Embed Size (px)
Citation preview

Review article
Emerging viral diseases of fish and shrimp
Peter J. WALKER1*, James R. WINTON
2
1 CSIRO Livestock Industries, Australian Animal Health Laboratory (AAHL), 5 Portarlington Road,Geelong, Victoria, Australia
2 USGS Western Fisheries Research Center, 6505 NE 65th Street, Seattle, Washington, USA
(Received 7 December 2009; accepted 19 April 2010)
Abstract – The rise of aquaculture has been one of the most profound changes in global food production ofthe past 100 years. Driven by population growth, rising demand for seafood and a levelling of productionfrom capture fisheries, the practice of farming aquatic animals has expanded rapidly to become a majorglobal industry. Aquaculture is now integral to the economies of many countries. It has providedemployment and been a major driver of socio-economic development in poor rural and coastal communities,particularly in Asia, and has relieved pressure on the sustainability of the natural harvest from our rivers,lakes and oceans. However, the rapid growth of aquaculture has also been the source of anthropogenicchange on a massive scale. Aquatic animals have been displaced from their natural environment, cultured inhigh density, exposed to environmental stress, provided artificial or unnatural feeds, and a prolific globaltrade has developed in both live aquatic animals and their products. At the same time, over-exploitation offisheries and anthropogenic stress on aquatic ecosystems has placed pressure on wild fish populations. Notsurprisingly, the consequence has been the emergence and spread of an increasing array of new diseases.This review examines the rise and characteristics of aquaculture, the major viral pathogens of fish andshrimp and their impacts, and the particular characteristics of disease emergence in an aquatic, rather thanterrestrial, context. It also considers the potential for future disease emergence in aquatic animals asaquaculture continues to expand and faces the challenges presented by climate change.
disease emergence / shrimp / fish / virus
Table of contents
1. Introduction........................................................................................................................................... 21.1. The rapid growth of aquaculture................................................................................................. 21.2. Diversity of farmed aquatic species ............................................................................................ 21.3. Characteristics of fish and shrimp production systems .............................................................. 21.4. The decline of capture fisheries .................................................................................................. 3
2. History of disease emergence in fish and shrimp ................................................................................ 32.1. Problems associated with emerging diseases .............................................................................. 32.2. Emerging viral diseases of fish ................................................................................................... 42.3. Emerging viral diseases of shrimp .............................................................................................. 8
3. Impacts of emerging diseases of fish and shrimp.............................................................................. 133.1. Economic and social impacts .................................................................................................... 133.2. Environmental impacts .............................................................................................................. 13
* Corresponding author: [email protected]
Vet. Res. (2010) 41:51DOI: 10.1051/vetres/2010022
� INRA, EDP Sciences, 2010
www.vetres.org
This is an Open Access article distributed under the terms of the Creative Commons Attribution-Noncommercial License(http://creativecommons.org/licenses/by-nc/3.0/), which permits unrestricted use, distribution, and reproduction in anynoncommercial medium, provided the original work is properly cited.
Article published by EDP Sciences

4. Factors contributing to disease emergence in aquatic animals.......................................................... 144.1. Activities related to the global expansion of aquaculture ........................................................ 154.2. Improved surveillance................................................................................................................ 164.3. Natural movement of carriers.................................................................................................... 164.4. Other anthropogenic factors ...................................................................................................... 16
5. Future disease emergence risks .......................................................................................................... 17
1. INTRODUCTION
1.1. The rapid growth of aquaculture
The farming of fish and other aquatic ani-mals is an ancient practice that is believed todate back at least 4 000 years to pre-feudalChina. There are also references to fish pondsin The Old Testament and in Egyptian hiero-glyphics of the Middle Kingdom (2050–1652BC). Fish farms were common in Europe inRoman times and a recent study of land formsin the Bolivian Amazon has revealed a complexarray of fish weirs that pre-date the Hispanic era[30, 49]. However, despite its ancient origins,aquaculture remained largely a low-level, sub-sistence farming activity until the mid-20thcentury when experimental husbandry practicesfor salmon, trout and an array of tropical fishand shrimp species were developed andadopted. Aquaculture is now a major globalindustry with total annual production exceeding50 million tonnes and estimated value of almostUS$ 80 billion [32]. With an average annualgrowth of 6.9% from 1970–2007, it has beenthe fastest growing animal food-producing sec-tor and will soon overtake capture fisheries asthe major source of seafood [31].
1.2. Diversity of farmed aquatic species
In contrast to other animal production sec-tors, aquaculture is highly dynamic and charac-terised by enormous diversity in both the rangeof farmed species and in the nature of produc-tion systems. Over 350 different species ofaquatic animals are farmed, including 34 finfish(piscean), 8 crustacean and 12 molluscan spe-cies each for which annual production exceeds100 000 tonnes [32]. Aquatic animals are
farmed in freshwater, brackishwater and marineenvironments, and in production systems thatinclude caged enclosures, artificial lakes,earthen ponds, racks, rafts, tanks and raceways.Farming can be a small-scale traditional activitywith little human intervention, through tosophisticated industrial operations in which ani-mals are bred and managed for optimal perfor-mance and productivity. The diversity ofaquaculture species and farming systems alsoextends geographically from tropical to sub-arctic climes and from inland lakes and riversto estuaries and open offshore waters.
Aquaculture production is heavily domi-nated by China and other developing countriesin the Asia-Pacific region which accounts for89% by volume of global production and77% by value [31]. The major farmed speciesare carp, oysters and shrimp of which 98%,95% and 88% of production, respectively, orig-inates in Asia. By contrast, Atlantic salmon pro-duction is dominated by Norway, Chile, theUnited Kingdom and Canada which togetheraccount for 88% by volume and 94% by value[32]. Capture fisheries and aquaculture, directlyor indirectly, play an essential role in the liveli-hoods of millions of people, particularly indeveloping countries. In 2006, an estimated47.5 million people were primarily or occasion-ally engaged in primary production of aquaticanimals [31].
1.3. Characteristics of fish and shrimpproduction systems
Although a significant component of thesmall-scale aquaculture sector continues to relyon traditional methods of natural recruitment ofseed into ponds, modern fish and shrimpproduction systems more typically involve
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 2 of 24 (page number not for citation purpose)

a hatchery/nursery phase in which broodstockare spawned or stripped for the collection andhatching of eggs, and in which larvae arenursed through to post-larvae or juvenile stages(i.e., fry, smolt, fingerling) for delivery to farms.Broodstock may be captured from wild fisheriesor produced in captivity from mature farmedstock or from closed-cycle breeding and geneticimprovement programs. The use of wild brood-stock that may be healthy carriers of viralpathogens is arguably the most significant bio-security risk in aquaculture. Farming systemsemployed for grow-out may be extensiveor intensive. Extensive farming employs fishor shrimp trapped at low density in natural orman-made enclosures utilising natural sourcesof feed with minimal human intervention.Intensive systems employ medium-to-highstocking densities in cages or ponds and artifi-cial or supplemental feeds. Intensive pond cul-ture usually requires aeration and controlledwater exchange to maintain water quality.Super-intensive recirculating aquaculture sys-tems (RAS) are also employed for some spe-cies. On-farm biosecurity measures to excludepathogens and minimise health risks are morecommonly employed in intensive and super-intensive systems.
1.4. The decline of capture fisheries
Whilst aquaculture has been on a steady pathof expansion, capture fishery production haslevelled since 1990 and many of the world’smajor fisheries have been driven into a stateof decline by unsustainable fishing practicesand environmental pressures. It has been esti-mated that 11 of the 15 major fishing areasand 80% of marine fishery resources are cur-rently overexploited or at their maximum sus-tainable limit [31]. Much of the pressure onwild stocks is due to commercial fishing butthe increasing popularity of recreational anglinghas led to a growing awareness of the need forregulation to ensure marine and inland fisherysustainability. Disease emergence is also a con-cern in wild fisheries due to environmentalpressures, the direct impact of human activitiesand the risk of pathogen spread from aquacul-ture [102].
2. HISTORY OF DISEASE EMERGENCEIN FISH AND SHRIMP
Whilst various forms of disease have beenreported among aquatic animals for centuries,most were either non-infectious (e.g., tumours),or caused by common endemic pathogens(mainly parasites and bacteria), and thus alreadyknown among observers and those engaged intraditional aquaculture.However, during the pastcentury, the rise of novel forms of intensive aqua-culture, increased global movement of aquaticanimals and their products, and various sourcesof anthropogenic stress to aquatic ecosystemshave led to the emergence of many new diseasesin fish and shrimp. In this review, we considerthese emerging diseases as: (i) new or previouslyunknowndiseases; (ii) knowndiseases appearingfor first time in a new species (expanding hostrange); (iii) knowndiseases appearing for thefirsttime in a new location (expanding geographicrange); and (iv) known diseases with a newpresentation (different signs) or higher virulencedue to changes in the causative agent.
2.1. Problems associated with emerging diseases
Emerging disease epizootics frequently causesubstantial, often explosive, losses among popu-lations of fish and shrimp, resulting in largeeconomic losses in commercial aquaculture andthreats to valuable stocks of wild aquatic ani-mals. However, the extent of disease spread andimpacts are often exacerbated by other problemsthat are typically encountered including: (i) delayin developing tools for the confirmatory diagno-sis of disease or identification of the causativeagent that allows infected animals to go unde-tected; (ii) poor knowledge of the current orpotential host range; (iii) inadequate knowledgeof the present geographic range; (iv) no under-standing of critical epidemiological factors (repli-cation cycle, mode of transmission, reservoirs,vectors, stability); and (v) poor understandingof differences among strains and/or relationshipsto established pathogens. Interventions to pre-vent pathogen spread are also often limited bypoor capacity in some developing countries forimplementation of effective quarantine and/orbiosecurity measures and the illegal or poorly
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 3 of 24

