Embed Size (px)
Citation preview

Plant Physiol. (1970) 45, 339-347
Events Surrounding the Early Development ofEuglena ChloroplastsI. INDUCTION BY PREILLUMINATION'
Received for publication September 26, 1969
ANDREW W. HOLOWINSKY2, I AND JEROME A. SCHIFFDepartment of Biology, Brandeis University, Waltham, Massachusetts 02154
ABSTRACT
Preillumination, followed by a dark period prior to ex-posure of dark-grown nondividing cells of Euglena gracilisvar. bacillaris to normal lighting conditions for chloroplastdevelopment, results in potentiation, or abolishment of theusual lag in chlorophyll accumulation. The degree ofpotentiation is a function of the length of the preillumina-tion period, the intensity of preilluminating light, and thelength of the dark period interposed before re-exposure tocontinuous light for development. The optimal conditionsare found to be: 90 minutes of preillumination with whitelight at an intensity greater than 30 microwatts per squarecentimeter (14 foot candles) followed by a dark period of atleast 12 hours. Reciprocity is not found between durationand intensity of preilluminating light. Preilluminationwith blue light and red light was found to be the most effec-tive in promoting potentiation, and the ratio of effectivenessof blue to green to red is consistent with protochlorophyll-(ide) being the photoreceptor. Although red light is effec-tive, there is no reversal by far red light, and these facts,taken together with the effectiveness of blue light, suggestthat the phytochrome system is not involved. The amount ofchlorophyll formed at the end of preillumination is propor-tional to the resulting potentiation, suggesting that theamount of protochlorophyll(ide) removed or chlorophyll-(ide) formed regulates this phenomenon. Potentiated andnonpotentiated cells show comparable rates of proto-chlorophyll (ide) resynthesis, suggesting that this is not thelimiting factor in nonpotentiated cells. Although light isrequired for protochlorophyll (ide) conversion in chlorophyllsynthesis, a brief preillumination seems also to initiate theproduction of components in the subsequent dark periodwhich, in nonpotentiated cells, are ordinarily synthesizedduring the lag period under continuous illumination. Thesecomponents are necessary to sustain maximal rates of sub-sequent chlorophyll accumulation.
Chlorophyll synthesis during chloroplast development in dark-grown seedlings of higher plants, induced by continuous illumina-
1 Work supported by Research Grant GM 14595 from the NationalInstitutes of Health.
1 Investigations reported on in this paper were conducted while thesenior author was on research leave from Harvard University.
3Present address: Division of Biological and Medical Sciences,Brown University, Providence, Rhode Island 02912.
tion, shows a 4- to 6-hr lag in the onset of rapid chlorophyll ac-cumulation (47). The existence and extent of this lag period inbean plants are dependent on the age and nutritional condition ofthe seedlings (40, 50). Over 10 years ago, Withrow et al. (49)using bean plants and later Virgin (44) working with wheatseedlings showed that this lag could be eliminated by exposing theetiolated leaves to a brief period of illumination followed by adark period prior to their re-exposure to continuous light. Inmost systems the optimal length of the dark period was found tocorrespond approximately in length to the lag period which isnormally observed in the absence of any experimental pretreat-ment (29, 44). Red light has been shown to be effective for preil-lumination (44, 46, 49), and in beans (32) reversibility of the redeffect by far red light has been clearly demonstrated, therebyimplicating phytochrome in this phenomenon. Phytochrome wasalso shown to regulate other parameters of chloroplast develop-ment in addition to its apparent effect on chlorophyll synthesis.Mego and Jagendorf (28) observed phytochrome control ofchloroplast size and protein nitrogen content. Klein et al. (21)and Holowinsky and O'Brien (unpublished), however, found noclear far red reversal of the red light induction of some structuralchanges during chloroplast development. The synthesis of severalphotosynthetic enzymes has been shown to be triggered or medi-ated by the phytochrome system (12, 18, 26, 27).
Since, in the higher plant system, phytochrome also influencesnumerous other morphogenetic and developmental responses(11), it is not clear whether its participation in lag elimination isdirect or whether it affects chloroplast development through moregeneral cellular responses. In the higher plant system, the lagelimination can be accomplished by other means (20, 40, 50),and, therefore, the developmental events associated with the lagperiod appear not to strictly require light stimulation. It is con-ceivable that phytochrome-mediated lag elimination could beexerted through phytochrome regulation of some nonchloroplastnutritional or developmental events in the leaf tissue. The in-crease in diameter of the chloroplast in the course of plastid de-velopment, for instance, is one such parameter which appears tobe closely linked to increase in cell size (16).To learn more about the control of early chloroplast develop-
ment, we turned to Euglena where we sought to reproduce thephenomenon of light-induced lag elimination. Euglena affords anexperimental system under defined nutrition without the multi-cellular complications found in higher plants.Dark-grown resting cells of Euglena, exposed to continuous
white light, exhibit a 10- to 12-hr lag period before the onset ofrapid chlorophyll accumulation (42). While chlorophyll is syn-thesized at a low rate during the lag period, the major accumula-tion occurs from the onset of the maximal rate at 12 hr until thecompletion of development at 72 hr. The earliest events whichcan be detected are the initial conversion of protochlorophyl-
339

HOLOWINSKY AND SCHIFF
1 (ide)4 to chlorophyll(ide) (31, 38), the large increase in respira-tory rate (35), and the formation of chloroplast membranes, allof which are associated with light induction (4, 36). Net photo-synthetic oxygen evolution and carbon dioxide fixation maketheir appearance at about 4 hr of development, at which time thefirst lamellae are also organized (4, 42). In general, all parametersof development, including the formation of several photosyn-thetic enzymes (36), are in step with one another and with pig-ment formation from 12 hr to the completion of development.
In this paper we explore the influence of preillumination onpigment formation in Euglena and show that induction of chloro-plast development can be achieved by a brief exposure to light.Such exposure triggers subsequent developmental events whichdo not necessarily require continuous light. The light-inducedstep is shown not to be related to the red-far red system.A preliminary report of this work has already appeared (17).
MATERIALS AND METHODS
Euglena gracilis Klebs var. bacillaris Pringsheim, cultured formany generations in the dark as previously described (25, 42),was used as the innoculum in the experiments to be described. Allmedia were sterilized by autoclaving as before (25, 42), and allmanipulations of dark-grown cells were performed under dimgreen safelights (25). For each experiment, 5-day-old dark-growncells (10 ml) on Hutner's pH 3.5 medium (13) were innoculatedinto 1 liter of Hutner's pH 8.0 medium (13). After 3 days ofgrowth in the dark on a shaker at 26 C, as previously described(25), 200 ml were aseptically mixed with 500 ml of pH 6.8 restingmedium and were returned to the shaker for 3 days to allow thecells to cease division (42). Resting medium at pH 6.8 (Changand Schiff, unpublished) is prepared by autoclaving mannitol(1.0%, w/v) in 0.01 M potassium phosphate buffer, pH 6.8. Toavoid precipitation, magnesium chloride (1.0 M) is autoclavedseparately and enough is added to the preceding solution whencool to yield a final concentration of 0.01 M MgCl2 in the restingmedium. Resting medium of a higher than usual pH (41) wasselected for the present experiments to preclude pheophytinformation from chlorophyll in the dark period following preil-lumination (see Greenblatt and Schiff [13]). In any particularexperiment several resting cultures were pooled prior to aliquot-ing for the various treatments.
