Embed Size (px)
Citation preview

Epigenetics and Developmental Origins of Health and Disease
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Aim
bull To highlight important issues relating to the epidemiological investigation of epigenetic mechanisms in the context of developmental origins of health and disease
Overview bull Change over time in the epigenome
bull Evidence for the influence of early life exposures on the epigenome
bull Inter-generational exposure versus trans-generational effects
bull Persistent versus transient epigenetic change
bull Temporal relationships
bull The problem of confounding in a DOHaD context
Epigenetic mechanisms and developmental programming
Many diseases of maturity have their origins early in life
Early development
Stroke
Obesity
Diabetes mellitus
Hypertension
Rheumatoid arthritis
Ischaemic heart disease
The dynamic epigenome
Germline epimutation
Parental genomic demethylation Epigenetic drift somatic epimutation
Developmental epigenetic programming
Waterland RA Nutr Rev 2008
bull 3 gene loci analysed (DRD4 SERT MAOA)
bull 46 MZ twin pairs
bull 45 DZ twin pairs
bull Total n = 182
bull Sampled at 5 and 10 years
bull [Modest] differences observed between genetically identical individuals
bull Variation not consistent across all loci
Age 5 Age 10
Age-related change in methylation
Manhattan plot showing association between methylation at individual CpG sites and chronological age Plotted are P-values indicating strength of association between DNA methylation levels at gt27 000 CpG sites and age in cerebellum (purple) frontal cortex (green) pons (blue) and temporal cortex (red) For each point a positive association between DNA methylation and chronological age is indicated by upward pointing triangles a negative association is indicated by downward pointing triangles
Note p-values give no indication of magnitude of change
Hernandez DG et al Hum Mol Genet 2011
Studies linking early life exposures to changes in DNA methylation using animal models
Early life exposure Animal
model Epigenetic change
Disease
association
Maternal nutrition
Low Protein Rat Mouse
Pig
and DNA methylation and histone
acetylation and histone methylation Obesity
Calorie restriction Sheep Rat DNA methylation histone acetylation and
histone methylation
Obesity
Diabetes
Periconceptional restriction B12
folate methionine Sheep Altered DNA methylation Obesity
High fat Macaque
Mouse
and DNA methylation and histone
acetylation and and histone methylation Obesity
Surgical models
IUGR ( uterine artery ligation) Rat Altered DNA methylation histone acetylation Diabetes
Environmental toxin
Arsenic Mouse DNA methylation Diabetes
Paternal effect
Low protein Mouse DNA methylation Obesity
Neonatal diet
Leptin treatment Rat DNA methylation Obesity
Extendin-4 Rat Hyperacetylation Diabetes
Reversal with folic acid
Methyl supplementation Avy mouse DNA methylation Obesity
Genistein supplementation +FA Avy mouse DNA methylation Obesity
Protein restriction + FA Rat Prevented or reversed hypomethylation Obesity
Seki Y et al Endocrinology 2012
The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Aim
bull To highlight important issues relating to the epidemiological investigation of epigenetic mechanisms in the context of developmental origins of health and disease
Overview bull Change over time in the epigenome
bull Evidence for the influence of early life exposures on the epigenome
bull Inter-generational exposure versus trans-generational effects
bull Persistent versus transient epigenetic change
bull Temporal relationships
bull The problem of confounding in a DOHaD context
Epigenetic mechanisms and developmental programming
Many diseases of maturity have their origins early in life
Early development
Stroke
Obesity
Diabetes mellitus
Hypertension
Rheumatoid arthritis
Ischaemic heart disease
The dynamic epigenome
