Embed Size (px)
Citation preview

47
Núm.20, pp.47-72, ISSN 1405-2768; México, 2005
ESTUDIO CITOGENÉTICO DEL MAÍZ HÍBRIDO SIMPLE H-311 ( ZEA MAYS SSP .MAYS), TEOCINTLE CHALQUEÑO (ZEA MAYS SSP. MEXICANA) Y SU HÍBRIDO
F1 (ZEA MAYS SSP . MAYS X ZEA MAYS SSP. MEXICANA)
Saúl Flores-Maya, Olivares-Carrillo José Luis, Flores-Crespo Lucina Ireney Rivera-Aguilar, Víctor
Unidad de Biología, Tecnología y Prototipos, Facultad de Estudios SuperioresIztacala, Avenida de los Barrios No 1, Los Reyes Iztacala, CP 54090 Tlalnepantla,
Edo. de México, Tel. 01(5) 56 23 11 34 y Fax 01(5) 56-231225.E-mail: saulsel@ servidor.unam.mx.
RESUMEN
Se analizó la frecuencia de los quiasmas enprofase y metafase I meiótica en células deplantas de maíz híbrido simple H-311 (Zeamays ssp. mays), teocintle chalqueño (Zeamays ssp. mexicana) y la F1(Zea mays ssp.mays X Zea mays ssp. mexicana) resultante.La frecuencia de quiasmas en la F1 fue me-nor que en sus progenitores. En el maíz hí-brido simple H-311 el promedio de quiasmastotales por célula fue de 21.88, en el teocintlechalqueño 21.31 y para la F1 16.63. En elmaíz híbrido simple H-311 el promedio dequiasmas totales por célula fue de 21.88, enel teocintle chalqueño 21.31 y para la F116.63. En las plantas híbridas F1, el aparea-miento entre los cromosomas homólogos nofue regular, se presentaron univalentes,trivalentes, tetravalentes, pentavalentes yhexavalentes. Los tres tipos de plantas pre-sentaron un número cromosómico diploidede 2n = 20. La longitud total promedio porgenomio fue de 33.51 µm para el maíz híbri-do simple H-311, 33.18 µm para el teocintlechalqueño y 33.33 µm para la F1. La longi-tud total de los cromosomas de los tresgenomios fue muy similar, presentaron unalongitud entre 2 y 5 µm. Los tres cariotipos
estuvieron formados por cromosomasmetacéntricos y submetacéntricos. El maíz hí-brido simple H-311 y el teocintle chalqueñopresentaron una estrecha r elaciónfilogenética, dado que tienen el mismo nú-mero cromosómico, así como una longitud yrelación de brazos similar en suscromosomas, además de haber apareamien-to e intercambio genético entre loscromosomas de ambas plantas cuando sepresentan en el híbrido F1.
Palabras clave: quiasma, cromosoma B,cariotipo, Zea mays ssp. mays, Zea maysssp. mexicana.
ABSTRACT
Meiotic prophase and metaphase I chiasmafrequency was analyzed in cells of maizehybrid simple H-311 (Zea mays ssp. mays),teosintle chalqueño (Zea mays ssp. mexi-cana) and their resulting hybrid F1(Zea maysssp. mays X Zea mays ssp. mexicana).Chiasma frequency was smaller in F
1 than in
its progenitors. Mean total chiasms per cellwas 21.88 in maize hybrid simple H-311,21.31 in teosintle chalqueño and 16.63 inthe F1.

48
Diciembre 2005Núm. 20:47-72
In the hybrids, the crossing betweenhomologous chromosomes was irregular,presenting univalents, trivalents, tetravalents,pentavalents and hexavalents. All three planttypes presented a diploid chromosomenumber of 2n = 20. The mean total length pergenome was 33.51 µm in maize hybrid simpleH-311, 33.18 µm in teosintle chalqueño and33.33 µm in the F1. The total length of thechromosomes of all three genomes was similar,with a length between 2 and 5 µm. The threekaryotypes constituted of metacentric andsubmetacentric chromosomes. Maize hybridsimple H-311 and teosintle chalqueñopresented a close phylogenetic relationship,as they had the same chromosome numberand similar relationships and lengths betweenchromosome arms. Moreover, the hybrid
F1
genome presents chromosome pairing andgenetic exchange between chromosomes ofboth plants.
Key words: chiasma, B chromosome, karyotype,Zea mays ssp. mays, Zea mays ssp. mexicana.
INTRODUCCIÓN
El maíz (Zea mays ssp. mays) estáestrechamente relacionado con la plantasilvestre llamada teocintle, de la cual seconocen: Zea perennis, teocintle perennetetraploide; Zea diploperennis, teocintleperenne anual; Zea mays ssp. parviglumisy Zea mays ssp. mexicana, son teocintlesanuales. La especie tetraploide tiene 40cromosomas somáticos, el teocintlediploperenne y los teocintles anuales tienenel mismo número cromosómico 2n = 20,como el maíz. Del teocintle Zea mays ssp.mexicana se reconocen las variedadesNovogame, Mesa central, Durango ychalqueño (Pääbo, 1999; Coe, 2001; Pressoiry Berthaud, 2004 ).
Los teocintles pueden ser cruzados con elmaíz y producir híbridos parcialmente fértilescomo sucede con la forma perenne, o híbridosfértiles al cruzar el maíz con las formas anuales.En estudios de recombinación se haobservado que entre los cromosomas delmaíz y sus respectivos homólogos en lasvariedades anuales Durango y Florida (Zeamays ssp. parviglumis), el entrecruzamientoentre los marcadores c-wx del brazo corto delcromosoma 9 es muy bajo o no lo hay,mientras en los cromosomas del maíz y delteocintle chalqueño el entrecruzamiento enla misma región es similar al que se presentaentre los cromosomas del maíz (Kato, 1984).Los cromosomas del maíz y del teocintle hansido estudiados en la fase de paquiteno,demostrando su gran similitud en la longitud,relación de brazos y ubicación delcentrómero, cromómeros conspicuos ypresencia de nudos cromosómicos, ademásde ciertos cromosomas adicionalesdenominados cromosomas B (González-Sánchez et al., 2003). Sin embargo, hay pocainformación sobre la morfología de loscromosomas del maíz, del teocintle y delhíbrido resultante en la metafase mitótica. Encuanto al apareamiento de los cromosomasse sabe que en la fase de diacinesis de lameiosis de los híbridos F1 (maíz X teocintleFlorida) falta apareamiento entre doscromosom as, encon trán dose comounivalentes o unidos por un solo extremocomo en los híbridos del maíz X teocintleDurango, indicando quizá una reducción dequiasmas. Sin embargo, hay escasainformación sobre apareamiento en diacinesis(profase meiótica) y frecuencia de quiasmasen la metafase I meiótica, especialmente enhíbridos F1 del maíz X teocintle chalqueño(Kato, 1984).
El estudio de las semejanzas cromosómicasentre maíz y teocintle es importante porque