regulated international trans-boundary move-ment of live aquatic animals [128, 164].
2.2. Emerging viral diseases of fish
Important emerging viral pathogens of fishare found among many families of vertebrateviruses that are well-known to include patho-gens of humans or domestic livestock. How-ever, there are significant differences betweenthe ecology of viral diseases of fish and thoseof humans or other terrestrial vertebrates. Themost significant amongst these differences arethat: (i) few fish viruses are known to be vec-tored by arthropods; (ii) water is a stabilizingmedium, but currents are less effective for longrange virus transmission than are aerosols;(iii) wild reservoir species are often at verylow densities (except for schooling and aggre-gate spawning stocks); (iv) fish are poikilo-therms and temperature has an exceptionallycritical role in modulating the disease processby affecting both the replication rate of the virusas well as the host immune response and otherphysiological factors involved in resistance;(v) few fish viruses are transmitted sexuallybetween adults, although high levels of someviruses are present in spawning fluids and afew viruses are transmitted vertically from adultto progeny, either intra-ovum or on the egg sur-face. However, as occurs for avian diseases,migratory fish can serve as carriers for long-range dispersal of viral pathogens.
The global expansion of finfish aquacultureand accompanying improvements in fish healthsurveillance has led to the discovery of severalviruses that are new to science. Many of theseare endemic among native populations andopportunistically spill-over to infect fish inaquaculture facilities. Other well-characterizedfish viruses (e.g., channel catfish virus,Onchorhynchus masou virus) can also causesignificant losses in aquaculture but do notseem to be increasing significantly in host orgeographic range. In the following sections,we consider the major emerging fish virus dis-eases that cause significant losses in aquacultureand are expanding in host or geographic range(Tab. I). Because of the risk of spread throughcommercial trade in finfish, many of the dis-
eases are listed as notifiable by the WorldOrganization for Animal Health (OIE).
2.2.1. Infectious haematopoietic necrosis
Infectious haematopoietic necrosis is oneof three rhabdovirus diseases of fish that arelisted as notifiable by the OIE. Originally ende-mic in the western portion of North Americaamong native species of anadromous salmon,infectious haematopoietic necrosis virus(IHNV) emerged in the 1970s to become animportant pathogen of farmed rainbow trout(Oncorhynchus mykiss) in the USA [11, 156].Subsequently, the virus was spread by the move-ment of contaminated eggs to several countriesof Western Europe and East Asia, where itemerged to cause severe losses in farmed rain-bow trout, an introduced species. Similar toother members of the genus Novirhabdovirusin the family Rhabdoviridae, IHNV contains anegative-sense, single-stranded RNA genome,approximately 11 000 nucleotides in lengthand encoding six proteins, packaged within anenveloped, bullet-shaped virion [67]. Isolatesof IHNV from North America show a strongphylogeographic signature with relatively lowgenetic diversity among isolates from sock-eye salmon (Oncorhynchus nerka) inhabitingthe historic geographic range of the virus[66]. However, isolates from trout in Europe,Japan or Korea, where the virus is emerging,show evidence of independent evolutionaryhistories following their initial introduction[29, 64, 106]. The emergence of IHNV in rain-bow trout aquaculture is accompanied bygenetic changes that appear to be related to ashift in host specificity and virulence [112].
2.2.2. Viral haemorrhagic septicaemia
Viral haemorrhagic septicaemia (VHS) isanother emerging disease caused by a fish rhab-dovirus. Similar to IHNV in morphology andgenome organization, viral haemorrhagic septi-caemia virus (VHSV) is also a member of thegenus Novirhabdovirus [67]. The virus wasinitially isolated and characterized in Europewhere it had become an important cause of lossamong rainbow trout reared in aquaculture
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 4 of 24 (page number not for citation purpose)
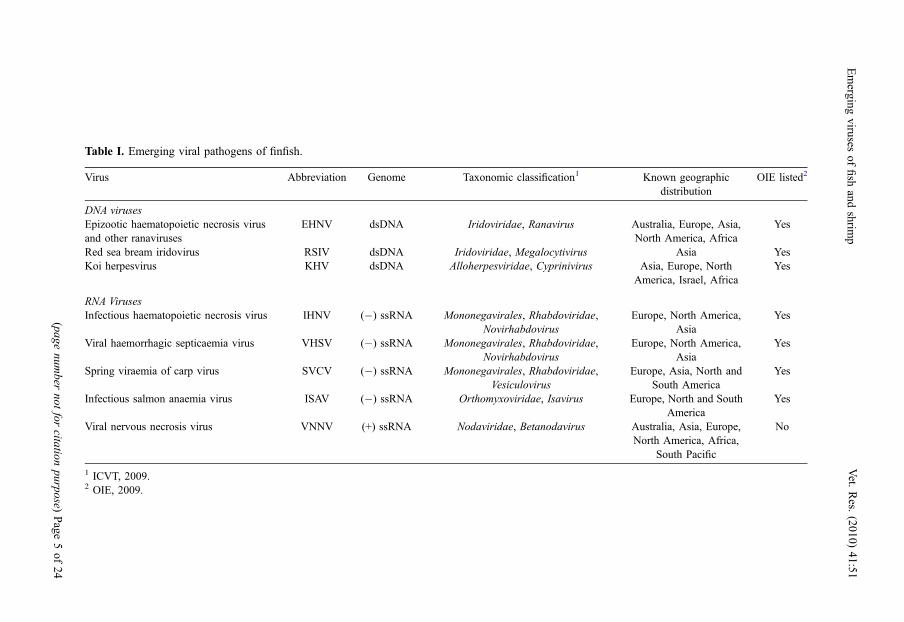
Table I. Emerging viral pathogens of finfish.
Virus Abbreviation Genome Taxonomic classification1 Known geographicdistribution
OIE listed2
DNA virusesEpizootic haematopoietic necrosis virusand other ranaviruses
EHNV dsDNA Iridoviridae, Ranavirus Australia, Europe, Asia,North America, Africa
Yes
Red sea bream iridovirus RSIV dsDNA Iridoviridae, Megalocytivirus Asia YesKoi herpesvirus KHV dsDNA Alloherpesviridae, Cyprinivirus Asia, Europe, North
America, Israel, AfricaYes
RNA VirusesInfectious haematopoietic necrosis virus IHNV (�) ssRNA Mononegavirales, Rhabdoviridae,
NovirhabdovirusEurope, North America,
AsiaYes
Viral haemorrhagic septicaemia virus VHSV (�) ssRNA Mononegavirales, Rhabdoviridae,Novirhabdovirus
Europe, North America,Asia
Yes
Spring viraemia of carp virus SVCV (�) ssRNA Mononegavirales, Rhabdoviridae,Vesiculovirus
Europe, Asia, North andSouth America
Yes
Infectious salmon anaemia virus ISAV (�) ssRNA Orthomyxoviridae, Isavirus Europe, North and SouthAmerica
Yes
Viral nervous necrosis virus VNNV (+) ssRNA Nodaviridae, Betanodavirus Australia, Asia, Europe,North America, Africa,
South Pacific
No
1 ICVT, 2009.2 OIE, 2009.
Emerging
virusesof
fish
andshrim
pVet.
Res.
(2010)41:51
(pagenum
bernot
forcitation
purpose)Page
5of
24

[123]. Prior to the 1980s, VHSV was assumedto be largely endemic among native freshwaterspecies of fish in Western Europe where itspilled-over to the introduced, and presumablymore susceptible, rainbow trout [156]. Subse-quently, an increasing number of virologicalsurveys of anadromous and marine fish in theNorth Pacific and North Atlantic Oceansrevealed a much greater host and geographicrange than previously recognized [35, 91, 99,121], and VHSV was shown to cause signifi-cant losses in both cultured and free-rangingspecies of marine fish [48, 56]. The understand-ing that VHS appeared to be an emerging dis-ease of marine fish as a result of both greatersurveillance efforts and the development ofnovel forms of marine fish aquaculture was fur-ther extended when VHS emerged for the firsttime in the Great Lakes of North America.The explosive losses among free-ranging nativespecies revealed how devastating the diseasecan be when first introduced into naive popula-tions of freshwater fish [28, 40, 84].
2.2.3. Spring viraemia of carp
Spring viraemia of carp (SVC) is also causedby a fish rhabdovirus (SVCV). However, unlikeIHNVand VHSV, SVCV is related to rhabdovi-ruses in the genus Vesiculovirus in having anenveloped, bullet-shaped virion with somewhatshorter morphology and lacking the non-viriongene characteristic of novirhabdoviruses [67].Initially believed to be endemic among com-mon carp (Cyprinus carpio) in Eastern andWestern Europe, the disease appeared in thespring to cause large losses among farm-rearedcarp [2, 156]. More recently, SVCV hasemerged in several regions of the world whereit has been associated with very large lossesin common carp and its ornamental form, thekoi carp. These outbreaks have occurred in bothfarmed and wild fish, suggesting a recent rangeexpansion. The emergence of SVC in NorthAmerica, Asia and in portions of Europe, for-merly free of the virus, appears to be a resultof both improved surveillance and the globalshipment of large volumes of ornamental fish,including koi carp. Genotyping of isolates ofSVCV and a closely related fish rhabdovirus
from Europe, pike fry rhabdovirus, fromvarious locations have revealed the isolatesform four major genetic clades [127], and thatthe isolates of SVCV representing the recentemergence and geographic range expansionappear to have links to the spread of the viruswithin China where common carp are rearedin large numbers for food and koi are rearedfor export [92].
2.2.4. Infectious salmon anaemia
Infectious salmon anaemia (ISA) is anemerging disease of farmed Atlantic salmon(Salmo salar) caused by a member of the fam-ily Orthomyxoviridae. The virus (ISAV) haseight independent genome segments ofnegative-sense, single-stranded RNA pack-aged within a pleomorphic, enveloped virion,approximately 100–130 nm in diameter, andis the type species of the genus Isavirus. Ini-tially identified as the causative agent of out-breaks and high rates of mortality amongAtlantic salmon reared in sea cages in parts ofNorway [109], ISAV subsequently emerged tocause losses in other areas of Western Europewhere Atlantic salmon are farmed [95]. Thevirus was also confirmed to be the cause ofan emerging hemorrhagic kidney disease offarmed Atlantic salmon along the Atlantic coastof Canada and the USA [82]. Isolates of ISAVform two major genotypes containing isolatesfrom Europe and North America, respectively[62]. More recently, ISAV has caused veryextensive losses in the Atlantic salmon farmingindustry in Chile. Genetic analysis has revealedthat the Chilean isolates group with those fromNorway and that the virus was likely transferredto Chile sometime around 1996 by the move-ment of infected eggs [63]. Although princi-pally known as a pathogen of Atlanticsalmon, ISAV has been isolated from naturallyinfected marine species that are apparent reser-voirs for virus spill-over to susceptible Atlanticsalmon in sea cages [115]. Investigation of vir-ulence determinants of ISAV has also revealedsignificant differences among isolates [88].Thus, the emergence of ISA appears to be aresponse to the farming of a susceptible speciesin an endemic area, evolution of the virus and
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 6 of 24 (page number not for citation purpose)

some degree of transmission via the movementof fish or eggs used in aquaculture.
2.2.5. Koi herpesvirus disease
The disease caused by koi herpesvirus(KHV) is amongst the most dramatic examplesof an emerging disease of fish. KHV is a mem-ber of the genus Cyprinivirus in the familyAlloherpesviridae. Koi herpesvirus disease isrelatively host-specific; although other cyprinidspecies have been shown to be susceptible, onlycommon carp (C. carpio) and its ornamentalsubspecies, the koi carp, have been involvedin the explosive losses that have been reportedglobally in areas where the virus has been firstintroduced [47]. The enveloped virion of KHV,formally classified as the species Cyprinid her-pesvirus 3, has a morphology typical of herpe-sviruses and contains a double-stranded DNAgenome of approximately 295 kbp [5]. Molecu-lar analysis has shown little variation amongisolates, as might be expected for a virus thatis being rapidly disseminated by the globalmovement of infected fish [37]; however, minorvariation has been reported that may reflect atleast two independent introductions or emer-gence events of KHV [68]. A significant prob-lem is that once fish are infected, the viruspersists for some period of time in a latent orcarrier state without obvious clinical signs[125]. It appears that the movement of suchcarriers via the extensive trade in cultured orna-mental fish has resulted in the rapid appearanceof the disease in many regions of the world[41]. In addition, the release or stocking ofornamental fish into ponds and other naturalwaters has resulted in the introduction ofKHV to naive wild populations, where the ini-tial exposure can result in substantial mortality.
2.2.6. Epizootic haematopoietic necrosisand other ranavirus diseases
Epizootic haematopoietic necrosis is causedby a large DNA virus (EHNV) which is classi-fied in the genus Ranavirus of the family Irido-viridae [152]. Initially discovered in Australiawhere it was identified as an important causeof mortality among both cultured rainbow trout
and a native species, the redfin perch [71], itlater became clear that EHNV was but one ofa large pool of ranaviruses having a broad hostand geographic range that included amphibians,fish and reptiles [53]. Isolated from sub-clinicalinfections as well as severely diseased fish ineither aquaculture or the wild, the geneticallydiverse, but related, ranaviruses have beengiven many names in different locations [86].Although, there is evidence of spread bythe movement of infected fish, either naturallyor via trade, an important driver of the emer-gence of ranavirus diseases in finfish aquacul-ture seems to be the spill-over of virus fromendemic reservoirs among native fish, amphib-ians or reptiles [152]. In this regard, ranavirusesrelated to the type species, Frog Virus 3, havebeen shown to be an important cause of emerg-ing disease among both cultured and wildamphibian populations and may be associated,at least in some areas, with their global decline[19]. Both the large, unregulated global trade inamphibians and the unintended movement ofranaviruses by humans, including anglers andbiologists, have been postulated to be importantmethods of dissemination of these relativelystable viruses to new aquatic habitats [114].
2.2.7. Red sea bream iridoviral disease and othermegalocytivirus diseases
Another group of emerging iridovirusescauses disease in marine as well as freshwaterfish species [19]. Initially identified in 1990 asthe cause of high rate of mortality among cul-tured red sea bream (Pagrus major) in south-western Japan [54], the causative agenttermed red sea bream iridovirus (RSIV) wasshown to affect at least 31 species of marinefish cultured in the region [60, 89]. Antigenicand molecular analyses revealed the causativeagent of these outbreaks differed from otherknown fish iridoviruses [69, 103]. Soon,reports began to emerge that similar viral dis-eases in new hosts and other geographic areasof Asia were associated with novel iridovirusesincluding: infectious spleen and kidney necro-sis iridovirus (ISKNV) from cultured manda-rinfish (Sinaperca chuatsi) in southern China,sea bass (Lateolabrax sp.) iridovirus (SBIV)
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 7 of 24

