Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 368~198-214 (1996)
Efferent Connectivity of the Hippocampal Formation of the Zebra Finch (Taenopygia guttata): An Anterograde Pathway Tracing
Study Using Phaseolus vulgaris leucoagglutinin
ANDREA D. SZEKELY AND JOHN R. KREBS Departments of Pharmacology and Zoology, University of Oxford, Oxford OX1 3PS,
United Kingdom
ABSTRACT The avian hippocampal formation (HP) is considered to be homologous to the mammalian
hippocampus, being involved in memory formation and spatial memory in particular. The subdivisions and boundaries of the pigeon hippocampus have been defined previously by various morphological methods to detect further similarities with the mammalian homologue. We studied the efferent projections of the zebra finch hippocampus by applying Phaseolus vulgaris leucoagglutinin, and three main subdivisions were distinguished on the basis of the connectivity patterns.
Dorsolateral injections gave rise to projections innervating the rostralmost extension of the HP, a laminar complex including the dorsal and ventral hyperstriata and the lamina frontalis superior, the rostra1 lobus parolfactorius, the medial and ventral paleostriatal regions, the lateral septal nucleus, the nucleus of the diagonal band, the dorsolateral corticoid area, the archistriatum posterius, and the nucleus taeniae in the telencephalon. In the diencephalon, labelled axons were seen in the periventricular and lateral hypothalamus, including the lateral mammillary nuclei, and in the dorsolateral and the dorsomedial posterior thalamic nuclei, whereas, in the midbrain, only the area ventralis of Tsai contained hippocampal fibres. With the exception of the bilateral archistriatal efferents, all projections were ipsilateral.
Dorsomedial injections gave rise to a local fibre system that was almost completely restricted to the ipsilateral hippocampal formation. In addition, lectin-containing fibres continued in the dorsal septal region and a thin band in the hyperstriatum accessorium, adjacent to the lateral ventricle. Ventral injections gave rise to axons innervating ipsilaterally the dorsolateral subdivision, and bilaterally the medial septal nuclei and the contralateral ventral hippocampus. a 1996 Wiley-Liss, Inc.
Indexing terms: avian, hippocampal efferents, limbic circuitry, memory
The avian hippocampal formation (HP) is considered to be homologous with the mammalian hippocampal complex (Campbell and Hodos, 1970) in terms of topography (Craigie, 1930; Ariens-Kappers et al., 1936) and developmental origin (Kuhlenbeck, 1938; Kallh, 1962). It also is thought to be similar in function to its mammalian counterpart, playing a role in memory formation and, in particular, in the processing of spatial memory (birds: Bingman et al., 1988; Krebs et al., 1989; Bingman and Yates, 1992; mam- mals: O’Keefe and Nadel, 1978; Morris et al., 1989; Squire, 1992). Long-term potentiation of synaptic efficacy, the putative physiological correlate of memory formation, has
also been described by Wieraszko and Ball (1991, 1993) in both the songbird and the pigeon HP, respectively.
However, the morphological organisation of the avian HP is very different from that of mammals, and components similar to the subdivisions of the mammalian structure are not clearly recognisable. Recent attempts to characterise the subdivisions of the avian HP and to draw parallels
Accepted November 24, 1995. Address reprint requests to Dr. Andrea D. Szekely, First Department of
Anatomy, Semmelweis University of Medicine, Budapest IX, Tuzolto utca 58, H-1450, Hungary. E-mail: adszekely(6 anal.sote.hu
O 1996 WILEY-LISS, INC.

HIPPOCAMPAL PROJECTIONS IN THE SONGBIRD 199
ventralHP: anterior, +1 - +2.8; lateral, (+) -O.l;ventral, 1; (n = 6).
The beak bar was 5 mm under zero level of the instru- ment. All of the anterior coordinates were measured from lambda, because Bregma is not distinguishable in adult songbirds; the lateral coordinates were measured from the midline, and the ventral coordinates were measured from the brain surface. Following iontophoresis, the skin was sutured, and the birds were allowed to recover in a hospital cage until their balance and flying abilities were normal.
between these subdivisions and those of the mammalian hippocampus have used cytoarchitectural (Montagnese et al., 1996) and immunocytochemical (Erichsen et al., 1991; Krebs et al., 1991) landmarks. In the present paper, we describe the efferent connections of the zebra finch HP by using Phaseolus vulgaris leucoagglutinin (PHAL), a sensi- tive anterograde tracer, to complement these other ap- proaches.
The connectivity of the avian HP has been mapped previously in pigeons (Columba livia; Benowitz and Karten, 1976; Krayniak and Siegel, 1978; Casini et al., 1986), an avian species that is used frequently in behavioural and morphological studies. However, much recent work on the functional role of the avian HP has focused on songbirds (Passeriformes). Therefore, we chose the zebra finch to establish whether the connections of the avian HP are identical in different avian orders.
MATERIALS AND METHODS Hippocampal injections
Sixteen adult zebra finches (Taenopygia guttata) of both sexes (weighing 13-16 g) were injected with PHAL. The iontophoretic injections were placed into either the right or the left hippocampi to reveal any possible differences in connectivity. The birds were anaesthetised with Equithesin (0.025 ml/lO g body weight) and mounted in a Kopf stereotaxic frame. Following incision of the skin and partial removal of either the right or the left parietal bone, the hippocampal formation was exposed. Unilateral injections of PHAL (Vector; 2.5%) were delivered by iontophoresis by using a current injector with 7 seconds on17 seconds off cycles at 5-8 FA for approximately 15-20 minutes. At first, the injection sites (Fig. 1A-C) were estimated empirically, because there is no stereotaxic atlas available for zebra finches. Subsequently, the following relative coordinates (given in millimeters) were established in the rostrocaudal central portion, where the hippocampal formation has its largest extension: dorsomedial HP: anterior, + 1 - +2.8; lateral, (+) -0.1; ventral, 0.3; (n = 4); dorsolateral HP: anterior, + 1 - +2.8; lateral, (+) -0.2; ventral, 0.7; (n = 6);
Control injections Four zebra finches received PHAL injections into the
adjacent telencephalic regions, the rostra1 hyperstriatum accessorium (two birds; Fig. 2A) and the medial hyperstria- tum ventrale (two birds; Fig. 2B). The birds were treated as described above.