49
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
puede confirmar la información acerca desus relaciones filogenéticas y probablementede mecanismos de evolución cromosómicaocurridos, ya que están estrechamenterelacionados (Kato, 1984). Aunado a esto,en los últimos años se ha dado auge a losestudios moleculares (Gourmet y Rayburn,1996; Goodnight, 2000; Stewart et al., 2003),los cuales dan información muy específicapor medio de marcadores en determinadoscromosomas. Sin embargo, esto no deberestar importancia a los estudioscitogenéticos con métodos clásicos, loscuales son un primer paso y permiten ademásobservar y analizar el comportamientoconjunto de todo el genomio.
El presente estudio tuvo como objetivocontr ibuir al conocimiento sobre lascaracterísticas cariotípicas mitóticas, elcom por t amien to del a pareamien tocromosómico en diacinesis y el análisis dela variación en la frecuencia de quiasmas enla metafase I en un híbrido F1 (maíz híbridosimple H-311 X teocintle chalqueño) conrespecto a sus progenitores.
MATERIAL Y MÉTODOS
Material genético
Para este estudio se utilizaron semillas delmaíz híbrido simple H-311 (Zea mays ssp.mays) y del teocintle chalqueño (Z. maysssp. mexicana). Las semillas del maíz fueronadquiridas en la empresa Berentsen (Centralde Abastos F-42-A, DF) y las del teocintlefueron recolectadas a orillas de un campode cultivo de maíz sobre la carretera Chalco-Amecameca frente al poblado de Chalco,Estado de México (fig. 1).
Parte de las semillas fue sembrada en elinvernadero de la Facultad de Estudios
Superiores Iztacala en el ciclo agrícolaprimavera-verano del 2002, de acuerdo a latécnica de Jugenheimer (1990). De estasplantas se recolectaron y almacenaron bajorefrigeración algunas espigas, de las cuales20 espigas del maíz y 20 del teocintle fueronanalizadas.
Hibridización
Por otra parte, se hicieron cruzas directas yrecíprocas. Las inflorescencias femeninas delas plantas del maíz fueron polinizadas congranos de polen de las plantas del teocintley viceversa para obtener las cruzasrecíprocas. Ambas polinizaciones se llevarona cabo con el siguiente procedimiento: antesde que los estilos de las inflorescenciasfemeninas fueran visibles, el jilote fuecubierto con una bolsa encerada para evitarfecundación por polen no deseado. Un díaantes de la polinización los jilotes fueronpreparados, lo cual se hizo cortando losestilos de cada jilote al ras de las hojas quelos envolvían. Esto se hizo con el objetode que al momento de efectuar laspolinizaciones, dichos estilos tuvieran uncrecimiento uniforme, tratando con esto quelos granos de polen quedaran situados enun plano más o menos uniforme y asegurarun mayor número de óvulos fecundados.Después los jilotes fueron cubiertosnuevamente hasta alcanzar la polinización.Cuando las inflorescencias masculinasempezaron a soltar polen, muy tempranoantes de que saliera el sol y se presenta-ra la dehiscencia, fueron tomadas de lasespigas algunas anteras y colocadas conbolsas enceradas, previamente prepara-das con tul, en la parte superior. Enton-ces el jilote fue descubierto y rápidamen-te se colocó una bolsa conteniendo lasanteras en la zona con tul, ésta fue fijadaa la base de la mazorca, doblada y

50
Diciembre 2005Núm. 20:47-72
Fig. 1. Semillas de maíz híbrido H-311 (Zea mays ssp. mays y teocintle chalqueño(Zea mays ssp. mexicana).
engrapada en la parte superior para pre-venir una posible contaminación conpolen dispersado por el viento o diver-sos insectos (Aguirre y Kato, 1979).
De las semillas híbridas obtenidas, una par-te fue nuevamente sembrada en el mismocampo de cultivo en el ciclo agrícola prima-vera-verano del 2003 y posteriormente delas plantas híbridas se recolectaron y alma-cenaron las espigas para su estudio.
Obtención del material citológico.Preparaciones mitóticas
Para la observación de los cromosomasmitóticos, las semillas del maíz híbrido simpleH-311, las del teocintle chalqueño y las de loshíbridos F1, fueron germinadas en cajas de petrien una incubadora a 30°C y después de tresdías se tomó el ápice radicular de cada una deellas. Estos ápices fueron tratados de acuerdoa la técnica citogenética de García (1988), paraobtener células en metafases iniciales.
Preparaciones meióticas
Para la observación de los cromosomas endiacinesis y metafase I, en las variedades delmaíz híbrido simple H-311, del teocintlechalqueño y de los híbridos resultantes, laspreparaciones fueron hechas con la técnicatradicional del carmín propiónico, siguiendola descripción dada por García (1988).
De cada cromosoma, se determinó la longi-tud de cada uno de los brazos. La relación debrazos se calculó dividiendo la longitud delbrazo largo entre la del brazo corto y fueronclasificados de acuerdo a la posición delcentrómero (Levan et al., 1964).
Para los cromosomas meióticos las observacio-nes se llevaron a cabo en 400 aumentos en lasfases de diacinesis y metafase I. Fueron obser-vadas 200 células de 20 plantas (10 células porcada una) para cada tipo de variedad y fase.