from Hong Kong, rock bream (Oplegnathusfasciatus) iridovirus (RBIV) from Koreaand the orange-spotted grouper (Epinepheluscoiodes) iridovirus (OSGIV) from China[27, 45, 83, 94]. Sequence analysis showedthese and several additional, but related,viruses formed a novel group and have beenassigned to the genus Megalocytivirus in thefamily Iridoviridae, with ISKNV as the typespecies [27, 46, 69, 83]. In addition to causingoutbreaks associated with severe necrosis andhigh mortality in a wide range of cultured mar-ine fish, these viruses have emerged to affectfreshwater species such as the African lampeye(Aplocheilichthys normani) and dwarf gourami(Colisa lalia) in which they have caused addi-tional losses [132]. Molecular epidemiologicalstudies have shown that megalocytivirusesform at least three genetic lineages. There isevidence that some of the initial outbreaks inmarine species were due to spill-over fromviruses endemic among free-ranging fishes; inother cases, there are clear links to the interna-tional movement of both marine and ornamen-tal fish [132, 150, 152].
2.2.8. Viral nervous necrosis and othernodavirus diseases
Viral nervous necrosis (VNN) has emergedto become a major problem in the culture of lar-val and juvenile marine fish worldwide [101].Initially described as a cause of substantial mor-tality among cultured barramundi (Lates calca-rifer) in Australia where the disease was termedvacuolating encephalopathy and retinopathy,the condition was shown to be caused by asmall, icosahedral virus that resisted cultivationin available cell lines, but appeared similar topicornaviruses [38, 100]. Around the sametime, efforts to expand marine fish aquaculturein other regions of the world revealed diseaseconditions associated with similar viruses inlarvae or juveniles from a range of speciesincluding turbot (Scophthalmus maximus) inNorway [8], sea bass (Dicentrarchus labrax) inMartinique and the French Mediterranean [12],and parrotfish (Oplegnathus fasciatus) and red-spotted grouper (Epinephelus akaara) in Japan[97, 163]. In Japan, the disease was termed ner-
vous necrosis and the virus infecting larvalstriped jack (Pseudocaranx dentex), nowassigned as the species Striped jack nervousnecrosis virus (SJNNV), was shown to be aputative member of the family Nodaviridae[98]. Following isolation in cell culture [34],sequence analysis of the coat protein genesupported the creation of a novel genus,Betanodavirus, within the family Nodaviridaeto include isolates of fish nodaviruses from var-ious hosts and geographic locations [104, 105].These and other phylogenetic analyses [23,122, 139] revealed that genetic lineages of thebetanodaviruses show low host specificity andgenerally correspond to geographic location,indicating they emerged due to spill-over fromreservoirs that include a broad range of wildmarine fish, although some isolates revealedlinks to commercial movement.
2.3. Emerging viral diseases of shrimp
Shrimp is the largest single seafood com-modity by value, accounting for 17% of allinternationally traded fishery products. Approx-imately 75% of production is from aquaculturewhich is now almost entirely dominated by twospecies – the black tiger shrimp (Penaeus mon-odon) and the white Pacific shrimp (Penaeusvannamei) that represent the two most impor-tant invertebrate food animals [32]. Diseasehas had a major impact on the shrimp farmingindustry. Since 1981, a succession of new viralpathogens has emerged in Asia and the Ameri-cas, causing mass mortalities and threateningthe economic sustainability of the industry[148]. Shrimp are arthropods and most shrimpviruses are either related to those previouslyknown to infect insects (e.g., densoviruses, di-cistroviruses, baculoviruses, nodaviruses, lute-oviruses) or are completely new to scienceand have been assigned to new taxa (Tab. II).
Several important characteristics are com-mon to shrimp diseases and distinguish themfrom most viruses of terrestrial or aquatic verte-brates. Firstly, as invertebrates, shrimp lack thekey components of adaptive and innate immuneresponse mechanisms (i.e., antibodies, lympho-cytes, cytokines, interferon) and, althoughToll-like receptors have been identified, there
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 8 of 24 (page number not for citation purpose)
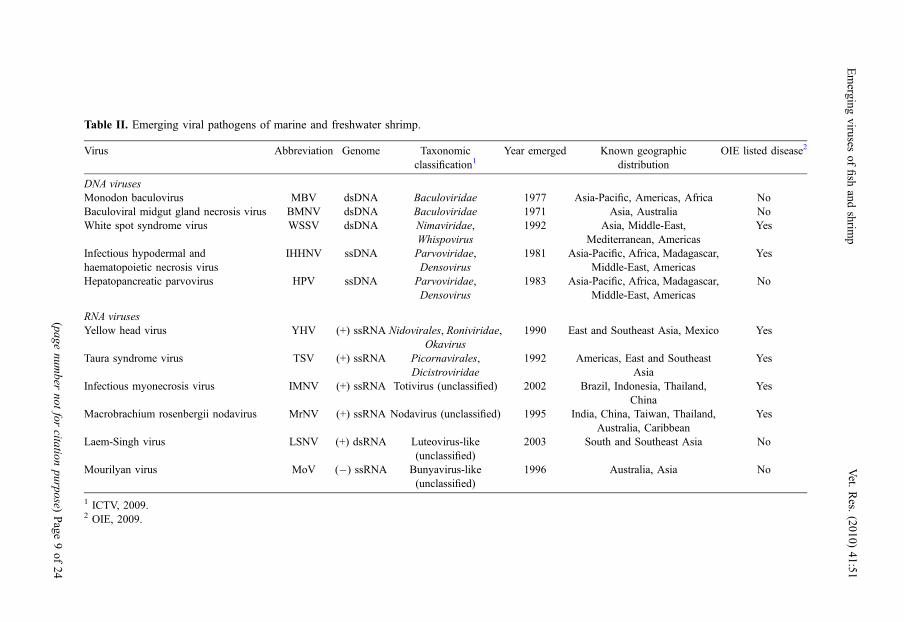
Table II. Emerging viral pathogens of marine and freshwater shrimp.
Virus Abbreviation Genome Taxonomicclassification1
Year emerged Known geographicdistribution
OIE listed disease2
DNA virusesMonodon baculovirus MBV dsDNA Baculoviridae 1977 Asia-Pacific, Americas, Africa NoBaculoviral midgut gland necrosis virus BMNV dsDNA Baculoviridae 1971 Asia, Australia NoWhite spot syndrome virus WSSV dsDNA Nimaviridae,
Whispovirus1992 Asia, Middle-East,
Mediterranean, AmericasYes
Infectious hypodermal andhaematopoietic necrosis virus
IHHNV ssDNA Parvoviridae,Densovirus
1981 Asia-Pacific, Africa, Madagascar,Middle-East, Americas
Yes
Hepatopancreatic parvovirus HPV ssDNA Parvoviridae,Densovirus
1983 Asia-Pacific, Africa, Madagascar,Middle-East, Americas
No
RNA virusesYellow head virus YHV (+) ssRNA Nidovirales, Roniviridae,
Okavirus1990 East and Southeast Asia, Mexico Yes
Taura syndrome virus TSV (+) ssRNA Picornavirales,Dicistroviridae
1992 Americas, East and SoutheastAsia
Yes
Infectious myonecrosis virus IMNV (+) ssRNA Totivirus (unclassified) 2002 Brazil, Indonesia, Thailand,China
Yes
Macrobrachium rosenbergii nodavirus MrNV (+) ssRNA Nodavirus (unclassified) 1995 India, China, Taiwan, Thailand,Australia, Caribbean
Yes
Laem-Singh virus LSNV (+) dsRNA Luteovirus-like(unclassified)
2003 South and Southeast Asia No
Mourilyan virus MoV (�) ssRNA Bunyavirus-like(unclassified)
1996 Australia, Asia No
1 ICTV, 2009.2 OIE, 2009.
Emerging
virusesof
fish
andshrim
pVet.
Res.
(2010)41:51
(pagenum
bernot
forcitation
purpose)Page
9of
24

is little evidence that they are involved in anti-viral immunity [70, 160]. RNA interference(RNAi) does appear to have a role in the anti-viral defensive response of shrimp and there isevidence that viral proteins can induce a short-lived protective immunity [57, 118]. However,as is the case for insects, the response of shrimpto viral infection is poorly understood and thesubject of intensive research. The second distin-guishing feature is that most of the major patho-genic viruses cause very low level persistentinfections that can occur at moderate to veryhigh prevalence in apparently health shrimppopulations [16, 51, 108, 133, 145]. Almostall shrimp pathogens are transmitted vertically(but usually not transovarially) and disease isthe result of a massive viral amplification thatfollows exposure to various forms of environ-ment or physiological stress [22, 81, 113, 119].Stressors can include handling, spawning, poorwater quality or abrupt changes in temperatureor salinity. Shrimp viruses can also commonlybe transmitted horizontally and, once viral loadsare high and disease is manifest, horizontaltransmission of infection is accompanied bytransmission of disease. The third significantcharacteristic is a logical consequence of the for-mer two in that shrimp commonly can beinfected simultaneously or sequentially withmultiple viruses [33], or even different strainsof the same virus [50]. These characteristicspresent a very different landscape for the inter-action of pathogen and host and significant chal-lenges for diagnosis, detection, pathogenexclusion and the use of prophylactics in healthmanagement. Viruses listed by the OIE as caus-ing notifiable diseases of marine and freshwatershrimp are reviewed briefly in this section.
2.3.1. White spot syndrome
White spot syndrome first emerged in FujianProvince of China in 1992 [165]. It was soonafter reported in Taiwan and Japan and hassince become panzootic throughout shrimpfarming regions of Asia and the Americas[148]. It is the most devastating disease offarmed shrimp with social and economicimpacts over 15 years on a scale that is seldomseen, even for the most important diseases of
terrestrial animals. White spot syndrome virus(WSSV) is a large, enveloped, ovaloid DNAvirus with a flagellum-like tail and helical nucle-ocapsid that has been classified as the onlymem-ber of the new family Nimaviridae, genusWhispovirus [144, 158]. The � 300 kbp viralgenomecontains at least 181ORF,most ofwhichencode polypeptides with no detectable homol-ogy to other known proteins [142, 159].Although first emerging in farmed kurumashrimp (Penaeus japonicus), WSSV has a verybroad host range amongst decapod crustaceans(e.g., marine and freshwater shrimp, crabs, lob-sters, crayfish, etc.), all of which appear to besusceptible to infection [72]. However, suscepti-bility to disease varies and some crustaceanspecies have been reported to develop very highviral loads in the absence of clinical signs[162]. All farmed marine (penaeid) shrimpspecies are highly susceptible to white spotdisease, with mass mortalities commonly reach-ing 80–100% in ponds within a period of 3–10days [20, 77]. Persistent, low level infections inshrimp and other crustaceans occur commonly,sometimes at levels that are not detectable, evenby nested PCR. The amplification of viral loadsand onset of disease can be induced by environ-mental or physiological stress [80, 113], or atambient temperatures below 30 �C [39, 143].
WSSV was not known prior to its emer-gence in China and the original source of infec-tion has not been determined. However, thespread of infection throughout most of Asiaduring the mid-1990s and subsequently to theAmericas from 2001 was explosive and wasalmost certainly the consequence of a prolificinternational trade in live shrimp and other crus-tacean seed and broodstock [76]. The suscepti-bility of all decapods and absence of evidenceof replication in other organisms suggests thevirus is of crustacean origin but it remains amystery why a virus with such broad host rangeand ease of transmission was not long estab-lished globally in crustacean populations priorto the advent of aquaculture [148].
2.3.2. Taura syndrome
Taura syndrome first emerged in whitePacific shrimp (P. vannamei) farms on the
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 10 of 24 (page number not for citation purpose)

Taura River near Guayaquil in Ecuador in1992, almost simultaneously with the emer-gence of WSSV in kuruma shrimp in China[43]. The disease spread rapidly throughoutmost shrimp farming regions of Central andSouth America [75]. In 1998, it was detectedin Taiwan and has now spread throughoutmuch of Asia [140]. Taura syndrome virus(TSV) is a small, naked (+) ssRNA virus thatis currently classified as an unassigned speciesin the family Dicistroviridae, order Picornavi-rales [21]. The most closely related knownviruses include insect viruses in the genusCripavirus such as cricket paralysis virus anddrosophila C virus [21, 87]. Acute, transitional(recovery) and chronic phases of TSV infectionhave been described [44]. Mortalities in theacute phase can be as high as 95% but surviv-ing shrimp remain infected and a potentialsource of virus transmission. The susceptiblehost range of TSV is far more restricted thanthat of WSSV but includes most farmed marineshrimp species. However, susceptibility to dis-ease varies and virulence varies for differentstrains of the virus. Other crustaceans includingfreshwater shrimp and crabs appear to be resis-tant to disease but may be potential carriers[61]. Birds and water-boatmen (Trichocorixareticulata) have been proposed as possiblemechanical vectors [14, 36, 126]. The focal ori-gin of the TSV panzootic and absence of evi-dence of infection prior to the first outbreaksuggest that, as for WSSV, penaeid shrimpare not the natural host. The rapid spread ofTSV in the Americas and then to Asia has alsobeen attributed to the international trade in liveshrimp.
2.3.3. Yellow head disease
Yellow head virus (YHV) is the most viru-lent of shrimp pathogens, commonly causingtotal crop loss within several days of the firstsigns of disease in a pond. It first emerged inblack tiger shrimp (P. monodon) in CentralThailand in 1990 and has since been reportedin most major shrimp farming countries inAsia, including India, Indonesia, Malaysia, thePhilippines, Sri Lanka, Vietnam and Taiwan[17, 145]. There is also a recent unconfirmed
report that YHV is present in farmed P. vanna-mei and P. stylirostris in Mexico [25]. YHV isan enveloped, rod-shaped (+) ssRNA viruswith a helical nucleocapsid and prominent gly-coprotein projections on the virion surface[157]. The particle morphology and organisa-tion and expression strategy of the � 26 kbgenome indicate that it is most closely relatedto vertebrate coronaviruses, toroviruses andarteriviruses, and it has been classified withinthe order Nidovirales in the family Ronivirus,genus Okavirus [147]. It is now known thatYHV is one of a complex of six closelyrelated viruses infecting P. monodon shrimp[153]. Gill-associated virus (GAV) is a far lessvirulent virus that emerged to cause mid-cropmortality syndrome in farmed P. monodon inAustralia in 1996 [124]. However, the preva-lence of GAV infection in healthy P. monodonbroodstock and farmed shrimp in Australia hasbeen reported to approach 100% and, althoughdisease can be transmitted by injection orexposure per os to moribund shrimp, out-breaks in ponds are most likely the result ofamplification of viral loads as a consequenceof environmental stress. The other four knowngenotypes in the complex have been detectedonly in healthy P. monodon in Asia and,although they occur at high prevalence inmany locations, they are not known to beassociated with disease [153]. Many other pen-aeid and palemonid shrimp species have beenshown to be susceptible to experimental infec-tion with YHV or GAV, but yellow-head-complex viruses are detected rarely in otherpenaeid shrimp species and P. monodonappears to be the natural host [148]. Neverthe-less, the very high virulence of YHV for pen-aeid shrimp does suggest that this genotypemay enter ponds via an alternative reservoirhost. Homologous genetic recombination isalso a feature of the yellow head complex.A recent study has indicated that � 30% ofyellow-head-complex viruses detected inP. monodon from across the Asia-Pacificregion are recombinants [154]. The prevalenceand geographic distribution of these recombi-nant viruses suggests that aquaculture and theinternational trade in live shrimp are the sourceof rapidly increasing viral genetic diversity.
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 11 of 24

