Embed Size (px)
Citation preview

Gastroenteritis ovina
21ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
Trabajos de Revisión
Dirección para correspondencia: Jorge Romero. Centro de Diagnóstico e Investigaciones VeterinariasCEDIVE, Facultad de Ciencias Veterinarias Universidad Nacional de La Plata. CC 147 (B 7130) Chascomús,ARGENTINA E-mail: [email protected]
Fecha de recepción: 11/04/00 Fecha de aprobación: 10/01/02
EPIDEMIOLOGÍA DE LA GASTROENTERITIS VERMINOSADE LOS OVINOS EN LAS REGIONES
TEMPLADAS Y CÁLIDAS DE LA ARGENTINA
JR Romero1, CA Boero2
1Centro de Diagnóstico e Investigaciones Veterinarias CEDIVEFacultad de Ciencias Veterinarias Universidad Nacional de La Plata.
2Director Técnico de Laboratorio Mesopotámico.
RESUMEN: Se presenta una discusión sobre epidemiología del parasitismo gastrointestinal delanares con especial énfasis en la problemática de áreas templadas. Se revisa la información dispo-nible en la Argentina y Uruguay, discutiendo situaciones sobre las que no se dispone de datosexperimentales. Además se analiza el problema de la resistencia a antihelmínticos con vistas almanejo integrado del parasitismo como alternativa.
Palabras clave: Gastroenteritis verminosa, ovinos, Argentina, epidemiología
EPIDEMIOLOGY OF PARASITIC GASTROENTERITIS OF SHEEP IN TEMPERATE AND WARM AREAS IN ARGENTINA
ABSTRACT: The paper discusses epidemiological aspects of gastrointestinal parasitism, particu-larly in temperate areas, and reviews available information on real situations of Argentina andUruguay. In addition some aspects of drug resistance are discussed pointing out the use of inte-grated management systems as an alternative measure to control parasitism.
Key words: Parasitic gastroenteritis, sheep, Argentina, epidemiology
El Sitio Argentino de Producción Animal
1 de 17

J. Romero y col.
22 ANALECTA VETERINARIA 2001; 21, 1: 21-37 ISSN 1514259-0
INTRODUCCIÓNLas existencias ovinas del país se estiman
actualmente en algo menos de 15 millones de ca-bezas, la mitad de las cuales está en la Patagonia.La provincia de Buenos Aires tiene unos 3 millo-nes, Corrientes y Entre Ríos tienen 1.800.000, yel resto está distribuido en explotaciones de im-portancia menor, o vinculadas a economías de sub-sistencia en todo el país.
En términos generales la explotación de laPatagonia es eminentemente lanera, especiali-zada en lanas finas, predominando raza Merino,salvo en las provincias de Santa Cruz y Tierra delFuego, donde tiene gran representatividad la razaCorriedale.
En Buenos Aires existe gran diversidad derazas y tamaños de explotación: en la región Su-deste y Sudoeste hay Merino y Corriedale, mien-tras que en la Zona Deprimida del Salado predo-minan las existencias de Romney y Corriedale,resultando insignificantes en la actualidad lasmajadas de Lincoln. Existen en la Pampa húmedagran cantidad de majadas pequeñas, de consumosin proyección comercial. Si bien no son numéri-camente importantes, las explotaciones lecherasson un interesante foco de atención, las razas masimportantes constituyen cruzas Frisonas y enmenor medida, Texel y Manchega. En la Mesopo-tamia existen aún importantes explotaciones co-merciales siendo predominantes las razas Corrie-dale, Romney e Ideal (Polward).
El negocio de carne, es el consumo interno(y de estancia). La depresión del mercado de la-nas en las últimas décadas, ha sido la causa de lareducción del stock en la Argentina y en otros paí-ses. El parasitismo gastrointestinal es la variablesanitaria más importante de explotación de lana-res en áreas templadas y por ello hemos puestoen éstas el enfoque de la presente revisión.
Principales especies que parasitan alanares en nuestro medio.
Los especies gastrointestinales más impor-tantes aislados en lanares en nuestro país revi-sadas por Johnstone (1), Lukovich (2), M. Suárez(3), V. Suárez (4, 5) entre otros son:
Estómago:Ostertagia ostertagi, O. lyrata, O. trifurcata,
Marshallagia marshalli, Teladorsagia circumcincta,T. davtiana.
Haemonchus contortus, H. placeiTrichostrongylus axei
Intestino delgado:T. colubriformis, T. vitrinus, T. longispicularis,
T. capricolaStrongyloides papillosusCooperia serrata, C. curticei ,C. oncophora, C.
mcmasteri, C. pectinataNematodirus filicollis, Nematodirus spathiger,
N. abnormalis, N. oriatianus, N. battus, N. helvetianusBunostomum trigonocephalumToxocara vitulorum
Intestino gruesoChabertia ovinaOesophagostomum columbianum, O.
venulosumTrichuris ovis
Reseña del ciclo evolutivo:No es intención de la presente revisión ex-
playarse en este sentido y sólo se hará una brevereferencia. Las hembras adultas de estas espe-cies depositan sus huevos en la luz del tracto gas-trointestinal, los que se eliminan con la materiafecal del hospedador.
Las larvas de Toxocara spp. (Orden Ascaridida)y Trichuris (Orden Trichurida), evolucionan dentrodel huevo. Cuando éstas maduran constituyen laforma infectante, existiendo en el primero, ade-más, transmisión transplacentaria. Las formas pa-rasitarias de Strongyloides spp. (Orden Rhabditida)son hembras partenogenéticas. Sus huevos, unavez en el medio, dan lugar a la formación de gene-raciones de adultos de vida libre y larvas 3 infec-tantes. Estas pueden también provenir de esasgeneraciones de vida libre. Las L3 de Strongyloi-des spp., infectan atravesando la piel o la mucosaoral de los animales, con lo que las infeccionessuelen ser muy tempranas. Luego de una migra-ción traqueal alcanzan el intestino delgado y enpocos días cumplen totalmente el período prepa-tente iniciando la oviposición.
El resto de los nematodes mencionados per-tenecen al orden Strongylida con un patrón de ci-clo evolutivo similar, en que los huevos liberadoscon la materia fecal evolucionan en el medioambiente (generalmente en el interior de la masade heces) eclosionando una L 1. Esta se alimentaen el medio fecal, crece y muda dos veces. La lar-va 3 surgida de esa segunda muda retiene la cutí-cula de la L2, lo que le da gran resistencia a lascondiciones ambientales (como excepción, Nema-todirus spp. alcanza el estado L3 dentro del huevoy luego eclosiona conservando su doble cutícula)Este proceso dura entre poco más de una y seissemanas, según la temperatura, pudiendo dete-
El Sitio Argentino de Producción Animal
2 de 17

Gastroenteritis ovina
23ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
nerse cuando las temperaturas son inferiores alumbral de cada especie, o retomarse si las condi-ciones no han sido letales. Las L1 y 2 son las mássusceptibles. La L3 es la infectante y su supervi-vencia y capacidad de dispersión es clave en elproceso de alcanzar al siguiente hospedador. Pue-den sobrevivir varios meses y en condiciones idea-les, hasta un año o más. Una vez ingeridas las L3deben fijarse en su órgano blanco, luego de lo cualvuelven a mudar. En su estado de L4 son histotró-ficas y, eventualmente, pasan un período en hi-pobiosis que prolonga a veces por varios meses. Eltiempo de prepatencia normalmente es de 3 a 4semanas en Trichostrongylidae y de 6 a 8 en Stron-gylidae para alcanzar el estado adulto en la luz delestómago o intestino.
El daño que producen los parásitos:Las lesiones en el estómago producidas por
Ostertagia spp, Teladorsagia circumcincta, y Trichos-trongylus axei, alteran la funcionalidad de las glán-dulas. Esto disminuye la producción de ácido clor-hídrico, elevando el pH gástrico e impidiendo laactivación del pepsinógeno, con la consiguientefalta de digestión proteica. La infiltración e infla-mación de la mucosa altera la continuidad de lasuniones desmosómicas entre células epitelialescon el consiguiente pasaje de proteínas hacia laluz de la víscera y eventualmente de pepsinógenoal plasma. Luego, la diarrea se debe al pasaje dequimo indigesto al intestino, y la hipoproteine-mia a la pérdida osmótica y a la falta de digestiónpépsica. Trichostrongylus axei, cuyos adultos intro-ducen su extremidad anterior en la lámina pro-pia y en distinto grado las formas juveniles de és-tos y de Haemonchus spp, producen el mismo cua-dro dominado por la reacción del hospedador (6).
Haemonchus contortus, tiene una acción pa-rasitaria expoliatriz hematófaga, el signo carac-terístico es la anemia. Como es muy elevado supotencial biótico suelen alcanzarse rápidamentealtas cargas parasitarias por esta razón el cuadrode anemia y muerte de los animales se presentatambién muy rápido (en esos brotes son comunesniveles de hematocrito de 15 %). Cuando los picosde infección coinciden con la parición, esos cua-dros se presentan también en animales adultos,y si la parición coincide con el final de un períodode hipobiosis, pueden registrarse cargas elevadasen las hembras paridas.
Cooperia spp. y Nematodirus spp., producenlesiones superficiales en la mucosa intestinal lasque revisten gravedad sólo en infecciones impor-tantes. Las infecciones intestinales por Nemato-dirus spp. son frecuentes en corderos y difícilmente
alcanzan cargas elevadas. De los huevos que so-breviven cómodamente inviernos fríos, eclosionanlarvas en primavera que infectan a animales jó-venes en los que rápidamente inducen inmuni-dad que, de alguna forma, limita una larga suce-sión de reinfecciones. La estacionalidad es rela-tivamente menos marcada en zonas cálidas perosu abundancia es siempre baja. Las especies deCooperia no alcanzan en nuestro medio cargas ele-vadas y varían según la cohabitación de potreroscon terneros y la existencia de infecciones cruza-das. Bunostomum sp. es un parásito de gran pato-genicidad por su acción expoliatriz y efecto trau-mático. No es frecuente hallar altas cargas.
Las especies de Trichostrongylus que habi-tan el intestino delgado, especialmente T.colubriformis son más patógenas y los adultos le-sionan mas profundamente en la mucosa suman-do a los efectos de la reacción inflamatoria los desu acción expoliatriz.
Las diarreas en corderos son típicas en laProvincia de Buenos Aires en invierno y se aso-cian a gastroenteritis causadas principalmentepor Ostertagia spp., Teladorsagia spp., Trichostron-gylus spp. y Cooperia spp. Las “cascarrias” que seobservan a fines de invierno se asocian a estetipo de infecciones gástricas o intestinales.
Finalmente, en el intestino grueso suelenhallarse cargas de varios cientos de Trichuris ovis,Oesophagostomum spp. cuya marcada patogenici-dad las hace relativamente graves . La reacciónde hipersensibilidad retardada en torno a los si-tios de ubicación histotrófica de las larvas de Oe-sophagostomum spp. se traduce en un trastornoclínico sólo con infecciones importantes en las quelos adultos también afectan la mucosa intestinaly ejercen su acción expoliatriz. Las cargas de Cha-bertia ovina no presentan grandes alteraciones,no porque el parásito no sea potencialmente agre-sivo sino porque las posibilidades de alcanzar al-tas cargas están limitadas. Los hallazgos de estaespecie en áreas templadas son siempre en es-casa cantidad.
Estudios de pérdidas en la región semiáridapampeana (4, 5, 6, 7), arrojaron diferencias de 3 a4 kg en la ganancia de peso en recría de borregos,agravada por mortandades y otras alteraciones pro-ductivas en los picos de parasitismo. Por otro ladoCastells y col. (8) demostraron también en el Uru-guay pérdidas potenciales por mortandad en recríade hasta el 50 %, de un 23 % en el peso vivo deanimales no tratados, más un 29 % en el peso delvellón, además de pérdidas en la calidad de lana.
El Sitio Argentino de Producción Animal
3 de 17