Germline epimutation
Parental genomic demethylation Epigenetic drift somatic epimutation
Developmental epigenetic programming
Waterland RA Nutr Rev 2008
bull 3 gene loci analysed (DRD4 SERT MAOA)
bull 46 MZ twin pairs
bull 45 DZ twin pairs
bull Total n = 182
bull Sampled at 5 and 10 years
bull [Modest] differences observed between genetically identical individuals
bull Variation not consistent across all loci
Age 5 Age 10
Age-related change in methylation
Manhattan plot showing association between methylation at individual CpG sites and chronological age Plotted are P-values indicating strength of association between DNA methylation levels at gt27 000 CpG sites and age in cerebellum (purple) frontal cortex (green) pons (blue) and temporal cortex (red) For each point a positive association between DNA methylation and chronological age is indicated by upward pointing triangles a negative association is indicated by downward pointing triangles
Note p-values give no indication of magnitude of change
Hernandez DG et al Hum Mol Genet 2011
Studies linking early life exposures to changes in DNA methylation using animal models
Early life exposure Animal
model Epigenetic change
Disease
association
Maternal nutrition
Low Protein Rat Mouse
Pig
and DNA methylation and histone
acetylation and histone methylation Obesity
Calorie restriction Sheep Rat DNA methylation histone acetylation and
histone methylation
Obesity
Diabetes
Periconceptional restriction B12
folate methionine Sheep Altered DNA methylation Obesity
High fat Macaque
Mouse
and DNA methylation and histone
acetylation and and histone methylation Obesity
Surgical models
IUGR ( uterine artery ligation) Rat Altered DNA methylation histone acetylation Diabetes
Environmental toxin
Arsenic Mouse DNA methylation Diabetes
Paternal effect
Low protein Mouse DNA methylation Obesity
Neonatal diet
Leptin treatment Rat DNA methylation Obesity
Extendin-4 Rat Hyperacetylation Diabetes
Reversal with folic acid
Methyl supplementation Avy mouse DNA methylation Obesity
Genistein supplementation +FA Avy mouse DNA methylation Obesity
Protein restriction + FA Rat Prevented or reversed hypomethylation Obesity
Seki Y et al Endocrinology 2012
The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352
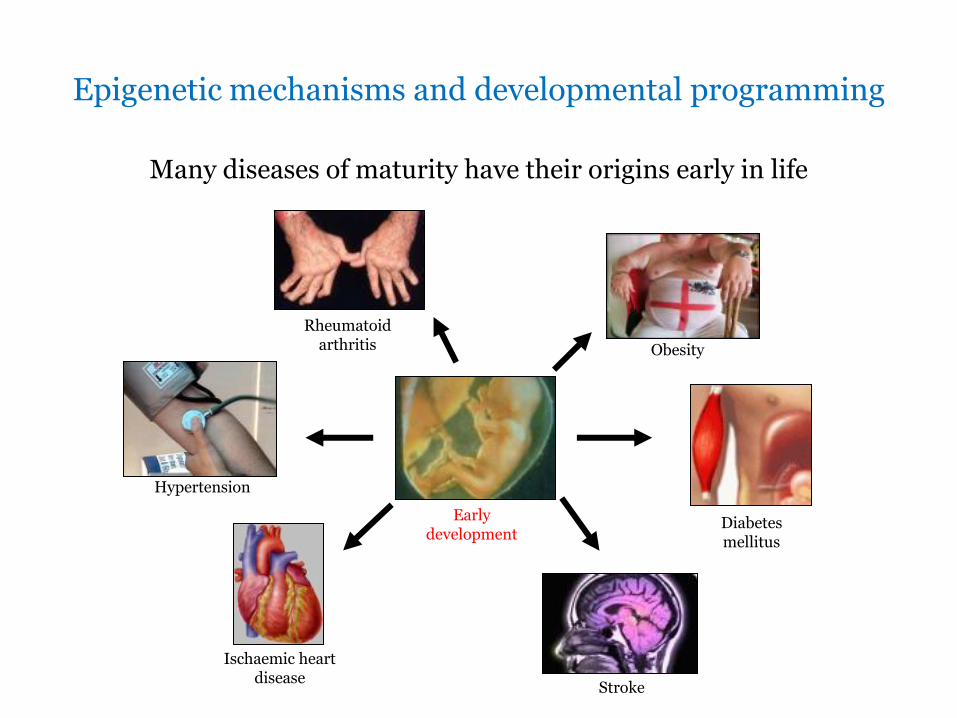
Epigenetic mechanisms and developmental programming
Many diseases of maturity have their origins early in life
Early development
Stroke
Obesity
Diabetes mellitus
Hypertension
Rheumatoid arthritis
Ischaemic heart disease
The dynamic epigenome
Germline epimutation
Parental genomic demethylation Epigenetic drift somatic epimutation
Developmental epigenetic programming
Waterland RA Nutr Rev 2008
bull 3 gene loci analysed (DRD4 SERT MAOA)
bull 46 MZ twin pairs
bull 45 DZ twin pairs
bull Total n = 182
bull Sampled at 5 and 10 years
bull [Modest] differences observed between genetically identical individuals
bull Variation not consistent across all loci
Age 5 Age 10
Age-related change in methylation
Manhattan plot showing association between methylation at individual CpG sites and chronological age Plotted are P-values indicating strength of association between DNA methylation levels at gt27 000 CpG sites and age in cerebellum (purple) frontal cortex (green) pons (blue) and temporal cortex (red) For each point a positive association between DNA methylation and chronological age is indicated by upward pointing triangles a negative association is indicated by downward pointing triangles
Note p-values give no indication of magnitude of change
Hernandez DG et al Hum Mol Genet 2011
Studies linking early life exposures to changes in DNA methylation using animal models
Early life exposure Animal
model Epigenetic change
Disease
association
Maternal nutrition
Low Protein Rat Mouse
Pig
and DNA methylation and histone
acetylation and histone methylation Obesity