51
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
factor la variedad. Se aplicó este análisistomando en cuenta que los datos fueronindependientes y se distribuían de maneranormal.
RESULTADOS Y DISCUSIÓN
Frecuencia de quiasmas
Los promedios del número de quiasmastotales, intercalares y distales, por célula delas variedades estudiadas en el presentetrabajo, se muestran en el cuadro 1. Losdatos del análisis de varianza para el númerode quiasmas en diferentes posiciones en lasvariedades del maíz híbrido simple H-311,del teocintle chalqueño y de la F1 aparecenen el cuadro 2. En este cuadro se observóque el valor de F calculado y el valorestablecido por la F de tablas indica quehubo diferencias significativas entre lasmedias del número de quiasmas, lo cualsugiere ciertas modificaciones en el genomade una o ambas variedades, como lo son:diversas inversiones encontradas envariedades de teocintles en México, entreellos el teocintle chalqueño (Kato, 1984);translocaciones, o bien por mutaciones enloci genéticos como los que marcan lasdiferencias morfológicas entre el maíz y elteocintle (Doebley y Stec, 1991; Doebley,1993; Paterson et al., 1995).
La frecuencia de quiasmas totales (cuadro1) obtenida para maíz híbrido simple H-311fue de 21.88 y 21.02 y de 23.31 y 21.63 parateocintle chalqueño en metafase I ydiacinesis, respectivamente. Estos datosconcuerdan con los rangos estimados parala frecuencia de quiasmas en Zea mays quevan de 17.4 a 25.0 por célula (Nils-Otto etal., 1993).
El conteo del número de quiasmas se hizobasándose en la interpretación dada porDarlington (1934) y Ward (1976, 1979) para laconfiguración de cada bivalente en lascélulas de cada planta analizada. Se contó elnúmero de quiasmas intercalares y distalesy el total por bivalente. De la suma del númerode quiasmas de los 10 bivalentes se obtuvosu frecuencia por célula.
Análisis estadístico
Con el fin de determinar si había diferenciasestadísticamente significativas entre lasvariedades progenitoras y sus híbridos, asícomo entre la fase de diacinesis y metafaseI, y tomando en cuenta que los datos sonindependientes y se distribuyen de maneranormal, se llevó a cabo un análisis devarianza para dos factores, los cuales fueronla variedad y la fase, con una variable derespuesta, el número de quiasmas (Duránet al., 2005). Para determinar cuál o cuálesmedias diferían, se aplicó una prueba decomparación múltiple de medias (prueba deTukey) (Socal y Rohlf, 1995).
Considerando que el número total dequiasmas lo determina la suma de quiasmasdistales e intercalares, se utilizó un análisisde correlación simple para determinar sihabía alguna diferencia en la asociación entreel número total de quiasmas con losintercalares y distales, las cuales son variablesaleatorias.
Para determinar si había diferenciasestadísticamente significativas entre lalongitud total de cromatina entre los trestipos de plantas así como entre la longitudtotal de cada uno de los 10 cromosomasmitóticos, con sus respectivos homólogos,se realizó un análisis de varianza para cadauno de los 10 cromosomas teniendo como

52
Diciembre 2005Núm. 20:47-72
Cuadro 1. Promedio y desviación estándar para el número de quiasmas por célula endiacinesis y metafase I meiótica. Datos obtenidos de 200 células observadas.
A pesar de que el promedio en teocintlechalqueño (5.36) para los quiasmasintercalares en la fase de diacinesis fuemayor a los del maíz híbrido simple H-311(4.26) y mientras que en los quiasmasdistales, en la misma fase, el promedio paralas plantas de maíz híbrido simple H-311(16.76) fue superior al teocintle chalqueño(16.27), la prueba de Tukey (comparaciónmúltiple de medias) señaló que en esta faselas variedades progenitoras no mostraronvariación significativa en la frecuencia dequiasmas intercalares, distales y quiasmastotales (cuadro 3).
Varios estudios han demostrado que loscambios en la frecuencia de entrecruzamientosson debidos a alteraciones en la estructuragenética (Nils-Otto et al., 1993; Doebley etal., 1995). En ambas fases, siendo los valoresde la frecuencia de quiasmas totales
y distales tan similares en teocintlechalqueño y maíz híbrido simple H-311(cuadro 3), podría esperarse que laconstitución de los genes que regulan elentrecruzamiento fuera muy parecidadebido a la estrecha relación filogenéticaentre ambas variedades, no obstante, lasdiferencias numéricas en la frecuencia dequiasmas intercalares, no desde el punto devista estadístico, podría estar dada porpequeñas modificaciones en la estructuragenética que se ha venido dando a travésdel tiempo en cada una de las dosvariedades, por la selección natural en elteocintle chalqueño y por la mano delhombre en el maíz híbrido simple H-311 alir seleccionando aquellas característicasacordes a sus necesidades.
En la metafase I, los quiasmas totales ein tercalares tuvieron di ferencias
__
FASE x ± SD Metafase I Diacinesis Metafase I Diacinesis Metafase I Diacinesis
VARIEDAD
Quiasmas totales Quiasmas intercalares Quiasmas distales Maíz híbrido simple H-311
21.88 ± 0.7 21.02 ±1.8 5.84 ± 1.40 4.26 ± 1.9 16.12 ± 0.6 16.76 ± 1.17
Teocintle chalqueño
23.31 ± 1.8 21.63 ± 2.4 7.1 ± 1.3 5.36 ± 1.9 16.20 ± 0.93 16.27 ± 1.0
Maíz híbrido simple H-311 X Teocintle chalqueño
16.63 ± 1.0
16.90 ± 0.4
1.71 ± 0.7
1.69 ± 0.4
14.9 ± 0.94
15.21 ± 0.34
Teocintle chalqueño X Maíz híbrido simple H-311
16.20 ± 0.5
16.69 ± 1.1
1.48 ± 0.42
1.65 ± 0.62
14.72 ± 0.42
15.02 ± 0.88

53
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
Cua
dro
2. A
náli
sis
de la
var
ianz
a pa
ra e
l núm
ero
de q
uias
mas
en
dife
rent
e po
sici
ón e
n la
s va
ried
ades
de
Zea
may
s ss
p. m
ays
(híb
rido
sim
ple
H-3
11),
Zea
may
s ss
p. m
exic
ana
(teo
cint
le c
halq
ueño
) y la
F1
(Zea
may
s ss
p. m
ays
X Z
ea m
ays
ssp.
mex
ican
a). *
Dif
eren
cia
sign
ific
ativ
a, a
=0
.05
.