J. Romero y col.
24 ANALECTA VETERINARIA 2001; 21, 1: 21-37 ISSN 1514259-0
Especie Fecundidad Tiempo de generación Potencial biótico(individuos totales de (período entre el inicio número de generacio-cada generación o de la vida de una generación nes y de individuosproducidos en una y el inicio de la siguiente) posibles a los 40 díasunidad de tiempo ) días-Producción sexualo asexual
a 100.000.000 40 1 generación: (100.000.000 1)(10 8 individuos)
b 10 5 8 generaciones (10 8)(10 8 individuos)
c 10.000 10 4 generaciones (10.000 4)(10 8 individuos)
BASES PARA LA COMPRENSIÓN DELA EPIDEMIOLOGÍA
La dinámica del parasitismo puede repre-sentarse por modelos conceptuales de la situaciónde campo que valoren la presencia, abundancia,distribución espacial (regional o dentro de un de-terminado nicho) y temporal (estacionalidad) delas principales especies. Estos parámetros estáninfluidos por 4 grupos de factores (9).
1. Dependientes de los parásitos:Los parásitos de ciclo directo habitan en dos
ambientes durante su ciclo vital: el medio exter-no, donde evolucionan expuestos a condiciones cli-máticas variables y a los factores de dispersión; yel hospedador, en el que enfrentan la localizaciónde su nicho de ubicación, la respuesta inmune,la competencia entre especies parasitarias y lostratamientos farmacológicos.
Los factores de adaptación son genéticos,predeterminados en cada especie o en cada cepay de ellos depende el comportamiento individual opoblacional.
Entre esos factores los principales son: po-tencial biótico, rango de adaptación a factores cli-máticos, aptitud para la hipobiosis y resistencia alos medicamentos. Los tres primeros se analiza-rán seguidamente, dedicándose a la resistenciaa los medicamentos un espacio más adelante.
Potencial bióticoSe denomina potencial biótico al potencial
de multiplicación de una especie en función deltiempo. Su expresión matemática es una relaciónentre el tiempo de generación y la fecundidad (elvalor de la fecundidad está condicionado al tipo dereproducción, sexual o asexual, la existencia de
hospedadores intermediarios y eventual multipli-cación en ellos, etc.). La expresión de este poten-cial está muy influida por los factores ambienta-les y del hospedador. En la tabla 1 se muestrantres especies hipotéticas, de igual potencial bióti-co.
En especies con sexos separados, la proba-bilidad de nacimiento de hembras modificará es-tos valores (reducirá a la mitad en cada genera-ción los individuos a multiplicar).
Las variaciones en la duración del períodode postura y la supervivencia potencial de las for-mas infectantes en el medio ambiente, modifica-rá tanto el valor de la fecundidad como el tiemporeal de cada generación.
El potencial biótico representa el valor ensituación ideal de adaptación al medio ambiente,y se modifica siempre reduciéndose según losambientes y las circunstancias hasta el límite dela supervivencia de la especie en cada región.
Los que siguen son ejemplos de diferenciasen la ovipostura diaria:
Haemonchus spp. 5.000- 10.000Ostertagia spp. 200 - 300Cooperia spp. 100 - 2.000Trichostrongylus spp. 100 - 200Nematodirus spp. < 100
Ejemplos de diferencia entre períodos pre-patentes de nematodes gastrointestinales de ru-miantes:
Haemonchus placei 26-28 díasOstertagia ostertagi 18-23 “Trichostrongylus axei 18-21 “Cooperia spp. 11-14 “
Tabla 1. Especies hipotéticas de potencial biótico equivalenteTable 1. Biotic potential equivalence of different hypotetic especies
El Sitio Argentino de Producción Animal
4 de 17

Gastroenteritis ovina
25ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
Nematodirus helvetianus 21-26 díasBunostomum phlebotomum 52-56 “Oesophagostomum radiatum 35-41 “Strongyloides papillosus 9 “
Rango de adaptación a condicionesambientales:
Las diferentes temperaturas umbrales parainiciar o detener el desarrollo de las formas devida libre o limitar su supervivencia, la humedadrelativa del ambiente (HRA), y la cantidad de ho-ras con temperatura adecuada, condicionan ladistribución regional y la tendencia estacional delparasitismo por cada especie. Si bien Haemonchuscontortus tiene un gran potencial biótico ve limi-tado su desarrollo con menos de 11 °C de tempe-ratura media, y la sobrevida de las larvas se com-promete severamente por temperaturas cercanasa los 0 °C. Ostertagia, y más aún Nematodirus (queevoluciona hasta L3 dentro del huevo) pueden evo-lucionar con temperaturas de apenas 6 °C, y so-brevivir cerca de los 0°C. A su vez las L 3 de estasespecies están menos adaptadas para soportarperíodos calurosos prolongados especialmente sila HRA es baja (< 60 %). En general las vainas delas larvas 3 de los trichostrongilidos y los huevos(en Trichuris-Toxocara) protegen a las formas in-fectantes, pero la resistencia es diferente entreespecies. Cooperia spp., Trichostrongylus spp., Hae-monchus spp. y Bunostomum spp., tienden a desa-rrollar y sobrevivir mejor en climas cálidos lo queles da mayor oportunidad y explica su distribuciónabundante en el Noreste del país, su estacionali-dad en áreas templadas y su inexistencia en laPatagonia.
Entre los factores de dispersión desde la bos-ta hacia el pasto, con el que serán ingeridas porsus hospedadores, la lluvia es el más importante,(también el pisoteo, especialmente en sistemasde pastoreo intensivo) y, en algunas áreas, el rie-go. Una vez fuera del refugio que representa labosta, las larvas están mucho más expuestas a ladesecación.
Hipobiosis:La hipobiosis en el estado de L4, dentro de
la mucosa del tracto gastrointestinal, es determi-nada genéticamente y su presentación está con-dicionada por factores ambientales soportados fue-ra del hospedador (10). En la zona templada de laArgentina, y Uruguay con veranos calurosos y re-lativamente secos, la inhibición se produce du-rante la primavera-verano en el caso de Osterta-gia spp. (11, 12) o en invierno para Haemonchuscontortus, menos resistente a las condiciones frías
(13, 14). La posibilidad de atravesar períodos pro-longados como larvas hipobióticas determina quesobrevivan a períodos desfavorables desde el pun-to de vista climático, y la reactivación masiva pue-de asociarse a trastornos clínicos severos, inclu-so en ovejas adultas cuando la desinhibición co-incide con el parto en ovejas. Es muy importanteel impacto sobre la infección temprana de los cor-deros que tienen estas poblaciones de helmintosadultos en las madres.
2. Dependientes del ambiente:Condicionan la mayor o menor abundancia
de parásitos en los sistemas. Se expresan en lavariación de los períodos de incubación de hue-vos, tasas de eclosión y sobrevivencia de los esta-díos de vida libre, y aún en los niveles y época dehipobiosis de las L4 en los tejidos de hospedado-res. El ambiente permite o no la instalación depoblaciones de diferente aptitud en distintas re-giones, y condiciona su estacionalidad, ademáslimita el potencial biótico con lo que determinadiferentes niveles de riesgo para las poblacionesde hospedadores. Gordon en Australia y Levine enEE. UU. (21), entre otros, han contribuido a la com-prensión de las relaciones del clima con la epide-miología. Mediante los bioclimatogramas han po-dido definir épocas de riesgo de enfermedad enfunción de las condiciones generales aptas parael desarrollo y traslación de formas infectantesen el suelo, determinando condiciones regiona-les y estacionales que explican y predicen la ten-dencia al establecimiento y abundancia de dife-rentes especies.
Los bioclimatogramas se construyen en undiagrama cartesiano donde las abscisas indicanlas precipitaciones mensuales y las ordenadas lastemperaturas medias de una región dada. Se in-dican los puntos de ocurrencia de ambas varia-bles para cada mes del año y se señalan los valo-res umbrales para el desarrollo de la fase exógenade las especies a analizar, considerando 50 mmde precipitaciones mensuales como suficientespara iniciar la dispersión de las larvas. Los me-ses encerrados entre los umbrales de ambas va-riables son epidemiológicamente aptos y tantomás cuanto más hacia la derecha y arriba deldiagrama se encuentren. No obstante, sobre lasvariables afectadas regularmente por el ambien-te influyen otras dependientes de las condicionesdel clima en cada año, del suelo, la vegetación y,especialmente de situaciones de manejo, por loque en el diagnóstico del riesgo dentro de cadaexplotación se requiere el monitoreo particularde cada lugar.
El Sitio Argentino de Producción Animal
5 de 17

