Embed Size (px)
Citation preview

Cells and Materials Cells and Materials
Volume 2 Number 1 Article 5
1992
Evaluation of Bacterial Glycocalyx Preservation and Staining by Evaluation of Bacterial Glycocalyx Preservation and Staining by
Ruthenium Red, Ruthenium Red-Lysine and Alcian Blue for Several Ruthenium Red, Ruthenium Red-Lysine and Alcian Blue for Several
Methanotroph and Staphylococcal Species Methanotroph and Staphylococcal Species
Theresa A. Fassel University of Wisconsin-Madison
James E. Van Over Medical College of Wisconsin, Milwaukee
Christopher C. Hauser Medical College of Wisconsin, Milwaukee
Lorie E. Buchholz University of Wisconsin-Milwaukee
Charles E. Edmiston Medical College of Wisconsin, Milwaukee
See next page for additional authors
Follow this and additional works at: https://digitalcommons.usu.edu/cellsandmaterials
Part of the Biological Engineering Commons
Recommended Citation Recommended Citation Fassel, Theresa A.; Van Over, James E.; Hauser, Christopher C.; Buchholz, Lorie E.; Edmiston, Charles E.; Sanger, James R.; and Remsen, Charles C. (1992) "Evaluation of Bacterial Glycocalyx Preservation and Staining by Ruthenium Red, Ruthenium Red-Lysine and Alcian Blue for Several Methanotroph and Staphylococcal Species," Cells and Materials: Vol. 2 : No. 1 , Article 5. Available at: https://digitalcommons.usu.edu/cellsandmaterials/vol2/iss1/5
This Article is brought to you for free and open access by the Western Dairy Center at DigitalCommons@USU. It has been accepted for inclusion in Cells and Materials by an authorized administrator of DigitalCommons@USU. For more information, please contact [email protected].

Evaluation of Bacterial Glycocalyx Preservation and Staining by Ruthenium Red, Evaluation of Bacterial Glycocalyx Preservation and Staining by Ruthenium Red, Ruthenium Red-Lysine and Alcian Blue for Several Methanotroph and Ruthenium Red-Lysine and Alcian Blue for Several Methanotroph and Staphylococcal Species Staphylococcal Species
Authors Authors Theresa A. Fassel, James E. Van Over, Christopher C. Hauser, Lorie E. Buchholz, Charles E. Edmiston, James R. Sanger, and Charles C. Remsen
This article is available in Cells and Materials: https://digitalcommons.usu.edu/cellsandmaterials/vol2/iss1/5

CELLS & MATERIALS, Vol. 2, No. 1, 1992 (Pages 37-48) 1051-6794/92$3. 00 +. 00 Scanning Microscopy International , Chicago (AMF O'Hare) , IL 60666 USA
EVALUATION OF BACTERIAL GLYCOCALYX PRESERVATION AND STAINING BY RUTHENIUM RED, RUTHENIUM RED-LYSINE AND ALCIAN BLUE
FOR SEVERAL METHANOTROPH AND STAPHYLOCOCCAL SPECIES _./ 9 8b1 ,/ v 3
Theresa A. Fassel 1•2 , James E. Van Over2 , Christopher C. Hauser2 , Lorie E. Buchholz , Charles E. Edmist9114 , James R. Sange1} · and Charles C. Remsen 3/
1 Present Address: Muscle Biology Laboratory , Univ. Wisconsin-Madison , Madison, Wisconsin 53706 2Dept. Plastic and Reconstructive Surgery, Medical College of Wisconsin, Milwaukee, WI 53226
3Center for Great Lakes Studies, Univ. Wisconsin-Milwaukee, Milwaukee, Wisconsin 53204 4Surgical Microbiology Res. Lab. , Dept. Surgery , Medical College of Wisconsin, Milwaukee, WI 53226
(Received for publication September 24 , 1991, and in revised form March 2, 1992)
Abstract
The cationic reagen ts , ruthenium red (RR), alcian blue (AB) and ruthenium red -lysine were compared for ultrastructural preservation and staining of the bacterial glycocalyx. En bloc RR , AB and RR-lysine procedures, and a glutaraldehyde/Os04 fixation wi thou t cationic reagent , were evaluated by transmission electron microscopy (TEM) for several methanotroph and staphylococcal species. The glutaraldehyde/Os04 fixation was total ly ineffective in preserving or staining the glycocalyx material in all species studied . The RR procedure was more effective than the glutaraldehyde/Os04 fixation but was generally less so than the RR-lysine procedure. By the RR-lysine procedure , extensive layers that were ofte n filamentous or fibrou s were observed in the meth anotrophs, and extended fibrous elaborations were seen in the staphylococci. By the AB procedure, staphylococci glycocalyx material consisted of condensed curved structures or net-like features not directly comparable to the RR-lysine images. In the methanotrophs, filamentous or fibrous layers were often comparable between AB and RR-lysine . Thus , ultrastructural observation of the bacterial glycocalyx in all species studied was improved by use of RR-lysine , whereas the use of AB showed species variation.
Key Words: alcian blue, ruthenium red, ruthenium redlysine, bacterial glycocalyx, polysaccharide , staphylococci , methanotroph , en bloc reagent , staining , transmission electron microscopy.
*Address for correspondence: C.C. Remsen, Center for Great Lakes Studies, University of Wisconsin-Milwaukee , 600 E. Greenfield Avenue , Milwaukee , WI 53204
Telephone No. : (414)-649-3000
37
Introduction
Extracellular polysaccharides are a major component of structures external to the outer surface of microbial cells (Sutherland, 1990). If these structures are of bacterial origin and lie outside the integral elements of the outer membrane of gram-negative cells and outside the peptidoglycan of gram-positive cells, then by defi ni tion we are dealing with the bacterial glycocalyx (Costerton et al., 1981). The term glycocalyx was first used in this context by Bennett (1963). In general , there are two types of bacterial glycocalyx (Costerton er al., 1981) . One is composed of globular glycoproteins, often in regular arrays. These are also known as S layers (Sleytr, 1972). The other consists of fibrous matri ces of polysaccharide with variation in thickness and other characteristics such as flexibility and strength of association with the bacterial cell body (Costerton et al., 1981; van Herson, 1984). The polysaccharide is highly poly meric in nature and consists of numerous anionic moieties of variable composition (Costerton et al ., 1981; van Herson, 1984). The bacterial glycocalyx is also highly hydrated , as much as 99 % water (Sutherland , 1972).
These properties make effective observation of the bacterial glycocalyx by TEM difficult and often subject to uncertainty of interpretation . The aldehydes and osmium tetroxide generally are considered to have a low affinity for polysaccharide (Hayat , 1981). Insufficient stabilization by these chemical fixatives may result in observation of glycocalyx material not preserved. An alternative interpretation is that due to a lack of innate electron density and an inability to stain with conventional reagents (Roth, 1977), any preserved glycocalyx material is not visible in the TEM. During dehydration, the step-wise removal of water from the highly hydrated bacterial glycocalyx (Sutherland, 1972) can cause deformation or condensation (Roth, 1977; Springer and Roth, 1973). This adverse modification may be decreased or minimized by reagents with multiple attachment sites or by cross-linking agents (Costerton et al. , 1981). Lectins (Molinari et al., 1988; Orefici et al ., 1986) and specific antibodies (Molinari et al., 1987; Progulske and Holt , 1980) are two possible cross-linking approaches.
Another approach is the use of cationic reagents

