Embed Size (px)
Citation preview

144
Journal of Scientific and Innovative Research 2016; 5(4): 144-148
Available online at: www.jsirjournal.com
Research Article
ISSN 2320-4818
JSIR 2016; 5(4): 144-148
© 2016, All rights reserved
Received: 21-06-2016
Accepted: 20-07-2016
Tin May Sev
Biotechnology Research
Department, Ministry of Education, Kyaukse, Myanmar
Aye Aye Khai
Professor, Biotechnology Research
Department, Ministry of Education,
Kyaukse, Myanmar
Ayme Aung
Biotechnology Research Department, Ministry of Education,
Kyaukse, Myanmar
San San Yu
Associate Professor, Biotechnology
Research Department, Ministry of Education, Kyaukse, Myanmar
Correspondence: Dr. San San Yu
Associate Professor, Biotechnology
Research Department, Ministry of
Education, Kyaukse, Myanmarr
Evaluation of endophytic bacteria from some rice
varieties for plant growth promoting activities
Tin May Sev, Aye Aye Khai, Ayme Aung, San San Yu*
Abstract
Eighteen endophytic bacteria were isolated from different rice varieties by using Jensen’s nitrogen free
medium. According to morphological and biochemical tests, isolates S1, R1, R2, R5 and R6 may be Rhizobium
spp. and the remaining isolates may be Azospirillum spp. Fourteen out of isolates had plant growth hormone
(Indole Acetic Acid) producing activity. R7 was the best Indole Acetic Acid producer among the isolated
strains at two days incubation period in Jensen’s nitrogen free medium with L-tryptophan 0.5mg /ml at 37˚C
and the concentration produced was 120.55 ppm. In nitrogen free semi-solid agar medium containing trace
amount of bromothymol blue as indicator, all isolates showed nitrogen fixing activity during seven days
incubation period. All isolates except S6 could solubilize phosphate on Pikovskaia’s medium supplemented
with (0.5% Ca3(PO4)2). S5 was the best P- solubilizer at 5 days incubation period both in plate screening
showing zone diameter 45mm and 539.78 ppm quantitatively. Fourteen isolates except (LS, R2, S4, S8) could
also decompose potassium (ranging from 2-30mm in diameter) on K- decomposing medium with 0.12% K-
mica. R3 was the best K-decomposer by plate screening (30mm in diameter) and 19.84 ppm quantitatively. S3
and R3 had good antagonistic activities against Rhizotonia solani and LS and R5 to Phythium spp. and S7 to
Fusarium oxysporum respectively.
Keywords: Endophytic bacteria, Jensen’s nitrogen free medium, Indole Acetic Acid producer, Nitrogen
fixing activity, P-solubilizer, K-decomposer.
INTRODUCTION
Plants are constantly involved in interactions with a wide range of bacteria. These plant associated
bacteria colonize the rhizosphere (rhizobacteria), the phyllosphere (epiphytes) and the inside of plant
tissues (endophytes). Endophytes are sheltered from environmental stresses and microbial competition
by the host plant and they seem to be ubiquitous in plant tissues, having been isolated from flowers,
fruits, leaves, stems, roots and seeds of various plant species [1].
Some endophytic bacteria exert several beneficial effects on host plants, such as stimulation of plant
growth by production of plant hormones [2], nitrogen fixation [3] and induction of resistance to plant
pathogens [4-6], enhanced nutrient uptake, enhanced grain quality and increased drought tolerance. Some
endophytic microorganisms are able to produce compounds of biotechnological value as antibiotics and
antitumor drugs. It is well established that plant bacterial endophytes are to be found in most healthy
plant tissues [6-8]. This particular host endophyte interaction has been variously defined as altruism,
commensalisms, mutualistic, symbiosis or passivity to pathogenicity. Whatever the specific
relationship(s) involved, internal plant colonization by bacteria constitutes a vast and, as yet, little
mapped ecological niche.
There is much debate as to how to define an endophyte. For example, [6] suggested that bacteria and
fungi that are isolated from surface sterilized plant tissues, and that do no apparent harm to the plant,
could be considered as endophytes. Endophytic microorganisms have been defined as those that reside at
some phase of their life cycle within living plant tissues [9, 10]. Endophytes have been found in all plants
that have been examined [11]. These microorganisms include both commensal species, which have no
direct effect on the host plant, and mutualistic symbionts, which could be used in the biological control
of pathogens or plant growth promotion. The intimate relationship between endophytic bacteria and their
host involves co-evolutionary processes, and may influence the physiological mechanisms of plants [12].