Calorie restriction Sheep Rat DNA methylation histone acetylation and
histone methylation
Obesity
Diabetes
Periconceptional restriction B12
folate methionine Sheep Altered DNA methylation Obesity
High fat Macaque
Mouse
and DNA methylation and histone
acetylation and and histone methylation Obesity
Surgical models
IUGR ( uterine artery ligation) Rat Altered DNA methylation histone acetylation Diabetes
Environmental toxin
Arsenic Mouse DNA methylation Diabetes
Paternal effect
Low protein Mouse DNA methylation Obesity
Neonatal diet
Leptin treatment Rat DNA methylation Obesity
Extendin-4 Rat Hyperacetylation Diabetes
Reversal with folic acid
Methyl supplementation Avy mouse DNA methylation Obesity
Genistein supplementation +FA Avy mouse DNA methylation Obesity
Protein restriction + FA Rat Prevented or reversed hypomethylation Obesity
Seki Y et al Endocrinology 2012
The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

The dynamic epigenome
Germline epimutation
Parental genomic demethylation Epigenetic drift somatic epimutation
Developmental epigenetic programming
Waterland RA Nutr Rev 2008
bull 3 gene loci analysed (DRD4 SERT MAOA)
bull 46 MZ twin pairs
bull 45 DZ twin pairs
bull Total n = 182
bull Sampled at 5 and 10 years
bull [Modest] differences observed between genetically identical individuals
bull Variation not consistent across all loci
Age 5 Age 10
Age-related change in methylation
Manhattan plot showing association between methylation at individual CpG sites and chronological age Plotted are P-values indicating strength of association between DNA methylation levels at gt27 000 CpG sites and age in cerebellum (purple) frontal cortex (green) pons (blue) and temporal cortex (red) For each point a positive association between DNA methylation and chronological age is indicated by upward pointing triangles a negative association is indicated by downward pointing triangles
Note p-values give no indication of magnitude of change
Hernandez DG et al Hum Mol Genet 2011
Studies linking early life exposures to changes in DNA methylation using animal models
Early life exposure Animal
model Epigenetic change
Disease
association
Maternal nutrition
Low Protein Rat Mouse
Pig
and DNA methylation and histone
acetylation and histone methylation Obesity
Calorie restriction Sheep Rat DNA methylation histone acetylation and
histone methylation
Obesity
Diabetes
Periconceptional restriction B12
folate methionine Sheep Altered DNA methylation Obesity
High fat Macaque
Mouse
and DNA methylation and histone
acetylation and and histone methylation Obesity
Surgical models
IUGR ( uterine artery ligation) Rat Altered DNA methylation histone acetylation Diabetes
Environmental toxin
Arsenic Mouse DNA methylation Diabetes
Paternal effect
Low protein Mouse DNA methylation Obesity
Neonatal diet
Leptin treatment Rat DNA methylation Obesity
Extendin-4 Rat Hyperacetylation Diabetes
Reversal with folic acid
Methyl supplementation Avy mouse DNA methylation Obesity
Genistein supplementation +FA Avy mouse DNA methylation Obesity
Protein restriction + FA Rat Prevented or reversed hypomethylation Obesity
Seki Y et al Endocrinology 2012
The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352
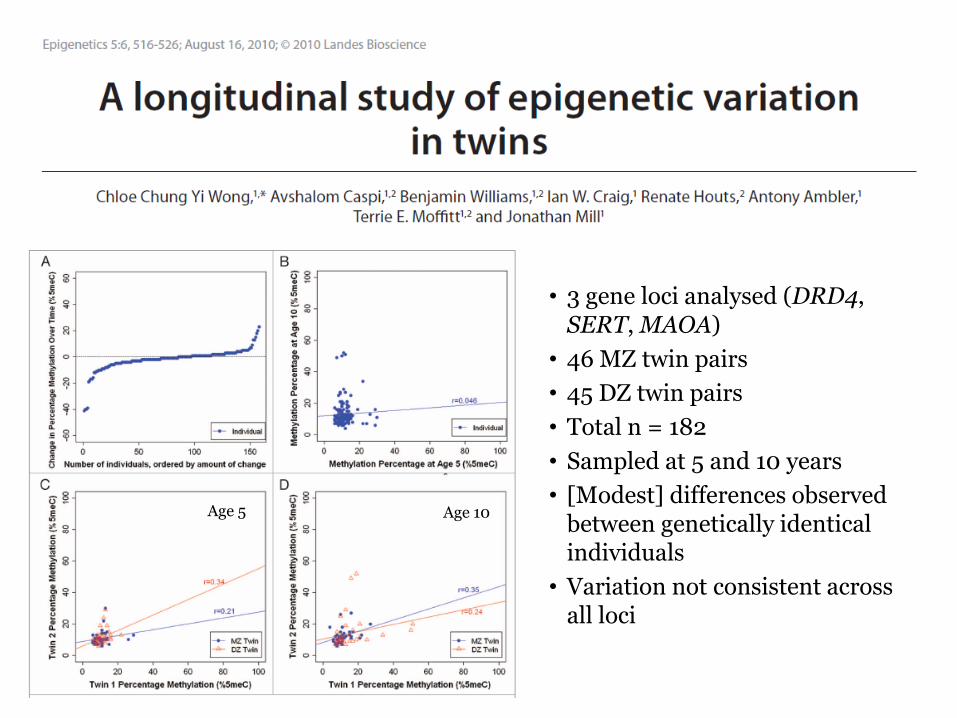
bull 3 gene loci analysed (DRD4 SERT MAOA)
bull 46 MZ twin pairs
bull 45 DZ twin pairs
bull Total n = 182
bull Sampled at 5 and 10 years
bull [Modest] differences observed between genetically identical individuals
bull Variation not consistent across all loci
Age 5 Age 10
Age-related change in methylation
Manhattan plot showing association between methylation at individual CpG sites and chronological age Plotted are P-values indicating strength of association between DNA methylation levels at gt27 000 CpG sites and age in cerebellum (purple) frontal cortex (green) pons (blue) and temporal cortex (red) For each point a positive association between DNA methylation and chronological age is indicated by upward pointing triangles a negative association is indicated by downward pointing triangles
