Embed Size (px)
Citation preview

Evaluating the performance of phylogeneticinference methods on heterogeneous data
Mark HolderDepartment of Ecology and Evolutionary BiologyUniversity of KansasLawrence, Kansas, USA

Thanks:
• Dr. Ziheng Yang, Dr. Nick Goldman and the Royal Society
• Dr. Derrick Zwickl
• Dr. Gavin Naylor, Dr. Junhyong Kim, Dr. David Hillis, Dr. Paul Lewis,Dr. Dave Swofford, Tracy Heath, Dr. A. Town Peterson
• National Science Foundation

Weaknesses of early models of sequence evolution
They assumed that
• the process of evolution is identical across all sites,
• the process of evolution is fixed over time,
• sites evolve independently of one another.
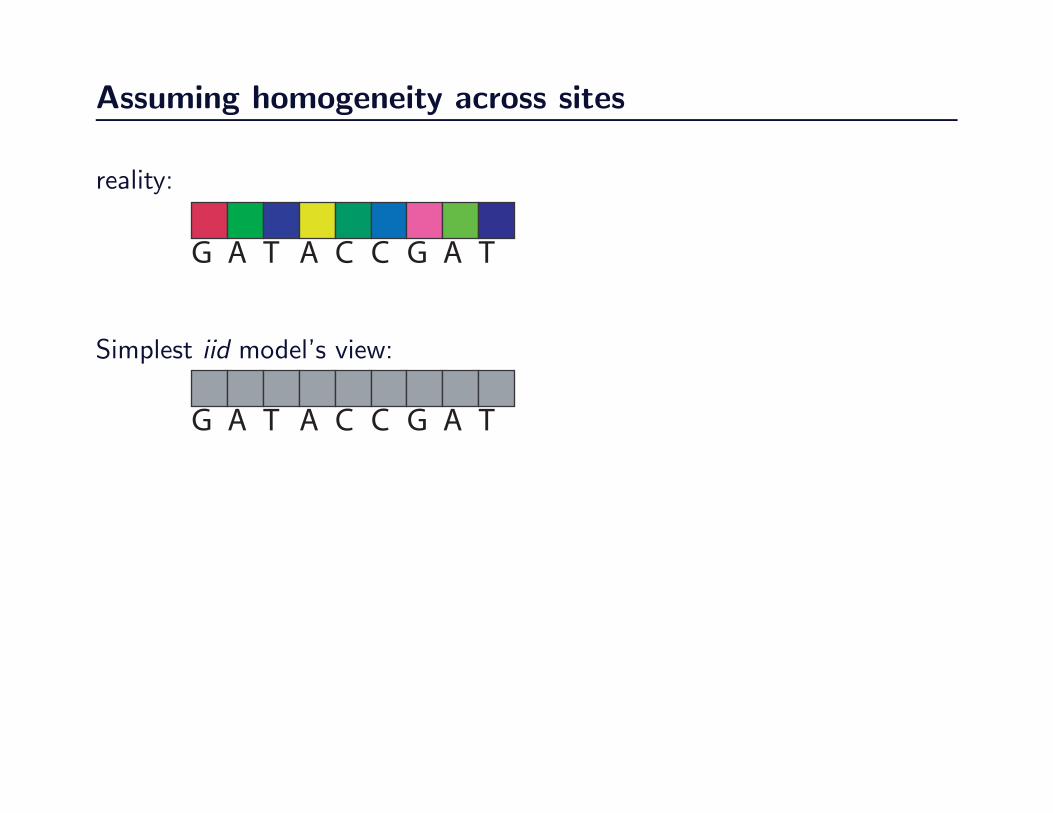
Assuming homogeneity across sites
G A T A C C G A T
G A T A C C G A T
reality:
Simplest iid model’s view:
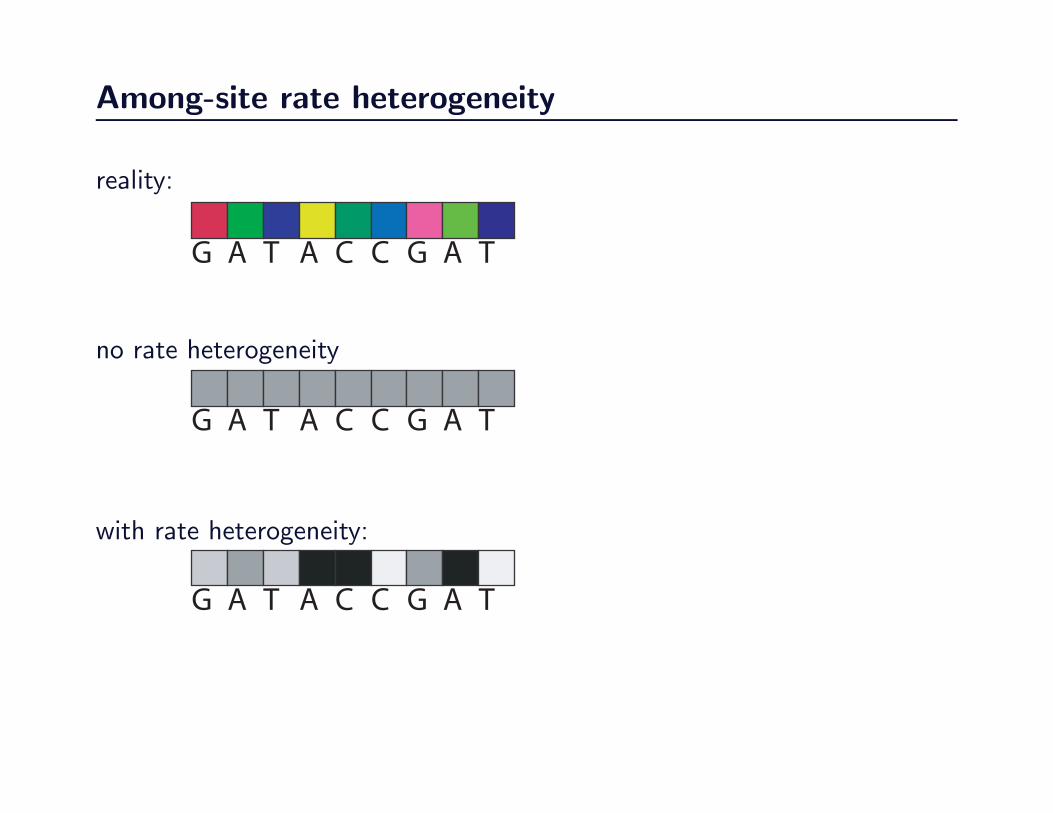
Among-site rate heterogeneity
G A T A C C G A T
G A T A C C G A T
G A T A C C G A T
reality:
no rate heterogeneity
with rate heterogeneity:

Among-site heterogeneity in substitution patterns
G A T A C C G A T
G A T A C C G A T
Reality:
Partitioned or mixure model:
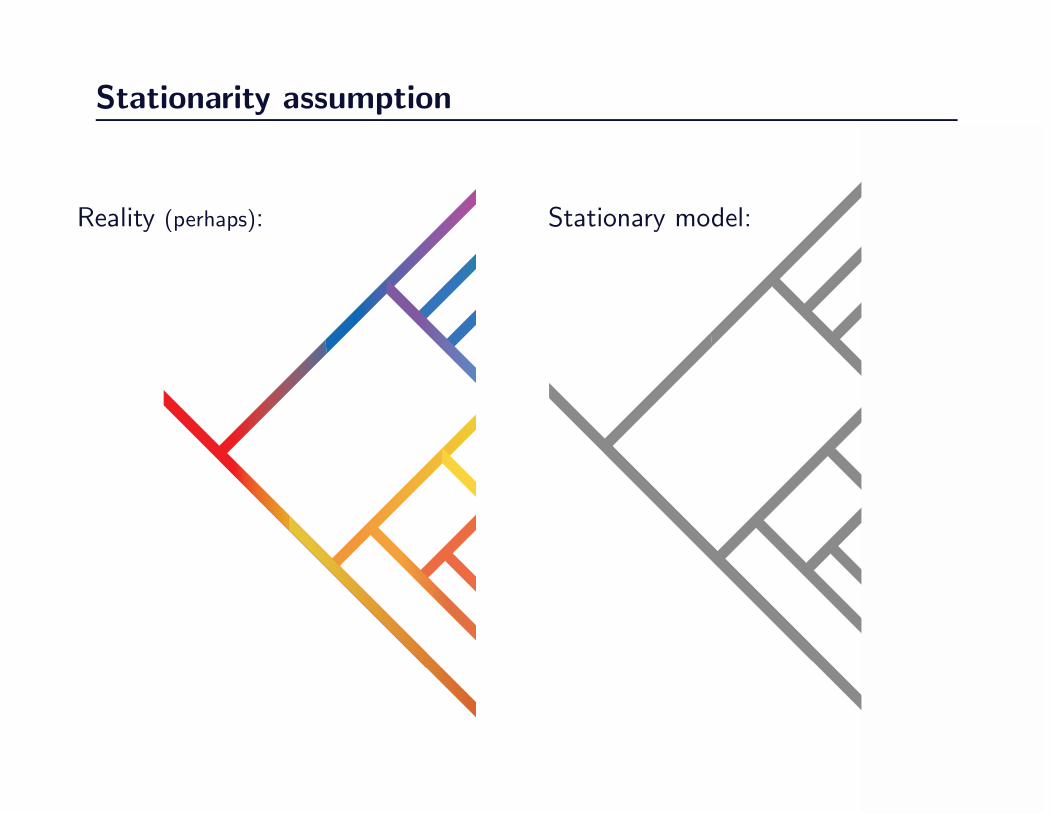
Stationarity assumption
Reality (perhaps): Stationary model:

Changes in evolutionary process over time
• Covarion models
• State frequency changes
• models that allow for changes to ω on a specific branch
• relaxed and local clock methods
. . .
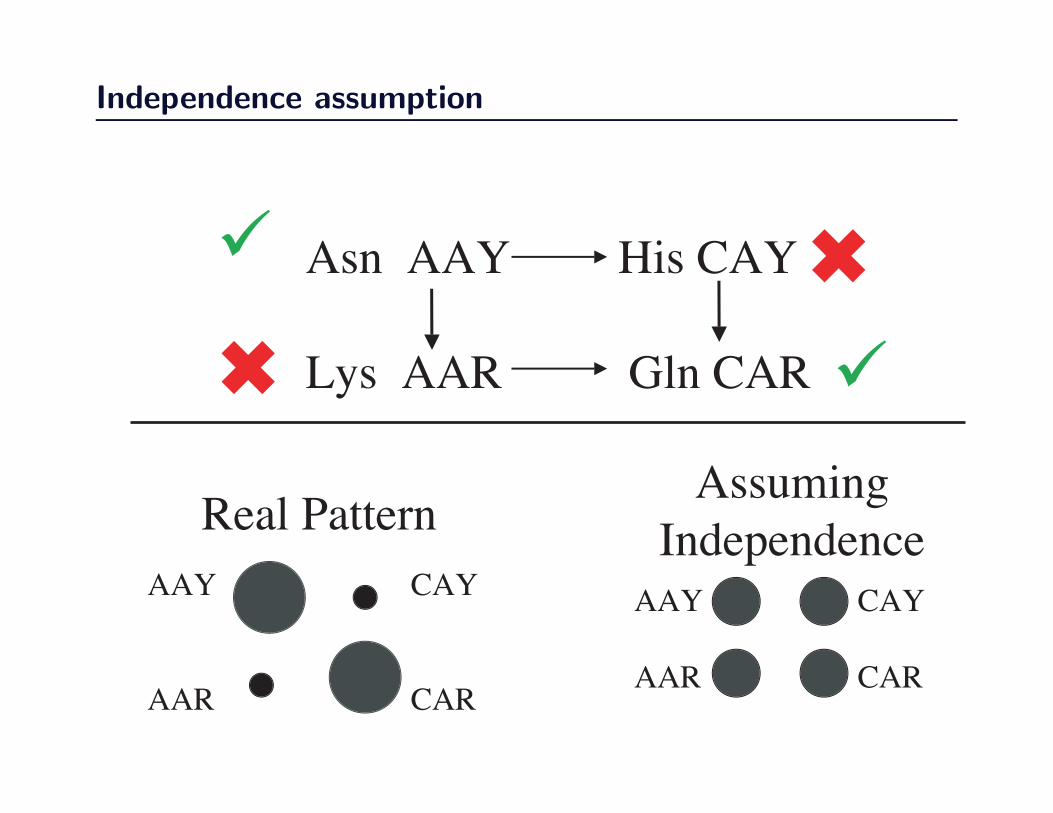
Independence assumption
Real PatternAssuming
Independence
Asn AAY His CAY
Lys AAR Gln CAR6
6¸
¸
AAY
AAR
CAY
CAR
CAY
CAR
AAY
AAR

Interactions among sites and neighborhood effects
• Codon models
• Doublet models for RNA
• Context-dependent models
. . .

Amazing progress in the development of models for phylogenetic inference,but . . .
• Are current models sufficient?
• How should we decide between alternative models?

