Embed Size (px)
Citation preview

Enhancement of Object Representations in Primate Perirhinal Cortex During aVisual Working-Memory Task
Sidney R. Lehky1 and Keiji Tanaka1,2
1Cognitive Brain Mapping Laboratory, RIKEN Brain Science Institute, Saitama; and 2Graduate School of Science and Engineering,Saitama University, Saitama, Japan
Submitted 16 February 2006; accepted in final form 6 November 2006
Lehky SR, Tanaka K. Enhancement of object representations inprimate perirhinal cortex during a visual working-memory task. JNeurophysiol 97: 1298–1310, 2007. First published November 15,2006; doi:10.1152/jn.00167.2006. We compared single-cell activitiesin perirhinal cortex (PRh) as well as adjacent visual cortex (area TE)across two tasks. One task required the monkey to identify anystimulus repetition within a sequence of object stimuli. In the othertask, the same stimuli were presented, but the monkey didn’t have toremember them. PRh responses during the object-memory task wereelevated relative to those during the second task. In TE, on the otherhand, there were no significant task-related differences in responses.We did not observe task-related differences related to repetitioneffects in either brain area. The onset of the enhanced signal in PRhduring the object-memory task occurred with a latency of 80 ms afterthe onset of the stimulus response, suggesting that it was the result oftop-down feedback.
I N T R O D U C T I O N
The inferior temporal cortex (IT) plays a central role invisual object processing. Neurons in dorsal and ventral regionsof anterior IT (TEad and TEav), the last unimodal visual areas,are selective for complex object features (Desimone 1984;Rolls and Tovee 1995; Tamura and Tanaka 2001; Tanaka1996), thus providing the capacity to perceptually identify anddiscriminate stimuli. Perirhinal cortex (PRh) is believed tohave an involvement in the mnemonic aspect of recognition forcomplex objects in addition to perhaps contributing further toperceptual processing. It is a polymodal structure mediallyadjacent to TEav on the ventral surface of IT. TE providesmajor anatomical inputs to PRh and in return receives strongfeedback projections from PRh (Lavenex et al. 2002; Saleemand Tanaka 1996; Suzuki and Amaral 1994; Van Hoesen1982).
PRh in particular appears to play a multifaceted role inobject processing. In the context of long-term visual memory,recognition of complex objects seems to preferentially involvethe PRh over neighboring areas. Lesions to PRh producegreater behavioral deficits for object recognition than lesions tonearby limbic structures, during delayed match to sample(DMS) or similar tasks (Alvarado and Bachevalier 2005; Bax-ter and Murray 2001; Meunier et al. 1993; Murray and Mishkin1998; Zola-Morgan et al. 1989). Lesions to PRh also producegreater deficits to recognition memory than lesions to theadjacent visual cortex, area TE (Buckley et al. 1997; Buffalo etal. 1999). Moreover, disrupting cholinergic input to PRh but
not TE or dentate gyrus impairs recognition memory (Tang etal. 1997; Turchi et al. 2005). As the delay tasks in these studiesused trial-unique stimuli and delays stretching into severalminutes, they were most likely primarily probing long-termmemory.
Long-term memory has also been examined at the single-celllevel in temporal cortex. Miyashita and colleagues studiedassociative memory between visual stimuli (Miyashita 1993;Naya et al. 2001; Takeda et al. 2005), finding a critical role forPRh in the formation of associations. Stimulus-reward asso-ciative memory was studied by Liu and Richmond (2000) andMogami and Tanaka (2006), who both found a larger involve-ment of PRh over TE. There is also an extensive literature onfamiliarity and recency effects on single-cell activity in TE andPRh (reviewed by Brown and Aggleton 2001), which to a largeextent also involve long-term memory.
In addition to long-term memory, working memory withintemporal cortex has been the subject of monkey studies (Baylisand Rolls 1987; Davachi and Goldman-Rakic 2001; Eskandaret al. 1992; Fuster 1990; Fuster and Jervey 1981; Holscher andRolls 2002; Miller and Desimone 1994; Miller et al. 1991;Miyashita and Chang 1988; Nakamura and Kubota 1995;Sobotka et al. 2005). Working memory is believed to beclosely related to attention (Awh and Jonides 2001; Awh et al.2006; Barnes et al. 2001; de Fockert et al. 2001; Desimone1996). In this study, we shall be examining single-cell activityin both TE and PRh associated with the performance of aworking-memory task.
Working-memory effects within temporal cortex are likelyto be closely tied to the perceptual properties of variousstructures within that region. Although the perceptual role ofarea TE is clear, that of PRh is less so. However, there ismounting evidence that in addition to mnemonic functions,PRh may play a role in perceptual processing. This evidenceindicates that PRh is necessary when object identificationrequires the conjunction of a more complex set of visualfeatures than TE neurons are capable of handling (Buckley andGaffan 1997; Buckley et al. 2001; Bussey et al. 2002, 2003,2005; Eacott et al. 1994; Lee et al. 2005; Murray and Bussey1999; Tyler et al. 2004; see also Gaffan 2002). This perspectivetreats PRh, in terms of its visual perceptual properties, as thenext link beyond TE in a chain of cortical areas stretching backto V1 with increasingly complex receptive fields. However,such a perceptual role for PRh remains controversial, andothers have suggested that the function of PRh can be under-
Present address and address for reprint requests and other correspondence:S. R. Lehky, Computational Neuroscience Lab, The Salk Institute, 10010 N.Torrey Pines Road, La Jolla, CA 92037 (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
J Neurophysiol 97: 1298–1310, 2007.First published November 15, 2006; doi:10.1152/jn.00167.2006.
1298 0022-3077/07 $8.00 Copyright © 2007 The American Physiological Society www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

stood purely in mnemonic terms (Hampton 2005; Hampton andMurray 2002; Levy et al. 2005; Squire et al. 2004).
The primary structure associated with working memory islateral prefrontal cortex. This association is based on lesionstudies extending as far back as Jacobsen (1936) and Mishkin(1957) as well as observations of delay activity in prefrontalneurons during delayed-match-to-sample or delayed-responsetasks (Fuster 1973; Fuster and Alexander 1971; Kubota andNiki 1971; Niki 1974). The portrayal of prefrontal cortex as aworking-memory buffer received an influential formulation byGoldman-Rakic (1987).
Although there is widespread agreement that prefrontal de-lay activity plays an important role in working memory, thereis disagreement about what that role is. On one hand, there isthe view that prefrontal delay activity actually represents thecontents of working memory, including specific stimulus fea-tures (reviewed by Courtney 2004). By this view, representa-tions in visual working memory do not engage the sameposterior neural structures that are involved in perceptualrepresentations of stimuli. Alternatively, there is the view thatthe contents of visual working memory are indeed representedin posterior sensory cortices. By this view, prefrontal delayactivity serves as a control signal engaged in the attentionalactivation, manipulation, or monitoring of the posteriorly heldcontents of working memory (Curtis and D’Esposito 2003;Lebedev et al. 2004; Petrides 1994, 2000; Postle 2006; Postleet al. 1999).
In the particular case of TE and PRh, the possibility offeedback interactions with prefrontal cortex of the sort outlinedin the preceding text is supported by the existence of extensiveanatomical connections running both ways (Lavenex et al.2002; Rempel-Clower and Barbas 2000; Suzuki and Amaral1994; Ungerleider et al. 1989; Webster et al. 1994). If prefron-tal cortex was indeed projecting a control or activating signalto posterior representations during a working-memory task, asthe later hypothesis in the preceding text indicates, then PRhand TE, given their physiological properties and anatomicalconnections, would be likely places to look for such a signal inthe context of remembering complex objects.
Delay activity is considered the primary neural substrate ofworking memory in prefrontal cortex. The fact that prefrontaldelay activity survives disruption by intervening visual stimulibetween sample and match in a DMS task (Miller et al. 1996)reinforces that belief. However, delay activity does not surviveintervening stimuli in PRh, making it unlikely that it is thesubstrate for working memory in that structure (Miller et al.1993). Subsequently it was suggested that a repetition enhance-ment effect for the match stimulus in a DMS task representedthe neural substrate of working memory in PRh cortex (Millerand Desimone 1994).
In this study, we have taken a different approach to identi-fying activity in temporal cortex associated with workingmemory. Rather than comparing neural responses during dif-ferent epochs within a single working-memory task, we madea paired comparison across two tasks. In one task, the monkeywas required to keep a set of serially presented object stimuliin working memory. In the second task, the monkey againviewed the same stimuli but without the memory requirementwhile being kept busy encoding into memory a different set ofunrelated stimuli. Instead of using a delayed-match-to-sampletask, which requires keeping just one item in working memory
(the sample), we used a serial-recognition task that requiredkeeping multiple stimulus images in working memory simul-taneously. If prefrontal cortex was providing a control signal torepresentations in temporal cortex, then it might be morevisible if prefrontal cortex was keeping track of multiple itemsrather than just one.
Although we used a working-memory task, there is evidencefor extensive overlap between mechanisms of working mem-ory and attention, as will be detailed in the DISCUSSION, and weshall consider the data within the context of both those factors.
M E T H O D S
Two female Japanese macaque monkeys (Macaca fuscata) wereused, weighing 7 and 9 kg. All experimental procedures conformed toNational Institutes of Health guidelines on the care and use oflaboratory animals. Recordings were conducted on the right side ofthe brain in both cases, ranging over anterior-posterior positionsA17–A19 in one monkey and A14–A17 in the other (Fig. 1). Cham-bers were angled at 10° so that electrodes slanted laterally as theyadvanced.