Note p-values give no indication of magnitude of change
Hernandez DG et al Hum Mol Genet 2011
Studies linking early life exposures to changes in DNA methylation using animal models
Early life exposure Animal
model Epigenetic change
Disease
association
Maternal nutrition
Low Protein Rat Mouse
Pig
and DNA methylation and histone
acetylation and histone methylation Obesity
Calorie restriction Sheep Rat DNA methylation histone acetylation and
histone methylation
Obesity
Diabetes
Periconceptional restriction B12
folate methionine Sheep Altered DNA methylation Obesity
High fat Macaque
Mouse
and DNA methylation and histone
acetylation and and histone methylation Obesity
Surgical models
IUGR ( uterine artery ligation) Rat Altered DNA methylation histone acetylation Diabetes
Environmental toxin
Arsenic Mouse DNA methylation Diabetes
Paternal effect
Low protein Mouse DNA methylation Obesity
Neonatal diet
Leptin treatment Rat DNA methylation Obesity
Extendin-4 Rat Hyperacetylation Diabetes
Reversal with folic acid
Methyl supplementation Avy mouse DNA methylation Obesity
Genistein supplementation +FA Avy mouse DNA methylation Obesity
Protein restriction + FA Rat Prevented or reversed hypomethylation Obesity
Seki Y et al Endocrinology 2012
The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Age-related change in methylation
Manhattan plot showing association between methylation at individual CpG sites and chronological age Plotted are P-values indicating strength of association between DNA methylation levels at gt27 000 CpG sites and age in cerebellum (purple) frontal cortex (green) pons (blue) and temporal cortex (red) For each point a positive association between DNA methylation and chronological age is indicated by upward pointing triangles a negative association is indicated by downward pointing triangles
Note p-values give no indication of magnitude of change
Hernandez DG et al Hum Mol Genet 2011
Studies linking early life exposures to changes in DNA methylation using animal models
Early life exposure Animal
model Epigenetic change
Disease
association
Maternal nutrition
Low Protein Rat Mouse
Pig
and DNA methylation and histone
acetylation and histone methylation Obesity
Calorie restriction Sheep Rat DNA methylation histone acetylation and
histone methylation
Obesity
Diabetes
Periconceptional restriction B12
folate methionine Sheep Altered DNA methylation Obesity
High fat Macaque
Mouse
and DNA methylation and histone
acetylation and and histone methylation Obesity
Surgical models
IUGR ( uterine artery ligation) Rat Altered DNA methylation histone acetylation Diabetes
Environmental toxin
Arsenic Mouse DNA methylation Diabetes
Paternal effect
Low protein Mouse DNA methylation Obesity
Neonatal diet
Leptin treatment Rat DNA methylation Obesity
Extendin-4 Rat Hyperacetylation Diabetes
Reversal with folic acid
Methyl supplementation Avy mouse DNA methylation Obesity
Genistein supplementation +FA Avy mouse DNA methylation Obesity
Protein restriction + FA Rat Prevented or reversed hypomethylation Obesity
Seki Y et al Endocrinology 2012
The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Studies linking early life exposures to changes in DNA methylation using animal models
Early life exposure Animal
model Epigenetic change
Disease
association
Maternal nutrition
Low Protein Rat Mouse
Pig
and DNA methylation and histone
acetylation and histone methylation Obesity
Calorie restriction Sheep Rat DNA methylation histone acetylation and
histone methylation
Obesity
Diabetes
Periconceptional restriction B12
folate methionine Sheep Altered DNA methylation Obesity
High fat Macaque
Mouse
and DNA methylation and histone