• Are current models sufficient?
Not for difficult “deep phylogeny” questions. For example, it is fairlycommon to see strong support for a grouping, but sensitivity to taxonsampling.
• How should we decide between alternative models?
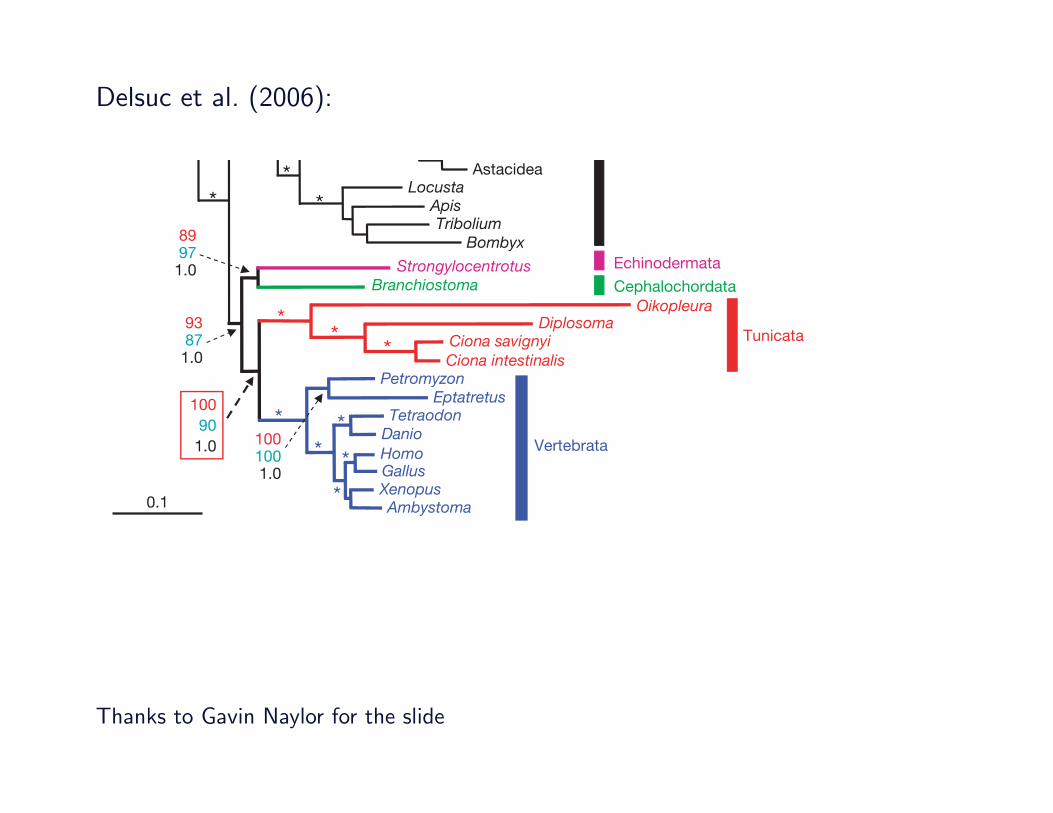
Delsuc et al. (2006):
©!2006!Nature Publishing Group!
!
alternative in which cephalochordates emerge before echinoderms.These two topologies appeared significantly worse that theML tree ofFig. 1, even for the conservative Shimodaira–Hasegawa test13.Our results therefore indicate a strong phylogenetic affinity
between tunicates and vertebrates to the exclusion of cephalo-chordates. However, obtaining high statistical support for a giventopology does not necessarily indicate that the phylogenetic inferenceis correct. Indeed, the phylogenetic analysis of large-scale data setsrequires particular attention to potential systematic biases associated,for instance, with differences in evolutionary rates among species,compositional biases and heterotachy8. In particular, a long-branchattraction (LBA) artefact14 may potentially occur since tunicatesinclude fast (Ciona ssp.) and very fast (O. dioica) evolving species(Fig. 1). A high evolutionary rate of tunicate genes was alreadynoticed in rRNA genes5 and in complete mitochondrial genomes15.Our results confirm these observations for a large number of nucleargenes. As fast evolutionary rates are also often associated withcompositional bias or with heterotachy, it is a necessary first stepto exclude the possibility that the observed grouping of tunicateswith vertebrates results from a tree reconstruction artefact.The most obvious potential artefact, an LBA, predicts that the fast-
evolving tunicates would be attracted towards the outgroup, and notby the slowly evolving vertebrates. This would produce a topologycompatible with the classical hypothesis of chordate evolution wherethe slow-evolving cephalochordates and vertebrates group together.This prediction is perfectly congruent with the lower support forOlfactores observed with MP (90%), a method known to be moresensitive to LBA than probabilistic methods14. Indeed, whenO. dioica
was used as the single representative of tunicates, MP unambiguouslysupported (BP ! 100) an aberrant position for this group whichemerged before cnidarians, disrupting the monophyly of bilaterians(Supplementary Fig. S1). By contrast, the less sensitive ML methodrecovered Olfactores, albeit with decreased bootstrap support(BP ! 84) (Supplementary Fig. S2). Therefore, despite its extremeevolutionary rate, O. dioica retained enough phylogenetic signal forits position to be recovered with ML. This demonstrates that LBAis not responsible for the inferred grouping of tunicates and verte-brates, and represents a strong argument in favour of the authenticityof Olfactores. In addition, neither compositional bias nor hetero-tachy significantly influenced phylogenomic inference with our dataset (see Supplementary Information). In fact, the compositionaleffect would act against Olfactores, because vertebrates and theamphioxus share similar amino acid compositions (SupplementaryFig. S3). In conclusion, the strongly supported monophyly ofOlfactores cannot be explained by any kind of identifiable systematicbias (LBA, compositional bias, and heterotachy) and thereforeconstitutes the best current hypothesis for chordate phylogeny.The monophyly of deuterostomes remained moderately sup-
ported in our phylogenomic analyses (Fig. 1). Also, the monophylyof chordates is not found in the ML tree, but is the only alternativenot significantly rejected by likelihood-based statistical tests(Table 1). A unique origin of chordates and their distinctive featuressuch as notochord and hollow nerve cord cannot be excluded. Ourresults nevertheless favoured the intriguing possibility of a sister-group relationship between cephalochordates and echinoderms thatseems robust to analyses aimed at avoiding compositional bias and
Figure 1 | Phylogenetic analyses of genomic data strongly support thegrouping of tunicates and vertebrates into Olfactores. ML tree obtainedfrom the analysis of 33,800 aligned amino acid positions under a WAGsubstitution matrix plus a four-category gamma rate correction (a ! 0.5)using two independent reconstruction algorithms (see SupplementaryInformation). Weighted maximum parsimony and bayesian inference using
the same WAG " F " G4 model and WAG " F " G4 plus covarion modelalso retrieved the same topology (see Supplementary Information).Bootstrap proportions obtained after 100ML (red) and 1,000 MP replicates(blue), as well as bayesian posterior probabilities (black) are shown forselected branches. A star indicates that all three values are maximal (100%,100% and 1.0). Scale bar indicates number of changes per site.
LETTERS NATURE|Vol 439|23 February 2006
966
Thanks to Gavin Naylor for the slide
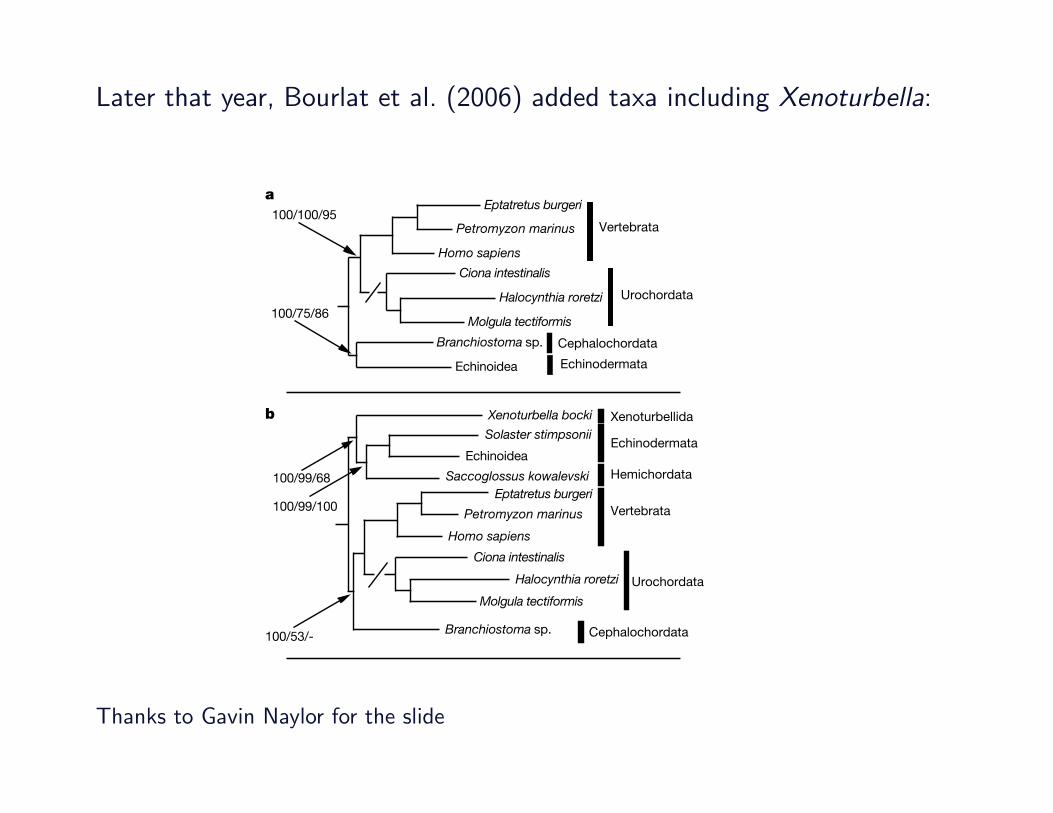
Later that year, Bourlat et al. (2006) added taxa including Xenoturbella:
the sampling of the echinoderm clade with data from the sun starSolaster stimpsoni (5,614 ESTs). We searched these ESTs for ortho-logues of the 146 genes used by Philippe et al.17. We then supplemen-ted these with 35 genes having a representative orthologue in ourXenoturbella ESTs, as this taxon had the fewestmatches to the data setfrom ref. 17.We culled 11 alignments where orthology was uncertain,leaving 170 concatenated, aligned orthologous genes (35,588 reliablyaligned amino acids). Saccoglossus had data from 153 of 170 genesand 29,777 amino acids, Xenoturbella had 63 genes and 8,370 aminoacids and Solaster had 67 genes and 11,033 amino acids.
We compared results from our extended data set of 170 genes tothose of the 146-gene data set of Delsuc et al.2 by performing phylo-genetic analyses without the new hemichordate,Xenoturbella and sunstar sequences. Phylogenetic analyses using maximum-likelihoodand bayesian approaches recapitulated previous results (Fig. 1a).All nodes received maximum support from bayesian analysis (baye-sian posterior probability (BPP) 5 100%). Maximum-likelihoodnon-parametric bootstrap support (MLBP) was lower for somenodes (see Supplementary Information). The overall topology of thistree conforms to the ‘new animal phylogeny’18 with monophyleticprotostomes, deuterostomes, lophotrochozoans (represented hereby molluscs and an annelid), and ecdysozoans (represented hereby arthropods and a nematode; see Supplementary Information).Within the deuterostomes, we find support for monophyletic groupsof vertebrates, cyclostomes (hagfish and lamprey) and Olfactores(vertebrates plus urochordates). Importantly, we recovered the rela-tionship linking cephalochordates not to other chordates but toechinoderms: BPP 5 100%, Treefinder19 LRSH support (TF) 575% and MLBP 5 81%. This relationship was unexpected and wewanted to test the possibility that it was caused by systematic error.Two powerful ways of addressing systematic errors are to includeadditional taxa, thereby enabling better detection of misleading mul-tiple substitutions, and to remove problematic taxa (for example,long branches); we have used both approaches20.
By adding sequences from three taxa that are likely to be related tothe echinoids (asteroid, hemichordate and Xenoturbella) we couldtest the possibility that the cephalochordate/echinoderm relationshipmight result from a systematic error such as long-branch attraction21.The effect of including these three taxa in our analysis is clear; sup-port for a cephalochordate plus echinoderm clade disappears andsupport for the canonical monophyletic chordates increases(Fig. 1b). The monophyletic chordate clade in our new analysis hasBPP 5 100% and TF5 53%, but no MLBP support. This change intopology on the addition of taxa that divide the echinoid branch isconsistent with the idea that the attraction between the cephalochor-date and the echinoid results from a systematic error; the addition ofnon-problematic taxa tends to improve the reliability of a phylo-geny21. We compared the support for the optimal topology to thatfor the alternative topology, which groups the cephalochordates withAmbulacraria and Xenoturbella using the Shimodaira–Hasegawa(SH) tests and Bayes factors. The SH test did not reject the alternativetopology as significantly worse than the optimal topology (SHP5 0.476), but the alternative topology was rejected as significantlyworse using the Bayes factors test (2loge (B10)5 24), although thistest might not be conservative. The inclusion of data from the othermajor taxon of cephalochordates—the Epigonichthyidae—willdivide the cephalochordate branch and, we predict, will furtherincrease support for the monophyletic chordates.
A parallel possibility is that rapidly evolving urochordates, alongwith their sister group the vertebrates, have been attracted towardsthe base of the tree by long-branch effects22. Two lines of evidencesupport this possibility. First, in the analyses described above, weused diploblast metazoans (ctenophore and cnidarian) as close out-groups; the inclusion of more distant outgroups (yeast and choano-flagellates), as in ref. 2, results in a more basal position for Olfactoreswith the corollary that cephalochordates group with echinoderms.This result is observed even when the asteroid, hemichordate
Solaster stimpsonii
Echinoidea
Saccoglossus kowalevski
Xenoturbella bocki
Ciona intestinalis
Halocynthia roretzi
Molgula tectiformis
Eptatretus burgeri
Petromyzon marinus
Homo sapiens
Branchiostoma sp.
Protopterus dolloi Lepidosiren paradoxaRaja porosa
Danio rerioChelonia mydas
Homo sapiens Boa constrictor
Myxine glutinosa Eptatretus burgeri
Lampetra fluviatilisPetromyzon marinus
Branchiostoma belcheriEpigonichthys lucayanus
Xenoturbella bockiAstropecten polyacanthusAsterina pectiniferaLuidia quinaliaAsterias amurensisPisaster ochraceusStrongylocentrotus purpuratusParacentrotus lividusArbacia lixula
Cucumaria miniataFlorometra serratissima
Balanoglossus carnosusSaccoglossus kowalevskii
Ciona intestinalis Ciona savignyi
Doliolum nationalis Halocynthia roretzi
Ecdysozoa + Lophotrochozoa
Vertebrata
Cephalochordata
Xenoturbellida
Echinodermata
Hemichordata
Urochordata
Vertebrata
Cephalochordata
Xenoturbellida
Echinodermata
Hemichordata
Urochordata
100/83/82
100/68/70
100/94/91
100/99/68
100/75/86
100/53/-
Ciona intestinalis
Halocynthia roretzi
Molgula tectiformis
Eptatretus burgeri
Petromyzon marinus
Homo sapiens
Branchiostoma sp.
Echinoidea
Vertebrata
Cephalochordata
Urochordata
Echinodermata
100/100/95
100/99/100
c
a
b
Figure 1 | Phylogenetic analyses of 170 nuclear proteins and 13mitochondrial proteins support a monophyletic chordate clade and anindependent deuterostome phylum of Xenoturbellida. a, Bayesian analysisof nuclear data before the addition of the new asteroid, hemichordate andxenoturbellid data. The cephalochordate groups with the echinoderm,implying that the chordates are paraphyletic. b, Bayesian analysis of nucleardata after the addition of asteroid, hemichordate and xenoturbellid data. Thenew sequences join the branch to the echinoderm, and the cephalochordatesnow join the chordate branch. This indicates that the previous result is dueto systematic error. Xenoturbella is the sister group of the Ambulacraria(echinoderms plus hemichordates). c, Bayesian analysis of mitochondrialdata with the amino acids M, I, N and K recoded as missing data places thecephalochordates with vertebrates; Xenoturbella is the sister group ofAmbulacraria. Support for critical nodes is shown in the order BPP (%), TF(%) and MLBP (%). Long branches leading to the urochordates have beenshortened as indicated by a break (/) in the branch (see SupplementaryInformation for unaltered tree and scale).
LETTERS NATURE |Vol 444 |2 November 2006
86Nature Publishing Group ©2006
Thanks to Gavin Naylor for the slide

