Embed Size (px)
Citation preview

90
1
Research ArticleReceived: 2 December 2008 Revised: 11 February 2009 Accepted: 13 February 2009 Published online in Wiley Interscience: 9 April 2009
(www.interscience.wiley.com) DOI 10.1002/jctb.2173
Exploiting olive oil mill effluents as arenewable resource for production ofbiodegradable polymers through a combinedanaerobic–aerobic processM. Beccari,a L. Bertin,b D. Dionisi,a F. Fava,b S. Lampis,c M. Majone,a∗F. Valentino,a G. Vallini,c and M. Villanoa
Abstract
BACKGROUND: The performance of a three-stage process for polyhydroxyalkanoate (PHA) bioproduction from olive oil milleffluents (OME) has been investigated. In the first anaerobic stage OME were fermented in a packed bed biofilm reactor intovolatile fatty acids (VFAs). This VFA-rich effluent was fed to the second stage, operated in an aerobic sequencing batch reactor(SBR), to enrich mixed cultures able to store PHAs. Finally, the storage response of the selected consortia was exploited in thethird aerobic stage, operated in batch conditions.
RESULTS: The anaerobic stage increased the VFA percentage in the OME from 18% to ∼32% of the overall chemical oxygendemand (COD). A biomass with high storage response was successfully enriched in the SBR fed with the fermented OME at anorganic load rate of 8.5 gCOD L−1 d−1, with maximum storage rate and yield (146 mgCOD gCOD−1 h−1 and 0.36 COD COD−1,respectively) very similar to those obtained with a synthetic VFA mixture. By means of denaturing gradient gel electrophoresis(DGGE) analysis, different bacterial strains were identified during the two SBR runs: Lampropedia hyalina and CandidatusMeganema perideroedes, with the synthetic feed or the fermented OMEs, respectively. In the third stage, operated at increasingloads, the maximum concentration of the PHA produced increased linearly with the substrate fed. Moreover, about half of thestored PHAs were produced from substrates other than VFAs, mostly alcohols.
CONCLUSION: The results obtained indicate that the process is effective for simultaneous treatment of OME and their valorizationas a renewable resource for PHA production.c© 2009 Society of Chemical Industry
Keywords: polyhydroxyalkanoates (PHAs); storage; olive mill effluents (OME); VFA production; DGGE
INTRODUCTIONBioplastics, such as polyhydroxyalkanoates (PHAs) and partic-ularly the copolymer poly(ß-hydroxybutyrate/ß-hydroxyvalerate)[P(HB/HV)], are attracting much interest as alternatives to oil-basedplastics, because they are biodegradable and can be formed fromrenewable resources. The main factor that has until now hamperedgreater diffusion of PHAs is the high cost of production.1 Indeed,present processes for PHA production are mostly based on the useof pure cultures (e.g. Ralstonia eutropha) grown on well-defined,nutrient-deficient synthetic media.2
In most organisms so far investigated, PHAs are synthesizedfrom acetate activation through acetyl coenzyme A (or propionyl-CoA, in the case of propionate). Acetyl-CoA is then drifted tothe synthesis of PHB by condensation to acetoacetyl-CoA andreduction to hydrobutyryl-CoA (consuming NAD(P)H), which isfinally incorporated in the growing chain of PHB. Under aerobicconditions, this pathway is usually triggered as an alternativesink of excess reducing power, when the growth is limited byeither external factors (i.e. nitrogen limitation) or internal factors(i.e. sudden excess of carbon source for microorganisms coming
from substrate limiting conditions). Alternation of anaerobicand aerobic conditions can also create the driving force forstorage for those microorganisms that have an alternative wayto provide energy themselves under anaerobic conditions, e.g.phosphorus or glycogen accumulating microorganisms.3 Thus,an interesting alternative to pure cultures would be the use ofactivated sludge from wastewater treatment plants, where suchtriggering conditions often occur.4 – 6 In particular, the storage is
∗ Correspondence to: M. Majone, Department of Chemistry, Sapienza Universityof Rome, P.le Aldo Moro 5, 00185 Rome, Italy.E-mail: [email protected]
a Department of Chemistry, Sapienza University of Rome, P.le Aldo Moro 5, 00185Rome, Italy
b Department of Applied Chemistry and Material Science, University of Bologna,viale Risorgimento 2, 40136 Bologna, Italy
c Department of Biotechnology, University of Verona, Strada Le Grazie 15 – Ca’Vignal, 37134 Verona, Italy
J Chem Technol Biotechnol 2009; 84: 901–908 www.soci.org c© 2009 Society of Chemical Industry

90
2
www.soci.org M Beccari et al.
more evident when the activated sludge is cultured under the socalled ‘feast and famine conditions’,7 e.g. the intermittent feedcreates the above mentioned alternation of excess and lack ofexternal substrate, as typical in sequencing batch reactors (SBRs).8
In ‘feast and famine’ systems, the microorganisms that are mostable to store the substrate during the initial feast phase possess acompetitive advantage over the other microbes, as they can usethe stored polymer as an internal carbon source during the faminephase. Thus, under the feast and famine regime the activatedsludge can be enriched with PHA-forming microorganisms and itsoverall PHA-storage potential is greatly increased. On the otherhand, in activated sludge processes the substrate consumptionrate and storage rate remain too low due to the high sludge ageand low organic load rate (OLR), which these systems are typicallyoperated at.
Therefore, in order to produce PHAs at high rates and yields,microorganisms most able to store PHAs have to be enriched fromactivated sludge under appropriate conditions, i.e. the process hasto be operated with concentrated influent, dynamic feeding, andat high organic load.