J. Romero y col.
26 ANALECTA VETERINARIA 2001; 21, 1: 21-37 ISSN 1514259-0
3. Dependientes de los hospedadores:Un hecho relevante es que la mayor parte
de los parásitos se encuentran en estado natural,en un número reducido de hospedadores más sus-ceptibles. Por lo tanto éstos condicionan por suabundancia y nivel de susceptibilidad el desarro-llo de las poblaciones de aquellos. El condiciona-miento se expresa en la tasa de establecimientode formas infectantes, sobrevivencia y prolifici-dad de los parásitos.
El primer ámbito de variabilidad en la sus-ceptibilidad de los hospedadores es filogenético.La adaptación mutua a lo largo de la evolución,que ha hecho que las especies parásitas desarro-llen “ajustes” con variables bioquímicas, anató-micas y fisiológicas de sus hospedadores. El se-gundo plano es de carácter inmunológico: los pa-rásitos exponen antígenos estructurales o somá-ticos y funcionales en los distintos estados de suevolución ontogénica, que inician, reaccionesinmunes en el hospedador. Las más caracteriza-das son: a) la eliminación de L3 que intentan es-tablecerse, por medio de reacciones de hipersen-sibilidad, gatillada por reacciones específicas me-diadas por IgE que se fijan a los mastocitos y des-encadenan la serie de eventos del fenómeno alér-gico; b) la eliminación de estados parasitarios endiferente nivel de desarrollo y adultos, gatilladopor mecanismos similares o de citotoxicidad me-diada por diferentes células efectoras (linfocitos,eosinófilos, etc.); c) reacciones de hipersensibili-dad retardada o producción de complejos de anti-cuerpos con antígenos funcionales parasitarios enáreas expuestas de los parásitos que neutraliceno condicionen el desarrollo, las migraciones, lacópula o hasta el ritmo de ovipostura de las hem-bras. La capacidad de respuesta inmune evolu-ciona con la edad y las experiencias de parasitis-mo y los individuos adultos alcanzan un elevadonivel de resistencia.
Los niveles de susceptibilidad individualvarían genéticamente entre individuos de igualcategoría y a los fines prácticos se estudian desdedos puntos de vista: la tolerancia, que es la capa-cidad de algunos hospedadores de albergar grancantidad de parásitos sin sufrir sus efectos en laproducción o la salud, y de resistencia que es laaptitud de algunos hospedadores para interrum-pir total o parcialmente el ciclo y la reproducciónde las poblaciones parásitas a que es expuesto (15).Estos dos factores incrementan aún más la hete-rogeneidad de la distribución de parásitos dentrode la majada (16). En la figura 1 se expresan dife-rentes niveles de resistencia de animales expues-tos desde el destete a igual nivel de infección con
trichostrongilidos y observados por su eliminaciónde huevos (HPG) en varios períodos de desafío na-tural (17).
Figura 1. Diferentes niveles de resistencia enanimales de recría de raza Polward tomados al azarFigure 1. Different levels of resistence in Polward ani-mals in randomized samples.
La correlación entre los recuentos de hue-vos en la materia fecal y la carga de nematodesadultos en animales de necropsia de líneas deovinos resistentes y no resistentes es elevada.Se ha demostrado una mayor infiltración deeosinófilos en la mucosa gastrointestinal en ani-males resistentes, lo que junto con otras eviden-cias inmunológicas fundamentan esa mejor re-acción. El pastoreo separado de poblaciones másresistentes determina una menor contaminacióny menor desafío ulterior que en el equivalente depoblaciones susceptibles. La heredabilidad de laresistencia, estimada en distintos trabajos estáentre 0,26 y 0,30. En ambos subgrupos puedenexistir individuos que soporten las cargas parasi-tarias con buenos niveles de producción en com-paración con otros que se resientan, la selecciónpor tolerancia ha sido muy discutida por ser apa-rentemente de menor heredabilidad que la de laresistencia y más difícil de evaluar. Uno de losprimeros parámetros estudiados en el fenotipo deestos animales fue la capacidad de mantener ele-vado hematocrito frente a infecciones por H.contortus. Se encontró en diferentes trabajos unaheredabilidad extremadamente baja de este ca-rácter (0,09). En cambio cuando se evaluó median-te un test de progenie a campo, en el cual las críasfueron tratadas sólo por evolución de parámetrosclínicos, los indicadores como edad al primer tra-tamiento, frecuencia o número de tratamientosen un tiempo dado; y productivos como gananciade peso hasta el destete o en un período dado, haresultado de gran expectativa y justifica el man-
El Sitio Argentino de Producción Animal
6 de 17

Gastroenteritis ovina
27ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
tenimiento de programas de investigación enNueva Zelanda (18). No existe correlación entrela selección por resistencia y selección realizadapor resiliencia o tolerancia, por lo que programassimultáneos o secuenciados de selección no de-berían interferir entre sí ni con la evolución de laselección por parámetros productivos.
Los animales adultos son más resistentesque los jóvenes pero ese estado de mayor resis-tencia puede variar en diferentes estados repro-ductivos. El “alza de la lactancia” en la elimina-ción de huevos en ovejas desde proximidades delparto y hasta el destete, ha sido observado y des-cripto por gran cantidad de autores y es de coti-diano conocimiento de los productores. Este au-mento en la susceptibilidad del lanar tiene quever con los cambios inmunológicos (especialmen-te en la actividad mitótica de los linfocitos) influi-dos por las concentraciones elevadas de glucocor-ticoides y de prolactina). En la tabla 2 se presen-tan las diferencias en los valores de HPG de ove-jas paridas y secas igualmente expuestas a infec-ción natural en la Provincia de Corrientes.
Las diferencias en resistencia genética ob-servada en corderas jóvenes se expresa tambiénen estado adulto frente a la relajación inmunedel periparto.
bilidad de los ovinos expuestos, y el pastoreo si-multáneo o alternado con especies no suscepti-bles, afectan la dinámica y diversidad natural delas poblaciones. Las cargas elevadas en sistemasintensivos generan altas tasas de contaminaciónaún con niveles iniciales bajos de eliminación dehuevos, lo que se traduce en una elevada trasla-ción e infección posterior en los animales.
EPIDEMIOLOGÍA EN DISTINTAS RE-GIONES.Un modelo de tendencias en la conta-minación de pasturas.
En la figura 2 se describe un modelo de in-fección mixta predominantemente otoño-invernalde áreas templadas (20) comparable lo que ocurri-ría, por ejemplo, en nuestra Pampa húmeda conuna majada de parición de primavera.
Durante la lactancia la oveja se convierteen una multiplicadora del pie de infección encon-trado. Cuando la parición coincide con el períodode desinhibición de larvas de Haemonchus contortus(primavera) y con abundancia de larvas de otrasespecies en el pasto, la oveja lactante produce laprimera multiplicación que funda la población dela temporada siguiente que encontrará en los cor-deros al pie y luego del destete los hospedadoressusceptibles necesarios. Cuando la parición ocu-rre en el pico de condiciones favorables al cicloexterno de Haemonchus contortus (otoño) la infec-ción en las madres tiende a ser elevada e inclusoproducir mortandad.
En nuestro medio no han sido agotados losestudios epidemiológicos por lo que el análisis debebasarse (por lo menos por ahora) en informaciónfehaciente obtenida en algunas regiones y en da-tos más o menos atomizados y observaciones nosistematizadas en otras áreas. Plantearemos amodo de hipótesis las tendencias en la contami-nación de las pasturas y las épocas de riesgo paracada región y principales géneros parasitarios ytrataremos de confrontarla con la información dis-ponible. Para la obtención de esa información, eltrabajo fundacional fue probablemente el releva-miento y consideraciones sobre las parasitosisovinas de la Patagonia, llevado a cabo porJohnstone y col. (1), aunque definió la problemá-tica de una región cuyo análisis no es objeto delpresente. La mayoría de las infecciones son sub-clínicas pero merecen interés los estudios pun-tuales en cada región. Encierran algún riesgo epi-demiológico las concentraciones de animales enlos mallines durante el verano. Estudios posterio-res demuestran que los sistemas de explotación
Tabla 2. HPG en ovejas de raza Polward a 15-30días de paridas y secas, igualmente expuestas ainfección natural.Table 2. Fecal egg count (FEC) in Polward ewes at 15-30days pospartum and dry, with the same exposition to naturalinfection.
hpg ovejas secas paridas
Mediana 0 1500Promedio 58 1664Valores extremos 0-200 150-7950
4. Dependientes del sistemaLas características emergentes de los sis-
temas no son sólo sumatoria de los factores pre-sentes, sino que resultan de la interacción de losmismos, por lo tanto el manejo y desenvolvimien-to de otras variables no estrictamente parasitoló-gicas contribuyen a determinar las tasas de in-fección y su efecto sobre la producción, los trata-mientos antihelmínticos, los cambios de potreroy los niveles de alimentación a que están someti-dos los animales (19). La variación en la suscepti-
El Sitio Argentino de Producción Animal
7 de 17