Conditions for normal chloroplast development and for post-illumination were the same as described previously (41, 51):150 ft-c of white light (provided by a mixture of white and redfluorescent tubes) illuminated resting dark-grown cultures on arotary shaker at 26 C. Preillumination was achieved with thesame conditions or with a 150-w Sylvania Spot-Lite, filtered byabout 2 inches of water, which delivered high light intensitiesto the bottom of the flask when necessary. For preilluminationwith colored light, the cultures (20 ml) were contained in 50-mlErlenmeyer flasks, which were placed in opaque containers witha Wratten filter screening L bottom aperture. Three filters wereemployed to divide the spectrum into three fairly equal portions:400 to 500 nm (Wratten No. 98); 480 to 600 nm (Wratten No.61); and 610 to 700 nm (Wratten No. 29) (22). The containerswere placed over a bank of Sylvania fluorescent tubes (daylight,in the case of blue and green illumination, and red, in the caseof 610-700 nm). Energy was measured with an Eppley linear 12-junction thermophile with bismuth-silver couples amplified by aListon Becker model 14 D.C. breaker amplifier and displayed ona Texas Instruments recorder (37). Light intensities were alsomeasured by means of a Weston Sunlight illumination meter and
4 It is not known whether protochlorophyllide or protochlorophyllis the photochemically transformed precursor of chlorophyll in dark-grown cells. The designation here of (ide) reflects this uncertainty. Afuller discussion of this problem is found in Schiff and Epstein (36).
expressed in terms of foot candles. Energy was adjusted by inter-posing neutral density screens between the containers and thelight source. In experiments where red or far red light was givenin preillumination, cultures under red illumination were exposedto Sylvania red fluorescent tubes with the ends taped to preventcontamination by non red light where the phosphor coating be-comes unreliable. Far red illumination was provided by a Syl-vania 150-w Spot-Lite filtered through 2 inches of water and aRohm and Haas Plexiglass V-58015 filter. Unless otherwise indi-cated, all incubations took place at 26 C.
Cell counts were made in a Coulter model A cell counter in0.4% (w/v) NaCl solution after suitable dilution (51). Chloro-phyll was determined as previously described (42), with appro-priate adjustments in the volume of acetone as demanded by cellnumber and chlorophyll concentration.
Protochlorophyll(ide) synthesis in intact cells was measuredwith a Biospect model 61, a commercial version of the Norris-Butler spectrophotometer (8, 9), produced by Agricultural Spe-cialties Co., Bethesda, Md. Cell suspensions of suitable density(approximately 1 X 108 cells/ml) were illuminated in a thin layerfor 3 min with white light of approximately 300 ft-c by the Spot-Lite source described above. The suspension was then returnedto darkness and maintained at 26 C in a water bath. Aliquotswere removed with time, frozen in cylindrical cuvettes to liquidnitrogen temperatures, and scanned in the Norris-Butler spectro-photometer. The base line has been previously adjusted to be flatwith a sample of calcium carbonate suspension. Typical scansare shown in Figure 1. Since the protochlorophyll(ide) to chlo-rophyll(ide) conversion in Euglena involves the conversion ofPC635 and since newly formed chlorophyll(ide) does not undergoany dramatic spectral shifts in this organism (7), the increase inabsorption at 635 nm was taken as an estimate of the resynthesisof protochlorophyll in the dark after light exposure. To calculate
WAVELENGTH (nM)FIG. 1. Absorption spectra of dark-adapted Euglena after 3 days on
resting medium. Each trace represents the spectrum obtained with 2ml of suspension containing 2.1 X 108 cells cooled to liquid nitrogentemperature. The positions of the 635 nm and the 675 nm calibrationcorresponding to the expected positions of the absorption peaks of pro-tochlorophyll(ide) and chlorophyll(ide), respectively, are marked byvertical lines.
340 Plant Physiol. Vol. 45, 1970
DEVELOPMENT OF EUGLENA CHLOROPLASTS. I
PREI LLUMINATIONPERIOD
lDARKI PERIODPOTENTIATEDJ
NORMAL
12-
-J
8
-J
I
CLL
0%
-J
a
-J
0
~-i2
-10 0 10
I PtC)?IT I IUIU ATInL lD01rin. I 1--
30TIME (HRS.)
50 70
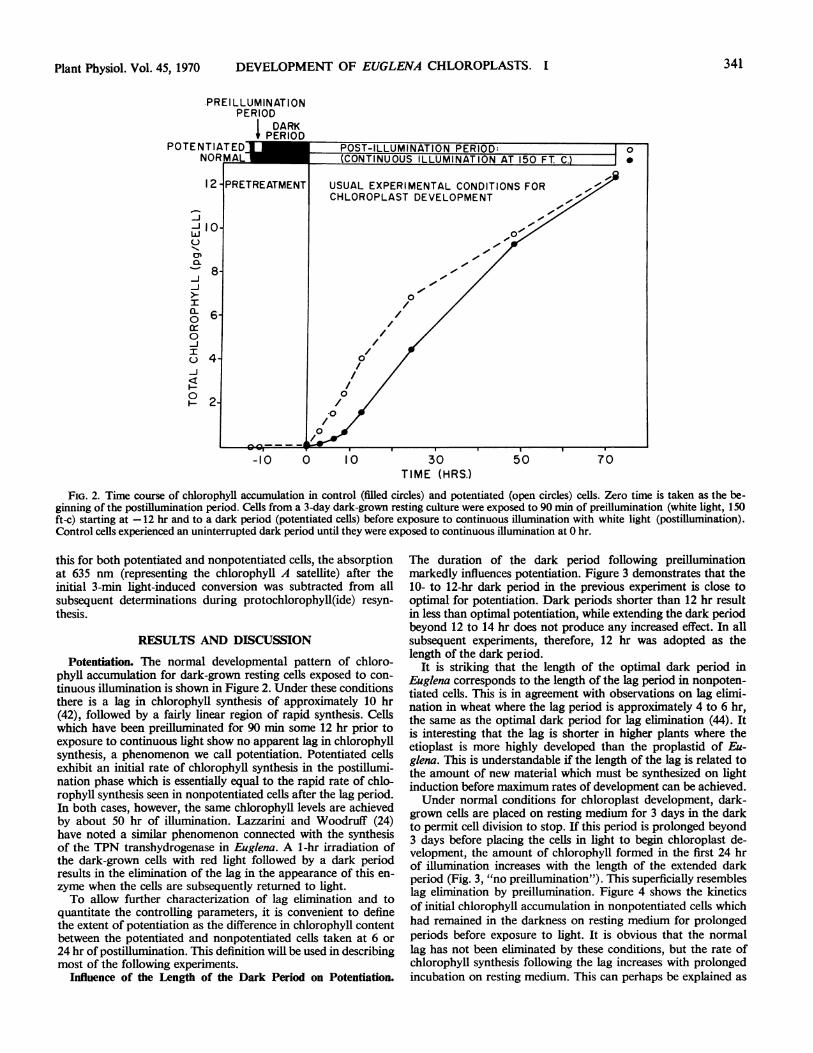
FIG. 2. Time course of chlorophyll accumulation in control (filled circles) and potentiated (open circles) cells. Zero time is taken as the be-ginning of the postillumination period. Cells from a 3-day dark-grown resting culture were exposed to 90 min of preillumination (white light, 150ft-c) starting at -12 hr and to a dark period (potentiated cells) before exposure to continuous illumination with white light (postillumination).Control cells experienced an uninterrupted dark period until they were exposed to continuous illumination at 0 hr.