• Are current models sufficient?
Not for difficult “deep phylogeny” questions.
• How should we decide between alternative models?
“Black-box” model choice methods (LRT, AIC, BIC, Bayes Factors,. . .) are very helpful at assessing the fit.
However, they may not do a good job of identifying the model thatis the best estimator of the parameters that we care about (e.g. thetree).
This particularly a concern when none of the models considered isclose to being “true.”
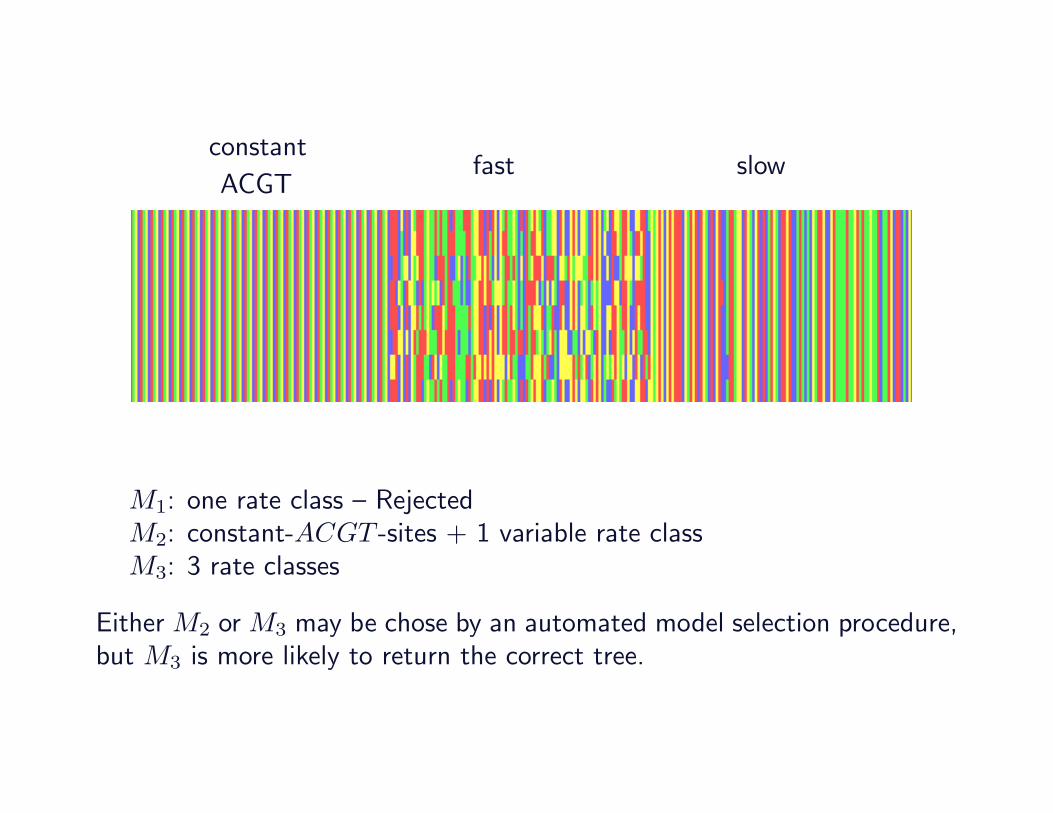
constantfast slow
ACGT
M1: one rate class – RejectedM2: constant-ACGT -sites + 1 variable rate classM3: 3 rate classes
Either M2 or M3 may be chose by an automated model selection procedure,but M3 is more likely to return the correct tree.
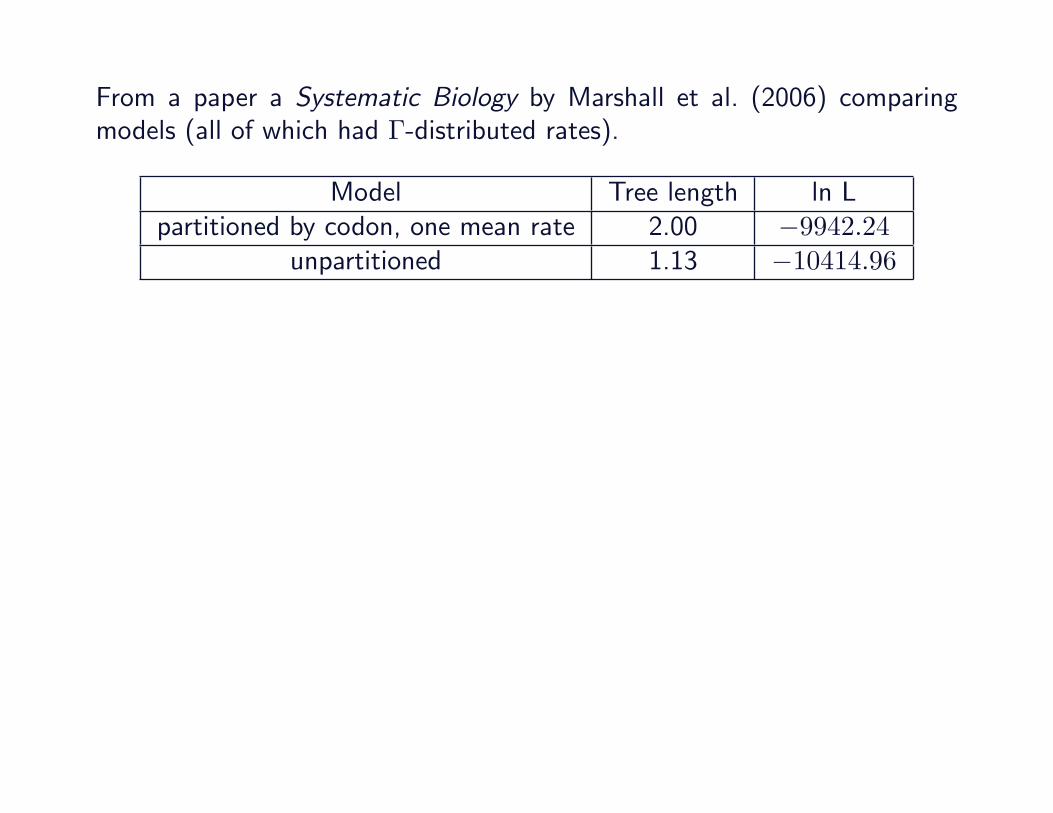
From a paper a Systematic Biology by Marshall et al. (2006) comparingmodels (all of which had Γ-distributed rates).
Model Tree length ln Lpartitioned by codon, one mean rate 2.00 −9942.24
unpartitioned 1.13 −10414.96
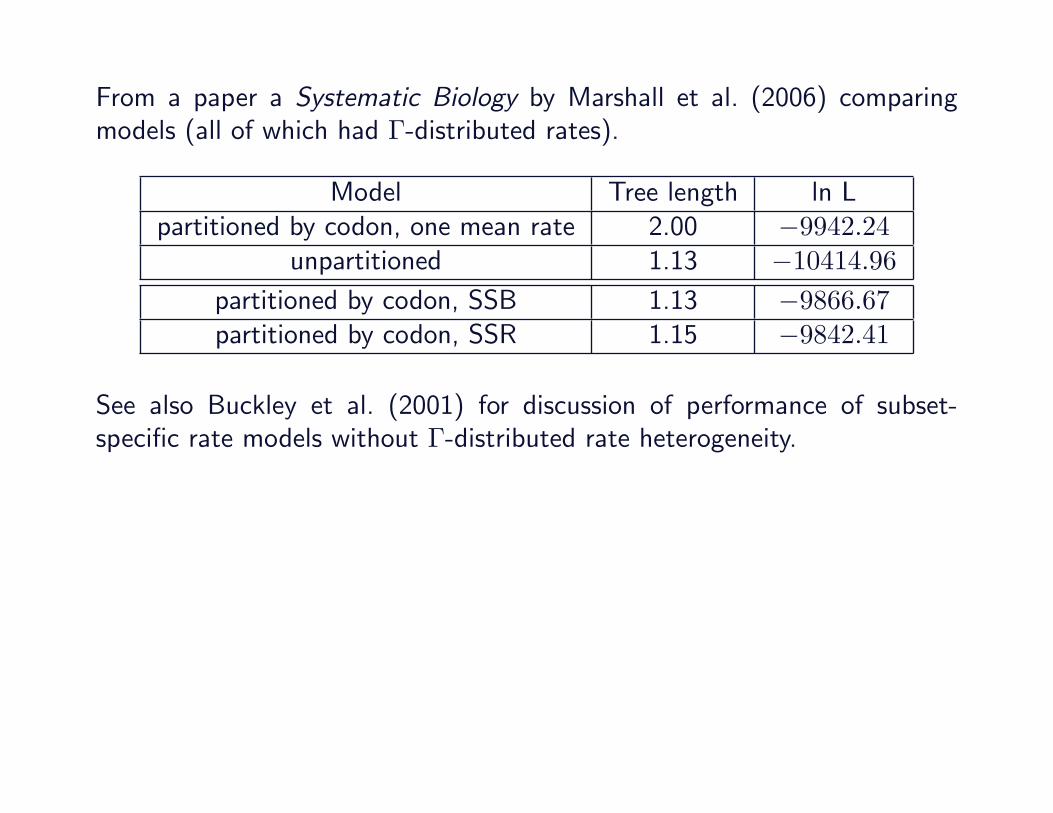
From a paper a Systematic Biology by Marshall et al. (2006) comparingmodels (all of which had Γ-distributed rates).
Model Tree length ln Lpartitioned by codon, one mean rate 2.00 −9942.24
unpartitioned 1.13 −10414.96partitioned by codon, SSB 1.13 −9866.67partitioned by codon, SSR 1.15 −9842.41
See also Buckley et al. (2001) for discussion of performance of subset-specific rate models without Γ-distributed rate heterogeneity.

How should we decide between alternative models?
• Standard model selection tools are certainly helpful,
• Performance-based model selection (e.g. Minin et al., 2003),
• Performance of a model on previous real data analyses,
• Performance of inference methods on data derived from computersimulations.

Using computer simulations to evaluate models and methods:
• flexible, fast, and transparent
but. . .
• simulations often generate data that is too “clean” (but see work ofJunhyong Kim and his collaborators).
• simulated data may bear little resemblance to the real world.

Evaluating performance using computer simulation
1. Choose a simulation model
2. Choose a tree shape
3. Simulate many data sets
4. Infer trees using a variety of methods
5. Compare inferred tree to true (model) tree.
In the following slides we’ll deal with a very complex, parameter-richsimulation model – Halpern and Bruno (1998) model of site-specific residuefrequencies.

Halpern and Bruno (1998) model of coding sequenceevolution
• All nucleotides share a set of parameters for a mutational model,
• Each amino acid residue has a set of equilibrium frequencies

Halpern and Bruno (1998) model properties
1. Extreme heterogeneity of process
2. Sites (within the same codon) are not independent
3. Among-site rate heterogeneity, but not following a simple distribution(such as the Γ-distribution).