J. Romero y col.
28 ANALECTA VETERINARIA 2001; 21, 1: 21-37 ISSN 1514259-0
intensivos, o bajo riego, pueden aumentar los ni-veles de riesgo.
Rosa y col. (21), estudiaron el parasitismogastrointestinal (PGI) en el Sudeste de la provin-cia de Buenos Aires, aunque no ofrecieron preci-siones señalaron que entre los meses entre ju-nio y octubre el predominio es de Trichostrongylusspp., Nematodirus spp., y Cooperia spp., siendo re-gular la abundancia de Ostertagia spp. en todo elaño.
Nuestra experiencia de estudios no siste-máticos coloca a Haemonchus contortus como laespecie prevalente desde mediados de verano yhasta bien entrado el otoño, y como responsablede los mayores trastornos. Las especies deestrongilidos de menor potencial biótico y mayor
adaptación invernal tienden a acumularse duran-te el otoño e invierno produciendo picos a fines deinvierno en los que tienden a presentarse Trichos-trongylus spp, Cooperia spp., Nematodirus spp. y enmenor medida Ostertagia spp. y Teladorsagia spp.
Figura 2. Modelo de infección de pasturas de zo-nas templadas ocupadas en forma permanente porlanares (adaptado de Mc Ewan 1994).Figure 2. Pasture-animal infection model, in permenetsheep occupation systems of temperate areas. (adapted fromMc Ewan 1994)
Al final de la preñez y durante la lactancia se produce el“alza de lactancia” (1), lo que genera un aumento en laoferta de larvas en poco tiempo (2). Esa población renovadaes la fuente principal de infección para los corderos, ha-cia fines de la lactancia y especialmente a partir del deste-te si se produce en el mismo ambiente(3). Los corderosjóvenes son altamente susceptibles y los parásitos tienenuna elevada producción de huevos que genera una mayorcontaminación de los pastos (4) la que, dependiendo de lascondiciones ambientales y del potencial biótico de las es-pecies dominantes, producen en una o dos generacionesuna elevada oferta de larvas infectantes que es responsa-ble de las mayores cargas parasitarias de los animales (5).
“Peripartum race” is coming on (1) later in pregnancy andin milking time and increace L3 disponibility in shorttime (2). This renovated larva population is source of lambinfection, later in lactation and after weaning (3). Younglambs are highly susceptible and the egg output is increas-ing (4).This, develope a high L3 disponibility after one ortwo generations, in depending on environment conditionsand predominant especies and led to major parasitic bur-dens in animals (5).
Figura 3: Tendencia de la distribución de la cargaparasitaria de larvas en las pasturas para la Pam-pa HúmedaFigure 3: Parasitic burden distribution in Humid Pampapastures
Haemonchus contortus adquiere su mayor proporción desdefin de verano y durante el otoño. Afecta a borregos de dien-tes de leche, y ovejas, especialmente cuando la parición esde abril-mayo. Con el correr de los meses, hacia fines deinvierno y principios de primavera predominan en la com-posición de las cargas los géneros Trichostrongylus, Nema-todirus y Cooperia, en mucho menor medida Teladorsagiacircumcincta y Ostertagia spp. que constituyen fuente deinfección de ovejas adultas de parición de primavera. Estepatrón tiende a modificarse o atenuarse en explotacionesmixtas con menor relación ovinos/bovinos.Haemonchus contortus frequency is highest later in sum-mer and early autum, affecting after-weaning lambs andpregnant sheep (autum parturition). In later winter andearly spring Trichostrongylus spp, Nematodirus spp andCooperia spp. are dominants and low number of overTeladorsagia circumcincta and Ostertagia spp.are pressent.They are sourse of infection for adults sheep of springparturition. Intensity of values in this pattern may changein farms with smaller ovine/bovine ratio.
(Figura 3).Balbi (14) (en INTA Castelar) demostró que
la evolución exógena de los huevos y larvas deHaemonchus contortus se interrumpe durante elinvierno siendo también mínima la superviven-cia de larvas y traslación en julio y agosto en laProvincia de Buenos Aires. En el Uruguay, Nari ycol. (22), realizaron un estudio sobre dinámica delPGI. Ellos encontraron un pico de parasitismo porHaemonchus contortus en borregos y en ovejas en-tre los meses de febrero y mayo. En borregos«tracers» de 6 a 12 meses, describen sin embargoun pico entre junio y septiembre, demostrandopara ese período la mayor tendencia a hipobiosisde este género. Aunque estos datos no son de laArgentina orientan el entendimiento de lo que ocu-rre en esa latitud.
Los primeros trabajos sistemáticos se rea-
El Sitio Argentino de Producción Animal
8 de 17

Gastroenteritis ovina
29ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
lizaron en La Pampa. En un conjunto de estudiosdetallados de seguimiento en ovejas y en corde-ros, describió la dinámica del PGI, en la RegiónSemiárida Pampeana (4, 5). De la necropsia deborregos tracers y naturalmente infectados surgea lo largo del año el predominio de Haemonchuscontortus ente enero y junio. Más lenta es la acu-mulación de otros géneros (Trichostrongylus, Ne-matodirus), que hacen pico entre los meses deagosto y setiembre. En el caso de este último pro-bablemente como consecuencia de la sobreviven-cia de larvas durante el invierno, fenómeno esteque también pudo ser observado en el Uruguay encorderos que pastorearon pasturas contaminadaspor borregos durante el otoño (24). En La Pampalas lluvias son estacionales, secos los inviernos yprimavera temprana, además desde fines de oto-ño no se dan las temperaturas suficientes para laevolución y mantenimiento de los estadíos am-bientales de H.contortus. En ovejas (7) se repite elmismo patrón con un pico a fines de primavera(asociado al parto). Las especies de Ostertagia noson prevalentes en las infecciones de esa región.El modelo de tendencias de disponibilidad de lar-vas puede resumirse gráficamente en la figura 4.
En la provincia de Corrientes Rosa y col. (25),indican una tendencia al aumento de la elimina-ción de huevos por parte de corderos entre noviem-
bre y junio con predominio de Haemonchus spp.,Desde fines de invierno la tendencia se reviertey para primavera se determina mayor prevalen-cia de Trichostrongylus spp.
Trabajos inéditos en Corrientes (Vasquez,com. pers) y nuestra experiencia en seguimientode majadas de la región mesopotámica permitendetectar que la estacionalidad es menos marcadaque en la Pampa Húmeda a medida que nos acer-camos desde el sur hacia el centro de la provinciade Corrientes. Las temperaturas medias de in-vierno no son suficientemente frías como paraimpedir la sobrevida de Haemonchus contortus, queestá presente todo el año en los recuentos de hue-vos y en las necropsias de animales de todas lasedades, aunque manteniendo el predominio defines de verano y principios de otoño. Tambiénlas especies de Trichostrongylus, si bien resultande lenta acumulación hasta fines de invierno yprimavera, se encuentran todo el año y con me-nor proporción en verano. Durante la primaveraaparecen otros géneros como: Oesophagostomum,Nematodirus y Cooperia (especialmente en corde-
Figura 4. Tendencia de la distribución de la cargaparasitaria de larvas en las pasturas para la re-gión Semiárida.Figure 4. Parasitic burden distribution in Humid Pampapastures
Definida con mayor precisión, está muy influenciada porlas lluvias estacionales de primavera a otoño, condicio-nando la evolución de haemonchosis. Durante la primave-ra las cargas predominantes son de Trichostrongylus, Ne-matodirus y en menor medida Teladosrsagia circuncincta. Losinviernos secos separan marcadamente estos dos períodosy grupos de parásitos. Las ovejas de parición de primaverason fuente de infección de pasturas.
In this area Haemonchus contortus burdens are conditionedby seasonality of spring and autumnal rainfall. In spring,Trichostrongylus spp. and Nematodirus spp. are predomi-nant, and Teladorsagia spp. is in low level. Dry wintersdivide this two periods and parasites groups. Spring lamb-ing sheep are source of spring pasture infections.
Figura 5. Tendencia de la distribución de la cargaparasitaria de larvas en las pasturas para el nor-te de la Mesopotamia.Figure 5. Parasitic burden distribution in HumidMesopotamic pastures
En el Nordeste, las temperaturas son más elevadas y elrégimen pluviométrico mayor que en la región pampeana,especialmente desde mediados de verano y en otoño. Conello los niveles potenciales de contaminación por Haemon-chus contortus en las pasturas son máximos, y no puedeesperarse una disminución importante ni aún en invier-no. Entre las especies de menor potencial biótico, el Tri-chostrongylus colubriformis tiene mayor presencia hacia fi-nes de invierno y durante la primavera. En general todo elaño pueden encontrarse infecciones clínicamente impor-tantes, mixtas o con predominio de Haemonchus contortus.
Temperatures and rainfall are higher in North East regionthan pampean area. Weather is humid specially in sum-mer and autumn. Haemonchus contortus contamination inthis two seasons is maximum, and it’s reduction is notimportant not even in winter. Trichostrongylus colubriformisis important later in winter and during spring, and lessabundant are Nematodirus spp. Oesophagostomum spp andCooperia spp. Any time in the year, clinically high bur-dens with Haemonchus contortus predominance may oc-cur.
El Sitio Argentino de Producción Animal
9 de 17