2.3.4. Infectious hypodermal and haematopoieticnecrosis
Infectious hypodermal and haematopoieticnecrosis was first detected in Hawaii in 1981,causing mass mortalities in blue shrimp(Penaeus stylirostris) farmed in super-intensiveraceways [74]. Infectious hypodermal and hae-matopoietic necrosis virus (IHHNV) is a small,naked, ssDNA virus that has been classifiedwith several insect viruses in the family Parvo-viridae, genus Brevidensovirus [21]. Followingits initial detection in Hawaii, IHHNV wasfound to be widely distributed in both P. styliros-tris and P. vannamei shrimp throughout farmingregions of the Americas and in the wild shrimppopulation of the Gulf of California wheresome reports suggest that it may have contrib-uted to the collapse of the capture fishery[96, 111]. Although it does not cause mortalitiesin P. vannamei, IHHNV has been shown toreduce growth by up to 30% and cause deformi-ties of the rostrum and anterior appendages in acondition called ‘‘runt deformity syndrome’’[59]. In Asia, IHHNV is endemic and occurscommonly in P. monodon shrimp which appearsto be the natural host and in which it does notcause disease and has no impact on growth orfecundity [18, 155]. Four genotypes of IHHNVhave been identified of which two have beenshown to be integrated into host genomicDNA and experimental transmission studiessuggest they may not be infectious for P. mon-odon or P. vannamei shrimp [65, 135, 136].The other two genotypes can be transmitted hor-izontally by injection, ingestion or exposure toinfected water, or vertically from infectedfemales [73]. Genetic evidence suggests thatP. monodon imported from the Philippines werethe source of the epizootic in the Americas, indi-cating that disease emergence has been the con-sequence of an expanded host range providingopportunities for pathogenicity and a vastlyexpanded geographic distribution [134].
2.3.5. Infectious myonecrosis
Infectious myonecrosis is the most recentlyemerging of the major viral diseases of marineshrimp. It first appeared in farmed P. vannamei
shrimp at Pernambuco in Brazil in 2002 andhas subsequently spread throughout coastalregions of north-east Brazil and to Indonesia,Thailand and Hainan Province in China[4, 79, 120]. The original source of infectionis unknown but the trans-continental spreadhas almost certainly been due to the volumi-nous trade in P. vannamei broodstock. Shrimpwith the acute form of the disease display vari-ous degrees of skeletal muscle necrosis, visibleas an opaque, whitish discolouration of theabdomen [137]. Surviving shrimp progress toa chronic phase with persistent low-level mor-talities. Infectious myonecrosis virus (IMNV)is a small, naked, icosahedral, dsRNAvirus thatis most closely related to members of the familyTotiviridae, genus Giardiavirus [116]. The onlyother known members of this family infectyeasts and protozoa. Several farmed marineshrimp species have been reported to besusceptible to infection but disease has onlybeen reported in white Pacific shrimp [137].The increasingly common practice in parts ofAsia of co-cultivation of white Pacific shrimpand black tiger shrimp is likely to presentopportunities for adaptation and further spreadof the disease.
2.3.6. White tail disease
White tail disease is an emerging infectionof the giant freshwater shrimp Macrobrachiumrosenbergii. It was first reported in 1995 fromthe island of Guadeloupe and then nearbyMartinique in the French West Indies, and hassince been reported from China, Taiwan,Thailand, India and Australia [6, 110, 117,141, 161]. The disease can affect larvae, post-larvae and early juvenile stages, causing up to100% mortalities within 5–7 days of the firstgross signs which include a white or milkyappearance of abdominal muscle [42, 117].Adults are resistant to the disease but can bepersistently infected and transmit the infectionvertically. Marine shrimp (Penaeus monodon,P. japonicus and P. indicus) have been shownto be susceptible to infection but did notdevelop disease, and artemia and some spe-cies of aquatic insects appear to be vectors[130, 131]. White tail disease is caused by a
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 12 of 24 (page number not for citation purpose)

small, naked (+) ssRNA virus that has beennamed Macrobrachium rosenbergii nodavirus(MrNV). Sequence alignments indicate that itis related to but distinct from nodaviruses ofinsects (genus Alphanodavirus) and nodavirus-es of fish (genus Betanodavirus) [9]. A verysmall satellite virus (extra small virus, XSV)appears to be universally associated with natu-ral MrNV infections but is not the direct causeof disease [117]. As the native endemic range ofMacrobrachium rosenbergii is restricted tosouth and south-east Asia, the wide geographicdistribution of the disease most likely has beendue to the movement of stock for aquaculturepurposes. Nevertheless, the detection in 2007of a distinct strain of MrNV in Macrobrachiumrosenbergii broodstock captured from theremote Flinders River in western Queensland,Australia [110], where there is a long-standingenforced prohibition on the importation of livecrustaceans, suggests that the virus is a naturalinfection of freshwater shrimp. Penaeus vanna-mei nodavirus (PvNV) is a distinct but relatedvirus that was detected in 2004 in cultured mar-ine shrimp in Belize displaying the gross signsof white tail disease [138].
3. IMPACTS OF EMERGING DISEASESOF FISH AND SHRIMP
3.1. Economic and social impacts
The impacts of emerging diseases of aquaticanimals have been substantial; all have affectedlivelihoods locally and many have impacted onregional or national economies. The most dev-astating economic and social impacts have beenin shrimp aquaculture for which it was esti-mated in 1996 that the global direct and indirectcosts of emerging diseases had reached $US 3billion annually or 40% of the total productioncapacity of the industry [55, 85]. The most sig-nificant production losses have immediately fol-lowed the emergence of each of the majorpathogens, with ensuing periods of poor pro-ductivity and reduced rates of industry expan-sion during which pathogens have beenidentified and characterised, diagnosis anddetection methods developed, and improved
biosecurity measures implemented [10, 148].In many cases, impacts have continued formany years, particularly for small low-incomefarmers in developing countries who lack theknowledge, skill and resources to respondeffectively. WSSV has been by far the mostdevastating of the shrimp pathogens. It has beenestimated that the impact of WSSV in Asiaalone during the 10 years after its emergencein 1992 was $US 4–6 billion [78]. In the Amer-icas, the emergence of WSSV in 1999–2000resulted in immediate losses estimated at $US1 billion. The combined impacts of TSV andIHHNVon aquaculture and wild shrimp fisher-ies in the Americas have been estimated at $US1.5–3 billion [44, 78]. The consequences of dis-ease emergence for some countries have beenso severe that shrimp production has never fullyrecovered. Beyond the direct effects on produc-tion and profitability, disease impacts on theincome and food-security of small-holdershrimp farmers and the job security of workerson larger farms and in feed mills and processingplants, with a flow-on effect to the sustaininglocal communities [148].
The global economic losses in fish aquacul-ture due to infectious diseases are of a lessermagnitude, but still highly important in severalways and can be crippling for farmers. Not onlyare many individual animals of greater commer-cial value (e.g., koi carp) but the disruption ofconsistent production schedules by companiesengaged in intensive aquaculture can result inloss of market share. The emergence of ISAVin Scotland in 1998–1999 is estimated to havecost the industry � $US 35 million andresulted in an ongoing annual loss of $US 25million to the industries in Norway and Canada.The estimated cost of the emergence of KHV inIndonesia was in excess of $US 15 million dur-ing the first 3 years [10], with ongoing socio-economic impacts on low-income, small-holderfarmers.
3.2. Environmental impacts
Environmental impacts of emerging diseasesof aquatic animals have been both direct andindirect. Disease can impact directly on wildpopulations and the ecosystem by changing
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 13 of 24

host abundance and predator/prey populations,reducing genetic diversity and causing localextinctions [7]. The emergence of pilchard her-pesvirus in Australasia during the 1990s wasone of the most dramatic examples of large-scale impact on ecosystems. Commencing inMarch 1995, mass mortalities occurred in wildpilchard (Sardinops sagax neopilchardus) pop-ulations across 5 000 km of the Australiancoastline and 500 km of the coastline ofNew Zealand [58]. The epizootic subsided inSeptember 1995 but a second more severe waveof mortalities occurred between October 1998and April 1999 with vast numbers of pilchardswashed onto southern Australian beaches andmortality rates in pilchard populations as highas 75% [151]. The causative agent was identi-fied as a previously unknown fish herpesvirus,but the source of the epidemic has never beenidentified [52]. Beyond the direct impact on pil-chard populations, wider secondary impacts onpiscivorous species were observed. Penguinssuffered increased mortalities and failed tobreed due to food shortage [24]. The contribu-tion of pilchards to the diet of the Australiangannet (Morus serrator) declined from 60%to 5% following the mortality event and wascompensated by feeding on species with lowercalorific value [15]. As pilchards are importantprey for seabirds, fish and marine mammals,other secondary impacts were likely. The recentintroduction of VHSV to the Great Lakes Basinhas also resulted in large-scale mortalities andspread to at least 25 native freshwater fish spe-cies with potential for similar broader environ-mental impacts [1].
Despite their panzoootic distribution in aqua-culture systems and wide crustacean host range,direct environmental impacts of shrimp viruseshave not been commonly observed. It has beenreported that IHHNV impacted on wildP. stylirostris fisheries in the Gulf of Mexico fol-lowing its introduction in 1987 [96] but rigorousenvironmental assessments of the impact of thisor other shrimp pathogens have not been con-ducted. Indirect environmental impacts of dis-ease in shrimp aquaculture are more clearlyevident. These include the destruction of man-grove habitats due to pond abandonment andrelocation to new sites, soil salinisation in inland
areas due to avoidance of disease-prone coastalzones, and use of antibiotics, disinfectants andother chemicals to prevent or treat disease inponds [3, 93, 148]. Difficulties in managing dis-ease in native Asian marine shrimp species havealso led to an extraordinary shift in productionsince 2001 to imported P. vannamei for whichSPF stock are readily available [13]. The naturalhabitat of this species is the west coast of CentralAmerica but it now accounts for 67.1% of totalfarmed shrimp production in Asia [32], repre-senting a massive species translocation forwhich the impacts on local biodiversity remainuncertain.
4. FACTORS CONTRIBUTING TO DISEASEEMERGENCE IN AQUATIC ANIMALS
The increasing rate of emergence of diseasesof fish and shrimp has been driven primarily byanthropogenic influences, the most profound ofwhich have been associated with the globalexpansion of aquaculture. Farming of aquaticanimals commonly involves displacement fromtheir natural habitat to an environment that isnew and sometimes stressful, the use of feedsthat are sometimes live and often unnatural orartificial, and culture in stocking densities thatare much higher than occur naturally. This hasprovided opportunities for exposure to newpathogens and conditions that can compromisedefensive responses and facilitate pathogen rep-lication and disease transmission [146, 148].Most importantly, the growth in aquacultureand increasing international trade in seafoodhas resulted in the rapid movement of aquaticanimals and their products, with associated risksof the trans-boundary movement of pathogens[129]. Other diseases have been spread by nat-ural or unintentional movement of infectedhosts or amplified by invasive species, whileanthropogenic environmental pressures havecaused changes in the severity of several ende-mic diseases. Better surveillance activitiesbased upon novel and more sensitive toolsand their application in new species and geo-graphic areas have also contributed to an appar-ent expansion of host or geographic range,sometimes dramatically.
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 14 of 24 (page number not for citation purpose)