Recordings
There were three target structures, TEad, TEav, and PRh, eachadjacent to the next as one moves down ventrally along the infero-temporal cortex. TEad is in the middle temporal gyrus, extendingventrally into the lateral lip of the anterior medial temporal sulcus(amts). TEav is located around the medial bank and medial lip ofamts. PRh, or area 35/36, is located on the lateral bank and lateral lipof the rhinal sulcus (rs) (Saleem and Tanaka 1996; Suzuki and Amaral1994).
Because these structures are located so close to each other in themediolateral direction, electrode positions had to be determined withprecision. To do this, stereotactic coordinates of the target structureswere first obtained from MRI scans. Then, after each recordingsession, an X-ray was taken with the electrode still in place within thebrain, using a portable veterinary X-ray machine. From the X-rayimages, the lateral distance between the electrode guide tube tip andthe brain midline was measured. This in turn allowed determination ofthe mediolateral position of the tip of the slanting electrode. Slightrotations of the head relative to the fronto-parallel plane of the X-raymachine were mathematically corrected using information from small
sts
amts
rs
hc
Monkey 1 Monkey 2
1 cm
1 cm
A18 A16
amtsrs
stshc
A C
B D
FIG. 1. Recording sites, indicated in gray. Gray areas on the lateral viewsof the brains indicate anterior-posterior ranges of recording. rs, the rhinalsulcus; amts, the anterior middle temporal sulcus; sts, the superior temporalsulcus.
1299ENHANCED PERIRHINAL ACTIVITY DURING A WORKING-MEMORY TASK
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

(1 mm) titanium markers implanted in the skull at known stereotacticcoordinates, which were visible on the X-ray images.
At the end of the experiments, the animals were killed, andrecording sites were verified histologically through examination ofelectrode tracks. In a few cases, tracks fell too far medially in therhinal sulcus, and the corresponding cells were excluded from thedata.
In an attempt to reduce selection bias, all isolated cells withdetectable activity, whether spontaneous or stimulated, were acceptedfor complete testing without informal prescreening of cells for respon-siveness.
Behavioral task and stimuli
Stimuli were a set of 110 color photographs of real-world objects,both natural and man-made (examples shown in Fig. 2). They werepresented against a neutral gray background that matched the generalbackground of the video display, set at 40 cd/m2, so that the stimulusboundary box was not visible. The boundary box subtended a visualangle of 6° (full width). The video display had a resolution of 1,024 �768 pixels and a frame rate of 75 Hz and was viewed from a distanceof 48 cm. Eye position was monitored with an optical eye tracker.
The monkeys were repeatedly exposed to the same set of 110 objectstimuli during several months of training, so all the stimuli werehighly familiar to them by the time recordings were started.
The general testing procedure for each cell was as follows. First, apreliminary test using the entire 110 object stimulus set was per-formed. Based on that, 10 stimuli were chosen for the main phase ofthe testing, in which two tasks, an object-memory task and a dot-memory distractor task, were interleaved. A serial-recognition taskwas chosen rather than a serial delayed-match-to-sample (sDMS) taskin an attempt to increase the response difference from the distractortask (see following text). A serial-recognition task requires keepingmultiple stimuli in memory simultaneously and not just a singlesample stimulus as in sDMS. Thus it provides a heavier memory loadthan a sDMS task.
The object-memory task and the dot-memory distractor task, as willbe described in the following text, were both essentially the sameworking-memory task operating on different stimulus sets. The strat-
egy was to select visual patterns for each task that stimulated differentpopulations of neurons, taking advantage of the narrow pattern selec-tivity of cells in inferotemporal cortex. As the monkey switched backand forth between remembering one set of stimuli or the other,memory-related activity would be expected to switch back and forthbetween the two neural populations. Because we would be recordingfrom just one of those two populations (those neurons stimulated bythe images for the “object-memory” task), the memory-related signalwould appear to switch on when the monkey had to remember thestimuli for that task and off when the memory related signal wasattached to a different population of neurons forming representationsfor the other class of stimuli. A “memory-related signal” in thiscontext could include an attentional aspect to it, as directing attentionto a stimulus may be a condition for the stimulus to enter workingmemory.
During the preliminary testing phase, the monkey was performingthe object-memory task. Each of the 110 objects was presented aminimum of two times (mean: 2.5). After that, stimuli were sorted inorder of the firing rate elicited (effectiveness), and the top four wereselected, as well as six additional stimuli evenly spaced over theremaining range of effectiveness.
In the object-memory task, which was a variant of a serial recog-nition task, the monkey viewed a series of object stimuli (Fig. 3A).The task was to report any repetition in the series by releasing a touchsensitive bar within 800 ms of repetition onset. The trial terminatedafter the first repetition occurred. The stimulus series length randomlyvaried from two to five images. Each object stimulus was presentedfor 600 ms, with a 700-ms inter-stimulus interval between them. Asquare fixation spot (0.3o) was presented at the start of each trial andduring interstimulus intervals within the series with no temporal gapsbetween the fixation spot and adjacent object stimuli.
In the second task, the dot-memory distractor task, the monkeyviewed the same serially presented object stimuli but was not requiredto remember them. To discourage the monkey from covertly remem-bering the object stimuli, the dot-memory task was performed as adistractor during each trial (Fig. 3B). During this task, a round dot(0.5° diam), the color of which was randomly selected from sixpossibilities, was displayed, starting before the first object stimulusand then afterward during each interstimulus interval between objectstimuli. The monkey’s task was to respond to any repetition in thecolor of the dot within 800 ms of stimulus onset. The object stimulihad no significance for correct performance of the task now.
As was the case for the object-memory task, two to five objectstimuli were presented during each trial but now with the colored dotdistractor displayed in the period preceding each object stimulus. Thelocation of the colored dot was randomly placed anywhere within 2°of fixation during each presentation within a trial, in an attempt todiscourage the monkey from maintaining a narrow focus of spatialattention.
For the distractor task, object stimulus presentations remained 600ms, but the interstimulus intervals were increased to 900 ms. Thecolor-dot distractor stimulus was displayed during the central 500 msof the interstimulus intervals, so that there was a 200-ms gap beforeand after the distractor dot separating it from the object stimuli. Asquare fixation spot was displayed simultaneously with the distractordot in exactly the same manner that the fixation spot had beendisplayed during the periods between object stimuli in the object-memory task (Fig. 3).
During the distractor task, 50% of the trials presented each objectstimulus just once within the trial, whereas the other 50% included arepeat presentation of one of the objects. For this task, however, arepeat presentation of an object stimulus had no significance to themonkey.
The two tasks were presented in interleaved blocks of trials, twoblocks for each task for a total of four blocks, while the activity ofeach neuron was recorded. Within each block, each object stimuluswas presented a minimum of 10 times (mean: 12). Trials were also
FIG. 2. Example stimulus images. These are a subset of the 110 imageseach cell was initially tested on. Based on responses during this preliminaryscreening, 10 images were selected for further testing (a different set for eachcell). Those 10 images were not the 10 most effective stimuli but included afull range of effectiveness from the best to the worst.
1300 S. R. LEHKY AND K. TANAKA
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

included which contained only the colored dots of the distractor task,with the object stimulus periods left blank, to test sensitivity of cellsto the color stimuli. The colored dots in the distractor task werepresented with the fixation spot on, using the same gray backgroundas that used for object stimuli.
For both tasks, the task-relevant stimulus repetition occurred withequal likelihood at the four possible time positions in the series(positions 2–5). We calculated chance performance conservatively byassuming that the monkeys had learned the basic structure of the task,so that they knew this probability of stimulus repetition at eachposition. In that case, if they ignore the stimuli and decide at the startof each trial to respond randomly at one of those four positions, theywould have 25% chance of correct performance.
Although the distractor task included an additional color dot in theperiods between object stimuli (Fig. 3), we believe it unlikely that thiswould result in a response difference between the two tasks due totemporal crowding or masking effects. A fixation spot, similar in sizeand shape to distractor color dot, preceded the object stimuli underboth tasks, with an ISI of 0 ms. Presenting an additional color dotwould likely have added little if any masking over that alreadyproduced by the fixation spot, particularly since the ISI between thecolor dot and object stimuli was 200 ms, and neural masking effectsterminate with ISIs of 100–150 ms (Kondo and Komatsu 2000;Kovacs et al. 1995; Macknik and Livingstone 1998; Rolls and Tovee1994; Rolls et al. 1999). Furthermore, in our data there was noperturbation to the early part of the object-stimulus response (Fig.7C), whereas suppression of the early response was a significantfeature in the physiological masking studies of Macknik and Living-stone (1998) as well as Kondo and Komatsu (2000). For these reasons,we believe the effects reported here reflect cognitive processing(attention, memory) rather than sensory responses to the stimuli,although the possibility of a sensory contribution cannot be com-pletely excluded.
Besides masking, another possible effect during target detectionwithin serial presentation of stimuli is attentional blink (Raymond etal. 1992). However, our tasks proceeded at a very slow pace relativeto the rapid serial visual presentation (RSVP) procedure used duringattentional blink experiments, with a 1,200-ms SOA (stimulus onsetasynchrony) between object stimuli in our object-memory task and
700-ms SOA between the dot stimuli and object stimuli in ourdot-memory task. Therefore we don’t think that limitations in thetemporal dynamics of deploying attention were a significant factor inour experiment.