T.A. Passel, et al.
traditional for specific or semi-specific staining of polyanions by light microscopy, such as ruthenium red (RR) and alcian blue (AB). Ruthenium red was first used to semi-specifically stain pectin (Jensen, 1962). This was extended to include acidic mucopolysaccharides, including acidic protein polysaccharides or glycoaminoglycans (Luft 1971a, 197lb). Many other substrates react with RR. The generalities concluded by Luft (197la, 1971b, 1976) were that reaction of RR is favorable with polyanions of high charge density. Neither high charge density alone, nor a high polymeric character alone, makes a favorable reaction. A relationship to the number of ionizable carboxylic acid groups of a polymer is possible. Neutral polysaccharides, polypeptides or proteins are generally unreactive (Luft, 197 la). Ruthenium red alone has been applied to study bacterial outer layers (Pate and Ordal 1967; Springer and Roth, 1973; Titus et al., 1982; Passel et al., 1990, 1992). For some bacterial species , considerable additional outer material of various descriptions are observed. In other cases , condensation or collapse, perhaps due to dehydration artifacts of the glycocalyx, is still reported (Costerton et al., 1981; Molinari et al., 1988). In general, where a high proteinaceous component (Buckmire and Murray, 1973; Mc Cowan et al., 1979; Costerton et al., 1974) or a glycocalyx well anchored to many attachment sites (Costerton et al., 1981) occurs, an extended structure is observed . Also, cross-linking antibodies applied in conjunction with RR have improved observation of extended fibrillar structures (Mackie et al., 1979; Patterson et al., 1975).
Ruthenium red was coupled with the diamine, lysine, and glutaraldehyde in another approach that makes use of the cross-linking concept. Jacques and Graham (1989), by inclusion of lysine in a prefixation with glutaraldehyde and RR, stabilized and visualized extensive glycocalyces for several bacterial species. This procedure has since been applied to ruminal bacteria (Akin and Rigsby, 1990), Clostridium difficile (Davies and Borriello, 1990) and Streptococcus suis (Jacques et al., 1990). It is based on the development of a glutaraldehyde-lysine fixation by Boyles (Boyles, 1982, 1984; Boyles et al., 1985). Lysine, which is positively charged at physiological pH, forms large polymers with glutaraldehyde. Cross-linkages of various lengths and directions with anionic sites of the glycocalyx could form stable configurations resistent to later stages of processing (Boyles, 1982, 1984; Boyles et al., 1985). Where the RR molecule exactly fits in this scheme, i.e., acting to stabilize or stain, is not yet established.
Another cationic reagent, alcian blue, was characterized by Scott (1972a, 1972b, 1972c, 1973, 1980). Alcian blue is thought to bind negatively charged moieties primarily through electrostatic or ionic linkages (Behnke and Zelander, 1970; Luft, 1971a; Scott et al., 1964). It has been used to stain acidic mucopolysaccharide (Luft, 1971a; Kiernan, 1990). For light microscopy, it reacts specifically only with sulphate-ester groups at pH 1.0 and with both carboxyl and sulfate groups at pH 2.5 (Kiernan, 1990; Jones and Reid, 1973).
38
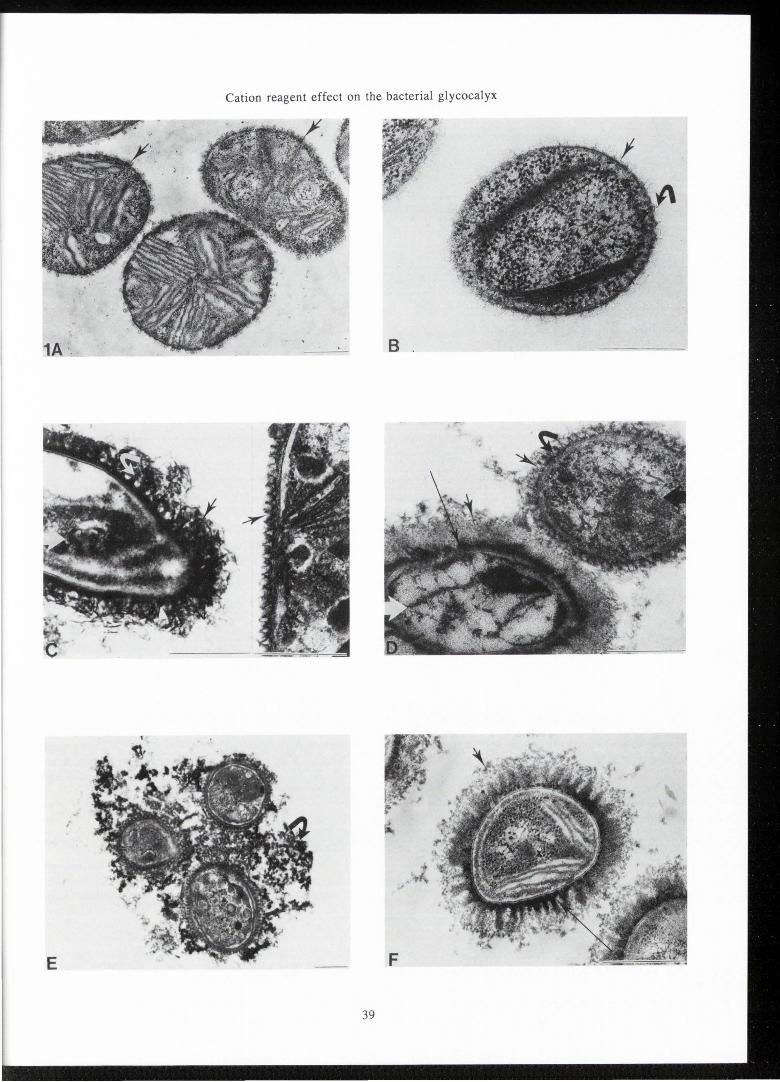
Figure 1. Transmission electron micrographs for Methylomonas albus BG8 (A, C, D, E) and for Methylosinus trichosporium Ob3b (B, C, D, F) are shown by the glutaraldehyde/Os04 fixation (A, B), the ruthenium red procedure (C), the alcian blue procedure (D) and the ruthenium red-lysine procedure (E,F). (A) Methylomonas albus BG8 by the glutaraldehyde/ Os04 fixation, shows cup-like features (arrows) in LR White resin. (B) Methylsinus trichosporium Ob3b by the glutaraldehyde/Os04 fixation, shows limited (arrow) and spikelike structures (curved arrow). (C) By the ruthenium red procedure, Me1hylomonas albus BG8 (cell indicated by large black arrow) cup features (arrow) with good definition are seen in a complete surrounding layer. For Methylosinus trichosporium Ob3b (cell indicated by large white arrow) round bead features (white arrowhead), thick filaments (curved arrow) and filamentous or fibrous material (arrow) are observed. (D) By the alcian blue procedure for Methylomonas albus BG8 (cell indicated by large black arrow), both the cup layer (arrow) and material within the cup (curved arrow) are retained. For Methylosinus trichosporium Ob3b (cell indicated by large white arrow), an inner electron dense layer (long arrow) is surrounded by an less dense outer fibrous layer (arrow). (E) By the ruthenium red-lysine procedure for Methylomonas albus BG8, a complete cup layer (arrow) with fibrous material and elaborate external material (curved arrow) between cells are observed. (F) By the ruthenium red-lysine procedure for Methylosinus trichosporium Ob3b, a dense thick innermost layer (long arrow), a gradation of material of varying electron density and a less dense fibrous outermost material (arrow) suggest complex layers.
All magnification bars are 0.5 µm.
In electron microscopy, it has been used in bacterial studies to demonstrate outer material in Eikenella strains (Progulske and Holt, 1980) , Pseudomonasfragi (Herald and Zottola, 1988) and several methanotrophs (Passel et al., 1992).
In this study, the use of RR and AB in previously reported procedures (Passel et al., 1990; 1991) are compared to the RR-lysine approach based on Jacques and Graham (1989). The comparison is carried out for several species of methanotrophs Methylomonas albus BG8, Methylosinus trichosporium Ob3b, Methylocystis parts OBBP, and Methylocystis species strain Lake Washington. They have been shown to play an important role in cycling of carbon in natural environments (Whittenbury and Dalton, 1981). Work with a totally unrelated group of organisms was desired to evaluate the utility of these approaches with other bacterial species. Species of the gram positive coagulase negative staphylococci were selected. Staphylococcal species studied are Staphylococcus aureus ATCC 25923, Staphylococcus epidermidis RP62 and Staphylococcus hominis SP2. These species
Cation reagent effect on the bacterial glycocalyx
E
39