54
Diciembre 2005Núm. 20:47-72C
uadr
o 3.
Pru
eba
de T
ukey
(DM
SH),
com
para
ción
múl
tipl
e de
las
med
ias
de la
frec
uenc
ia d
e qu
iasm
as to
tale
s (Q
t), i
nter
cala
res
(Qi)
y d
ista
les
(Qd)
ent
re la
s va
ried
ades
(let
ras)
y e
n m
etaf
ase
I y d
iaci
nesi
s.
*Dif
eren
cia
sign
ific
ativ
a, a
=0
.05
.

55
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
Fig. 2. Diferentes configuraciones en bivalentes (metafse I) determinados por el númeroy posición de quiasmas. A) Zea mays ssp. mays; B) Zea mays ssp. mexicana; C y D) F1
(Zea mays ssp. mays X Zea mays ssp. mexicana) (80X, campo claro A y B; contraste de fases C y D).
A
B
C
D

56
Diciembre 2005Núm. 20:47-72
estadísticamente significativas en lasvariedades de teocintle chalqueño (23.31y 7.10) comparando con la frecuenciaobtenida del maíz híbrido simple H-311(21.88 y 5.84), respectivamente (cuadro 3,fig. 2), mientras que en los quiasmasdistales no hubo diferencias significativas,lo cual indicó que la diferencia en losquiasmas totales entre el maíz híbridosimple H-311 y el teocintle chalqueño estádada por los quiasmas intercalares.Posiblemen te, el incremento en lafrecuencia de quiasmas intercalares enmetafase I del teocintle chalqueño conrespecto a la diacinesis, podría ser debidoa errores en la interpretación de losquiasmas, pues en la metafase I incrementala condensación de los cromosomasreduciendo la resolución citológica.
Las variedades híbridas mostraron unafrecuencia muy similar entre sí en losquiasmas totales, intercalares y distalestanto en la fase de diacinesis como en lametafase I (cuadros 1 y 2), con base en losresultados obtenidos del análisis de varianzase procedió aplicar la prueba de Tukey, quepermitió observar que no existe diferenciaestadísticamente significativa en las mediasde los tres tipos de la frecuencia de quiasmasentre el maíz híbrido simple H-311 Xteocintle chalqueño y teocintle chalqueñoX maíz híbrido simple H-311 (cuadro 3).
Al comparar la frecuencia de quiasmasintercalares entre progenitores y lasvariedades híbridas, se observó unadisminución significativa en éstas. Tomandocomo el 100% la frecuencia de quiasmasintercalares de los progenitores, hubo unadisminución en los híbridos de 70.92 %. Porotro lado, en los quiasmas distales hubo unadisminución de aproximadamente 6% conrespecto a los progenitores.
Fue muy notorio que los quiasmas intercalaressufrieran una mayor disminución que losquiasmas distales. Resultados muy similareshan sido obtenidos en trabajos de hibridizaciónde maíz con teocintle, por ejemplo: Kato (1984)observó en la fase de diacinesis en híbridosmaíz X teocintle Durango, maíz X teocintleFlorida una gran proporción de configuracionesen forma de aro, indicando que estabanpresentes quiasmas en la parte terminal de lascromátidas.
Esto sugiere que la zona distal de loscromosomas es menos susceptible amodificaciones en su estructura genéticaque la zona intercalar de un cromosoma, obien, que es más factible el apareamientode zonas homólogas localizadas en la regióndistal de cromosomas diferentes, quecuando dichas regiones homólogas estánen posiciones intercalares.
Al analizar la frecuencia de quiasmas en laF1 del maíz híbrido simple H-311 X teocintlechalqueño y de la F1 del teocintle chalqueñoX maíz híbrido simple H-311 en diacinesis ymetafase I, no se encontraron diferenciassignificativas, por tanto, se observó queno hubo terminalización, que es elmovimiento de los quiasmas desde el sitiodonde se originaron en un cromosoma hacialos extremos del mismo, pues en metafase Ino se vieron incrementados los quiasmasdistales o disminuidos los intercalares(cuadros 2 y 3 ). De lo contrario, si hubieraterminalización, la frecuencia total dequiasmas se ve reducida en metafase I conrespecto a diacinesis, y los quiasmasintercalares disminuyen y aumentan losdistales de una fase a otra. Los resultadosdel presente estudio concuerdan con losobtenidos en los trabajos de Ward (1976,1979) y Nils-Otto et al. (1993), ellosmencionaron que no hay evidencias de

57
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
terminalización en maíz al no encontrardiferencias significativas en la frecuenciade quiasmas entre diacinesis y metafase I.Al realizar el análisis de correlación simple,para observar qué tipo de quiasmas(intercalares o distales) afectan princi-palmente la frecuencia total, se obtuvo uníndice de correlación de 0.65 entre losdistales contra los totales, lo cual indicó quesí hay correlación entre estos dos tipos dequiasmas. Sin embargo, los quiasmasdistales no modifican a los totales tannotoriamente como los intercalares, pues lacorrelación fue más alta entre la frecuenciade quiasmas intercalares contra totales,siendo de 0.93 su índice de correlación, loque indica que al incrementar o disminuirlos quiasmas intercalares se ven afectadosde la misma manera los quiasmas totales(cuadro 1).
Apareamiento cromosómico
En las variedades híbridas, el apareamientoentre los cromosomas homólogos no fueregular, estuvieron presentes univalentes,trivalentes, tetravalentes, pentavalentes yhexavalentes en un 41% de las célulasexaminadas (fig. 3). Estas configuracionescromosómicas afectan tanto la regularidadde la meiosis como la fertilidad de losgametos que se producen y al formarsegametos no viables se observa disminuidanotoriamente la probabilidad de que seestablezca en la naturaleza una variedadhíbrida con estas características. En el 59%restante de las células examinadas, sepresentaron 10 bivalentes, de los cualesalgunos presentaban como máximo dosquiasmas y otros estaban unidos sólo porun extremo (fig. 4 A).
Como se observa en el cuadro 4 laproporción de univalentes y trivalentes no
fue muy semejante en el híbrido (maízhíbrido simple H-311 X teocintle chalqueño)y el híbrido (teocintle chalqueño X maízhíbrido simple H-311). En este último caso,se presentó un mayor número de estasasociaciones cromosómicas. La presenciade los univalentes es atribuible a la falta dehomología entre algunos cromosomashomólogos no apareados, probablementepor la falta o en la diferente distribuciónde segmentos homólogos en talescromosomas. Desafortunadamente, por elgrado de compactación de los cromosomasen diacinesis y metafase I no es fácil saberqué cromosomas son. Sin embargo, no sedescarta el hecho de que el maíz híbridosimple H-311, utilizado para este estudio,haya contribuido genéticamente a la faltade apareamiento entre sus cromosomas ylos correspondientes homólogos enteocintle chalqueño, pues en célulasmeióticas de cuatro plantas de maíz essabido que el apareamiento entrehomólogos está controlado por genes enlos mismos cromosomas y una mutaciónde estos loci puede ocasionar una parcial ototal inhibición de este evento en algunosindividuos (Doebley y Iltis, 1980; González-Sánchez et al., 2003).
La formación de univalentes significa queal menos un cromosoma tiene regioneshomólogas con otros cromosomas nohomólogos (Darlington, 1934). En estecaso los t r ivalentes, tetr avalentes,pentavalentes y hexavalentes sonindicadores de translocaciones enel genomio de una o las dos plantas en eltranscurso del tiempo. Las translocacionesque más favorecen las configuracionesmultivalentes son las terminales, puespermiten la formación de las cadenas o aros(Aguiar-Perecin y Vosa, 1985) que fue locomúnmente observado en este estudio.