Based on these considerations, a three-stage process for PHAproduction has been proposed based on studies with syntheticsubstrates9,10 and successfully verified with real wastewater.11,12
In the first anaerobic stage, a high-concentration biodegradablewaste is fermented to produce a volatile fatty acid (VFA)-richeffluent. This effluent is then fed to a second (aerobic) stage, inwhich an activated sludge process is operated at high organic loadby periodic feeding in a SBR, in order to enrich and producea sludge with high storage response. The storage responseof the sludge produced is then exploited in a third (aerobic)stage, which is operated in batch mode in order to increase theamount of polymer produced. Finally, the PHA-rich sludge canbe sent to the traditional processes for polymer extraction andpurification.13 – 15 A schematic drawing of the three-stage processis depicted in Fig. 1.
I Fermentative Stage:Acidogenic Fermentation
of OMEs
II Enrichment Stage:Production of biomass with high
storage ability (SBR reactor)
III Accumulation Stage:PHAs accumulation
(BATCH tests)
Olive Oil Mill Effluents(OMEs)
Fermented OMEs
To PHA extraction
Biomass with highPHA content
Enriched biomass withhigh storage ability
Sludge to treatmentand disposal
Figure 1. Schematic drawing of the three-stage process for PHA produc-tion from OMEs (solid lines refer to liquid fluxes, dotted lines to biomassfluxes).
A further advantage of employing enriched mixed culturesselected under dynamic feeding is the possibility to use organicwastes as feedstock for PHAs production instead of more expensivecarbon sources (e.g. well-defined synthetic substrates such asglucose and propionate mixtures). Several alternative feedstocksare currently being studied, such as swine waste liquor, palmoil mill effluents, and vegetable and fruit wastes.6,16 – 21 In aprevious research,16 the use of olive mill effluents (OMEs) hasbeen investigated, based on the possibility of fermenting OMEs athigh rates to produce VFAs, which are the most direct substrates forPHAs production. Moreover, OMEs cause serious environmentalproblems due to their high levels of production (about 30 millionm3 year−1 in the Mediterranean area) and high COD content.
The cited paper16 was aimed at determining, through batch tests(third process stage), whether fermented OMEs were a suitablesubstrate for PHA production by mixed cultures that had beenpreviously enriched on synthetic substrates. No attempt was madeto use OME for the enrichment stage (second stage).
The aim of this study was to investigate the possibility of usingOMEs as a no-cost substrate for the production of PHAs by meansof the overall proposed process. Moreover, in order to minimize theprocess costs and control procedures, the possibility of employinga non-pretreated OME was verified. The main attention was paid toinvestigating the factors and conditions affecting the acidogenicfermentation of OMEs into VFAs in the packed-bed biofilm reactor,as well as the enrichment of PHA-forming biomass in the aerobicSBR, and the accumulation of PHAs in the third stage of the process.
EXPERIMENTALThe olive mill effluent (OME)The OME was taken from the three phase Sant’Agata d’Onegliaolive mill, Imperia, Italy and it was fed to the anaerobic reactorwithout previous pretreatment, addition of nutrients, or dilution.The OME COD was about 37 g L−1, partially due to VFAs (about 7gCOD L−1), its suspended and volatile solids content was 22.6 and16.6 g L−1, respectively, and the Kjeldhal nitrogen amount was4.97 g L−1. The OME phenolic fraction was about 1.5 g L−1. Thewastewater pH was 5.5.
The first anaerobic stage: packed bed biofilm reactor (PBBR)The acidogenic fermentation of OMEs was performed in ananaerobic PBBR. The reactor consisted of about 2.5 L, hermeticallyclosed and thermostated glass column equipped with a recycleline in which the cultural broth was continuously recycled fromthe top to the bottom of the column, at a recycling ratio ofabout 0.97. High-porous alumina-based ceramic cubes namedVukopor S10 (Lanik, CZ) were employed as the packing material.Cube dimensions and porosity were 25 × 25 × 18 mm and 10ppi, respectively. In the presence of support, the actual reactionvolume of the PBBR was about 2.22 L. More details related to thesystem configuration are reported elsewhere.22 The reactor wasoriginally inoculated with the anaerobic, OME-digesting microbialconsortium employed in previous research22 and operated forabout 1 month at 25 ◦C and at an organic load rate (OLR) of about26 gCOD L−1 d−1. During the operating period, the PBBR wassampled daily through sampling ports placed along the PBBR inletand outlet lines. The samples collected were filtered on 0.22 µmcellulose-nitrate filters (Millipore, MO, USA) and then analysedfor chemical oxygen demand (COD), VFAs, alcohols and phenolsconcentration. pH was monitored daily using a pH probe (81-04 model, ATI Orion, Boston, USA) located on the top of the
www.interscience.wiley.com/jctb c© 2009 Society of Chemical Industry J Chem Technol Biotechnol 2009; 84: 901–908

90
3
Bioproduction of PHA from olive mill effluents www.soci.org
reactor. The amount and composition of the biogas producedwere determined as described in the analytical methods section.
The second aerobic stage: sequencing batch reactor (SBR)The second stage of the process (for the aerobic enrichmentof the PHA-producing biomass) was studied in a lab-scale SBR(working volume 1 L, temperature 25 ◦C). The SBR cycle was 2 hlong, consisting of a reaction phase (119 min), including an initialfeed phase (10 min), and a withdrawal phase of the mixed liquor(1 min). In contrast to typical SBR operation,8 there was no settlingphase and all excess biomass was withdrawn with the mixedliquor. In this way, the biomass retention time (SRT) was equal tothe hydraulic retention time (HRT, 1 d). During the overall cycle,the SBR was aerated by means of a membrane compressor andstirred by a mechanical impeller. The pH of the SBR was controlledat 7.5 by CO2 bubbling from a compressed gas cylinder.
After initial inoculation with activated sludge from a civilwastewater treatment plant, the SBR was fed with a syntheticmixture of VFAs (acetic and propionic acid, in mineral medium),at an overall concentration of 8.5 gCOD L−1 and an organicload rate (OLR) of 8.5 gCOD L−1 d−1 (85% from acetic and 15%from propionic acid). Subsequently, the feed was switched to theeffluent from the OME fermentation stage, diluted about fourtimes with mineral medium, in order to maintain the same OLR interms of overall COD, as for the synthetic feed.