T .A. Passel, et al.
c
Figure 2. Transmission electron micrographs for Methylocystts species strain Lake Washington and Methylocystts parts OBBP are shown for the ruthenium red procedure (A), the alcian blue procedure (B) and the ruthenium red-lysine procedure (C, D). (A) By the ruthenium red procedure, an electron dense fibrous layer (arrow) surrounds cells of Methylocystts parts OBBP. (B) By the alcian blue procedure, material in a fibrous layer (arrow) surrounds cells of Methylocystts species strain Lake Washington. (C, D) By the ruthenium red-lysine procedure, an extensive fibrous layer (arrow) is seen for (C) Methylocystts species strain Lake Washington and (D) Methylocystts parts OBBP. Some fibrous and/or filamentous material is observed near cells (curved arrow) of Methylocystts parts OBBP.
All magnification bars are 0.5 µm.
40
are widespread potential opportunistic pathogens and infectious agents (Edmiston et al., 1989). Through improved visualization of the glycocalyces of both groups of bacterial species, our environmental and medical studies will be facilitated. We hope other laboratories will also find this useful in their applications.
Material and Methods
Cell Culture
Methanotrophic species studied are Methylomonas albus BG8, Methylostnus trtchosportum Ob3b, Methylocystts parts OBBP, and Methylocystts species strain Lake Washington. They were grown as suspension cultures at 30 °C in nitrate mineral salts medium under 50:50 air: methane atmospheres until mid-to late log phase, 36 hours (Whittenbury and Dalton, 1981). Staphylococcal species are Staphylococcus aureus ATCC 25923, Staphy-

