Embed Size (px)
Citation preview

Expression and purification of bacteria membrane proteins for structural studies

To solve a membrane protein structure is very hard for the following
reasons:• Hard to over-express large quantity, functional protein;
• To get the protein into solution for purification it needs to be solublized with detergent, which might destroy the protein, we need protein that is functional, folded and monodisperse, many detergents need to be screened;
• Hard to crystallize: with detergent around the molecule it’s much harder to make contacts for crystal packing;
• Crystals often don’t diffract well because of high solvent contents.

Start with multiple homologues
Multiple Homologues
Plasmid Construction
Expression Screening
DetergentScreening
Crystallization
It is considered a good starting point if we could get 1mg of protein after passing through one column from one liter cell culture.
To find the right detergent/detergent mix to keep the protein stable and homogenous.

Expression Assay
21.514.46.0
kD
Eleven out of fifty targets overexpressed using arabinose induction.
Par
a6-aa
Pml I
6-hisCAC GTG TM gene TTA ATT AA
Pac I
Pme I
Nco I
RBS Sma IPT
7
Expression Vector
96 x 65 mL Fermentor
From Linda Columbus

Special considerations when over-expressing membrane proteins for
crystallographic studies Difficulties Solutions
Over-expressed protein often toxic to host cells
Choose a tight promoter to prevent leaky expression. Induce at mid-log phase so the cell will keep growing after induction, monitor the cell growth after induction.
Problems with misfolded protein in inclusion bodies
Lower growth temperature, avoid over-loading translocon and chaparones.

First purification step: isolate the membrane fraction
• Disrupt the harvested cells by mechanical force (high pressure) using equipment such as French Press or Xpress;
• Remove unbroken cells with a “low speed” centrifugation (10k xg). This step removes cell debris that are not membrane, as well as some cell organelles such as inclusion bodies;
• Collect the membrane fraction by centrifuge the supernatant from the last step with a “high speed” ultra-centrifugation (35-40k xg) . This step removes all the soluble proteins.

Properties of detergent molecules
From Anatrace.com
detergent = very expensive soap

Critical micelle concentration (CMC)
From Anatrace.com
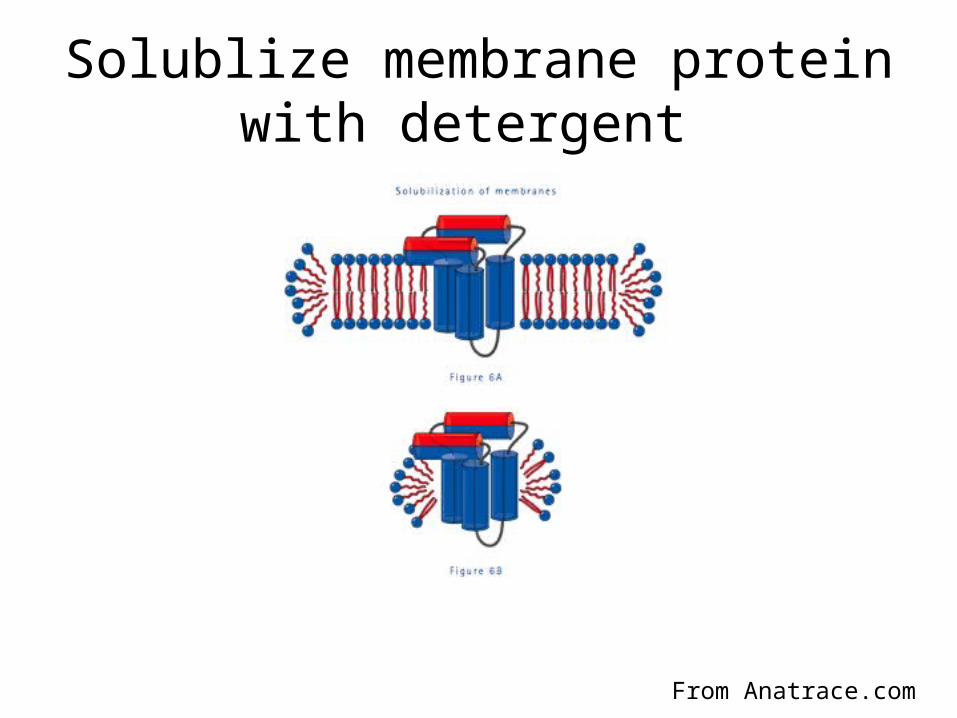
Solublize membrane protein with detergent
From Anatrace.com

Lipopeptide detergents
McGregor CL et. al. Nature Biotechnology, 2003
detergent
LPD
phospholipid

Special considerations when purifying membrane proteins
Difficulties SolutionsExtracting protein from its lipid bilayer often result in unstable/inactive proteins
Test a few detergent chosen by their reported success and price. Consider detergent exchange.
The amount of de-lipidation is often important for the protein’s stability and homogeneity
Keep in mind every column the protein passes is a de-lipidation process as well. Sometimes it’s important to add back lipid to keep the protein happy.
Because detergent forms micelles, it often concentrates with the protein sample, when the concentration is too high, the empty detergent micelles pack into crystals and making it impossible to get well-diffracting protein crystals.
Use 2-3 times CMC in all the buffers used for purification; control the protein : detergent ratio in the last purification step; Dialyze the sample to lower detergent concentration if necessary.

