Embed Size (px)
Citation preview

Central Nervous System Development Objectives1.To describe how the human nervous
system develops.2.To outline the origin of the cells of the
nervous system.3.To discuss the embryological basis of
the congenital malformations of the central nervous system.
Bonus point:1.To review applications of Central
Nervous System development.

Development of the Central Nervous System The central nervous system develops
from the neural tube.

neurulation
Commences after gastrulation (3rd week). It is the formation of the neural tube. First indication of the developing nervous
system is the formation of the neural plate.
The folding commences in the cervical region of the embryo and proceeds cranio-caudally ( i.e. Cranially and caudally).{level of the 5th cervical somite}.
The neural plate folds under influence of the notochord to form the neural tube.


Neurulation continued
Through the process of neurulation the neural plate forms the neural tube.
The stages of neural tube and neural plate occur in different regions of the body: in one region of the body the neural ectoderm is at neural tube stage and in the other region it is at neural plate stage.
Neuromeres are segments of the neural tube more apparent in the body wall regions than the head.

Result of neurulation
Formation of the neural tube and brain vesicles.
NB: it is not all of the neuro-ectoderm that is incorporated into the neural tube some of it is detached and forms the neural crest.
The neural crest migrates in 3 directions1.Ventral steam: autonomic ganglia (pre-
vertebral, para-vertebral & enteric)2.Dorsal stream : joins dermo-my0tomal pert
of somite to form melanocytes and DRG.3.Cranial stream: forms ecto-mesenchyme
that contributes to musculoskeletal structures of the head and neck

Vesicle formation
Initially the neural tube is open @ its cranial and caudal end via the anterior/superior neuropore and caudal/inferior neuropore.
The neuropores close Cranial(day 24) caudal(day 27).
At the cranial end dilations arise forming the 3 primary brain vesicles:
Prosencephalon Mesecencephalon Rhomboencephalon

Spinal cord
The spinal cord is formed after closure of the neuropores.
Initially it is co-extensive with the vertebral column but due to differential growth rates the vertebral column outgrows it.
The interior of the spinal canal is lined with neuro-epithelium consisting of neuroblasts.
The neural cells migrate out of the neuroepithelium and form the cells of the adult nervous system
NB: ALL THE CELLS OF THE CENTRAL NERVOUS SYSTEM ARISE FROM THE NEUROEPITHELIUM EXCEPT MICROGLIA.



Basal and alar plates
Neuroblasts migrate out of the neuro-epithelium and surround the neural canal.
Those that are situated ventrally form the basal plates (anterior horn cells[motor]).
Those that are posterior form the alar plates( dorsal horn cells[sensory]).
Those that remain as part of the neuro-epithelium form the ependymal layer.



Development of the Spinal Cord

Neural Tube Histogenesis
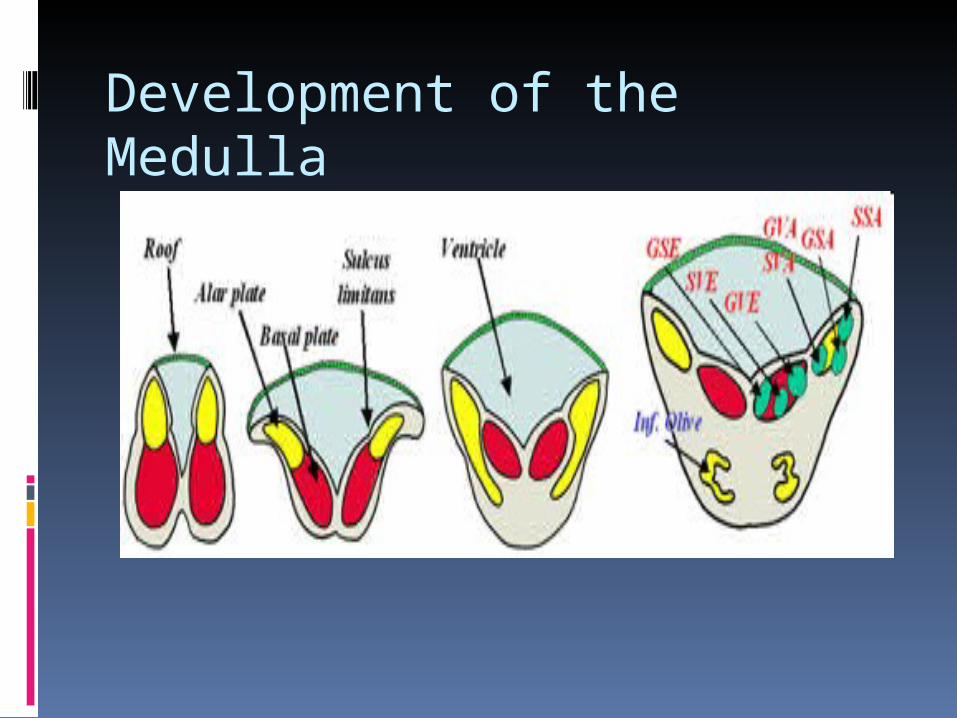
Development of the Medulla

