reverse transcribed by incubation at 40°C for 1 hr with 200 units MMLV reverse transcriptase in 20 pl containing 50 mM Tris-HCI, pH 8.3, 75 mM KCl, 3 mM MgCI2, 4 mM each of dATP, dCTP, dGTP and dTTP, 5 mM dithiothreitol and 250 ng random hexamers. The reaction was terminated by heating to 95°C for 5 min, and the resulting cDNA was stored at -70°C prior to PCR amplifica- tion. Blank samples were prepared in which the RNA was omitted.
PCR amplijkation Aliquots (4 pl) of cDNA were added to 94 pI containing (in
final concentration) 67 mM Tris-HCI, pH 8.8, 16 mM (NH&S04, 1.5 mM MgCIZ, 0.45% Triton X-100, 200 kg/ml gelatin, 200 p M each of dATP, dCTP, dGTP and dTTP and 1 unit Taq polymerase. This mix was divided equally into 2 tubes containing either 1 p1 (200 ng each) FGFRl primers or 1 p1 GAPDH primers (Table 11). Samples were overlaid with mineral oil and subjected to either 28 (for FGFR1) or 18 (for GAPDH) cycles of amplification by sequential steps of denatur- ation at 94°C for 30 sec, annealing at 50°C for 30 sec and extension at 72°C for 30 sec (10 min for the final cycle). Initial experiments were performed to ensure that both genes were amplified linearly under these conditions as described before (Luqmani et af, 1992).
Electrophoresis, blotting and hybridisation PCR products were chloroform extracted and 10 p1 were
electrophoresed in a 1.5% agarose gel and then capillary blotted onto Hybond N+ membrane by overnight alkali trans- fer using 0.4 M NaOH. Filters for FGFRl were prehybridised
in roller bottles (Hybaid, Teddington, UK) at 42°C for 4-6 hr in buffer containing5 x Denhardts (0.1% each of polyvinylpyr- rolidine, BSA and Ficoll), 5 x SSPE, 0.2% SDS and 100 pg/ml sonicated denatured salmon sperm DNA. For the GAPDH filters, the buffer composition was 5 x Denhardts, 0.1% SDS, 50% (v/v) deionised formamide, 5 x SSPE and 100 pg/ml salmon sperm DNA. Hybridisation was carried out overnight (18 hr) with fresh buffer supplemented with the appropriate 32-P-labelled probe. For FGFRl we used a polynucleotide kinase end-labelled oligonucleotide (encoding sequence just 3' of the acidic box) and for GAPDH a random primer-labelled cDNA. The FGFRl filters were subsequently washed in 4 x SSPE at 20°C for 30 min with 3 buffer changes and then in 2 x SSPE at 42°C for 30 min with 2 buffer changes. The GAPDH filters were washed in 2 x SSC, 0.1% SDS at 20°C for 25 rnin with 4 buffer changes and then in 0.1% SSC, 0.1% SDS at 60°C for 40 rnin with 2 buffer changes. Filters were autoradio- graphed by exposure to Hyperfilm (Amersham) and resulting bands quantified using laser densitometry.
Microdissection This was performed on 3 breast biopsies, consisting of
normal tissue as well as in situ and invasive ductal carcinoma, as described previously (Luqmani et al., 1991). Briefly, 5-7 pm frozen sections were examined under phase contrast using an inverted microscope and with reference to a parallel section stained with eosin and haematoxylin. Normal and cancerous cells were scraped off with a needle and resuspended in water before addition to the RT mix and subsequent PCR amplifica- tion as described above.
Oligonucleotides Oligonucleotides were synthesised on an ABI (Winnipeg,
Canada) DNA Synthesiser using phosphoramidite chemistry, deprotected by treatment with NH40H for 5-6 hr at 55"C, vacuum dried, resuspended in water and used without further purification. In some cases an additional step involving filtra- tion through an NAP-10 column was included, but was not generally found to be necessary. Primer design was optimised using the OLIGO software program. Primer sequences are shown in Table 11.