The composition of the mineral medium was the following(in mg L−1): (NH4)2SO4 (2000), K2HPO4 (334), KH2PO4 (259),CaCl2 ·2H2O (50), MgSO4 ·7H2O (100), FeCl3 ·6H2O (2), Na2EDTA (3),ZnSO4 · 7H2O (0.1), MnCl2 · 4H2O (0.03), H3BO3 (0.3), CoCl2 · 6H2O(0.2), NiCl2 · 6H2O (0.02), CuCl2 · 2H2O (0.01), NaMoO4 · 2H2O (0.03).Thiourea (20) was also added to inhibit nitrification. During eachSBR cycle, the dissolved oxygen (DO) was continuously monitoredand its change of slope from negative to positive values indicatedthat the substrate had been depleted. This determined the feastphase duration and the initiation of the famine phase. The SBRperformance was also characterized by measurements of volatilesuspended solids (VSS) and of PHAs, by taking samples duringtypical cycles on different days. As for the SBR run with thesynthetic feed, the PHA sampling was carried out at the time ofsubstrate depletion and at the end of the cycle, whereas the VSSsampling was performed only at the time of substrate depletion.On the other hand, during the run with the fermented OMEs, thecomplex nature of the COD contained in the fermented effluentdid not allow easy detection of the time of substrate depletionfrom the DO profile. Thus, the VSS and PHAs were determined at10 min (end of the feed), 30 min (arbitrarily chosen), and 120 min(end of the cycle). Moreover, during the latter run, samples of theliquid phase were taken in order to measure substrate removal(i.e. VFAs, alcohols, and COD), at the same time intervals (10, 30,120 min).
During both SBR runs, samples of biomass were also taken at theend of the cycle at different days for biomolecular characterization,as described later.
The third aerobic stageThe third stage of the process was simulated through tests directlycarried out within the aerobic SBR reactor by changing its usualfeed for just one cycle. Thus, the SBR was fed with differentconcentrations of fermented OMEs, corresponding to about 1, 3,and 9 times the usual load adopted in a single SBR cycle (feedingtime remaining 10 min). During the tests, the reactor was sampled
at regular intervals for the analysis of substrates (i.e. VFAs, alcohols,and total COD) and PHAs. Sufficient time was left between tests torestore normal SBR performance with the usual organic load.
Analytical methods and calculationsThe total COD determination was carried out with a CODcell test (Merck, Darmstadt, Germany). VFAs and alcohols weremeasured on filtered samples (0.22 µm porosity) by GC-FID (PerkinElmer 8410, Beaconsfield, England; stationary phase Carbowax20 M 4% on Carbopack B-DA). Amount and composition of thebiogas produced within the anaerobic stage were determinedthrough a Mariotte system and by GC-TCD, respectively, asreported elsewhere.22 OME total and volatile suspended solids(TSS and VSS, respectively), along with volatile suspended solidsrelated to the aerobic stages, where determined accordingto standard methods.23 Total OME nitrogen was measuredaccording to Kjeldhal method. OME phenols concentration werespectrophotometrically analyzed following the Folin-Chocalteuprocedure.22
For PHA determination, the sludge sample was treatedimmediately with a NaClO solution (7% of active Cl2). PHAswere extracted, hydrolysed and esterified to 3-hydroxyacyl methylesters, and determined by gas chromatography.24 The storedpolymer was the copolymer P(HB/HV), and the relative abundanceof HB and HV monomers were quantified by using a polymerP(HB/HV) of known HV content, as standard. The non-polymerbiomass was calculated from the difference between VSS and PHAin the sample and converted into COD according to a conversionfactor of 1.42 mgCOD mg−1 biomass. PHAs were converted intoCOD according to oxidation stoichiometry: 1.67 mgCOD mg−1
PHB, and 1.92 mgCOD mg−1 PHV.In the SBR run with synthetic feed, specific PHAs production
rate was calculated as the ratio of stored PHA and the length of thefeast phase, per unit of non-polymer biomass. The storage yieldduring the feast phase, was determined as the ratio between theamount of produced PHA (as COD) and the amount of removedsubstrate (as COD) at the substrate depletion. On the other hand,in the SBR run with OMEs, both PHA storage rate and yield werecalculated after 10 min from the beginning of the cycle, whenmost of the readily biodegradable COD had been removed andbefore that COD removal rate started to decrease.
In the tests carried out at increasing loads of fermentedOME, total substrate removal rate and PHA storage rate werecalculated by linear regression of concentrations vs time dataand were expressed as specific rates with respect to the initialbiomass concentration. The PHA content of the biomass wascalculated by dividing the measured PHA concentration bybiomass concentration (both expressed as COD).
Molecular techniquesTotal DNA extraction from SBR sludge samples was carriedout using the FAST DNA Spin Kit for Soil (MO BIO, Carlsbad,CA) according to the manufacturer’s instructions. Approximately0.5 mL of material was used per extraction and the extracted DNAwas polymerase chain reaction (PCR) amplified and then analyzedby denaturing gradient gel electrophoresis (DGGE).
The whole 16S rDNA was first amplified using primers F8 andR11.25 Then a nested PCR was performed starting from 16S rDNAamplicons obtained from the first amplification. In this second PCRreaction, the iper-variable V3 region of the 16S rRNA gene wasamplified using primers p3 (with a GC clamp) and p2.26 The PCR
J Chem Technol Biotechnol 2009; 84: 901–908 c© 2009 Society of Chemical Industry www.interscience.wiley.com/jctb

90
4
www.soci.org M Beccari et al.
reactions master mix composition and temperature conditionsfor PCR were described by Zocca et al.27 The PCR products werequantified using Low DNA Mass Ladder (Celbio, Italy) in a 2.0%agarose gel.