4.1. Activities related to the global expansionof aquaculture
Emerging viral diseases of fish or shrimpare usually caused either by (i) viruses that nat-urally infect the target species but are unob-served or not normally pathogenic in wild orunstressed populations, or (ii) the spill-over ofviruses from other species that may not beencountered naturally. Virus infections occurcommonly in apparently healthy populationsof wild fish and shrimp and, although diseaseoutbreaks may occasionally occur, these oftenpass undetected and are not essential to sustainthe natural cycle of transmission [149]. Inshrimp, IHHNV, yellow-head-complex virusesand possibly MrNVappear to be naturally ende-mic in healthy wild populations and haveemergedas significantpathogensonly asa conse-quence of aquaculture practices. In the case ofIHHNV, disease emergence has been due to thetranslocation of the natural host, P. monodon,from the Philippines to Hawaii and theAmericasfor use in aquaculture breeding programs,allowing spill-over into susceptible westernhemisphere shrimp species [134]. For virusesin the yellow head complex, the natural preva-lence can approach 100% in some healthy wildP. monodon populations but stressful cultureconditions, in combination with yet uncharac-terised virulence determinants, appear to triggerdisease outbreaks [26, 145, 153]. Similarly, theemerging fish pathogen ISAV is endemic at rel-atively high prevalence in wild salmonid popu-lations in Norway and Canada but has beenassociated with disease only in farmed Atlanticsalmon. It has been suggested that intensive cul-ture serves to provide concentrations of suscep-tible animals in which virulent strains of ISAVemerge due to a combination of high mutationrate and increased opportunity for virus replica-tion [63]. Other major emerging pathogens areclearly not naturally endemic in aquaculturespecies but have spilled-over into both farmedand wild populations as a consequence of theexposure opportunities provided by the rapidgrowth of a large and diverse industry. WSSV,TSV and IMNV each appear to have beeninitially introduced to shrimp populations fromunidentified sources that could potentially
include experimental live or frozen feeds orco-inhabitants of terrestrial pond environmentssuch as insects or aquatic invertebrates. Never-theless, each of these viruses now occurs com-monly as low-level persistent infections inhealthy shrimp populations and disease out-breaks are precipitated by environmental stres-ses associated with aquaculture. In the case ofWSSV, aquaculture has also provided opportu-nities for spread of infection to a very widerange of new wild crustacean hosts in whichthe virus has now become endemic across avast coastal area of Asia and the Americas.
Amongst finfish pathogens, the ranavirusesand nodaviruses provide good examples ofthe emergence of disease due to spill-overfrom wild reservoirs. Prior to the availabilityof large populations of susceptible speciesreared in aquaculture, the betanodavi-ruses were endemic but undetected amongfree-ranging populations of marine fish andvarious ranaviruses were present in a broadrange of native species that included fish,amphibians and reptiles. The introduction ofthese viruses to commercial aquaculture andthe ensuing emergence of disease were likelydue to the use of open water supplies andwild-caught broodstock.
The intentional or unintentional movementof infected hosts or pathogens by individualsor companies involved in global aquacultureor the ornamental fish trade has also been animportant driver of viral disease emergence inaquatic animals [150]. Exotic viruses translo-cated into new geographic areas have spreadto cause major disease outbreaks in populationsof both native and cultured species and thetrans-boundary movement of aquatic animalscontinues to be one of the greatest threats tothe productivity and profitability of aquacultureworldwide. For finfish, this is exemplified bythe spread of IHNV to Europe and Asia viathe shipment of contaminated eggs of troutand salmon, the global spread of SVCV andKHV via the ornamental fish trade and theintroduction of the pilchard herpesvirus intonative populations of pilchards in Australiavia baitfish. For shrimp, the prolific interna-tional trade in live broodstock has been themajor driver of the explosive trans-boundary
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 15 of 24

spread of emerging viral pathogens. The rapidspread of WSSV throughout Asia and then tothe Americas has been attributed to the move-ment of live crustaceans [78]. The magnitudeof this problem is most clearly exemplified bythe mass translocation of many hundreds ofthousands of white Pacific shrimp broodstockfrom the Americas to Asia, accompanied bythe introduction of TSV, IMNV and possiblyother exotic pathogens [13]. The internationaltrade in frozen commodity shrimp and shrimpproducts has also been recognised as a potentialmechanism of trans-boundary spread of disease[76]. Several reports have indicated the pres-ence of infectious WSSV and YHV in frozencommodity shrimp imported to the USA andAustralia [90, 107]. However, it is yet to beestablished that the trade in infected frozenproduct has contributed to disease outbreaksin wild or cultivated shrimp [33b].
4.2. Improved surveillance
Apparent changes in the prevalence and dis-tribution of viral diseases of fish and shrimp canalso be attributed to improved surveillance as aresult of the availability of new cell lines forvirus isolation and characterization, the devel-opment of more sensitive molecular diagnosticmethods, enhanced testing activity in naturalecosystems and greatly improved aquatic ani-mal health diagnostic capability in some devel-oping countries. For some viral pathogens, theavailability of improved detection and surveil-lance capabilities has resulted in reports of agreater host or geographic range than previ-ously known; in other cases, diseases in differ-ent species or geographic regions have beenshown to be due to the same or similar viruses.For fish diseases, this is exemplified by devel-opment of the salmon head kidney (SHK) cellline to detect and characterize ISAV in Norwaywhich subsequently led to the identification ofpotential carriers and the observation thatemerging diseases of Atlantic salmon in Canadaand Chile were due to isolates of the same virus[82]. Enhanced surveillance activities alsoresulted in the detection and isolation of VHSVfrom normal returning adult salmon on the westcoast of North America in 1988. While initially
appearing as a large geographic range expan-sion, increased surveillance activities showedthat VHSV is naturally endemic in the NorthPacific and North Atlantic Oceans among awide range of marine species [121]. Similarly,investigations in Australia have identified thatseveral viruses that were previously assumedto be exotic, occur commonly in healthy shrimppopulations [110]. The increasing number ofpreviously unknown viruses that have beenidentified through enhanced surveillance ofhealthy wild fish and shrimp populations illus-trates how little is currently known of the aqua-tic virosphere.
4.3. Natural movement of carriers
The emergence of some viral diseases of fishand shrimp has occurred through expansion ofthe known geographic range via the naturalmovement of infected hosts, vectors or carrierswith subsequent exposure of naive, and oftenhighly susceptible, species. While the mecha-nism is not known with certainty, it is consid-ered plausible that VHSV was introduced tothe Great Lakes Basin of North America viathe natural movement of fish up the SaintLawrence River from the near shore areas oftheAtlantic coast ofCanada. The initial introduc-tion of the virus into the Great Lakes precededthe emergence of massive epidemics amongseveral species by at least two years. This hasgiven rise to speculation that a single introduc-tion of VHSV resulted in infection of the mostsusceptible species such as the native muskel-lunge (Esox masquinongy) or the invasive roundgobi (Neogobius melanostomus) which couldamplify the virus sufficiently to increase theinfection pressure on a broad range of naivespecies within the larger ecosystem [40].
4.4. Other anthropogenic factors
The emergence of viral disease in aquaticsystems can also been driven by anthropogenicfactors unrelated to aquaculture such as themovement of pathogens or hosts via ballastwater in ships, movement of bait by anglersand unintentional movement in other biotic orabiotic vectors. The environmental impacts of
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 16 of 24 (page number not for citation purpose)

increasing global population such as increasingloads of pollutants, contaminants and toxins inaquatic habitats also threaten the health anddisease-resistance of both native and farmedfish populations. Finally, water diversions,impoundments, cooling water inputs and otherwater-use conflicts can result in environmentalchanges that may be detrimental to the healthof aquatic species.
5. FUTURE DISEASE EMERGENCE RISKS
The increasing global population, increasingdemand for seafood and limitations on produc-tion from capture fisheries will inevitably leadthe continued global expansion of aquaculturewith associated risks of disease emergence andspread. Africa, in particular, which currentlycontributes less than 1.3% of global aquacultureproduction [32], offers significant developmentopportunities for aquaculture but also presentsa rich biodiversity that may be the source ofemergence of new aquatic animal pathogens.There are also disease emergence risks associ-ated with the increasing diversity of farmed spe-cies, the introduction of species into newfarming areas and the increasing trend amongstsmall-holder farmers in some countries towardspolyculture or alternate cropping of several fishand crustacean species to improve productivity.
As is the case for terrestrial pathogens, cli-mate change also looms as a likely future driverof disease emergence in aquatic animals. Cli-mate change and variability are likely to bringecological disturbance on a global scale thatwill influence the suitability of aquaculturefarming sites and has potential to influencethe growth rate of pathogens and the hostimmune response to infection, generate thermalstress that will increase susceptibility to disease,change the distribution of vectors, carriers andreservoirs and the density or distribution of sus-ceptible wild species, and alter the physical hab-itat and farming practices in ways that affectdisease ecology. Nevertheless, there is a grow-ing awareness of the importance of emergingdiseases of aquatic animals and it is likely thatthe risks of future disease emergence will bemitigated somewhat by the development of
improved diagnostic methods and surveillanceefforts, increased regulatory oversight of aqua-culture with greater levels of health inspectionfor fish, shrimp and their products involved ininternational trade, and the development ofnovel vaccines and therapeutics.
REFERENCES
[1] Abraham D., Greig L., Viral haemorrhagic septi-caemia in the Great Lakes region: management andscience needs workshop proceedings. Prepared for theInternational Joint Commission, the Ontario Ministryof Natural Resources and Great Lakes Fishery Com-mission, ESSATechnologies, Richmond Hill, Ontario,2008, p. 11.
[2] Ahne W., Bjorklund H.V., Essbauer S., Fijan N.,Kurath G., Winton J.R., Spring viremia of carp (SVC),Dis. Aquat. Org. (2002) 52:261–272.
[3] Ali A.M.S., Rice to shrimp: land use land coverchanges and soil degradation in SouthwesternBangladesh, Land Use Policy (2006) 23:421–435.
[4] Andrade T.P.D., Redman R.M., Lightner D.V.,Evaluation of the preservation of shrimp samples withDavidson’s AFA fixative for infectious myonecrosisvirus (IMNV) in situ hybridization, Aquaculture(2008) 278:179–183.
[5] Aoki T., Hirono I., Kurokawa K., Fukuda H., NaharyR., Eldar A., et al., Genome sequences of three koi her-pesvirus isolates representing the expanding distributionof an emerging disease threatening koi and commoncarp worldwide, J. Virol. (2007) 81:5058–5065.
[6] Arcier J.M., Herman F., Lightner D.V., RedmanR.M., Mari J., Bonami J.R., A viral disease associatedwith mortalities in hatchery-reared postlarvae of thegiant freshwater prawn Macrobrachium rosenbergii,Dis. Aquat. Org. (1999) 38:177–181.
[7] Arthur J.R., Subasinghe R.P., Potential adversesocio-economic and biological impacts of aquaticanimal pathogens due to hatchery-based enhancementof inland open-water systems, and possibilities fortheir minimisation, in: Arthur J.R., Phillips M.J.,Subasinghe R.P., Reantaso M.B., MacRae I.H. (Eds.),Primary Aquatic Animal Health Care in Rural, Small-scale, Aquaculture Development FAO Fisheries,Technical Paper, No. 406, 2002, pp. 113–126.
[8] Bloch B., Gravningen K., Larsen J.L., Encepha-lomyelitis among turbot associated with a picorna-likeagent, Dis. Aquat. Org. (1991) 10:65–70.
[9] Bonami J.R., Shi Z., Qian D., Sri Widada J.,White tail disease of the giant freshwater prawn, Mac-robrachium rosenbergii: separation of the associated
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 17 of 24