I wrote software to:
1. find MLEs of parameters of the Halpern and Bruno (1998) model, and
2. simulate under the Halpern-Bruno model (on a user-defined tree).
Estimated parameters on a dataset of 1610 mammalian cytochrome-bsequences (376 amino acid residues, ≈ 7, 150 substitution parameters)
http://nladr-cvs.sdsc.edu/svn/CIPRES/cipresdev/trunk/python-example/org.cipres.bull
username: guest
password: guest
ORNITHORHYNCHUS ANATINUSMACROPUS ROBUSTUSMACROPUS GIGANTEUSMACROPUS RUFUSDROMICIOPS GLIROIDESPHASCOLARCTOS CINEREUSVOMBATUS URSINUSPSEUDOCHIROPS CUPREUSTRICHOSURUS VULPECULASMINTHOPSIS YOUNGSONISMINTHOPSIS OOLDEASMINTHOPSIS VIRGINIAE1SMINTHOPSIS VIRGINIAE2SMINTHOPSIS DOUGLASISMINTHOPSIS BINDISMINTHOPSIS MACROURASMINTHOPSIS CRASSICAUDATASMINTHOPSIS PSAMMOPHILASMINTHOPSIS LEUCOPUS1SMINTHOPSIS LEUCOPUS2SMINTHOPSIS GILBERTISMINTHOPSIS MURINASMINTHOPSIS DOLICHURASMINTHOPSIS ARCHERISMINTHOPSIS HIRTIPESSMINTHOPSIS LONGICAUDATANINGAUI YVONNAESMINTHOPSIS GRISEOVENTERSMINTHOPSIS AITKENISMINTHOPSIS GRANULIPESPLANIGALE MACULATA1PLANIGALE MACULATA2PLANIGALE SPECIESPLANIGALE INGRAMIPLANIGALE GILESIPLANIGALE TENUIROSTRISNINGAUI TIMEALEYININGAUI RIDEIANTECHINOMYS LANIGERMYRMECOBIUS FASCIATUSMUREXIA LONGICAUDATAANTECHINUS NASOMUREXIA ROTHSCHILDIANTECHINUS MELANURUSANTECHINUS SPECIESANTECHINUS LEOANTECHINUS BELLUSANTECHINUS FLAVIPESANTECHINUS STUARTII1ANTECHINUS STUARTII2ANTECHINUS GODMANIANTECHINUS MINIMUSANTECHINUS SWAINSONII1ANTECHINUS SWAINSONII2ANTECHINUS HABBEMAPHASCOGALE CALURAPHASCOGALE TAPOATAFADASYURUS VIVERRINUSDASYURUS ALBOPUNCTATUSDASYURUS SPARTACUSDASYURUS GEOFFROIIDASYURUS MACULATUSDASYURUS HALLUCATUSSARCOPHILUS HARRISIINEOPHASCOGALE LORENTZIIPHASCOLOSOREX DORSALISPARANTECHINUS BILARNIPARANTECHINUS APICALISDASYKALUTA ROSAMONDAEDASYUROIDES BYRNEIPSEUDANTECHINUS WOOLLEYAEPSEUDANTECHINUS MACDONNELLENSISPSEUDANTECHINUS NINGBINGMYOICTIS MELASMYOICTIS WALLACEIDASYCERCUS CRISTICAUDATHYLACINUS CYNOCEPHALUSNOTORYCTES TYPHLOPSISOODON MACROURUSPERAMELES NASUTAECHYMIPERA KALUBUCAENOLESTES FULIGINOSUSLESTOROS INCADIDELPHIS VIRGINIANADIDELPHIS MARSUPIALISDIDELPHIS ALBIVENTRISPHILANDER OPOSSUM1PHILANDER OPOSSUM2PHILANDER MCILHENNYIMETACHIRUS NUDICAUDATUS1METACHIRUS NUDICAUDATUS2MONODELPHIS DOMESTICAGLIRONIA VENUSTAMONODELPHIS ADUSTA1MONODELPHIS ADUSTA2MICOUREUS REGINAMICOUREUS DEMERARAE1MICOUREUS DEMERARAE2MARMOSA RUBRACALUROMYS LANATUS1CALUROMYS LANATUS2MARMOSOPS IMPAVIDUS1MARMOSOPS IMPAVIDUS2SIGMODON HISPIDUS1SIGMODON HISPIDUS2IRENOMYS TARSALISCHELEMYS MACRONYXGEOXUS VALDIVIANUSAKODON LONGIPILISAKODON JELSKIIAKODON OLIVACEUS1AKODON OLIVACEUS6AKODON OLIVACEUS8AKODON OLIVACEUS7AKODON OLIVACEUS2AKODON OLIVACEUS3AKODON OLIVACEUS5AKODON OLIVACEUS15AKODON OLIVACEUS20AKODON OLIVACEUS21AKODON OLIVACEUS23AKODON OLIVACEUS19AKODON OLIVACEUS22AKODON OLIVACEUS24AKODON OLIVACEUS4AKODON OLIVACEUS9AKODON OLIVACEUS12AKODON OLIVACEUS17AKODON OLIVACEUS18AKODON OLIVACEUS10AKODON OLIVACEUS11AKODON OLIVACEUS14AKODON OLIVACEUS13AKODON OLIVACEUS16ABROTHRIX ANDINUSANDINOMYS EDAXBOLOMYS URICHIBOLOMYS LASIURUSBOLOMYS AMOENUSAKODON TOBAAKODON TORQUESAKODON BOLIVIENSISAKODON NIGRITAOXYMYCTERUS PARAMENSISJUSCELINOMYS SPECIESLENOXUS APICALISOXYMYCTERUS IHERINGIBLARINOMYS BREVICEPSKUNSIA TOMENTOSUSSCAPTEROMYS TUMIDUSRHIPIDOMYS LEUCODACTYLUSRHIPIDOMYS NITELARHIPIDOMYS MACCONNELLIRHIPIDOMYS WETZELITHOMASOMYS AUREUSTHOMASOMYS NOTATUSTHOMASOMYS ISCHYURUSTHOMASOMYS SPECIESTHOMASOMYS OREASTHOMASOMYS GRACILISTHOMASOMYS DAPHNECHILOMYS INSTANSOECOMYS BICOLORORYZOMYS MEGACEPHALUSMICRORYZOMYS MINUTUSSCOLOMYS JURUAENSEREITHRODON AURITUSWILFREDOMYS SPECIESWILFREDOMYS PICTIPESDELOMYS SUBLINEATUSDELOMYS DORSALISAULISCOMYS PICTUSAULISCOMYS MICROPUSSALINOMYS DELICATUSANDALGALOMYS ROIGIANDALGALOMYS PEARSONITAPECOMYS PRIMUS1TAPECOMYS PRIMUS2PHYLLOTIS XANTHOPYGUSELIGMODONTIA TYPUSELIGMODONTIA MORGANIELIGMODONTIA PUERULUSCALOMYS LEPIDUS1CALOMYS LEPIDUS2CALOMYS CALLOSUSGRAOMYS DOMORUMGRAOMYS GRISEOFLAVUSREITHRODONTOMYS SUMICHRASTI1REITHRODONTOMYS MEXICANUSREITHRODONTOMYS SUMICHRASTI2REITHRODONTOMYS SUMICHRASTI22REITHRODONTOMYS SUMICHRASTI6REITHRODONTOMYS SUMICHRASTI21REITHRODONTOMYS SUMICHRASTI25REITHRODONTOMYS SUMICHRASTI8REITHRODONTOMYS SUMICHRASTI20REITHRODONTOMYS SUMICHRASTI10REITHRODONTOMYS SUMICHRASTI3REITHRODONTOMYS SUMICHRASTI16REITHRODONTOMYS SUMICHRASTI11REITHRODONTOMYS SUMICHRASTI18REITHRODONTOMYS SUMICHRASTI17REITHRODONTOMYS SUMICHRASTI23REITHRODONTOMYS SUMICHRASTI4REITHRODONTOMYS SUMICHRASTI9REITHRODONTOMYS SUMICHRASTI13REITHRODONTOMYS SUMICHRASTI19REITHRODONTOMYS SUMICHRASTI24REITHRODONTOMYS SUMICHRASTI31REITHRODONTOMYS SUMICHRASTI26REITHRODONTOMYS SUMICHRASTI28REITHRODONTOMYS SUMICHRASTI5REITHRODONTOMYS SUMICHRASTI7REITHRODONTOMYS SUMICHRASTI14REITHRODONTOMYS SUMICHRASTI30REITHRODONTOMYS SUMICHRASTI27REITHRODONTOMYS SUMICHRASTI15REITHRODONTOMYS SUMICHRASTI12REITHRODONTOMYS MEGALOTISPEROMYSCUS MANICULATUSPEROMYSCUS TRUEISCOTINOMYS XERAMPELINUSSCOTINOMYS TEGUINANEOTOMA ALBIGULACRICETULUS GRISEUSPHODOPUS CAMPBELLIPHODOPUS SUNGORUSMESOCRICETUS AURATUSMICROTUS ARVALIS1MICROTUS ARVALIS2MICROTUS ARVALIS9MICROTUS ARVALIS3MICROTUS ARVALIS8MICROTUS ARVALIS6MICROTUS ARVALIS7MICROTUS ARVALIS4MICROTUS ARVALIS5MICROTUS ROSSIAEMERIDIONALIS1MICROTUS ROSSIAEMERIDIONALIS2MICROTUS ROSSIAEMERIDIONALIS5MICROTUS ROSSIAEMERIDIONALIS6MICROTUS ROSSIAEMERIDIONALIS3MICROTUS ROSSIAEMERIDIONALIS4MICROTUS OECONOMUSMICROTUS MONTEBELLIMICROTUS AGRESTIS1MICROTUS AGRESTIS2MICROTUS MONTANUSMICROTUS PENNSYLVANICUSMICROTUS TOWNSENDIIMICROTUS OREGONIMICROTUS LONGICAUDUS1MICROTUS LONGICAUDUS2MICROTUS LONGICAUDUS3MICROTUS LONGICAUDUS14MICROTUS LONGICAUDUS4MICROTUS LONGICAUDUS6MICROTUS LONGICAUDUS23MICROTUS LONGICAUDUS7MICROTUS LONGICAUDUS10MICROTUS LONGICAUDUS8MICROTUS LONGICAUDUS9MICROTUS LONGICAUDUS11MICROTUS LONGICAUDUS24MICROTUS LONGICAUDUS12MICROTUS LONGICAUDUS20MICROTUS LONGICAUDUS26MICROTUS LONGICAUDUS13MICROTUS LONGICAUDUS17MICROTUS LONGICAUDUS15MICROTUS LONGICAUDUS21MICROTUS LONGICAUDUS25MICROTUS LONGICAUDUS5MICROTUS LONGICAUDUS16MICROTUS LONGICAUDUS22MICROTUS LONGICAUDUS19MICROTUS LONGICAUDUS27MICROTUS LONGICAUDUS38MICROTUS LONGICAUDUS28MICROTUS LONGICAUDUS29MICROTUS LONGICAUDUS35MICROTUS LONGICAUDUS30MICROTUS LONGICAUDUS33MICROTUS LONGICAUDUS37MICROTUS LONGICAUDUS34MICROTUS LONGICAUDUS40MICROTUS LONGICAUDUS31MICROTUS LONGICAUDUS32MICROTUS LONGICAUDUS39MICROTUS LONGICAUDUS36MICROTUS LONGICAUDUS41MICROTUS LONGICAUDUS43MICROTUS LONGICAUDUS46MICROTUS LONGICAUDUS48MICROTUS LONGICAUDUS49MICROTUS LONGICAUDUS50MICROTUS LONGICAUDUS44MICROTUS LONGICAUDUS45MICROTUS LONGICAUDUS18MICROTUS LONGICAUDUS47MICROTUS LONGICAUDUS42MICROTUS LONGICAUDUS52MICROTUS LONGICAUDUS53MICROTUS LONGICAUDUS58MICROTUS LONGICAUDUS54MICROTUS LONGICAUDUS56MICROTUS LONGICAUDUS55MICROTUS LONGICAUDUS59MICROTUS LONGICAUDUS51MICROTUS LONGICAUDUS65MICROTUS LONGICAUDUS57MICROTUS LONGICAUDUS64MICROTUS LONGICAUDUS60MICROTUS LONGICAUDUS61MICROTUS LONGICAUDUS62MICROTUS LONGICAUDUS63MICROTUS RICHARDSONIMICROTUS PINETORUMMICROTUS XANTHOGNATHUSMICROTUS MIURUSMICROTUS OCHROGASTERELLOBIUS TANCREIELLOBIUS FUSCOCAPILLUSCLETHRIONOMYS GLAREOLUS1CLETHRIONOMYS GLAREOLUS2CLETHRIONOMYS RUTILUS1CLETHRIONOMYS RUTILUS2ALTICOLA MACROTISCLETHRIONOMYS REXCLETHRIONOMYS RUFOCANUS1CLETHRIONOMYS RUFOCANUS10CLETHRIONOMYS RUFOCANUS3CLETHRIONOMYS RUFOCANUS4CLETHRIONOMYS RUFOCANUS2CLETHRIONOMYS RUFOCANUS5CLETHRIONOMYS RUFOCANUS6CLETHRIONOMYS RUFOCANUS7CLETHRIONOMYS RUFOCANUS8CLETHRIONOMYS RUFOCANUS9CLETHRIONOMYS RUFOCANUS15CLETHRIONOMYS RUFOCANUS16CLETHRIONOMYS RUFOCANUS11CLETHRIONOMYS RUFOCANUS12CLETHRIONOMYS RUFOCANUS13CLETHRIONOMYS RUFOCANUS14CLETHRIONOMYS RUFOCANUS17CLETHRIONOMYS RUFOCANUS18CLETHRIONOMYS RUFOCANUS25CLETHRIONOMYS RUFOCANUS26CLETHRIONOMYS RUFOCANUS20CLETHRIONOMYS RUFOCANUS21CLETHRIONOMYS RUFOCANUS23CLETHRIONOMYS RUFOCANUS24CLETHRIONOMYS RUFOCANUS19CLETHRIONOMYS RUFOCANUS22CLETHRIONOMYS RUFOCANUS28CLETHRIONOMYS RUFOCANUS27ARVICOLA TERRESTRIS1ARVICOLA TERRESTRIS2ONDATRA ZIBETHICUSDICROSTONYX TORQUATUSDICROSTONYX GROENLANDICUSPHENACOMYS INTERMEDIUSLEMMUS TRIMUCRONATUSMYOPUS SCHISTICOLORSYNAPTOMYS BOREALISMYSTROMYS ALBICAUDATUS1MYSTROMYS ALBICAUDATUS2PETROMYSCUS COLLINUS1PETROMYSCUS COLLINUS2CALOMYSCUS BAILWARDITOKUDAIA OSIMENSIS1TOKUDAIA OSIMENSIS2MICROMYS MINUTUS1MICROMYS MINUTUS2DIPLOTHRIX LEGATANESOKIA INDICARATTUS NORVEGICUS1RATTUS ARGENTIVENTERRATTUS NORVEGICUS2RATTUS NORVEGICUS3RATTUS NORVEGICUS4APODEMUS SYLVATICUS1APODEMUS SYLVATICUS3APODEMUS MICROPSAPODEMUS ALPICOLA1APODEMUS ALPICOLA2APODEMUS FLAVICOLLIS1APODEMUS FLAVICOLLIS2APODEMUS SYLVATICUS2APODEMUS MYSTACINUSAPODEMUS SEMOTUSAPODEMUS PENINSULAE2APODEMUS SPECIOSUSAPODEMUS AGRARIUSAPODEMUS PENINSULAE1APODEMUS ARGENTEUSLEMNISCOMYS STRIATUS1LEMNISCOMYS STRIATUS2LEMNISCOMYS ROSALIALEMNISCOMYS MACCULUSLEMNISCOMYS BELLIERILEMNISCOMYS ZEBRAARVICANTHIS SPECIES1ARVICANTHIS SPECIES5ARVICANTHIS SPECIES6ARVICANTHIS SPECIES7ARVICANTHIS SPECIES8ARVICANTHIS SPECIES9ARVICANTHIS SPECIES10ARVICANTHIS SPECIES2ARVICANTHIS SPECIES3ARVICANTHIS SPECIES4ARVICANTHIS SOMALICUS1ARVICANTHIS SOMALICUS2ARVICANTHIS NILOTICUS1ARVICANTHIS NILOTICUS2ARVICANTHIS NILOTICUS3ARVICANTHIS NILOTICUS4ARVICANTHIS DEMBEENSISARVICANTHIS ABYSSINICUS1ARVICANTHIS ABYSSINICUS2PELOMYS CAMPANAEMYLOMYS DYBOWSKIIDASYMYS INCOMTUSDASYMYS RUFULUSAETHOMYS SPECIESRHABDOMYS PUMILIODESMOMYS HARRINGTONIHYBOMYS UNIVITTATUSAETHOMYS NAMAQUENSISMILLARDIA MELTADAGRAMMOMYS SPECIESMASTOMYS HUBERTIPAROTOMYS LITTLEDALEIOTOMYS SLOGGETTIPAROTOMYS BRANTSIIOTOMYS IRRORATUSMUS SPRETUS1MUS SPRETUS2MUS MUSCULUS1MUS MUSCULUS2MUS MUSCULUS3MUS POSCHIAVINUSMUS SPICILEGUSMUS CAROLIMUS MUSCULOIDESLOPHUROMYS SIKAPUSIACOMYS SUBSPINOSUSACOMYS SPECIES1ACOMYS SPECIES4ACOMYS SPECIES6ACOMYS SPECIES13ACOMYS SPECIES12ACOMYS SPECIES11ACOMYS SPECIES5ACOMYS SPECIES8ACOMYS SPECIES7ACOMYS SPECIES10ACOMYS SPECIES9ACOMYS JOHANNISACOMYS DIMIDIATUS1ACOMYS DIMIDIATUS2ACOMYS DIMIDIATUS3ACOMYS DIMIDIATUS6ACOMYS DIMIDIATUS4ACOMYS DIMIDIATUS10ACOMYS DIMIDIATUS5ACOMYS DIMIDIATUS7ACOMYS DIMIDIATUS9ACOMYS DIMIDIATUS8ACOMYS CAHIRINUS1ACOMYS CAHIRINUS4ACOMYS CAHIRINUS5ACOMYS CAHIRINUS7ACOMYS CAHIRINUS2ACOMYS CAHIRINUS3ACOMYS CAHIRINUS6ACOMYS NESIOTESACOMYS MINOUS1ACOMYS MINOUS2ACOMYS MINOUS3ACOMYS CILICICUSACOMYS IGNITUS1ACOMYS IGNITUS2ACOMYS RUSSATUS1ACOMYS RUSSATUS2ACOMYS SPECIES2ACOMYS SPECIES3ACOMYS WILSONIACOMYS SPINOSISSIMUS1ACOMYS SPINOSISSIMUS3ACOMYS SPINOSISSIMUS2ACOMYS SPINOSISSIMUS4ACOMYS SPINOSISSIMUS5MERIONES UNGUICULATUS1MERIONES UNGUICULATUS2GERBILLUS NIGERIAETATERA KEMPINESOMYS AUDEBERTI1NESOMYS RUFUS1NESOMYS RUFUS7NESOMYS RUFUS2NESOMYS RUFUS3NESOMYS RUFUS6NESOMYS RUFUS5NESOMYS RUFUS4NESOMYS AUDEBERTI2BRACHYUROMYS BETSILEOENSIS1BRACHYUROMYS BETSILEOENSIS2BRACHYUROMYS RAMIROHITRAHYPOGEOMYS ANTIMENAMACROTARSOMYS BASTARDIMONTICOLOMYS KOOPMANI1MONTICOLOMYS KOOPMANI4MONTICOLOMYS KOOPMANI2MONTICOLOMYS KOOPMANI3CRICETOMYS GAMBIANUSCRICETOMYS EMINI1CRICETOMYS EMINI2CRICETOMYS EMINI3BEAMYS HINDEI1BEAMYS HINDEI2STEATOMYS PARVUSSTEATOMYS SPECIESVOALAVO GYMNOCAUDUSELIURUS GRANDIDIERI1ELIURUS GRANDIDIERI2ELIURUS ELLERMANIELIURUS WEBBI2ELIURUS WEBBI1ELIURUS TANALA1ELIURUS TANALA2ELIURUS TANALA3ELIURUS TANALA5ELIURUS TANALA4ELIURUS TANALA8ELIURUS TANALA6ELIURUS TANALA7ELIURUS MYOXINUS1ELIURUS MYOXINUS6ELIURUS MYOXINUS7ELIURUS MYOXINUS2ELIURUS MYOXINUS3ELIURUS MYOXINUS4ELIURUS MYOXINUS5ELIURUS MINOR1ELIURUS MINOR6ELIURUS MINOR3ELIURUS MINOR2ELIURUS MINOR4ELIURUS MINOR5ELIURUS MINOR8ELIURUS MINOR9ELIURUS MINOR7ELIURUS MAJORI1ELIURUS MAJORI2ELIURUS MAJORI3ELIURUS MAJORI4ELIURUS MAJORI11ELIURUS MAJORI12ELIURUS MAJORI7ELIURUS MAJORI10ELIURUS MAJORI9ELIURUS MAJORI5ELIURUS MAJORI6ELIURUS MAJORI8ELIURUS WEBBI3ELIURUS WEBBI7ELIURUS WEBBI8ELIURUS WEBBI9ELIURUS WEBBI6ELIURUS WEBBI5ELIURUS WEBBI4GYMNUROMYS ROBERTI1GYMNUROMYS ROBERTI2GYMNUROMYS ROBERTI3GYMNUROMYS ROBERTI4BRACHYTARSOMYS ALBICAUDAGEOMYS PINETISGEOMYS BURSARIUS1GEOMYS BURSARIUS4GEOMYS BURSARIUS5GEOMYS BURSARIUS8GEOMYS BURSARIUS7GEOMYS BURSARIUS9GEOMYS BURSARIUS2GEOMYS BURSARIUS3GEOMYS BURSARIUS6GEOMYS BURSARIUS10GEOMYS BREVICEPS1GEOMYS BREVICEPS2GEOMYS BREVICEPS3CRATOGEOMYS CASTANOPS1CRATOGEOMYS CASTANOPS2CRATOGEOMYS GOLDMANI1CRATOGEOMYS GOLDMANI2CRATOGEOMYS MERRIAMICRATOGEOMYS TYLORHINUSCRATOGEOMYS GYMNURUSCRATOGEOMYS FUMOSUSCRATOGEOMYS BULLERIORTHOGEOMYS HETERODUSTHOMOMYS UMBRINUS1THOMOMYS UMBRINUS4THOMOMYS UMBRINUS5THOMOMYS UMBRINUS6THOMOMYS UMBRINUS7THOMOMYS UMBRINUS8THOMOMYS UMBRINUS2THOMOMYS UMBRINUS3THOMOMYS BOTTAE29THOMOMYS TOWNSENDII1THOMOMYS BOTTAE14THOMOMYS TOWNSENDII2THOMOMYS BOTTAE16THOMOMYS BOTTAE19THOMOMYS BOTTAE24THOMOMYS TOWNSENDII3THOMOMYS BOTTAE15THOMOMYS BOTTAE17THOMOMYS BOTTAE18THOMOMYS BOTTAE27THOMOMYS BOTTAE28THOMOMYS BOTTAE25THOMOMYS BOTTAE7THOMOMYS BOTTAE9THOMOMYS BOTTAE12THOMOMYS BOTTAE13THOMOMYS BOTTAE20THOMOMYS BOTTAE11THOMOMYS BOTTAE8THOMOMYS BOTTAE10THOMOMYS BOTTAE26THOMOMYS BOTTAE30THOMOMYS BOTTAE32THOMOMYS BOTTAE31THOMOMYS BOTTAE1THOMOMYS BOTTAE2THOMOMYS BOTTAE3THOMOMYS BOTTAE4THOMOMYS BOTTAE21THOMOMYS BOTTAE23THOMOMYS BOTTAE22THOMOMYS BOTTAE5THOMOMYS BOTTAE6THOMOMYS TALPOIDESTHOMOMYS MONTICOLA1THOMOMYS MONTICOLA2PEROGNATHUS LONGIMEMBRISPEROGNATHUS AMPLUSDIPODOMYS AGILISTACHYORYCTES SPLENDENSZAPUS TRINOTATUSCAVIA PORCELLUSMYOPROCTA PRATTICTENOMYS SOCIABILISCTENOMYS HAIGICTENOMYS COYHAIQUENSIS1CTENOMYS COYHAIQUENSIS2CTENOMYS COYHAIQUENSIS3CTENOMYS MENDOCINUSCTENOMYS FLAMARIONICTENOMYS RIONEGRENSIS1CTENOMYS RIONEGRENSIS4CTENOMYS RIONEGRENSIS5CTENOMYS RIONEGRENSIS3CTENOMYS RIONEGRENSIS2CTENOMYS TORQUATUS1CTENOMYS TORQUATUS2CTENOMYS PEARSONICTENOMYS SPECIES1CTENOMYS SPECIES5CTENOMYS GOODFELLOWI1CTENOMYS BOLIVIENSIS2CTENOMYS GOODFELLOWI2CTENOMYS BOLIVIENSIS1CTENOMYS STEINBACHI1CTENOMYS STEINBACHI2CTENOMYS SPECIES2CTENOMYS SPECIES3CTENOMYS SPECIES4CTENOMYS CONOVERICTENOMYS LEWISICTENOMYS FRATER1CTENOMYS FRATER2CTENOMYS OPIMUS1CTENOMYS OPIMUS2CTENOMYS LEUCODONSPALACOPUS CYANUSOCTODON DEGUSTYMPANOCTOMYS BARRERAEPROECHIMYS SIMONSIPROECHIMYS AMPHICHORICUSEURYZYGOMATOMYS SPINOSUSTRINOMYS SPECIES1TRINOMYS IHERINGITRINOMYS DIMIDIATUS1TRINOMYS DIMIDIATUS2TRINOMYS DIMIDIATUS3TRINOMYS DIMIDIATUS4TRINOMYS SPECIES2TRINOMYS PARATUSTRINOMYS ELIASITRINOMYS ALBISPINISNELOMYS BRASILIENSISECHIMYS CHRYSURUSECHIMYS DIDELPHOIDES1ECHIMYS DIDELPHOIDES4ECHIMYS DIDELPHOIDES2ECHIMYS DIDELPHOIDES3DACTYLOMYS BOLIVIENSISDACTYLOMYS DACTYLINUSMESOMYS SPECIES1MESOMYS SPECIES2MESOMYS HISPIDUS1MESOMYS HISPIDUS2ISOTHRIX BISTRIATUSTRINOMYS SETOSUSTHRICHOMYS APEREOIDESCOENDOU BICOLOR1COENDOU BICOLOR2GEORYCHUS CAPENSISBATHYERGUS SUILLUSBATHYERGUS JANETTACRYPTOMYS HOTTENTOTUS1CRYPTOMYS HOTTENTOTUS2CRYPTOMYS HOTTENTOTUS5CRYPTOMYS HOTTENTOTUS6CRYPTOMYS HOTTENTOTUS4CRYPTOMYS CHOMACRYPTOMYS AMATUSCRYPTOMYS DAMARENSIS1CRYPTOMYS DAMARENSIS2CRYPTOMYS DAMARENSIS3CRYPTOMYS DAMARENSIS4CRYPTOMYS DAMARENSIS5CRYPTOMYS DAMARENSIS6CRYPTOMYS DAMARENSIS7CRYPTOMYS DAMARENSIS8CRYPTOMYS DAMARENSIS9CRYPTOMYS DAMARENSIS10CRYPTOMYS DARLINGICRYPTOMYS MECHOWI1CRYPTOMYS MECHOWI2CRYPTOMYS BOCAGEIHELIOPHOBIUS ARGENTEOCINEREUSHETEROCEPHALUS GLABER1HETEROCEPHALUS GLABER2HETEROCEPHALUS GLABER4HETEROCEPHALUS GLABER3HETEROCEPHALUS GLABER5HYSTRIX AFRICAEAUSTRALISMYOXUS GLIS1MYOXUS GLIS2GRAPHIURUS MURINUSMUSCARDINUS AVELLANARIUSELIOMYS QUERCINUSDRYOMYS NITEDULAGLAUCOMYS SABRINUS1GLAUCOMYS