this for both potentiated and nonpotentiated cells, the absorptionat 635 nm (representing the chlorophyll A satellite) after theinitial 3-min light-induced conversion was subtracted from allsubsequent determinations during protochlorophyll(ide) resyn-thesis.
RESULTS AND DISCUSSION
Potentiation. The normal developmental pattern of chloro-phyll accumulation for dark-grown resting cells exposed to con-
tinuous illumination is shown in Figure 2. Under these conditionsthere is a lag in chlorophyll synthesis of approximately 10 hr(42), followed by a fairly linear region of rapid synthesis. Cellswhich have been preilluminated for 90 min some 12 hr prior toexposure to continuous light show no apparent lag in chlorophyllsynthesis, a phenomenon we call potentiation. Potentiated cellsexhibit an initial rate of chlorophyll synthesis in the postillumi-nation phase which is essentially equal to the rapid rate of chlo-rophyll synthesis seen in nonpotentiated cells after the lag period.In both cases, however, the same chlorophyll levels are achievedby about 50 hr of illumination. Lazzarini and Woodruff (24)have noted a similar phenomenon connected with the synthesisof the TPN transhydrogenase in Euglena. A 1-hr irradiation ofthe dark-grown cells with red light followed by a dark periodresults in the elimination of the lag in the appearance of this en-zyme when the cells are subsequently returned to light.To allow further characterization of lag elimination and to
quantitate the controlling parameters, it is convenient to definethe extent of potentiation as the difference in chlorophyll contentbetween the potentiated and nonpotentiated cells taken at 6 or24 hr of postillumination. This definition will be used in describingmost of the following experiments.
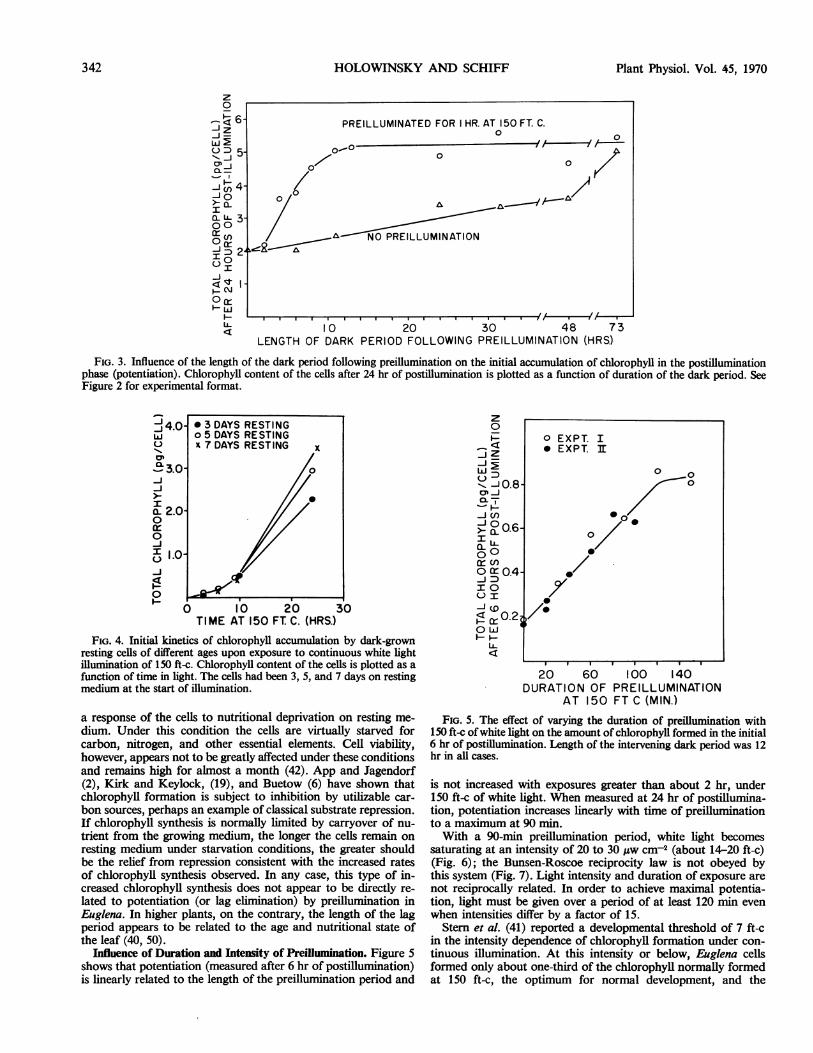
Influence of the Length of the Dark Period on Potentiation.
The duration of the dark period following preilluminationmarkedly influences potentiation. Figure 3 demonstrates that the10- to 12-hr dark period in the previous experiment is close tooptimal for potentiation. Dark periods shorter than 12 hr resultin less than optimal potentiation, while extending the dark periodbeyond 12 to 14 hr does not produce any increased effect. In allsubsequent experiments, therefore, 12 hr was adopted as thelength of the dark period.