Data analysis
Response was defined as average firing rate over the stimulusduration shifted by latency. Latency was determined as time tohalf-height of the first peak in the grand average PSTH, incorporatingdata from all cells within a given cortical area to all stimuli. Onsettime for the stimulus presentation was corrected for the time it tookthe video electron beam to reach the center of the screen, �8 ms.Estimates of response standard errors were made using Bayesianmethods (Lehky 2004), and the SEs were used to determine thosestimuli producing significant responses. Spontaneous activity wasdetermined from a 150-ms window preceding each object stimuluspresentation. Cells that showed �2 spike/s change in responses to thecolored dot distractor stimuli in trials of the distractor task with noobject stimulus presentation were excluded from the analysis regard-less of the significance of that response. This was done to reduce thepossibility that object-stimuli responses would be contaminated byafterdischarges from responses to the dot stimuli. In some cases, theactivity in the excluded cells may have reflected above-averagevariability in the background firing rather than responses to the dotstimuli per se.
For grand average comparisons of neural activities between the twotasks, the last stimulus presentation within each trial, which over-lapped the monkeys’ behavioral responses, were omitted from theanalysis. For comparisons between repeat presentations of a stimuluswithin single trials, all stimulus presentations were considered in theanalysis, but with inclusion of only the first 300 ms of the PSTH ineach case, so that there was no overlap with behavioral responses.
Visual selectivity can be measured in terms of the shape of theprobability density function (pdf) of a neuron’s responses over thestimulus set (Lehky et al. 2005; Simoncelli and Olshausen 2001).Here we define selectivity SE as the change in entropy of a neuralresponse pdf relative to the entropy of a Gaussian distribution, whichhas maximum entropy for a fixed variance
SE � HG � H�r� (1)
where HG is the entropy of a Gaussian, and H(r) is the entropy of acell’s response distribution taken from the data (Lehky et al. 2005).Equating high selectivity with low entropy captures the characteristicof a highly selective cell of having little response to most stimuli anda large response to a few.
The entropy of the Gaussian reference distribution is given byRieke et al. (1997)
HG �1
2log2�2�e�2� � 2.074 bits (2)
with variance normalized to one. Entropy of the probability distribu-tion of the data are
H�r� � � �p�r�log2�p�r��dr (3)
where r is the response and p(r) is the response probability densityfunction (pdf). For practical calculations Eq. 3 is discretized to
H�r� � � �j�1
M
p�rj�log2�p�rj���r (4)
where responses from n stimulus images have been placed in M bins.Expressing the selectivity index SE (Eq. 1) in terms of Eqs. 2 and 4,we get
releasebar
releasebar
A Object-memory task
B Dot-memory distractor task
Fixation spot onObject-stimulus onDistractor-stimulus on
Time
FIG. 3. A: object-memory task: monkey responded to any repetition withina series (2–5) of object stimuli. The duration of object stimuli was 600 ms andthe interstimulus interval was 700 ms. B: dot-memory distractor task: monkeyviewed a series of object stimuli, but was not required to remember them. Todiscourage the monkey from covertly retaining object stimuli in memory, adistractor task was included. The distractor involved displaying a small dotwith a randomly selected color during the inter-stimulus intervals betweenimage presentations. Each dot presentation was at a random location todiscourage the monkey from maintaining a narrow focus of attention. Themonkey responded to any repetition in dot color. Duration of object stimuliwas 600 ms, duration of distractor dot stimuli was 500 ms, and the intervalbetween object stimuli was 900 ms.
1301ENHANCED PERIRHINAL ACTIVITY DURING A WORKING-MEMORY TASK
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

SE � 2.074 � �j1
M
p�rj�log2�p�rj���r (5)
The pdf determined from the data must be normalized to a variance of1.0 before performing this calculation. Bin size �r was set by dividingthe data range in M bins equal to the square root of the number ofimages in the stimulus set.
To examine repetition effects, we calculated, for each trial, the ratioof the response during the second presentation of a stimulus imagerelative to the first presentation. The array of response ratios was thenlog transformed to normalize the distribution, and a t-test was con-ducted on the data for each cell, examining if log(ratio) was signifi-cantly different from zero. For plots of the probability distribution ofresponse ratios, the average ratio for each cell was calculated as thegeometric mean over all trials, and then response ratios were incor-porated into the probability plots on a cell-by-cell basis.
R E S U L T S
Data were analyzed from 65 cells in TEad, 181 cells inTEav, and 115 cells in PRh, collected while the macaquemonkey performed two tasks, an object-memory task (Fig. 3A)and a dot-memory distractor task (Fig. 3B). Histology showedthat the TEad cells came from the middle temporal gyrus nearthe lateral lip of the anterior medial temporal sulcus (amts),TEav cells from the medial bank and medial lip of amts, andPRh cells from the lateral bank and lateral lip of the rhinalsulcus (rs; Fig. 1).
Each cell underwent preliminary testing with a large set ofobject images (110 images) to select effective stimuli for themain phase of the experiment. ANOVA analysis of this dataindicated visual selectivity in 100% of TEad cells, 97% ofTEav cells, and 98% of PRh cells (P 0.01).
The probability density function of the response magnitudesfor each cell to all 110 stimuli was determined using Gaussiankernel smoothing methods. Figure 4 shows the median pdf ineach cortical area. They were determined by finding the me-dian probability density value at each firing rate, over all cells.The PRh cells differ from those in TE primarily by having
more symmetrical pdf”s with less of a tail of large responsesextending to the right.
Visual selectivity to complex stimuli (natural images, ob-jects) can be quantified by measures of the overall shape of theresponse probability density function (Lehky et al. 2005). Herewe define selectivity SE as the change in entropy of a responsepdf relative to a Gaussian pdf. The Gaussian is referenced asthe density function for a minimally selective neuron (seeMETHODS). Applying this selectivity measure to the pdf of eachcell and taking median values, we find SE � 0.33 for TEad,SE � 0.32 for TEav, SE � 0.22 for PRh, on a scale whereSE � 0 for the low-selectivity Gaussian distribution.
After preliminary testing of each cell with 110 object stim-uli, 10 objects were selected for use in the main phase of theexperiment comparing responses during the object-memorytask (Fig. 3A) and dot-memory distractor task (Fig. 3B).
Overall correct performance for the object-memory task was84% and for the dot-memory distractor task it was 74%. Thiswas calculated on the basis of trials that were not abortedbecause of broken fixation. If one includes aborted trials,correct performance for the object-memory task was 74% andfor the dot-memory task 68%. Both monkeys showed poorerperformance on the distractor task.
Performance on distractor trials that included object stimulusrepetition (78% correct) was close to performance during thosetrials that did not include repetition (85% correct). This indicatesthat the monkeys were indeed selectively responding to the colordot stimuli during the distractor task and not indiscriminately toany stimulus repetition that appeared on the screen.
Behavioral statistics are presented in more detail in Fig. 5.Performance declined as the length of the stimulus seriesincreased (Fig. 5A). On the other hand, performance as afunction of the separation (number of items) between the firstpresentation (cue) and second presentation (match) of thetarget stimulus formed a U-shaped curve (Fig. 5B). Althoughthe distractor curves are shifted down slightly, the shapes of thecurves for both tasks are similar, and there is no indication herethat the monkeys were using different strategies for performingthese two serial recognition tasks. Statistics for trials abortedbecause the monkeys broke fixation are also similar for the twotasks with the abort rate slightly lower for the dot-memory task(Fig. 5C). Yakovlev et al. (2004) provide a detailed consider-ation of behavioral statistics for monkeys performing essen-tially the same serial recognition task we used.
The U-shaped curves in Fig. 5B are examples of well-known serial position effects that occur when a list ofsequentially presented items is held in short-term memory.The upturn of the curve at the left is an indication of therecency effect, as it occurs when the cue immediatelyprecedes the match stimulus. The upturn at the right is anexample of the primacy effect, as it occurs when the cuestimulus is the first item presented in the stimulus series.Primacy and recency memory effects occur in both humansubjects (Atkinson and Shiffrin 1968, 1971; Glanzer andCunitz 1966; Murdock 1962) and monkeys (Castro andLarsen 1992; Sands and Wright 1980; Wright et al. 1985).
The poorer correct identification of the match stimulus onthe dot-memory distractor task may be related to lower sa-liency of the stimuli for that task. The stimuli in the dot-memory task were small (0.5°), simple geometric shapes thatdiffered along only one dimension (color). On the other hand,
0 10 20 300
0.02
0.04
0.06
0.08
0.10
spikes/sec
prob
abili
ty d
ensi
ty
TEadTEavPRh
FIG. 4. Probability density functions (pdf’s) of neural responses to objectstimuli. Responses of each cell to 110 stimuli were used to calculate that cell’spdf, and then the median pdf was determined for the different cortical areas.These curves show that PRh neurons have much lower probabilities ofproducing large responses than TE cells do. Peaks of the distributions approx-imately coincide with spontaneous activity. The number of units underlyingthese curves are TEad: 65, TEav: 181, and PRh: 115.
1302 S. R. LEHKY AND K. TANAKA
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

the object-memory patterns were large (up to 6°), with eachcomprising a complex multidimensional mixture of shape,texture, and color. The roughly equal propensity for the mon-keys to abort trials by breaking fixation under the dot- andobject-memory tasks suggests that the overall attentional ormotivational state of the monkeys did not differ during the twotasks. Those factors are therefore not likely to account fordifferences in correct performance or the task-related differ-ences in neural activities we observed.
We checked sensitivities of cells to the colored dot distrac-tors presented during the distractor task. The concern was thatresponses to distractors might overlap responses to objectstimuli. Trials were included which presented the colored dotsin the normal task setting (Fig. 3B) but with the “objectstimuli” periods held blank. Figure 6C shows the averagePSTH in PRh to the colored dots, indicating low sensitivity ofthis population to the distractor stimuli. TEad and TEav pro-duced similar results (Fig. 6, A and B).