J. Romero y col.
30 ANALECTA VETERINARIA 2001; 21, 1: 21-37 ISSN 1514259-0
ros). Resulta necesario contar con trabajos com-pletos en la región mesopotámica y en la provin-cia de Buenos Aires, que resuman la dinámicadel PGI, en función de mejorar los programas decontrol (Figura 5).
El elevado nivel de infección, la estacionali-dad menos marcada en la distribución de la ofertade larvas y la mayor relación ovinos/bovinos enestablecimientos correntinos, dificulta la com-prensión del fenómeno por parte de los producto-res, que realizan estos tratamientos sin contem-plar diferencias en la composición de la poblaciónde parásitos en cada época, en el espectro de lasdrogas utilizadas, o en la susceptibilidad de lasdiferentes categorías de hospedadores (hasta 6 y7 tomas anuales en algunos casos).
RESISTENCIA A ANTIHELMÍNTICOSDesde 1957, cuando se comunicó por prime-
ra vez la resistencia a la Fenotiazina en EE.UU., yluego en 1964 al Thiabendazole en Haemonchuscontortus, las citas se han reproducido en todo elmundo (26). El manejo irracional de medicamen-tos, sumado a la mala práctica de los tratamien-tos (subdosificación por fallas en el instrumental,la estimación de peso de los animales o mala ca-lidad de los productos), han conducido a la selec-ción de cepas con resistencia a antihelmínticos(RA) como en otras partes del mundo. En nuestromedio se comunicó por primera vez la sospechade un caso en 1988 (27), haciéndose la primeracomprobación en 1993 (28), ambos casos en la Pro-vincia de Buenos Aires. La aparición e intensi-dad de este fenómeno está en relación directa conla intensidad de los tratamientos. La RA es unfactor limitante de la producción en los países don-de la cría del lanar está más desarrollada. A nivelnacional en más del 60 % de las majadas se pre-senta resistencia a algún grupo químico (29, 30).
En la Mesopotamia (31) comprobaron que el90 % de las majadas presenta resistencia a losbencimidazoles, en la mitad de los casos con efi-cacias inferiores al 50 %. El 73 % de los estableci-mientos presentan resistencia a dos o más gru-pos químicos. Las parásitos más frecuentementeinvolucradas fueron Trichostrongylus spp. y Hae-monchus spp.
En Uruguay, donde los estudios de resisten-cia tienen más antigüedad el 92,5 % de los esta-blecimientos presentan algún nivel de resisten-cia a alguno de los grupos. Se ha comprobado re-sistencia de Haemonchus contortus a todos los gru-pos de drogas de amplio espectro y, hasta 1996, sehabía informado resistencia a ivermectinas en
Trichostrongylus y Ostertagia (32).El problema también es evidente en el sur
de Brasil (33) donde la subdosificación de drogasdurante muchos años tuvo origen, en parte, enlas propias indicaciones de los laboratorios pro-ductores de medicamentos de aplicar por debajode los niveles recomendados internacionalmen-te. En Paraguay la alta prevalencia de Haemon-chus contortus durante casi todo el año genera altafrecuencia de tratamientos (34), lo que ha contri-buido a seleccionar cepas resistentes a todos losgrupos de amplio espectro.
Naturaleza de la resistencia a antihel-mínticos
Los antihelmínticos se distribuyen en cin-co grupos según sus mecanismos de acción: elgrupo I, lo componen los bencimidazoles yprobencimidazoles, incorporados al mercado en losaños ‘60. Su mecanismo de acción se relacionacon la disrrupción de los microtúbulos y husosmitóticos, fijándose a la tubulina. Su efectividadrequiere de un prolongado tiempo de acción hastaque esa interferencia termine por matar al pará-sito, y eso es clave para los diferentes estadíos delos helmintos particularmente para formas inhi-bidas. Se presentan para aplicación oral, intrarru-minal o subcutánea, tienen efectividad contranematodes adultos y en dosis mayores contra lar-vas inhibidas.
El grupo II, reúne al Morantel, Pirantel y elLevamisol (cuyo efecto es limitado frente a for-mas inhibidas de Ostertagia spp.). Si bien no esclaro el mecanismo exacto de acción se colocanjuntos debido a la existencia de resistencia cru-zada entre ellos. Estas drogas producen una pará-lisis espástica de los helmintos que no los mataen forma inmediata, pero facilita su expulsión.
El grupo III las Avermectinas y Milbemicinas(Moxidectin), Doramectina y Epiromectina. Variosmecanismos de acción se han descripto para es-tas drogas en relación a la interacción con recep-tores de los canales de cloro y con algunos trans-portadores de membrana. Estas drogas matan len-tamente a los nematodes y no poseen poder ovici-da.
El grupo IV está conformado por losorganofosforados como el Triclorfón y el Naftalofós(cuyas dosis terapéuticas suelen tener efectivi-dad limitada, y resultan tóxicos en forma aguda ycrónica a veces en niveles de dosis cercanos a losindicados).
En el grupo V se encuentran, entre otros,
El Sitio Argentino de Producción Animal
10 de 17

Gastroenteritis ovina
31ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
las Salicilanilidas entre las que son conocidas elClosantel y el Rafoxanide. El espectro de Closanteles limitado a nematodes hematófagos, pero tienela propiedad de proteger contra la infección deHaemonchus spp. por más de 50 días si se lo usaen dosis elevadas (12 mg/kg peso vivo) ya que semantiene ligado a proteínas plasmáticas.
Algunos individuos en las poblaciones deparásitos tienen aptitud para evitar que las dro-gas los maten (esos mecanismos son muy varia-dos y no son igualmente frecuentes en cada espe-cie).
Se ha demostrado la naturaleza genética dela resistencia y se han identificado en algunoscasos con precisión las mutaciones del genomade los helmintos resistentes respecto de los sus-ceptibles a diferentes drogas (35). Por ejemplo, elefecto de los bencimidazoles (BZ) sobre los orga-nismos es la interferencia en la fijación de tubu-lina en el ensamble de los microtúbulos. El geno-ma de H. contortus tiene un locus específico parael Isotipo 1 de β-tubulina con muchas variantesalélicas en poblaciones susceptibles a losbencimidazoles. El incremento en la frecuenciade uno de esos alelos tiene una fuerte correla-ción con el aumento en la resistencia de las ce-pas. Se han identificado cambios en aminoácidosen diferentes posiciones de la estructura de la β-tubulina (36); y se ha comprobado que las muta-ciones son similares en la estructura de la β-tu-bulina de los microtúbulos de otros nematodes yhongos resistentes a bencimidazoles (37).
Si se repiten con frecuencia los tratamien-tos con el mismo grupo químico sobre la poblaciónde parásitos que contenga a esos individuos, enpoco tiempo predominará la descendencia de losque han soportado al medicamento, su caráctergenético habrá sido seleccionado y multiplicado.Generalmente cuando en una población se desa-rrolla la resistencia a una droga, la cepa resultaresistente a todo el grupo.
El diagnóstico clínico es evidente cuando elnivel de resistencia a las drogas es grave y lamayoría de los parásitos sobrevive a los tratamien-tos. Recién entonces el productor percibe la fallay generalmente cambia de grupo químico aunquesea por un tiempo. La repetición del proceso conel nuevo grupo tiende a seleccionar nuevamenteindividuos resistentes, en este caso al nuevo gru-po. Cuando la resistencia es “cruzada” (a dos o másgrupos), el problema se complica porque se redu-cen las alternativas de control. Debe destacarseque la reversión del proceso es muy poco probable
ya que no puede presionarse en forma inversa yuna vez establecido el genotipo con elevada fre-cuencia en la población difícilmente se erradique.Se ha comunicado una reducción en los nivelesde resistencia a Levamisol luego de interrumpirlos tratamientos, pero en general son controver-tidas las comunicaciones en ese sentido y en es-pecial respecto a bencimidazoles no habiendo evi-dencias experimentales de tal regresión (26).
Si bien para que el proceso de selección decomienzo es necesaria la preexistencia de indi-viduos resistentes, los contactos con dosis meno-res a las indicadas para cada producto pueden fa-cilitar la selección de individuos portadores degenes relacionados con la resistencia, pero conexpresión fenotípica de menor intensidad, contri-buyendo a una mayor frecuencia posterior de di-chos genes.
Subdosificación:Surge por fallas en el cálculo del peso de los
animales a tratar, defectuoso funcionamiento delos dosificadores y especialmente por fallas en laadministración. Los productos inyectables debenaplicarse en áreas libres de lana (cara internadel muslo) ya que suele no penetrar correctamentela piel en individuos con abundante lana. Estosucede cuando la aplicación la realiza personalno entrenado debidamente.
Con las dosificaciones orales el problema esdiferente, la permanencia de las drogas en el tubodigestivo guarda correlación con el mantenimien-to de niveles plasmáticos en el tiempo. Se ha de-mostrado que el nivel de llenado del rumen afectala velocidad del pasaje de los productos y con ellola disponibilidad.
El ayuno de 12 horas previo a la dosificaciónoral permite una mayor retención de droga en elrumen y un lento pasaje del contenido ruminalcon lo que la disponibilidad y absorción del pro-ducto resulta más lenta, y los niveles plasmáticosde concentración son más prolongados, permitien-do una exposición más prolongada de los parási-tos (38). Las estimaciones de resistencia realiza-das por el test de reducción de recuento de hue-vos varían si los tratamientos se realizan en con-diciones de ayuno previo. Cuando los productosno tienen la concentración adecuada del elemen-to activo, o cuando la formulación no permite al-canzar correctamente su biodisponibilidad luegode aplicado, también se incurre en sub dosifica-ción.
El comienzo del proceso de selección de pa-
El Sitio Argentino de Producción Animal
11 de 17