DGGE analyses were carried out on V3 region amplicons. Gels(8% acrylamide/bisacrylamide 19 : 1, BioRad) were cast using adenaturing gradient of 30–60%, with 100% denaturant definedas 7 mol L−1 urea and 20% (v/v) formamide. Electrophoresis wasperformed at 50 V for 16 h at 65 ◦C with the Dcode UniversalDetection System (Biorad) and gels were stained with EtBr (1 mgL−1). The PCR products corresponding to the different time pointssampled for both treatments were loaded in the same DGGE gel.Equal masses of the PCR products were loaded in each lane toallow for a semi-quantitative comparison among lanes.
DGGE bands containing DNA to be sequenced were excisedand incubated for 4 h in 100 µL of sterile water. Afterwards, PCRamplification was carried out as described before, except for theuse of non-GC-clamped primers. PCR products were transformedin E. coli DH5α using the pGEM-T vector system following themanufacturer’s instructions (Promega, Italy), sequenced on bothstrands, and subsequently searched for homology using theBLASTN database.28
RESULTS AND DISCUSSIONPerformance of the first anaerobic stage with OMEsPBBR technology was chosen to develop the anaerobic stage,since immobilized cells systems allow reduction of the risks ofshock loading and/or washout problems, which can negativelyaffect the productivity of suspended growth reactors operatingwith low-growth rate biomass and high and fluctuating organicloads, as typically happens in the anaerobic digestion of OMEs.22
Temperature and OLR applied in this research (25 ◦C and 26.2 gCODL−1 d−1, respectively) were chosen on the basis of preliminarywork carried out with the same OME and technology. Suchparameters were the ones which corresponded to the highestVFA production (data not shown). The low amount of OME solidsallowed operation without any problems. The COD concentrationof the fermented OMEs was of 33.9 g L−1, and very close tothe influent one (36.9 g L−1), indicating that only a small fraction(about 8%) of the influent substrate was converted into biogas andbiomass growth. No significant phenols depletion was observedthroughout the experiment. Such compounds, once fed to theaerobic stages, could inhibit the metabolic activity of the PHA-producing consortium. On the other hand, phenols are consideredadded value substances: thus, the possibility of recovering thephenolic fraction from the anaerobic effluent before it is fed tothe aerobic stages could represent an interesting task for furtherstudies. The VFA concentration in the effluent was 10.7 gCOD L−1;therefore, the anaerobic fermentation stage increased the VFApercentage in the OME from about 18% to about 32% (Table 1).pH slightly decreased to 5.2, still compatible with acidogenicfermentation processes.
The acetic, butyric and propionic acid percentages in the VFAmixture produced were 62%, 22% and 12%, respectively. As shownin Table 1, fermented OME also contained alcohols, accounting forabout 22% of the overall COD; namely ethanol (∼80% of theoverall alcohols), butanol (∼13%), and methanol (∼7%).
Performance of the second aerobic stage with fermented OMEsThe SBR reactor was operated for a period of approximately1 month with the synthetic feed of acetic and propionic acid.
Table 1. Operating conditions and performance of the first stage ofthe process (given intervals refer to standard deviation of the mean)
Parameter Value
HRT (day) 1.41
CODin (gCOD L−1) 36.9 ± 1.5
VFAin (gCOD L−1) 6.75 ± 0.60
R-OHin (gCOD L−1) N.A.
CODout (gCOD L−1) 33.9 ± 0.53
VFAout (gCOD L−1) 10.7 ± 0.45
VFAout/CODout (COD COD−1, %) 31.6 ± 1.4
R-OHout (gCOD L−1) 7.3 ± 0.24
R-OHout/CODout (COD COD−1, %) 21.6 ± 0.62
R-OH: total alcohols; N.A. not available
0500
100015002000250030003500400045005000
0 5 10 15 20 25 30
Time (d)
VSS
(m
g L
-1)
A
0
200
400
600
800
1000
1200
1400
0 5 10 15 20 25 30
Time (d)
PHA
(m
gCO
D L
-1)
end of the feast phaseend of the cycle
B
Figure 2. Biomass (A) and polymer (B) concentrations during the SBR runwith the synthetic mixture of acetic and propionic acid.
After an initial period of fluctuations of about 8 days, the VSSconcentration in the SBR, measured at the time of substratedepletion (end of the feast phase), stabilized at an average valueof 3137 ± 99 mg L−1(Fig. 2(A)), giving a biomass productivity of3137 mg L−1 d−1. Similar initial fluctuations were also observedfor PHAs concentrations measured at the end of the feast phaseand at the end of the cycle (Fig. 2(B)). Subsequently, the PHAsconcentrations also stabilized, with the PHA concentration atthe end of the feast phase being always higher than at theend of the cycle. This indicated that the polymer was producedduring the feast phase and consumed during the famine phase.During the SBR feast phase the average values of specificPHA storage rate and yield were 175 ± 22 mgCOD gCOD−1 h−1,and 0.36 ± 0.030 COD COD−1, respectively. This finding clearly
www.interscience.wiley.com/jctb c© 2009 Society of Chemical Industry J Chem Technol Biotechnol 2009; 84: 901–908

90
5
Bioproduction of PHA from olive mill effluents www.soci.org
0500
100015002000250030003500400045005000
0 5 10 15 20 25 30 35
Time (d)
VSS
(m
g L
-1)
end of the feed phase30 min. from the beginning of the cycleend of the cycle
A
0
50
100
150
200
250
300
0 5 10 15 20 25 30 35
Time (d)
PHA
(m
gCO
D L
-1)
end of the feed phase30 min. from the beginning of the cycleend of the cycle
B
Figure 3. Biomass (A) and polymer (B) concentrations during the SBR runwith the fermented OMEs.
indicated that biomass with a high storage response was enrichedin the SBR.