58
Diciembre 2005Núm. 20:47-72
Fig. 3. Células meióticas de la F1 (Zea mays ssp. mays X Zea mays ssp. mexicana).En diacinesis con 7 bivalentes y un hexavalente (1000X, contraste de fases
A y campo claro B).
Fig. 4. Cromosomas en diacinesis, diez pares bivalentes. Las flechas señalan alcromosoma 6 unido al nucleolo. A) Zea mays ssp. mays; B) Zea mays ssp. mexicana;
C y D) Zea mays ssp. mays X Zea mays ssp. mexicana (400X, campo claro A y B;contraste de fases C y D).

59
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
La formación de trivalentes pudo haber sidomenor que la de otras configuraciones,principalmente por la competencia deapareamiento entre bivalentes homólogos.Los resultados de este trabajo contrastan conlos obtenidos por Kato (1984), este autormenciona que el apareo de los cromosomasdel teocintle Chalco con los cromosomas delmaíz es completo.
El apareamiento irregular entre teocintlechalqueño y el maíz híbrido simple H-311utilizado, mostró falta de homologíaestructural entre algunos cromosomasdebido, por un lado, a translocaciones queprovocan que porciones de los cromosomasdel maíz tengan sus contrapartes encromosomas no homólogos del teocintle, ypor otro lado tal vez a inversiones, las cualesprevienen la formación de quiasmas entrealgunos segmentos de cromosomashomólogos. Esto puede interpretarse comoun mecanismo que tiene un importantepapel evolutivo porque reduce elentrecruzamiento, sirviendo como unmecanismo de aislamiento parcial entre maízhíbrido simple H-311 y teocintle chalqueño.
Cariotipo
Se analizaron 10 células en metafase mitóticatanto para el maíz híbrido simple H-311, elteocintle chalqueño y el híbrido resultante(fig. 6). Como es usual, los cromosomasfueron numerados en orden decreciente desu longitud, de tal manera que el cromosomamás pequeño es el número 10 en el cariotipode Zea (figs. 7A, 7B y 7C).
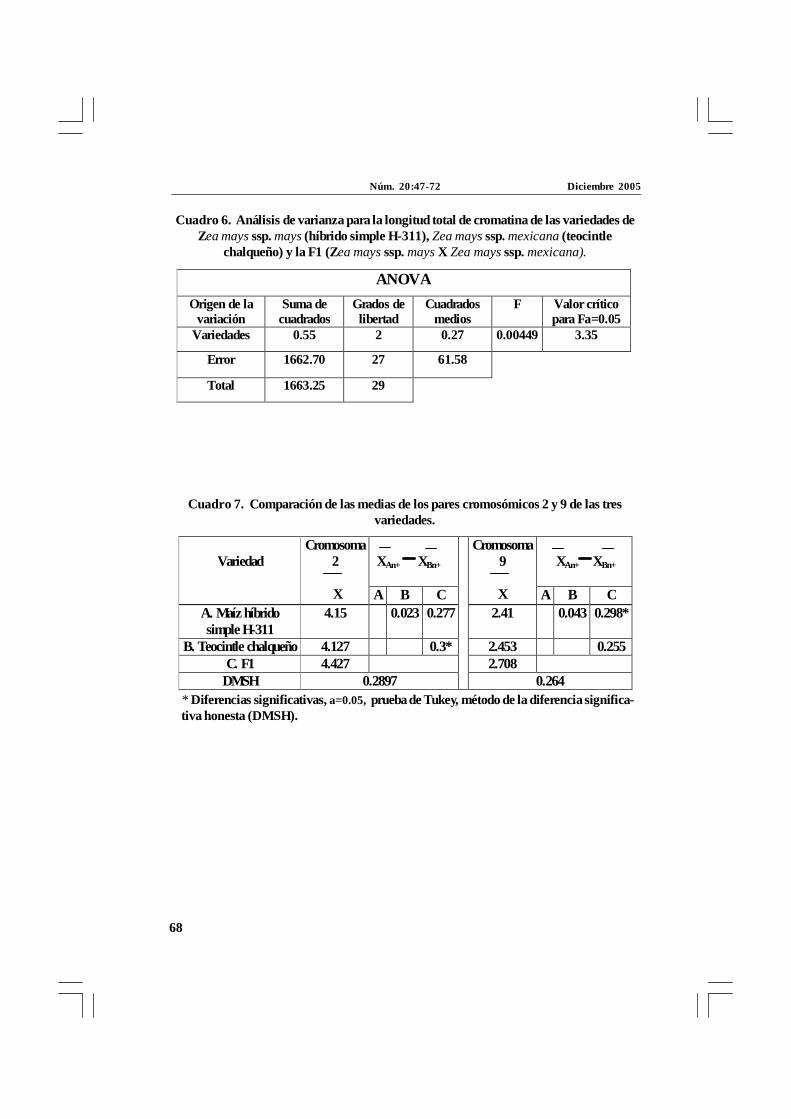
Los datos del análisis estadístico para loscromosomas del 1 al 10 y de la cromatina totalaparecen en los cuadros 5 y 6 de ANOVA,respectivamente. Se observa en estoscuadros que el valor de F calculado y el valorestablecido por la F de tablas nos indica queno hay diferencias significativas entre lasmedias de la longitud total de los cromosomashomólogos entre las tres variedades, exceptolos cromosomas 2 y 9 que presentarondiferencias significativas, y en la com-paración de las medias por la prueba deTukey se observó una separación de losprogenitores con el híbrido F1 (cuadro 7).Así, en el par cromosómico 2 hubo unadiferencia significativa entre teocintlechalqueño y el híbrido F1 y en el parcromosómico 9 esta diferencia se dio entre el
Cuadro 4. Frecuencias de distintas configuraciones cromosómicas en la F1.Datos obtenidos de 200 células observadas.
Maíz híbrido X teocintle
Simple H-311 chalqueño
Teocintle X maíz híbrido
Chalqueño simple H-311
METAFASE DIACINESIS METAFASE DIACINESIS Univalentes 17 18 25 38 Trivalentes 0 4 18 19
Tetravalentes 31 31 35 33 Pentavalentes 0 0 1 1 Hexavalentes 4 1 4 1