virions and characterization of MrNV as a new type ofnodavirus, J. Fish Dis. (2005) 28:23–31.
[10] Bondad-Reantaso M.G., Subasinghe R.P., ArthurJ.R., Ogawa K., Chinabut S., Adlard R., et al., Diseaseand health management in Asian aquaculture, Vet.Parasitol. (2005) 132:249–272.
[11] Bootland L.M., Leong J.C., Infectious hemato-poietic necrosis virus, in: Woo P.T.K., Bruno D.W.(Eds.), Fish diseases and disorders, Volume 3, Viral,bacterial and fungal infections, CAB International,Oxon, UK, 1999, pp. 57–121.
[12] Breuil G., Bonami J.R., Pepin J.F., Pichot Y.,Viral infection (picorna-like virus) associated withmass mortalities in hatchery-reared sea bass (Dicen-trarchus labrax) larvae and juveniles, Aquaculture(1991) 97:109–116.
[13] Briggs M., Funge-Smith S., Subasinghe R.,Phillips M., Introductions and movement of Penaeusvannamei and Penaeus stylirostris in Asia and thePacific, RAP publication 2004/10, FAO RegionalOffice for Asia and the Pacific, Bangkok, 2004.
[14] Brock J.A., Gose R., Lightner D.V., Hasson K.,An overview on Taura syndrome, an important diseaseof farmed Penaeus vannamei, Swimming throughtroubled water: Proceedings of the Special Session onShrimp Farming, Aquaculture ’95, World AquacultureSociety, Baton Rouge, 1995, pp. 84–94.
[15] Bunce A., Norman F.I., Changes in the diet ofthe Australasian gannet (Morus serrator) in responseto the 1998 mortality of pilchards (Sardinops sagax),Mar. Freshw. Res. (2000) 51:349–353.
[16] Chakraborty A., Otta S.K., Kumar B.J.S.,Hossain M.S., Karunasagar I., Venugopal M.N.,Karunasagar I., Prevalence of white spot syndromevirus in wild crustaceans along the coast of India, Curr.Sci. (2002) 82:1392–1397.
[17] Chantanachookin C., Boonyaratpalin S.,Kasornchandra J., Direkbusarakom S., EkpanithanpongU., Supamataya K., et al., Histology and ultrastructurereveal a new granulosis-like virus in Penaeus monodonaffected by yellow-head disease, Dis. Aquat. Org.(1993) 17:145–157.
[18] Chayaburakul K., Lightner D.V., SriurairattanaS., Nelson K.T., Withyachumnarnkul B., Differentresponses to infectious hypodermal and hematopoieticnecrosis virus (IHHNV) in Penaeus monodon andP. vannamei, Dis. Aquat. Org. (2005) 67:191–200.
[19] Chinchar V.G., Hyatt A., Miyazaki T., WilliamsT., Family Iridoviridae: poor viral relations no longer,Curr. Top. Microbiol. Immunol. (2009) 328:123–170.
[20] Chou H.Y., Huang C.Y., Wang C.H., ChiangH.C., Lo C.F., Pathogenicity of a baculovirus infectioncausing white spot syndrome in cultured penaeid
shrimp in Taiwan, Dis. Aquat. Org. (1995) 23:165–173.
[21] Christian P., Carstens E., Domier L., Johnson J.,Johnson K., Nakashima N., et al., Family Dicistrovi-ridae, in: Fauquet C.M., Mayo M.A., Maniloff J.,Desselberger U., Ball L.A. (Eds.), Virus Taxonomy,VIIIth Report of the International Committee onTaxonomy of Viruses, Elsevier, Academic Press,Amsterdam, 2005, pp. 783–788.
[22] Cowley J.A., Hall M.R., Cadogan L.C., SpannK.M., Walker P.J., Vertical transmission of gill-asso-ciated virus (GAV) in the black tiger prawn Penaeusmonodon, Dis. Aquat. Org. (2002) 50:95–104.
[23] Dalla Valle L., Negrisolo E., Patarnello P.,Zanella L., Maltese C., Bovo G., Colombo L.,Sequence comparison and phylogenetic analysis offish nodaviruses based on the coat protein gene, Arch.Virol. (2001) 146:1125–1137.
[24] Dann P., Norman F.I., Cullen J.M., Neira F.J.,Chiaradia A., Mortality and breeding failure of littlepenguins, Eudyptula minor, in Victoria, 1995–96,following a widespread mortality of pilchard, Sardin-ops sagax, Mar. Freshw. Res. (2000) 51:355–362.
[25] De la Rosa-Velez J., Cedano-Thomas Y.,Cid-Becerra J., Mendez-Payan J.C., Vega-Perez C.,Zambrano-Garcia J., Bonami J.R., Presumptive detec-tion of yellow head virus by reverse transcriptase-polymerase chain reaction and dot-blot hybridizationin Litopenaeus vannamei and L. stylirostris culturedon the Northwest coast of Mexico, J. Fish Dis. (2006)29:717–726.
[26] De la Vega E., Degnan B.M., Hall M.R., CowleyJ.A., Wilson K.J., Quantitative real-time RT-PCR demon-strates that handling stress can lead to rapid increases ofgill-associated virus (GAV) infection levels in Penaeusmonodon, Dis. Aquat. Org. (2004) 59:195–203.
[27] Do J.W., Moon C.H., Kim H.J., Ko M.S., KimS.B., Son J.H., et al., Complete genomic DNAsequence of rock bream iridovirus, Virology (2004)325:351–363.
[28] Elsayed E., Faisal M., Thomas M., Whelan G.,Batts W., Winton J., Isolation of viral haemorrhagicsepticaemia virus from muskellunge, Esox masqui-nongy (Mitchill), in Lake St Clair, Michigan, USAreveals a new sublineage of the North Americangenotype, J. Fish Dis. (2006) 29:611–619.
[29] Enzmann P.J., Kurath G., Fichtner D., BergmannS.M., Infectious hematopoietic necrosis virus: mono-phyletic origin of European isolates from NorthAmerican genogroup M, Dis. Aquat. Org. (2005)66:187–195.
[30] Erickson C.L., An artificial landscape-scalefishery in the Bolivian Amazon, Nature (2000)408:190–193.
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 18 of 24 (page number not for citation purpose)

[31] FAO, State of World Fisheries and Aquaculture(SOFIA), Food and Agricultural Organisation of theUnited Nations, Rome, 2008.
[32] FAO, Fishstat Plus, Food and AgriculturalOrganisation of the United Nations, Rome, 2009.
[33] Flegel T.W., Nielsen L., Thamavit V., KongtimS., Pasharawipas T., Presence of multiple viruses innon-diseased, cultivated shrimp at harvest, Aquacul-ture (2004) 240:55–68.
[33b] Flegel T.W., Review of disease transmissionrisks from prawn products exported for humanconsumption, Aquaculture (2009) 209:179–189.
[34] Frerichs G.N., Rodger H.D., Peric Z., Cellculture isolation of piscine neuropathy nodavirus fromjuvenile sea bass, Dicentrarchus labrax, J. Gen. Virol.(1996) 77:2067–2071.
[35] Gagne N., MacKinnon A.M., Boston L., SouterB., Cook-Versloot M., Griffiths S., Olivier G., Isola-tion of viral haemorrhagic septicaemia virus frommummichog, stickleback, striped bass and brown troutin eastern Canada, J. Fish Dis. (2007) 30:213–223.
[36] Garza J.R., Hasson K.W., Poulos B.T., RedmanR.M., White B.L., Lightner R.V., Demonstration ofinfectious Taura syndrome virus in the feces ofseagulls collected during an epizootic in Texas,J. Aquat. Anim. Health (1997) 9:156–159.
[37] Gilad O., Yun S., Adkison M.A., Way K., WillitsN.H., Bercovier H., Hedrick R.P., Molecular compar-ison of isolates of an emerging fish pathogen, koiherpesvirus, and the effect of water temperature onmortality of experimentally infected koi, J. Gen. Virol.(2003) 84:2661–2668.
[38] Glazebrook J.S., Heasman M.P., Debeer S.W.,Picorna-like viral particles associated with massmortalities in larval barramundi, Lates calcariferBloch, J. Fish Dis. (1990) 13:245–249.
[39] Granja C.B., Vidal O.M., Parra G., Salazar M.,Hyperthermia reduces viral load of white spot syn-drome virus in Penaeus vannamei, Dis. Aquat. Org.(2006) 68:175–180.
[40] Groocock G.H., Getchell R.G., Wooster G.A.,Britt K.L., Batts W.N., Winton J.R., et al., Detectionof viral hemorrhagic septicemia in round gobies inNew York State (USA) waters of Lake Ontario andthe St. Lawrence River, Dis. Aquat. Org. (2007)76:187–192.
[41] Haenen O.L.M., Way K., Bergmann S.M., ArielE., The emergence of koi herpesvirus and its signif-icance to European aquaculture, Bull. Eur. Assoc. FishPathol. (2004) 24:293–307.
[42] Hameed A.S.S., Yoganandhan K., Widada J.S.,Bonami J.R., Experimental transmission and tissue
tropism of Macrobrachium rosenbergii nodavirus(MrNV) and its associated extra small virus (XSV),Dis. Aquat. Org. (2004) 62:191–196.
[43] Hasson K.W., Lightner D.V., Poulos B.T.,Redman R.M., White B.L., Brock J.A., Bonami J.R.,Taura syndrome in Penaeus vannamei – Demonstra-tion of a viral etiology, Dis. Aquat. Org. (1995)23:115–126.
[44] Hasson K.W., Lightner D.V., Mohney L.L.,Redman R.M., Poulos B.T., White B.M., Taurasyndrome virus (TSV) lesion development and thedisease cycle in the Pacific white shrimp Penaeusvannamei, Dis. Aquat. Org. (1999) 36:81–93.
[45] He J.G., Wang S.P., Zeng K., Huang Z.J., ChanS.-M., Systemic disease caused by an iridovirus-likeagent in cultured mandarinfish, Sinaperca chuatsi(Basilewshy), in China, J. Fish Dis. (2000) 23:219–222.
[46] He J.G., Deng M., Weng S.P., Li Z., Zhou S.Y.,Long Q.X., et al., Complete genome analysis of themandarin fish infectious spleen and kidney necrosisiridovirus, Virology (2001) 291:126–139.
[47] Hedrick R.P., Gilad O., Yun S., SpangenbergJ.V., Marty G.D., Nordhausen R.W., et al., A herpes-virus associated with mass mortality of juvenile andadult koi, a strain of common carp, J. Aquat. Anim.Health (2000) 12:44–57.
[48] Hedrick R.P., Batts W.N., Yun S., Traxler G.S.,Kaufman J., Winton J.R., Host and geographic rangeextensions of the North American strain of viralhemorrhagic septicemia virus, Dis. Aquat. Org. (2003)55:211–220.
[49] Higginbothan J., Piscinae: artificial fishponds inRoman Italy, University of North Carolina Press,Chapel Hill, 1997.
[50] Hoa T., Hodgson R., Oanh D., Phuong N.,Preston N., Walker P., Genotypic variations in tandemrepeat DNA segments between ribonucleotide reduc-tase subunit genes of white spot syndrome virus(WSSV) isolates from Vietnam, in: Walker P., LesterR., Reantaso M. (Eds.), Diseases in Asian Aquacul-ture, Asian Fisheries Society, Manila, 2005,pp. 395–403.
[51] Hsu H.C., Lo C.F., Lin S.C., Liu K.F., Peng S.E.,Chang Y.S., et al., Studies on effective PCR screeningstrategies for white spot syndrome virus (WSSV)detection in Penaeus monodon brooders, Dis. Aquat.Org. (1999) 39:13–19.
[52] Hyatt A.D., Hine P.M., Jones J.B., WhittingtonR.J., Kearns C., Wise T.G., et al., Epizootic mortalityin the pilchard Sardinops sagax neopilchardus inAustralia and New Zealand in 1995. 2. Identificationof a herpesvirus within the gill epithelium, Dis. Aquat.Org. (1997) 28:17–29.
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 19 of 24

[53] Hyatt A.D., Gould A.R., Zupanovic Z.,Cunningham A.A., Hengstberger S., Whittington R.J.,et al., Comparative studies of piscine and amphibianiridoviruses, Arch. Virol. (2000) 145:301–331.
[54] Inouye K., Yamano K., Maeno Y., Nakajima K.,Matsuoka M., Wada Y., Sorimachi M., Iridovirusinfection of cultured red sea bream, Pagrus major,Fish Pathol. (1992) 27:19–27.
[55] Israngkura A., Sae-Hae S., A review of eco-nomic impacts of aquatic animal disease, in: ArthurJ.R., Phillips M.J., Subasinghe R.P., Reantaso M.B.,McCrae I.H. (Eds.), Primary Aquatic Animal HealthCare in Rural, Small-scale Aquaculture Development,Technical Proceedings of the Asia Regional ScopingWorkshop. FAO Fisheries Technical Paper 406, FAO,Rome, 2002, pp. 55–61.
[56] Isshiki T., Nishizawa T., Kobayashi T., NaganoT., Miyazaki T., An outbreak of VHSV (viral hemor-rhagic septicemia virus) infection in farmed Japaneseflounder Paralichthys olivaceus in Japan, Dis. Aquat.Org. (2001) 47:87–99.
[57] Johnson K.N., van Hulten M.C.W., Barnes A.C.,‘‘Vaccination’’ of shrimp against viral pathogens:phenomenology and underlying mechanisms, Vaccine(2008) 26:4885–4892.
[58] Jones J.B., Hyatt A.D., Hine P.M., WhittingtonR.J., Griffin D.A., Bax N.J., Australasian pilchardmortalities, World J. Microbiol. Biotechnol. (1997)13:383–392.
[59] Kalagayan G., Godin D., Kanna R., Hagino G.,Sweeney J., Wyban J., Brock J., IHHN virus as anetiological factor in runt-deformity-syndrome of juve-nile Penaeus vannamei cultured in Hawaii, J. WorldAquac. Soc. (1991) 22:235–243.
[60] Kawakami H., Nakajima K., Cultured fish spe-cies affected by red sea bream iridoviral disease from1996 to 2000, Fish Pathol. (2002) 37:45–47.
[61] Kiatpathomchai W., Jaroenram W., Arunrut N.,Gangnonngiw W., Boonyawiwat V., Sithigorngul P.,Experimental infections reveal that common Thaicrustaceans are potential carriers for spread of exoticTaura syndrome virus, Dis. Aquat. Organ. (2008)79:183–190.
[62] Kibenge F.S.B., Munir K., Kibenge M.J.T.,Moneke T.J., Moneke E., Infectious salmon anaemiavirus: causative agent, pathogenesis and immunity,Anim. Health Res. Rev. (2004) 5:65–78.
[63] Kibenge F.S.B., Godoy M.G., Wang Y.W.,Kibenge M.J.T., Gherardelli V., Mansilla S., et al.,Infectious salmon anaemia virus (ISAV) isolated fromthe ISA disease outbreaks in Chile diverged fromISAV isolates from Norway around 1996 and wasdisseminated around 2005, based on surface glyco-protein gene sequences, Virol. J. (2009) 6:88.
[64] Kim W.S., Oh M.J., Nishizawa T., Park J.W.,Kurath G., Yoshimizu M., Genotyping of Koreanisolates of infectious hematopoietic necrosis virus(IHNV) based on the glycoprotein gene, Arch. Virol.(2007) 152:2119–2124.
[65] Krabsetsve K., Cullen B.R., Owens L., Redis-covery of the Australian strain of infectious hypoder-mal and haematopoietic necrosis virus, Dis. Aquat.Org. (2004) 61:153–158.
[66] Kurath G., Garver K.A., Troyer R.M., Emme-negger E.J., Einer-Jensen K., Anderson E.D., Phylo-geography of infectious haematopoietic necrosis virusin North America, J. Gen. Virol. (2003) 84:803–814.
[67] Kurath G., Winton J., Fish rhabdoviruses, in: MahyB.W.J., van Regenmortel M.H.V. (Eds.), Encyclopediaof Virology, Elsevier, Oxford, 2008, pp. 221–227.
[68] Kurita J., Yuasa K., Ito T., Sano M., HedrickR.P., Engelsma M.Y., et al., Molecular epidemiologyof koi herpesvirus, Fish Pathol. (2009) 44:59–66.
[69] Kurita J.K., Nakajima I.H., Aoki T., Completegenome sequencing of red sea bream iridovirus(RSIV), Fish. Sci. (2002) 68:1113–1115.
[70] Labreuche Y., O’Leary N.A., de la Vega E.,Veloso A., Gross P.S., Chapman R.W., et al., Lack ofevidence for Litopenaeus vannamei Toll receptor(lToll) involvement in activation of sequence-indepen-dent antiviral immunity in shrimp, Dev. Comp.Immunol. (2009) 33:806–810.
[71] Langdon J.S., Humphrey J.D., Epizootic hema-topoietic necrosis, a new viral disease in redfin perch,Perca fluviatilis L, in Australia, J. Fish Dis. (1987)10:289–297.
[72] Leu J.H., Yang F., Zhang X., Xu X., Kou G.H.,Lo C.F., Whispovirus, Curr. Top. Microbiol. Immunol.(2009) 328:197–277.
[73] Lightner D.V., Redman R.M., Bell T.A., Infec-tious hypodermal and hematopoietic necrosis, a newlyrecognized virus disease of penaeid shrimp, J. Inver-tebr. Pathol. (1983) 42:62–70.
[74] Lightner D.V., Redman R.M., Bell T.A., BrockJ.A., Detection of IHHN virus in P. stylirostris andP. vannamei imported into Hawaii, J. World Maricul-ture Society (1983) 14:212–225.
[75] Lightner D.V., Epizootiology, distribution andthe impact on international trade of two penaeidshrimp viruses in the Americas, Rev. Sci. Tech. Off.Int. Epizoot. (1996) 15:579–601.
[76] Lightner D.V., Redman R.M., Poulos B.T.,Nunan L.M., Mari J.L., Hasson K.W., Risk of spreadof penaeid shrimp viruses in the Americas by theinternational movement of live and frozen shrimp,Rev. Sci. Tech. Off. Int. Epizoot. (1997) 16:146–160.
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 20 of 24 (page number not for citation purpose)