Perfusion and PHAL immunocytochemistry Fourteen days postsurgery, the birds were terminally
anaesthetised with Sagatal and intracardially perfused first with heparinised saline (1 ml heparin in 250 ml 0.75% NaC1) and then with a fixative containing 2% paraformalde- hyde, 1% glutaraldehyde, and 0.5% picric acid in 0.1 M phosphate buffer, pH 7.4. The brains were dissected out and immersed in 30% sucrose for cryoprotection. Frozen sections of 60-80 pm thickness were cut on a freezing microtome and processed further according to the method of Gerfen and Sawchenko (1984). Briefly, free-floating sections were rinsed in 0.05 M Tris buffered saline (TBS), pH 7.5, reacted in the presence of 0.5% Triton X-100 with a biotinylated primary antiserum against PHAL (Vector, 1:200) at 4°C for 48 hours, and then reacted with the avidin-biotin complex (1: 100; ABC Vectakit Elite; Vector) at room temperature for 2 hours. All of the antibodies were diluted in TBS. The immunoprecipitate was visualised by using 3,3'-diaminobenzidine (Sigma) intensified with nickel- ammonium sulphate (Merck) in the presence of 0.001%
A AP APH AVT B CA Cb CO CPa DL DLC DM DMP E EM FA FDB FPL GCT GLv HA HD HP
archistriatum archistriatum posterius area parahippocampalis area ventralis tegmentalis (Tsai) nucleus basalis commissura anterior cerebellum chiasma opticum commissura pallii dorsolateral subdivision of HP area corticoidea dorsolateralis dorsomedial subdivision of HP nucleus dorsomedialis posterior thalami ectostriatum nucleus ectomammillaris tractus frontoarchistriaticus fasciculus diagonalis Brocae fasciculus prosencephali lateralis substantia grisea centralis nucleus geniculatus lateralis, pars ventralis hyperstriatum accessorium hyperstriatum dorsale hippocampus proper (by Karten and Hodos, 1967)
Abbreviations
HP Hv J lfs lfsm lh lmd LPO NEO OM OT ov PA PVT R S SCE SL SM Tn TSM
V V
hippocampal formation in the present text hyperstriatum ventrale septohippocampal junction lamina frontalis superior lamina frontalis suprema lamina hyperstriatica lamina medullaris dorsalis lobus parolfactorius neostriatum tractus occipitomesencephalicus tectum opticum nucleus ovoidalis paleostriatum augmentatum paleostriatum ventrale nucleus rotundus septum stratum cellulare externum nucleus septalis lateralis nucleus septalis medialis nucleus taeniae tractus septomesencephalicus ventriculus lateralis ventral subdivision of HP
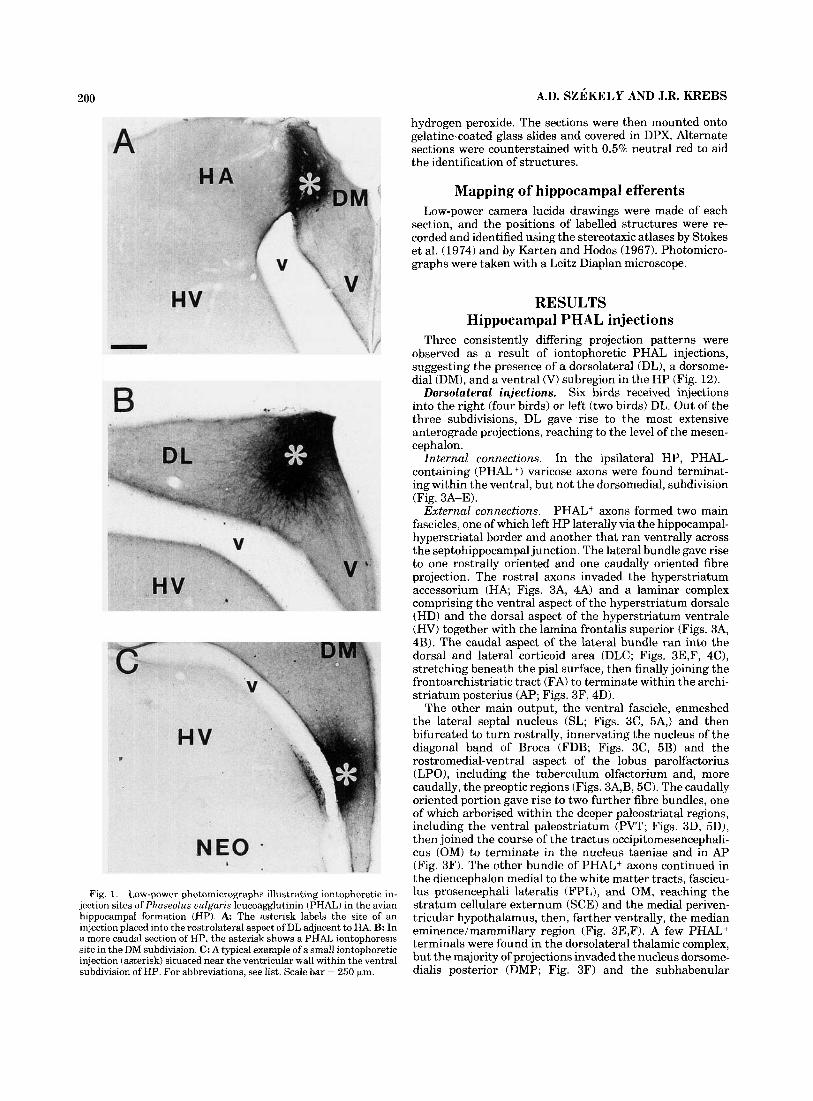
200 A.D. SZEKELY AND J.R. KREBS
Fig. 1. Low-power photomicrographs illustrating iontophoretic in- jection sites of Phaseolus uulgaris leucoagglutinin (PHAL) in the avian hippocampal formation (HP). A The asterisk labels the site of an injection placed into the rostrolateral aspect of DL adjacent to HA. B: In a more caudal section of HP, the asterisk shows a PHAL iontophoresis site in the DM subdivision. C: A typical example of a small iontophoretic injection (asterisk) situated near the ventricular wall within the ventral subdivision of HP. For abbreviations, see list. Scale bar = 250 km.
hydrogen peroxide. The sections were then mounted onto gelatine-coated glass slides and covered in DPX. Alternate sections were counterstained with 0.5% neutral red to aid the identification of structures.