It is striking that the length of the optimal dark period inEuglena corresponds to the length of the lag period in nonpoten-tiated cells. This is in agreement with observations on lag elimi-nation in wheat where the lag period is approximately 4 to 6 hr,the same as the optimal dark period for lag elimination (44). Itis interesting that the lag is shorter in higher plants where theetioplast is more highly developed than the proplastid of Eu-glena. This is understandable if the length of the lag is related tothe amount of new material which must be synthesized on lightinduction before maximum rates of development can be achieved.Under normal conditions for chloroplast development, dark-
grown cells are placed on resting medium for 3 days in the darkto permit cell division to stop. If this period is prolonged beyond3 days before placing the cells in light to begin chloroplast de-velopment, the amount of chlorophyll formed in the first 24 hrof illumination increases with the length of the extended darkperiod (Fig. 3, "no preillumination"). This superficially resembleslag elimination by preillumination. Figure 4 shows the kineticsof initial chlorophyll accumulation in nonpotentiated cells whichhad remained in the darkness on resting medium for prolongedperiods before exposure to light. It is obvious that the normallag has not been eliminated by these conditions, but the rate ofchlorophyll synthesis following the lag increases with prolongedincubation on resting medium. This can perhaps be explained as
L.- (CONTINUOUS ILLUMINATION AT 150 FT C.)
USUAL EXPERIMENTAL CONDITIONS FORCHLOROPLAST DEVELOPMENT
0//////
//
00
/////0/o
/_
= . I I I I I I
I
341Plant Physiol. Vol. 45, 1970
9-9 1-% -II II MI ffW LL I III IrU I 00
PRETREATMENT
HOLOWINSKY AND SCHIFF
PREILLUMINATED FOR HR. AT 150 FT. C.0o
00
6/ A T
/ ^0~~N PREILLUMINATION
10 20 30 48 73LENGTH OF DARK PERIOD FOLLOWING PREILLUMINATION (HRS.)
FIG. 3. Influence of the length of the dark period following preillumination on the initial accumulation of chlorophyll in the postilluminationphase (potentiation). Chlorophyll content of the cells after 24 hr of postillumination is plotted as a function of duration of the dark period. SeeFigure 2 for experimental format.
-i,4.0-wuCP
..1: 3.0-J-JI
X 2.0-0ck:0-J
-J0-0
I-
O 10 20 30
TIME AT 150 FT C. (HRS.)
FIG. 4. Initial kinetics of chlorophyll accumulation by dark-grownresting cells of different ages upon exposure to continuous white lightillumination of 150 ft-c. Chlorophyll content of the cells is plotted as afunction of time in light. The cells had been 3, 5, and 7 days on restingmedium at the start of illumination.
a response of the cells to nutritional deprivation on resting me-dium. Under this condition the cells are virtually starved forcarbon, nitrogen, and other essential elements. Cell viability,however, appears not to be greatly affected under these conditionsand remains high for almost a month (42). App and Jagendorf(2), Kirk and Keylock, (19), and Buetow (6) have shown thatchlorophyll formation is subject to inhibition by utilizable car-bon sources, perhaps an example of classical substrate repression.If chlorophyll synthesis is normally limited by carryover of nu-trient from the growing medium, the longer the cells remain onresting medium under starvation conditions, the greater shouldbe the relief from repression consistent with the increased ratesof chlorophyll synthesis observed. In any case, this type of in-creased chlorophyll synthesis does not appear to be directly re-lated to potentiation (or lag elimination) by preillumination inEuglena. In higher plants, on the contrary, the length of the lagperiod appears to be related to the age and nutritional state ofthe leaf (40, 50).
Influence of Duration and Intensity of Preillumination. Figure 5
shows that potentiation (measured after 6 hr of postillumination)is linearly related to the length of the preillumination period and
z0
_ ZJ E
.EI D" -0.8-c'JCI-_ ,_.
J U)
> ° 0.6-IQ- 000cr Ct0 r 0.4-I O)
m 0.2So ui
U- I
20 60DURATION OF
AT 150
100 140PRE ILLUMINATIONFT C (MIN.)
FIG. 5. The effect of varying the duration of preillumination with150 ft-c ofwhite light on the amount of chlorophyll formed in the initial6 hr of postillumination. Length of the intervening dark period was 12hr in all cases.
is not increased with exposures greater than about 2 hr, under150 ft-c of white light. When measured at 24 hr of postillumina-tion, potentiation increases linearly with time of preilluminationto a maximum at 90 min.With a 90-min preillumination period, white light becomes
saturating at an intensity of 20 to 30 ,uw cm-2 (about 14-20 ft-c)(Fig. 6); the Bunsen-Roscoe reciprocity law is not obeyed bythis system (Fig. 7). Light intensity and duration of exposure arenot reciprocally related. In order to achieve maximal potentia-tion, light must be given over a period of at least 120 min evenwhen intensities differ by a factor of 15.
Stern et al. (41) reported a developmental threshold of 7 ft-cin the intensity dependence of chlorophyll formation under con-tinuous illumination. At this intensity or below, Euglena cellsformed only about one-third of the chlorophyll normally formedat 150 ft-c, the optimum for normal development, and the
z0
_- 6-* Z
U D 5Co,-I-
I0-LI 3-O 0
() I
-J sX 2IOiox
I.I--
U-
* 3 DAYS RESTINGo 5 DAYS RESTINGx 7 DAYS RESTING x
/o
I -r
0 EXPT. I* EXPT. ir
0 o0
0
0
O/
342 Plant Physiol. Vol. 45, 1970

DEVELOPMENT OF EUGLENA CHLOROPLASTS. I
z e =
I-<JZ
llJ E
_L I
J(n 0.3-
a-ti-00cr~CI)1D.
-Jc-) 0
<(DI-
c
I .
< 60 120 180 240 509INTENSITY OF PREILLUMINATING LIGHT (ELWATTS/cm2)
FIG. 6. The effect of varying the intensity of white light given in 90min of preillumination on the amount of chlorophyll formed in theinitial 6 hr of postillumination. Duration of the intervening dark periodwas 12 hr.
z
0
<iZ1--
LLJCv D
P_ 0.6-,
I-
.10
I X 0.4-
00
J a: 0.2-uoO
OLs
A 300 FT. C. = 509 MW/cm20 20 FT. C.= 27MW/cm2
AAo/0
20 60 00o I1ODURATI ON OF PRE ILLUMINATION (MIN.)
FIG. 7. The effects of varying the duration of preillumination withwhite light of two widely different intensities on the amount of chloro-phyll formed in the initial 6 hr of postillumination. Duration of the in-tervening dark period was 12 hr.
plastids exhibited abnormalities suggesting inhibition of normallamella formation from discs. On subsequent exposure of suchcells to light of 150 ft-c, chlorophyll increased beyond the nor-
mal level, resulting in hyperdevelopment. The fact that the pre-illumination intensity curve in the present experiments saturatesat a value (14 ft-c) quite close to the threshold reported by Sternet al. (41) suggests that these phenomena may be closely related.Both preillumination at 14 ft-c and prolonged development at7 ft-c appear to induce the system to accumulate a developmentalpotential which is only realized when the cells are subsequentlyexposed to optimal light intensities for chloroplast development.The Nature of the Photoreceptor for Preillumination. The effec-
tive light quality for preillumination was measured as a broadband action spectrum with Wratten filters. For this purpose itwas necessary to pool data from several experiments performedat different times and on different cultures. Since there is some
degree of variability in the absolute level of responsiveness ofcells from different cultures to identical conditions of preillumi-nation, it was necessary to have an internal control for each
set of determinations. The control in each instance is exposed tonormal potentiating conditions, i.e., 90 min of preilluminationwith white light at 150 ft-c, followed by 12 hr of darkness beforere-exposure to continuous postilluminating light. The amount ofpotentiation after 6 hr of postillumination in this control wastaken as the 100% value for that experiment, and the responsesto all other preillumination light treatments within a given ex-periment were normalized to this value.