Cation reagent effect on the bacterial glycocalyx
3A
lococcus epidermidis RP62 and Staphylococcus hominis SP2 . These organisms were recovered from frozen storage (-70 °C) and plated on blood agar plates for test of viability. After 24 hrs the organisms were inoculated to trypticase soy broth and incubated for 18 hrs at 35 °C.
Cells were handled as pellets until all fixation/ wash procedures were completed. Then, they were enrobed in 4 % agar and handled as 1 mm 3 blocks.
Glutaraldehyde/Os04 fixation
Fixation was in 2.5% glutaraldehyde in buffer for 2 hours, followed by washes of 3X 10 minutes each. Postfixation was in 1 % (w/v) Os04 in buffer for 2.5 hours, followed by washes of 3X 10 minutes duration each. The buffer for these solutions was 0.1 M cacodylate pH 7. 0-7. 3. Dehydration was in a graded ethanol series of 10%, 25% , 50%, 70%, 95%, 100% anhydrous, and 100 % anhydrous. Infiltration and embedment was in Spurr resin, Standard Medium Mixture (Polysciences
41
Figure 3. Transmission electron micro graphs for Staphylococcus aureus ATCC 25933 is shown by the glutaraldehyde/Os04 fixation (A), the ruthenium red procedure (B), the alcian blue procedure (C) and the ruthenium red-lysine procedure (D). For Staphylococcus aureus ATCC 25933: (A) no glycocalyx material is seen by a glutaraldehyde/Os04 fixation. This is also observed for Staphylococcus hominis SP2 and Staphylococcus epidermidis RP62 (electron micrographs not shown). (B) By the ruthenium red procedure, sparse fibrous material (arrow) appears irregularly around cells. (C) By the alcian blue procedure, curved irregular electron dense structures (arrow) occur between and are frequently continous with cells. A net-like structure of fine strands (curved arrow) also is observed. (D) By the ruthenium red-lysine procedure, fibrous material (arrow) is extensively elaborated and surrounds cells.
All magnification bars are 0.5 µm.

T.A. Fassel , et al .
42

Cation reagent effect on the bacterial glycocalyx
Figure 4. Transmission electron micrographs for Staphylococcus hominis SP2 (A, C, E) and Staphylococcus epidermidis RP62(B, D, F) are shown by the ruthenium red procedure (A, B), the alcian blue procedure (C, D) and the ruthenium red-lysine procedure (E, F). (A, B) By the ruthenium red procedure , Staphylococcus hominis SP2 shows some fibrous material (arrow). Staphylococcus epidermidis RP62 shows considerably more glycocalyx material (arrow) around and between cells. (C, D) By the alcian blue procedure for both species, curved electron dense (arrow) structures occur that are frequently continuous with cells (shown for Staphylococcus hominis SP2 in C). A fine netlike structure (curved arrow) is continuous with cells of Staphylococcus hominis SP2 (C). It is shown completely surrounding a cell of Staphylococcus epidermidis RP62 (D). (E, F) By the ruthenium red-lysine procedure, the fibrous material completely surrounds each cell of Staphylococcus hominis SP2 (E). The fibrous material (arrow) is more extended and elaborated for Staphylococcus epidermidis RP62 (F). Additional extended material is seen between cells for both species (E and F).
All magnification bars are 0.5 µm .
Data Sheet #127), except for Methylomonas albus BG8 , where an LR White embedment proves more effective. This processing followed London Resin Company Limited, London, England; Data Sheet 305A. Thin sections were cut on a Porter-Blum MT-II using a diamond knife, and post-stained in 25 % ethanolic 2 % uranyl acetate and Reynolds lead citrate. Sections were studied on a Hitachi H-600 at 75 kV.
Ruthenium red en bloc procedure
Prefixation was in 0. 2 % glutaraldehyde and O. I5 % RR in buffer for 30 minutes. Fixation immediately followed in I% glutaraldehyde and 0.05 % RR in buffer for 2 hours. The washes were 3X 10 minutes each with 0. 05 % RR included in the wash buffer. Postfixation was in 2 % Os04 and 0.05 % RR in buffer for 2.5 hours. The washes were 3X 10 minutes each with 0.05 % RR included in the wash buffer. The buffer for these solutions was O. IM cacodylate pH 7.0-7.3. Dehydration in the graded ethanol series included 0.05 % RR through the 70 % stage. All remaining steps were carried out as above.
Alcian blue en bloc procedure
Prefixation was in 0.2 % glutaraldehyde and 0.15 % AB in buffer for 30 minutes. Immediately following fixation was in I% glutaraldehyde and 0.05 % AB in buffer for 2 hours. The washes were 3X 10 minutes each with 0.05 % AB included in the wash buffer. Postfixation was in 2 % Os04 and 0.05 % AB in buffer for 2.5 hours. The washes were 3X 10 minutes each with 0.05 % AB included in the wash buffer. The buffer for these solutions was 0.1 M cacodylate pH 7.0-7.3. Dehydration in the graded ethanol series included 0.05 % AB
43
through the 70% stage. All remaining steps were carried out as above.
Ruthenium red-lysine en bloc procedure
Prefixation was in 0.075 % RR, 2.5 % glutaraldehyde and 75 mM lysine in buffer for 20 minutes. Fixation was in 0.075 % RR and 2.5 % glutaraldehyde in buffer for 2 hours (Jacques and Graham, I989). This was followed by washing in buffer 3X 10 minutes each. Postfixation was in I% Os04 for 2 hours followed by 3X 10 minutes each washes. The buffer for these solutions was O.I M cacodylate pH 7.0-7.3. Further processing was identical to the glutaraldehyde/Os04 fixation procedure above.
Results
By the glutaraldehyde/Os04 fixation, in the absence of en bloc reagent, glycocalyx material is absent or limited for all species examined. In general, improvement is seen with cationic reagents.
In Figure I Methylomonas albus BG8 and Methylosinus trichosporium OB3b are shown. For Methylomonas alb us BG8 (Figure IA), accessory cup features (arrow) are observed external to the outer membrane of the gram-negative cell wall for a glutaraldehyde/osmium tetroxide fixation in LR White resin. These cup features are not preserved in Spurr resin without cationic reagents ruthenium red or alcian blue as previously shown (Passel et al., I990, 1992). For Methylosinus trichosporium OB3b (Figure lB), a few spike-like structures (curved arrow) and suggestion of some limited material (arrow) are observed external to the outer membrane irrespective of embedding resin. This figure and all following electron micrographs are from Spurr resin embed men ts.
Figure lC shows the ruthenium red procedure. The Methylomonas albus BG8 cell is indicated by a large black arrow. Cup features (arrows) are retained with good definition . As previously reported (Passel et al., I990), cups are seen in a gap-less layer surrounding the outer membrane. The Methylosinus trichosporium OB3b cell in Figure IC is indicated by large white arrow. Features seen with ruthenium red include round beadlike structures (white arrowhead), thickened electron dense filaments (curved arrow), and some fibrous material (arrow). To aid description of glycocalyx, the term filamentous is used where indivjdual strands or filaments can be easily identified. The term fibrous is used for outer material where separate and distinct strands or individual filaments are not easily discerned.
By the alcian blue procedure (Figure ID) for Methylomonas a!bus BG8 (cell indicated by large black arrow), fibrous material (curved arrow) appears in cups (arrow). For Methylosinus trichosporium OB3b (cell indicated by large white arrow), two layers of different electron density are observed. A dense inner layer (long arrow) is completely surrounded by a more amorphous or fibrous outer layer (arrow), as detailed previously (Passel et al., I 992).