Mapping of hippocampal efferents Low-power camera lucida drawings were made of each
section, and the positions of labelled structures were re- corded and identified using the stereotaxic atlases by Stokes et al. (1974) and by Karten and Hodos (1967). Photomicro- graphs were taken with a Leitz Diaplan microscope.
RESULTS Hippocampal PHAL injections
Three consistently differing projection patterns were observed as a result of iontophoretic PHAL injections, suggesting the presence of a dorsolateral (DL), a dorsome- dial (DM), and a ventral (V) subregion in the HP (Fig. 12).
Six birds received injections into the right (four birds) or left (two birds) DL. Out of the three subdivisions, DL gave rise to the most extensive anterograde projections, reaching to the level of the mesen- cephalon.
In the ipsilateral HP, PHAL- containing (PHAL+) varicose axons were found terminat- ing within the ventral, but not the dorsomedial, subdivision (Fig. 3A-E).
PHAL+ axons formed two main fascicles, one of which left HP laterally via the hippocampal- hyperstriatal border and another that ran ventrally across the septohippocampal junction. The lateral bundle gave rise to one rostrally oriented and one caudally oriented fibre projection. The rostra1 axons invaded the hyperstriatum accessorium (HA; Figs. 3A, 4A) and a laminar complex comprising the ventral aspect of the hyperstriatum dorsale (HD) and the dorsal aspect of the hyperstriatum ventrale ( H V ) together with the lamina frontalis superior (Figs. 3A, 4B). The caudal aspect of the lateral bundle ran into the dorsal and lateral corticoid area (DLC; Figs. 3E,F, 4C), stretching beneath the pial surface, then finally joining the frontoarchistriatic tract (FA) to terminate within the archi- striatum posterius (AP; Figs. 3F, 4D).
The other main output, the ventral fascicle, enmeshed the lateral septal nucleus (SL; Figs. 3C, 5A,) and then bifurcated to turn rostrally, innervating the nucleus of the diagonal band of Broca (FDB; Figs. 3C, 5B) and the rostromedial-ventral aspect of the lobus parolfactorius (LPO), including the tuberculum olfactorium and, more caudally, the preoptic regions (Figs. 3A,B, 5C). The caudally oriented portion gave rise to two further fibre bundles, one of which arborised within the deeper paleostriatal regions, including the ventral paleostriatum (PVT; Figs. 3D, 5D), then joined the course of the tractus occipitomesencephali- cus (OM) to terminate in the nucleus taeniae and in AP (Fig. 3F). The other bundle of PHAL+ axons continued in the diencephalon medial to the white matter tracts, fascicu- lus prosencephali lateralis (FPL), and OM, reaching the stratum cellulare externum (SCE) and the medial periven- tricular hypothalamus, then, farther ventrally, the median eminenceimammillary region (Fig. 3E,F). A few PHAL+ terminals were found in the dorsolateral thalamic complex, but the majority of projections invaded the nucleus dorsome- dialis posterior (DMP; Fig. 3F) and the subhabenular
Dorsolateral injections.
Internal connections.
External connections.

HIPPOCAMPAL PROJECTIONS IN THE SONGBIRD 201
Fig. 2. Low-power photomicrographs of small iontophoretic PHAL injections in the telencephalic control regions. The asterisks indicate the sites of injection in the rostra1 aspect of HA (A) and in the medial part of H V (B) from a more caudal section. For abbreviations, see list. Scale bar = 250 km.
region (not shown). A small number of varicose hippocam- pal fibres were present in the midbrain central gray and in the area ventralis tegmentalis of Tsai (AVT; Fig. 3F), but no further efferents were detected within or below the mesen- cephalon. All DL projections were ipsilateral; however, there was a small contingent of varicose PHAL+ fibres in the ventral portion of the archistriatal complex contralat- eral as well as ipsilateral to the injection (Fig. 3F). One PHAL injection that was placed in the DL subdivision of HP resulted in retrogradely filled perikarya (not shown) in an adjacent region previously identified as the substance P field in the pigeon (Erichsen et al., 1991).
Four birds were injected into the left (two birds) and right (two birds) HP. The projec- tions gave the impression of a local fibre system that was almost completely restricted to the ipsilateral HP (Fig. 6A-C). In addition to the dorsolateral and ventral subdivi- sions of HP, the dorsalmost aspect of the septum (Figs. 6B, 7A) and a thin band of HA along the dorsal wall of the lateral ventricle contained PHAL+ axon terminals (Figs. 6B, 7B).
Six zebra finches received iontopho- resis of PHAL into the left (four birds) and right (two birds) HP. The injections gave rise to internal axons invading the DL, but not the DM, subfield of HP (Figs. 8A-C, 9A). Labelled fibres passed through the septohippocampal junc- tion, profusely arborised within the medial septa1 nucleus (SM; Figs. 8B, 9B), and invaded the contralateral septum via the commissurapallii (CPa; Figs. BC, 9B). The contralat- eral SM and the ventral HP also contained PHAL+ varicose efferents (Figs. 8B-C, 9B). One PHAL injection resulted in retrogradely filled neurones in the DM and DL subdivisions of HP, and, in another case, the ipsilateral ventromedial archistriaturn contained a few PHAL+ axons (not shown).
Dorsomedial injections.
Ventral injections.
Control injections We do not attempt to give an exhaustive description of
efferents arising from the two hyperstriatal regions HA and HV. Only the characteristic projections that differ from those of HP injections are mentioned. HA and HV were
selected as control regions, because they lie adjacent to the hippocampal complex, and their projections partially over- lap with the distribution of hippocampal efferents. Injec- tions to DL or V may also have covered parts of the adjacent HA and HV, respectively, thus, possibly resulting in a composite of labelling from both regions. In an effort to distinguish between HP and HAiHV axons, we compared the patterns of efferent projections arising from hippocam- pal or hyperstriatal regions.