Figure 8 shows the relationship between light intensity andresponse in the blue, green, and red regions of the spectrum. Theresponse is essentially linear from zero to 10 ,uw cm-2 for allthree spectral regions. With increasing intensities, blue light ap-proaches 95% of the effectiveness of white light, while the maxi-mum obtained with red light is only about 60%. One possibleexplanation for this is that there are two pigment systems oper-ating, one of which absorbs only in the blue while the other ab-sorbs in both the blue and the red. If the effectiveness observedresults from synergism between the two systems, blue light atsuitable intensities should be as effective as white light, while redlight alone at saturation should be less effective as observed.Any complicating influence of far red light is excluded since thered fluorescent lamps employed emit little or no far red light.
In Table I we compare the relative effectiveness of the three
100
Z 80-0
a-z0 o
240z
o 20 o GREa-
0
INTENSITY OF PREILLUMI'NATING LIGHT (MW/cm2)FIG. 8. The effect of varying intensities of light of different spectral
qualities given in 90 min of preillumination on the amount of chloro-phyll formed in the initial 6 hr of postillumination with white light. Theintervening dark period was 12 hr. The degree of stimulation resultingfrom preillumination is expressed in terms of percentage potentiation.For a detailed explanation, refer to the text.
Table I. Comparison of Spectral Effectiveness for Preilluminationwith Absorption of Protochlorophyllide Holochrome
Energy Relative Effective-Required ness
Filter Transmission Range for 50%Potentia-
Dtr xiotenti mined pected'
Mw/cm'
Wratten 98 Blue, 40500 nm 5.5 6.3 6.0Wratten 61 Green, 480-600 nm 35.0 1.0 1.0Wratten 29 Red, 610-700 nm 8.0 4.4 3.0
1 The expected effectiveness was calculated as follows. For eachspectral range the integrated filter transmission was multiplied bythe integrated absorption of the protochlorophyllide holochromefrom beans (39). Red and blue were normalized to green taken as1 for both relative effectiveness and expected effectiveness.
8 A
0
0~~~~~~~~~~~oA
O EXPT. I/ EXPT. Jr
343Plant Physiol. Vol. 45, 1970

HOLOWINSKY AND SCHIFF
spectral regions in eliciting 50% potentiation (thereby excludingsaturation phenomena) with the effectiveness expected when thepreillumination photoreceptor has the same absorption envelopeas the protochlorophyllide holochrome from beans. As may beseen, there is excellent agreement between the expected and ex-perimentally determined values, and there is every reason to be-lieve that potentiation employs protochlorophyllide or a verysimilar pigment as the effective photoreceptor for preillumination.A secondary blue receptor may be involved, as discussed above,which makes its presence known as an additive phenomenon inthe saturation region.
z0
_jW E 0.6-
FL-jG
Jcn
o 0.4 _0.400Jr 0.2
_O0
o WL D R FR RFRFR,RI-i- PREILLUMINATING LIGHT
FIG. 9. The effect of preillumination with red and far red light on therate of chlorophyll formation in the initial 6 hr of postillumination.Duration of the intervening dark period was 12 hr. Preilluminationconditions were: WL: 90 min white light of 150 ft-c (213,uw cm<);D: no preillumination; R: 90 min red light at about 200gAw cm7; FR:90 min far red light at about 400 w cm2; R, FR: 90 min red followedby 90 min far red; FR, R: 90 min far red followed by 90 min red.
I0Z
0
ICkJ0
8
6
4
) 20 40 60 8oDURATION OF DARK INTERVALS
(MIN.)
It is unlikely that the red far red phytochrome system charac-teristic of higher plants is involved here. In Euglena, blue light ishighly effective, and preillumination must be given over a pro-longed period even at saturating intensities. Consistent with this,we have been unable to demonstrate a far red reversal of redlight-induced potentiation (Fig. 9). Preillumination of the cellswith red light for 90 min results at best in some 70% potentiationas compared to white light. Far red light alone is completely in-active. Curiously, when cells are exposed to far red immediatelyprior to or immediately after red light, the resulting potentiationis 100%. Clearly, far red does not reverse the effect of red lightunder our experimental conditions, and no red far red system ofthe usual type can be implicated in mediating the potentiationphenomenon in Euglena.
Potentiation is clearly a function of the quality and durationof light administered in the preillumination period. Since theaction spectrum indicates that the protochlorophyll(ide) to chlo-rophyll(ide) conversion is implicated, it is interesting to inquirewhether the degree of potentiation obtained is related to theamount of protochlorophyll(ide) converted during preillumina-tion. Figures lOa and lOb show that there is a direct correlationbetween the amount of chlorophyll accumulated during preillu-mination and the resulting degree of potentiation. In these ex-periments chlorophyll synthesis in the 90-min preilluminationphase was limited through the use of interrupted illuminationwith regularly spaced dark periods of various durations (closedcircles). Another means of observing this correlation is to usethe inherent variability in responsiveness of different culturespreilluminated with continuous light (open circles, Fig. lOb). Itis noteworthy that the points obtained from cultures preillumi-nated continuously fall on the same line described by the pointsobtained using different regimes of equally spaced alternatinglight-dark periods within the 90-min preillumination (filledcircles, Fig. lOb). The linear correlation is striking and suggeststhat the preillumination controls the events which are to follow,either by protochlorophyll(ide) removal or by chlorophyll(ide)
0 CONTINUOUS PREILLUMINATIONz * INTERRUPTED PREILLUMINATION0I-.4i0.60o-JO0~~~~~~~~_jJ
04 0C,)
0 Q40-0
0~~~0~~~
0.20
0.100 0.02 004 0.06 0.08TOTAL CHLOROPHYLL (pg/CELL)
AFTER 90 MINS. OF PREILLUMINATION
FIG. 10. Relationship between the amount of chlorophyll formed in the preillumination period and the extent of resulting potentiation as meas-ured by the amount of chlorophyll synthesized by the cells after 6 hr of postillumination. The amount of chlorophyll formed in the 90-min pre-illumination period was controlled by exposing the cells to a total of 10 min of white light at 300 ft-c, which was delivered as brief regularlyspaced "flashes" (ranging from 20 flashes each M min in duration to 2 exposures of 5 min each) with interposed dark intervals ranging from 4 to80 min. a: Total chlorophyll formed in the 90-min preillumination phase (open triangles) and after 6 hr of postillumination (filled triangles) as afunction of the duration of the regularly spaced dark intervals is shown. The values obtained for cells preilluminated with 90 min continuous whitelight were taken as 100%. b: A plot of the amount of chlorophyll formed in 6 hr of postillumination as a function of the amount of chlorophyllformed during the 90-min preillumination phase as a result of both continuous (open circles) and interrupted (filled circles) illumination.