Development of the medulla The most important feature that occurs at the
spino-medullary junction is ALAR PLATE EVERSION.
This occurs due to the presence of a thin roof plate by virtue of expansion of the central canal to form the IVth ventricle.
Thus the alar plates come to lie lateral to the basal plates.
This explains why the sensory nuclei of the cranial nerves are lateral to the motor nuclei.
I.E: SENSORY NUCLEI ARE EITHER DORSAL OR LATERAL WITH MOTOR NUCLEI BEING MEDIAL.

Cranial Nerve Components - Cell Columns
Show Efferent Cell Columns

Spinal nerves contain 4 functional fiber types: general somatic afferents (GSA), general visceral afferents (GVA), general somatic efferents (GSE) and general visceral efferents (GVE).
In cranial nerves there are 3 additional functional components; special somatic afferents (SSA), special visceral (branchial) efferents (SVE) and special visceral (branchial) afferent (SVA).
Cranial nerves may contain anywhere from 1 to 5 fiber types.
Hence, representatives of their cell columns will only be present at those levels of the brain where there is a cranial nerve that requires them i.e., many cell columns will be discontinuous.
In the spinal cord, the components are represented by columns of cells, extending longitudinally, that are more or less continuous.
The basal plate gives rise to the GSEs and GVEs in the ventral gray horn.
The GSE cell column is a continuous cell column (supplying skeletal muscle of somite origin) throughout the cord.

The GVE cell column is an interrupted one - it is represented by the intermediolateral cell column from a) C8 - L2 and b) S2 - S4.
The alar plate gives rise to the: a) continuous GSA cell column represented in
the dorsal gray by nucleus posteromarginalis, substantia gelatinosa, and nucleus proprius,
b) nucleus dorsalis (of Clarke) found between C8 - L3 at the base of the dorsal gray horn, and
c) nucleus gracilis and cuneatus which is present in the low medulla but which receives discriminative touch and proprioception from spinal cord levels.
Sensory fibers from the viscera enter the cord and synapse on the GVA cell column at the base of the dorsal gray horn between C8 - L2 and S2 - S4.

The brainstem develops similarly to the cord i.e., basal and alar plates, but as the roof of the 4th ventricle thins and the medulla and pons “open up”, alar plate derivatives come to lie laterally to those of the basal plate.
Also, the addition of special sensory and special motor components introduce certain changes from the plan of the cord.
None-the-less, the concept of cell columns of specific neuronal components, continuous from the cord and extending into the brain stem, is a useful one.
The concept allows for the anticipation of the location of even discontinuous cell columns; functional cell columns will appear in approximately the same dorsal-ventral or medial-lateral positions in whatever level of the brainstem they appear.
This concept provides a logical scheme to the CNS in spite of what at first appears to be random.

1 - The continuous GSA cell column of the spinal cord dorsal gray horn which handles pain, temperature and touch sensations is continued without interruption into the medulla, pons and midbrain.
In these locations it is variably known as the spinal nucleus of V, principal sensory nucleus of (of CN V) and the mesencephalic nucleus (of CN V).
Any CN carrying GSA information will terminate in these trigeminal nuclei, derived from the alar plate.
Proprioceptive GSA information destined for the cerebellum from the lower extremity terminate in the nucleus dorsalis of the cord.
Similar proprioceptive information from the upper extremity passes to the analagous rostral representation called the accessory cuneate nucleus in the low medulla.