Despite population insensitivity to distractors, cells wereexamined individually and any cell whose average responseshowed a deviation from baseline activity of �2 spike/s (eitherpositive or negative) was excluded from further analysis re-gardless if that activity was significant or not. That left 86%(56/65) of the cells in TEad and 76% (137/181) of TEav cellsremained, whereas 94% (108/115) of PRh cells remained.From the pool of remaining cells, we included in the analysisonly those stimuli that produced activity elevation in theobject-memory task that was both significant (P 0.05) andlarge (�2 spike/s above spontaneous). For some cells none ofthe 10 object stimuli met those criteria, so that after this second
stimulus series length
0
20
40
60
80
100
perc
ent c
orre
ct
2 3 4 5
A obj-memory distractorchance
2 3 4 50
10
20
30
40
50
stimulus series length
perc
ent b
roke
fixa
tion
C
1 2 30
20
40
60
80
100
perc
ent c
orre
ct
match position difference
B
FIG. 5. Behavioral statistics for the 2 tasks. A: percentage correct as afunction of the length of the stimulus time series, which varied randomlybetween 2 and 5 in different trials. Percentages were calculated excludingaborted trials. Chance performance is set at 25% on the assumption that foreach trial the monkey picks at random one of the four possible time positionsof the target stimulus. B: percentage correct as a function of the difference inposition along the stimulus time series between the 1st presentation of thetarget stimulus and the 2nd presentation (match). C: percentage of trials thatwere aborted because the monkey broke fixation as a function of the length ofthe stimulus time series.
0 200 400 6000
8
16
24
spik
es/s
ec
n=65 TEadA
0 200 400 6000
8
16
24
spik
es/s
ec
n=181 TEavB
0 200 400 6000
8
16
24
msec
spik
es/s
ec
n=115 PRhC
FIG. 6. Population averages of response spike trains (PSTHs) of cells inTEad, TEav, and PRh to the colored dot displayed during the distractor task,pooled over all cells responses. The plots have been smoothed using aGaussian kernel with � � 5 ms.
1303ENHANCED PERIRHINAL ACTIVITY DURING A WORKING-MEMORY TASK
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from
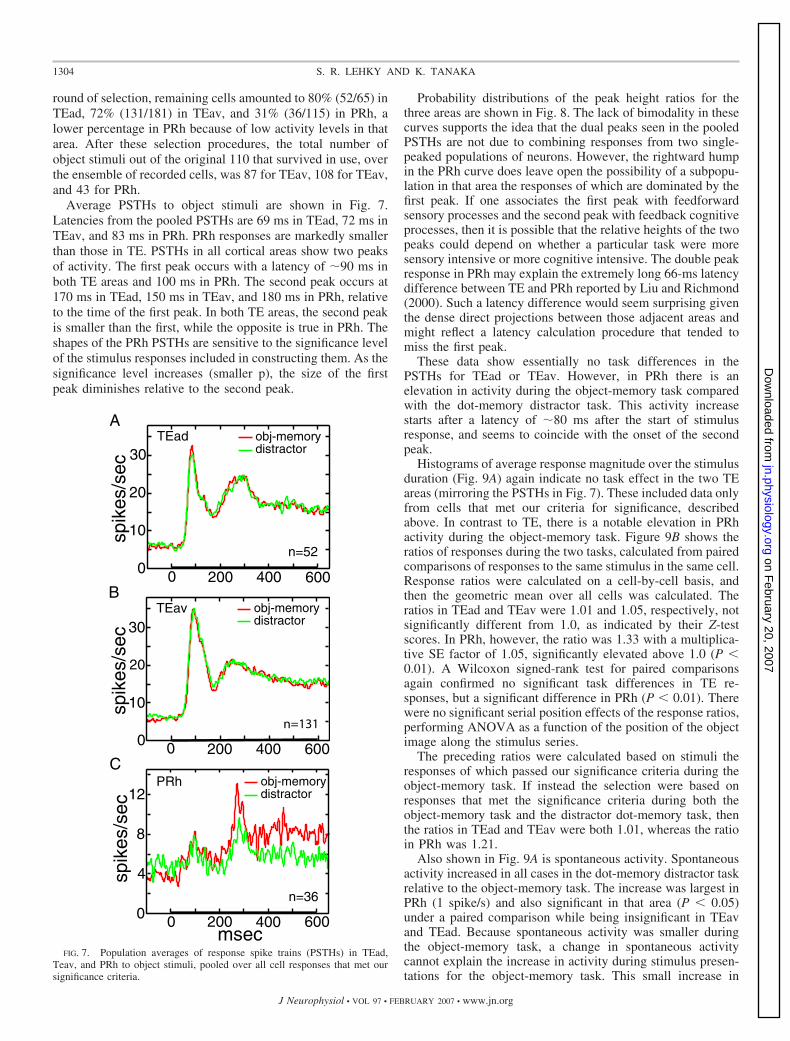
round of selection, remaining cells amounted to 80% (52/65) inTEad, 72% (131/181) in TEav, and 31% (36/115) in PRh, alower percentage in PRh because of low activity levels in thatarea. After these selection procedures, the total number ofobject stimuli out of the original 110 that survived in use, overthe ensemble of recorded cells, was 87 for TEav, 108 for TEav,and 43 for PRh.
Average PSTHs to object stimuli are shown in Fig. 7.Latencies from the pooled PSTHs are 69 ms in TEad, 72 ms inTEav, and 83 ms in PRh. PRh responses are markedly smallerthan those in TE. PSTHs in all cortical areas show two peaksof activity. The first peak occurs with a latency of �90 ms inboth TE areas and 100 ms in PRh. The second peak occurs at170 ms in TEad, 150 ms in TEav, and 180 ms in PRh, relativeto the time of the first peak. In both TE areas, the second peakis smaller than the first, while the opposite is true in PRh. Theshapes of the PRh PSTHs are sensitive to the significance levelof the stimulus responses included in constructing them. As thesignificance level increases (smaller p), the size of the firstpeak diminishes relative to the second peak.
Probability distributions of the peak height ratios for thethree areas are shown in Fig. 8. The lack of bimodality in thesecurves supports the idea that the dual peaks seen in the pooledPSTHs are not due to combining responses from two single-peaked populations of neurons. However, the rightward humpin the PRh curve does leave open the possibility of a subpopu-lation in that area the responses of which are dominated by thefirst peak. If one associates the first peak with feedforwardsensory processes and the second peak with feedback cognitiveprocesses, then it is possible that the relative heights of the twopeaks could depend on whether a particular task were moresensory intensive or more cognitive intensive. The double peakresponse in PRh may explain the extremely long 66-ms latencydifference between TE and PRh reported by Liu and Richmond(2000). Such a latency difference would seem surprising giventhe dense direct projections between those adjacent areas andmight reflect a latency calculation procedure that tended tomiss the first peak.
These data show essentially no task differences in thePSTHs for TEad or TEav. However, in PRh there is anelevation in activity during the object-memory task comparedwith the dot-memory distractor task. This activity increasestarts after a latency of �80 ms after the start of stimulusresponse, and seems to coincide with the onset of the secondpeak.
Histograms of average response magnitude over the stimulusduration (Fig. 9A) again indicate no task effect in the two TEareas (mirroring the PSTHs in Fig. 7). These included data onlyfrom cells that met our criteria for significance, describedabove. In contrast to TE, there is a notable elevation in PRhactivity during the object-memory task. Figure 9B shows theratios of responses during the two tasks, calculated from pairedcomparisons of responses to the same stimulus in the same cell.Response ratios were calculated on a cell-by-cell basis, andthen the geometric mean over all cells was calculated. Theratios in TEad and TEav were 1.01 and 1.05, respectively, notsignificantly different from 1.0, as indicated by their Z-testscores. In PRh, however, the ratio was 1.33 with a multiplica-tive SE factor of 1.05, significantly elevated above 1.0 (P 0.01). A Wilcoxon signed-rank test for paired comparisonsagain confirmed no significant task differences in TE re-sponses, but a significant difference in PRh (P 0.01). Therewere no significant serial position effects of the response ratios,performing ANOVA as a function of the position of the objectimage along the stimulus series.
The preceding ratios were calculated based on stimuli theresponses of which passed our significance criteria during theobject-memory task. If instead the selection were based onresponses that met the significance criteria during both theobject-memory task and the distractor dot-memory task, thenthe ratios in TEad and TEav were both 1.01, whereas the ratioin PRh was 1.21.
Also shown in Fig. 9A is spontaneous activity. Spontaneousactivity increased in all cases in the dot-memory distractor taskrelative to the object-memory task. The increase was largest inPRh (1 spike/s) and also significant in that area (P 0.05)under a paired comparison while being insignificant in TEavand TEad. Because spontaneous activity was smaller duringthe object-memory task, a change in spontaneous activitycannot explain the increase in activity during stimulus presen-tations for the object-memory task. This small increase in
B
C
A
FIG. 7. Population averages of response spike trains (PSTHs) in TEad,Teav, and PRh to object stimuli, pooled over all cell responses that met oursignificance criteria.
1304 S. R. LEHKY AND K. TANAKA
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

spontaneous activity during the dot-memory task may be acognitive effect, perhaps reflecting increased resources (mne-monic/attentional) directed at cells during the periods betweenobject stimuli presentations. Alternatively, it could be a sen-sory effect reflecting responses to the distractor dot stimuli,although this seems improbable as there is no evidence for suchsensory responses in Fig. 6.