SABRINUS2GLAUCOMYS SABRINUS5GLAUCOMYS SABRINUS6GLAUCOMYS SABRINUS7GLAUCOMYS SABRINUS3GLAUCOMYS SABRINUS4GLAUCOMYS VOLANSSPERMOPHILOPSIS LEPTODACTYLUSSCIURUS ABERTI1SCIURUS ABERTI2SCIURUS ABERTI4SCIURUS ABERTI8SCIURUS ABERTI5SCIURUS ABERTI6SCIURUS ABERTI7SCIURUS ABERTI9SCIURUS ABERTI11SCIURUS ABERTI10SCIURUS ABERTI17SCIURUS ABERTI19SCIURUS ABERTI14SCIURUS ABERTI15SCIURUS ABERTI16SCIURUS ABERTI18SCIURUS ABERTI20SCIURUS ABERTI3SCIURUS ABERTI13SCIURUS ABERTI12SCIURUS SPECIESSCIURUS NIGERMARMOTA MONAX1MARMOTA MONAX3MARMOTA MONAX2MARMOTA MONAX4MARMOTA MONAX5MARMOTA BOBAK1MARMOTA BOBAK2MARMOTA BOBAK3MARMOTA BAIBACINA1MARMOTA BAIBACINA3MARMOTA BAIBACINA2MARMOTA CAMTSCHATICA1MARMOTA CAMTSCHATICA3MARMOTA CAMTSCHATICA2MARMOTA SIBIRICAMARMOTA HIMALAYANAMARMOTA CAUDATA1MARMOTA CAUDATA2MARMOTA MENZBIERI1MARMOTA MENZBIERI2MARMOTA BROWERI1MARMOTA BROWERI2MARMOTA MARMOTA1MARMOTA MARMOTA2MARMOTA CALIGATA1MARMOTA CALIGATA2MARMOTA VANCOUVERENSIS1MARMOTA VANCOUVERENSIS2MARMOTA FLAVIVENTRIS1MARMOTA FLAVIVENTRIS2SPERMOPHILUS BRUNNEUS1SPERMOPHILUS BRUNNEUS3SPERMOPHILUS BRUNNEUS2SPERMOPHILUS BRUNNEUS4SPERMOPHILUS TOWNSENDI1SPERMOPHILUS TOWNSENDI9SPERMOPHILUS TOWNSENDI2SPERMOPHILUS TOWNSENDI6SPERMOPHILUS TOWNSENDI7SPERMOPHILUS TOWNSENDI8SPERMOPHILUS BELDINGI1SPERMOPHILUS BELDINGI2SPERMOPHILUS ARMATUS1SPERMOPHILUS ARMATUS2SPERMOPHILUS COLUMBIANUS1SPERMOPHILUS COLUMBIANUS2SPERMOPHILUS COLUMBIANUS3SPERMOPHILUS COLUMBIANUS4SPERMOPHILUS PARRYI1SPERMOPHILUS RICHARDSONI1SPERMOPHILUS RICHARDSONI2SPERMOPHILUS RICHARDSONIISPERMOPHILUS ELEGANS1SPERMOPHILUS ELEGANS2SPERMOPHILUS PARRYI2SPERMOPHILUS PARRYI3SPERMOPHILUS UNDULATUS1SPERMOPHILUS UNDULATUS2SPERMOPHILUS WASHINGTONI1SPERMOPHILUS WASHINGTONI2SPERMOPHILUS TOWNSENDI3SPERMOPHILUS TOWNSENDI4SPERMOPHILUS TOWNSENDI5SPERMOPHILUS PEROTENSISSPERMOPHILUS SPILOSOMA3SPERMOPHILUS SPILOSOMA4SPERMOPHILUS SPILOSOMA1SPERMOPHILUS SPILOSOMA2SPERMOPHILUS TERETICAUDUS1SPERMOPHILUS TERETICAUDUS2SPERMOPHILUS MOHAVENSIS1SPERMOPHILUS MOHAVENSIS2CYNOMYS GUNNISONI1CYNOMYS GUNNISONI2CYNOMYS LEUCURUS1CYNOMYS LEUCURUS2CYNOMYS PARVIDENS1CYNOMYS PARVIDENS2CYNOMYS LUDOVICIANUS1CYNOMYS LUDOVICIANUS2CYNOMYS MEXICANUS1CYNOMYS MEXICANUS2CYNOMYS MEXICANUS3SPERMOPHILUS FRANKLINII1SPERMOPHILUS FRANKLINII2SPERMOPHILUS TRIDECEMLINEATUSSPERMOPHILUS SPECIES1SPERMOPHILUS SPECIES2SPERMOPHILUS MEXICANUS1SPERMOPHILUS MEXICANUS2SPERMOPHILUS MEXICANUS3SPERMOPHILUS FULVUSSPERMOPHILUS MAJOR2SPERMOPHILUS ERYTHROGENYS3SPERMOPHILUS ERYTHROGENYS4SPERMOPHILUS CITELLUS1SPERMOPHILUS PALLIDICAUDASPERMOPHILUS ALASHANICUSSPERMOPHILUS CITELLUS2SPERMOPHILUS CITELLUS3SPERMOPHILUS ERYTHROGENYS1SPERMOPHILUS ERYTHROGENYS2SPERMOPHILUS RELICTUS1SPERMOPHILUS RELICTUS2SPERMOPHILUS CITELLUS4SPERMOPHILUS CITELLUS5SPERMOPHILUS XANTHOPRYMNUS1SPERMOPHILUS XANTHOPRYMNUS2SPERMOPHILUS DAURICUSSPERMOPHILUS SUSLICUS1SPERMOPHILUS SUSLICUS2SPERMOPHILUS SUSLICUS3SPERMOPHILUS PYGMAEUS1SPERMOPHILUS PYGMAEUS2SPERMOPHILUS MAJOR1SPERMOPHILUS MUSICUSSPERMOPHILUS LATERALIS1SPERMOPHILUS LATERALIS2SPERMOPHILUS MADRENSIS1SPERMOPHILUS MADRENSIS2SPERMOPHILUS SATURATUS1SPERMOPHILUS SATURATUS2SPERMOPHILUS ATRICAPILLUS1SPERMOPHILUS ATRICAPILLUS2SPERMOPHILUS BEECHYI1SPERMOPHILUS BEECHYI2SPERMOPHILUS VARIEGATUS1SPERMOPHILUS VARIEGATUS2SPERMOPHILUS VARIEGATUS3SPERMOPHILUS ANNULATUS1SPERMOPHILUS ANNULATUS2SPERMOPHILUS ADOCETUS1SPERMOPHILUS ADOCETUS2AMMOSPERMOPHILUS HARRISIIEUTAMIAS DORSALISORYCTOLAGUS CUNICULUS1ORYCTOLAGUS CUNICULUS2ORYCTOLAGUS CUNICULUS3LEPUS OIOSTOLUS1LEPUS OIOSTOLUS2LEPUS CAPENSIS2LEPUS CAPENSIS3LEPUS CAPENSIS6LEPUS CAPENSIS5LEPUS CAPENSIS7LEPUS CORSICANUS1LEPUS CORSICANUS2LEPUS TIMIDUS1LEPUS TIMIDUS3LEPUS MANDSHURICUS1LEPUS TIMIDUS2LEPUS OIOSTOLUS3LEPUS GRANATENSISLEPUS EUROPAEUSLEPUS CAPENSIS1LEPUS SINENSIS2LEPUS CAPENSIS8LEPUS SINENSIS3LEPUS SINENSIS1LEPUS MANDSHURICUS2LEPUS SINENSIS4LEPUS CAPENSIS4SYLVILAGUS FLORIDANUSSYLVILAGUS TRANSITIONALISOCHOTONA RUFESCENSLOXODONTA AFRICANA1LOXODONTA AFRICANA3LOXODONTA AFRICANA4LOXODONTA AFRICANA5LOXODONTA AFRICANA6LOXODONTA AFRICANA2MAMMUTHUS PRIMIGENIUS1MAMMUTHUS PRIMIGENIUS2ELEPHAS MAXIMUS1ELEPHAS MAXIMUS2ELEPHAS MAXIMUS6ELEPHAS MAXIMUS5ELEPHAS MAXIMUS3ELEPHAS MAXIMUS4HYDRODAMALIS GIGASTRICHECHUS MANATUSDUGONG DUGONPROCAVIA CAPENSISORYCTEROPUS AFER1ORYCTEROPUS AFER2ELEPHANTULUS RUFESCENSDASYPUS NOVEMCINCTUSPONGO PYGMAEUS1PONGO PYGMAEUS4PONGO PYGMAEUS2PONGO PYGMAEUS3GORILLA GORILLAPAN PANISCUSPAN TROGLODYTES1PAN TROGLODYTES2PAN TROGLODYTES3HOMO SAPIENS1HOMO SAPIENS2HOMO SAPIENS4HOMO SAPIENS3HYLOBATES LAR1HYLOBATES LAR2HYLOBATES MUELLERIHYLOBATES AGILIS1HYLOBATES AGILIS2HYLOBATES MOLOCHHYLOBATES PILEATUSHYLOBATES KLOSSIIHYLOBATES GABRIELLAEHYLOBATES LEUCOGENYSHYLOBATES HOOLOCK1HYLOBATES HOOLOCK2HYLOBATES SYNDACTYLUS1HYLOBATES SYNDACTYLUS2PAPIO HAMADRYAS1PAPIO HAMADRYAS2MACACA MULATTAPRESBYTIS ENTELLUSNASALIS LARVATUSCOLOBUS GUEREZASAIMIRI SCIUREUSLEMUR CATTA1LEMUR CATTA2LEMUR CATTA3LEMUR CATTA4LEMUR CATTA6LEMUR CATTA5HAPALEMUR GRISEUSEULEMUR RUBRIVENTEREULEMUR MONGOZEULEMUR FULVUS1EULEMUR FULVUS2EULEMUR FULVUS3EULEMUR MACACO1EULEMUR MACACO2VARECIA VARIEGATA1VARECIA VARIEGATA2MICROCEBUS MURINUSMIRZA COQUERELICHEIROGALEUS MAJORPROPITHECUS TATTERSALLIDAUBENTONIA MADAGASCARIENSISTARSIUS BANCANUSOTOLEMUR CRASSICAUDATUSNYCTICEBUS COUCANGLORIS TARDIGRADUSOVIS ARIES1OVIS ARIES2OVIS ARIES5OVIS ARIES4OVIS VIGNEIOVIS AMMONOVIS DALLI1OVIS DALLI2OVIS CANADENSISBUDORCAS TAXICOLOR1BUDORCAS TAXICOLOR2OVIS ARIES3OVIBOS MOSCHATUS1OVIBOS MOSCHATUS2OVIBOS MOSCHATUS3OVIBOS MOSCHATUS4OVIBOS MOSCHATUS5CAPRICORNIS CRISPUS2CAPRICORNIS CRISPUS1NEMORHAEDUS CAUDATUSOREAMNOS AMERICANUS1OREAMNOS AMERICANUS2AMMOTRAGUS LERVIARUPICAPRA PYRENAICARUPICAPRA RUPICAPRAHEMITRAGUS JEMLAHICUS1HEMITRAGUS JEMLAHICUS2CAPRA SIBIRICACAPRA NUBIANACAPRA AEGAGRUS1CAPRA CYLINDRICORNISCAPRA CAUCASICACAPRA FALCONERI1CAPRA FALCONERI2CAPRA AEGAGRUS2CAPRA HIRCUS1CAPRA HIRCUS6CAPRA HIRCUS2CAPRA HIRCUS3CAPRA HIRCUS5CAPRA HIRCUS4CAPRA AEGAGRUS3CAPRA HIRCUS7CAPRA HIRCUS8CAPRA IBEXPSEUDOIS NAYAURPANTHOLOPS HODGSONIDAMALISCUS PYGARGUSDAMALISCUS LUNATUSBEATRAGUS HUNTERIALCELAPHUS BUSELAPHUS1ALCELAPHUS BUSELAPHUS2ALCELAPHUS LICHTENSTEINISIGMOCEROS LICHTENSTEINIICONNOCHAETES TAURINUS1CONNOCHAETES GNOUCONNOCHAETES TAURINUS2ORYX LEUCORYXORYX DAMMAHADDAX NASOMACULATUSHIPPOTRAGUS NIGER1HIPPOTRAGUS NIGER3HIPPOTRAGUS NIGER2HIPPOTRAGUS EQUINUSANTILOPE CERVICAPRA1ANTILOPE CERVICAPRA2GAZELLA SUBGUTTUROSAGAZELLA GAZELLAGAZELLA DAMAGAZELLA GRANTIANTIDORCAS MARSUPIALIS1ANTIDORCAS MARSUPIALIS2SAIGA TATARICA1SAIGA TATARICA2MADOQUA GUENTHERIMADOQUA KIRKIIRAPHICERUS CAMPESTRISRAPHICERUS SHARPEIRAPHICERUS MELANOTISKOBUS ELLIPSIPRYMNUS2KOBUS ELLIPSIPRYMNUS3KOBUS VARDONIKOBUS KOBKOBUS MEGACEROS1KOBUS MEGACEROS2KOBUS LECHE1KOBUS LECHE2KOBUS LECHE3REDUNCA FULVORUFULA1REDUNCA FULVORUFULA2REDUNCA ARUNDINUMREDUNCA REDUNCAPELEA CAPREOLUSOREOTRAGUS OREOTRAGUS1OREOTRAGUS OREOTRAGUS2CEPHALOPHUS DORSALISCEPHALOPHUS MAXWELLIICEPHALOPHUS LEUCOGASTERAEPYCEROS MELAMPUS1AEPYCEROS MELAMPUS3AEPYCEROS MELAMPUS2NEOTRAGUS MOSCHATUS1NEOTRAGUS MOSCHATUS3NEOTRAGUS MOSCHATUS2BOS TAURUSBISON BONASUSBOS GRUNNIENSBISON BISONBOS JAVANICUS1BOS JAVANICUS2PSEUDORYX NGHETINHENSISTRAGELAPHUS STREPSICEROS1TRAGELAPHUS STREPSICEROS2TRAGELAPHUS ORYX1TRAGELAPHUS ORYX2TRAGELAPHUS SCRIPTUS1TRAGELAPHUS SCRIPTUS2TRAGELAPHUS EURYCERUS1TRAGELAPHUS EURYCERUS2TRAGELAPHUS SPEKIITRAGELAPHUS DERBIANUSTRAGELAPHUS ANGASII1TRAGELAPHUS ANGASII2TRAGELAPHUS IMBERBIS1TRAGELAPHUS IMBERBIS2TETRACERUS QUADRICORNISBOSELAPHUS TRAGOCAMELUSSYNCERUS CAFFER1SYNCERUS CAFFER2BUBALUS DEPRESSICORNIS1BUBALUS DEPRESSICORNIS2BUBALUS DEPRESSICORNIS6BUBALUS DEPRESSICORNIS3BUBALUS DEPRESSICORNIS4BUBALUS DEPRESSICORNIS5BUBALUS QUARLESIBUBALUS ARNEE3BUBALUS ARNEE15BUBALUS ARNEE14BUBALUS ARNEE16BUBALUS ARNEE4BUBALUS ARNEE5BUBALUS ARNEE12BUBALUS ARNEE13BUBALUS ARNEE17BUBALUS ARNEE18BUBALUS ARNEE1BUBALUS ARNEE2BUBALUS ARNEE6BUBALUS ARNEE7BUBALUS ARNEE8BUBALUS ARNEE9BUBALUS ARNEE10BUBALUS ARNEE11BUBALUS MINDORENSISMEGAMUNTIACUS VUQUANGENSISMUNTIACUS MUNTJAK1MUNTIACUS MUNTJAK2MUNTIACUS MUNTJAK4MUNTIACUS MUNTJAK3MUNTIACUS REEVESIMUNTIACUS SPECIESCERVUS ELAPHUS1CERVUS ELAPHUS6CERVUS ELAPHUS5CERVUS ELAPHUS2CERVUS ELAPHUS4CERVUS ELAPHUS3CERVUS NIPPON1CERVUS NIPPON2CERVUS NIPPON3CERVUS NIPPON5CERVUS NIPPON6CERVUS NIPPON4CAPREOLUS CAPREOLUS1CAPREOLUS CAPREOLUS2CAPREOLUS PYGARGUSHYDROPOTES INERMISALCES ALCESODOCOILEUS HEMIONUS1ODOCOILEUS HEMIONUS2MAZAMA SPECIESRANGIFER TARANDUSMOSCHUS LEUCOGASTERMOSCHUS FUSCUSMOSCHUS CHRYSOGASTERMOSCHUS BEREZOVSKII1MOSCHUS BEREZOVSKII2MOSCHUS MOSCHIFERUSOKAPIA JOHNSTONIGIRAFFA CAMELOPARDALISANTILOCAPRA AMERICANA1ANTILOCAPRA AMERICANA2TRAGULUS JAVANICUSTRAGULUS NAPUSUS SCROFA1SUS SCROFA5SUS SCROFA22SUS SCROFA24SUS SCROFA25SUS SCROFA27SUS SCROFA32SUS SCROFA36SUS SCROFA34SUS SCROFA35SUS SCROFA4SUS SCROFA2SUS SCROFA3SUS SCROFA21SUS SCROFA33SUS SCROFA26SUS SCROFA23SUS SCROFA6SUS SCROFA9SUS SCROFA29SUS SCROFA7SUS SCROFA8SUS SCROFA28SUS SCROFA20SUS SCROFA31SUS SCROFA15SUS SCROFA16SUS SCROFA17SUS SCROFA19SUS SCROFA30SUS SCROFA18SUS SCROFA10SUS SCROFA11SUS SCROFA12SUS SCROFA14SUS SCROFA13SUS BARBATUSPHACOCHOERUS AFRICANUSBABYROUSA BABYRUSSAPECARI TAJACU1PECARI TAJACU2TAYASSU PECARICATAGONUS WAGNERISTENELLA LONGIROSTRIS1STENELLA LONGIROSTRIS5STENELLA LONGIROSTRIS6STENELLA LONGIROSTRIS7STENELLA LONGIROSTRIS3STENELLA LONGIROSTRIS2STENELLA LONGIROSTRIS4STENELLA ATTENUATA1STENELLA ATTENUATA4STENELLA ATTENUATA2STENELLA ATTENUATA3STENELLA FRONTALIS1STENELLA FRONTALIS2STENELLA CLYMENESTENELLA COERULEOALBA1STENELLA COERULEOALBA2DELPHINUS TROPICALISDELPHINUS CAPENSIS1DELPHINUS CAPENSIS2DELPHINUS DELPHIS2DELPHINUS DELPHIS1TURSIOPS ADUNCUS1TURSIOPS ADUNCUS2TURSIOPS TRUNCATUS1TURSIOPS TRUNCATUS4TURSIOPS TRUNCATUS2TURSIOPS TRUNCATUS3SOUSA CHINENSIS1SOUSA CHINENSIS2LAGENODELPHIS HOSEI1LAGENODELPHIS HOSEI2LAGENORHYNCHUS ACUTUSGRAMPUS GRISEUS1GRAMPUS GRISEUS2PSEUDORCA CRASSIDENSGLOBICEPHALA MELAS1GLOBICEPHALA MELAS2GLOBICEPHALA MACRORHYNCHUS1GLOBICEPHALA MACRORHYNCHUS2PEPONOCEPHALA ELECTRAFERESA ATTENUATAORCINUS ORCA1ORCINUS ORCA3ORCINUS ORCA2ORCAELLA BREVIROSTRIS1ORCAELLA BREVIROSTRIS3ORCAELLA BREVIROSTRIS2LAGENORHYNCHUS AUSTRALISLAGENORHYNCHUS CRUCIGERLAGENORHYNCHUS OBLIQUIDENSLAGENORHYNCHUS OBSCURUSCEPHALORHYNCHUS COMMERSONIICEPHALORHYNCHUS EUTROPIACEPHALORHYNCHUS HECTORICEPHALORHYNCHUS HEAVISIDIILISSODELPHIS PERONIILISSODELPHIS BOREALISSOTALIA FLUVIATILISSTENO BREDANENSIS1STENO BREDANENSIS2LAGENORHYNCHUS ALBIROSTRIS1LAGENORHYNCHUS ALBIROSTRIS2PHOCOENA SINUSPHOCOENA PHOCOENA1PHOCOENA PHOCOENA2MONODON MONOCEROS1MONODON MONOCEROS2DELPHINAPTERUS LEUCAS1DELPHINAPTERUS LEUCAS2PONTOPORIA BLAINVILLEIINIA GEOFFRENSISKOGIA BREVICEPS1KOGIA BREVICEPS2PHYSETER CATODONZIPHIUS CAVIROSTRISMESOPLODON EUROPAEUSMESOPLODON DENSIROSTRISMESOPLODON BIDENSHYPEROODON AMPULLATUSBERARDIUS BAIRDIIPLATANISTA MINORBALAENOPTERA MUSCULUSBALAENOPTERA EDENIBALAENOPTERA BOREALISBALAENOPTERA PHYSALUSMEGAPTERA NOVAEANGLIAEESCHRICHTIUS ROBUSTUSBALAENOPTERA BONAERENSISBALAENOPTERA ACUTOROSTRATACAPEREA MARGINATABALAENA GLACIALISBALAENA MYSTICETUSHIPPOPOTAMUS AMPHIBIUS1HIPPOPOTAMUS AMPHIBIUS2HIPPOPOTAMUS AMPHIBIUS3HEXAPROTODON LIBERIENSISVICUGNA VICUGNALAMA PACOSLAMA GLAMALAMA GUANICOE1LAMA GUANICOE2CAMELUS BACTRIANUSCAMELUS DROMEDARIUS1CAMELUS DROMEDARIUS2CERATOTHERIUM SIMUMDICEROS BICORNISRHINOCEROS UNICORNISTAPIRUS TERRESTRISTAPIRUS INDICUSEQUUS ASINUSEQUUS GREVYIEQUUS CABALLUS1EQUUS CABALLUS3EQUUS CABALLUS2HERPESTES AUROPUNCTATUSFELIS CATUS1FELIS CATUS4FELIS CATUS3FELIS CATUS2PANTHERA TIGRIS1PANTHERA TIGRIS3PANTHERA TIGRIS8PANTHERA TIGRIS9PANTHERA TIGRIS4PANTHERA TIGRIS5PANTHERA TIGRIS6PANTHERA TIGRIS7PANTHERA TIGRIS2PANTHERA LEO1PANTHERA LEO2CANIS FAMILIARIS1CANIS FAMILIARIS2VULPES VULPESHALICHOERUS GRYPUSPHOCA HISPIDAPHOCA VITULINA1PHOCA VITULINA2PHOCA LARGHACYSTOPHORA CRISTATAPHOCA GROENLANDICAPHOCA FASCIATAERIGNATHUS BARBATUSMONACHUS SCHAUINSLANDILEPTONYCHOTES WEDDELLIHYDRURGA LEPTONYXMIROUNGA LEONINAZALOPHUS CALIFORNIANUSEUMETOPIAS JUBATUSARCTOCEPHALUS FORSTERIARCTOCEPHALUS GAZELLAODOBENUS ROSMARUSAILURUS FULGENSSELENARCTOS THIBETANUSURSUS THIBETANUSURSUS AMERICANUS1URSUS AMERICANUS2HELARCTOS MALAYANUSURSUS ARCTOS1URSUS ARCTOS2URSUS ARCTOS3URSUS ARCTOS22URSUS ARCTOS24URSUS ARCTOS25URSUS ARCTOS26URSUS ARCTOS4URSUS ARCTOS5URSUS ARCTOS7URSUS ARCTOS8URSUS ARCTOS9URSUS ARCTOS10URSUS ARCTOS11URSUS ARCTOS14URSUS ARCTOS12URSUS ARCTOS13URSUS ARCTOS15URSUS ARCTOS16URSUS ARCTOS18URSUS ARCTOS19URSUS ARCTOS21URSUS ARCTOS23URSUS ARCTOS27URSUS ARCTOS28URSUS ARCTOS20URSUS ARCTOS17URSUS ARCTOS6URSUS ARCTOS29URSUS ARCTOS31URSUS ARCTOS32URSUS ARCTOS33URSUS ARCTOS30THALARCTOS MARITIMUS1THALARCTOS MARITIMUS2MELURSUS URSINUS1MELURSUS URSINUS2TREMARCTOS ORNATUSAILUROPODA MELANOLEUCA1AILUROPODA MELANOLEUCA2MARTES FLAVIGULA1MARTES FLAVIGULA2MARTES ZIBELLINA1MARTES ZIBELLINA2MARTES ZIBELLINA4MARTES MELAMPUS1MARTES ZIBELLINA3MARTES ZIBELLINA5MARTES MARTESMARTES AMERICANA1MARTES AMERICANA2MARTES AMERICANA3MARTES AMERICANA4MARTES AMERICANA5MARTES AMERICANA6MARTES AMERICANA7MARTES AMERICANA8MARTES AMERICANA10MARTES AMERICANA9MARTES MELAMPUS2MARTES MELAMPUS3MARTES MELAMPUS7MARTES MELAMPUS4MARTES MELAMPUS5MARTES MELAMPUS11MARTES MELAMPUS8MARTES MELAMPUS13MARTES MELAMPUS15MARTES MELAMPUS12MARTES MELAMPUS9MARTES MELAMPUS14MARTES MELAMPUS10MARTES MELAMPUS16MARTES MELAMPUS6MARTES PENNANTIGULO GULOMUSTELA VISONMUSTELA PUTORIUSMUSTELA ERMINEAPTERONURA BRASILIENSISLUTRA MACULICOLLISLUTRA LUTRA1LUTRA LUTRA2AMBLONYX CINEREUSAONYX CAPENSISENHYDRA LUTRIS1ENHYDRA LUTRIS2LONTRA LONGICAUDISLONTRA FELINALONTRA CANADENSISTAXIDEA TAXUSMELES MELESPROCYON LOTORBASSARICYON GABBIISPILOGALE PUTORIUSMEPHITIS MEPHITISARTIBEUS JAMAICENSIS1ARTIBEUS JAMAICENSIS3ARTIBEUS OBSCURUS1ARTIBEUS OBSCURUS2ARTIBEUS PLANIROSTRISARTIBEUS LITURATUSARTIBEUS INTERMEDIUSARTIBEUS FIMBRIATUSARTIBEUS INOPINATUSARTIBEUS HIRSUTUSARTIBEUS FRATERCULUSARTIBEUS TOLTECUSARTIBEUS PHAEOTISARTIBEUS GLAUCUS1ARTIBEUS AZTECUSARTIBEUS GLAUCUS2ARTIBEUS GLAUCUS3ARTIBEUS CINEREUSARTIBEUS ANDERSENIARTIBEUS CONCOLOR1ARTIBEUS CONCOLOR2ARTIBEUS HARTIIERINACEUS EUROPAEUS1ERINACEUS EUROPAEUS2TALPA EUROPAEA1TALPA EUROPAEA2TALPA ALTAICAMOGERA WOGURA1MOGERA WOGURA2MOGERA WOGURA3MOGERA IMAIZUMIIMOGERA TOKUDAEMOGERA INSULARISEUROSCAPTOR MIZURAUROTRICHUS TALPOIDESCROCIDURA RUSSULASOREX TUNDRENSIS2SOREX ASPER1SOREX ASPER2SOREX CORONATUS1SOREX CORONATUS2SOREX GRANARIUS1SOREX GRANARIUS2SOREX ARANEUS1SOREX ARANEUS2SOREX ARANEUS3SOREX ARCTICUS1SOREX ARCTICUS2SOREX SAMNITICUS1SOREX SAMNITICUS2SOREX UNGUICULATUSSOREX ISODON1SOREX ISODON2SOREX SHINTO1SOREX SHINTO2SOREX CAECUTIENS1SOREX CAECUTIENS2SOREX MINUTUSSOREX MINUTISSIMUS1SOREX MINUTISSIMUS2SOREX RADDEI1SOREX RADDEI2SOREX EXCELSUS1SOREX EXCELSUS2SOREX ALPINUS1SOREX ALPINUS2SOREX CINEREUS2SOREX HAYDENI1SOREX HAYDENI2SOREX VAGRANSSOREX PACIFICUS1SOREX PACIFICUS2SOREX MONTICOLUS1SOREX MONTICOLUS2SOREX PALUSTRIS1SOREX PALUSTRIS2SOREX HOYISOREX FUMEUS1SOREX FUMEUS2SOREX TROWBRIDGII1SOREX TROWBRIDGII2NEOMYS ANOMALUSNEOMYS FODIENS
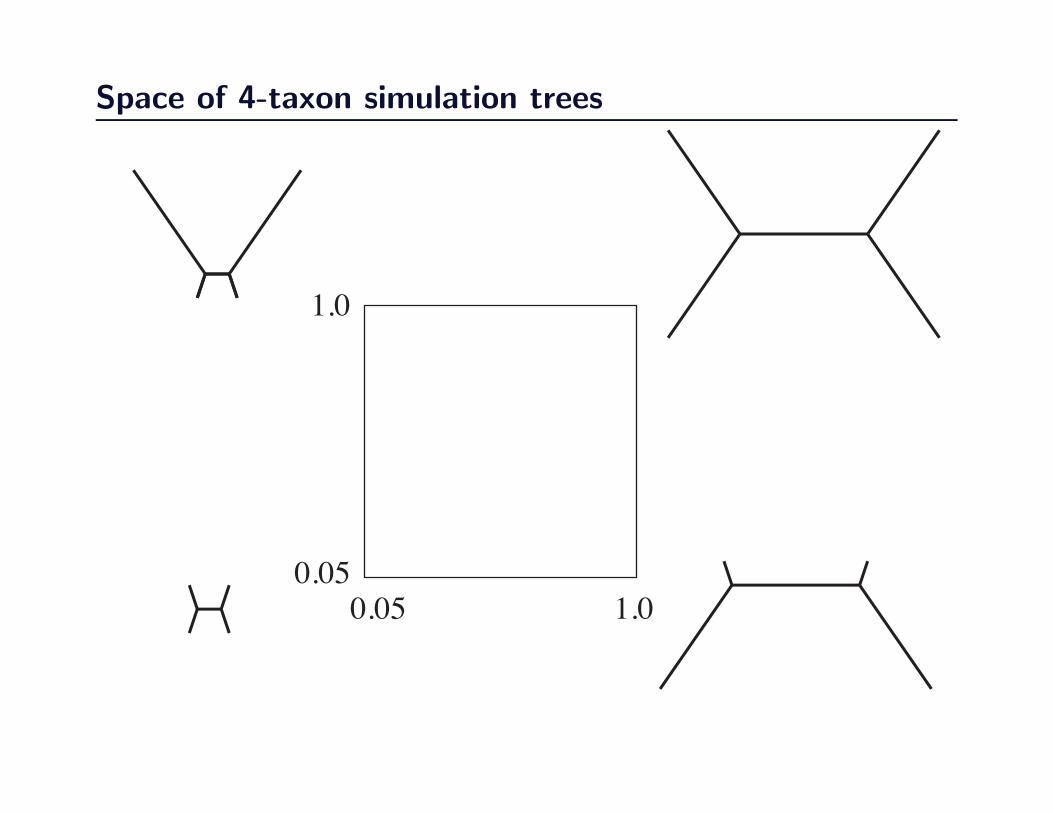
Space of 4-taxon simulation trees
0.05 1.0
1.0
0.05
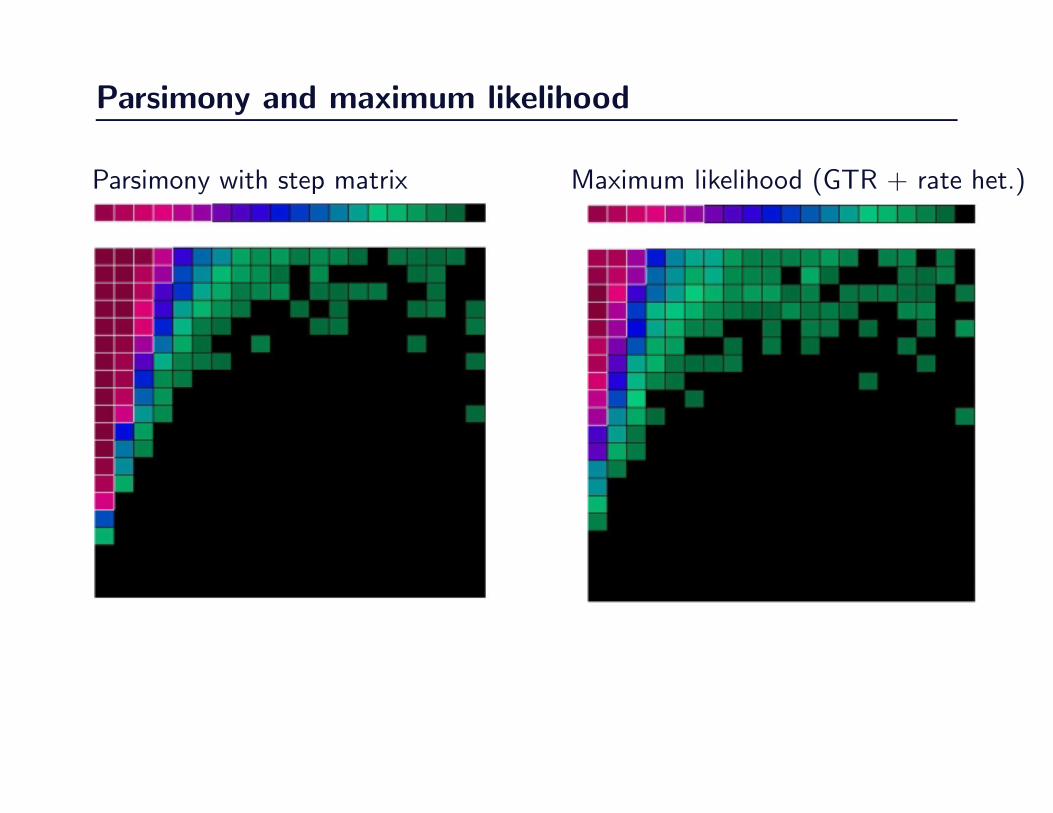
Parsimony and maximum likelihood
99
Figure 3.6
100
Figure 3.7
Parsimony with step matrix Maximum likelihood (GTR + rate het.)