2 - The discontinuous GVA cell column of the spinal cord dorsal gray horn is recognized throughout the medulla when it resumes as the nucleus of the tractus solitarius. Any CN carrying GVA information will terminate in this cell column.
3 - The continuous GSE cell column of the spinal cord ventral gray horn is represented throughout the brainstem by an interrupted cell column of neurons supplying skeletal muscle of head somite origin.
In the medulla there is the motor nucleus of CN XII, in the pons there is the motor nucleus of CN VI, in the midbrain it is represented by the the nearly continuous cell column of CNs III and IV.
4 - The discontinuous GVE cell column of the spinal cord ventral gray horn resumes in the medulla as the dorsal motor nucleus of the vagus and the slightly more rostral inferior salivatory nucleus (of CN IX).
In the caudal pons this discontinuous cell column is represented by the superior salivatory nucleus (of CN VII).
In the midbrain, this interrupted cell column appears again as the nucleus of Edinger-Westphal (of CN III) at the level of the superior colliculus.
5 - Special visceral (branchial) afferents (SVA) are fibers from taste buds and are carried by CNs VII, IX and X.
Upon entering the brainstem, they travel with the GVAs, in the tractus solitarius to the most rostral end of its nucleus (in the pons) where it is called the gustatory portion of the solitary nucleus.
6 - Special visceral (branchial) efferents (SVE) are represented in the brainstem by an interrupted cell column that has no equivalent in the cord.
These neurons innervate skeletal muscle that developed from the mesoderm of the branchial arches e.g., larynx, pharynx, facial muscles, muscles of mastication.
In the medulla it is represented by nucleus ambiguus (CN IX and X) while in the pons it is represented by the motor nuclei of CN VII and V.
7 - Special somatic afferents (SSA) are found only in supraspinal locations and are represented by the laterally-lying vestibular and auditory nuclei in the medulla and pons. They develop from branchial arch structures.
Cranial nerves I and II serve vision (SSA) and olfaction (SVA). However, these are not true nerves. Rather they develop as evaginations of the telencephalon (olfactory nerve) and the diencephalon (optic nerve).

Cranial Nerve Components - Cell Columns
Show Efferent Cell Columns

Development of the Medulla

Development of the Pons The metencephalon is the more rostral component of the
hindbrain and develops into 2 parts, the pons and cerebellum. The pons consists of 2 parts: a) the phylogenetically older dorsal
portion called the pontine tegmentum which lies in the floor of the 4th ventricle and is continuous with the medulla and exhibits a similar organization and
b) the phylogenetically newer basis pontis which develops later. Alar and basal plates, with their afferent and efferent nuclei, lie
medial and lateral to the sulcus limitans , respectively. Cortically-originating fibers synapse on nuclei of the basis pontis
(which migrate there from the alar plate)[cortico-pontine tract]
\\

Cerebellar Development. The cerebellum is first noted at 5-6
weeks as the rhombic lips of the cranial edge of the thinned roof of the 4th ventricle of the hindbrain.
The lips develop from the dorsolateral portions of the alar plates forming the cerebellar primordia.
Growth causes the 2 rhombic lips to fuse in the midline to form the cerebellar plate which covers the 4th ventricle caudal to the mesencephalon.

Development of the Cerebellum


Cerebellar dvpmnt contd. The fused rhombic lips assume a dumbbell
shape - the central unpaired part is the vermis while the lateral knobs are the hemispheres.
By the end of the 4th month growth causes fissures on the surface.
The first fissure to develop is the posterolateral fissure which separates the flocculonodular lobe from the corpus cerebelli which is rostral to it.
The corpus cerebelli grows more rapidly than the flocculonodular lobes and becomes divided by the primary fissure into the anterior and posterior lobes.
The surface of the lobes are further marked by the development of many closely packed transverse gyri called folia.

Cerebellar development

Histogenesis of the cerebellum Histogenesis of the cerebellar cortex - In the 2nd
month of development the cerebellum consists of a ventricular (inner germinal layer), mantle, and marginal zones.
By 19 weeks neuroblasts dividing in inner germinal layer migrate to the marginal layer, proliferate and form the external granular (germinal) layer.
The outer germinal layer produces basket, granule and stellate cells.
The inner germinal layer produces the Purkinje and Golgi cells and the cells of the deep cerebellar nuclei (dentate, globose, emboliform and fastigii).
Radial glial cells extend from the ventricular layer to the surface of the marginal layer and guide the migration of the developing neurons.
Neuroblasts of both dividing cell layers produce glia.