SDS-PAGE and Western blotting Frozen tissue samples were pulverised to a fine powder,
lysed in standard SDS-PAGE sample buffer and sonicated for 30 sec using a sonicator at maximum output. Fifty micrograms of protein were electrophoresed through a 7.5% polyacryl- amide gel and the separated proteins were transferred onto nitrocellulose membranes by overnight blotting at 4°C. After blocking with 3% milk powder in PBS-T (PBS containing 0.1% Tween 20) for 1 hr at room temperature, the membranes were incubated with a commercially available murine monoclonal anti-FGFR1 antibody (Upstate Biotechnology, Lake Placid, NY) raised against amino acids ProZ2-HidZ5 and reacting with both the (Y (3-loop) and p (2-loop) forms of the receptor. The blots were then incubated, after the standard washes, for 1 hr with an anti-mouse IgG antibody conjugated to horseradish peroxidase and after further washing with PBS-T, bands were visualised using the ECL method (Amersham) as described in the manufacturer's protocol.
Data calculation and statistics PCR products were quantitated as described previously
(Luqmani et af, 1992) and normalisation carried out using the GAPDH signals. Relative expression of 2- and 3-loop RNA was expressed as a ratio of the respective densitometric signals. Different blots were normalised by inclusion of an arbitrary standard sample in every PCR run and every blot. The clinical data were analysed using the non-parametric Mann-Whitney

276 LUQMANI ETAL.
TABLE I1 - OLIGONUCLEOTIDE PRIMERS
Size (hp)* Location Sequence (5’-3’) in cDNA’ RNA
FGFRl Sense CCTCCTCTTCTGGGCTGTGCT 18-3 8 464 3-IOOp Antisense TCTTTTCTGGGGATGTCC 482-464 197 t2-loop] Probe AACAGATAACACCAAACC 420437
Sense TCCCATCACCATCTTCCA3 379 Antisense CATCACGCCACAGTTTCC
1Nucleotides.-2Base pairs.-’Luqmani et al. (1992).
GAPDH
test and survival analyses were carried out using the Kaplan- Meier method and the log rank test.
RESULTS
PCR conditions were optimised as described before (Luq- mani et aL, 1992) such that the range of input cDNA resulted in linear amplification, allowing for quantitative comparisons (data not shown). As the FGFRl and the GAPDH primers would not work together, manipulations were stringently controlled to minimise tube variation and checked by duplicate runs.
Expression of FGFRl in normal and cancer tissues All 177 samples produced assessable products for GAPDH.
Southern blotting analysis of the FGFRl PCR products showed the expected band reflecting a 464 bp product. In addition, we also observed a smaller band corresponding to an approxi- mately 197 bp fragment, suggesting exclusion of the 267 bp cxon encoding the 1st Ig domain. In all except 3 cancers and 1 normal sample, the smaller product was predominant, as illustrated by the examples in Figure 1. In 6 of the malignant samples we obtained only the smaller 2-loop product and the 3-loop form was undetectable. In the remaining 137 cancers analysed, the median value for the densitometric ratio of 2- and 3-loop bands was 9.2 (range 0.42494). This was signifi- cantly greater ( p = 0.02) than the median of 6.7 (range 0.90-23.6) observed for the 34 non-malignant samples (Fig. 2a). The total amount of FGFRl 2- and 3-loop products normalized to GAPDH (Fig. 2b) was, however, not signifi- cantly different ( p = 0.06) between the 2 groups (cancers: median 1.1, range 0.02-35; normals: median 1.9, range 0.01- 56). We also amplified RNA from microdissected cell popula- tions from 3 breast biopsies and found that the 2:3 loop F G F R l ratio was much higher in RNA from both in situ and invasive ductal carcinoma cells than from the non-malignant cells. Figure 3 shows an example in which we obtained all 3 types of cells from different areas of the same biopsy specimen. It can be seen that the smaller band corresponding to the 2-loop form is predominant in both the invasive and in situ cancer cells (lanes I and D) whereas the normal cells (lane N) have similar amounts of the 2 forms in this case. The plasmid as expected gave only the full 3-loop product (lane p).