A AT END OF PREILLUMINATIONA AFTER 6 HOURS OF POST-
ILLUMINATION
0° k)o
'10-I A
i0 %%N
%A~~~
O A^\"O-I_,
344 Plant Physiol. Vol. 45, 1970

DEVELOPMENT OF EUGLENA CHLOROPLASTS. I
40 60
TIME (MIN.)
FIG. 11. Kinetics of protochlorophyll(ide) resynthesis in controlcells (dark-grown cells after 3.5 days on resting medium in the dark)and in maximally potentiated cells (90-min preillumination with whitelight followed by 12 hr of darkness). Protochlorophyll(ide) resynthesisin darkness following initial total conversion by light is plotted as afunction of time. Protochlorophyll(ide) is expressed as the change inoptical density at 635 nm for 1010 cells. The experimental regime isdepicted graphically in the upper portion of the figure. For a detailedexplanation, refer to the "Materials and Methods" section of the text.
or chlorophyll accumulation, but it is not clear which is the con-trolling species. Protochlorophyll(ide), then, could be an inhibitorof some critical initial synthesis essential to plastid development,and its constant removal by illumination might be required fora sufficiently long period to allow threshold amounts of thecritical product(s) to accumulate (36). Alternatively, accumu-lating chlorophyll might act as an inducer.
Events of the Dark Period after Preillumination. Potentiation isa result of both preillumination and the events taking place inthe dark period. It is not clear at present what the critical eventsare or what components might be synthesized as a result ofpreillumination. It is unlikely, however, that the rate of proto-chlorophyll(ide) resynthesis is the limiting factor in determiningpotentiation in Euglena. As seen in Figure 11, protochloro-phyll(ide) resynthesis is kinetically similar in both potentiatedand nonpotentiated cells. Protochlorophyll(ide) resynthesis inetiolated wheat seedlings, however, is accelerated, and the lag iseliminated, as a result of preillumination (3). Akoyunoglou andSiegelman (1) reported a changing pattern of protochloro-phyll(ide) resynthesis with increasing age of etiolated bean leaves.The kinetic pattern characteristic of juvenile bean leaf tissue isvery similar to that observed by us for Euglena and by Augusti-nussen and Madsen (3) for preilluminated barley. The pattern inunilluminated etiolated wheat resembles that for aged beanleaves.
CONCLUSIONS
The classical experiments on lag elimination in chlorophyllsynthesis in higher plants (3, 32, 44, 45, 49), including the actionspectrum determinations of Virgin (46), established the follow-ing: (a) the optimal length of dark period following preillumi-nation is approximately 4 hr, which is about the length of thenormal lag period in these organisms; (b) the saturation durationfor preillumination is a few minutes at most; and (c) preillumi-nation by blue light is ineffective in lag elimination. Effectivenesswas concentrated in a single broad peak in the red-orange regionof the spectrum, suggesting the participation of the red-far red
system, although the reversal by far red light was ambiguous inthe case of wheat (46). This action spectrum measured by Virgin(45, 46) and the far red light reversal observed in other plants(29, 32) exclude protochlorophyll(ide) as the unique mediator inlag elimination since blue light would be expected to be highlyeffective if protochlorophyll(ide) were the photoreceptor.
Euglena has several similarities to and several differences fromthe higher plant system:
1. The optimal length of the dark period is about 12 hr, whichcorresponds to the length of the normal lag period in develop-ment in continuous light. It is tempting to explain the longer lag(and, therefore, the longer optimal dark period for potentiation)by correlating this with the extent of development of the pro-plastid in the dark. The higher plant etioplast is about one-thirdthe size it will achieve as the mature chloroplast and has appre-ciable levels of many plastid constituents (5, 10). It must increasein volume about 10-fold during chloroplast development. TheEuglena proplastid, on the other hand, must increase about 40-fold in becoming a chloroplast. It is surely no coincidence thatthe volume changes during development as well as the length ofthe respective dark periods for potentiation and the length of thelag in continuous light (4 hr for higher plants and 12 hr forEuglena) all show a ratio of between 3 and 4 to 1. A reasonableexplanation is that all of these parameters are an external reflec-tion of the starting point of the respective proplastids and theamount of material and organization which must be achieved tocomplete development. If light serves as an inducer for darksynthesis to follow, the amounts of dark synthesis required shouldbe greater in Euglena than in higher plants.
2. The preillumination period in Euglena must be at least 90min at saturating intensities to achieve full potentiation, in con-trast to the very biief preillumination (2 to 5 min) effective inhigher plants (32). This suggests that the two phenomena arevery different in mechanism. In Euglena the final effect appearsto be governed (during the preillumination period) by limitingdark reactions and the requirement that enough time elapse topermit sufficient products to accumulate. If dark reactions doindeed limit the over-all rate and thereby determine the optimalperiod for preillumination, the optimal length of preilluminationshould be related to temperature via the Arrhenius relationship,an experiment which remains to be done.