145
It has also been shown that in some cases endophytes can accelerate
seed emergence and promote plant establishment under adverse
conditions [13] as well as increase plant growth and hasten plant
development [14]. On the other hand, the effects of endophytes may also
be deleterious, possibly contributing to disease status [15,16], or some
endophytes may interact with other endophytic populations [15]. Causes
of these contrasting outcomes are mostly unclear, but they are very
likely affected by the complex dynamics of interactions among
endophytes, which are in turn affected by environmental conditions,
plant species, and soil type.
MATERIALS AND METHODS
Sampling
The healthy and disease symptomless rice plants were collected from
different rice viz. (Shwe thwe yin, Ma jan taw, Ma naw thukha, Paw san
baykyar and Sin thwe latt) from different places (around Mandalay
Technological University and Kyaukse, Myanmar).
Isolation of Endophytic Bacteria
Endophytic bacteria used in this study were isolated from root and stem
tissues of healthy and disease symptomless rice plants which were
collected from various places. They were washed with tap water to
remove soil and other impurities and then roots and stems were cut into
2-3 cm pieces with sterile surgical blade.
These pieces were dipped and shaked in 96% ethanol for 10 seconds
and then they were rinsed in sterile distilled water for three times.
After that they were dipped and shaked for 10 seconds in 0.1 %
mercuric chloride and rinsed with sterile distilled water for three times.
Then, these sterilized pieces were crushed with sterile motor and pestle.
Finally, the Jensen’s Nitrogen free agar medium was prepared and it
was poured into the sterilized petridish and divided into three parts; one
part for control without inoculation, one for rolling of sterilized tissue
pieces for checking efficient sterilization and one for inoculation of
crushed sample. These were incubated at 37˚C for three to five days.
Screening for Indole Acetic Acid (IAA) Producing Activity
Indole acetic acid (IAA) productions of the isolated strains were
determined by colorimetrically using Salkowski’s reagent (50ml of 35%
perchloric acid: 1ml of 0.5 N FeCl3). To measure IAA qualitatively,
isolated strains were aseptically cultured in Jensen’s nitrogen free broth
containing 0.5 mg L-tryptophan per ml and then incubated at 37ºC for 7
days.
After harvesting, these were centrifuged at 10,000 rpm for 20 mins.
One millilitre of supernatant was mixed with two ml of Salkowski’s
reagent and one drop of orthophosphoric acid and incubated at room
temperature for 25-30 min. Development of pink colour showed IAA
production.
Quantitative Analysis on IAA Production by UV-Vis
Spectrophotometer
Isolated endophytic bacteria were inoculated in Jensen’s nitrogen free
broth containing 0.5 mg L-tryptophan/ml and then incubated at 37°C for
1 to 10 days. After harvesting, these were centrifuged at 10,000 rpm for
20 mins. One ml of supernatant was mixed with 2 ml of Salkowski’s
reagent and one drop of orthophosphoric acid and incubated at room
temperature for 25-30 min.
The absorbance of developed pink colour was measured at 530nm and
then IAA concentrations are calculated by using IAA standard curve
(generated by plotting absorbance on Y-axis and authentic IAA
concentration (ppm) on X-axis).
Screening of Nitrogen-Fixing Activity
All isolates were inoculated in nitrogen free semi-solid agar medium
containing trace amount of BTB as indicator.
Determination of Phosphate Solubilizing Activity by Plate screening
method
A total of 18 bacterial isolates were tested on solid media to select the
most efficient strain. The Pikovskaia's medium was used as a basal
medium and tricalcium phosphate was used as substrate in this study.
After autoclaving, the medium was adjusted to pH 8.6. All isolated
endophytic bacteria were inoculated on this medium and incubated at
37°C for 5 days. After that, the solubilizing activity was measured by
diameter of halo zone (millimetre) around the colony.
Quantitative Determination of Phosphate Solubilizing Activity by
Spectrophotometric Method
The best five phosphorus solubilizing strains out of eighteen isolates
(SR, S5, R6, S7, R8) determined by plate screening method were further
evaluated quantitatively for their P- solubilizing ability in Pikovskaia’s
liquid medium. After autoclaving the medium, it was left to be cool and
0.5 % tricalcium phosphate was added as substrate to avoid its
denaturation by heat. These five strains were inoculated in their
respective broth and incubated at 37˚C and 150 rpm. Uninoculated
medium was used as the control.