[77] Lightner D.V., Hasson K.W., White B.L.,Redman R.M., Experimental infection of westernhemisphere penaeid shrimp with Asian white spotsyndrome virus and Asian yellow head virus, J. Aquat.Anim. Health (1998) 10:271–281.
[78] Lightner D.V., The penaeid shrimp viral pan-demics due to IHHNV, WSSV, TSV and YHV: historyin the Americas and current status, Proceedings of the32nd Joint UJNR Aquaculture Panel Symposium,Davis and Santa Barbara, California, USA, 2003,pp. 17–20.
[79] Lightner D.V., Pantoja C.R., Poulos B.T., TangK.F.J., Redman R.M., Andrade T.P.D., Bonami J.R.,Infectious myonecrosis: a new disease in Pacific whiteshrimp, Global Aquaculture Advocate (2004) 7:85.
[80] Liu B., Yu Z.M., Song X.X., Guan Y.Q.,Jian X.F., He J.F., The effect of acute salinitychange on white spot syndrome (WSS) outbreaks inFenneropenaeus chinensis, Aquaculture (2006) 253:163–170.
[81] Lotz J.M., Anton L.S., Soto M.A., Effect ofchronic Taura syndrome virus infection on salinitytolerance of Litopenaeus vannamei, Dis. Aquat.Organ. (2005) 65:75–78.
[82] Lovely J.E., Dannevig B.H., Falk K., Hutchin L.,MacKinnon A.M., Melville K.J., et al., First identifi-cation of infectious salmon anaemia virus in NorthAmerica with haemorrhagic kidney syndrome, Dis.Aquat. Organ. (1999) 35:145–148.
[83] Lu L., Zhou S.Y., Chen C., Weng S.P., ChanS.-M., He J.G., Complete genome sequence analysisof an iridovirus isolated from the orange-spottedgrouper, Epinephelus coioides, Virology (2005)339:81–100.
[84] Lumsden J.S., Morrison B., Yason C., Russell S.,Young K., Yazdanpanah A., et al., Mortality event infreshwater drum Aplodinotus grunniens from LakeOntario, Canada, associated with viral haemorrhagicsepticemia virus, Type IV, Dis. Aquat. Organ. (2007)76:99–111.
[85] Lundin C.G., Global attempts to address shrimpdisease, Report of the Land, Water and NaturalHabitats Division, Environment Department, TheWorld Bank, Washington DC, 1995.
[86] Mao J.H., Hedrick R.P., Chinchar V.G., Molec-ular characterization, sequence analysis, and taxo-nomic position of newly isolated fish iridoviruses,Virology (1997) 229:212–220.
[87] Mari J., Poulos B.T., Lightner D.V., BonamiJ.R., Shrimp Taura syndrome virus: genomic charac-terization and similarity with members of the genusCricket paralysis-like viruses, J. Gen. Virol. (2002)83:915–926.
[88] Markussen T., Jonassen C.M., Numanovic S.,Braaen S., Hjortaas M., Nilsen H., Mjaaland S.,Evolutionary mechanisms involved in the virulence ofinfectious salmon anaemia virus (ISAV), a piscineorthomyxovirus, Virology (2008) 374:515–527.
[89] Matsuoka S., Inouye K., Nakajima K., Culturedfish species affected by red sea bream iridoviraldisease from 1991 to 1995, Fish Pathol. (1996)31:233–234.
[90] McColl K., Slater J., Jeyasekaran G., Hyatt A.D.,Crane M.S., Detection of white spot syndrome virusand yellow head virus in prawns imported intoAustralia, Aust. Vet. J. (2004) 82:69–74.
[91] Meyers T.R., Winton J.R., Viral hemorrhagicsepticemia virus in North America, Annu. Rev. FishDis. (1995) 5:3–24.
[92] Miller O., Fuller F.J., Gebreyes W.A., LewbartG.A., Shchelkunov I.S., Shivappa R.B., et al., Phylo-genetic analysis of spring virema of carp virus revealsdistinct subgroups with common origins for recentisolates in North America and the UK, Dis. Aquat.Org. (2007) 76:193–204.
[93] Miller P., Flaherty M., Szuster B., Inland shrimpfarming in Thailand, Aquaculture Asia (1999) 4:27–32.
[94] Miyata M., Matsuno K., Jung S.J., Danayadol Y.,Miyazaki T., Genetic similarity of iridoviruses fromJapan and Thailand, J. Fish Dis. (1997) 20:127–134.
[95] Mjaaland S., Rimstad E., Cunningham C.O.,Molecular diagnosis of infectious salmon anaemia,in: Cunningham C.O. (Ed.), Molecular diagnosisof salmonid diseases, Kluwer Academic Publishers,Dordrecht, The Netherlands, 2002, pp. 1–22.
[96] Morales-Covarrubias M.S., Nunan L.M.,Lightner D.V., Mota-Urbina J.C., Garza-AguirreM.C., Chavez-Snachez M.C., Prevalence of IHHNVin wild broodstock of Penaeus stylirostris from theupper Gulf of California, Mexico, J. Aquat. Anim.Health (1999) 11:296–301.
[97] Mori K., Nakai T., Nagahara M., Muroga K.,Mekuchi T., Kanno T., A viral disease in hatchery-reared larvae and juveniles of redspotted grouper, FishPathol. (1991) 26:209–210.
[98] Mori K.I., Nakai T., Muroga K., Arimoto M.,Mushiake K., Furusawa I., Properties of a new virusbelonging to Nodaviridae found in larval striped jack(Pseudocaranx dentex) with nervous necrosis, Virol-ogy (1992) 187:368–371.
[99] Mortensen H.F., Heuer O.E., Lorenzen N., OtteL., Olesen N.J., Isolation of viral haemorrhagicsepticaemia virus (VHSV) from wild marine fishspecies in the Baltic Sea, Kattegat, Skagerrak and theNorth Sea, Virus Res. (1999) 63:95–106.
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 21 of 24

[100] MundayB.L.,LangdonJ.S.,HyattA.,HumphreyJ.D., Mass mortality associated with a viral-inducedvacuolating encephalopathy and retinopathy of larvaland juvenile barramundi, Lates calcarifer Bloch,Aquaculture (1992) 103:197–211.
[101] Munday B.L., Kwang J., Moody N., Betanoda-virus infections in teleost fish: a review, J. Fish Dis.(2002) 25:127–142.
[102] Naish K.A., Taylor J.E., Levin P.S., Quinn T.P.,Winton J.R., Huppert D., Hilborn R., An evaluation ofthe effects of conservation and fishery enhancementhatcheries on wild populations of salmon, Adv. Mar.Biol. (2008) 53:61–194.
[103] Nakajima K., Maeno Y., Yokoyama K., Kaji C.,Manabe S., Antigen analysis of red sea breamiridovirus and comparison with other fish iridoviruses,Fish Pathol. (1998) 33:73–78.
[104] Nishizawa T., Mori K.I., Furuhashi M., NakaiT., Furusawa I., Muroga K., Comparison of coatprotein genes of 5 fish nodaviruses, the causativeagents of viral nervous necrosis in marine fish, J. Gen.Virol. (1995) 76:1563–1569.
[105] Nishizawa T., Furuhashi M., Nagai T., Nakai T.,Muroga K., Genomic classification of fish nodavirusesby molecular phylogenetic analysis of the coat proteingene, Appl. Environ. Microbiol. (1997) 63:1633–1636.
[106] Nishizawa T., Kinoshita S., Kim W.S., HigashiS., Yoshimizu M., Nucleotide diversity of Japaneseisolates of infectious hematopoietic necrosis virus(IHNV) based on the glycoprotein gene, Dis. Aquat.Org. (2006) 71:267–272.
[107] Nunan L.M., Poulos B.T., Lightner D.V., Thedetection of white spot syndrome virus (WSSV) andyellow head virus (YHV) in imported commodityshrimp, Aquaculture (1998) 160:19–30.
[108] Nunan L.M., Arce S.M., Staha R.J., LightnerD.V., Prevalence of infectious hypodermal and hema-topoietic necrosis virus (IHHNV) and white spotsyndrome virus (WSSV) in Litopenaeus vannamei inthe Pacific Ocean off the coast of Panama, J. WorldAquac. Soc. (2001) 32:330–334.
[109] Nylund A., Hovland T., Watanabe K., EndresenC., Presence of infectious salmon anemia virus (ISAV)in tissues of Atlantic salmon, Salmo salar L, collectedduring 3 separate outbreaks of the disease, J. Fish Dis.(1995) 18:135–145.
[110] Owens L., La Fauce K., Juntunen K.,Hayakijkosol O., Zeng C., Macrobrachiumrosenbergii nodavirus disease (white tail disease) inAustralia, Dis. Aquat. Org. (2009) 85:175–180.
[111] Pantoja C.R., Lightner D.V., Holtschmit K.H.,Prevalence and geographic distribution of infectious
hypodermal and hematopoietic necrosis virus(IHHNV) in wild blue shrimp Penaeus stylirostrisfrom the Gulf of California, Mexico, J. Aquat. Anim.Health (1999) 11:23–34.
[112] Penaranda M.M.D., Purcel M.K., Kurath G.,Differential virulence mechanisms of infectious hema-topoietic necrosis virus in rainbow trout (Oncorhyn-chus mykiss) include host entry and virus replicationkinetics, J. Gen. Virol. (2009) 90:2172–2182.
[113] Peng S.E., Lo C.F., Liu K.F., Kou G.H., Thetransition from pre-patent to patent infection of whitespot syndrome virus (WSSV) in Penaeus monodontriggered by pereiopod excision, Fish Pathol. (1998)33:395–400.
[114] Picco A.M., Collins J.P., Amphibian commerceas a likely source of pathogen pollution, Conserv. Biol.(2008) 22:1582–1589.
[115] Plarre H., Devold M., Snow M., Nylund A.,Prevalence of infectious salmon anaemia virus (ISAV)in wild salmonids in western Norway, Dis. Aquat. Org.(2005) 66:71–79.
[116] Poulos B.T., Tang K.F., Pantoja C.R., BonamiJ.R., Lightner D.V., Purification and characterizationof infectious myonecrosis virus of penaeid shrimp,J. Gen. Virol. (2006) 87:987–996.
[117] Qian D., Shi Z., Zhang S., Cao Z., Liu W., LiL., et al., Extra small virus-like particles (XSV) andnodavirus associated with whitish muscle disease inthe giant freshwater prawn, Macrobrachium rosen-bergii, J. Fish Dis. (2003) 26:521–527.
[118] Robalino J., Bartlett T.C., Chapman R.W.,Gross P.S., Browdy C.L., Warr G.W., Double-strandedRNA and antiviral immunity in marine shrimp:inducible host mechanisms and evidence for theevolution of viral counter-responses, Dev. Comp.Immunol. (2007) 31:539–547.
[119] Sanchez-Martinez J.G., Aguirre-Guzman G.,Mejia-Ruiz H., White spot syndrome virus in culturedshrimp: a review, Aquac. Res. (2007) 38:1339–1354.
[120] Senapin S., Phewsaiya K., Briggs M., FlegelT.W., Outbreaks of infectious myonecrosis virus(IMNV) in Indonesia confirmed by genome sequenc-ing and use of an alternative RT-PCR detectionmethod, Aquaculture (2007) 266:32–38.
[121] Skall H.F., Olesen N.J., Mellergaard S., Viralhaemorrhagic septicaemia virus in marine fish and itsimplications for fish farming – a review, J. Fish Dis.(2005) 28:509–529.
[122] Skliris G.P., Krondiris J.V., Sideris D.C., ShinnA.P., Starkey W.G., Richards R.H., Phylogenetic andantigenic characterization of new fish nodavirusisolates from Europe and Asia, Virus Res. (2001)75:59–67.
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 22 of 24 (page number not for citation purpose)