3. Preillumination in Euglena is most effective when mediatedby the blue and red regions of the spectrum, with the blue andred effectiveness ratio consistent with the expected integratedabsorptions of protochlorophyll(ide). Furthermore, no evidencefor phytochrome involvement was obtained. Thus the two sys-tems differ markedly in the photosensitizing pigment utilized inthe preillumination period. In higher plant tissues a red-far redsystem is implicated on the basis of (a) a single peak of effective-ness in the red and (b) depending on the organism, a small tofairly complete reversal by far red light. Euglena appears toeschew the red-far red morphogenetic control system entirelyand entrusts developmental induction of the chloroplast to thepenultimate step in chlorophyll biosynthesis. The action spec-trum also excludes the exclusive participation of a high energyblue response, which would be expected to lack a red effectivenesspeak and to possess one in the far red (30). A synergistic blue-absorbing pigment may be invoked to explain the saturationphenomena in Figure 8. Henningsen (15) reported that vesicledispersal in etioplasts is elicited by blue light with red and greenshowing no effectiveness. This suggests a photoreceptor with ablue absorption peak only, quite unlike protochlorophyll orphytochrome (48). Pigments other than protochlorophyll(ide)have been postulated to act in Euglena plastid development andin respiration control in other organisms (23, 52).The findings presented in this paper have some evolutionary
implications. The primitive condition which apparently persists
Plant Physiol. Vol. 45, 1970 345

HOLOWINSKY AND SCHIFF
in most green algal groups renders the organisms capable ofchlorophyll and chloroplast formation in the dark. Since one canobtain what appear to be single step mutants of green algaewhich require light for chlorophyll and chloroplast formation(33, 34), these organisms probably contain two systems forachieving the protochlorophyll(ide) to chlorophyll(ide) trans-formation and the concomitant induction of chloroplast develop-ment: a system which requires light and one which does not.The mutants have probably lost the dark system, rendering themlight-dependent for these processes. Euglena seems to be muchlike these mutants of the green algae in that it requires light topromote the formation of chlorophyll and chloroplasts althoughfurther adaptation has provided it with a true proplastid which theorganism maintains under conditions of darkness. In the line ofevolution leading to higher plants, the phytochrome system seemsto have become implicated as a limiting control step quite early inthe evolutionary sequence (14, 43) and exhibits its most elaborateform in the angiosperms. The ability to convert proto-chlorophyll(ide) to chlorophyll(ide) in the dark has apparentlybeen lost more slowly but is completely absent in the angio-sperms, which are dependent on light for chlorophyll and chloro-plast development. In these organisms, however, the concomitantelaboration of the phytochrome system has apparently resulted inthis system's becoming an additional and often overriding controlpoint in the induction of chloroplast formation (3, 28, 29, 32, 45,46, 49), with the light-induced protochlorophyll(ide) to chloro-phyll(ide) conversion assuming a secondary position. In Euglena,of course, the phytochrome system appears not to have beenevolved, and the induction of chlorophyll and chloroplast forma-tion is largely dependent on the light-induced conversion of proto-chlorophyll(ide) to chlorophyll(ide).
Turning to the potentiation phenomenon in its broad develop-mental sense, it seems likely that a brief light induction is sufficientto initiate syntheses which would ordinarily be carried out duringthe first 12 hr of continuous light. Furthermore, the synthesis ofthese components appears to be required to sustain subsequenthigh rates of chloroplast development in the postilluminationperiod. Attention focuses, then, on the nature of the induction bylight during the preillumination period and on the post inductionevents occupying the dark period. A model for induction ofchloroplast development has been presented previously (36, 51)where protochlorophyll(ide), aside from controlling its own bio-synthesis through feedback inhibition, also mediates unblockingof protein synthesis at some level in transcription and translation,in both the cytoplasm and the developing plastid. Now that it ispossible to separate the inductive step (as preillumination) fromthe subsequent early developmental events (in the subsequent12-hr dark period), both phenomena become amenable to studywith tracers and inhibitors of RNA and protein synthesis. Thuspotentiation should be a valuable tool in obtaining the datanecessary to define the level of control and its cellular localization.
Ackntowledgment-The technical assistance of Mrs. Lorna West in the initial experi-ments is greatly appreciated.
LITERATURE CITED
1. AKOYUNOGLOU, G. A. AND H. W. SIEGELMAN. 1968. Protochlorophyllide re-
synthesis in dark-grown bean leaves. Plant Physiol. 43: 66-68.2. ApP, A. A. AND A. T. JAGENDORF. 1963. Repression of chloroplast develop-
ment in Euglena gracilis by substrates. J. Protozool. 10: 340-343.3. AUGUSTINUSSEN, E. AND A. MADSEN. 1965. Regeneration of protochlorophyll in
etiolated barley seedlings following different light treatments. Physiol. Plant.18: 828-837.
4. BEN-SHAUL, Y., J. A. SCHIFF, AND H. T. EPSTEIN. 1964. Studies of chloroplastdevelopment in Euglena. VII. Fine structure of the developing plastid. PlantPhysiol. 39: 231-240.
5. BOGORAD, L. 1967. Chloroplast structure and development. In: A. Sani Pietroand F. A. Greer, eds., Harvesting the Sun. Academic Press, New York. pp.191-210.
6. BUETOW, D. E. 1967. Acetate repression of chlorophyll synthesis in Euglenagracilis. Nature 213, 1127-1128.
7. BUTLER, W. L. AND W. R. BRIGGS. 1966. The relation between structure and pig-ments during the first stages of proplastid greening. Biochim. Biophys. Acta112: 45-53.
8. BUTLER, W. L. AND K. H. NORRIS. 1960. Spectrophotometry of dense lightscattering materials. Arch. Biochem. Biophys. 87: 31-40.
9. BUTLER, W. L. AND K. H. NORRIS. 1962. Plant spectra: Absorption and action.In: H. F. Linskens and M. V. Tracy, eds., Modern Methods of Plant Analysis,Vol. V. Springer-Verlag, Berlin. pp. 51-72.
10. CHEN, S., D. MCMAHON, AND L. BoGORAD. 1967. Early effects of illumination onthe activity of some photosynthetic enzymes. Plant Physiol. 42: 1-5.
11. DOWNS, R. J. 1955. Photoreversibility of leaf and hypocotyl elongation of dark-grown red kidney bean seedlings. Plant Physiol. 30: 468-473.
12. FILNER, B. AND A. 0. KLEIN. 1968. Changes in enzymatic activities in etiolatedbean seedling leaves after a brief illumination. Plant Physiol. 43: 1587-1595.
13. GREENBLATT, C. L. AND J. A. SCHIFF. 1959. A pheophytin-like pigment in dark-adapted Euglena gracilis. J. Protozool. 6: 23-28.
14. HAuPT, W. 1959. Die Chloroplastendrehung bei Mougeotia. 1. Uber den quan-titativen und qualitativen Lichtbedarf der Schwachlichtbewegung. Planta 53:484-501.
15. HENNINGSEN, K. W. 1966. An action spectrum for vesicle dispersal in bean plas-tids. In: T. W. Goodwin, ed., Biochemistry of Chloroplasts, Vol. II. AcademicPress, New York. pp. 453-457.
16. HOLOWINSKY, A. W., P. B. MOORE, AND J. G. TORREY. 1965. Regulatory as-pects of chloroplast growth in leaves of Xanithiunz pennsylvanzicum and etiolatedred kidney bean seedling leaves. Protoplasma 60: 94-110.
17. HOLOWINSKY, A. W. AND J. A. SCHIFF. 1968. Potentiation of chloroplast de-velopment in Euglenta by preillumination. Plant Physiol. 43: S-7.
18. KEiSTER, D. L., A. T. JAGENDORF, AND A. SAN PIETRO. 1962. Development ofbean leaf transhydrogenase in etiolated leaves. Biochim. Biophys. Acta 62:332-357.
19. KIRK, J. T. 0. AND M. J. KEYLOCK. 1967. Control of chloroplast formationin Euglena gracilis: dependence of rate of chlorophyll synthesis on previous nu-tritional history of cells. Biochem. Biophys. Res. Commun. 28: 927-931.