Hyperstriatum accessorium injections. Two zebra finches received iontophoretic injections into the rostrome- dial aspect of HA. The projection pattern of PHAL+ effer- ents was in agreement with previous observations in the pigeon (Miceli et al., 1987). All projections were ipsilateral.
Intratelencephalic HA projections were seen in all layers of the visual Wulst, including HA, HD (Fig. lOA), and hyperstriatum intercalatum supremum (HIS) (not shown). In HV and in the neostriatum (NEO), PHAL+ fibres were observed along the entire rostrocaudal axis (Fig. 10A-D). The parvicellular portions of the avian basal ganglia, the paleostriatum augmentatum (PA), and especially the cen- tral-dorsal LPO were also heavily enmeshed with HA efferents (Fig. 1OA-C). A small number of labelled fibres innervated the dorsolateral aspect of the archistriatal com- plex (Fig. 10D).
Extratelencephalic projections left HA via HP and joined the tractus septomesencephalicus (TSM; Fig. 10B). Termi- nating and/or passing axow were observed in the dience- phalic and mesencephalic retinorecipient visual nuclei, including the dorsolateral nuclear complex and the ventral portion of the lateral geniculate nucleus. Also, the perirotun- dal region (thought to be equivalent to the intergeniculate leaflet of mammals; Hamassaki and Britto, 1990; Gun- turkun and Karten 1991; Szkkely et al., 19921, the pretectal nuclei, the nucleus ectomammillaris, and the tectum opti- cum (Fig. 10C,D) contained HA efferents.
Hyperstriatum ventrale injections. Two zebra finches received PHAL injections into the centromedial aspect of HV that gave rise to exclusively ipsilateral intratelence- phalic projections. Our results are in agreement with the
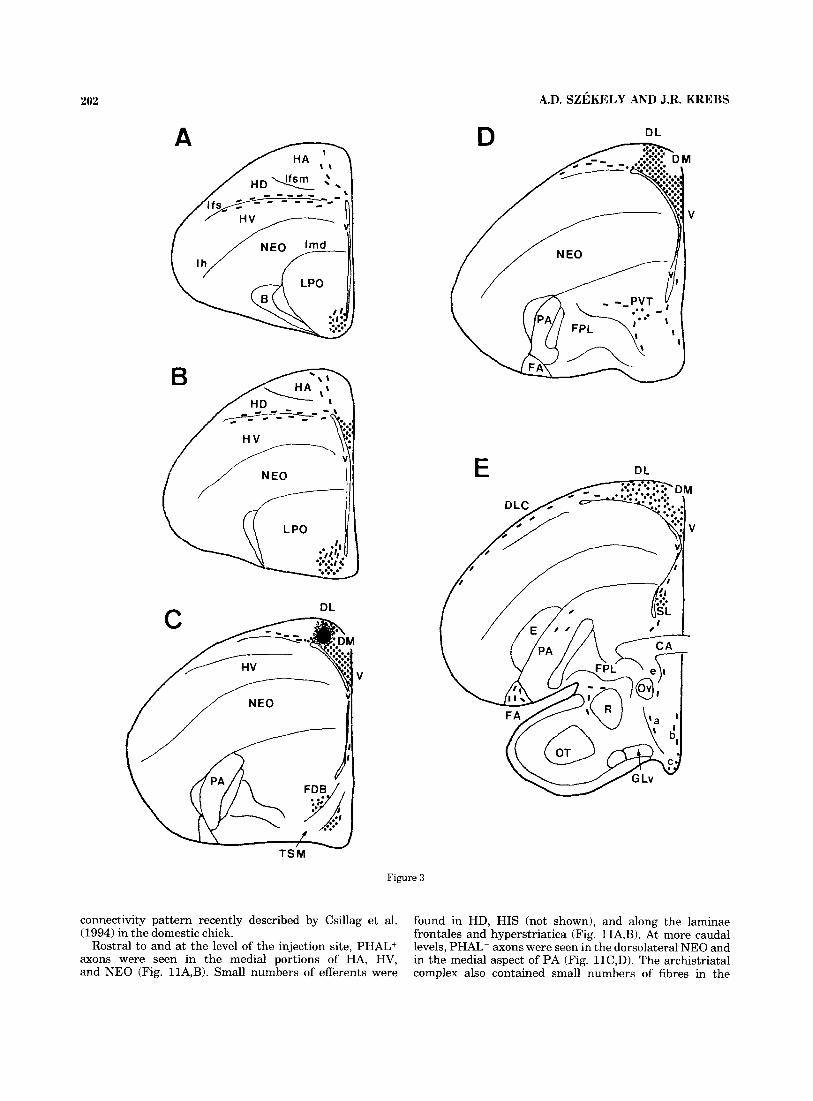
202 A.D. SZEKELY AND J.R. KREBS
DL
DL
V
TS M
Figure 3
connectivity pattern recently described by Csillag et al. (1994) in the domestic chick.
Rostral to and at the level of the injection site, PHAL+ axons were seen in the medial portions of HA, HV, and NEO (Fig. 11A,B). Small numbers of efferents were
found in HD, HIS (not shown), and along the laminae frontales and hyperstriatica (Fig. 11A,B). At more caudal levels, PHAL+ axons were seen in the dorsolateral NEO and in the medial aspect of PA (Fig. 11C,D). The archistriatal complex also contained small numbers of fibres in the

HIPPOCAMPAL PROJECTIONS IN THE SONGBIRD 203
Fig. 3. A-G: Schematic diagram of the distribution of antero- gradely labelled fibres (dashed lines) and terminal fields (dotted areas) in the zebra finch brain after PHAL injection (solid filled area) in the DL subdivision of HP. The illustration demonstrates a single representa-
tive case that has been repeated successfully in five other birds. a, SCE; b, periventricular hypothalamic nuclei; c, mammillary region; d, nucleus pretectalis; e, OM; f, nucleus ruber; g, nervus oculomotorius. The white matter laminae are named only in A. For abbreviations, see list.