Ten-milliliter samples were withdrawn every 24 hours for 1 week and
then each sample was passed through the cation exchange resin. Sodium
molybdate solution and hydrazine sulphate solution were used to form
blue color complex and P- solubilizing activities of these samples were
measured at 830nm by UV-Vis spectrophotometric method. The
intensity of the blue color is propotional to the amount of phosphate
initially corporated in the heteropoly acid.
Determination of Potassium Decomposing Activity by Plate
screening method
The potassium decomposing activity of the isolates was also determined
on K decomposing medium with 0.12% insoluble potassium mica. The
mica sample was washed to remove soluble potassium (K) and dried at
40oC in microwave oven.
All isolated strains were cultured on K decomposing medium
supplemented with insoluble potassium mica and incubated at 37˚C for
5 days. The decomposing activity was measured by diameter of clear
zone around the colony after this incubation period.
Quantitative Determination of K- decomposing Activity
The mica sample was supplemented after cooling the sterilized K-
decomposing broth, and inoculated the best two potassium decomposing
bacteria, S3 and R3, and incubated on shaker at 37˚C and 150 rpm. The
samples were withdrawn every day for one week and the insoluble mica
was removed by centrifugation at 6000 rpm for 10 minutes and then the
supernatants were analyzed by AAS (Atomic Absorption Spectrometry)
method.

146
Study on the Antagonistic Activity of Isolated Endophytes
Isolated strains were screened for their antagonistic activities against
common soil borne plant pathogenic fungi like Fusarium oxysporum,
Pythium spp. and Rhizoctonia soloni. This was carried out by hyphal
growth inhibition assay. Czapeak’s medium was used for screening of
antifungal activity. Inhibition of hyphal growth was observed.
Tests for Characterization of Isolated Endophytic Bacteria
These strains were identified on the basis of their colonial morphology,
microscopic characteristics and some biochemical tests etc.
RESULTS AND DISCUSSIONS
Isolation and Identification of Rice Endophytic Bacteria
Eighteen endophytic bacteria were isolated from different rice viz.
(Shwe thwe yin, Ma jan taw, Ma naw thukha, Paw san bay kyar and Sin
thwe latt) by using Jensen’s nitrogen free medium.
Colonial morphologies of all isolates were round raised, mucoid and
transparent on Jensen’s nitrogen free medium. All isolates were gram-
negative and cell diameters are in the range of 0.5-1.5 x 2-4 µm rods.
According to morphological and biochemical tests, isolates S1, R1, R2,
R5 and R6 may be Rhizobium spp. and the remaining isolates may be
Azospirillum spp.
Although Rhizobium association had been extensively explored in the
root nodules of legumes where they fix atmospheric nitrogen, recent
studies also suggest that Rhizobium can exhibit plant growth promoting
(PGP) activities with non-legumes such as rice [17].
Azospirillum spp have broad host range and these two species can
promote plant growth via secretion of plant growth hormones (IAA,
GA), vitamins, solubilization of insoluble phosphates, decomposition of
potassium, biological nitrogen fixation, induced resistance to some plant
pathogens via secretion of antimicrobial compounds that may hinder
colonization of hosts by phytopathogens, thereby suppressing the
diseases they cause, enhancement in stress resistance to abiotic factors [18] such as salt tolerance, drought resistance and production of
siderophores under iron limiting condition.
Figure 1: Growth of Endophytic Bacteria on Jensen’s medium (Azospirillum
spp.) and on YEM Medium (Rhizobium spp.)
Figure 2: Microscopic Morphology of Some Isolated Endophytes
Table 1: Biochemical Characteristics of Isolated Endophytic Bacteria
R3, R7, R8, S2, S3, S4, S5,
S6, S7, S8, LS, SS, SR
S1, R1, R2, R5, R6
Gram reaction - -
Colonial morphology
Small, white, dense
colonies on semi-
solid NFb agar
medium
Circular, convex,
mucous, opaque
or transparent on
YEM
Microscopic
morphology
Clump shape, slightly
curved and straight
rod, single and pairs
(0.6–1.7 x 2.1–
3.8µm)
Small rod, x, y
shape
(0.5–1 x 1.2–3
µm)
Motility + +
Gelatin agar - -
Starch hydrolysis - -
Catalase + +
Citrate utilization + -
Carbon sources utilization
Mannitol + +
Glucose + +
Sucrose + +
Screening on Indole Acetic Acid (IAA) Producing Activity
Four isolates, R2, SS, SR, LS couldn’t produce IAA by screening them
with IAA detecting reagent (Salkowski’s reagent) after 7 days
incubation.