Expression of FGFRl protein in breast cancer Five breast cancer samples (different from the ones used in
the PCR experiments) were analysed for F G F R l protein by SDS-PAGE and Western blotting. As shown in Figure 4a, all 5 cancers expressed F G F R l protein. Two bands were visualised, a 135 kDa band corresponding to the 3-loop form of F G F R l and a 115 kDa one corresponding to the 2-loop form. The identity of these bands was confirmed by transfecting Cos 1 cells with DNA encoding the 3- and 2-loop forms of F G F R l and then analysing lysates of these cells by Western blotting (Fig. 4b). In 3 out of the 5 cancers examined, the predominant variant of F G F R l was the 2-loop form and in 1 cancer the 2
FIGURE 1 - Autoradiograph showing Southern blot of PCR products from 11 (out of 34) normal (a) and 11 (out of 143) malignant (b) breast biopsies. RNA was reverse transcribed and amplified using FGFRl primers. PCR products were electropho- resed through 1.5% agarose, ca illary blotted onto Hybond N+ membrane and hybridised to a 3FP-labelled internal oligonucleo- tide. The upper band corresponds to the expected 464 bp 3-loop PCR product and the lower one to the 197 bp 2-loop form missing a 267 bp exon. Lane marked p had PCR product generated from a plasmid template containing the full length FGFRl sequence, and lane n was a negative control.
isoforms were present in equal amounts, as shown by the intensity of the 2 bands.
Clinical correlations We evaluated clinical parameters in terms of both the total
FGFRl content (k., sum of 2- and 3-loop values), as well as the 2 3 loop ratio. In patients for whom data was available we found that there was no significant correlation between total FGFRl PCR product levels and clinical and pathological stage. However, oestrogen receptor-negative tumours had a higher level of F G F R l when compared with ER-positive ones and the difference reached statistical significance ( p = 0.02). Higher levels of FGFRl were also found in node-positive tumours, but this was not statistically significant ( p = 0.29). The 2:3 loop ratio appeared to be unrelated to oestrogen receptor status and clinical and pathological stage. The only correlation was between high 2:3 loop ratios and lymph node positivity (statistically not significant,p = 0.23).
Overall and disease-free survival analyses were performed by dividing the patients into 2 equal groups using the median value. The follow-up period ranged from 5 to 10 years. When considering the total FGFRl content, we found no difference in survival between high and low expressors. However, the patient group whose 2:3 loop F G F R l ratio was greater than the median (i.e., 8.7) relapsed earlier than the group whose ratio was below this value (Fig. 5) . Overall survival was not significantly different. The sub-group of patients (n = 19) with ER-negative tumours also showed significantly reduced relapse- free survival if they were in the group expressing a higher amount of the 2-loop form ( p = 0.004).
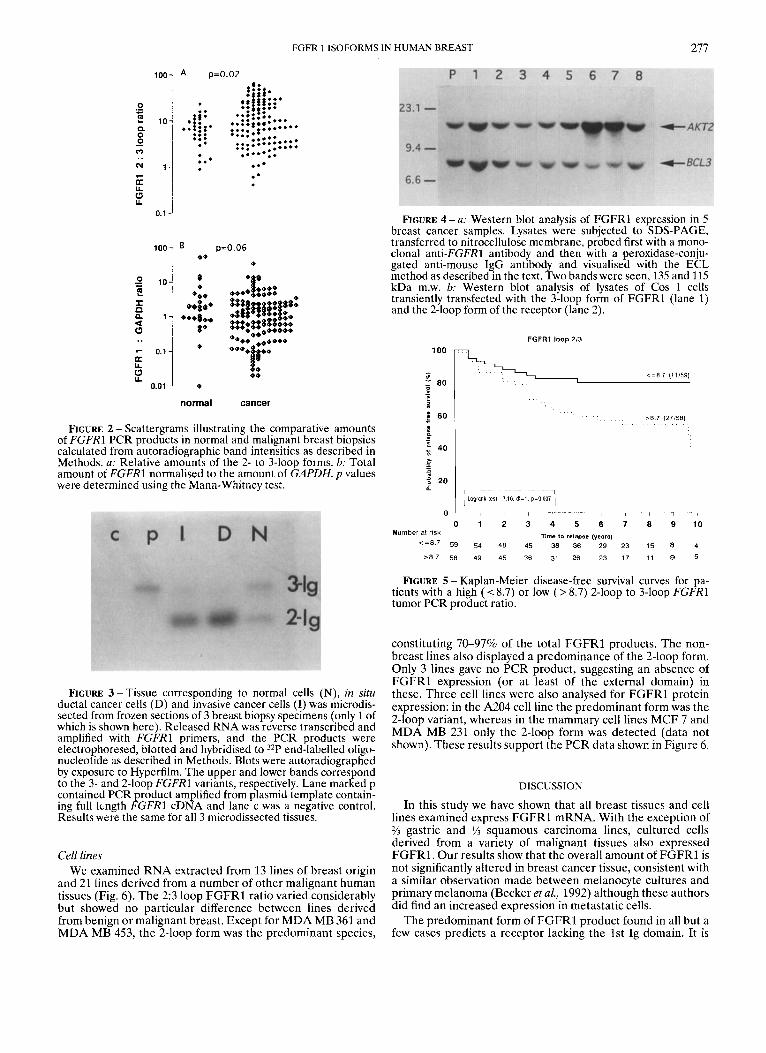
FGFR 1 ISOFORMS IN HUMAN BREAST 277
100
0 .- L
2 10 Q
0 m -
" 1 1
R Y
7-
2 0.1
100
2 10 2 n I 0 1 4 0
7 0.7
Y a
0.01 9
A p=o.oz
.*
FIGURE 2 - Scattergrams illustrating the comparative amounts of FCFRl PCR products in normal and malignant breast biopsies calculated from autoradiographic band intensities as described in Methods. a: Relative amounts of the 2- to 3-loop forms. b: Total amount of FGFRl normalised to the amount of GAPDH. p values were determined using the Mann-Whitney test.