Contrasting parsimony and maximum likelihood
115
Figure 3.22
A contrast of DNA-based weighted parsimony and maximum likelihood using GTR
with preferred rate heterogeneity made by subtracting the number of successes by one
method from the number of successes by the other method across the paremeter space
shown in Figure 3.2. The maximum value is 100 (shown in deep blue), the minimum
value is –100 shown as deep red. White indicates equivalent performance.
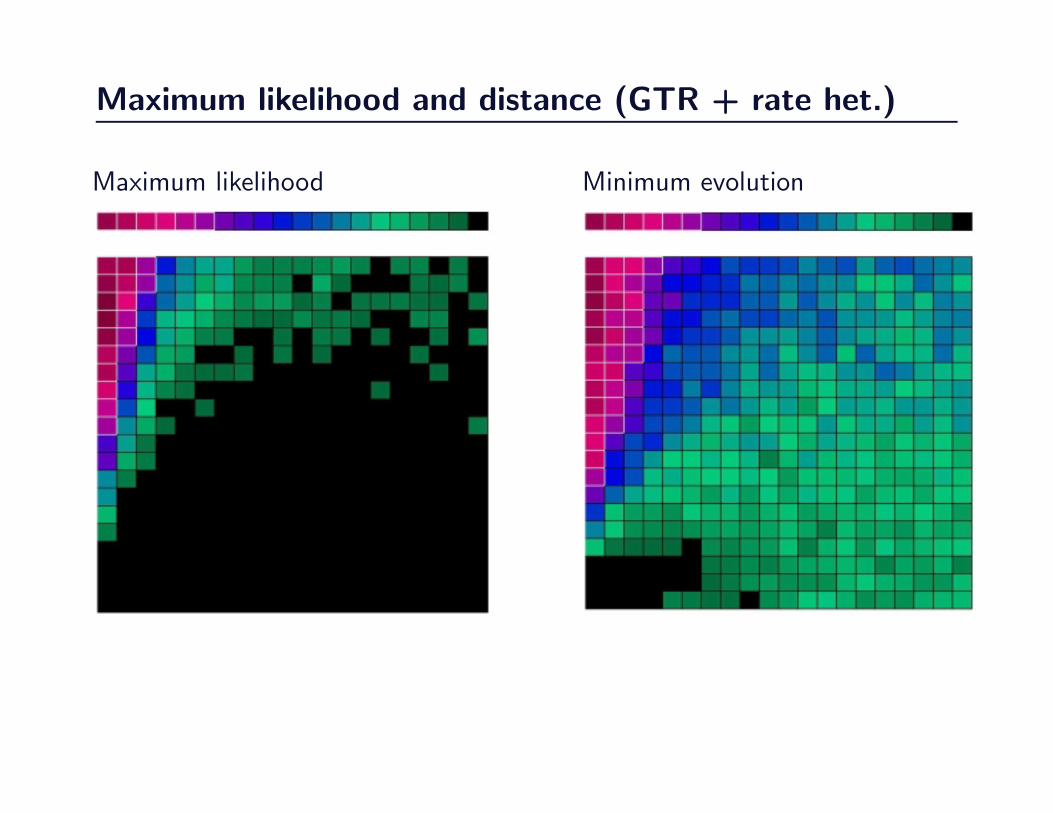
Maximum likelihood and distance (GTR + rate het.)
100
Figure 3.7
106
Figure 3.13
Maximum likelihood Minimum evolution
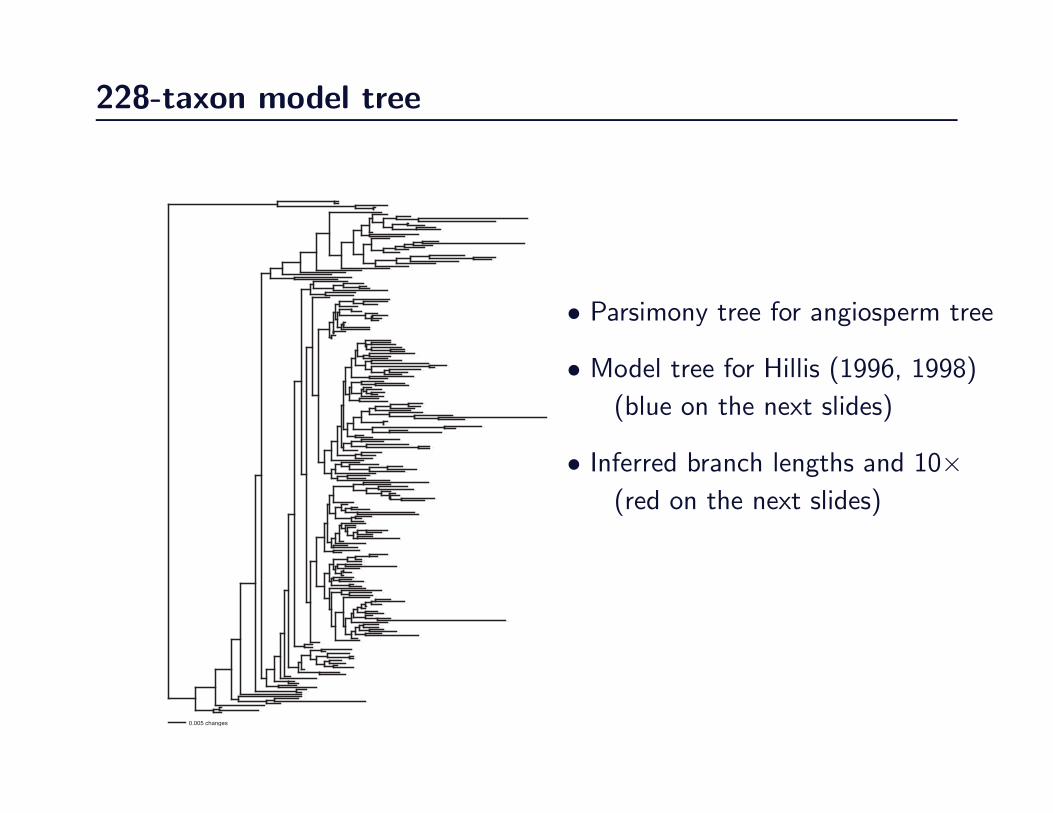
228-taxon model tree
t1
t2
t3
t4
t5
t6
t7
t8
t13
t27
t10
t11
t15
t16
t9
t12
t28
t20
t29
t31
t30
t34
t32
t33
t21
t24
t25
t26
t22
t23
t14
t18
t17
t35
t91
t136
t37
t47
t48
t49
t52
t44
t70
t71
t73
t80
t82
t81
t74
t75
t76
t77
t79
t78
t83
t85
t86
t84
t89
t90
t87
t88
t101
t102
t108
t106
t133
t135
t109
t111
t110
t104
t105
t112
t113
t149
t150
t153
t103
t107
t120
t134
t152
t151
t155
t156
t157
t158
t137
t165
t114
t115
t159
t163
t118
t160
t161
t162
t116
t117
t129
t130
t131
t132
t148
t164
t124
t154
t125
t126
t127
t128
t121
t122
t123
t138
t139
t142
t144
t145
t147
t140
t141
t143
t146
t92
t93
t94
t95
t96
t97
t98
t99
t100
t189
t175
t177
t179
t180
t182
t183
t184
t185
t169
t173
t178
t181
t193
t192
t187
t188
t174
t191
t186
t190
t170
t171
t166
t194
t195
t196
t202
t212
t221
t217
t223
t218
t203
t225
t226
t227
t228
t167
t168
t197
t198
t172
t176
t201
t210
t208
t207
t205
t206
t204
t213
t222
t199
t209
t211
t216
t219
t214
t220
t215
t200
t224
t65
t66
t54
t119
t55
t56
t57
t59
t60
t61
t62
t58
t63
t64
t72
t67
t68
t69
t42
t43
t45
t46
t19
t36
t50
t53
t51
t39
t38
t40
t41
0.005 changes
• Parsimony tree for angiosperm tree
• Model tree for Hillis (1996, 1998)
(blue on the next slides)
• Inferred branch lengths and 10×(red on the next slides)
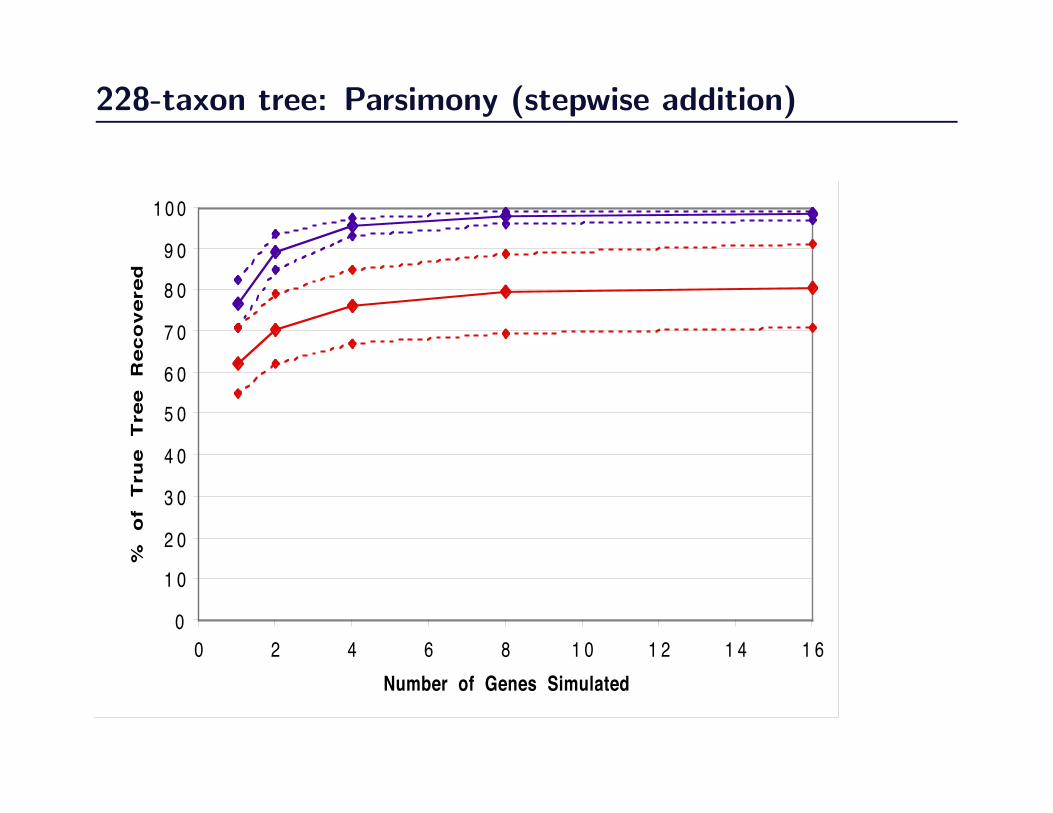
228-taxon tree: Parsimony (stepwise addition)
1
Parsimony Stepwise Addition
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
0 2 4 6 8 1 0 1 2 1 4 1 6Number of Genes Simulated
%
of
Tru
e
Tre
e
Re
co
ve
red
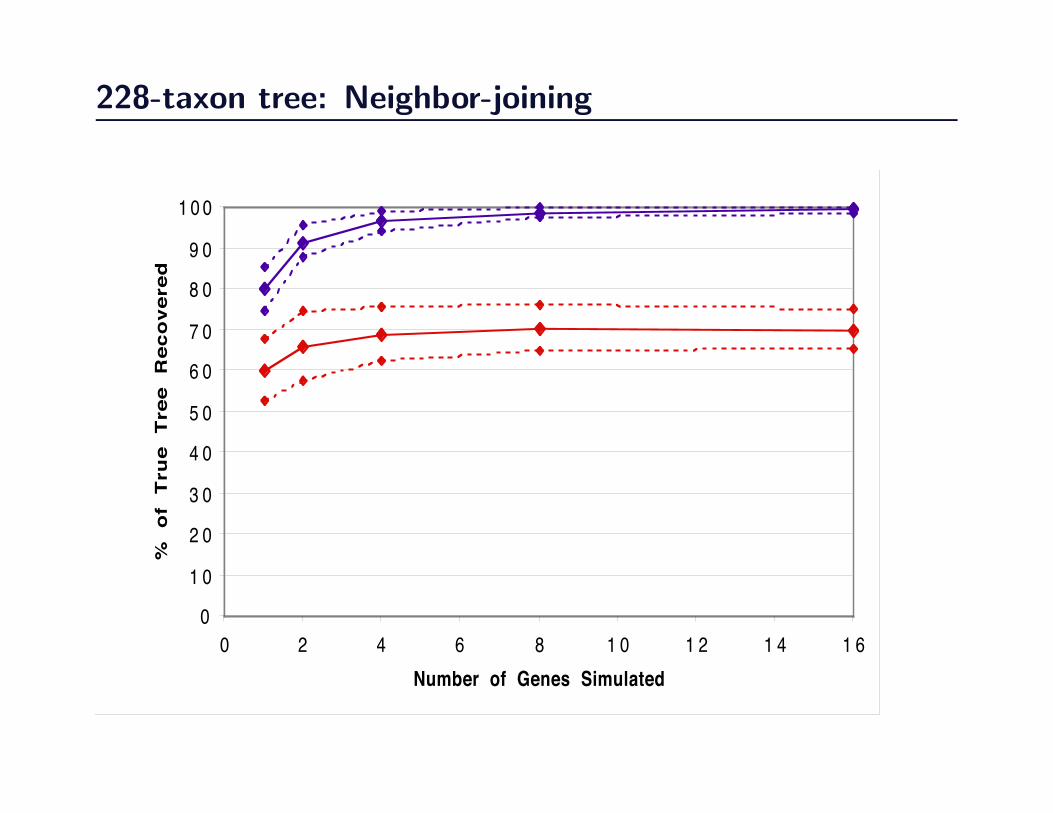
228-taxon tree: Neighbor-joining
1
Neighbor Joining
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
0 2 4 6 8 1 0 1 2 1 4 1 6Number of Genes Simulated
%
of
Tru
e
Tre
e
Re
co
ve
red
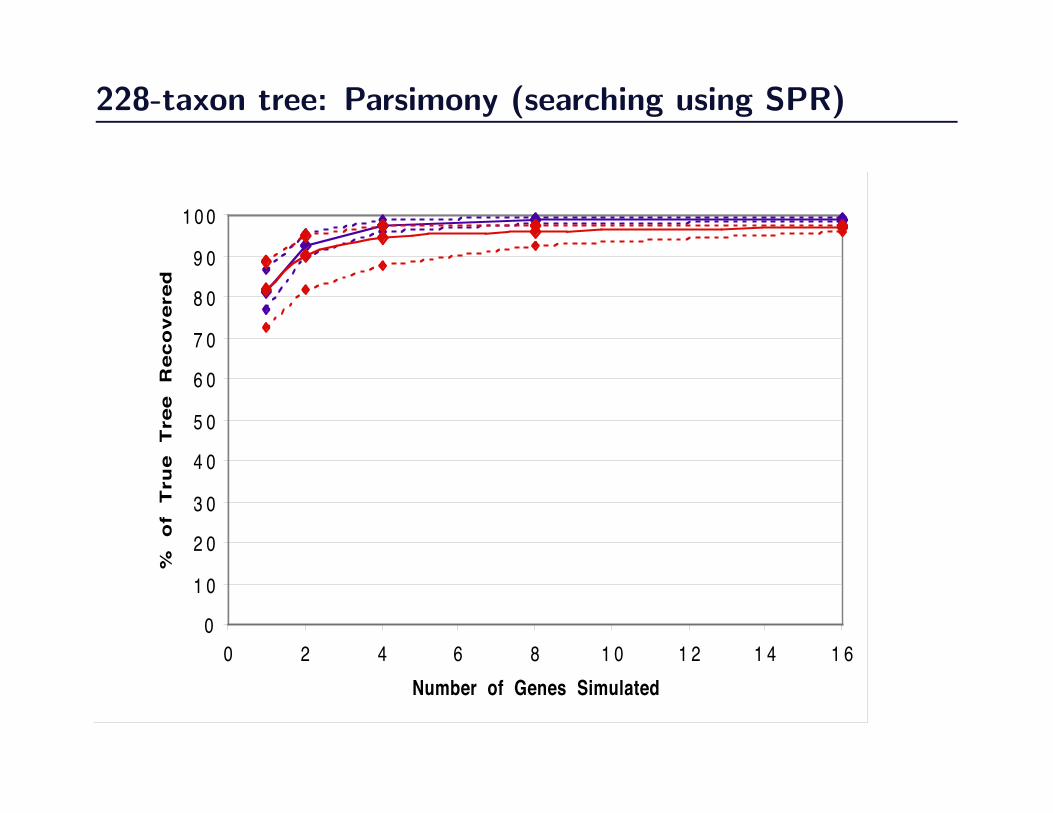
228-taxon tree: Parsimony (searching using SPR)
1
Parsimony Search
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
0 2 4 6 8 1 0 1 2 1 4 1 6Number of Genes Simulated
%
of
Tru
e
Tre
e
Re
co
ve
red
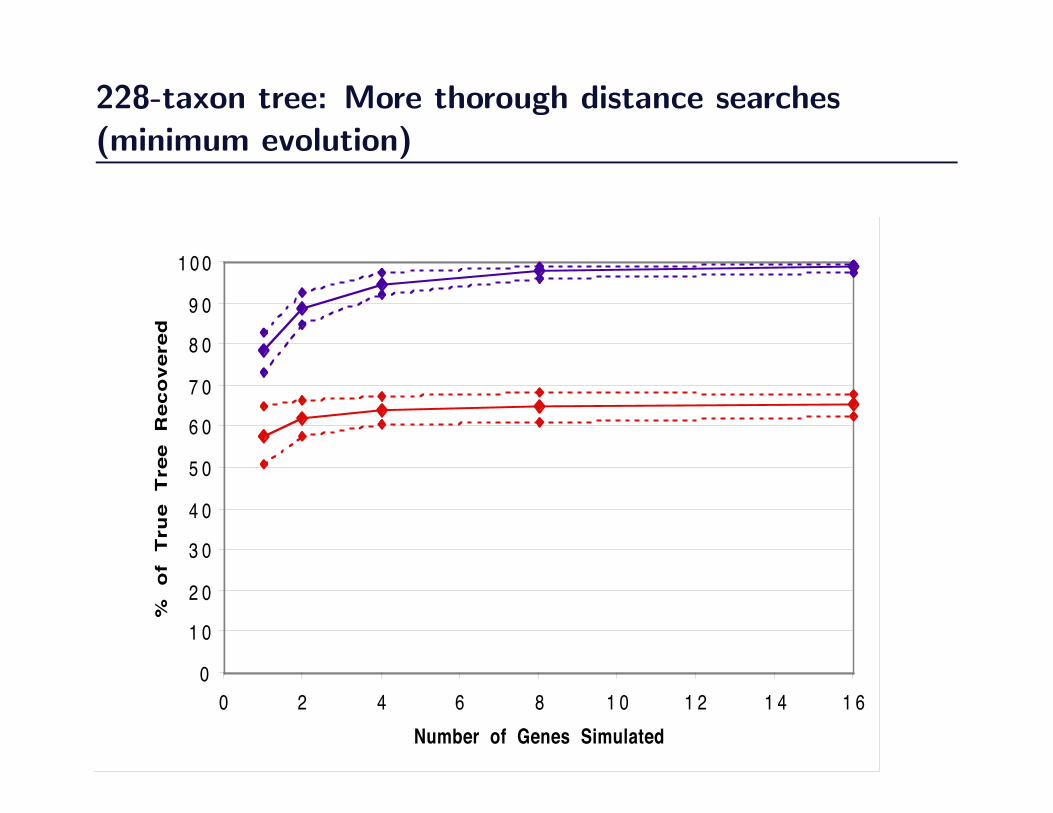
228-taxon tree: More thorough distance searches(minimum evolution)
1
Minimum Evolution Search
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
0 2 4 6 8 1 0 1 2 1 4 1 6Number of Genes Simulated
%
of
Tru
e
Tre
e
Re
co
ve
red