In the following run, the SBR feed was switched to the fermentedOMEs, which were diluted in order to operate the SBR at the sameOLR (8.5 gCOD L−1 d−1). It is worth noting that in the fermentedOMEs, VFAs were about 32% of the overall COD; accordingly,the OLR referred only to VFAs was significantly lower than inthe previous run and about 2.7 gCOD L−1 d−1. Also under theseconditions, the reactor was operated for about 1 month.
During this run, the VSS concentrations in the SBR, measuredat the end of the feed phase, at 30 min from the beginning ofthe cycle, and at the end of the cycle were very similar and about3722 ± 146 mg L−1 (Fig. 3(A)). Accordingly, the correspondingbiomass productivity was about 3722 mg L−1 d−1. The slightlyhigher VSS concentration obtained with the fermented OME thanwith the synthetic feed might be due to the different compositionof the substrate and therefore to the occurrence of differentmechanisms of substrate removal.
Figure 3(B) shows the concentration of PHAs during the SBR runwith the fermented OMEs, in the reactor at different times duringthe cycle. Also in this case the PHA concentration both at 10 min(end of the feed phase) and at 30 min from the beginning of thecycle was higher than at the end of the cycle, confirming that thepolymer was stored during the feast phase and consumed duringthe following famine phase. This finding indicates that, even ifVFAs were only a fraction of the overall COD of the fermentedOMEs, a selective pressure was maintained in the SBR towards theenrichment of mixed cultures able to store PHAs. It is worth notingthat during a typical SBR cycle the maximum PHA concentrationwas reached after 10 min (end of the feed phase) when about 50%of the overall COD was removed.
Table 2. Performance of the second aerobic stage on synthetic VFAmixture and on fermented OMEs (given intervals refer to standarddeviation of the mean)
ParameterSynthetic mixture
of VFAFermented
OMEs
Influent concentration(gCOD L−1)
8.5 8.5
Overall OLR (gCOD L−1 d−1) 8.5 8.5
OLRVFA (gCOD L−1 d−1) 8.5 2.7
VSS (mg L−1) 3137 ± 99 3722 ± 146
Storage rate (mgCODgCOD−1 h−1)
175 ± 22 146 ± 12
Storage yield (COD COD−1) 0.36 ± 0.030 0.36 ± 0.035
Effluent concentration(gCOD L−1)
0 1.3 ± 0.04
Removed COD (%) 100 85 ± 1
0
1000
2000
3000
4000
5000
6000
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5
Time (h)
Rem
oved
CO
D a
nd F
orm
ed P
HA
(mgC
OD
L-1
)
Removed CODFormed PHA
Figure 4. Time course of the removed substrate and the stored polymerduring the test carried out with the fermented OME at 9 times the usualfeed load in a single SBR cycle.
Maximum PHA storage rate and yield obtained duringthis SBR run were 146 ± 12 mgCOD gCOD−1 h−1 and 0.36 ±0.035 COD COD−1, respectively. It is important to note that suchvalues were very similar to those obtained with the synthetic mix-ture of VFAs, thus indicating that the composition of the fermentedOME was suitable for the enrichment of a biomass with a high stor-age capacity. Moreover, the SBR effluent contained a residual CODof approximately 1.3 g L−1, indicating that the process resulted ina significant reduction (85%) of the polluting load of the OMEs.
A summary of the performance of the second aerobic stage onboth the synthetic feed and the fermented OMEs is reported inTable 2.
Performance of the third aerobic stage with fermented OMEsTo simulate the third stage of the process, tests were carried outwith the fermented OMEs by feeding the SBR at several loads corre-sponding to 1, 3, and 9 times its usual load. Figure 4, shows profilesof the removed COD and the stored PHA in the reactor during thetest carried out at the highest load investigated. During the initial2.5 h, the substrate was rapidly consumed at a constant rate andpolymer was produced almost linearly, reaching the maximumconcentration of about 1620 mgCOD L−1. Subsequently, a sharpdecrease in substrate removal rate was observed and the PHAconcentration started to decrease. Probably, the initial faster COD
J Chem Technol Biotechnol 2009; 84: 901–908 c© 2009 Society of Chemical Industry www.interscience.wiley.com/jctb
90
6
www.soci.org M Beccari et al.