FIGURE 3 -Tissue corresponding to normal cells (N), in situ ductal cancer cells (D) and invasive cancer cells (I) was microdis- sected from frozen sections of 3 breast biopsy specimens (only 1 of which is shown here). Released RNA was reverse transcribed and amplified with FGFRl primers, and the PCR products were electrophoresed, blotted and hybridised to 32P end-labelled oligo- nucleotide as described in Methods. Blots were autoradiographed by exposure to Hyperfilm. The upper and lower bands correspond to the 3- and 2-loop FGFRl variants, respectively. Lane marked p contained PCR product amplified from plasmid template contain- ing full length FGFRl cDNA and lane c was a negative control. Results were the same for all 3 microdissected tissues.
Cell lines We examined RNA extracted from 13 lines of breast origin
and 21 lines derived from a number of other malignant human tissues (Fig. 6). The 2:3 loop FGFRl ratio varied considerably but showed no particular difference between lines derived from benign or malignant breast. Except for MDA MB 361 and MDA MB 453, the 2-loop form was the predominant species,
FIGURE 4 -a: Western blot analysis of FGFRl expression in 5 breast cancer samples. Lysates were subjected to SDS-PAGE, transferred to nitrocellulose membrane, probed first with a mono- clonal anti-FGFR1 antibody and then with a peroxidase-conju- gated anti-mouse IgG antibody and visualised with the ECL method as described in the text. Two bands were seen, 135 and 115 kDa m.w. b: Western blot analysis of lysates of Cos 1 cells transiently transfected with the 3-loop form of FGFRl (lane 1) and the 2-loop form of the receptor (lane 2).
FGFRl loop 213
100 r - l
... . . . . . . >8 7 (27158) . . . . . . . .
B ; I 20
I Logranh lest 7 16 dl 1 p - 0 0 4
0 \ - I - I 7 1 1
0 1 2 3 4 5 6 7 8 9 10 T h e to relapse (years)
: I Number at risk
< = 8 7 59 54 48 45 38 36 29 23 15 8 4 > 8 7 58 49 45 36 31 26 23 17 11 9 5
FIGURE 5 - Kaplan-Meier disease-free survival curves for pa- tients with a high ( I 8.7) or low ( > 8.7) 2-loop to 3-loop FGFRl tumor PCR product ratio.
constituting 70-97% of the total F G F R l products. The non- breast lines also displayed a predominance of the 2-loop form. Only 3 lines gave no PCR product, suggesting an absence of FGFRl expression (or at least of the external domain) in these. Three cell lines were also analysed for F G F R l protein expression: in the A204 cell line the predominant form was the 2-loop variant, whereas in the mammary cell lines MCF 7 and MDA MB 231 only the 2-loop form was detected (data not shown). These results support the PCR data shown in Figure 6.
DISCUSSION
In this study we have shown that all breast tissues and cell lines examined express FGFRl mRNA. With the exception of 2/2 gastric and % squamous carcinoma lines, cultured cells derived from a variety of malignant tissues also expressed FGFRl . Our results show that the overall amount of FGFRl is not significantly altered in breast cancer tissue, consistent with a similar observation made between melanocyte cultures and primary melanoma (Becker et al., 1992) although these authors did find an increased expression in metastatic cells.