ventral and, more caudally, in the dorsomedial regions (Fig. such as Nissl staining, fail to reveal a subdivisional system l lC,D) . identical to the one evident from anterograde tracing
studies. Although the outer boundaries of the hippocampal complex were rather well defined. especially the septohippo- Comparison to cytoarchitecture
The present results obtained by PHAL tracing do not harmonise fully with previous parcellation schemes of avian HP based on cytoarchitecture or on the distribution of neurochemical markers. Routine histological techniques,
campal junction, the only visible citoarchitectural dfffer- ence within HP (Fig. 12) was represented by a group of large neurones in the “v”-shaped structure (see below). The internal borderlines, as superimposed over the Nissl-

204 A.D. SZEKELY AND J.R. KREBS
Fig. 4. Photomicrographs demonstrating PHAL-labelled, sparsely varicose fibres from DL arborising and partially terminating in the ipsilateral hemisphere of the zebra finch forebrain. A: A fine web of stained axons (arrowheads) arises from the neighbouring DL of HP and invades HA. Some of the fibres join a laminar complex of PHAL+ axons (arrowheads in B) overlapping the HDiHV interface together with the
separating white matter lamina, lfs. C: More caudally, PHAL-labelled varicose axons form a dense network within the DLC (arrowheads) while travelling towards the tractus frontoarchistriaticus. D: The posterior aspect of archistriaturn is one of the largest termination fields of DL, being densely packed with varicose PHAL+ axons (arrowheads). For abbreviations, see list. Scale bar = 150 bm.
stained section in Figure 12, arbitrarily separate the territo- ries of tracer deposit sites that gave distinct patterns of efferent projections. In summary, intrahippocampal, in- tratelencephalic, and subtelencephalic projections were typi- cal of the dorsolateral subdivision. The ventral subdivision gave rise to intrahippocampal and intratelencephalic projec- tions, but no long descending pathways were observed to arise from this area. The dorsomedial subdivision, which is defined for the first time in the present study, was found to innervate almost exclusively the ipsilateral hippocampal formation.
DISCUSSION The present study describes the efferent connectivity of
the hippocampal formation in the zebra finch brain. Our results largely agree with previous findings in the pigeon (Krayniak and Siegel, 1978; Casini et al., 19861, with some
notable differences. These earlier studies distinguished two main, although not very accurately defined, subregions, which are termed the hippocampus proper (Hp) and the area parahippocampalis (APH). In our experiments, we could differentiate three (dorsolateral, dorsomedial, and ventral) subdivisions with characteristic projection pat- terns.
Comparison to previous findings in birds Following PHAL injections into the DL
subregion, the observed projection pattern resembled the efferents ascribed to APH by Krayniak and Siegel (1978) and Casini et al. (1986). We found that DL injections gave rise to mostly ipsilateral projections, except for the weak innervation of the contralateral archistriatum. Similar to our observations, in the pigeon, efferents from APH (par- tially coextensive with DL as defined in our study) were distributed in CDL, in HD, and in the lateroventral hypo-
Similarities.

HIPPOCAMPAL PROJECTIONS IN THE SONGBIRD 205
Fig. 5. Photomicrographs illustrating passing and terminating PHAI-labelled DL fibres in the ipsilateral septal and ventrobasal forebrain regions. Arrowheads delineate the regions containing labelled axons. A SL is heavily enmeshed by varicose PHAJ-immunoreactive axons and terminals, whereas a proportion of labelled fibres crosses the ventral border to leave the nucleus. B: A thick bundle of DL efferents
surrounds the TSM and pulls farther ventral, partially terminating in FDB. C: The most medial and ventral portion of LPO is one of the main termination fields of DL comprising a dense meshwork of PHAL+ varicose fibres. D: Labelled DL axons travelling through the lateral septum enmesh PVT and partially terminate there. For abbreviations, see list. Scale bar = 150 wm.
thalamus (Casini et al., 1986), and further axons termi- nated in FDB, in the lateroventral septal regions, in the nucleus taeniae (Tn), and in the caudal aspect of the archistriatum (Krayniak and Siegel, 1978, Casini et al., 1986).
Injections into the ventral subdivision resulted in label- ling that was essentially identical to the previous findings (Krayniak and Siegel, 1978; Casini et al., 1986). Terminat- ing axons were observed in SM bilaterally and also in the contralateral HP, with the fibres crossing the midline via CPa.
We could distinguish a third DM subdivi- sion with almost exclusively internal efferents. Casini et al. (1986) did not mention projections similar to those of DM, whereas Krayniak and Siegel (1978) described a similar pattern arising from “the anterior third of the dorsomedial hippocampus” in the pigeon. However, those projections
Differences.
were more extensive than the fibre pattern observed in the present study, because these authors reported labelled fibres also in FDB and in the ventral septal areas stretching into the hypothalamus.
Conversely, in the zebra finch, DL injections resulted in more extensive projections at the diencephalomesence- phalic levels, particularly in the medial hypothalamus, in the dorsomedial and dorsolateral thalamic nuclei, and in AVT adjacent to the oculomotor nerve. None of these projections were mentioned in previous studies, although Casini et al. (1986) described afferents from AVT distrib- uted within HP. Nevertheless, unlike Casini et al. (19861, we did not observe labelled efferents crossing the midline around the median eminence.