60
Diciembre 2005Núm. 20:47-72
maíz híbrido simple H-311 con híbrido F1. Porotra parte, también se determinó que noexisten diferencias significativas entre lasmedias de la longitud total de sus genomios(cuadro 6, figs. 6 y 7).
En todas las plantas de las tres variedadesanalizadas se observó el número somático2n = 2x = 20. El tamaño de los cromosomasen los tres tipos de plantas resultó ser muysimilar (cuadros 8, 9 y 10). La longitud totalde los cromosomas fue de 2.26 a 4.76, 2.22 a4.64 y de 2.11 a 4.59 µm para el maíz híbridosimple H-311, el teocintle chalqueño y la F1,respectivamente (fig. 7). Estos datosconcuerdan con lo reportado porVishnyakov y Kometiani (1988) y Chen(1969). Ellos reportan una longitud total delos cromosomas de maíz entre 2 y 5 µm.
La longitud total de cromatina por genomiose obtuvo mediante la suma de laslongitudes de los 10 cromosomas en cadacariotipo, se observó una longitud muysimilar para los tres cariotipos. La longitudmáxima total se observó en el maíz híbrido
Fig. 5. Célula en diacinesis de Zea mays ssp. mays (híbrido simple H-311) con nuevebivalentes y dos univalentes (400X).
simple H-311 siendo de 33.51 µm y la mínimapara el teocintle chalqueño con una longitudde 33.18 µm. La longitud de cromatina totalpara la F1 (maíz híbrido simple H-311 Xteocintle chalqueño) fue intermedia con unalongitud de 33.33 µm (cuadros 8, 9 y 10).
Estadísticamente la longitud total de los tresgenomios, así como la longitud total de cadauno de los 10 cromosomas que integran elgenomio de estas plantas no presentandiferencias significativas, es decir son muysimilares; aunado a esto, a pesar de habersepresentado univalentes, tr ivalentes,tetravalentes y hexavalentes en un 59% delas células analizadas, se observaron 10bivalentes aun cuando la frecuencia dequiasmas se observó reducida a sólo uno odos quiasmas por bivalente. Esta homologíaentre los cromosomas del teocintlechalqueño y del maíz híbrido simple H-311indica que estas dos variedades tienen unaestrecha relación filogenética, y dado queel teocintle presenta característicasmorfológicas más primitivas como son ladesarticulación de sus semillas para

61
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
Fig. 6. Células radiculares en metafase, 2n = 20. Las flechas señalan la posición delsatélite en el par 6. A) Zea mays ssp. mays. mays; B) Zea mays ssp. mexicana;
C) F1 (Zea mays ssp. mays X Zea mays ssp. mexicana); (1000X, campo claro A y B;contraste de fases C). Barra = 6 µm

62
Diciembre 2005Núm. 20:47-72
Fig. 7. Cariogramas. A) Zea mays ssp. mays (maíz híbrido simple H-311); B) Zea mays ssp.mexicana (teocintle chalqueño); C) F1 (Zea mays ssp. mays X Zea mays ssp. mexicana).
A cromosomas
micrómetros
B cromosomas
micrómetros
C cromosomas
micrómetros

63
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
dispersarse por sí sola y la pequeña masade sus semillas que les permite penetrarfácilmente en el suelo (Paterson et al.,1995), y como lo mencionan diversosinvestigadores, éste es probablemente elprogenitor del maíz.
La relación de brazos es muy importantepara poder clasificar los cromosomas yobtener el cariotipo completo (Levan, et al.,1964). Basándose en las mediciones de lalongitud de los cromosomas se calculó larelación de brazos (BL/BC) (cuadros 8, 9 y10). Estos valores concuerdan con losobtenidos en metafase mitótica por Chen(1969) y Aguiar-Perecin y Vosa (1985).
Comparando las posiciones del centrómerode cada uno de los cromosomas con susrespectivos homólogos en los tres tipos devariedades, se observó lo siguiente:solamente los cromosomas 1, 2, 6 y 8presentan la misma nomenclatura para laposición del centrómero (m, m, sm y sm,respectivamente) (cuadro 11). En los otrospares cromosómicos, varía de metacéntricoa submetacéntr ico. En los trabajosanteriormente citados obtuvieron unavariación similar entre variedades de maíz.Aguiar-Perecin y Vosa (1985) en su estudiode bandeo específico para nudoscromosómicos obtuvieron diferencias en larelación de brazos del cromosoma 2 en elcual era heterocigótico para un nudomediano en el brazo largo. El cromosoma connudo mediano tuvo una relación de brazos de1.67 ± 0.16, mientras que el homólogo sin nudotuvo una relación de 1.24± 0.13, mencionandoque estos datos demuestran que los nudosmedianos y grandes afectan la longitud de losbrazos de los cromosomas mitóticos, estopodría explicar las diferencias significativasencontradas en los pares cromosómicos 2 y9, además, explica la variación en la relación
de brazos y por tanto la variación en lanomenclatura del centrómero para loscromosomas 3, 4, 5, 7, 9 y 10 del maíz híbridosimple H-311 con respecto a sus homólogosen el teocintle chalqueño y la F1 (maíz híbridosimple H-311 X teocintle chalqueño), noolvidando que en algunas ocasiones puededeberse al grado de compactación diferencialde los cromosomas como una respuestadiferente de las plantas a los reactivosutilizados en la técnica citogenética o bien poruna pequeña diferencia en la etapa mitóticaen que se encon traban las célulasmeristemáticas radiculares en el momento delpretratamiento y fijación para su observacióncitológica.
Sin embargo, las diferencias en la relación debrazos entre los cromosomas mitóticos noson tan marcadas como cuando se comparala posición del centrómero de cromosomasen metafase mitótica con cromosomas enpaquitene. Para estos últimos se danposiciones subterminales para loscromosomas 6 y 8 del maíz y teocintle (Kato,1984). Los pares cromosómicos 6 y 8 de lastres variedades aquí estudiadas, resultaronsubmetacéntricos, y con la presencia desatélites sólo en el par 6, estos resultadoscoinciden con Chen (1969), Aguiar-Perecin yVosa (1985) (figs. 6 y 7).
Por lo anteriormente citado, y al menosrefiriéndonos a las características visiblesdel genomio (número cromosómico,longitud cromosómica, posición delcentrómero y constricciones secundarias)del maíz híbrido simple H-311 y del teocintlechalqueño, se observó que estas variedadesno han mostrado cambios evolutivossignificativos.