The predominant form of F G F R l product found in all but a few cases predicts a receptor lacking the 1st Ig domain. It is

278
PAP- m - 1
m - 2 8 - KATOIII -
LUQMANI E T A L
-m * *
FIGURE 6 - Histograms showing the relative amounts of 2-loop to 3-loop FGFR1 PCR product obtained from a variety of cell lines derived from normal breast (HBL100, HBR SV1.6.1, MCFlOA), breast cancer (ZR75-1, MDA MB 231, 361, 415, 453, 157, BT20, MCF7, SKRRIII, PMC42), gastric carcinoma (KATOIII, MJSN- 28, MKN-l), squamous carcinoma (PAP, SMN, GEE), skin (SK+, SK-), B cells (MALM6, M0417, JM), hepatoma (HEPG2), cervical carcinoma (HELA, CASKI, SIHA), choriocarcinoma (JAR), rhabdomyosarcoma (A204), colon (COL0320), neuroblas- toma (KELLY) and foetal lung fibroblast (MRC-5). *, Lines which did not give any FGFRl PCR product.
probable that this results from the alternate use of a 267 bp exon located YO bp upstream of the consensus translation initiation site (Johnson et al., 1991). The finding that normal breast tissues also expressed this truncated transcript suggests that a 2-loop receptor is not a consequence of malignancy, but may be a normal form of the receptor. Indeed, in the normal rat kidney, this form far exceeds the 3-loop form at all developmental stages (Kim et al., 1993). Another report showed almost exclusive presence of the 2-loop FGFRl isoform using domain-specific antibodies (Xu et al., 1992) in HepG2 cells. Murine muscle and heart tissue and cell lines from several other tissucs were reported to express both forms, while the 2-loop form was undetectable by RNase protection assays in both embryonic and adult brain (Bernard et aL, 1991). Interestingly, these authors found that immortalised neural precursor cells did show 2-loop expression. In the human pancreas, both FGFRl forms are reported to be present in approximately equivalent amounts in the normal tissues, but there is a preponderance of the 2-loop form in adenocarcino- mas (Kobrin et al., 1993). Using the microdissection technique, we were able to confirm that our findings with RNA extracted from whole tissue homogenates accurately reflect the differ-
ence between normal breast cells and cancer cells. We found that in situ and invasive cancer cells both expressed a prepon- derance of the shorter 2-loop variant, in contrast to normal cells from the same tissue, suggesting that the relative reduc- tion in 3-loop FGFRl (from normal cells) occurs at a stage before cells acquire propensity for invasion.
In transfection experiments, both 2- and 3-loop FGFRl constructs have been found to bind FGF 1 and 2, and both appear to be biologically responsive to these 2 peptides (Johnson et aL, 1990). Using chimeric constructs, Shi et al., (1991) have demonstrated that intact loops 1 and 2 with a loop 3 defective in its carboxyterminal half, loops 1 and 3 with a defective loop 2 and a tandem arrangement of loops 1 and 3 without loop 2 are all ineffective in binding of FGF ligand. An alternatively spliced exon forming the carboxyl half of loop 3 appears to be the major determinant of ligand specificity in both FGFRl and FGFR2 (Werner et al., 1992; Miki et al., 1992). Xu et al. (1992) have proposed a number of hypothetical models for the tertiary arrangement of the Ig domains and while it appears likely that loops 2 and 3 are sufficient to form the functional binding site, and loop 1 does not form an independent binding site either by itself or with either of the other 2 loops, they suggest it is nevertheless functionally interactive with them. It is noteworthy that both loop 2 and 3 amino acid sequences are well conserved between the 5 FGF receptors, while there is considerable divergence in the loop 1 sequence domains (Jaye et aZ., 1992); indeed, this is the region of least homology in the entire receptor, suggesting that it may confer functional differences in receptor subtypes, and at least partly account for the remarkable redundancy in this family.