We examined scatterplots that compared firing rates ofindividual cells during the two tasks (Fig. 10). This showsagain that responses in TEav during the two tasks were almostthe same, as was the case in TEad, but responses in PRh wereelevated during the object-memory task. The linear regressionslope in TEav was 0.98 � 0.01, in TEad, 0.97 � 0.01, and inPRh, 1.25 � 0.04. Correlation coefficients between responsesfor the two tasks were 0.99 in both TEav and TEad, and 0.94in PRh. Cells whose response ratios were significantly differentfrom 1.0 at the P � 0,01 level under a Z-test are shown in greenin the figure. The proportion of such significant cells was 6%(2/52) in TEav, 4% (5/131) in TEav, and 44% (16/36) in PRh.
The 10 object stimuli used during the task comparison wereselected for each cell, from a preliminary set of 110, to span arange of stimulus effectiveness from high to low. To determineif stimulus selectivities are affected by the task, we examinedresponse magnitudes over the ordered set of the 10 stimuli(Fig. 11). These curves were constructed by ranking the stimulifor each cell by response magnitude in the object-memory taskand then averaging over all cells. As before, there was no task
effect in either TEad or TEav. In PRh, on the other hand, thecurve for the object-memory task was shifted upward parallelto that of the distractor task. The parallel nature of the shiftindicates that stimulus selectivity has not been changed by thetask.
Under the object-memory task, repetition of a stimuluswithin a trial indicated a memory target. Under the distractortask, object stimulus repetition had no significance. We exam-ined the ratio of responses to the second stimulus presentationcompared with the first within a trial and determined thesignificance of the ratio for each cell by means of a t-test, usingonly cells whose responses were large enough to meet the
prob
abili
ty
1/16 1/4 1 4 160
0.1
0.2
0.3
0.4
0.5 TEadTEavPRh
ratio 1st/2nd peakFIG. 8. Probability distributions of the relative heights of the 2 peaks seen
in the PSTHs (Fig. 7). The ratio of the peaks was determined for each cell thatmet our significance criteria, and the pdf was then generated by kernelsmoothing estimation. The number of units in each brain area is the same asindicated in Fig. 7.
TEad0
5
10
15
20
25
spik
es/s
ec
obj-memory distractor
TEav0
5
10
15
20
25
PRh0
2
4
6
8
A
TEad TEav PRh
0.2
0.6
1.0
1.4
Bspont.
obj-m
emor
y/di
stra
ctor
FIG. 9. A: average response magnitudes in different cortical areas under the2 tasks. Spontaneous levels of activity are shown in gray. Data were pooledover stimulus duration shifted by latency. B: ratios of responses under theobject-memory and distractor tasks. Error bars indicate SE. The number ofcells in each brain area is the same as indicated in Fig. 7.
0 20 40 60 800
20
40
60
80
obje
ct ta
sk (
spik
es/s
ec) TEad
0 20 40 60 800
20
40
60
80
obje
ct ta
sk (
spik
es/s
ec) TEav
0 5 10 15 20 250
5
10
15
20
25
obje
ct ta
sk (
spik
es/s
ec) PRh
distractor task (spikes/sec)
A
B
C
FIG. 10. Responses of individual cells plotted as average firing rate duringthe object-memory task vs. average firing rate during the distractor task. Inboth areas of TE, responses to the two tasks were almost equal, whereas inperirhinal cortex there was greater activity during the memory task. Greenpoints indicate cells whose response ratios for the 2 tasks were different from1.0 at the P � 0.01 level of significance.
1305ENHANCED PERIRHINAL ACTIVITY DURING A WORKING-MEMORY TASK
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

criteria we set. For TEav, repetition enhancement was seen in1% (1/131) of the units during the object-memory task and 3%(4/131) during the distractor task. TEav repetition suppressionwas seen in 19% (25/131) for the object-memory task and 11%(15/131) for the distractor task. For TEad, enhancement wasseen in 2% (1/52) for the object-memory task and 8% (4/52)for the distractor task. Suppression occurred in 11% (6/52) forboth tasks. Finally, for PRh, enhancement occurred for 0%(0/36) during the object-memory task and 6% (2/36) during thedistractor task, whereas suppression was seen in 6% (2/36) forthe object-memory task and 3% (1/29) during the distractortask. These data are broadly consistent with previous reports(Baylis and Rolls 1987; Fahy et al. 1993; Miller and Desimone1994; Miller et al. 1993) in which repetition effects wereobserved in only small fractions of the recorded cells.
The pdf’s over all response ratios for units that met theresponse criteria described in the preceding text are plotted inFig. 12. In no case is the distribution average significantlydifferent from 1.0. The most notable difference between TEand PRh in these plots is the greater variance of the PRhdistribution. That may be due to the lower activity in PRh,leading to greater uncertainty in estimating firing rates withina fixed time window. The pdf’s for the two tasks are notsignificantly different in any brain area, under a Kolmogorov-Smirnov test, nor are the median response ratios significantlydifferent for the two tasks under a Wilcoxon signed rank pairedcomparison. Furthermore, there were no significant serial po-sition effects on the magnitude of the ratio, performingANOVA as a function of the number of items separating thefirst and second presentation of the stimulus image. Thus wesee no influence of the memory task on repetition effects.
D I S C U S S I O N
Dealing first with the perceptual aspect of perirhinal re-sponses, our data indicate lower stimulus selectivity in PRhcompared with TE. Selectivity determinations were based oncalculating entropies of response probability density functions(Fig. 4). The pdf of PRh lacks a rightward tail of largeresponses, making the distribution closer to normal than that ofTE. Under an entropy measure of selectivity (as well as akurtosis measure), that is indicative of lower selectivity. Incontrast, Naya et al. (2003) report that they found essentiallyno difference in stimulus selectivity between PRh and TE.
Whether PRh is less selective than TE, as we observed, orequally selective as observed by Naya et al. (2003), the
significant point is that PRh is not more selective than TE.Thus these data seemingly fail to support the perceptual-mnemonic model of PRh function (Buckley et al. 2001; Busseyet al. 2005; Murray and Bussey 1999), which holds that PRhplays a perceptual role in vision by forming more complexconjunctions of features than TE in addition to its mnemonicrole. If PRh was forming more complex conjunctions offeatures, one would expect it to show greater visual selectivitythan TE.
Nevertheless, we shall offer two suggestions how observa-tion of lower selectivity in PRh could still be compatible withthe perceptual-mnemonic model.
The first possibility is that the measured selectivity in PRhhas been underestimated because of the limited size of thestimulus set (110 objects). To see how this might happen,imagine how the response statistics of a very highly selectivecell might appear if the (rare) effective stimuli for that cellweren’t included in the stimulus set. Lacking any large re-sponses, the cell’s pdf would likely be quasi-normally distrib-
obj-memory distractor
spik
es/s
ec
2 4 10
5
10
15
20
25TEav
6 8response rank
2 4 10
5
10
15
20
25
response rank
TEad
6 8 2 4 10
4
5
6
7
8 PRh
6 8response rank
FIG. 11. Average response as a function of response rank for the 10 stimulipresented to each cell. Each cell had a different set of 10, selected from 110objects based on preliminary testing. The parallel shift in PRh indicates thatmemory processing does not change the stimulus selectivity of cells. Thenumber of units in each brain area is the same as indicated in Fig. 7.
0.25 0.5 1.0 2.0 4.00
2.0
TEad
prob
abili
ty d
ensi
ty
1.0
distractorobj-memory
0.25 0.5 1.0 2.0 4.00
TEav
prob
abili
ty d
ensi
ty
1.0
2.0distractorobj-memory
0.25 0.5 1.0 2.0 4.00
0.2
0.4
0.6PRh
prob
abili
ty d
ensi
ty
ratio 2nd/1st presentation
distractorobj-memory
A
B
C
FIG. 12. Repetition suppression and repetition enhancement. These arepdf’s of the ratios of response magnitudes during 1st and 2nd presentations ofthe same stimulus within the same trial. The pdf’s show that stimulus repetitioneffects are not significantly different for the 2 tasks in any of the cortical areas.The number of units in each brain area is the same as indicated in Fig. 7.
1306 S. R. LEHKY AND K. TANAKA
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

uted around some low firing rate rather similar to what we seein the PRh data, giving the false appearance of low selectivity.Including just one effective stimulus dramatically changes theselectivity calculations, but it is unlikely that we would bychance include something approximating that image within asmall stimulus set. Because of this potential problem, themeasurement here of lower selectivity in PRh relative to TE isopen to question. It may take thousands of object stimuli toproperly characterize neurons whose receptive fields have thislevel of complexity.
Alternatively, the lower selectivity observed here in PRhrelative to TE may not be a sampling artifact, as suggested inthe preceding text, but rather may reflect the possibility thatPRh is involved in visual categorization to a greater degreethan TE. Object categorization requires cells to generalizeacross stimuli and therefore be less selective, contrary to thehigh selectivity required for object identification. Cells ininferotemporal cortex are known to be involved in visualcategorization (Freedman et al. 2003; Hung et al. 2005; Sigalaand Logothetis 2002; Vogels 1999), although that aspect oftheir properties has not been compared between PRh and TE.Therefore under this view, the advance from TE to PRh, ratherthan continuing the pattern of increasing selectivity as oneascends in visual pathways, may mark the transition to aqualitatively distinct and more abstract functional mode inwhich object-based rather than feature-based representationspredominate. Studies indicating that PRh is involved in pro-cessing more complex visual stimuli than TE (Buckley et al.2001; Bussey et al. 2002, 2003, 2005; Eacott et al. 1994;Murray and Bussey 1999; Tyler et al. 2004), together with theobservation here that PRh neurons are visually less selectivethan those of TE, are consistent with the idea that PRh ispreferentially involved in object-based categorization ratherthan feature-based processing.