T.A. Passel , et al.
By the ruthenium red-lysine procedure , several observations similar to the above are seen for Methylomonas albus BG8 (Figure lE) . A well defined and complete cup layer is preserved (arrow). Fibrous material appears within cups. Additionally, more elaborate material frequently extends between cells (curved arrow). For Methylosinus trichosporium OB3b (Figure IF) by the ruthenium red-lysine procedure , an electron dense inner material (long arrow) is frequently seen. A gradation of electron density in the other glycocalyx material occurs with frequent suggestion of an outermost fibrous layer (arrow) around many cells.
For Methylocystis species strain Lake Washington and Methylocystis paris OBBP by a glutaraldehyde/Os04 tetroxide fixation, observation of glycocalyx is limited to remnants of spikes , or strands , or is completely absent (electron micrographs not shown), as was seen for Methylosinus trichosporium OB3b. By the ruthenium red procedure (Figure 2A) , a matted fibrous layer (arrow) is found for some cells , as shown here for Methylocystis paris OBBP. For other cells , abundant filamentous material is often seen . Additionally , for some cells of Methylocystis species strain Lake Washington , discrete bead-like structures were observed (Passel et al. , 1992). By the alcian blue procedure , a fibrous layer (arrow) completely surrounds cells of Methylocystis species strain Lake Washington (Figure 2B) as is similarly found in cells of Methylocystts parts OBBP (Passel et al. , 1992).
By the RR-lysine procedure , cells are also surrounded by an extensive fibrous layer (arrow) for Methylocystts species strain Lake Washington (Figure 2C) and Methylocystis parts OBBP (Figure 2D) . For the latter , filamentous or fibrous material (curved arrow) is also seen frequently in a mass between cells. An electron translucent region, possibly artifact, between the fibrous layer and the cell wall is noted for some cells.
For Staphylococcus aureus ATCC 25923 (Figure 3A) and the other staphylococci species studied, no suggestion of glycocalyx material is seen by a glutaraldehyde/Os04 fixation. In S. aureus (Figure 3B-3D) , by the ruthenium red procedure (Figure 3B) sparse fibrous material (arrow) occurs irregularly around cells. Where there is a grouping of cells , more material is frequently seen on the outermost cells. By the alcian blue procedure (Figure 3C), curved irregular electron dense structures (arrow) appear continuous with cells and extend between cells. A netlike structure that appears to consist of fine strands (curved arrow) is occasionally seen . In contrast, by ruthenium red-lysine (Figure 3D) , an elaborate fibrous material (arrow) surrounds cells and extends between cells.
By the ruthenium red procedure for Staphylococcus hominis SP2 (Figure 4A), some fibrous material (arrow) is seen. For Staphylococcus epidermidis RP62 (Figure 4B), more fibrous material (arrow) is observed. By the alcian blue procedure, for Staphylococcus hominis SP2 (Figure 4C) and Staphylococcus epidermidis RP62 (Figure 4D), curved electron dense structures
44
(arrow) are seen. A netlike structure (curved ran"Jw) is shown continuous with some Staphylococcus hJminis SP2 cells (Figure 4C) and completely surroundiin~ a cell of Staphylococcus epidermidis RP62 (Figure t4D). By the ruthenium red-lysine procedure for Staphy1lowccus hominis SP2 (Figure 4E) , fibrous material (arrrov) surrounds the cell body as a more complete layer. I addition, more elaborate material extends between c1el ls . For Staphylococcus epidermidis RP62 (Figure 4F), <elaborate and extensive fibrous material surrounds cellls md is maintained between cells (arrow).
Discussion
The bacterial glycocalyx is defined as s;tnctures containing polysaccharide of bacterial origin arnc lying outside the integral elements of the outer merrnbrane of gram-negative cells and outside the peptidog ly:an of gram-positive cells (Costerton et al. , 1981) . It is rich in highly varied and diverse polymeric polysac ct.arides (Costerton et al. , 1981 ; van Herson , 1984) an d! is also a highly hydrated structure containing 99 % water (Sutherland , 1972) . These characteristics make:s ptimizing their observation for transmission electron microscopy challenging. One approach is the use of cationic reagents , RR and AB , in en bloc procedures wi tth chemical fixatives . A previous successful method util izing RR or AB improved observation of glycocalyx mate rial over a glutaraldehyde/Os04 fixation for several sp ecies of methanotrophs. This material bore a resemblance to freeze etch images without cationic reagent or chemical fixative (Passel et al ., 1990; 1992). The use of a crosslinking reagent , the diamine lysine , with glutar aldehyde was developed by Boyles (Boyles , 1982, 1984; Boyles et al. , 1985) and applied to fi xation of actin filaments, extracellular matrix and the mammalian gly cocalyx. This approach was combined with the use of RR by Jacques and Graham (1989) and applied to the bacterial glycocalyx by themselves and several authors (Akin and Rigsby, 1990; Davies and Borriello, 1990 ; Jacques et al. , 1990).
Two general types of bacterial glycocalyx have been described. One is composed of Slayers of globular glycoproteins often in regular arrays (Sleytr, 1972). The other is a complex fibrous matrix (Costerton et al., 1981). Both are represented in the species of the present study.
Many gram-negative species have regular arrays of wine glass- , globet- or cup-shaped subunits that are considered by Sleytr (1972) as special types of S layers. Examples include Flexibacter polymorphus (Ridgway, 1977), several species of Chromattum (Remsen et al., 1970; Hageageand Gherna, 1970, 1971) andMethylomonas alb us. The cup layer of Methylomonas albus strain BG8 , fits the Slayer definition of Sletyr (1972), and was identified by negative staining and freeze etch electron microscopy by Haubold (1978) and in thin section by Wilkinson (1971).
By TEM , the cup layer was not observed in Spurr