An example: Major facilitator superfamily (MFS)
• Largest family of secondary active transporters, > 5,000 members Including:
- Glucose-6-phosphate transporter from E. coli - UhpT- Glucose-6-phosphate transporter from human - G6PT- Glucose transporters in erythrocyte and muscle - Glut1, Glut4 - Vesicular glutamate transporters in presynapse - VGlut1, VGlut2
Transporters Families MFS ABC
E. coli 297 59 66 67
S. cerevisiae 258 42 78 22
Human 1,247 69 104 94

Fifteen MSF proteins are expressed in E. coliSearching crystallization space by cloning homologues
pBAD-MycHis
MCS-Myc-HispBAD term
AMPrAraC
pUC ori
SDS-PAGE Western blot (INDIA anti-His)

Flexible tail is identified and removedby proteolysis and mass spectrometry
C
N
N
C
Wile-type: 1-452Final construct: 2-452
G2->L2, C-terminal RNGG->LVPR (thrombin site)
R449

GlpT is a monomer in detergent solutionas measured by size-exclusion column on HPLC
In DDM
Void

Commercially-available MP screens
•MembFac (Hampton)
• MemStart & MembSys, MemGold (Molecular Dimensions)
• JBScreen Membrane 1-3 (Jena Biosciences)
• The MbClass (Nextal)
Statistics show most membrane protein crystalizes in PEG based
Conditions, a through PEG/pH screen should be performed as well.
Crystallization

Web resourcesMembrane protein structure databases:Topology databasehttp://blanco.biomol.uci.edu/mptopo/
Known structureshttp://blanco.biomol.uci.edu/Membrane_Proteins_xtal.html
Known structures with statistics of crystallization conditionshttp://www.mpibp-frankfurt.mpg.de/michel/public/memprotstruct.html
PDB database for membrane proteinshttp://pdbtm.enzim.hu/

Lemieux MJ et. al. Protein Science, 2003

Table 2. Endogenous phospholipid co-purified with GlpT
Ni2+-NTA column Preparative size-exclusion column
Detergent Mono-dispersity
Lipid /GlpT
Crystal Detergent Mono- dispersity
Lipid /GlpT
Crystal
DDM& Yes 41±4 (n=8)
No DDM& Yes 22±1 (n=8)
Yes 7.0 Å
DDM/C12E9*
Yes 24±1 (n=4)
Yes 3.2 Å
Notes: Lipid/protein ratio represents mol Pi/mol GlpT determined using a phosphorous assay, with ‘n’ being the number of samples analyzed. Protein monodispersity was determined by analytical size-exclusion chromatography on HPLC. &: 0.075 % DDM; *: 0.075 % DDM and 0.025 % C12E9.
Lemieux MJ et. al. Protein Science, 2003


Structure determination to 3.3 Å
P3221, a = b = 97.6 Å, c = 175.2 Å, 72 % sol
W15 SIRAS phasing
R = 29.65 %, Rfree = 32.53 %
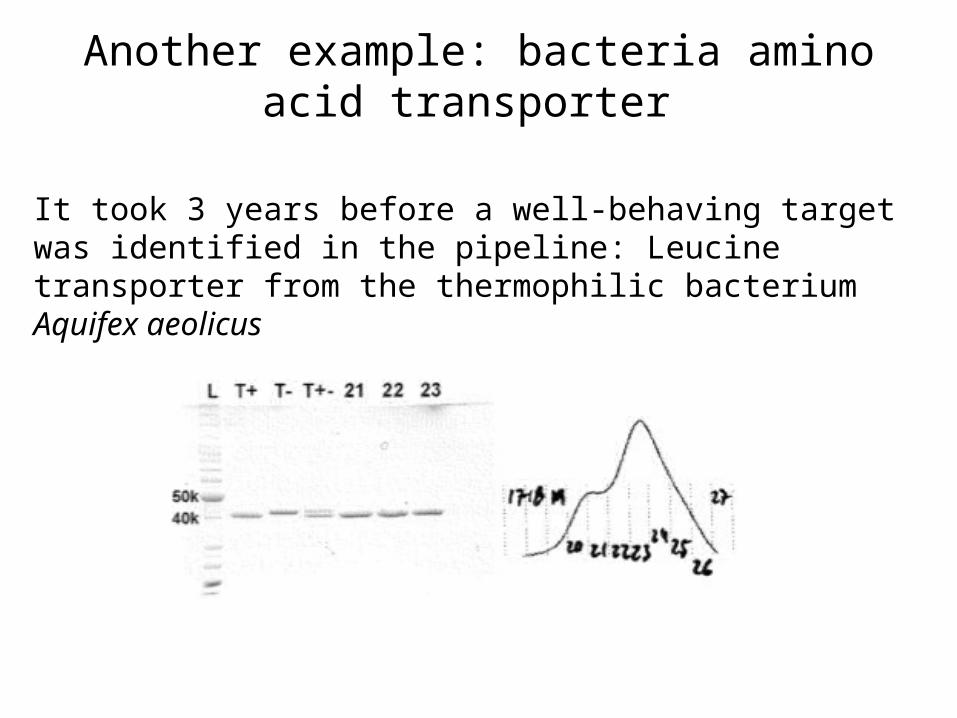
Another example: bacteria amino acid transporter
It took 3 years before a well-behaving target was identified in the pipeline: Leucine transporter from the thermophilic bacterium Aquifex aeolicus

By varying the cell culture temperature, the protein became very homogeneous shown by size exclusion
chromatography

Yamashita A et. al. Nature, 2005

It often takes a long time before a suitable target is identified, Can
we speed up this process?

Green Fluorescent Protein (GFP) has existed for more than one hundred and sixty million years in one species of jellyfish, Aequorea victoria. The protein is found in the photoorgans of Aequorea.
Green Fluorescent Protein

Kawate & Gouaux Structure. 2006 Apr;14(4):673-81





