[123] Smail D.A., Viral haemorrhagic septicaemia,in: Woo P.T.K., Bruno D.W. (Eds.), Fish diseases anddisorders, Volume 3: Viral, bacterial and fungalinfections, CAB International, Oxon, UK, 1999,pp. 123–147.
[124] Spann K.M., Cowley J.A., Walker P.J., LesterR.J.G., A yellow-head like virus from Penaeusmonodon cultured in Australia, Dis. Aquat. Org.(1997) 31:169–179.
[125] St-Hilaire S., Beevers N., Way K., Le DeuffR.M., Martin P., Joiner C., Reactivation of koiherpesvirus infections in common carp Cyprinuscarpio, Dis. Aquat. Org. (2005) 67:15–23.
[126] Stentiford G.D., Bonami J.R., Alday-Sanz V.,A critical review of susceptibility of crustaceans toTaura syndrome, yellowhead disease and white spotdisease and implications of inclusion of these diseasesin European legislation, Aquaculture (2009) 291:1–17.
[127] Stone D.M., Ahne W., Denham K.L., DixonP.F., Liu C.T.Y., Sheppard A.M., et al., Nucleotidesequence analysis of the glycoprotein gene of putativespring viraemia of carp virus and pike fry rhabdovirusisolates reveals four genogroups, Dis. Aquat. Org.(2003) 53:203–210.
[128] Subasinghe R.P., Bondad-Reantaso M.G., Bio-security in aquaculture: International agreements andinstruments, their compliance, prospects, and chal-lenges for developing countries, in: Scarfe A.D., LeeC.S., O’Bryen P.J. (Eds.), Aquaculture biosecurity:prevention, control, and eradication of aquatic animaldisease, Blackwell Publishing, 2006, pp. 9–16.
[129] Subasinghe R.P., Bondad-Reantaso M.G., TheFAO/NACA Asia regional technical guidelines for theresponsible movement of live aquatic animals: lessonslearned from their development and implementation,Rev. Sci. Tech. Off. Int. Epizoot. (2008) 27:54–63.
[130] Sudhakaran R., Musthaq S.S., Haribabu P.,Mukherjee S.C., Gopal C., Hameed A.S.S., Experi-mental transmission of Macrobrachium rosenbergiinodavirus (MrNV) and extra small virus (XSV) inthree species of marine shrimp (Penaeus indicus,Penaeus japonicus and Penaeus monodon), Aquacul-ture (2006) 257:136–141.
[131] Sudhakaran R., Haribabu P., Kumar S.R.,Sarathi M., Ahmed V.P.I., Babu V.S., et al., Naturalaquatic insect carriers of Macrobrachium rosenbergiinodavirus (MrNV) and extra small virus (XSV), Dis.Aquat. Org. (2008) 79:141–145.
[132] Sudthongkong C., Miyata M., Miyazaki T.,Viral DNA sequences of genes encoding the ATPaseand the major capsid protein of tropical iridovirusisolates which are pathogenic to fishes in Japan, SouthChina Sea and Southeast Asian countries, Arch. Virol.(2002) 147:2089–2109.
[133] Tan Y., Xing Y., Zhang H., Feng Y., Zhou Y.,Shi Z.L., Molecular detection of three shrimp virusesand genetic variation of white spot syndrome virus inHainan Province, China, in 2007, J. Fish Dis. (2009)32:777–784.
[134] Tang K.F., Lightner D.V., Low sequence var-iation among isolates of infectious hypodermal andhematopoietic necrosis virus (IHHNV) originatingfrom Hawaii and the Americas, Dis. Aquat. Org.(2002) 49:93–97.
[135] Tang K.F., Poulos B.T., Wang J., RedmanR.M., Shih H.H., Lightner D.V., Geographic varia-tions among infectious hypodermal and hematopoieticnecrosis virus (IHHNV) isolates and characteris-tics of their infection, Dis. Aquat. Org. (2003) 53:91–99.
[136] Tang K.F., Lightner D.V., Infectious hypoder-mal and hematopoietic necrosis virus (IHHNV)-relatedsequences in the genome of the black tiger prawnPenaeus monodon from Africa and Australia, VirusRes. (2006) 118:185–191.
[137] Tang K.F.J., Pantoja C.R., Poulos B.T., RedmanR.M., Lightnere D.V., In situ hybridization demon-strates that Litopenaeus vannamei, L. stylirostris andPenaeus monodon are susceptible to experimentalinfection with infectious myonecrosis virus (IMNV),Dis. Aquat. Org. (2005) 63:261–265.
[138] Tang K.F.J., Pantoja C.R., Redman R.M.,Lightner D.V., Development of in situ hybridizationand RT-PCR assay for the detection of a nodavirus(PvNV) that causes muscle necrosis in Penaeusvannamei, Dis. Aquat. Org. (2007) 75:183–190.
[139] Thiery R., Cozien J., de Boisseson C., Kerbart-Boscher S., Nevarez L., Genomic classification of newbetanodavirus isolates by phylogenetic analysis of thecoat protein gene suggests a low host-fish speciesspecificity, J. Gen. Virol. (2004) 85:3079–3087.
[140] Tu C., Huang H.T., Chuang S.H., Hsu J.P., KuoS.T., Li N.J., et al., Taura syndrome in Pacific whiteshrimp Penaeus vannamei cultured in Taiwan, Dis.Aquat. Org. (1999) 38:159–161.
[141] Tung C.W., Wang C.S., Chen S.N., Histologicaland electron microscopic study on Macrobrachiummuscle virus (MMV) infection in the giant freshwaterprawn,Macrobrachium rosenbergii (de Man), culturedin Taiwan, J. Fish Dis. (1999) 22:319–323.
[142] Van Hulten M.C.W., Witteveldt J., Peters S.,Kloosterboer N., Tarchini R., Fiers M., et al., Thewhite spot syndrome virus DNA genome sequence,Virology (2001) 286:7–22.
[143] Vidal O.M., Granja C.B., Aranguren F., BrockJ.A., Salazar M., A profound effect of hyperthermia onsurvival of Litopenaeus vannamei juveniles infected
Emerging viruses of fish and shrimp Vet. Res. (2010) 41:51
(page number not for citation purpose) Page 23 of 24

with white spot syndrome virus, J. World Aquac. Soc.(2001) 32:364–372.
[144] Vlak J.M., Bonami J.R., Flegel T.W., KouG.H., Lightner D.V., Lo C.F., et al., Family Nimavir-idae, in: Fauquet C.M., Mayo M.A., Maniloff J.,Desselberger U., Ball L.A. (Eds.), Virus Taxonomy:VIIIth Report of the International Committee onTaxonomy of Viruses, Elsevier, Academic Press,London, 2005, pp. 187–192.
[145] Walker P.J., Cowley J.A., Spann K.M.,Hodgson R.A.J., Hall M.R., Withychumnarnkul B.,Yellow head complex viruses: transmission cycles andtopographical distribution in the Asia-Pacific region,in: Browdy C.L., Jory D.J. (Eds.), The new wave:Proceedings of the Special Session on SustainableShrimp Culture, Aquaculture 2001, World Aquacul-ture Society, Baton Rouge, 2001, pp. 292–302.
[146] Walker P.J., Disease emergence and foodsecurity: global impact of pathogens on sustainableaquaculture production, in: Brown A.G. (Ed.), Fish,Aquaculture and Food Security, The ASTE CrawfordFund, Canberra, 2004, pp. 44–50.
[147] Walker P.J., Bonami J.R., Boonsaeng V., ChangP.S., Cowley J.A., Enjuanes L., et al., Family Roni-viridae, in: Fauquet C.M., Mayo M.A., Maniloff J.,Desselberger U., Ball L.A. (Eds.), Virus Taxonomy,VIIIth Report of the International Committee onTaxonomy of Viruses, Elsevier, Academic Press,Amsterdam, 2005, pp 975–979.
[148] Walker P.J., Mohan C.V., Viral disease emer-gence in shrimp aquaculture: origins, impacts and theeffectiveness of health management strategies, Rev.Aquac. (2009) 1:125–154.
[149] Ward J.R., Lafferty K.D., The elusive baselineof marine disease: are diseases in ocean ecosystemsincreasing?, PLOS Biol. (2004) 2:542–547.
[150] Whittington R.J., Chong R., Global trade inornamental fish from an Australian perspective: thecase for revised import risk analysis and managementstrategies, Prev. Vet. Med. (2007) 81:92–116.
[151] Whittington R.J., Crockford M., Jordan D.,Jones B., Herpesvirus that caused epizootic mortalityin 1995 and 1998 in pilchard, Sardinops sagaxneopilchardus (Steindachner), in Australia is nowendemic, J. Fish Dis. (2008) 31:97–105.
[152] Whittington R.J., Becker J.A., Dennis M.M.,Iridovirus infections in finfish – critical review withemphasis on ranaviruses, J. Fish Dis. (2010) 33:95–122.
[153] Wijegoonawardane P.K., Cowley J.A., Phan T.,Hodgson R.A., Nielsen L., Kiatpathomchai W.,Walker P.J., Genetic diversity in the yellow headnidovirus complex, Virology (2008) 380:213–225.
[154] Wijegoonawardane P.K., Sittidilokratna N.,Petchampai N., Cowley J.A., Gudkovs N., WalkerP.J., Homologous genetic recombination in the yellowhead complex of nidoviruses infecting Penaeus mono-don shrimp, Virology (2009) 390:79–88.
[155] Withyachumnarnkul B., Chayaburakul K.,Supak L.A., Plodpai P., Sritunyalucksana K., NashG., Low impact of infectious hypodermal and hema-topoietic necrosis virus (IHHNV) on growth andreproductive performance of Penaeus monodon, Dis.Aquat. Org. (2006) 69:129–136.
[156] Wolf K., Infectious hematopoietic necrosis,Fish viruses and fish viral diseases, Cornell UniversityPress, Ithaca, New York, 1988.
[157] Wongteerasupaya C., Sriurairatana S., VickersJ.E., Akrajamorn A., Boonsaeng V., Panyim S., et al.,Yellow head virus of Penaeus monodon is an RNAvirus, Dis. Aquat. Org. (1995) 22:45–50.
[158] Wongteerasupaya C., Vickers J.E., SriurairatanaS., Nash G.L., Akarajamorn A., Boonsaeng V., et al.,A non-occluded, systemic baculovirus that occurs incells of ectodermal and mesodermal origin and causeshigh mortality in the black tiger prawn Penaeusmonodon, Dis. Aquat. Org. (1995) 21:69–77.
[159] Yang F., He J., Lin X.H., Li Q., Pan D., ZhangX.B., Xu X., Complete genome sequence of theshrimp white spot bacilliform virus, J. Virol. (2001)75:11811–11820.
[160] Yang L.S., Yin Z.X., Liao J.X., Huang X.D.,Guo C.J., Weng S.P., et al., A Toll receptor in shrimp,Mol. Immunol. (2007) 44:1999–2008.
[161] Yoganandhan K., Leartvibhas M., SriwongpukS., Limsuwan C., White tail disease of the giantfreshwater prawn Macrobrachium rosenbergii inThailand, Dis. Aquat. Org. (2006) 69:255–258.
[162] Yoganandhan K., Hameed A.S., Tolerance towhite spot syndrome virus (WSSV) in the freshwaterprawn Macrobrachium rosenbergii is associated withlow VP28 envelope protein expression, Dis. Aquat.Org. (2007) 73:193–199.
[163] Yoshikoshi K., Inoue K., Viral nervous necrosisin hatchery-reared larvae and juveniles of Japaneseparrotfish, Oplegnathus fasciatus (Temminck andSchlegel), J. Fish Dis. (1990) 13:69–77.
[164] Yoshimiza M., Disease problems of salmonidfish in Japan caused by international trade, Rev. Sci.Tech. Off. Int. Epizoot. (1996) 15:533–549.
[165] Zhan W.B., Wang Y.H., Fryer J.L., Yu K.K.,Fukuda H., Meng Q.X., White spot syndrome virusinfection of cultured shrimp in China, J. Aquat. Anim.Health (1998) 10:405–410.
Vet. Res. (2010) 41:51 P.J. Walker, J.R. Winton
Page 24 of 24 (page number not for citation purpose)





