Development of the Midbrain

Mesencephalon The adult mesencephalon (midbrain) is the least modified of the
brainstem structures with regard to basal and alar plates.{MOST IMPORTANT AS FAR AS BEHAVIOUR IS CONCERNED}
Neuroblasts of alar plates migrate to form inferior and superior colliculi (tectum-corpora quadrigemina).
Inferior and superior colliculi are related to the auditory and visual systems respectively.
The general somatic afferent portion of CN V, the mesencephalic nucleus, is also found here.
Oculomotor neurons (GSE) arise from mesencephalic neuroblasts while trochlear motor (GSE) neurons migrate to this location from the metencephalon[ remember: medial longitudinal fasciculus].
The basal plates and the floor plate expand. They form the tegmentum of the cerebral peduncles in which the
motor nuclei of CN III and IV are found in the central gray along with the general visceral efferents to the eye i.e., the Edinger-Westphal nucleus.

Corticofugal fiber tracts help form ventrolateral bulges of the cerebral peduncles.
The embryologic origin of the red nucleus and substantia nigra (from alar or basal plates) in the cerebral peduncles are uncertain.
The cavity of the original neural tube is little modified in the adult midbrain except to be narrowed by growth of the surrounding midbrain structures; it remains as the narrow cerebral aqueduct.

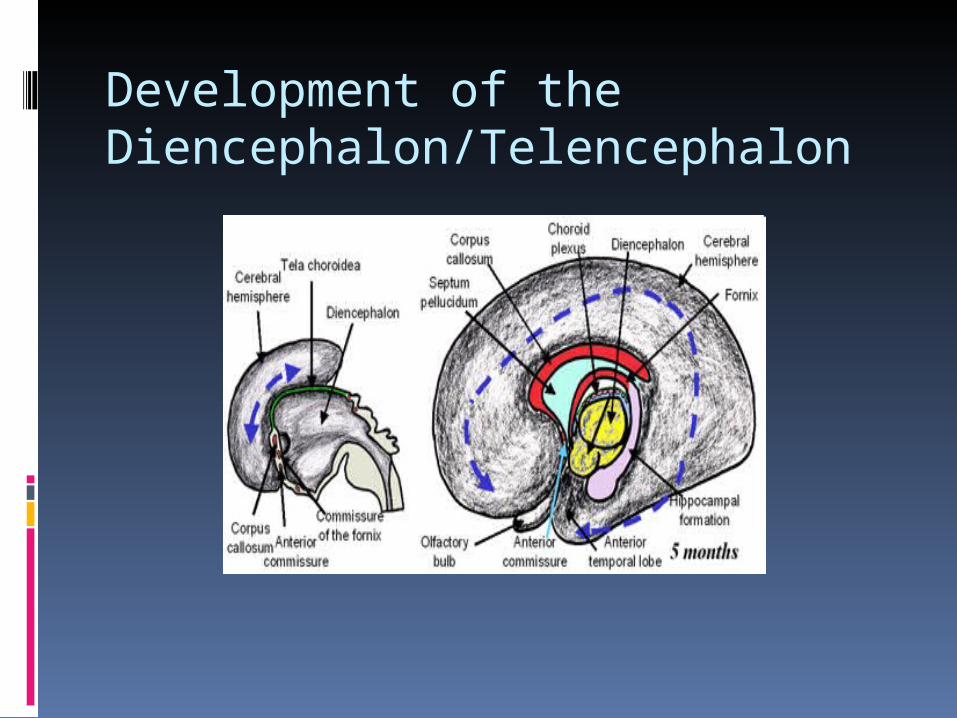
Development of the Diencephalon/Telencephalon

The structure of the early prosencephalon closely resembles the early neural tube i.e., thick lateral walls are connected by thinner floor and roof plates.
The early prosencephalon develops an ocular cup from its ventrolateral walls in the position of the future diencephalon.
In the 5th week, when the embryo reaches the 7 mm stage, the simple plan of the prosencephalon changes; paired telencephalic vesicles and the diencephalon begin to form; the optic cup continues its development.
Rapid bilateral expansion of the telencephalon results in rearward overgrowth of the diencephalon.
An early separation of the telencephalon and diencephalon occurs through a shallow tele-diencephalic sulcus which deepens and persists in the adult as the horizontal transverse cerebral fissure (filled with duplicated pia and blood vessels, called velum interpositum).
As development progresses the growing hemispheres continue their caudal expansion so as to cover the mesencephalon