0
300
600
900
1200
1500
1800
0 1000 2000 3000 4000 5000 6000 7000
Total Substrate fed (mgCOD L-1)
Max
imum
PH
Aco
ncen
trat
ion
(mgC
OD
L-1
)
0.00
0.10
0.20
0.30
0.40
0.50
0.60
Stor
age
Yie
ld (
CO
D C
OD
-1)PHA
Storage yield
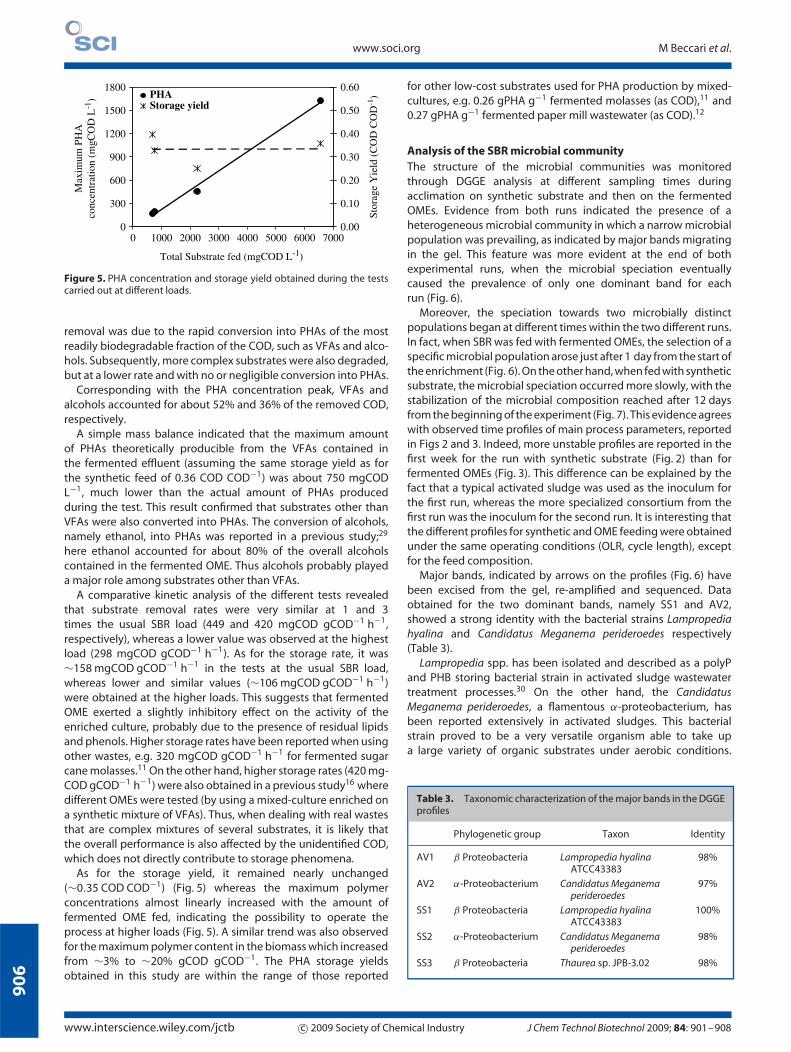
Figure 5. PHA concentration and storage yield obtained during the testscarried out at different loads.
removal was due to the rapid conversion into PHAs of the mostreadily biodegradable fraction of the COD, such as VFAs and alco-hols. Subsequently, more complex substrates were also degraded,but at a lower rate and with no or negligible conversion into PHAs.
Corresponding with the PHA concentration peak, VFAs andalcohols accounted for about 52% and 36% of the removed COD,respectively.
A simple mass balance indicated that the maximum amountof PHAs theoretically producible from the VFAs contained inthe fermented effluent (assuming the same storage yield as forthe synthetic feed of 0.36 COD COD−1) was about 750 mgCODL−1, much lower than the actual amount of PHAs producedduring the test. This result confirmed that substrates other thanVFAs were also converted into PHAs. The conversion of alcohols,namely ethanol, into PHAs was reported in a previous study;29
here ethanol accounted for about 80% of the overall alcoholscontained in the fermented OME. Thus alcohols probably playeda major role among substrates other than VFAs.
A comparative kinetic analysis of the different tests revealedthat substrate removal rates were very similar at 1 and 3times the usual SBR load (449 and 420 mgCOD gCOD−1 h−1,respectively), whereas a lower value was observed at the highestload (298 mgCOD gCOD−1 h−1). As for the storage rate, it was∼158 mgCOD gCOD−1 h−1 in the tests at the usual SBR load,whereas lower and similar values (∼106 mgCOD gCOD−1 h−1)were obtained at the higher loads. This suggests that fermentedOME exerted a slightly inhibitory effect on the activity of theenriched culture, probably due to the presence of residual lipidsand phenols. Higher storage rates have been reported when usingother wastes, e.g. 320 mgCOD gCOD−1 h−1 for fermented sugarcane molasses.11 On the other hand, higher storage rates (420 mg-COD gCOD−1 h−1) were also obtained in a previous study16 wheredifferent OMEs were tested (by using a mixed-culture enriched ona synthetic mixture of VFAs). Thus, when dealing with real wastesthat are complex mixtures of several substrates, it is likely thatthe overall performance is also affected by the unidentified COD,which does not directly contribute to storage phenomena.
As for the storage yield, it remained nearly unchanged(∼0.35 COD COD−1) (Fig. 5) whereas the maximum polymerconcentrations almost linearly increased with the amount offermented OME fed, indicating the possibility to operate theprocess at higher loads (Fig. 5). A similar trend was also observedfor the maximum polymer content in the biomass which increasedfrom ∼3% to ∼20% gCOD gCOD−1. The PHA storage yieldsobtained in this study are within the range of those reported
for other low-cost substrates used for PHA production by mixed-cultures, e.g. 0.26 gPHA g−1 fermented molasses (as COD),11 and0.27 gPHA g−1 fermented paper mill wastewater (as COD).12
Analysis of the SBR microbial communityThe structure of the microbial communities was monitoredthrough DGGE analysis at different sampling times duringacclimation on synthetic substrate and then on the fermentedOMEs. Evidence from both runs indicated the presence of aheterogeneous microbial community in which a narrow microbialpopulation was prevailing, as indicated by major bands migratingin the gel. This feature was more evident at the end of bothexperimental runs, when the microbial speciation eventuallycaused the prevalence of only one dominant band for eachrun (Fig. 6).
Moreover, the speciation towards two microbially distinctpopulations began at different times within the two different runs.In fact, when SBR was fed with fermented OMEs, the selection of aspecific microbial population arose just after 1 day from the start ofthe enrichment (Fig. 6). On the other hand, when fed with syntheticsubstrate, the microbial speciation occurred more slowly, with thestabilization of the microbial composition reached after 12 daysfrom the beginning of the experiment (Fig. 7). This evidence agreeswith observed time profiles of main process parameters, reportedin Figs 2 and 3. Indeed, more unstable profiles are reported in thefirst week for the run with synthetic substrate (Fig. 2) than forfermented OMEs (Fig. 3). This difference can be explained by thefact that a typical activated sludge was used as the inoculum forthe first run, whereas the more specialized consortium from thefirst run was the inoculum for the second run. It is interesting thatthe different profiles for synthetic and OME feeding were obtainedunder the same operating conditions (OLR, cycle length), exceptfor the feed composition.