Moving from perception to memory, there was a significantincrease in activity in perirhinal cells during the object-mem-ory task compared with the distractor task (Figs. 7, 9, and 10).Similar task-related changes in responses were not seen in theadjacent visual areas that project directly to PRh, areas TEadand TEav. As our task involved the use of a small set (10) ofhighly familiar stimuli, in which every member of the setappeared repeatedly within a short period of time (severaltimes per minute), solving the task would invoke predomi-nantly working-memory mechanisms rather than long-termmechanisms. That being the case, we would interpret theincreased PRh activity during the object-memory task as re-flecting some aspect of the neural activity required when visualobjects are being processed by working memory.
Although we observed a task-specific increase in neuralresponses for objects being held in working memory, we didnot observe any task-specific changes in repetition effects,either repetition enhancement or repetition suppression. Thusthere is no basis in these data for associating such effects withprocessing during our working-memory task. A dissociationbetween performance during a working-memory DMS task andrepetition suppression has previously been shown by Millerand Desimone (1993). During systemic application of a cho-linergic blocker, performance in the task was disrupted, butthere was no change in repetition suppression in PRh. As yet,this test has not been applied to repetition enhancement orother putative working memory correlates.
The designs of the tasks used here do not allow us toassociate the elevated PRh activity with specific stages withinthe working-memory process. Unlike the delayed match tosample task, in which encoding, maintenance and retrieval areseparated in time, in our serial recognition task all threeprocesses occur concurrently, as the sequence of stimulusitems are presented within each trial. In compensation for thisloss of temporal separability among working-memory subpro-cesses, our task offered a more complex memory load thatincreased the contrast between the memory and nonmemory(distractor) conditions.
Working-memory effects have been detected in a number ofearlier visually responsive areas (V1, V4, MT), using simplesynthetic stimuli such as spots, bars, and stripes (Bisley et al.2004; Haenny et al. 1988; Maunsell et al. 1991; Motter 1994b;Pasternak and Greenlee 2005; Super et al. 2001). Given thoseobservations, the question arises why the memory-related ef-fects we saw were specific to PRh and did not include theearlier area TE. A possible explanation arises when one con-siders psychophysical evidence that for complex stimuli, visualworking-memory stores integrated objects rather than individ-ual features (Lee and Chun 2001; Luck and Vogel 1997). Itwas suggested in the preceding text that PRh is the site wherefeature clusters are integrated into more holistic object repre-sentations, whereas TE still maintains representations at thefeature level. If that were so, then working-memory effectsusing more naturalistic complex objects might then be prefer-entially associated with PRh.
Response PSTHs in all cortical areas had two peaks. Thefirst peak most likely represents feedforward flow of responsesup from the retina. The second peak appears around 150–180ms after the first peak and may represent recurrent input.Elevated PRh activity during the object-memory task is asso-ciated with the second peak (Fig. 7C), starting �80 ms after theonset of the stimulus response. The latency between the onsetof the stimulus response and the onset of the putative memory-related activity suggests the possibility that such activity isdependent on top-down feedback. Given the close associationbetween working memory and lateral prefrontal cortex that wasoutlined earlier, a likely source of this feedback would beprefrontal cortex, although these data cannot exclude othersources. We therefore view these data as consistent withmodels of working memory postulating that object representa-tions within posterior cortices (forming the contents of workingmemory) are controlled or activated by feedback signals fromprefrontal cortex (Curtis and D’Esposito 2003; Petrides 1994,2000; Postle 2006; Postle et al. 1999). This prefrontal cortexsignal may serve to tag particular posterior visual representa-tions as behaviorally significant for the task at hand.
Thus under the interpretation here, as the monkey switchesback and forth between the object-memory task and the dot-memory distractor task (both basically the same task, but withdifferent stimuli), the memory-related feedback signal fromprefrontal cortex switches back and forth between the neuralpopulations forming the representations of the two sets ofstimuli (Fig. 13). The cells we recorded from were selected tobe responsive to just one stimulus class (the complex stimuli ofthe object-memory task) and not the other (the simple patternsof the dot-memory-distractor task). Therefore we observed thememory related signal turning on and off depending on whichneural representation was the target of prefrontal control.
1307ENHANCED PERIRHINAL ACTIVITY DURING A WORKING-MEMORY TASK
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

As there is a close connection between working memory andattention (Awh and Jonides 2001; Awh et al. 2006; Barnes etal. 2001; de Fockert et al. 2001; Desimone 1998), the mecha-nisms underlying the memory-related activation we observedin PRh could extensively overlap with mechanisms involved inattentional selection. The changes in PRh activity cannot beascribed to shifts in spatial attention (for example, see Moranand Desimone 1985), as the stimuli used in the object-memoryand distractor tasks were in overlapping locations. On the otherhand, it is more difficult to exclude the possibility of a contri-bution from object attention, if indeed working memory forobjects and selective attention for objects are independentmechanisms. It is well established that attention can selectfeatures or objects nonspatially as demonstrated psychophysi-cally (Awh et al. 2001; Duncan 1984; Lee and Chun 2001;Vecera and Farah 1994), neurophysiologically (Chelazzi et al.1993; Hayden and Gallant 2005; McAdams and Maunsell2000; Motter 1994a; Sereno and Amador 2006; Treue andTrujillo 1999) and by brain imaging (Corbetta et al. 1991; Liuet al. 2003; O’Craven et al. 1999; Serences et al. 2004).
Also, given that relevant and nonrelevant stimuli in our taskoccurred at predictable times, another possible contributor toour results is temporal attention, the ability to direct attentionto particular points in time. Although not as extensively studiedas object attention, temporal attention has been demonstratedwith human brain imaging (Coull and Nobre 1998; Griffin etal. 2002; Nobre 2001) as well as monkey single-cell recordings(Ghose and Maunsell 2002).
Again suggestive of an overlap between working memoryand attentional mechanisms, an analogy may be drawn be-tween what appears to be feedback activation of PRh duringour working-memory task and feedback activation of variousposterior cortices during attention (reviewed by Pessoa et al.2003).
To summarize, in a paired comparison between two tasks wehave observed activity in PRh associated with the requirementfor keeping complex objects in visual working memory. Theactivity did not extend into the adjacent visual cortex, area TE.There was a long latency between the onset of the stimulus
response and the onset of the memory-related activation, con-sistent with a top-down feedback source of the activation. Thenature of this feedback activation during working memory mayoverlap extensively with feedback activation associated withattentional selection. These results support models of visualworking memory in which prefrontal cortex exerts a control-ling influence on visual representations held within posteriorcortices.
A C K N O W L E D G M E N T S
We thank K. Saleem for assistance with histology, J. Maunsell for providingdata-acquisition software as well as M. Mishkin, A. Sereno, and L. Chen forcomments on the manuscript.
G R A N T S
This research was partly supported by a Grant-in-Aid for Scientific Re-search on Priority Areas 17022047 from Japanese Ministry of Education,Sports, Science and Technology.
R E F E R E N C E S
Alvarado MC, Bachevalier J. Comparison of the effects of damage to theperirhinal and parahippocampal cortex on transverse patterning and locationmemory in rhesus macaques. J Neurosci 25: 1599–1609, 2005.
Atkinson RC, Shiffrin RM. The control of short-term memory. Sci Am 225:82–90, 1971.
Atkinson RC, Shiffrin RM. Human memory: A proposed system and itscontrol processes. In: The Psychology of Learning and Motivation: Ad-vances in Research and Theory, edited by Spence KW and Spence JT. SanDiego, CA: Academic, 1968, vol. 2, p. 89–195.
Awh E, Dhaliwal H, Christensen S, Matsukara M. Evidence for twocomponents of object-based selection. Psychol Sci 12: 329–334, 2001.
Awh E, Jonides J. Overlapping mechanisms of attention and spatial workingmemory. Trends Cogn Sci 5: 119–126, 2001.
Awh E, Vogel EK, and Oh S-H. Interactions between attention and workingmemory. Neuroscience 139: 201–208, 2006.
Barnes LL, Nelson JK, Reuter-Lorenz PA. Object-based attention andobject working memory: overlapping processes revealed by selective inter-ference effects in humans. Prog Brain Res 134: 471–481, 2001.
Baxter MG, Murray EA. Opposite relationship of hippocampal and rhinalcortex damage to delayed nonmatching-to-sample deficits in monkeys.Hippocampus 11: 61–71, 2001.
Baylis GC, Rolls ET. Responses of neurons in the inferior temporal cortex inshort term and serial recognition memory tasks. Exp Brain Res 65: 614–622, 1987.
Bisley JW, Zaksas D, Droll JA, Pasternak T. Activity of neurons in corticalarea MT during a memory for motion task. J Neurophysiol 91: 286–300,2004.
Brown MW, Aggleton JP. Recognition memory: what are the roles of theperirhinal cortex and hippocampus? Nat Rev Neurosci 2: 51–61, 2001.
Buckley MJ, Booth MCA, Rolls ET, Gaffan D. Selective perceptual impair-ments after perirhinal cortex ablation. J Neurosci 21: 9824–9836, 2001.
Buckley MJ, Gaffan D. Impairment of visual object-discrimination learningafter perirhinal cortex ablation. Behav Neurosci 111: 467–475, 1997.
Buckley MJ, Gaffan D, Murray EA. Functional double dissociation betweentwo inferior temporal cortical areas: perirhinal cortex versus middle tempo-ral gyrus. J Neurophysiol 77: 587–598, 1997.
Buffalo EA, Ramus S, Clark R, Teng E, Squire LR, Zola SM. Dissociationbetween the effects of damage to perirhinal cortex and area TE. Learn Mem6: 572–599, 1999.
Bussey TJ, Saksida LM, Murray EA. Perirhinal cortex resolves featureambiguity in complex visual discriminations. Eur J Neurosci 15: 365–374,2002.