Development of the Diencephalon/Telencephalon

Development of the Diencephalon/Telencephalon

In the embryo, as in the adult, the roof of the diencephalon is very thin comprised only of ependyma plus adjacent pia called the tela choroidea.
When blood vessels invade the tela choroidea the choroid plexus of the 3rd (and lateral) ventricles develop and invade the ventricles.
The thin medial wall of the ventricle is the choroid fissure; its most rostral point is at the interventricular foramen. In the adult the choroid fissure is “C”-shaped and continuous in the medial walls of the parietal and temporal lobes.
Early in development, the diencephalon develops 2 pairs of prominent swellings in the walls of the 3rd ventricle.
The swellings represent only the alar plate; there is no basal plate.
The largest mass is the thalamus dorsally, separated by the hypothalamic sulcus from the ventral hypothalamus.
Where the roof plate thickens along the medial wall of the thalamus are the smaller swellings of the epithalamus comprised of the a) midline pineal gland, b) paired habenular nuclei, and c) paired stria medullaries.

Development of the Diencephalon/Telencephalon

Other diencephalic structures The lamina terminalis (represents the membrane formed at
the point of closure of the anterior neuropore) is the most rostral structure of the early telencephalon.
By 10 weeks it contains the rudiments of the commissural bundles i.e., corpus callosum, optic chiasm and anterior commissure.
The lamina terminalis provides the only location where nerves interconnect the cerebral hemispheres.
Crossing fibers of the optic chiasm develop in the lamina ventrally, the anterior commissure connects olfactory bulbs and temporal lobes, the commissure of the fornix joins the hippocampal formations, and most dorsally is the corpus callosum which connects non-olfactory cortical areas.
Other crossing fibers do not connect the hemispheres e.g., the posterior commissure connects preoptic areas, the habenular commissure connects habenular nuclei.
The interventricular foramina, which connects the 3rd ventricle with the lateral ventricles, lies just caudal to the lamina terminalis.).
The stria medullaris and tela choroidea form the roof of the 3rd ventricle.
Development of the Diencephalon/Telencephalon

Corpus callosum The largest commissural bundle is the corpus
callosum. It begins as a small bundle but it grows as the
cerebral hemispheres expand, encircling the diencephalon and becoming “C”-shaped.
First, the hemispheres form parietal and frontal lobes; posterior expansion next forms occipital lobes followed by growth in an inferior direction producing temporal lobes.
By its caudalward growth the hemispheres arch over the tela choroidea and choroid plexus of the roof of the diencephalon.
This “C”-shaped expansion causes many underlying structures to also become “C”-shaped e.g., corpus callosum, lateral ventricle, choroid plexus, caudate nucleus, fornix.
The insula is a lobe of the brain that remains relatively undeveloped as it retains a fixed position while the rest of the hemisphere grows around and over it.

Early in development hippocampal ridges form on the medial aspect of the telencephalic vesicles.
These ridges are joined by the commissure of the fornix.
When the temporal lobes grow downward and forward in the older embryo, the hippocampal formation forms a ring of gray matter (just above the choroid fissure) on the medial aspect of the hemispheres which is continuous with the “C”-shaped fornix.
As both the fornix and corpus callosum grow rearward they become separated and the tissue remaining between them is the septum pellucidum.
The anterior commissure connect olfactory bulbs and anterior temporal lobes.

Development of the Diencephalon/Telencephalon

The corpus striatum is comprised of two structures a) caudate-putamen and b) globus pallidus.
In the adult the caudate and putamen are partially separated by fibers of the anterior limb of the internal capsule.
The posterior limb of the internal capsule separates the globus pallidus from the diencephalon (thalamus).
Early in the development of the cerebral vesicles. The caudate-putamen develops from neuroblasts of the
floor of the developing telencephalic vesicle called the striatal ridge while the globus pallidus originates from neuroblasts in the wall of the 3rd ventricle of the diencephalon.
Pallidal neuroblasts eventually migrate laterally to join the caudate-putamen.
Both parts of the corpus striatum originate near the foramen of Monro.
At first, during the caudal expansion of the telencephalic vesicles, the hemispheres are separate from the diencephalon (except at the foramen of Monro).