acetylation and and histone methylation Obesity
Surgical models
IUGR ( uterine artery ligation) Rat Altered DNA methylation histone acetylation Diabetes
Environmental toxin
Arsenic Mouse DNA methylation Diabetes
Paternal effect
Low protein Mouse DNA methylation Obesity
Neonatal diet
Leptin treatment Rat DNA methylation Obesity
Extendin-4 Rat Hyperacetylation Diabetes
Reversal with folic acid
Methyl supplementation Avy mouse DNA methylation Obesity
Genistein supplementation +FA Avy mouse DNA methylation Obesity
Protein restriction + FA Rat Prevented or reversed hypomethylation Obesity
Seki Y et al Endocrinology 2012
The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352
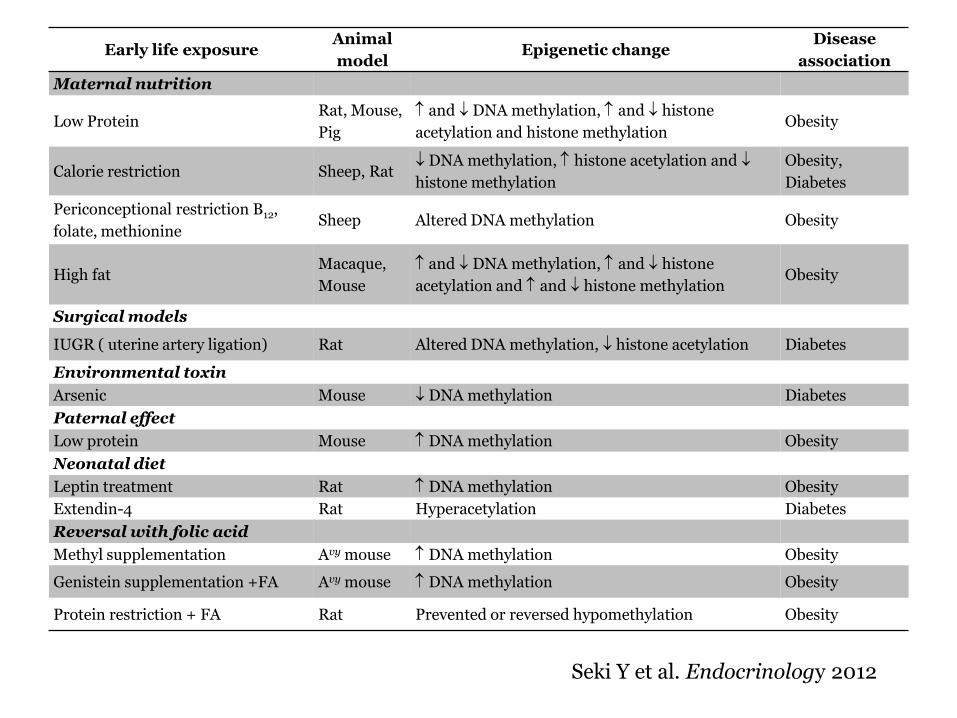
Early life exposure Animal
model Epigenetic change
Disease
association
Maternal nutrition
Low Protein Rat Mouse
Pig
and DNA methylation and histone
acetylation and histone methylation Obesity
Calorie restriction Sheep Rat DNA methylation histone acetylation and
histone methylation
Obesity
Diabetes
Periconceptional restriction B12
folate methionine Sheep Altered DNA methylation Obesity
High fat Macaque
Mouse
and DNA methylation and histone
acetylation and and histone methylation Obesity
Surgical models
IUGR ( uterine artery ligation) Rat Altered DNA methylation histone acetylation Diabetes
Environmental toxin
Arsenic Mouse DNA methylation Diabetes
Paternal effect
Low protein Mouse DNA methylation Obesity
Neonatal diet
Leptin treatment Rat DNA methylation Obesity
Extendin-4 Rat Hyperacetylation Diabetes
Reversal with folic acid
Methyl supplementation Avy mouse DNA methylation Obesity
Genistein supplementation +FA Avy mouse DNA methylation Obesity
Protein restriction + FA Rat Prevented or reversed hypomethylation Obesity
Seki Y et al Endocrinology 2012
The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

The component parts of a gene
Intron Exon
Gene body
Promoter Enhancer
Transcription start site
Transcription factor binding sites
Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Environmentally induced epigenetic changes to promoter-enhancer interaction
bull Sub-optimal nutrition in early life modifies a promoter-enhancer interaction at the Hnf4 locus
bull Role in fetal pancreatic development
bull Implicated in type 2 diabetes aetiology
bull Modest impact upon DNA methylation
bull Pronounced effects upon histone marks
Sandovici I et al Proc Natl Acad Sci 2011
Pro
mo
ter
En
ha
nce
r
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352
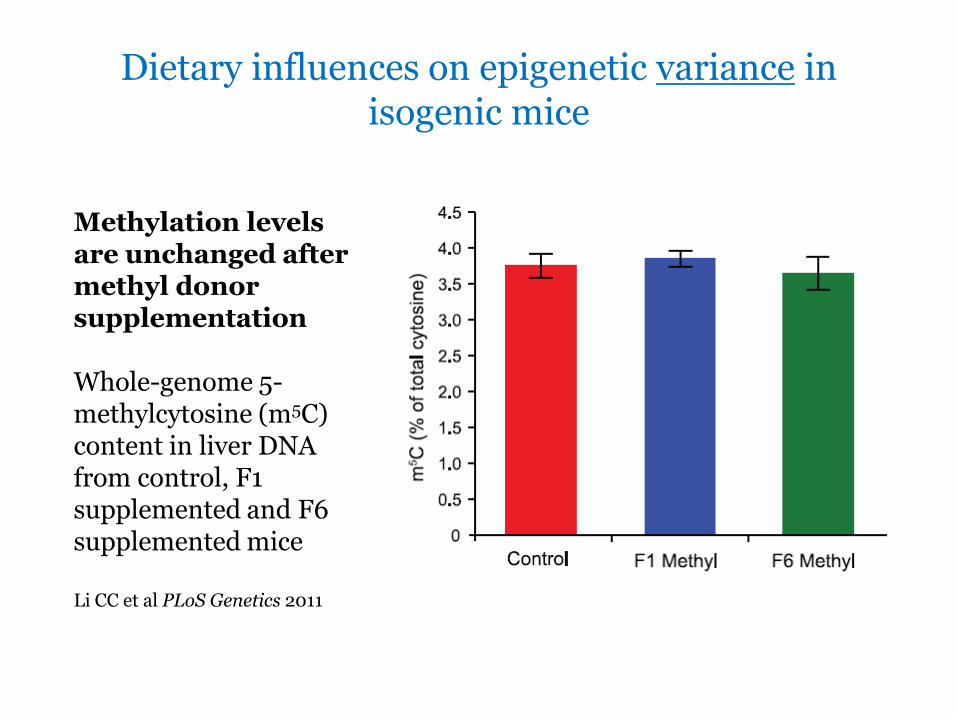
Dietary influences on epigenetic variance in isogenic mice
Methylation levels are unchanged after methyl donor supplementation Whole-genome 5-methylcytosine (m5C) content in liver DNA from control F1 supplemented and F6 supplemented mice Li CC et al PLoS Genetics 2011
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352
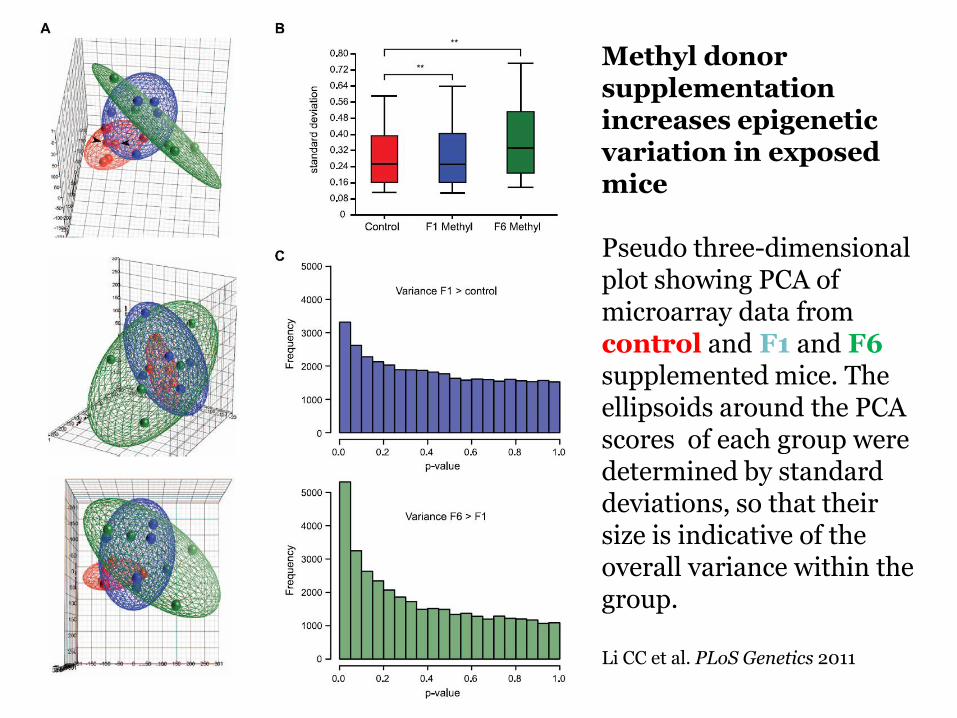
Methyl donor supplementation increases epigenetic variation in exposed mice Pseudo three-dimensional plot showing PCA of microarray data from control and F1 and F6 supplemented mice The ellipsoids around the PCA scores of each group were determined by standard deviations so that their size is indicative of the overall variance within the group Li CC et al PLoS Genetics 2011
Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Evidence from human studies
Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Trans-generational effects vs inter-generational exposure
bull DOHaD is largely concerned with inter-generational exposure ie exposure of the developing fetus whilst in utero via dietary lifestyle and behavioural exposures to the mother
bull A lack of clarity in the literature has led to mis-interpretation of inter-generational exposures as trans-generational effects ie those inherited through altered germ line epigenetics