64
Diciembre 2005Núm. 20:47-72
CONCLUSIONES
Los cromosomas mitóticos en los tres tiposde genomios presentaron una longitud queva de 2 a 5 µm, confirmando datos reportadospor Chen (1969) y Aguiar-Perecin y Vosa(1985).
El cariotipo mitótico del maíz híbridosimple H-311, del teocintle chalqueñoy de la F1 está constituido únicamentepor cr om osom a s m et a cén t r i cos ysubmetacén tr icos. Presentándose lossatélites en el cromosoma 6.
En la fase de diacinesis el apareamiento enla F1 no fue regular, mostrando univalentes,trivalentes, tetravalentes, pentavalentes yhexavalentes.
Existen «probables» diferenciasestructurales entre los cromosomas del maízhíbrido simple H-311 con sus respectivoshomólogos en el teocintle chalqueño.
La frecuencia de quiasmas intercalares enla F1 disminuye más notoriamente en losdistales, tanto en metafase I, meiótica ydiacinesis.
La frecuencia de quiasmas totales se veafectada directamente por el aumento odisminución de los quiasmas intercalares enambas fases.
En los híbridos F1 (maíz híbrido simple H -311 X teocintle chalqueño y teocintlechalqueño X maíz híbrido simple H –311)no hubo diferencias estadísticamentesignificativas en la frecuencia de quiasmasentre la diacinesis y metafase I, indicandoque no hay terminalización.
El maíz híbrido simple H-311 y el teocintlechalqueño presentan una estrecha relaciónfilogenética, dado que tienen el mismonúmero cromosómico, longitud y relaciónde brazos cromosómicos similares, ademásexhiben apareamiento e intercambiogenético entre los cromosomas de ambasvariedades cuando se encuentran en elhíbrido F1 (maíz híbrido simple H-311 Xteocintle chalqueño).
AGRADECIMIENTOS
Deseamos manifestar un sinceroagradecimiento al doctor Diódoro GranadosSánchez por la r evisión crí tica delmanuscrito, y a los M. en C. Agustín VargasV. , Antonio E. Cisneros C. y Edgar Urrutiaen la revisión del trabajo estadístico.
LITERATURA CITADA
Aguiar-Perecin, M .L. R. y C. G. Vosa, 1985. C-Banding in maize II. Identification ofsomatic chromosomes. Heredity, 54: 37-42.
Aguirre, G. C. D. y T. A. Y. Kato, 1979. Com-petencia entre el polen de maíz y deteocintle durante la fecundación.Agrociencia, 37: 109-121.
Coe, H. E., 2001. The origins of maizegenetics. Nat. Rev. Genet., 2: 898-905.
Chen, C.C., 1969. The somatic chromosomes ofmaize. Can. J. Genet. Cyt., 11: 752-754.

65
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
Darlington, C.D., 1934. The origin andbehavior of chiasmata, VII. Zea mays.Z.Verebungsl, 67:96-114.
Doebley, J.F. y H.H. Iltis, 1980. Taxonomy ofZea (gramínea). I. A. subgenericclassification with key to taxa. Am. J.Bot., 67: 982-993.
Doebley, J.F. y A. Stec, 1991. Geneticanalysis of the morphologicaldifferences between Maize andTeosinte. Genetics, 129: 285-295.
Doebley, J.F., 1993. Inheritence of themorphological differences betweenmaize and teosinte: comparison ofresults for two F2 populations.Genetics, 134: 559-570.
Doebley, J.F., A. Stec y Ch. Gustus, 1995.Teosinte branchedl and the origin ofmaize: evidence for epistasis and theevolution of dominance. Genetics,141: 333-346.
Durán, D. A., A.E. C. Cisneros y A. V. Vargas,2005. Bioestadística. Facultad de Es-tudios Superiores Iztacala, UniversidadNacional Autónoma de México, Méxi-co. 222 pp.
García, V.A., 1988. Técnicas y procedimien-tos de citogenética vegetal.3ª ed.Universidad Autónoma de Chapingo.Chapingo, México. 144 pp.
González-Sánchez, M., E. González-González,F. Molina, A.M. Chiavarino, M. Rosatoy M. J. Puerta, 2003. One gene determi-nes maize B chromosome accumulationby preferential fertilisation; anothergene(s) determines their meiotic loss.Heredity, 90: 122-129.
Gourmet, C. y A. L. Rayburn, 1996. Identificationof RAPD markers associated with thepresence of B chromosomes in maize.Heredity, 77:240-244.
Goodnight, J. Ch., 2000. Quantitative trait lociand gene interaction: the quantitativegenetics of metapopulations. Heredity,84: 587-598.
Jugenheimer, W. R., 1990. Maíz: Varieda-des mejoradas Métodos de Cultivo yProducción de Semilla. Trad. R PiñaG, Cuarta reimpresión, Limusa, Méxi-co. 841 pp.
Kato, Y.T.A., 1984. Chromosomemorphology and the origin of maize andits races. Evol. Biol., 17: 219-253.
Levan, A., K. Fredga y A.A. Sandberg., 1964.«Nomenclature for centromericposition on chromosomes».Hereditas, 52: 201-219.
Nils-Otto, N., T. Säll y B. O. Bengtsson, 1993.Chiasma and recombination data inplants: are they compatible?. Trendsgenet. 9: 334-338.
Pääbo, S. , 1999. Neol ith ic genet icengineering. Nature, 398: 194-195.
Paterson, A.H., Y. Lin, Z. Li, K. F. Scherz, F.Doebley, Sh. R.M. Pinson, S. Liu, J.W.Stansel, J.E. Iruine, 1995. Convergentdomestication of cereak crops byindependent mutations at correspondinggenetic loci. Science, 269: 1714-1718.