Bussey TJ, Saksida LM, Murray EA. Impairments in visual discriminationafter perirhinal cortex lesions: testing “declarative” vs. “perceptual-mne-monic” views of perirhinal cortex function. Eur J Neurosci 17: 649–660,2003.
Bussey TJ, Saksida LM, Murray EA. The perceptual-mnemonic/featureconjunction model of perirhinal cortex function. Q J Exp Psychol 58B:269–282, 2005.
Castro CA, Larsen T. Primacy and recency effects in nonhuman primates. JExp Psychol Anim Behav Process 18: 335–340, 1992.
Prefrontal Cortex
Perirhinal Cortex
Object-memory task Dot-memory distractor task
Neural representation of object stimulus
Neural representation of dot stimulus
FIG. 13. Model of prefrontal-perirhinal interactions during visual workingmemory. During the object-memory task, feedback from prefrontal cortexactivated population-coded neural representations of the object stimuli inperirhinal cortex. During the dot-memory task, the prefrontal feedback signalswitched to neural representations of the dot stimuli. As we recorded only fromneurons that were part of the object representations, the prefrontal signalappeared to turn on and off as it was switched back and forth between the 2neuronal populations.
1308 S. R. LEHKY AND K. TANAKA
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

Chelazzi L, Miller EK, Duncan J, Desimone R. A neural basis for visualsearch in inferior temporal cortex. Nature 363: 345–347, 1993.
Corbetta M, Miezin FM, Dobmeyer S, Shulman GL, Petersen SE. Selec-tive and divided attention during visual discrimination of shape, color, andspeed: functional anatomy by positron emission tomography. J Neurosci 11:2383–2402, 1991.
Coull JT, Nobre AC. Where and when to pay attention: the neural systems fordirecting attention to spatial locations and to time intervals as revealed byboth PET and fMRI. J Neurosci 18: 7426–7435, 1998.
Courtney SM. Attention and cognitive control as emergent properties ofinformation representation in working memory. Cogn Affect Behav Neurosci4: 501–516, 2004.
Curtis CE, D’Esposito M. Persistent activity in the prefrontal cortex duringworking memory. Trends Cogn Sci 7: 415–423, 2003.
Davachi L, Goldman-Rakic P. Primate rhinal cortex participates in bothvisual recognition and working memory tasks: functional mapping with2-DG. J Neurophysiol 85: 2590–2601, 2001.
de Fockert JW, Rees G, Frith CD, Lavie N. The role of working memory invisual selective attention. Science 291: 1803–1806, 2001.
Desimone R. Stimulus-selective properties of inferior temporal neurons in themacaque. J Neurosci 4: 2051–2062, 1984.
Desimone R. Neural mechanisms for visual memory and their role in attention.Proc Natl Acad Sci USA 93: 13494–13499, 1996.
Desimone R. Visual attention mediated by biased competition in extrastriatevisual cortex. Philos Trans R Soc Lond B Biol Sci 353: 1245–1255, 1998.
Duncan J. Selective attention and the organization of visual information. JExp Psychol Gen 113: 501–517, 1984.
Eacott MJ, Gaffan D, Murray EA. Preserved recognition memory for smallsets, and impaired stimulus identification for large sets, following rhinalcortex ablations in monkeys. Eur J Neurosci 6: 1466–1478, 1994.
Eskandar EN, Richmond BJ, Optican LM. Role of inferior temporalneurons in visual memory. I. Temporal encoding of information about visualimages, recalled images, and behavioral context. J Neurophysiol 68: 1277–1295, 1992.
Fahy FL, Riches IP, Brown MW. Neuronal activity related to visual recog-nition memory: long-term memory and the encoding of recency and famil-iarity information in the primate anterior and medial inferior temporal andrhinal cortex. Exp Brain Res 96: 457–472, 1993.
Freedman DJ, Riesenhuber M, Poggio T, Miller EK. A comparison ofprimate prefrontal and inferior temporal cortices during visual categoriza-tion. J Neurosci 23: 5325–5246, 2003.
Fuster JM. Unit activity in prefrontal cortex during delayed-response perfor-mance: neuronal correlates of transient memory. J Neurophysiol 36: 61–78,1973.
Fuster JM. Inferotemporal units in selective visual attention and short-termmemory. J Neurophysiol 64: 681–697, 1990.
Fuster JM, Alexander GE. Neuron activity related to short term memory.Science 173: 652–654, 1971.
Fuster JM, Jervey JP. Inferotemporal neurons distinguish and retain behav-iorally relevant features of visual stimuli. Science 212: 952–955, 1981.
Gaffan D. Against memory systems. Philos Trans R Soc Lond B Biol Sci 357:1111–1121, 2002.
Ghose GM, Maunsell JH. Attentional modulation in visual cortex depends ontask timing. Nature 419: 616–620, 2002.
Glanzer M, Cunitz AR. Two storage mechanisms in free recall. J VerbalLearn Verbal Behav 5: 351–360, 1966.
Goldman-Rakic P. Circuitry of the prefrontal cortex and the regulation ofbehavior by representational memory. In: Handbook of Physiology.TheNervous System. Higher Functions of the Brain., edited by Plum F. Be-thesda: Am. Physiol. Soc., 1987, sect. 1, vol. V, part 1, p. 373–417.
Griffin IC, Miniussi C, Nobre AC. Multiple mechanisms of selective atten-tion: differential modulation of stimulus processing by attention to space ortime. Neuropsychologia 40: 2325–2340, 2002.
Haenny PE, Maunsell JH, Schiller PH. State dependent activity in monkeyvisual cortex. II. Retinal and extraretinal factors in V4. Exp Brain Res 69:245–259, 1988.
Hampton RR. Monkey perirhinal cortex is critical for visual memory, but notfor visual perception: reexamination of the behavioural evidence frommonkeys. Q J Exp Psychol 58B: 283–299, 2005.
Hampton RR, Murray EA. Learning of discriminations is impaired, butgeneralization to altered views is intact, in monkeys (Macaca mulatta) withperirhinal cortex removal. Behav Neurosci 116: 363–377, 2002.
Hayden BY, Gallant JL. Time course of attention reveals different mecha-nisms for spatial and feature-based attention in area V4. Neuron 47:637–643, 2005.
Holscher C, Rolls ET. Perirhinal cortex neuronal activity is actively related toworking memory in the macaque. Neural Plas 9: 41–51, 2002.
Hung CP, Kreiman G, Poggio T, DiCarlo JJ. Fast readout of object identityfrom macaque inferior temporal cortex. Science 310: 863–866, 2005.
Jacobsen CF. The functions of the frontal association areas in monkeys. CompPsychol Monogr 13: 1–60, 1936.
Kondo H, Komatsu H. Suppression on neuronal responses by a metacontrastmasking stimulus in monkey V4. Neurosci Res 36: 27–33, 2000.
Kovacs G, Vogels R, Orban GA. Cortical correlate of pattern backwardmasking. Proc Natl Acad Sci USA 92: 5587–5591, 1995.
Kubota K, Niki H. Prefrontal cortical unit activity and delayed alternationperformance in monkeys. J Neurophysiol 34: 337–347, 1971.
Lavenex P, Suzuki WA, Amaral DG. Perirhinal and parahippocampal cor-tices of the macaque monkey: projections to the neocortex. J Comp Neurol447: 394–420, 2002.
Lebedev MA, Messinger A, Kralik JD, Wise SP. Representation of attendedversus remembered locations in prefrontal cortex. PLOS Biol 2: e365, 2004.
Lee ACH, Bussey TJ, Murray EA, Saksida LM, Epstein RA, Kapur N,Hodges JR, Graham KS. Perceptual deficits in amnesia: challenging themedial temporal lobe “mnemonic” view. Neuropsychologia 43: 1–11, 2005.
Lee D, Chun MM. What are the units of visual short-term memory, objects orspatial locations? Percept Psychophys 63: 253–257, 2001.
Lehky SR. Bayesian estimation of stimulus responses in Poisson spike trains.Neural Comput 16: 1325–1343, 2004.
Lehky SR, Sejnowski TJ, Desimone R. Selectivity and sparseness in theresponses of striate complex cells. Vision Res 45: 57–73, 2005.
Levy DA, Shrager Y, Squire LR. Intact visual discrimination of complex andfeature-ambiguous stimuli in the absence of perirhinal cortex. Learn Mem12: 61–66, 2005.
Liu T, Slotnick SD, Serences JT, Yantis S. Cortical mechanisms of feature-based attentional control. Cereb Cortex 13: 1334–1443, 2003.
Liu Z, Richmond BJ. Response differences in monkey TE and perirhinalcortex: stimulus association related to reward schedules. J Neurophysiol 83:1677–1692, 2000.
Luck SJ, Vogel EK. The capacity of visual working memory for features andconjunctions. Nature 390: 279–281, 1997.
Macknik SL, Livingstone MS. Neuronal correlates of visibility and invisi-bility in the primate visual system. Nat Neurosci 1: 144–149, 1998.
Maunsell JH, Sclar G, Nealey TA, DePriest DD. Extraretinal representationsin area V4 in the macaque monkey. Vis Neurosci 7: 561–573, 1991.
McAdams CJ, Maunsell JH. Attention to both space and feature modulatesneuronal responses in macaque area V4. J Neurophysiol 83: 1751–1755,2000.
Meunier M, Bachevalier J, Mishkin M, Murray EA. Effects on visualrecognition of combined and separate ablations of the entorhinal andperirhinal cortex in rhesus monkeys. J Neurosci 13: 5418–5432, 1993.
Miller EK, Desimone R. Scopolamine affects short-term memory but notinferior temporal neurons. Neuroreport 4: 81–84, 1993.