Thus, fourteen out of isolates had plant growth hormone, IAA,
producing activity determined by the development of pink colour.
Figure 3: Screening on IAA Producing Activities of Fourteen Isolates
Quantitative Determination of IAA Producing Activity of Isolated
Endophytic Bacteria
Among fourteen isolates, R7 was the best IAA producer and the
concentration produced was 120.55 ppm in two days incubation period.
S6 produced 104.58 ppm and R8 produced 95.55 ppm. R6 had the lowest
C R1 S4 R5 S6 R7 S8 S5 R8 S3 R3 S7 S2 S1 R6C R1 S4 R5 S6 R7 S8 S5 R8 S3 R3 S7 S2 S1 R6
Biochemical
characteristics
Isolated
Bacteria
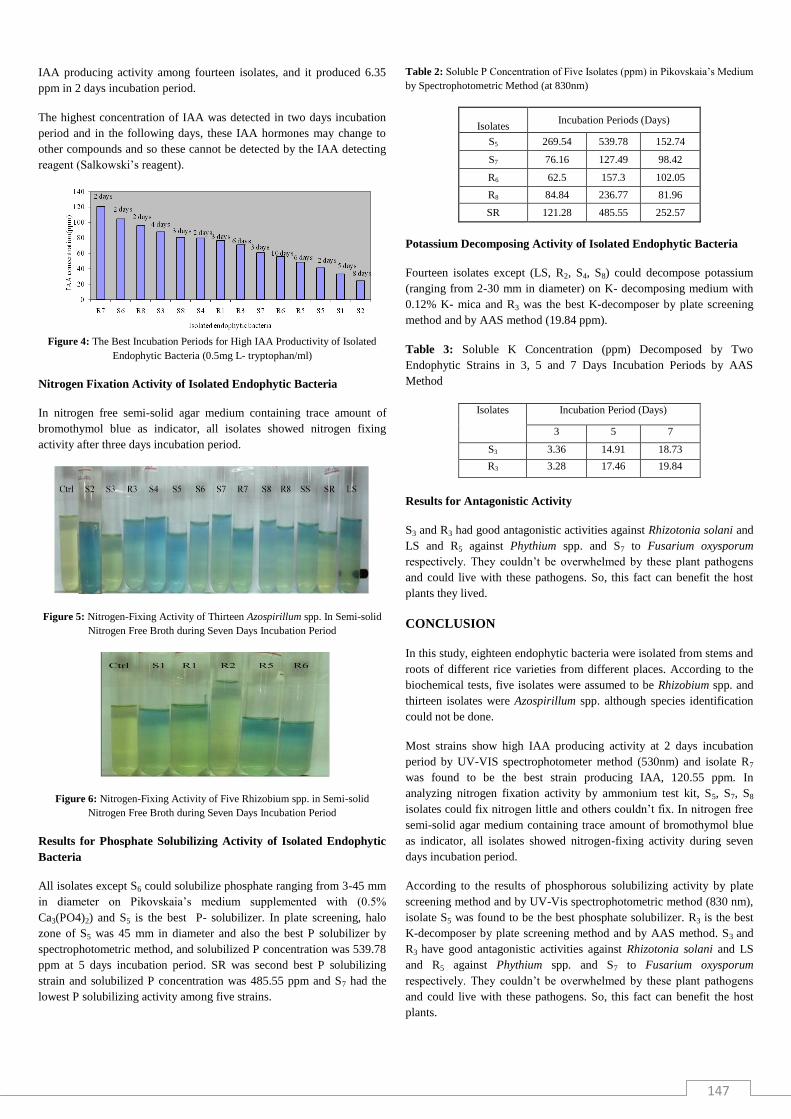
147
IAA producing activity among fourteen isolates, and it produced 6.35
ppm in 2 days incubation period.
The highest concentration of IAA was detected in two days incubation
period and in the following days, these IAA hormones may change to
other compounds and so these cannot be detected by the IAA detecting
reagent (Salkowski’s reagent).