bull Interest in epigenetics in the context of evolution adaptation and selection means language is used across disciplines but with differing definitions and in different contexts
bull Trans-generational effects are likely to play an extremely small role in disease pathogenesis
Genome Res 2010 2(12) 1623-8 Int J Epidemiol 2012 41(1) 236-47
Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Persistence versus transient epigenetic changes
bull Metabolic programming
hellipthe concept that a stimulus or insult operating at a critical or sensitive period of development could result in a long-standing or life-long effect on the structure or function of the organism Lucas A Human milk and infant feeding In Battaglia F Boyd R eds Perinatal medicine London Butterworths 1983172ndash200
Transient
Persistent
Met
hy
lati
on
ch
an
ge
Time
bull Acute or chronic exposure
bull Long term epigenetic change required
bull Transient epigenetic change with lasting physiological impact
bull Implications for the age of population studied and the inferences that can be made
Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Temporal relationships between exposures and epigenetic patterns
bull The DOHaD paradigm is based upon the premise of a temporal relationship between exposure (in early life) and an outcome (later in the life course)
bull Longitudinal studies can assist in defining whether methylation changes occur before the onset of phenotype
bull HOWEVER a temporal relationship does not necessarily infer causation (but it helps)
bull Confounding structures within data can persist across the lifecourse
Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Temporal relationships in epigenetics the problem of confounding
Debbie Lawlor
Centre for Causal Analyses in Translational Epidemiology
University of Bristol UK
Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Confounding
bull Affects is associated with exposure
bull Affects outcome
bull Is not on the causal pathway between exposure and outcome
bull Fools (confounds) us into believing an association is causal
bull Can distort associations in either direction eg smoking may mask (negative confounding) a stronger effect of BMI on CHD
Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Confounders
eg Dietary fat
Cigarette smoking
Physical activity
Risk protective factor
eg Vitamin C
Disease outcome
eg CHD
Confounding
What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

What this means
bull If interested in best causal estimate must
bull Have knowledge of all possible confounders
bull Measure these accurately
bull Correctly control for them (eg correctly modelled in multivariable analyses)
bull Ideally should be measured before (or at same time) as exposure ndash since to confound the confounder had to influence the exposure amp outcome
bull But also need to understand plausible confounding causal pathways ndash eg smoking as confounder between birthweight amp CHD
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352
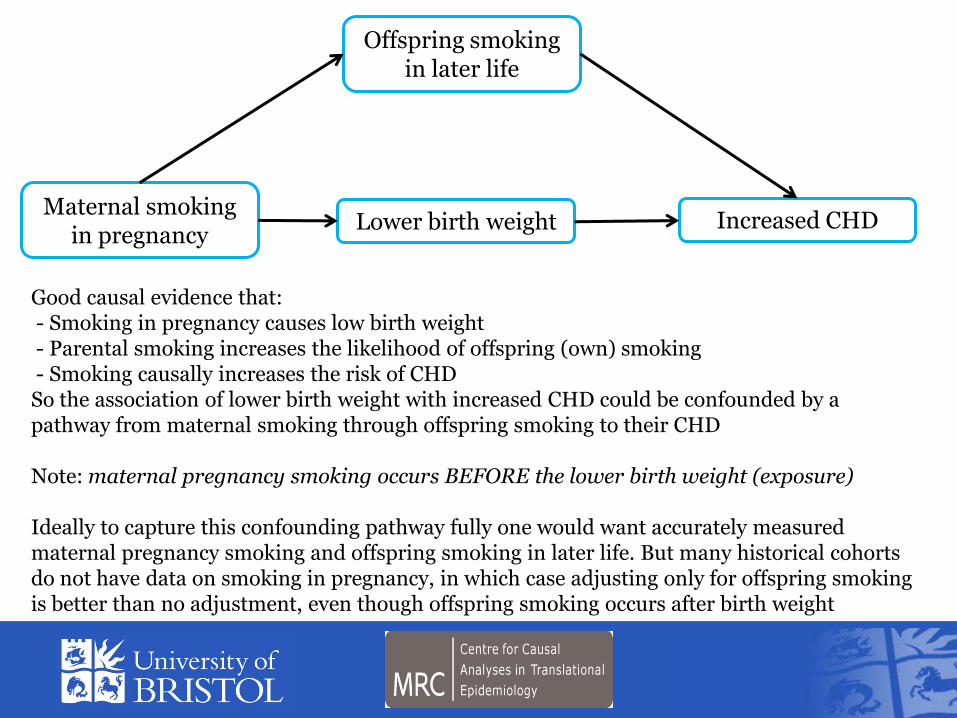
Lower birth weight Increased CHD Maternal smoking
in pregnancy
Offspring smoking in later life
Good causal evidence that - Smoking in pregnancy causes low birth weight - Parental smoking increases the likelihood of offspring (own) smoking - Smoking causally increases the risk of CHD So the association of lower birth weight with increased CHD could be confounded by a pathway from maternal smoking through offspring smoking to their CHD Note maternal pregnancy smoking occurs BEFORE the lower birth weight (exposure) Ideally to capture this confounding pathway fully one would want accurately measured maternal pregnancy smoking and offspring smoking in later life But many historical cohorts do not have data on smoking in pregnancy in which case adjusting only for offspring smoking is better than no adjustment even though offspring smoking occurs after birth weight
Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Difficulties in controlling confounding
bull Unmeasured confounding
bull Very difficult to measure all factors associated with both treatment and outcome (confounders from across the life course)
bull Residual confounding
bull If confounders are measured with error then they wonrsquot be fully controlled in regression models
bull lsquoAssociational worldrsquo difficult to think of every possible confounder and include in statistical model
bull May model confounders incorrectly
Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Associational World
Pair-wise associations
Expected significant at
p lt 001
Observed significant at
p lt 001
P for null observed =
expected
96 non-genetic traits
4560 456 (1) 2036 (45) lt 0000001
Davey Smith G Lawlor DA Harbord R et al PLoS-Med 2007
Including traits from across the life course eg birth weight childhood social class leg length (marker of childhood nutrition) associated with various adult lsquorisk factorsrsquo including HRT use serum vitamin C amp E levels etc
bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

bull Characteristics with lsquosmallrsquo associations in data driven confounder selection often excluded from final model
bull Sensitivity analyses examine how strong one potential confounder would need to be related to exposure and outcome to nullify the best adjusted association
bull BUT the more plausible situation is that many many confounders from across the life course each with lsquosmallrsquo association combine to produce a big confounding effect
Real sensitivity analyses
Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Difference between top and bottom frac14
Vit C
Independent OR for CHD
Predicted OR comparing top to
bottom frac14 Vit C
Child NM social class 95 079 098
Child car access 76 075 098
Full time education gt 18 113 065 095
Adult NM social class 170 077 096
Not living in council house 15 064 099
Adult car access 132 077 097
State plus other pension 123 088 099
None smoker 112 068 096
Regular activity 118 067 095
Low fat diet 62 063 097
High fibre diet 22 086 099
Not obese 104 076 097
Reg Moderate alcohol 111 080 098
Leg length per cm 0095 075 097
FEV1 per litre 019 055 089
Total confounding effect
060
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352
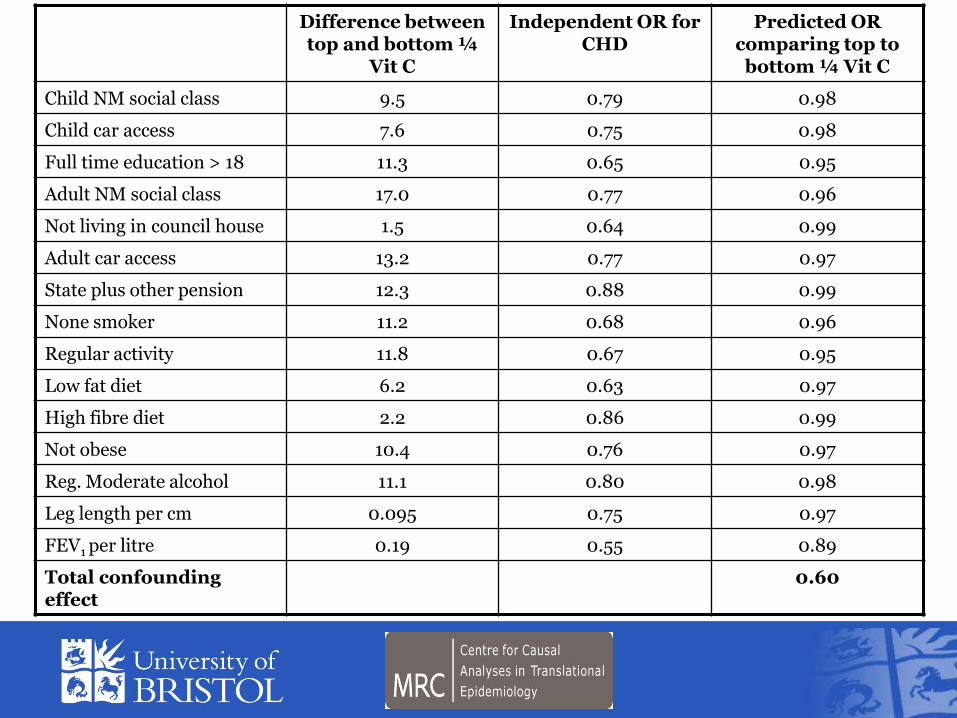
Observed vitamin C - CHD association in a cohort and an RCT
HR (95CI) incident CHD per 157micromoll
Cohort no adjustment
Cohort adult confounder adjustment
Cohort adult amp childhood
confounder adjustment
RCT
088 (080 097) 090 (082 099) 096 (085 105) 102 (094 111)
bull 157micromoll ndash is the difference in vitamin C achieved in the RCT by supplementation
bull Associations in the cohort study progressively attenuate from 12 reduction per dose to 4 reduction as go from no adjustment to adjustment for all available confounders from childhood and adulthood
bull Given lsquoassociational worldrsquo cannot be certain that residual confounding remains
bull Well conducted RCTs will not be confounded because randomisation breaks the association of confounder with exposure (so do not need to measure all confounders)
hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

hellip back to hellip
Caroline Relton
Institute of Genetic Medicine
Newcastle University UK
Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Epidemiological strategies for strengthening causality in a DOHaD context
bull Observed associations between an exposureDNA methylation and DNA methylationoutcome represent a first step in identifying a robust mechanistic pathway
bull Additional strategies can be applied using epidemiological approaches
ndash Replication in an independent sample
ndash Cohort comparison in particular where the second cohort is not subject to the same confounding influences