Miller EK, Desimone R. Parallel neuronal mechanisms for short-term mem-ory. Science 263: 520–522, 1994.
Miller EK, Erickson CA, Desimone R. Neural mechanisms of visual workingmemory in prefrontal cortex of the macaque. J Neurosci 16: 5154–5167,1996.
Miller EK, Li L, Desimone R. A neural mechanism for working andrecognition memory in inferior temporal cortex. Science 254: 1377–1379,1991.
Miller EK, Li L, Desimone R. Activity of neurons in anterior inferiortemporal cortex during a short-term memory task. J Neurosci 13: 1460–1478, 1993.
Mishkin M. Effects of small frontal lesions on delayed alternation in monkeys.J Neurophysiol 20: 615–622, 1957.
Miyashita Y. Inferior temporal cortex: where visual perception meets mem-ory. Annu Rev Neurosci 16: 245–263, 1993.
Miyashita Y, Chang HS. Neuronal correlate of pictorial short-term memoryin the primate temporal cortex. Nature 331: 68–70, 1988.
Mogami T, Tanaka K. Reward association affects neuronal responses tovisual stimuli in macaque te and perirhinal cortices. J Neurosci 26: 6761–6770, 2006.
Moran J, Desimone R. Selective attention gates visual processing in theextrastriate cortex. Science 229: 782–784, 1985.
1309ENHANCED PERIRHINAL ACTIVITY DURING A WORKING-MEMORY TASK
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from

Motter BC. Neural correlates of attentive selection for color or luminance inextrastriate area V4. J Neurosci 14: 2178–2189, 1994a.
Motter BC. Neural correlates of feature selective memory and pop-out inextrastriate area V4. J Neurosci 14: 2190–2199, 1994b.
Murdock BB. The serial position effect in free recall. J Exp Psychol 64:482–488, 1962.
Murray EA, Bussey TJ. Perceptual–mnemonic functions of the perirhinalcortex. Trends Cogn Sci 3: 142–151, 1999.
Murray EA, Mishkin M. Object recognition and location memory in mon-keys with excitotoxic lesions of the amygdala and hippocampus. J Neurosci18: 6568–6582, 1998.
Nakamura K, Kubota K. Mnemonic firing of neurons in the monkeytemporal pole during a visual recognition memory task. J Neurophysiol 74:162–178, 1995.
Naya Y, Yoshida M, Miyashita Y. Backward spreading of memory-retrievalsignal in the primate temporal cortex. Science 291: 661–664, 2001.
Naya Y, Yoshida M, Miyashita Y. Forward processing of long-term asso-ciative memory in monkey inferotemporal cortex. J Neurosci 23: 2861–2871, 2003.
Niki H. Prefrontal unit activity during delayed alternation in the monkey. I.Relation to direction of response. Brain Res 68: 185–196, 1974.
Nobre AC. Orienting attention to instants in time. Neuropsychologia 39:1317–1328, 2001.
O’Craven KM, Downing PE, Kanwisher N. fMRI evidence for objects as theunits of attentional selection. 401: 584–587, 1999.
Pasternak T, Greenlee MW. Working memory in primate sensory systems.Nat Rev Neurosci 6: 97–107, 2005.
Pessoa L, Kastner S, Ungerleider LG. Neuroimaging studies of attention:from modulation of sensory processing to top-down control. J Neurosci 23:3990–3998, 2003.
Petrides M. Frontal lobes and working memory: evidence from investigationsof the effects of cortical excisions in nonhuman primates. In: Handbook ofNeuropsychology, edited by Boller F and Grafman J. New York: Elsevier,1994, p. 59–82.
Petrides M. Dissociable roles of mid-dorsolateral prefrontal and anteriorinferotemporal cortex in visual working memory. J Neurosci 20: 7496–7503, 2000.
Postle BR. Working memory as an emergent property of the mind and brain.Neuroscience 139: 23–38, 2006.
Postle BR, Berger JS, D’Esposito M. Functional neuroanatomical doubledissociation of mnemonic and executive control processes contributing toworking memory performance. Proc Natl Acad Sci USA 96: 12959–12964,1999.
Raymond JE, Shapiro KI, Arnell KM. Temporary suppression of visualprocessing in an RSVP task: an attentional blink? J Exp Psychol HumPercept Perform 18: 849–860, 1992.
Rempel-Clower NL, Barbas H. The laminar pattern of connections betweenprefrontal and anterior temporal cortices in the Rhesus monkey is related tocortical structure and function. Cereb Cortex 10: 851–865, 2000.
Rieke F, Warlan D, de Ruyter van Steveninck R, Bialek W. Spikes:Exploring the Neural Code. Cambridge, MA: MIT Press, 1997.
Rolls ET, Tovee MJ. Processing speed in the cerebral cortex and theneurophysiology of visual masking. Proc R Soc Lond B Biol Sci 257: 9–15,1994.
Rolls ET, Tovee MJ. Sparseness of the neuronal representation of stimuli inthe primate temporal visual cortex. J Neurophysiol 73: 713–726, 1995.
Rolls ET, Tovee MJ, Panzeri S. The neurophysiology of backward visualmasking: information analysis. J Cogn Neurosci 11: 300–311, 1999.
Saleem KS, Tanaka K. Divergent projections from the anterior inferotempo-ral area TE to the perirhinal and entorhinal cortices in the macaque monkey.J Neurosci 16: 4757–4775, 1996.
Sands SF, Wright AA. Primate memory: retention of serial list items by arhesus monkey. Science 209: 938–940, 1980.
Serences JT, Schwarzbach J, Courtney SM, Golay X, Yantis S. Control ofobject-based attention in human cortex. Cereb Cortex 14: 1346–1357, 2004.
Sereno AB, Amador SC. Attention and memory related responses of neuronsin the lateral intraparietal area during spatial and shape delayed match-to-sample tasks. J Neurophysiol 95: 1078–1098, 2006.
Sigala N, Logothetis NK. Visual categorization shapes feature selectivity inthe primate temporal cortex. Nature 415: 318–320, 2002.
Simoncelli EP, Olshausen BA. Natural image statistics and neural represen-tation. Annu Rev Neurosci 24: 1193–1216, 2001.
Sobotka S, Diltz MD, Ringo JL. Can delay-period activity explain workingmemory? J Neurophysiol 93: 128–136, 2005.
Squire LR, Stark CEL, Clark RE. The medial temporal lobe. Annu RevNeurosci 27: 279–306, 2004.
Super H, Spekreijse H, Lamme VAF. A neural correlate of working memoryin the monkey primary visual cortex. Science 293: 120–124, 2001.
Suzuki WA, Amaral DG. Perirhinal and parahippocampal cortices of themacaque monkey: cortical afferents. J Comp Neurol 350: 497–533, 1994.
Takeda M, Naya Y, Fujimichi R, Takeuchi D, Miyashita Y. Activemaintenance of associative mnemonic signal in monkey inferior temporalcortex. Neuron 48: 839–848, 2005.
Tamura H, Tanaka K. Visual response properties of cells in the ventral anddorsal parts of the macaque inferotemporal cortex. Cereb Cortex 11:384–399, 2001.
Tanaka K. Inferotemporal cortex and object vision. Annu Rev Neurosci 19:109–139, 1996.
Tang Y, Mishkin M, Aigner TG. Effects of muscarinic blockade in perirhinalcortex during visual recognition. Proc Natl Acad Sci USA 94: 12667–12669,1997.
Treue S, Trujillo JC. Feature-based attention influences motion processinggain in macaque visual cortex. Nature 399: 575–579, 1999.
Turchi J, Saunders RC, Mishkin M. Effects of cholinergic deafferentation ofthe rhinal cortex on visual recognition memory in monkeys. Proc Natl AcadSci USA 102: 2158–2161, 2005.
Tyler LK, Stamatakis EA, Bright P, Acres K, Abdallah S, Rodd JM, MossHE. Processing objects at different levels of specificity. J Cogn Neurosci 16:351–362, 2004.
Ungerleider LG, Gaffan D, Pelak VS. Projections from inferior temporalcortex to prefrontal cortex via the uncinate fascicle in rhesus monkeys. ExpBrain Res 76: 473–484, 1989.
Van Hoesen GW. The parahippocampal gyrus: new observations regarding itscortical connections in the monkey. Trends Neurosci 5: 345–350, 1982.
Vecera SP, Farah MJ. Does visual attention select objects or locations? J ExpPsychol Gen 123: 146–160, 1994.
Vogels R. Categorization of complex visual images by rhesus monkeys. II.Single-cell study. Eur J Neurosci 11: 1239–1255, 1999.
Webster MJ, Bachevalier J, Ungerleider LG. Connections of inferiortemporal areas TEO and TE with parietal and frontal cortex in macaquemonkeys. Cereb Cortex 4: 470–483, 1994.
Wright AA, Santiago HC, Sands SF, Kendrick DF, Cook RG. Memoryprocessing of serial lists by pigeons, monkeys, and people. Science 229:287–289, 1985.
Yakovlev V, Bernacchia A, Orlov T, Hochstein S, Amit D. Multi-itemworking memory–a behavioral study. Cereb Cortex 15: 602–615, 2004.
Zola-Morgan S, Squire LR, Amaral DG, Suzuki WA. Lesions of perirhinaland parahippocampal cortex that spare the amygdala and hippocampalformation produce severe memory impairment. J Neurosci 9: 4335–4370,1989.
1310 S. R. LEHKY AND K. TANAKA
J Neurophysiol • VOL 97 • FEBRUARY 2007 • www.jn.org
on February 20, 2007
jn.physiology.orgD
ownloaded from





