Figure 4: The Best Incubation Periods for High IAA Productivity of Isolated
Endophytic Bacteria (0.5mg L- tryptophan/ml)
Nitrogen Fixation Activity of Isolated Endophytic Bacteria
In nitrogen free semi-solid agar medium containing trace amount of
bromothymol blue as indicator, all isolates showed nitrogen fixing
activity after three days incubation period.
Figure 5: Nitrogen-Fixing Activity of Thirteen Azospirillum spp. In Semi-solid
Nitrogen Free Broth during Seven Days Incubation Period
Figure 6: Nitrogen-Fixing Activity of Five Rhizobium spp. in Semi-solid
Nitrogen Free Broth during Seven Days Incubation Period
Results for Phosphate Solubilizing Activity of Isolated Endophytic
Bacteria
All isolates except S6 could solubilize phosphate ranging from 3-45 mm
in diameter on Pikovskaia’s medium supplemented with (0.5%
Ca3(PO4)2) and S5 is the best P- solubilizer. In plate screening, halo
zone of S5 was 45 mm in diameter and also the best P solubilizer by
spectrophotometric method, and solubilized P concentration was 539.78
ppm at 5 days incubation period. SR was second best P solubilizing
strain and solubilized P concentration was 485.55 ppm and S7 had the
lowest P solubilizing activity among five strains.
Table 2: Soluble P Concentration of Five Isolates (ppm) in Pikovskaia’s Medium
by Spectrophotometric Method (at 830nm)
Isolates Incubation Periods (Days)
S5 269.54 539.78 152.74
S7 76.16 127.49 98.42
R6 62.5 157.3 102.05
R8 84.84 236.77 81.96
SR 121.28 485.55 252.57
Potassium Decomposing Activity of Isolated Endophytic Bacteria
Fourteen isolates except (LS, R2, S4, S8) could decompose potassium
(ranging from 2-30 mm in diameter) on K- decomposing medium with
0.12% K- mica and R3 was the best K-decomposer by plate screening
method and by AAS method (19.84 ppm).
Table 3: Soluble K Concentration (ppm) Decomposed by Two
Endophytic Strains in 3, 5 and 7 Days Incubation Periods by AAS
Method
Isolates Incubation Period (Days)
3 5 7
S3 3.36 14.91 18.73
R3 3.28 17.46 19.84
Results for Antagonistic Activity
S3 and R3 had good antagonistic activities against Rhizotonia solani and
LS and R5 against Phythium spp. and S7 to Fusarium oxysporum
respectively. They couldn’t be overwhelmed by these plant pathogens
and could live with these pathogens. So, this fact can benefit the host
plants they lived.
CONCLUSION
In this study, eighteen endophytic bacteria were isolated from stems and
roots of different rice varieties from different places. According to the
biochemical tests, five isolates were assumed to be Rhizobium spp. and
thirteen isolates were Azospirillum spp. although species identification
could not be done.
Most strains show high IAA producing activity at 2 days incubation
period by UV-VIS spectrophotometer method (530nm) and isolate R7
was found to be the best strain producing IAA, 120.55 ppm. In
analyzing nitrogen fixation activity by ammonium test kit, S5, S7, S8
isolates could fix nitrogen little and others couldn’t fix. In nitrogen free
semi-solid agar medium containing trace amount of bromothymol blue
as indicator, all isolates showed nitrogen-fixing activity during seven
days incubation period.
According to the results of phosphorous solubilizing activity by plate
screening method and by UV-Vis spectrophotometric method (830 nm),
isolate S5 was found to be the best phosphate solubilizer. R3 is the best
K-decomposer by plate screening method and by AAS method. S3 and
R3 have good antagonistic activities against Rhizotonia solani and LS
and R5 against Phythium spp. and S7 to Fusarium oxysporum
respectively. They couldn’t be overwhelmed by these plant pathogens
and could live with these pathogens. So, this fact can benefit the host
plants.

148
Acknowledgements
I would like to thank my supervisor, Dr. Aye Aye Khai and Dr. San San
Yu, Biotechnology Research Department, Kyaukse, for their generous
supports and helpful discussions throughout my research work. I also
deeply thank to my colleagues at our department for their kindly help
and suggestions.
REFERENCES
1. Kobayashi, D. Y. and Palumbo, J. D.: Bacterial endophytes and their
effects on plants and uses in agriculture, In: Bacon, C.W. and White, J. F.