J. Romero y col.
32 ANALECTA VETERINARIA 2001; 21, 1: 21-37 ISSN 1514259-0
rásitos resistentes en general es el descripto, peroen un campo también pueden ingresar las cepasresistentes: con el ingreso de animales prove-nientes de otros establecimientos, reproductoresde cabaña, compra de núcleos, o liquidaciones demajadas, arrendamientos de pastoreos, etc. (even-tos muy frecuentemente implicados en Sud Afri-ca) (Van Wyk J.A. com. pers).
Población en refugio:Cuando se aplica un tratamiento antipara-
sitario se afecta con él a la porción de la poblaciónque se encuentra en los animales. Según la si-tuación epidemiológica en ese momento, es posi-ble que un número mayor de individuos se en-cuentre en estado de huevo, o larva en el suelo yno se exponga a la presión del medicamento. Cuan-do los sistemas de manejo de tratamientos ejer-cen gran presión, por su frecuencia y por involu-crar a todos las categorías de hospedadores, la po-blación en refugio se ve reducida, y el impacto dedicha presión es mayor. Cuando ante la apariciónde síntomas en algunos animales, los producto-res tratan toda la majada es posible que expongana las drogas de amplio espectro a poblaciones deparásitos cuyo número no afectan la salud o laproducción en todos los individuos. Incluso expo-ne a especies que componen la fauna parasitariapero que no requieren ser eliminadas. En estasespecies puede aparecer resistencia sin que sehubiera justificado tratarlas. Existen sistemas es-tratégicos de control que, a pesar de ser raciona-les, pueden presionar la selección hacia la resis-tencia sólo por ser relativamente fijos (ver másadelante).
Un sistema de protección de población enrefugio lo constituye el programa “FAMACHA” de-sarrollado en Sud Africa. Luego de evaluar pormedio del entrenamiento de los operadores, lacorrelación entre el hematocrito y el grado de pa-lidez de las conjuntivas de animales infectadoscon Haemonchus spp. se establecen rutinas de con-trol visual en las cuales se decide la dosificaciónsólo de los individuos con mayores grados de ane-mia. Las apreciaciones se realizan frecuentemen-te (cada 15 días). Si bien supone pérdidas subclí-nicas y un gran esfuerzo del personal, es una pro-puesta interesante en áreas con altos niveles deresistencia como las que se encuentran en aquelpaís y, por ejemplo, en Paraguay resultando apli-cable por lo menos en majadas chicas y la rutinapueden ejercerla campesinos sólo con el entre-namiento adecuado (39). Este método de control,que en realidad podría constituirse en una rutinapara la selección de individuos tolerantes enáreas de predominio de Haemonchus, tiende a rea-
lizar sólo tratamientos selectivos a mantener ele-vada la población en refugio de los antiparasita-rios.
Diagnóstico de estatus de resistenciaen cada establecimiento:
La situación general sobre resistencia encada región ofrece pocas variantes mientras lascondiciones de manejo sean comparables entreestablecimientos. Hoy en día las majadas debensometerse a tratamientos que sean efectivos, nosólo para evitar el efecto de los parásitos, sino parano complicar la disponibilidad de drogas eficaces.
Si bien para establecer con precisión losniveles de resistencia pueden realizarse pruebasque incluyan la necropsia de animales tratados,se han utilizado los ensayos de reducción del re-cuento de huevos en materia fecal de animalestratados con mucho éxito en encuestas regiona-les y en el monitoreo de establecimientos bajocontrol. Uno de ellos es el modelo de Presidente,descripto por Nari (40) y de aceptación generaliza-da, según el cual el día “0” se tratan 10-15 anima-les con cada droga o dosis a evaluar, tomándosemuestras individuales de materia fecal de cadagrupo y de un grupo control, y realizándose el re-cuento de huevos por gramo de materia fecal, ycoprocultivo para estimar la composición genéri-ca de la carga en cada grupo. El día 10 se vuelvena muestrear todos los individuos tratados y los delgrupo control, para realizar nuevamente el estu-dio de huevos por gramo, coprocultivo. La estima-ción de la eficacia expresada como porcentaje dereducción del recuento de huevos (RRH%), se rea-liza con la fórmula:
RRH% = {1 - (T2 /C2) x (C1 /T1 )} x 100
donde C y T son las medias geométricas delos HPG de los grupos “control” y “tratados” respec-tivamente, representando los números 1 y 2 lacorrespondencia a los días “0” y “10” del ensayo.
Cuando el valor medio de reducción es me-nor al 95 %, o cuando el extremo inferior del in-tervalo de confianza (error µ < 5 %) está por debajodel 90 % puede fundarse la sospecha de resisten-cia para ese grupo químico y dosis, y para ese gé-nero parasitario.
Con esta metodología se han realizado losrelevamientos y elaborado estrategias de manejoen todo el mundo. Con un método de cálculo aúnmás simple (comparación de medias aritméticas)se evaluó la situación en el Cono Sur (Argentina,Uruguay, Brasil y Paraguay) utilizando un progra-
El Sitio Argentino de Producción Animal
12 de 17
Gastroenteritis ovina
33ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
ma denominado RESO (anon.) elaborado por CSIROen Australia.
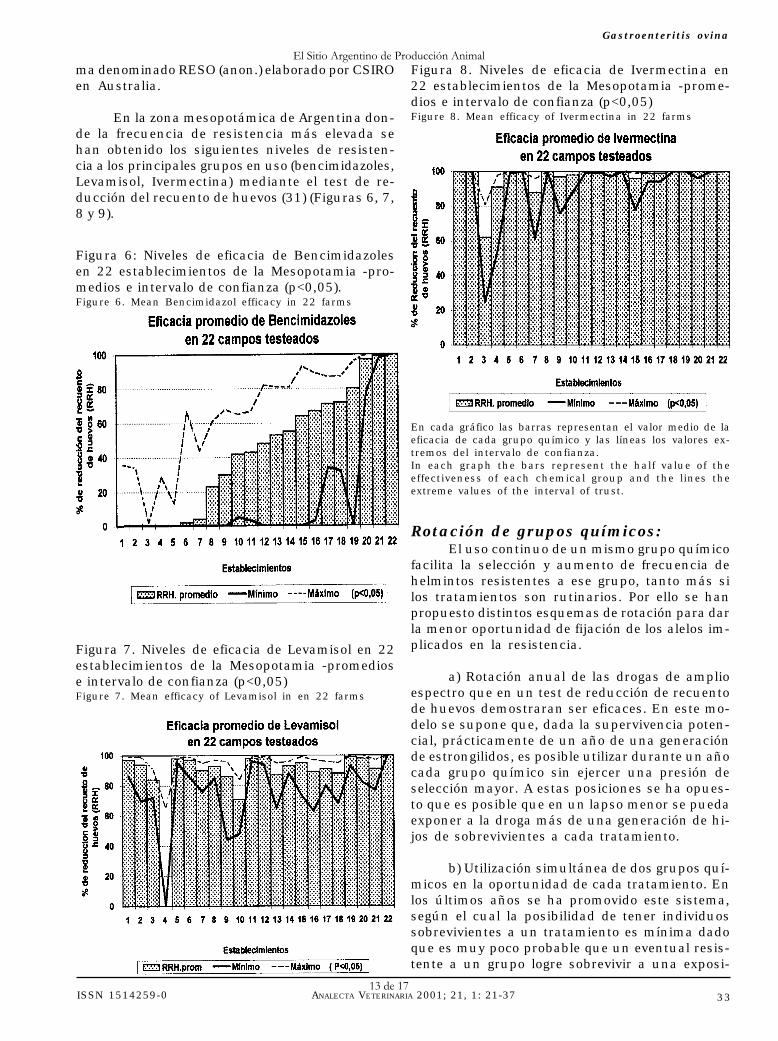
En la zona mesopotámica de Argentina don-de la frecuencia de resistencia más elevada sehan obtenido los siguientes niveles de resisten-cia a los principales grupos en uso (bencimidazoles,Levamisol, Ivermectina) mediante el test de re-ducción del recuento de huevos (31) (Figuras 6, 7,8 y 9).
Rotación de grupos químicos:El uso continuo de un mismo grupo químico
facilita la selección y aumento de frecuencia dehelmintos resistentes a ese grupo, tanto más silos tratamientos son rutinarios. Por ello se hanpropuesto distintos esquemas de rotación para darla menor oportunidad de fijación de los alelos im-plicados en la resistencia.
a) Rotación anual de las drogas de amplioespectro que en un test de reducción de recuentode huevos demostraran ser eficaces. En este mo-delo se supone que, dada la supervivencia poten-cial, prácticamente de un año de una generaciónde estrongilidos, es posible utilizar durante un añocada grupo químico sin ejercer una presión deselección mayor. A estas posiciones se ha opues-to que es posible que en un lapso menor se puedaexponer a la droga más de una generación de hi-jos de sobrevivientes a cada tratamiento.
b) Utilización simultánea de dos grupos quí-micos en la oportunidad de cada tratamiento. Enlos últimos años se ha promovido este sistema,según el cual la posibilidad de tener individuossobrevivientes a un tratamiento es mínima dadoque es muy poco probable que un eventual resis-tente a un grupo logre sobrevivir a una exposi-
Figura 6: Niveles de eficacia de Bencimidazolesen 22 establecimientos de la Mesopotamia -pro-medios e intervalo de confianza (p<0,05).Figure 6. Mean Bencimidazol efficacy in 22 farms
Figura 7. Niveles de eficacia de Levamisol en 22establecimientos de la Mesopotamia -promediose intervalo de confianza (p<0,05)Figure 7. Mean efficacy of Levamisol in en 22 farms
Figura 8. Niveles de eficacia de Ivermectina en22 establecimientos de la Mesopotamia -prome-dios e intervalo de confianza (p<0,05)Figure 8. Mean efficacy of Ivermectina in 22 farms
En cada gráfico las barras representan el valor medio de laeficacia de cada grupo químico y las líneas los valores ex-tremos del intervalo de confianza.In each graph the bars represent the half value of theeffectiveness of each chemical group and the lines theextreme values of the interval of trust.
El Sitio Argentino de Producción Animal
13 de 17