Major bands, indicated by arrows on the profiles (Fig. 6) havebeen excised from the gel, re-amplified and sequenced. Dataobtained for the two dominant bands, namely SS1 and AV2,showed a strong identity with the bacterial strains Lampropediahyalina and Candidatus Meganema perideroedes respectively(Table 3).
Lampropedia spp. has been isolated and described as a polyPand PHB storing bacterial strain in activated sludge wastewatertreatment processes.30 On the other hand, the CandidatusMeganema perideroedes, a flamentous α-proteobacterium, hasbeen reported extensively in activated sludges. This bacterialstrain proved to be a very versatile organism able to take upa large variety of organic substrates under aerobic conditions.
Table 3. Taxonomic characterization of the major bands in the DGGEprofiles
Phylogenetic group Taxon Identity
AV1 β Proteobacteria Lampropedia hyalinaATCC43383
98%
AV2 α-Proteobacterium Candidatus Meganemaperideroedes
97%
SS1 β Proteobacteria Lampropedia hyalinaATCC43383
100%
SS2 α-Proteobacterium Candidatus Meganemaperideroedes
98%
SS3 β Proteobacteria Thaurea sp. JPB-3.02 98%
www.interscience.wiley.com/jctb c© 2009 Society of Chemical Industry J Chem Technol Biotechnol 2009; 84: 901–908

90
7
Bioproduction of PHA from olive mill effluents www.soci.org
Figure 6. DGGE profiles of SBR fed with synthetic substrate (Panel A) or with the real effluent from the OMEs fermentation stage (Panel B). Numbers atthe top of each lane indicate the sampling time in days.
Figure 7. DGGE profiles of SBR fed with synthetic substrate. Numbers atthe top of each lane indicate the sampling time in days.
Moreover, it has been shown to have a remarkably high storagecapacity, forming polyhydroxyalkanoates from a wide variety ofsubstrates.31
CONCLUSIONSThe aim of this work was to investigate the possibility of using non-pretreated OMEs as a renewable resource for PHAs production ina three-stage process. The first fermentative stage was carriedout in a packed-bed biofilm reactor using alumina-based high-porous ceramic cubes as packing material, at 25 ◦C and 26.2 gCODL−1 d−1. Under these conditions, the percentage of VFAs (the mostdirect substrates for PHAs production), in the OMEs increased fromabout 18% to about 32%. Moreover, the fermented effluent alsocontained other substrates easily convertible into PHAs, such asalcohols, which represented about 22% of the overall COD. The
second stage was carried out in a SBR, inoculated with activatedsludge from a wastewater treatment plant, at an overall OLR of 8.5gCOD L−1 d−1 (corresponding to an OLR of 2.7 gCOD L−1 d−1 interms of VFAs only). Even if VFAs were only a fraction of the overallCOD, selective pressure on the enrichment of mixed cultures ableto store PHAs was maintained in this stage. The maximum valuesof PHA storage rate and yield with the fermented OMEs were about146 mgCOD gCOD−1 h−1, and 0.36 COD COD−1 respectively, verysimilar to those obtained using a synthetic mixture of acetic andpropionic acid. The SBR reactor also allowed a significant reductionof the polluting load of the fermented effluent, with a COD removalefficiency of about 85%. The biomass enriched during each SBR runwas monitored through DGGE analysis at different sampling times.Data obtained showed that the different composition of substratesfed to the reactor strongly influenced the selection of microbialpopulations responsible for PHA storage. Lampropedia hyalinaand Candidatus Meganema perideroedes were the predominantbacterial strains enriched with the synthetic mixture of VFA or thefermented OME, respectively.
Finally, the third stage was investigated by means of tests atincreasing loads (up to about 9 times the load of a single SBR cycle).The maximum concentration of PHAs produced increased almostlinearly with the amount of fermented OMEs fed, indicating thepossibility to operate the process at higher OLR. These tests alsorevealed a significant contribution to the PHA storage of substratesother than VFAs, mostly alcohols and especially ethanol.
Overall, the proposed process is effective for the simultaneoustreatment of OMEs and their valorization as a renewable resourcefor PHA production.
AKNOWLEDGEMENTSThis research was supported financially by the Italian Ministry ofUniversity and Research (PRIN 2005). The Authors L. Bertin and F.Fava acknolewdge the FONDAZIONE DEL MONTE DI BOLOGNA ERAVENNA (Bologna, Italy) for the economical support provided totheir Unit.
J Chem Technol Biotechnol 2009; 84: 901–908 c© 2009 Society of Chemical Industry www.interscience.wiley.com/jctb

90
8
www.soci.org M Beccari et al.
REFERENCES1 Reddy CSK, Ghai R, Rashmi and Kalia VC, Polyhydroxyalkanoates: an
overview. Biores Technol 87:137–146 (2003).2 Lee SY, Bacterial polyhydroxyalkanoates. Biotechnol Bioeng 49:1–14
(1996).3 Mino T, Van Loosdrecht MCM and Heijnen JJ, Microbiology and
biochemistry of the enhanced biological phosphate removalprocess. Water Res 32:3193–3207 (1998).
4 Beccari M, Majone M, Massanisso P and Ramadori R, A bulking sludgewith high storage response selected under intermittent feeding.Water Res 32:3403–3413 (1998).
5 Chua ASM, Takabatake H, Satoh H and Mino T, Production ofpolyhydroxyalkanoates (PHA) by activated sludge treatingmunicipal wastetwater: effect of pH, sludge retention time (SRT),and acetate concentration in the influent. Water Res 37:3602–3611(2003).
6 Reis MAM, Serafim LS, Lemos PC, Ramos AM, Aguiar FR and VanLoosdrecht MCM, Production of polyhydroxyalkanoates by mixedmicrobial cultures. Bioprocess Biosystems Eng 25:377–385 (2003).