Randomly generated trees
Model trees for 50 leaves generated:
• by a pure birth process, followed by selection of 50% of the taxa.
• rate of evolution then allowed to evolve along the tree Kishino et al.(2001)
• mean branch length has an expected # changes per site ≈ 0.02
The following results are very preliminary.
t36
t27
t45
t 6
t 4
t11
t22
t17
t39
t25
t31
t20
t40
t49
t15
t44
t10
t41
t28
t16
t26
t 2
t38
t50
t 8
t29
t32
t46
t 5
t 3
t21
t34
t35
t19
t42
t12
t24
t43
t37
t18
t30
t47
t13
t14
t33
t48
t 9
t 1
t23
t 7
0.03
t46
t30
t28
t10
t38
t37
t45
t36
t 9
t41
t47
t21
t22
t33
t25
t34
t27
t32
t14
t23
t40
t 8
t 7
t16
t11
t49
t39
t42
t17
t24
t44
t19
t18
t 2
t 1
t12
t 6
t26
t13
t29
t35
t 3
t48
t31
t20
t 4
t50
t 5
t43
t15
0.03
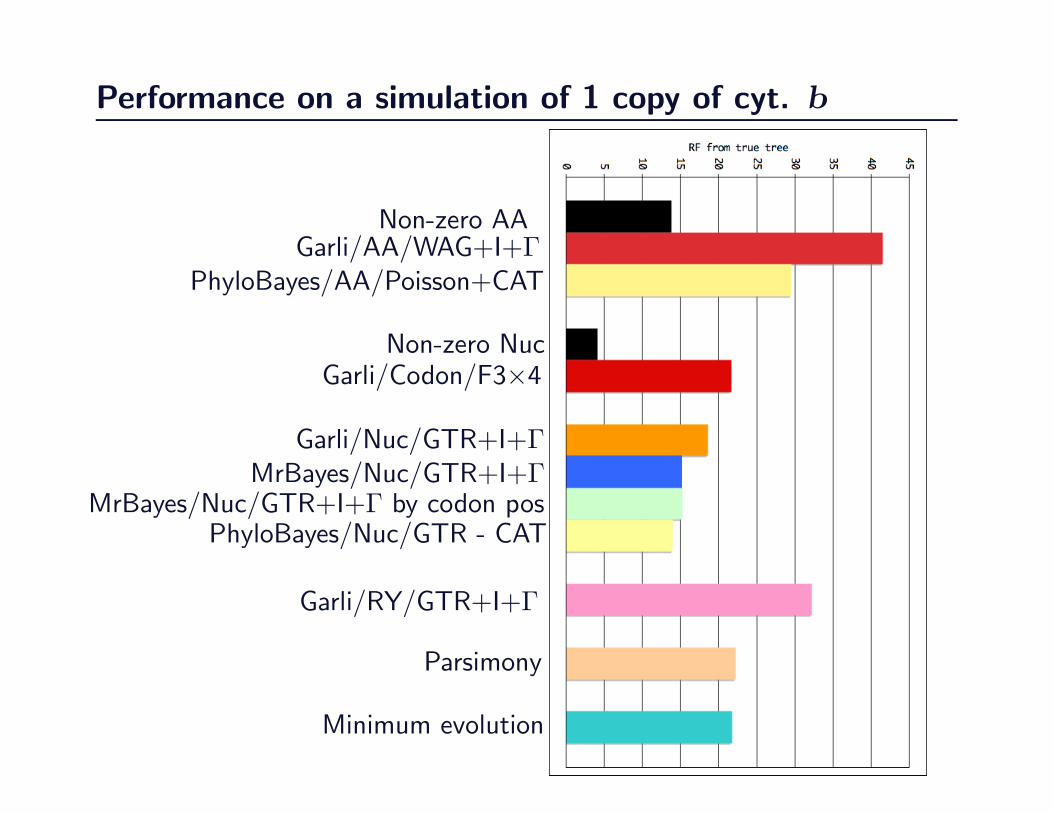
Performance on a simulation of 1 copy of cyt. b
Non-zero AAGarli/AA/WAG+I+Γ
PhyloBayes/AA/Poisson+CAT
Non-zero NucGarli/Codon/F3×4
Garli/Nuc/GTR+I+ΓMrBayes/Nuc/GTR+I+Γ
MrBayes/Nuc/GTR+I+Γ by codon posPhyloBayes/Nuc/GTR - CAT
Garli/RY/GTR+I+Γ
Parsimony
Minimum evolution
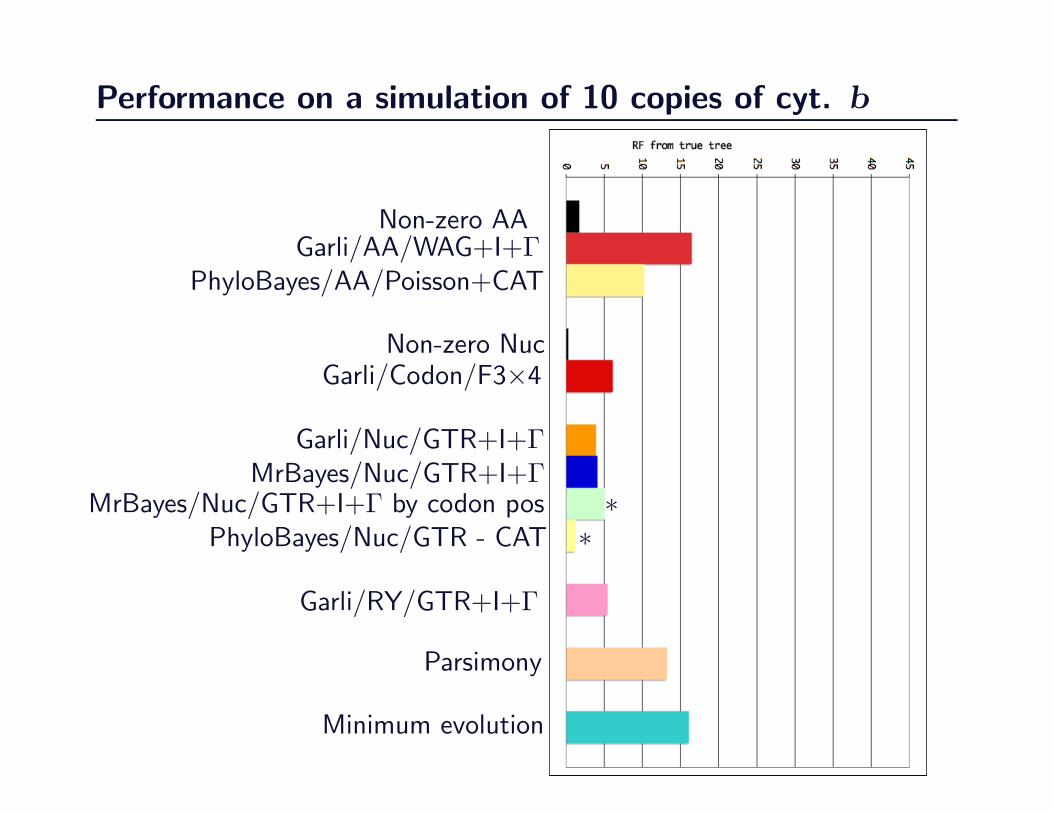
Performance on a simulation of 10 copies of cyt. b
Non-zero AAGarli/AA/WAG+I+Γ
PhyloBayes/AA/Poisson+CAT
Non-zero NucGarli/Codon/F3×4
Garli/Nuc/GTR+I+ΓMrBayes/Nuc/GTR+I+Γ
MrBayes/Nuc/GTR+I+Γ by codon pos ∗PhyloBayes/Nuc/GTR - CAT ∗
Garli/RY/GTR+I+Γ
Parsimony
Minimum evolution