J. Romero y col.
34 ANALECTA VETERINARIA 2001; 21, 1: 21-37 ISSN 1514259-0
ción simultánea a otro grupo químico. Esa expec-tativa es relativa cuando se enfrentan poblacio-nes donde la resistencia está ya establecida a cadauna de las drogas. En la figura 9 puede observarseel cuadro de situación de Bencimidazoles y de Le-vamisol cuando se aplicaron en forma simultá-nea en los mismos establecimientos descriptoscon anterioridad (31).
inicio de cada temporada tratando de impedir lafundación de poblaciones parasitarias de tenden-cia estacional (especialmente Haemonchuscontortus). Estos programas “Wormkill” o“Drenchplan” impuestos por el CSIRO (43, 44), convariantes según la región de aplicación, se basa-ron en el conocimiento previo de la dinámica delas especies parasitarias y buscaron el aprovecha-miento racional del espectro de las drogas, comoel Closantel, que aplicado en dosis elevadas cuentacon un período prolongado de acción sobre Hae-monchus spp. Cada dosificación con Closantel (12mg/kg) elimina la población de adultos en los ani-males y evita la reinfección por casi dos meses;con 3 a 4 semanas más de período prepatente, seinterrumpe la siembra de huevos por prácticamen-te 3 meses. En regiones con prevalencia de esteparásito a lo largo de todo el año, 4 tratamientosanuales tenderían teóricamente a erradicarlo. Enzonas donde la tendencia sea marcadamente es-tacional pueden programarse otras rutinas quecontemplen este uso de Closantel en momentosestratégicos. En cada tratamiento, el uso simul-táneo o alternativo de otras drogas mejorará porun lado el efecto de la primera y se orientará alcontrol de otras especies. Estos programas de tra-tamientos estratégicos han sido exitosos dismi-nuyendo al mínimo la expresión poblacional de lasespecies implicadas, con un mínimo de dosifica-ciones. Esquemas de 4 tratamientos anuales handemostrado su eficacia en el manejo de poblacio-nes expuestas a Haemonchus contortus en Uruguay(45), pero se debe considerar que al dejar una muyreducida población “en refugio”, y obviamente nolograr el exterminio de los parásitos, son procli-ves a seleccionar muy eficientemente cepas re-sistentes.
Pastoreo alterno con bovinos u otrasespecies.
Los lanares no comparten sus principalesparásitos con los bovinos, Haemonchus contortuspuede desarrollar en bovinos con dificultad, y Os-tertagia ostertagi desarrolla con mucho menos efi-ciencia en el lanar. Las especies de Cooperia pue-den ser compartidas y componer parte importan-te de la infección en corderos que pastorean po-treros donde lo hicieron antes terneros (45). Enregiones templadas se ha demostrado que la rota-ción programada puede disminuir la carga en lospastos de larvas de especies de riesgo en Uruguay(46). También en Uruguay Castells y col. (com. per-sonal), estudiaron la evolución de peso y HPG decorderos a partir del destete y pastoreando en po-treros que en primavera fueron utilizados pormajada de cría, capones o novillos. Demostraron
Figura 9. Niveles de eficacia de Bencimidazoles yLevamisol utilizados en forma simultánea, en 22establecimientos de la Mesopotamia -promediose intervalo de confianza (p<0,05)Figure 9.Mean efficacy of mix (BZM+LVM) in 22 farms
En estos casos era conocida la resistencia acada uno de los grupos químicos y el cuadro desituación cambia sólo parcialmente. En la actua-lidad la propuesta de manejo simultáneo, se orientaal uso de drogas de conocida eficacia al iniciarseel programa. Modelos de simulación desarrolladosen Australia (42) hacen suponer que la utiliza-ción simultánea de dos grupos químicos evolucio-na más lentamente hacia la resistencia a cadauno de ellos que si se rotaran anualmente. Noobstante la estabilidad de estos sistemas fijos deadministración de medicamentos combinados notiene aún validación experimental o de campo porlo que requeriría monitoreo permanente.
OTROS RECURSOS DE MANEJOProgramas estratégicos de dosifica-ción
Durante muchos años en Australia y enotros lugares se promovieron sistemas de despa-rasitaciones de gran presión de tratamiento en el
El Sitio Argentino de Producción Animal
14 de 17

Gastroenteritis ovina
35ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
en un informe preliminar que es posible que losHPG se mantengan en niveles entre 5 y 8 vecespor debajo y las ganancias de peso fueran de un15 y un 25 % mayores en lotes cuyo antecesorfueron novillos respecto de lotes que pastorearonparcelas en las que pastorearon antes majadasde cría o capones. Esta propuesta puede llevarse acabo a campo.
Selección de individuos tolerantes y re-sistentes:
Se explicó el significado de esos términos.Ambas características son heredables y segreganen forma independiente. Puede emprenderse laselección por resistencia en base a la evaluaciónde niveles de recuentos de huevos tanto en ma-chos como hembras de majada y evaluar ambasvariables para machos del núcleo o cabaña. Parael manejo a campo la herramientas aún no estántotalmente desarrolladas desde lo metodológico. Semencionó el “FAMACHA” como método de control,la susceptibilidad de los animales puede medirsepor el hematocrito en el caso de exposición a lahaemonchosis.
Puede demostrase en poblaciones no selec-cionadas que el 25 % de los animales pueden ge-nerar mas del 75 % de la contaminación con locual queda absolutamente justificada la segrega-ción de estas poblaciones susceptibles.
Teniendo en cuenta que la resistencia ytolerancia a los parásitos son una fundamentalvariable de adaptación al ambiente, constituyenobjetos de selección importantes además de losproductivos.
Las rutinas de selección pueden variar entorno a la evaluación de los niveles de HPG entretratamientos a partir del destete y en varios de-safíos. En hembras de reposición podría ser sufi-ciente para no retener susceptibles. La presiónde selección en los núcleos de carneros puede sermayor que en los de hembras, y la tolerancia pue-de evaluarse con un manejo individual de trata-mientos y registrando parámetros como edad alprimer tratamiento requerido, número de trata-mientos recibidos, niveles de ganancia de peso(18). Presionando en la selección tanto de machoscomo de hembras en la reposición de la majadageneral y aplicando programas exigentes entre losindividuos del núcleo puede obtenerse el mayorimpacto que utilizando sólo carneros selecciona-dos, que por otra parte no se disponen en el país.(17).
MANEJO INTEGRADO Y CONCLUSIÓNEn general las comunidades dedicadas a la
explotación de lanares comparten los problemasligados al control de parásitos y todas han halladoen los medicamentos antiparasitarios el recursode mayor impacto. La resistencia a drogas anti-helmínticas quita la herramienta de mayor po-der. El enfoque integrado del manejo de las hel-mintiasis es la expresión con mayor presenciaen los foros internacionales y su sola menciónconvoca un gran consenso. Sin embargo es en lapráctica un punto crítico en la evolución de la pro-fesión veterinaria. Los recursos alternativos nopueden incluirse sin un actor que los integre enun sistema de decisiones que minimice el im-pacto de los parásitos sin dependencia total de laindustria farmacéutica.
Ese actor debe echar mano a todo el arsenalde recursos y procesos antes mencionados y com-binarlos racionalmente según las circunstanciasde cada majada y explotación Los programas deextensión de organismos oficiales, y las comuni-caciones de marketing de la industria no logranestablecer los procesos necesarios al manejo in-tegrado. Por otro lado esa integración debe incluirla consideración de selección de animales porcaracteres productivos, manejo de otras enferme-dades, manejo reproductivo, administración deintereses comerciales y, seguramente la asocia-ción de explotación de lanares con otras especies.
Todos los recursos modernos son de comple-jidad creciente y sólo los provenientes de la in-dustria se presentan acabados y listos para el usopor parte del productor. Los otros reclaman al pro-fesional para su administración, y exigen otrasestrategias de convencimiento:
* Elección del ciclo reproductivo de la maja-da según conveniencia real de la explotación (fe-chas según interés de venta o mejor adaptación ala oferta forrajera, conveniencia de selección demelliceras, disponibilidad de suplementos, etc.).Al manejo productivo, se subordinará el control detodos los problemas sanitarios en general y el pa-rasitario en particular, estableciendo épocas y si-tuaciones de riesgo. Esa subordinación es sinóni-mo de integración.
* Diagnóstico del estatus de resistencia yregistro de drogas y espectros disponibles.
* En explotaciones con un núcleo definido,establecimiento de un programa de selección queademás de caracteres productivos, contemple laselección de individuos resistentes y, de ser posi-ble, tolerantes a los parásitos en la medida que lavariación interna de la población lo permita, es-
El Sitio Argentino de Producción Animal
15 de 17