66
Diciembre 2005Núm. 20:47-72
Pressoir, G. y J. Berthaud, 2004. Patterns ofpopulation structure in maize landracesfrom the Central Valleys of Oaxaca inMexico. Heredity, 92: 88-94.
Sokal, R. R. y F. J. Rohlf, 1995. Biometry. Theprinciples and practice of statistics inbiological research. 3a ed. W. H.Freeman and company. Nueva York.887 pp.
Stewart, C. N., M. D. Halfhill y S. I. Warwick,2003. Genetic modifications: transgeneintrogression from genetically modifiedcrops to their wild relatives. Nat. Rev.Genet., 4: 806-817.
Ward, E.J., 1976. The effect of accessorychromatin on chiasma distribution inmaize. Can. J. Genet. Cyt., 18: 479-484.
Ward, E.J., 1979. Chiasma frecuency anddistribution in maize family segregationfor K10 and trisomy 10. Genetics, 92:223-230.
Vishnyakov, D.A. y D.G. Kometiani., 1988.Karyotipic characterization of someplants of southern regions of URSS.Sov. Agric. Sci., 6: 13-16.

67
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
Cua
dro
5. A
náli
sis
esta
díst
ico
para
la lo
ngit
ud to
tal d
e lo
s cr
omos
omas
de
las
vari
edad
es d
e Z
ea m
ays
ssp.
may
s (m
aíz
híbr
ido
sim
ple
H-3
11),
Zea
may
s ss
p. m
exic
ana
(teo
cint
le c
halq
ueño
) y la
F1
(Zea
may
s ss
p. m
ays
X Z
ea m
ays
ssp.
mex
ican
a).
*Dif
eren
cia
sign
ific
ativ
a, a
= 0
.05.
68
Diciembre 2005Núm. 20:47-72
Cuadro 6. Análisis de varianza para la longitud total de cromatina de las variedades deZea mays ssp. mays (híbrido simple H-311), Zea mays ssp. mexicana (teocintle
chalqueño) y la F1 (Zea mays ssp. mays X Zea mays ssp. mexicana).
Cuadro 7. Comparación de las medias de los pares cromosómicos 2 y 9 de las tresvariedades.
* Diferencias significativas, a=0.05, prueba de Tukey, método de la diferencia significa-tiva honesta (DMSH).
ANOVA
Origen de la variación
Suma de cuadrados
Grados de libertad
Cuadrados medios
F Valor crítico para Fa=0.05
Variedades 0.55 2 0.27 0.00449 3.35
Error 1662.70 27 61.58
Total 1663.25 29
XAn+ XBn+
XAn+ XBn+
Variedad
Cromosoma 2
X A B C
Cromosoma 9
X A B C A. Maíz híbrido simple H-311
4.15 0.023 0.277 2.41 0.043 0.298*
B. Teocintle chalqueño 4.127 0.3* 2.453 0.255 C. F1 4.427 2.708
DMSH 0.2897
0.264

69
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
Cua
dro
8. C
arac
terí
stic
as d
el c
ario
tipo
mitó
tico
prom
edio
(2n
= 2
0) d
e Ze
a m
ays
ssp.
may
s (hí
brid
o si
mpl
e H
-311
). L
os v
alor
essu
peri
ores
son
los l
ímite
s de
vari
ació
n m
ínim
a y
máx
ima,
y lo
s inf
erio
res s
on e
l val
or p
rom
edio
de
los
dato
s ob
teni
dos d
e 10
cél
ulas
.
Valor crítico para Fa=0.05
Bn+
C 0.298*
0.255

70
Diciembre 2005Núm. 20:47-72C
uadr
o 9.
Car
acte
ríst
icas
del
car
iotip
o m
itótic
o pr
omed
io (2
n =2
0) d
e Ze
a m
ays s
sp. m
exic
ana
(teo
cint
le c
halq
ueño
). L
os v
alor
essu
peri
ores
son
los l
ímite
s de
vari
ació
n m
ínim
a y
máx
ima,
y lo
s inf
erio
res
son
el v
alor
pro
med
io d
e lo
s dat
os o
bten
idos
de
10 c
élul
as.

71
Flores et al. Estudio citogenético del maíz híbrido simple H-311, teocintle chalqueño y su híbrido F1
Cua
dro
10.
Car
acte
ríst
icas
del
car
iotip
o m
itótic
o pr
omed
io (2
n =
20) d
e la
F1
(Zea
may
s ssp
. may
s X Z
ea m
ays s
sp. m
exic
ana.
Los
val
ores
supe
rior
es so
n lo
s lím
ites d
e var
iaci
ón m
ínim
a y
máx
ima,
y lo
s inf
erio
res s
on e
l val
or p
rom
edio
de
los d
atos
obt
enid
os d
e 10
cél
ulas
.

72
Diciembre 2005Núm. 20:47-72
Cuadro 11. Posición del centrómero y satélites en cromosomas de maíz híbrido H-311,teocintle chalqueño y la F1.
*Presencia de satélites: m, metacéntrico; sm, submetacéntrico; M, metacéntrico(posición estrictamente media) (Levan et al., 1964).
Cromosoma
Maíz híbrido simple h-311
Teocintle chalqueño
F1
1 m m m 2 m m m 3 m m sm 4 m m sm 5 m m M 6 sm* sm* sm* 7 sm m M 8 sm sm sm 9 sm m m
10 sm m m





