The differences between our results and previous studies might be explained by the fact that we applied PHAL iontophoresis, which produces smaller deposits of tracer
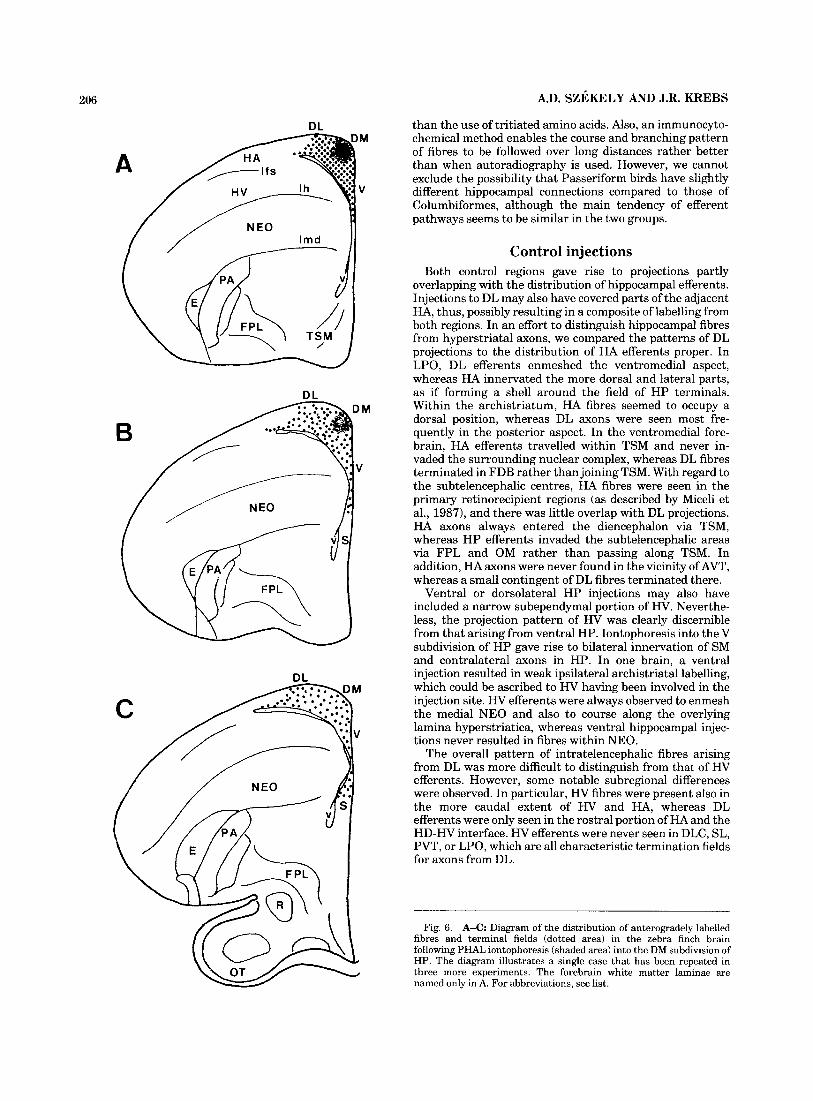
206 A.D. SZEKELY AND J.R. KREBS
B
C
DL than the use of tritiated amino acids. Also, an immunocyto- chemical method enables the course and branching pattern of fibres to be followed over long distances rather better than when autoradiography is used. However, we cannot exclude the possibility that Passeriform birds have slightly different hippocampal connections compared to those of Columbiformes, although the main tendency of efferent pathways seems to be similar in the two groups.
M
V
Control injections Both control regions gave rise to projections partly
overlapping with the distribution of hippocampal efferents. Injections to DL may also have covered parts of the adjacent HA, thus, possibly resulting in a composite of labelling from both regions. In an effort to distinguish hippocampal fibres from hyperstriatal axons, we compared the patterns of DL projections to the distribution of HA efferents proper. In LPO, DL efferents enmeshed the ventromedial aspect, whereas HA innervated the more dorsal and lateral parts, as if forming a shell around the field of HP terminals. Within the archistriatum, HA fibres seemed to occupy a dorsal position, whereas DL axons were seen most fre- quently in the posterior aspect. In the ventromedial fore- brain, HA efferents travelled within TSM and never in- vaded the surrounding nuclear complex, whereas DL fibres terminated in FDB rather than joining TSM. With regard to the subtelencephalic centres, HA fibres were seen in the primary retinorecipient regions (as described by Miceli et al., 1987), and there was little overlap with DL projections. HA axons always entered the diencephalon via TSM, whereas HP efferents invaded the subtelencephalic areas via FPL and OM rather than passing along TSM. In addition, HA axons were never found in the vicinity of AVT, whereas a small contingent of DL fibres terminated there.
Ventral or dorsolateral HP injections may also have included a narrow subependymal portion of H V . Neverthe- less, the projection pattern of HV was clearly discernible from that arising from ventral HP. Iontophoresis into the V subdivision of HP gave rise to bilateral innervation of SM and contralateral axons in HP. In one brain, a ventral injection resulted in weak ipsilateral archistriatal labelling, which could be ascribed to HV having been involved in the injection site. HV efferents were always observed to enmesh the medial NEO and also to course along the overlying lamina hyperstriatica, whereas ventral hippocampal injec- tions never resulted in fibres within NEO.
The overall pattern of intratelencephalic fibres arising from DL was more difficult to distinguish from that of HV efferents. However, some notable subregional differences were observed. In particular, HV fibres were present also in the more caudal extent of HV and HA, whereas DL efferents were only seen in the rostral portion of HA and the HD-HV interface. HV efferents were never seen in DLC, SL, PVT, or LPO, which are all characteristic termination fields for axons from DL.
DL
Fig. 6. A-C: Diagram of the distribution of anterogradely labelled fibres and terminal fields (dotted area) in the zebra finch brain following PHAL iontophoresis (shaded area) into the DM subdivision of HP. The diagram illustrates a single case that has been repeated in three more experiments. The forebrain white matter laminae are named only in A. For abbreviations, see list.

HIPPOCAMPAL PROJECTIONS IN THE SONGBIRD
B 207
Fig. 7. Photomicrographs of PHAL-labelled terminating fibres arising from DM. In the ipsilateral hemisphere, immunoreactive axons are found in the entire hippocampal complex and in the dorsal septum (A, delineated by arrowheads). Also, there is a thin band of PHAL+ axons (B, arrowheads) in the dorsal wall of the lateral ventricle outside HP. For abbreviations, see list. Scale bars = 200 pm in A, 150 pm in B.