Development of the Diencephalon/Telencephalon

Later, when the hemispheres begin their complex bending, the medial aspects of the growing hemispheres approach the diencephalon and their apposing surfaces fuse.
After the fusion, corticofugal fibers heading to and from the cerebral cortex incompletely divide the caudate nucleus from the putamen.
Later, then putamen merges with the globus pallidus forming the lenticular nucleus.
As the hemisphere grows backward, the body of the caudate nucleus portion of the striatal ridge in the floor of the lateral ventricle follows the wall of the developing lateral ventricle downward into the roof of the inferior horn of the lateral ventricle where it forms the tail of the caudate nucleus.


Development of the Diencephalon/Telencephalon

Fetal neurogenesis The layered structure of the adult cerebral
cortex forms from the telencephalon as the surface area increases. At first there are 3 zones to the cortex:
1) germinal zone, immediately surrounding the lateral ventricle,
2) intermediate zone, becomes the white matter, and
3) marginal zone, becomes the gray matter. Neuroblasts of the germinal zone divide and
migrate into the marginal zone to become neurons and glial cells.
The marginal zone is formed by cells migrating a) early, which become the deeper layers of the cortex while those that migrate b) later, form the more superficial layers.

Between 6 and 8 months 6 layers are observed in the marginal zone of the neocortex, recognized by cellular and fiber laminae.
The 6-layered cortex is further distinguished as having two main divisions:
1) the deeper highly cellular pyramidal layer are layers II - VI and
2) the more superficial molecular layer (layer I) is comprised mostly of fibers.
The intermediate zone becomes the white matter of the cerebral hemispheres which is traversed by the processes of the cells migrating toward the surface.
At birth the neocortex has a 6-layered structure while cortical areas dealing with olfaction (paleocortex) and the hippocampal formation (archicortex) do not have a six-layered structure.

Development of the Diencephalon/Telencephalon

Early in the development e.g., 12 weeks, of the cerebral hemispheres the surface is smooth, without sulci.
The various lobes of the brain are present.
By about 7 months rapid growth of the hemispheres results in the development of gyri and sulci.
At birth, the main gyri and sulci of adult brain are recognized.

Development of the Ventricular System (Late)

The pattern of the ventricular system is established early. The prosencephalon separates into 2 expanding
telencephalic vesicles (lateral ventricles) and a slit-like diencephalic vesicle (3rd ventricle).
Thickening of its walls narrows the lumen of this segment to a vertical slit, the 3rd ventricle.
The lumina of the lateral and 3 ventricles connect just behind the lamina terminalis via the paired foramina of Monro.
Expansion of the mesencephalon narrows its vesicle forming the cerebral aqueduct, which connects the 3rd and 4th ventricle of the rhombencephalon.
The rhombencephalic roof is thin and covers the 4th ventricle which forms a shallow, diamond-shaped depression.
The expansion of the cerebral hemispheres affects the shape of the lateral ventricles which become “C”-shaped.

Development of the Choroid Plexus

Choroid plexus
A rich capillary plexus develops in the connective tissue around the neural tube of the 6-8 mm embryo.
The roofs of the prosencephalon and rhombencephalon are thin, composed of ependyma that becomes invaded by the capillaries.
Ependyma plus pia mater is called tela choroidea; vascularized tela choroidea is choroid plexus.

“Holoprosencephaly” is an anomaly resulting from disturbances of early forebrain development.
Because of the influence of the brain on surrounding structures facial defects are common.
Fully developed cases frequently include facial defects, microcephaly, and closely set eyes.
Cyclopia: During early normal development the eye fields form on either side of the diencephalon.
Failure of development of the ventral midline portion of the prosencephalon allows the eye fields to form in close proximity to one another resulting in cyclopia.
In the example shown, the near absence of upper and mid-facial tissue allowed the fusion of the optic primordia below a tubular proboscis (nose).

Development of the EyeCyclopia

Defects
Anencephaly; Hydrocephaly; Hydramcepahaly; Spina bifida cystica / occulta
/with/without meningocele or myelomeningocele;
Lissencephaly- absence of gyri; Microcephaly; Machiafava-Bignami Syndrome; Craniosynostosis; Myeloschisis ;