(Eds.) Microbial endophytes. Marcel Dekker, Inc., N.Y., New York, 2000,
pp. 199-233.
2. Sturz, A. V., Christie, B. R., Matheson, B. G. and Nowak, J.: Biodiversity
of endophytic bacteria which colonize red clover nodules, roots, stems and
foliage and their influence on host growth, Biol. Fertil. Soils, 1997;25:13 –
19.
3. Kirchhorf, G., Reis, V. M., Baldani, J. I., Eckert, B., Döbereiner, J. and
Hartmann, A.: Occurrence, physiological and molecular analysis of
endophytic diazotrophic bacteria in gramineous energy plants, Plant Soil
1997;194:45-55.
4. Chen, C., Bauske, E. M., Mussan, G., Rodriguez-Kabana, R. and Kloepper,
J. W.: Biological control of Fusarium wilt on cotton by use of endophytic
bacteria, Biol. Control, ;9955:83-91.
5. Liu, L., Kloepper, J. W. and Tuzun, S.: Induction of systemi resistance in
cucumber against Fusarium wilt by plant growth-promoting rhizobacteria,
PhytoPath, 1995;5:695-698.
6. Sturz, A. V.: The role of endophytic bacteria during seed piece decay and
potato tuberization, Plant Soil, 1995;175:257-263.
7. Frommel, M. I., Nowak, J. and Lazorovits, G.: Treatment of potato tubers
with a Pham Quang Hung et al. growth promoting Pseudomonas sp.: plant
growth responses and bacterium distribution in the rhizosphere, Plant Soil,
1993;150:51-60.
8. McInroy, J. A. and Kloepper, J. W.: Survey of indigenous bacterial
endophytes from cotton and sweet corn, Plant Soil, 1995;173:337- 342.
9. Carroll, G. C.: The Biology of Endophytism in Plants with Particular
Reference to Woody Perennials. In: Microbiology of Phyllosphere
(Fokkema, N., J. and van den Heuvel, J., eds.), Cambridge University
Press, London, 1986;205-222.
10. Petrini, O.: Fungal Endophyte of Tree Leaves. In: Microbial Ecology of
Leaves (Andrews J and Hirano SS, eds.), Spring-Verlag, New York,
1991;179-197.
11. Saikkonen, K., Faeth, S. H., Helander, M. and Sullivan T. J.: Fungal
endophytes: a continuum of interactions with host plants, Ann. Rev. Ecol.
Syst, 1998;29: 319-343.
12. Misaghi, I. J. and Donndelinger, C. R.: Endophytic bacteria in symptom-
free cotton plants, Phytopathology, 1900;80: 808-811.
13. Chanway, C. P.: Inoculation of tree roots with plant growth promoting soil
bacteria: an emerging technology for reforestation, Forest Sci, 1997;43: 99-
112.
14. Ting, A. S. Y., Meon, S., Kadir, J. and Radu, S.: Endophytic
microorganisms as potential growth promoters of banana, BioControl,
2008;53: 541-553.
15. Araújo, W. L., Marcon, J., Maccheroni, W. J. and Van Elsas, J. D.:
Diversity of endophytic bacterial populations and their interaction with
Xylella fastidiosa in citrus plants, Appl. Environ. Microbiol, 2002;68:
4906-4914.
16. Lacava, P. T., Araújo, W. L., Marcon, J. and Maccheroni, W. J.: Interaction
between endophytic bacteria from citrus plants and the phytopathogenic
bacteria Xylella fastidiosa, causal agent of citrus-variegated chlorosis, Lett.
Appl. Microbiol, 2004;39: 55-59.
17. Yanni, Y. G., Rizk, R. Y., Corich, V., Squartini, A., Ninke, K., Philip-
Hollingsworth, S., Orgambide, G., Bruijn, F. D., Stoltzfus, R., Buckley, D.,
Schmidt, T., Mateos, P. F., Ladha, J. K. and Dazzo, F. B.: Natural
endophytic association between Rhizobium leguminosarum bv. trifolii and
rice roots and assessment of its potential to promote rice growth. Plant and
Soil, 1997;194: 99-114.
18. Mayak, S., Tirosh, T. and Glick, B. R.: Plant growth promoting bacteria
confer resistance in tomato plants to salt stress, Plant Physiology and
Biochemistry, 2004;42: 565-572.





