Cation reagent effect on the bacterial glycocalyx
embedments without RR (Passel et al., 1990), AB (Passel et al., 1992) or RR-lysine. It was seen without these reagents in LR White. However, the cups lacked strong defi nition as individual structures and were often not in a complete layer around the outer membrane (Passel et al., 1990). Further, the improvement in the cup layer by RR is not increased by either AB or the use of lysine. The appearance of the cup layer by RR compares directly with the images obtained by freeze etch, where cationic reagents or chemical fixatives were not used (Passel et al., 1990). Based on the Spurr observations alone , one cannot differentiate between the improved preservation of the cups or the alternative possibility of improved staining of otherwise invisible cup features. However , the inferior appearance of cups in LR White (Passel et al. , 1990), their improvement with RR in either resin (Passel et al., 1990) and the agreement with the freeze etch cup layer (Haubold , 1978; Passel et al., 1990) suggest an improvement in preservation of the cup layer by RR processing. This improvement also agrees with the suggestions of a cup layer composition that includes acidic mucopolysaccharides, acidic protein polysaccharides or glycoprotein.
Additionally, fibrous material appears within cups by the AB or RR-lysine procedure. Elaborate material that frequently extends between cells is seen by RRlysine processing. For Methylomonas a/bus BG8, RR alone is inadequate for observation of this glycocalyx material. Perhaps this composition is sufficiently different from the cup layer that RR alone is insufficient to effect its preservation or staining.
Glycocalyces of the remaining three methanotrophs appear to fall under the fibrous matrix type. In previous work, freeze etch was compared to a glutaraldehyde/Os04 fixation without cationic reagent. For Methylosinus trichosporium Ob3b and the two Methylocystis species, large filamentous and complex layers were revealed outside the outer membrane of these gram-negative cells by freeze etch after ultra-rapid freezing (Passel et al, 1990; 1992). When fixed by a glutaraldehyde/ Os04 procedure, spike-like structures or a complete lack of any glycocalyx material was observed. This can be interpreted as suggestive of collapse and/or condensation perhaps due to an ethanol dehydration artifact (Bayer and Thurow, 1977) that is minimized or avoided by ultra-rapid freezing and by freeze etch. An alternative explanation is this material lacks innate electron density and fails to stain by conventional procedures (Roth, 1977). Perhaps some combination of both dehydration artifact and inability to stain may occur.
When cells of Methylosinus trichosporium Ob3b and the two Methylocystis species were processed in a procedure with AB extensive layers that were filamentous or fibrous were observed. For Methylosinus trichosporium Ob3b, this glycocalyx material appears as two layers of distinct electron density. This was interpreted as suggesting a segregation of different polyanions (Passel et al., 1992). By the RR procedure for these three species, although some fibrous layers were ob-
45
served, a variety of other features have consistently been seen for some cells. This includes round bead-like structures, thick filaments, and filamentous layers (Passel et al., 1990, 1992; Titus et al., 1982).
The use of lysine with RR, resulted in images that were markedly similar to the AB fibrous layer surrounding the two Methylocystis species. However, there was a clear zone between the cell outer membrane and the fibrous layer. This may be suggestive of an artifactual cell shrinkage, an area of washout, lack of stain or other artifact. Where cells were observed with a fibrous layer by RR processing, or for all cells by AB processing, this clear zone was not seen. However, a few cells on occasion had evidence of shrinkage that was internal to the outer membrane . Also, for Methylocystis paris by RRlysine a more massive congregation of filamentous or fi brous material was noted between cells . For Methylosinus trichosporium Ob3b by RR-lysine, the gradation of electron density in glycocalyx material may again suggest a separation of polyanions. The clear zone was not seen for this species. This may suggest a greater vulnerability to this type of artifact by the Methylocystis species. On the basis of the observation of bead-like structures and thick filaments by RR and their absence by RR-lysine or AB , one could conclude these features are perhaps due to collapse of more filamentous or fibrous material, and their insufficient stabilization or stain reaction with RR. However , the bead-like structures have been observed by scanning electron microscopy (SEM) in the absence of RR or other cationic reagent (Passel et al., 1990; Titus et al. , 1982). There is some suggestion of this structure by freeze etch (Passel et al., 1990). Alternatively, their lack of observation by RRlysine or AB might mean an insufficient stabilization or other damage that results in loss of their structural integrity. Perhaps, the composing polymers of the beads are split apart and the structures broken to merge visually with other fibrous or filamentous material.
It was desirable to study the effects of these procedures on glycocalyx preservation or staining on a different group of bacteria. For the staphylococcal species studied, there was no evidence of a glycocalyx by the glutaraldehyde/Os04 fixation even in species known for production of external polysaccharide as a slime layer. By RR some fibrous material was observed for all species studied. A considerable increase in the amount of fibrous material in extended configuration around and between cells was seen in RR-lysine processed cells for all species. For SP2, an incomplete layer close to the cell body by RR was replaced by a more complete layer surrounding the cell by RR-lysine. This suggests an improvement in preservation and/or staining of this glycocalyx, possibly by a more efficient crosslinkage and stabilization. For RP62, the difference between RR and RR-lysine processing is less obvious with considerable material observed for both.
A very different image was seen by AB processing for all species of staphylococci studied . Curved electron dense structures and net-like structures may be