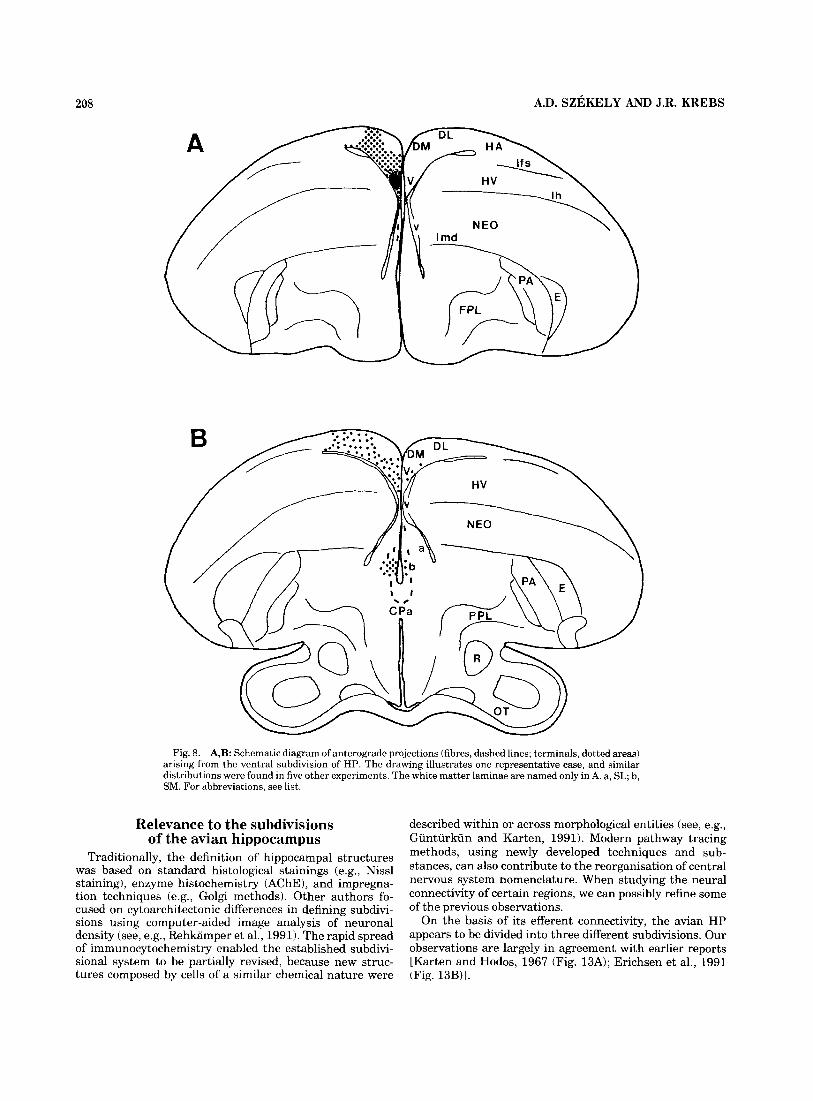
208 A.D. SZEKELY AND J.R. KREBS
Fig. 8. A,B: Schematic diagram of anterograde projections (fibres, dashed lines; terminals, dotted areas) arising from the ventral subdivision of HP. The drawing illustrates one representative case, and similar distributions were found in five other experiments. The white matter laminae are named only in A. a, SL; b, SM. For abbreviations, see list.
Relevance to the subdivisions of the avian hippocampus
Traditionally, the definition of hippocampal structures was based on standard histological stainings (e.g., Nissl staining), enzyme histochemistry (AChE), and impregna- tion techniques (e.g., Golgi methods). Other authors fo- cused on cytoarchitectonic differences in defining subdivi- sions using computer-aided image analysis of neuronal density (see, e.g., Rehkamper et al., 1991). The rapid spread of immunocytochemistry enabled the established subdivi- sional system to be partially revised, because new struc- tures composed by cells of a similar chemical nature were
described within or across morphological entities (see, e.g., Gunturkun and Karten, 1991). Modern pathway tracing methods, using newly developed techniques and sub- stances, can also contribute to the reorganisation of central nervous system nomenclature. When studying the neural connectivity of certain regions, we can possibly refine some of the previous observations.
On the basis of its efferent connectivity, the avian HP appears to be divided into three different subdivisions. Our observations are largely in agreement with earlier reports [Karten and Hodos, 1967 (Fig. 13A); Erichsen et al., 1991 (Fig. 13B)j.
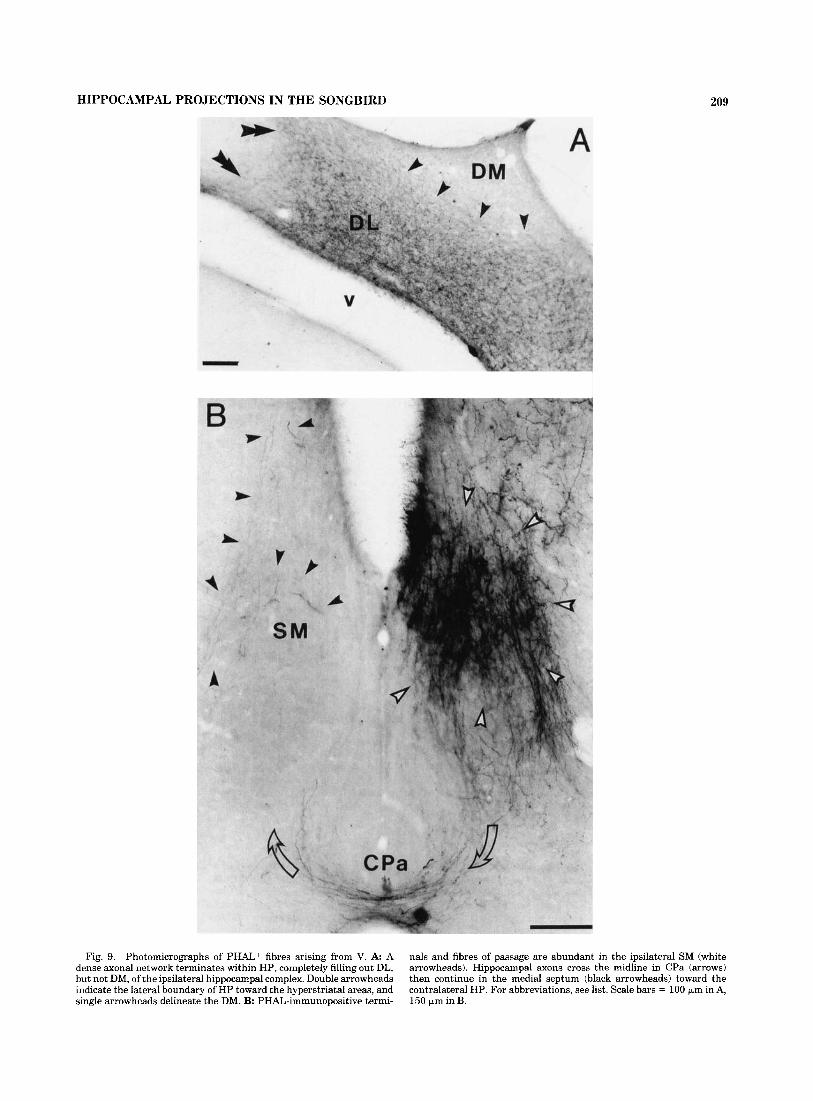
HIPPOCAMPAL PROJECTIONS IN THE SONGBIRD 209
Fig. 9. Photomicrographs of PHAL+ fibres arising from V. A A dense axonal network terminates within HP, completely filling out DL, but not DM, of the ipsilateral hippocampal complex. Double arrowheads indicate the lateral boundary of HP toward the hyperstriatal areas, and single arrowheads delineate the DM. B: PHAL-immunopositive termi-
nals and fibres of passage are abundant in the ipsilateral SM (white arrowheads). Hippocampal axons cross the midline in CPa (arrows) then continue in the medial septum (black arrowheads) toward the contralateral HP. For abbreviations, see list. Scale bars = 100 km in A, 150 pm in B.