At least 2 groups have also demonstrated by transfection studies that the 2-loop constructs of FGFRl (Shi et al., 1993) and of FGFR2 (Cheong et aL, 1994) have a greatly increased affinity for acidic and basic FGF and for acidic FGF and keratinocyte growth factor, respectively, compared with the 3-loop construct. Our results would support the notion that overexpression of the 2-loop form should therefore result in a more aggressive tumor, which indeed seems to be the case. As with other receptor tyrosine kinases (Ullrich and Schlessinger, 1990), it seems likely that FGFR-mediated activation involvcs receptor dimerisation. Experiments demonstrating that trans- fection of a cytoplasmically truncated FGFRl construct into Xenopus result in inhibition of intracellular calcium mobilisa- tion induced by co-transfected wild type FGFR1, FGFR2 and FGFR3, but not PDGF receptor, provide evidence for specific FGFR heterodimerisation (Ueno et al., 1992). It could be envisaged that a 2-loop FGFR in association with a 3-loop form or indeed another 2-loop FGFR, could have a quite different binding specificity from that of a 3-loop homodimer and, depending on what cytoplasmic domains are present (Jaye et al., 1992), could confer substantial growth advantages. Future studies may also highlight other binding specificities with respect to the other members of the FGF family. Both the breast carcinomas in our study and pancreatic tumours (Ko- brin et al., 1993) show an increased proportion of the 2-loop form. An elevation in the 2:3 loop ratio was also associated with increasing degree of malignancy in human astrocytic tumours (Yamagouchi et al., 1994), suggesting a significant association of the 2-loop FGFRl with the transformed pheno- type. In this study we have also shown that disease progression is correlated with the relative proportion of 2- and 3-loop FGFR1, as breast cancer patients with a higher amount of 2-loop receptor relapsed earlier. It is interesting, given the small number, that the sub-group of oestrogen receptor- negative patients also showed this trend. Expression of total FGFRl was inversely correlated with oestrogen receptor status.

FGFR 1 ISOFORMS IN HUMAN BREAST 279
The mechanism of exon splicing is clearly differentially regulated in a manner allowing different tissues to express variable amounts of variant isoforms. The pattern of expres- sion of these extracelluar FGFRl variants in established cell lines did not appear to be related to either tissue type or malignancy. It may be that culture conditions can determine the selection; if so, care should be taken to interpret such observations made on cell lines. It is also clear that compara- tive measurements relying on total receptor contents could be misleading, and that it is more appropriate to examine the relative amounts of the receptor isoforms. This is another
mechanism whereby the target cell can modulate response to a ligand without overexpression.
ACKNOWLEDGEMENTS
We thank the Cancer Research Campaign for financial support in the form of a Programme Grant, and the Buckle Family Trust for funding C.M. and C.Y. We are also grateful to Dr C. Dionne for his kind gifts of the FGFRl and -2 cDNA clones.
REFERENCES
BAIRD, A. and WALICKE, P.A., Fibroblast growth factors. Brit. med. Bull., 45,438452 (1989). BECKER, D., LEE, L.L., RODECK, U. and HERLYN, M., Inhibition of the fibroblast growth factor receptor 1 (FGFR-I) gene in human melano- cytes and malignant melanomas leads to inhibition of proliferation and signs indicative of differentiation. Oncogene, 7,2303-2313 (1992). BELLOT, F., CRUMLEY, G., UPLOW, J.M., SCHLESSINGER, J., JAYE, M. and DIONNE, C.A., Ligand induced transphosphorylation between different FGF receptors. EMBO J., 10,2849-2854 (1991). BERNARD, O., LI, M. and REID, H.H., Expression of two different forms of fibroblast growth factor receptor 1 in different mouse tissues and cell lines. Proc. nut. Acad. Sci. (Wash.), 88,7625-7629 (1991). BURGESS, W.H. and MACIAG, T., The heparin-binding fibroblast growth factor family of proteins. Ann. Rev. Biochem., 58, 575-606 (1989). CHEONG, G., LAROCHELLE, W.J., BOTTARO, D.P., BURGESS, W.H. and AARONSON, S.A., High affinity binding sites for related fibroblast growth factor ligands within different receptor immunoglobulin-like domains. Proc. nut. Acad. Sci. (Wash.), 91,989-993 (1994). Hou, J., KAN, M., MCKEEHAN, K., MCBRIDE, G., ADAMS, P. and MCKEEHAN, W.L., Fibroblast growth factor receptors from liver vary in three structural domains. Science, 251,665-668 (1991). HUGHES, S.E. and HALL, P.A., Immunolocalisation of fibroblast growth factor receptor 1 and its ligands in human tissues. Lab. Invest., 69, 173-182 (1993). JAYE, M., SCHLESSINGER, J. and DIONNE, C.A., Fibroblast growth factor receptor tyrosine kinases: molecular analysis and signal transduc- tion. Biochim. biophys. Acta, 1135, 185-199 (1992). JOHNSON, D.E., LEE, P.L., Lu, J. and WILLIAMS, L.T., Diverse forms of a receptor for acidic and basic fibroblast growth factors. Mol. cell. Bid., 10,47284736 (1990). JOHNSON, D.E., Lu, J., CHEN, H., WERNER, S. and WILLIAMS, L.T., The human fibroblast growth factor genes: a common structural arrange- ment underlies the mechanisms for generating receptor forms that differ in their third immunoglobulin domain. Mol. cell. Biol., 11, 4627-4634 (1991). KIM, E.G., KWON, H.M., BURROW, C.R. and BALLERMA”, B.R., Expression of rat fibroblast growth factor receptor 1 as three splicing variants during kidney development. Amer. J. Physiol., 264, F66-F73 (1993). KOBRIN, M.S., YAMANAKA, Y., FREISS, H., LOPEZ, M.E. and KORC, M., Aberrant expression of type 1 fibroblast growth factor receptor in human pancreatic adenocarcinoma. Cancer Res., 53,47414744 (1993).