Conclusions from simulations under Halpern-Brunomodel fit to cytochrome b data
• in general, likelihood-based inference on the nucleotide level appearsrobust,
• mixture/partitioned approaches performing well,
• parsimony’s performance was quite variable (depending on model treeshape),
• distance methods are much more sensitive to model violation,
• analyses at the amino acid level were substantially less accurate
however. . .

Caveats about simulations under Halpern-Bruno modelfit to cytochrome b data
• On small trees, the simulator generates amino sequences with relativelylittle variation
– cyt b is constrained,– overfitting of amino acid parameters could be decreasing the variability
• Still too simple:
– homogeneity of mutational process (over space and time),– lack of codon bias
Could cause the nucleotide-level conclusions to be too optimistic.
These drawbacks can be addressed by fitting parameters from more genes,and using parameter estimates from other clades estimate the rate of changein parameter values over time.

Perspective and Discussion questions
Devising realistic, complex simulators still an open area of research. Suchtools could allow us to compare fundamentally different analysis styles (e.g.codon vs. nucleotide vs amino acid), and could provide sound guidanceabout the appropriateness of a model – evidence to be used in conjuctionwith standard model choice techniques.
• Is it feasible to develop compelling simulators? Can we really make themcomplex, and yet realistic without knowing the “true” model?
• Suppose that AIC (or your-favorite-model-selection) framework stronglyprefers model X over model Y. Would a simulation study saying thatmodel Y is more robust and reliable sway you?

References
Bourlat, S., Juliusdottir, T., Lowe, C., and Freeman, R. (2006).Deuterostome phylogeny reveals monophyletic chordates and the newphylum xenoturbellida. Nature.
Buckley, T. R., Simon, C., and Chambers, G. K. (2001). Exploring among-site rate variation models in a maximum likelihood framework usingempirical data: effects of model assumptions on estimates of topology,branch lengths, and bootstrap support. Systematic Biology, 50(1):67–86.
Delsuc, F., Brinkmann, H., Chourrout, D., and Philippe, H. (2006).Tunicates and not cephalochordates are the closest living relatives ofvertebrates. Nature.
Halpern, A. L. and Bruno, W. J. (1998). Evolutionary distances for protein-coding sequences: modeling site-specific residue frequencies. MolecularBiology and Evolution, 15(7):910–917.

Hillis, D. M. (1996). Inferring complex phylogenies. Nature, 383(6596):130–131.
Hillis, D. M. (1998). Taxonomic sampling, phylogenetic accuracy, andinvestigator bias. Systematic Biology, 47(1):3–8.
Kishino, H., Thorne, J. L., and Bruno, W. J. (2001). Performance of adivergence time estimation method under a probabilistic model of rateevolution. Molecular Biology and Evolution, 18:352–361.
Marshall, D., Simon, C., and Buckley, T. (2006). Accurate branch lengthestimation in partitioned bayesian analyses requires accommodation ofamong-partition rate variation and attention to branch length priors.Systematic Biology, 55(6):993–1003.
Minin, V., Abdo, Z., Joyce, P., and Sullivan, J. (2003). Performance-based selection of likelihood models for phylogeny estimation. SystematicBiology, 52(5):674–683.





