ndash Paternal versus maternal associations to decipher true in utero effects
ndash Using genetic proxies for exposure andor methylation levels (Mendelian randomization)
bull As well as other tools ndash More details later
Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Using genetic information
Diabetes 2012 61(2) 391-400
TACSTD2 methylation
Childhood adiposity Postnatal growth TACSTD2 expression
TACSTD2 SNP
Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Differential gene expression and DNA methylation are associated with postnatal growth and
childhood adiposity
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
Rho = -055 p = 0016
n = 20
Rho = -022 p = 0037
n = 91
Rho = 044 p = 0061 n = 20
Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

Reverse causation and confounding
Differential gene expression
0-12 weeks 11 years
Differential postnatal growth
Adiposity
Differential gene methylation
bull Are changes in methylation caused by childhood phenotype
bull Are changes in methylation caused by early growth patterns
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352
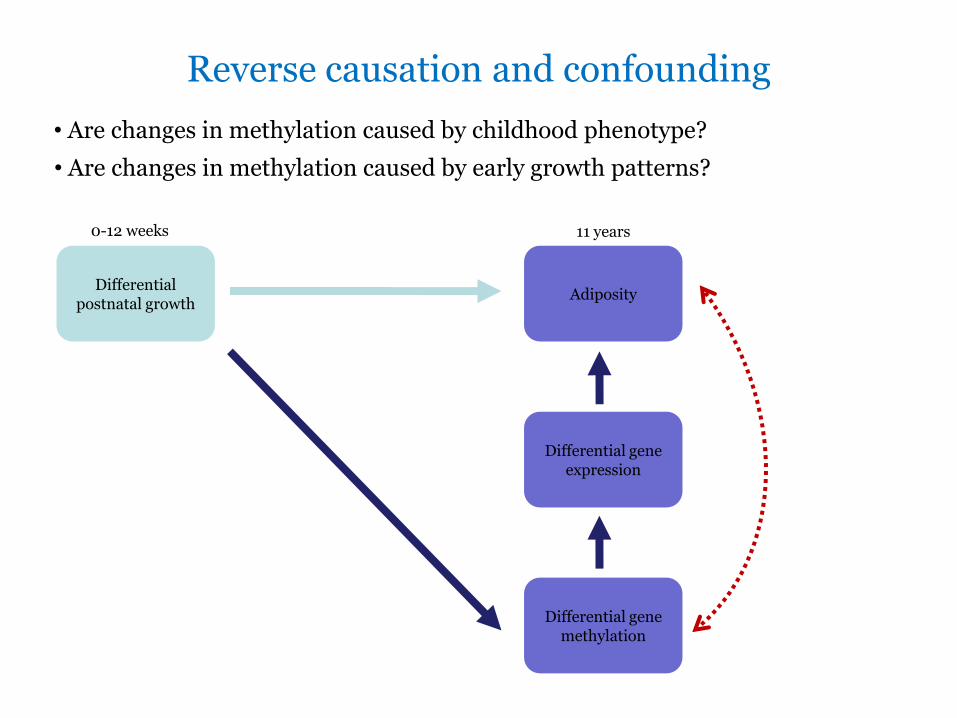
Summary
bull Evidence suggests that epigenetic processes are likely to play a role in developmental programming
bull Animal evidence is more compelling than human
bull Care is required not to confuse inter-generational exposure with trans-generational inheritance
bull We know little about the persistence of epigenetic marks
bull Identifying the role of epigenetic processes in the context of developmental programming faces all of the same challenges as other epidemiological analyses
bull Temporal relationships between exposure mediator and outcome are a pre-requisite but not a guarantee of an un-confounded association
References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352

References bull Waterland RA Epigenetic epidemiology of obesity application of epigenomic technology Nutr Rev 2008 66 (Suppl 1)
S21-3
bull Wong C et al A longitudinal study of epigenetic variation in twins Epigenetics 2010 5(6) 516-26
bull Hernandez DG et al Distinct DNA methylation changes highly correlated with chronological age in the human brain Hum Mol Genet 2011 20(6) 1164-72
bull Seki Y et al Minireview Epigenetic programming of diabetes and obesity Animal models Endocrinology 2012 153(3) 1031-8
bull Sandovici et al Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets Proc Natl Acad Sci USA 2011 108(13) 5449-54
bull Li CC et al A sustained dietary change increases epigenetic variation in isogenic mice PLoS Genet 2011 7(4) e1001380
bull Tobi EW et al Prenatal famine and genetic variation are independently and additively associated with DNA methylation and regulatory loci within IGF2H19 PLoS ONE 2012 7(5) e37933
bull Relton CL et al DNA methylation patterns in cord blood DNA and body size in childhood PLoS ONE 2012 7(3) e31821
bull Godfrey KM et al Epigenetic gene promoter methylation at birth is associated with childrsquos later adiposity Diabetes 201160(5)1528-34
bull McKay JA et al Genetic and non-genetic influences during pregnancy on infant global and site specific DNA methylation role for folate variants and vitamin B12 PLoS ONE 2012 7(3) e33290
bull Groom A et al Postnatal growth and DNA methylation are associated with differential gene expression of the TACSTD2 gene and childhood fat mass Diabetes 2012 61(2) 391-400
bull Daxinger L Whitelaw E Transgenerational epigenetic inheritance more questions than answers Genome Res 2010 2(12) 1623-8
bull Davey Smith G Epigenesis for epidemiologists does evo-devo have implications for population health research and practice Int J Epidemiol 201241(1)236-47
bull Davey Smith G et al Clustered environments and randomized genes a fundamental distinction between conventional and genetic epidemiology PLoS Med 2007 4(12) e352





