7 Van Loosdrecht MCM, Pot M and Heijnen JJ, Importance of bacterialstorage polymers in bioprocesses. Water Sci Technol 35:41–47(1997).
8 Wilderer PA, Irvine RL and Goronszy MC, Sequencing Batch ReactorTechnology. Scientific and Technical Report No.10. IWA Publishing,London (2001).
9 Dionisi D, Majone M, Papa V and Beccari M, Biodegradable polymersfrom organic acids by using activated sludge enriched by aerobicperiodic feeding. Biotechnol Bioeng 85:569–579 (2004).
10 Dionisi D, Majone M, Vallini G, Di Gregorio S and Beccari M, Effect of theapplied organic load rate on biodegradable polymer production bymixed microbial cultures in a sequencing batch reactor. BiotechnolBioeng 93:76–88 (2006).
11 Albuquerque MGE, Eiroa M, Torres C, Nunes BR and Reis MAM,Strategies for the development of a side stream process forpolyhydroxyalkanoate (PHA) production from sugar cane molasses.J Biotechnol 130:411–421 (2007).
12 Bengtsson S, Werker A, Christensson M and Welander T, Productionof polyhydroxyalkanoates by activated sludge treating paper millwastewater. Biores Technol 99:509–516 (2008).
13 Ghatnekar MS, Pai JS and Ganesh M, Production and recovery of poly-3-hydrxybutyrate from Methylobacterium sp V49. J Chem TechnolBiotechnol 77:444–448 (2002).
14 Hahn SK, Chang YK, Kim BS and Chang HN, Optimization ofmicrobial poly(3-hydroxybutyrate) recover using dispersions ofsodium hypochlorite solution and chloroform. Biotechnol Bioeng44:256–261 (1994).
15 Choi JC and Lee SY, Efficient and economical recovery of poly (3-hydroxybutyrate) from recombinant Escherichia Coli by simpledigestion with chemicals. Biotechnol Bioeng 62:546–553 (1999).
16 Dionisi D, Carucci G, Petrangeli Papini M, Riccardi C, Majone M andCarrasco F, Olive oil mill effluents as a feedstock for production ofbiodegradable polymers. Water Res 39:2076–2084 (2005).
17 Hassan MA, Shirai N, Kusubayashi N, Abdul Karim MI, Nakanishi K andHashimoto K, Effect of organic acid profiles during anaerobic
treatment of palm oil mill effluent on the production ofpolyhydroxyalkanoates by Rhodobacter sphaeroides. J FermentBioeng 82:151–156 (1996).
18 Hassan MA, Shirai N, Kusubayashi N, Abdul Karim MI, Nakanishi Kand Hashimoto K, The production of polyhydroxyalkanoates frompalm oil mill effluent by Rhodobacter spheroides. Ferment Bioeng83:485–488 (1997).
19 Hassan MA, Shirai N, Umeki H, Abdul Karim MI, Nakanishi K andHashimoto K, Acetic acid separation from anaerobically treatedpalm oil mill effluent for the production of polyhydroxyalkanoateby Alcaligenes eutrophus. Biosci Biotechnol Biochem 61:1465–1468(1997).
20 Meesters KHP, Production of poly (3 hydroxyalkanoates) from wastestreams. Report of Technical University of Delft (1998).
21 Salehizadeh H and Van Loosdrecht MCM, Production ofpolyhydroxyalkanoates by mixed cultures: Recent trends andbiotechnological importance. Biotechnol Adv 22:261–279 (2004).
22 Bertin L, Colao MC, Ruzzi M and Fava F, Performances and microbialfeatures of a granular activated carbon packed-bed biofilm reactorcapable of an efficient anaerobic digestion of olive mill wastewaters.FEMS Microbiol Ecol 48:413–423 (2004).
23 APHA, Standard Methods for the Examination of Water and Wastewater,19th edn. American Public Health Association, Washington DC(1995).
24 Braunegg G, Sonnleitner B and Lafferty RM, A rapid gaschromatographic method for the determination of poly-ß-hydroxybutyric acid in microbial biomass. Eur J Appl Microbiol6:29–37 (1978).
25 Weisburg WG, Berns SM, Pelletier DA and Lane DJ, 6 16S ribosomalDNA amplification for phylogenetic study. J Bacteriol 173:697–703(1991).
26 Muyzer G, de Waal EC and Uitterlinden AG, Profiling of complexmicrobial populations by denaturing gradient gel electrophoresisanalysis of polymerase chain reaction-amplified genes coding for16S rRNA. Appl Environ Microbiol 59:695–700 (1993).
27 Zocca C, Di Gregorio S, Visentini F and Vallini G, Biodiversity amongstcultivable polycyclic aromatic hydrocarbon-transforming bacteriaisolated from an abandoned industrial site. FEMS Microbiol Lett238:375–382 (2004).
28 Altshul SF, Madden TL, Shaffer AA, Zhang J, Zhang Z, Miller W, et al,Gapped BLAST and PSI-BLAST: a new generation of protein databasesearch programs. Nucleic Acids Res 25:3389–3402 (1997).
29 Dionisi D, Levantesi C, Renzi V, Tandoi V and Majone M, PHA storagefrom several substrates by different morphological types in ananoxic/aerobic SBR. Water Sci Technol 46:337–344 (2002).
30 Mino T, Microbial selection of polyphosphate-accumulating bacteriain activated sludge wastewater treatment processes for enhancedbiological phosphate removal. Biochemistry (Moscow) 65:341–348(2000).
31 Kragelund C, Nielsen JL, Thomsen TR and Nielsen PH, Ecophysiologyof the filamentous Alphaproteobacterium Menagema perideroedesin activated sludge. FEMS Microbiol Ecol 54:111–122 (2005).
www.interscience.wiley.com/jctb c© 2009 Society of Chemical Industry J Chem Technol Biotechnol 2009; 84: 901–908





