T.A . Passel , et al.
the result of condensations or distortions of glycocalyx material for these cells. This is very likely due to insufficient stabilization and subsequent collapse. The lack of agreement with the extended configurations seen with RR-lysine also suggests the inability of AB to interact as effectively with the components of the staphylococci glycocalyx .
In summary , for the methanotrophs , extended filamentous or fibrous configurations were generally similar between AB and RR-lysine procedures. The additional cross-linking ability of the lysine procedure does not appear superior to the interaction of alcian blue with these glycocalyces. For the staphylococci , the most abundant and extended glycocalyces were observed for cells by RR-lysine processing. Thus perhaps , the glycocalyx for staphylococci are of a biochemical and/or charge distri bution that are preferentially preserved and/or stained by processing that cross-links glutaraldehyde , lysine and polysaccharide polymers and interacts with RR . The difference in effectiveness of the methods using RR , AB or RR-lysine appears somewhat genera/species dependent. Di fferences in complex polyanionic polysaccharide composition of the glycocalyx for the different bacteria could account for these observations .
Acknowledgements
Th e authors wish to acknowledge Dr. Ken Seigesmund , Medical College of Wisconsin , for access to electron mi croscopy instrumentation and Marilyn J . Schaller for her continued and valuable insight and positive support. This work was supported in part by grant 2022-IRP-79 from NASA and in part by grant MCB-9118653 from NSF . This is contribution No. 360 from the Center for Great Lakes Studies.
References
Akin DE, Rigsby LL (1990) Preservation of ruminal bacterium capsules by using lysine in the electron microscopy fixative. Appl. Envir. Microbiol. 56, 2933-2935.
Bayer ME, Thurow H (1977) Polysaccharide capsule of Eschericha coli: Microscope study of its size , structure, and sites of synthesis. J. Bacteriol. 130, 911 -936.
Behnke 0 , Zelander T ( 1970) Preservation of intercellular substances by the cationic dye alcian blue in preparative procedures for electron microscopy . J. Ultrastructure Research, 31, 424-438.
Bennet HS (1963) Morphological aspects of extracellular polysaccharides. J. Histochem. Cytochem. 11, 14-25.
Boyles JK (1982) A modified fixation for the preservation of microfilaments in cells and isolated Factin. J. Cell Biol. 95 , 287a (abstract).
Boyles JK, Anderson L , Hutcherson P (1985) A new fixative for the preservation of actin filaments: fixation of pure actin filament pellets. J. Histochem.
46
Cytochem. 33, 1116- 1128. Boyles JK (1984) The use of primary amines to
improve glutaraldehyde fixation. In: Science of Biological Specimen Preparation for Microscopy and Microanalysis , SEM , Inc. , AMR O'Hare (Chicago), IL 60666-0507 , USA , p. 7-21.
Buckmire FLA , Murray RGE (1973) Studies on the cell wall of Spirillum serpens. II. Chemical characterization of the outer structured layer. Can. J. Microbiol. 19 , 59-66.
Costerton JW , Damgaard HN , Cheng KJ (1974) Cell envelope morphology of rumen bacteria. J. Bacteriol. 118 , 1132- 1143.
Costerton JW , Irvin RT , Cheng KJ (1981) The bacterial glycocalyx in nature and disease. Ann . Rev. Microbiol. 35, 299-324.
Davies HA , Borriello SP (1990) Detection of capsule in strains of Clostridiun difficile of varying virulence and toxigenicity. Microbial Pathogenesis . 9 , 141-146 .
Edmiston CE, Schmitt DD , Seabrook GR (1989) Coagulase-negative staphylococcal infections in vascular surgery: epidemiology and pathogenesis. Infect. Control Hosp. Epidemiol. 10, 111 -117 .
Fassel TA , Schaller MJ, Lidstrom ME and Remsen CC (1990) Effect of fixation -resin combinations and ruthenium red on elucidating outer envelope structure and surface morphology of two methanotrophic bacteria . J. Electron Microsc. Tech. 14, 52-62 .
Passel TA , Schaller MJ , Remsen CC. (1992) Comparison of alcian blue and ruthenium red effects on preservation of outer envelope ultrastructure in methanotrophic bacteria. Microscopy Research and Technique 20 , 87-94.
Hageage GJ, Gherna RL ( 1970) Electron microscopy of the cell envelope of Chromatium warimgii. In: Bacteriol. Proc . Am. Soc. Microbiology, Sulkin SE, Wood WA (eds.) , Annual Reviews Inc. , Palo Alto , CA , p. 57.
Hageage GJ , Gherna RL (1971) Surface structure of Chromatium okenii and Chromatium weissei. J. Bact. I 06 , 687-690.
Haubold R (1978) Two different types of surface structures of methane utilizing bacteria. Zeitschrift fi.ir Allgemeine Mikrobiologie 18, 511-515.
Hayat MA (1981) Fixation for electron microscopy. Academic Press , New York, p. 64-182.
Herald PJ, Zottola EA (1988) The use of transmission electron microscopy to study the composition of Pseudomonas fragi attachment material. Food Microstructure 7, 53-57.
Jacques M, Graham L (1989) Improved preservation of bacterial capsule for electron microscopy. J. Electron Microscopy Technique. 11, 167-169.
Jacques M , Gottschalk M , Foiry B, Higgins R (1990) Ultrastructural study of surface components of Steptococcus suis. J. Bacteriol. 172, 2833-2838.
Jensen WA (1962) Carbohydrates and cell wall constituents . In: Botanical Histochemistry: Principles