20. KLEIN, E. AND S. KLEIN. 1962. The effect of light intensity and sucrose feedingon the fine structure in chloroplasts and on the chlorophyll content of etiolatedleaves. J. Cell Biol. 14: 169-182.
21. KLEIN, S., G. BRYAN, AND L. BOGORAD. 1964. Early stages in the developmentof plastid fine structure in red and far-red light. J. Cell Biol. 22: 433-442.
22. KODAK WRATT`EN FILTERS, Ed. 20, 1960, Kodak Publication No. B3, EastmanKodak Company.
23. KOWALLIK, W. 1967. Action spectrum for an enhancement of endogenousrespiration by light in Chlorella. Plant Physiol. 42: 672-676.
24. LAZZARINI, R. A. AND M. WOODRUFF. 1964. The photoinduction of transhy-drogenase in Euglenta. Biochim. Biophys. Acta 79: 412-415.
25. LYMAN, H., H. T. EPSTEIN, AND J. A. SCHIFF. 1961. Studies of chloroplast de-velopment in Euglena. 1. Inactivation of green colony formation by UV light.Biochim. Biophys. Acta 50: 301-309.
26. MARCUS, A. 1960. Photocontrol of formation of red kidney bean leaf triphos-phopyridine nucleotide linked triose-phosphate dehydrogenase. Plant Physiol.35: 126-128.
27. MARGULIES, M. M. 1965. Relationship between red light mediated glyceralde-hyde-3-phosphate dehydrogenase formation and light dependent development ofphotosynthesis. Plant Physiol. 40: 57-60.
28. MEGO, J. L. AND A. T. JAGENDORF. 1961. Effect of light on growth and BlackValentine bean plastids. Biochim. Biophys. Acta 53: 237-254.
29. MITRAKOS, K. 1961. The participation of the red far-red reaction system inchlorophyll-metabolism. Physiol. Plant. 14: 497-503.
30. MOHR, H. 1962. Primary effects of light on growth. Ann. Rev. Plant Physiol.13: 465-488.
31. NISHIMURA, M. AND H. HUZISIGE. 1959. Studies on the chlorophyll formationin Euglena gracilis with special reference to the action spectrum of the process.J. Biochem. 46: 225-234.
32. PRICE, L. AND W. H. KLEIN. 1961. Fed, far-red response and chlorophyll syn-thesis. Plant Physiol. 36: 733-755.
33. SAGER, R. 1955. Inheritance in the green alga Chlamydonozo,as reilnhardi.Genetics 40: 476-489.
34. SAGER, R. 1958. The architecture of the chloroplast in relation to its photo-synthetic activities. Brookhaven Symp. Biol. 11: 101-116.
35. SCHIFF, J. A. 1963. Oxygen exchange by Euglenza cells undergoing chloroplastdevelopment. Carnegie Inst. Wash. Year B. 62: 375-378.
36. SCHIFF, J. A. AND H. T. EPSTEIN. 1965. The continuity of the chloroplast inEuglena. In: Michael Locke, ed., Reproduction: Molecular, Subcellular, andCellular. Academic Press, New York. pp. 131-189.
37. SCHIFF, J. A., H. LYMAN, AND H. T. EPSTEIN. 1961. Studies of chloroplastdevelopment in Euglena. II. Photoreversal of the UV inhibition of green colonyformation. Biochim. Biophys. Acta 50: 310-318.
38. SCHIFF, J. A., H. LYMAN, AND H. T. EPSTEIN. 1961. Studies of the chloroplastdevelopment in Euglena. III. Experimental separation of chloroplast developmentand chloroplast replication. Biochim. Biophys. Acta 51: 340-346.
39. SCHOPFER, P. AND H. W. SIEGELMAN. 1968. Purification of protochlorophyllideholochrome. Plant Physiol. 43: 990-996.
40. SISLER, E. C. AND W. H. KLEIN. 1963. The effect of age and various chemicalson the lag phase of chlorophyll synthesis in dark-grown bean seedlings. Physiol.Plant. 16: 315-322.
346 Plant Physiol. Vol. 45, 1970

Plant Physiol. Vol. 45, 1970 DEVELOPMENT OF EUGLENA CHLOROPLASTS. I
41. STERN, A. I., H. T. EPSTEIN, AND J. A. SCHIFF. 1964. Studies of chloroplast de-velopment in Euglena. VI. Light intensity as a controlling factor in development.Plant Physiol. 39: 226-240.
42. STERN, A. I., J. A. SCHIFF, AND H. T. EPSTEIN. 1964. Studies of chloroplastdevelopment in Euglena. V. Pigment biosynthesis, photosynthetic 02 evolutionand CO2 fixation during chloroplast development. Plant Physiol. 39: 220-226.
43. TAYLOR, A. 0. AND B. A. BONNER. 1967. Isolation of phytochrome from thealga Mesotaenium and liverwort Sphaerocarpos. Plant Physiol. 42: 762-766.
44. VIRGIN, H. I. 1957. Chlorophyll a content and transpiration of etiolated wheatleaves after pretreatment with a short light impulse followed by dark periods ofvarying lengths. Physiol. Plant. 10: 445-453.
45. VIRGIN, H. I. 1958. Studies on the formation of protochlorophyll and chloro-phyll a under varying light treatments. Physiol. Plant. 11: 347-362.
46. VIRGIN, H. I. 1961. Action spectrum for the elimination of the lag phase inchlorophyll formation in previously dark-grown leaves of wheat. Physiol. Plant.14: 439-452.
47. VIRGIN, H. I., A. KAHN, AND D. VON WErSTEIN. 1963. The physiology ofchlorophyll formation in relation to structural changes in chloroplasts. Photo-chem. Photobiol. 2: 83-91.
48. VON WETI5TEIN, D. 1966. On the physiology of chloroplast structures. In: T. W.Goodwin, ed., Biochemistry of Chloroplasts, Vol. I. Academic Press, New York.pp. 19-21.
49. WITHROW, R. B., J. B. WOLFF, AND L. PRICE. 1956. Elimination of the lagphase of chlorophyll synthesis in dark-grown bean leaves by a pretreatment withlow irradiances of monochromatic energies. Plant Physiol. 31: xiii.
50. WOLFF, J. B. AND L. PRICE. 1960. The effect of sugars on chlorophyll biosynthe-sis in higher plants. J. Biol. Chem. 235: 1603-1608.
51. ZELDIN, M. H. AND J. A. SCHIFF. 1967. RNA metabolism during light-inducedchloroplast development in Euglena. Plant Physiol. 42: 922-932.
52. ZELDIN, M. H. AND J. A. SCHIFF. 1968. A comparison of light-dependent RNAmetabolism in wild-type Euglena with that of mutants impaired for chloroplastdevelopment. Planta 81: 1-15.
347





