J. Romero y col.
36 ANALECTA VETERINARIA 2001; 21, 1: 21-37 ISSN 1514259-0
pecialmente segregación de individuos más sus-ceptibles en cualquiera de los estados de evolu-ción del programa.
* Diagnóstico parasitológico durante los pe-ríodos de mayor riesgo de infección y en las cate-gorías susceptibles.
* Diseño de estrategias de desparasitaciónestratégicas tendientes a mantener reducido elriesgo de categorías muy susceptibles y la infecti-vidad de las praderas y tácticos sobre la base dediagnóstico, para evitar la repetición innecesariade dosificaciones masivas o en animales que nolas necesitan.
* Consideración del pastoreo alterno con bo-vinos.
* Monitoreo de todos los subprogramas enfunción de no instituir sistemas que tiendan aseleccionar cepas resistentes a los medicamen-tos.
BIBLIOGRAFÍA1. Johnstone Ian L. Enfoque ecológico para el controlde la parasitosis ovina. INTA. Agropecuaria 1971. 20:113 páginas.
2. Lukovich R. Identificación de las formas adultas delos nematodos gastrointestinales y pulmonares de losrumiantes en la República Argentina. Publicación mis-celánea de INTA, 1981 Buenos Aires.
3. Suárez MC. Nematodirus oiratianus (Raevskaya 1929)en la República Argentina (Primer hallazgo). Rev InvAgr INTA, Bs.As. 1982; XVII (2): 169.
4. Suárez VH. Parasitosis gastrointestinal en ovinosCorriedale en la región semiárida pampeana (1) Re-sultados de los primeros 16 meses de observaciones.Rev Med Vet 1985; 66 (3): 140-149.
5. Suarez VH Parasitosis Gastrointestinal en ovinosCorriedale en la región semiárida pampeana. II resul-tados período 1981-1983. Rev Prod An. 1985; 5 (3-4):243-55.
6. Bueno L, Honde Ch, Luffau G, Fioramonti J. Originof the early digestive disturbances induced by Haemon-chus contortus infection in lambs. Am J Vet Res 198243 (7): 194/99.
7. Suarez VH. Epizootiología de los parásitos gastroin-testinales en ovejas en la región semiárida pampea-na. Rev Med Vet. 1986; 67 (4); 190-202.
8. Castells D, Nari A, Rizzo E, Mármol E, Acosta D.Incidencia de los nematodes gastrointestinales en laproducción de carne y lana. 3º encuentro de Veterina-rios endoparasitólogos rioplatenses. Arapey, Uruguay24 al 26 de mayo de 1994.
9. Kennedy CR. Ecological Animal Parasitology”Blackwell Scientific Publications. Oxford-LondonEdinburg- Melburne. 1975
10. Michel JF, Lancaster MB, Hong C. Arrested devel-opment of Ostertagia ostertagi, and Cooperia oncophora,
Effect of temperature at the living third stage. J ComphPath. 1975 85:133-138.
11. Fiel CA, Steffan PE, Vercesi HM, Ambrústolo RR,Catania P, Casaro AP, Entrocasso CM, Biondani CA.Variación estacional del parasitismo interno de bovi-nos en el sudeste de la provincia de Buenos Aires (Ar-gentina) con especial referencia al fenómeno de hipo-biosis”. Rev Med Vet (Bs.As.) 1988. 69 (1): 57-64.
12. Fernández AS, Fiel CA, . Estudio de Factores queinducen la hipobiosis de Ostertagia ostertagi, en bovi-nos. Premio R. Niec. Sociedad de Medicina Veterina-ria, Buenos Aires. 1996.
13. Nari A, Petraccia C, Cardozo H. La inhibición deldesarrollo larvario en nematodes gastrointestinalesde ovinos con especial referencia a Haemonchuscontortus. Veterinaria 1982. 18 (81): 78-88.
14. Balbi A. Ecología de la fase libre del ciclo de Hae-monchus contortus del ovino en la Pampa Húmeda. En“Aportes a la Parasitología Veterinaria por Eddi yCaracostantógolo 1993. (II). INTA CICV. Castelar.
15. Woolaston R, Greeff J. NEMESIS Merino BreedersWorm Control Network, recommendations for breed-ing Sheep for worm resistance Miscellaneous publica-tion of CSIRO- UNE, WADA, y la Wool research andpromotion Organization Australia, 1994.
16. Albers GA, Gray GD, Piper LR, Barker JSF, Le JambreLF, Barger IA. The genetics of resistance and resil-ience to Haemonchus contortus infection in young me-rino sheep. Int J for Parasitol 1987. 17 (7):1355-1363.
17. Romero J, Boero CA, Prando AJ, Baldo A, Griffin B,Silvestrini MP. Selection of trichostrongylid resistantsheep in argentine farms. 17th International confer-ence of the World Association for the advancement ofVeterinary Parasitology . Copenhagen 15-19 august1999 Abst.f.304.
18. Morris CA, Bisset SA. Breeding Sheep which re-quire minimal anthelmintic treatment: A review of thegenetics of resistance and resilience of sheep to nema-tode parasites In Managing Anthelmintic Resistancein Endoparasites. Workshop held at the 16th Interna-tional conference of de World Association for the ad-vancement of Veterinary Parasitology Sun City 10-15august. van Wyk. Schalkwyc ed. 1997: 21-28.
19. Coop RL, Kyriazakis I. Nutrition-parasite interac-tion. Vet Parasitol 199; 17 (7):187-204
20. McEwan J. Breeding sheep resistant o roundworminfection: Wormfec Beeder’s Manual. edited by NewZealand Pastoral Agriculture Research Institute. 1994.
21. Rosa W, Lukovich R, Niec R. Parasitismo -gastro-intestinal de los ovinos y bovinos en la zona sur de laProvincia de Buenos Aires (Tres Arroyos, Cnel. Pringles,y Cnel. Dorrego). Rev Inv Agrop INTA. Serie 4. Pat. An.1971. VIII (3): 71-83.
22. Nari A, Cardozo H, Berdie J, Canábez F, Bawden R.Dinámica de población para nematodes gastrointesti-nales de ovinos en el Uruguay. Veterinaria 1977. 14(66): 11-24.
El Sitio Argentino de Producción Animal
16 de 17

Gastroenteritis ovina
37ANALECTA VETERINARIA 2001; 21, 1: 21-37ISSN 1514259-0
23. Suarez VH, Larrea S, Busetti MR, Bedotti DO,Bulman GM, Ambrustolo RR. Nematodes Gastrointes-tinales de ovinos: Su control y efectos sobre los pará-metros epizootiológicos y productivos en la región se-miárida pampeana (Argentina). Therios 1990. 15 (73):156-173.
24. Nari A, Cardozo H, Rizzo E, Solari MA, Petraccia C.Efecto del parasitismo gastrointestinal en la perfor-mance de corderos sometidos a diferentes planos denutrición y edad de destete. Veterinaria 1983. 29 (85):57-63
25. Rosa W, Niec R, Lukovich R, Dindart J, BarberanR. Etiología y variación estacional del parasitismo gas-trointestinal de los ovinos y bovinos en la zona deMercedes (Provincia de Corrientes). Rev Inv Agrop INTA.Serie 4 Pat. An. 1973: X (1): 25-40.
26. Borgsteede FHM, en Coles GC, Borgsteede FHM,Geerts S. Anthelmintic resistance in Nematodes offarm animals. Editing of Seminar in the European Com-mission of coordination for agricultural research, Brus-sels Belgium 8/9 November. 1993:1-16.
27. Entrocasso C, Lange R, Fernández Ripoll. Resis-tencia antiparasitaria al oxfendazole en ovinos en unestablecimiento de la provincia de Buenos Aires. Me-morias del VI Congreso Arg de Ciencias Vet. BuenosAires. 1988. Com. libre 208.
28. Romero JR, Espinosa G, Valera AR, Demostraciónde Resistencia al Oxfendazole en Trichostrongylidosde ovinos de la Zona deprimida del Salado. Rev de MedVet. 1992. 73 (2): 82-86.
29. Eddi C. Resistencia. Memorias del VII CongresoArgentino de Cs. Veterinarias Buenos Aires 6-11 deNoviembre de 1994
30. Eddi C, Caracostantogolo J, Peña M, Shapiro J,Marangunich L, Waller O, Hansen JW. The prevalenceof anthelmintic resistance in nematode parasites ofsheep in southern Latin America: Argentina.” VetParasitol 1996. 62 : 89-197.31. Romero J, Boero C, Vazquez R, Aristizábal MT, BaldoA. Estudio de resistencia a antihelmínticos en maja-das de la Mesopotamia. Rev Med Vet. 1998; 79 (5):342-346.
32. Nari A, Salles J, Gil A, Waller PJ, Hansen JW. Theprevalence of anthelmintic resistance in nematodeparasites of sheep in Southern Latin America: Uru-guay. Veterinary Parasitology 1996. 62: 213-222.
33. Echeverría F, Borba MFS, Pinheiro AC, Waller PJ,Hansen JW. The prevalence of anthelmintic resistancein nematode parasitesof sheep in Southern LatinAmerica: Brazil. Vet Parasitol 1996. 62:199-206.
34. Maciel S, Gimenes AM, Gaona C, Waller P, HansenJW. The prevalence of anthelmintic resistance in nema-tode parasites of sheep in southern Latin America:Paraguay Vet Parasitol 1996. 62: 207.212.35. Roos MH. The role of drugs in the control of para-sitic nematode infections: must we do without?”. Para-sitology 1997; 114: 137-144.
36. Kwa MSG, Veenstra JG, Roos MH. Molecular char-acterization of β-tubulin genes present in benzimida-zole resistant populations of Haemonchus contortus.Molecular and Biochemical Parasitology 1993. 60: 133-144.
37. Kwa MSG, Veenstra JG, Roos MH. Benzimidazoleresistance in Haemonchus contortus is correlated witha conserved mutation at amino acid 200 in b-tubulinisotype 1. Mol and Biochem parasitol 1994. 63: 299-303.
38. Hennessy DR. The disposition of antiparasiticdrugs in relation to the development of resistance byparasites of livestock Acta Tropica 1994. 56:125-141.
39. Van Wyk JA, Malan FS, Bath GF. Rampant anthel-mintic resistance in sheep in South Africa - What arethe options?. Workshop held at the 16th Internationalconference of de World Association for the advance-ment of Veterinary Parasitology Sun City 10-15 au-gust. van Wyk. Schalkwyc ed. 1996: 51-63.
40. Nari AH. Enfoque epidemiológico sobre el diagnós-tico y control de resistencia a antihelmínticos en ovi-nos. Ed. Hemisferio Sur Montevideo Uruguay 1987 59pág.
41. Waller P, Echevarría F, Eddi C, Maciel S, Nari A,Hansen JW. The prevalence of anthelmintic resistancein nematode parasites of sheep in Southern LatinAmerica: General overview. Vet Parasitol. 1996; 62:181-187.
42. Hennessy DR. The impact of pharmacokinetic prop-erties on development of resistance to antiparasiticcompounds. 17th International conference of the WorldAssociation for the advancement of Veterinary Parasi-tology . Copenhagen 15-19 august 1999
43. Dash K, Waller P. Denchplan research-based wormcontrol. Rural Research 1987. 135: 4-9.
44. Waller PJ. Resistance in nematode parasites oflivestock to the benzimidazole antihelmintics. Parasi-tology today 1990; 6 (4): 127-129.
45. Nari A, Franchi M, Rizzo E, Marmol E, Mautone G.Evaluación de un programa de control de nematodesgastrointestinales en ovinos. Medidas para dilatar laaparición de resistencia antihelmíntica. XXV Jorna-das de buiatría, Junio 18-22 1977. Paysandú Uruguay.
46. Quintana S. Manejo parasitario del cordero de des-tete en campo natural y pastoreo alterno con bovinosen un área de basalto superficial. Veterinaria 1987. 23(97): 6-14.
El Sitio Argentino de Producción Animal
17 de 17





