Cation reagent effect on the bacterial glycocalyx
and Practice. W.H. Freeman, San Francisco , CA , p. 175-208.
Jones R, Rtid L (1973) The effect of pH on alcian blue staining of epithelial acid glycoproteins. I. Sialomucins and sulfomuc.ns (singly or in simple combinations). Histochem. J. 5, )-18.
Kiernan JA (1990) Carbohydrate histochemistry. In: Histological ard Histochemical Methods: Theory and Practice. p. 170-197.
Luft JH (197 la) Ruthenium red and violet. I. Chemistry, purification , methods of use for electron microscopy and mechanism of action. Anat. Rec. 171 , 347-368.
Luft JH (1 7lb) Ruthenium red and violet. II. Fine structural localization in animal tissue. Anat. Rec . 171, 369-415.
Luft JH (1976) The structure and properties of the cell surface coat. rnternat. Rev. Cytol. 45, 291-382.
Mackie EB, Brown KN, Lam J, Costerton JW (1979) Morphological stabilization of capsules of group B streptococci, t ~pes Ia, lb, II and III with specific antibody. J. Bact.138 , 609-617.
Mc Cowan lP, Cheng KJ, Costerton JW ( 1979) Colonization of a Portion of the bovine tongue by unusual filamentous Jacteria. Appl. Envir. Biol. 37, 1224-1229.
Molinari A Orefici G, Donelli G, Von Hunolstein C, Paradisi S, Ara1cia G. ( 1988) Preservation of capsular material of streptccoccal cells by specific lectins determined by immunoelectron microscopy. Histochem . J. 20 , 526-530.
Molinari /-,, Von Hunolstein C, Donelli G, Paradisi S, Aranci.3. G, Orefici G (1987) Effects of some capsular componeits on pathogenicity of type IV and provisional type V group B streptococci. FEMS Microbial. Lett. 41, 69-72.
Orefici G, Molinari A, Donelli G, Paradisi S, Teti G, Arancia G 1986) Immunolocation of lipoteichoic acid on group B st1eptococcal surface. FEMS Microbiology Letters. 34, 111-115.
Pate J, Or al EJ ( 1967) The fine structure of Chondrococcus cd umnaris. J. Cell Biol. 35, 37-50.
Patterson I-:, Irvin R, Costerton JW, Cheng KJ (1975) Ultrastruct1re and adhesion properties of Ruminococcus a/bus. J. 3act. 122, 278-287.
Progulske A, Holt SC (1980) Transmissionscanning electron microscopic observations of selected Eikenella coroden: strains. J. Bact. 143, 1003-1018.
Remsen CC, Watson SW, Truper HG (1970) Macromolecular s bunits in the walls of marine photosynthetic bacteria. J. Bact. 103, 254-258.
Ridgway H? (1977) Ultrastructural characterization of globlet-sh1ped particles from the cell wall of Flexibacter polymorphus. Can. J. Microbial. 23, 1201-1213.
Roth IL (1977) Physical structure of surface carbohydrates. It: Surface carbohydrates of the prokaryotic cell, ~ utherland I (ed.), Academic Press, New York, p. 5-20.
47
Scott JE, Quintarelli G, Dellovo MC (1964) The chemical and histochemical properties of alcian blue. I. the mechanism of alcian blue staining. Histochemie. 4, 73-85.
Scott JE (1972a) The histochemistry of alcian blue. Note on the presence and removal of boric acid as the major diluent in alcian blue 8 GX . Histochemie. 29, 129-133.
Scott JE (1972b) Histochemistry of alcian blue. II. The structure of alcian blue 8 GX. Histochemie. 30, 215-234 .
Scott JE (1972c) Histochemistry of alcian blue. III. The molecular biological basis of staining by alcian blue 8GX and analogous phthalocyanins. Histochemie. 30 , 191-212.
Scott JE (1973) Alcian dyes: LC.I. Cease manu-facture and release details of composition. Histochemie. 37 ' 379-380.
Scott JE (1980) The molecular biology of histochemical staining by cationic phthalocyanin dyes: the design of replacements for alcian blue . J. Micros. 119, 373-381.
Sleytr UB (1972) Regular arrays of macromolecules on bacterial cell walls: structure, chemistry, assembly, and function. Internat. Rev. Cytol. 53, 1-64.
Springer EL, Roth IL (1973) The ultrastructure of the capsules of Diplococcus pneumoniae and Klebsiella pneumoniae stained with ruthenium red. J. Gen. Microbial. 74, 21-31.
Sutherland IW (1972) Bacterial exopolysaccharides. In: Advances in Microbial Physiology, Rose AH, Tempest DW (eds.), Academic Press , New York. Vol. 8, p. 143-213.
Sutherland IW (1990) Biotechnology of microbial exopolysaccharides. Cambridge University Press. p. 1-11.
Titus JA, Reed WM, Pfister RM, Dugan PR (1982) Exospore formation in Methylosinus trichosporium. J. Bacterial. 149, 354-360.
van Herson W (1984) Coverings of the outer cell wall surface. In: Outer Structures of Bacteria, Van Nostrand Reinhold Company, New York, p. 155-200.
Whittenbury R, Dalton H (1981) The methylotrophic bacteria. In: The Prokaryotes, Starr MP, Stolp H, Truper HG, Balows A, Schegel HG, (eds.), SpringerVerlag, Heidelberg, p. 894-902.
Wilkinson JF (1971) Hydrocarbons as a source of single-cell protein . Symp. Soc. Gen. Microbial. 21, 15-46.
Discussion with Reviewers
T. Zelander: Having in mind the highly hydrated condition of the glycocalyx material, it occurred to me, that the spinning of unfixed bacteria to a pellet might influence the distribution and appearance of the glucocalyx material as seen in some of the pictures. Did you consider this possible influence or have I totally misunderstood your paragraph on methods?

T.A. Passel, et al.
Authors: We cannot conclusively rule out any effect of the centrifugation on the glycocalyces. However, of the two types of glycocalyces we feel the S layers to be less likely effected. The fibrous/filamentous matrices may be the more susceptible in the extended nature of their layers. Previous freeze etch comparisons for the methanotrophs with fibrous/filamentous matrices suggested a good correspondence between freeze etch and images obtained by the alcian blue procedure (Passel et al., 1992). Therefore, we feel confident with our methods.
48










![Functionalizing the glycocalyx of living cells with ... · Functionalizing the glycocalyx of living cells with supramolecular guest ligands for cucurbit[8]uril-mediated assembly](https://img.dokumen.tips/doc/110x75/5ec159ef491c257e8647d3c4/functionalizing-the-glycocalyx-of-living-cells-with-functionalizing-the-glycocalyx.jpg)


















![Vinther, J. (2016). Fossil melanosomes or bacteria? A wealth of …€¦ · highlighted the importance of bacterial activity in exceptional fossil preservation [2-4]. The role of](https://img.dokumen.tips/doc/110x75/5f0fb35a7e708231d44574ab/vinther-j-2016-fossil-melanosomes-or-bacteria-a-wealth-of-highlighted-the.jpg)