LUQMANI, Y.A., SMITH, J. and COOMBES, R.C., PCR aided analysis of gene expression in frozen tissue sections. Anal. Biochem., 200,291-295 (1992). LUQMANI, Y.A., GRAHAM, M. and COOMBES, R.C., Expression of basic fibroblast growth factor, FGFRl and FGFR2 in normal and malignant human breast, and comparison with other normal tissues. Brit. J. Cancer, 66,273-280 (1992). MIKI, T., BO~TARO, D., FLEMING, T., SM~TH, C., BURGESS, W., CHAN, A. and AARONSON, S., Determination of ligand binding specificity by alternative splicing: two distinct growth factor receptors encoded by a single gene. Proc. nut. Acad. Sci. (Wash.), 89,246-250 (1992). SHI, E., KAN, M., Xu, J., MORRISON, R. and MCKEEHAN, W.L., Direct linkage of fibroblast growth factor to a fragment of its receptor extracellular domain after receptor-dependent internalization. J. Cell. Bid., 115,416~ (1991). SHI, E., KAN, M., Xu, J., WANG, F., Hou, J. and MCKEEHAN, W.L., Control of fibroblast growth factor receptor kinase signal transduction by heterodimerisation of combinatorial splic variants. Mol. cell. B i d , 13,3907-3918 (1993). STEWART, A.J., WESTLEY, B.R. and MAY, F.E.B., Modulation of the proliferative response of breast cancer cells to growth factors by oestrogen. Brit. J. Cancer, 66,640-648 (1992). TAKAHASHI, K., SUZUKI, K., KAWAHARA, S. and ONO, T., Growth stimulation of human breast epithelial cells by basic fibroblast growth factor in serum-free medium. In/. J. Cancer, 43,870-874 (1989). UENO, H., GUNN, M., DELL, K., TSENG, A. and WILLIAMS, L.A., Truncated form of fibroblast growth factor receptor 1 inhibits signal transduction by multiple types of fibroblast growth factor receptors. J. b id . Chem., 267,1470-1476 (1992). ULLRICH, A. and SCHLESSINGER, J., Signal transduction by receptors with tyrosine kinase activity. Cell, 61,203-212 (1990). WERNER, S., SHUHN, D., DUAN, R., DEVRIES, C., PETERS, K.G., JOHNSON, D.E. and WILLIAMS, L.T., Differential splicing in the extracellular domain of fibroblast growth factor receptor 1 generates receptor variants with different ligand binding specificities. Mol. cell. B i d , 12,82-88 (1992). XU, J., NAKAHARA, M., CRABB, J.W., SHI, E., MATUO, Y., FRASER, M., KAN, M., Hou, J. and MCKEEHAN, W.L., Expression and immunochemi- cal analysis of rat and human fibroblast growth factor receptor (flg) isoforms. J. biol. Chem., 267,17792-17803 (1992). YAMAGOUCHI, F., SAYA, H., BRUNER, J.M. and MORRISON, R.S., Differential expression of two fibroblast growth factor receptor genes is associated with malignant progression in human astrocytomas. Proc. nut. Acad. Sci. (Wash.), 91,484488 (1994).





