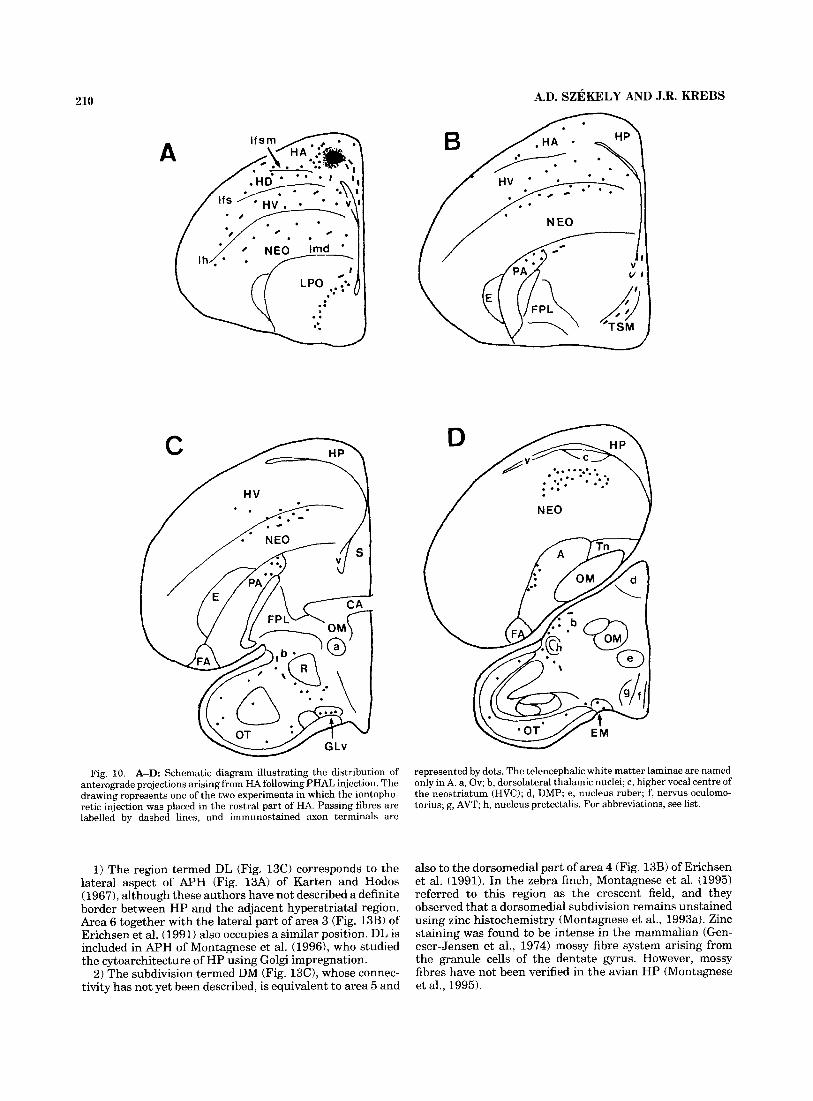
210
A
J.R. KREBS
Fig. 10. A-D: Schematic diagram illustrating the distribution of anterograde projections arisingfrom HA following PHAL injection. The drawing represents one of the two experiments in which the iontopho- retic injection was placed in the rostra1 part of HA. Passing fibres are labelled by dashed lines, and immunostained axon terminals are
represented by dots. The telencephalic white matter laminae are named only in A. a, Ov; b, dorsolateral thalamic nuclei; c, higher vocal centre of the neostriatum ( H V C ) ; d, DMP; e, nucleus ruber; f, nervus oculomo- torius; g, AVT; h, nucleus pretectalis. For abbreviations, see list.
1) The region termed DL (Fig. 13C) corresponds to the lateral aspect of APH (Fig. 13A) of Karten and Hodos (1967), although these authors have not described a definite border between HP and the adjacent hyperstriatal region. Area 6 together with the lateral part of area 3 (Fig. 13B) of Erichsen et al. (1991) also occupies a similar position. DL is included in APH of Montagnese et al. (1996), who studied the cytoarchitecture of HP using Golgi impregnation.
2) The subdivision termed DM (Fig. 13C), whose connec- tivity has not yet been described, is equivalent to area 5 and
also to the dorsomedial part of area 4 (Fig. 13B) of Erichsen et al. (1991). In the zebra finch, Montagnese et al. (1995) referred to this region as the crescent field, and they observed that a dorsomedial subdivision remains unstained using zinc histochemistry (Montagnese et al., 1993a). Zinc staining was found to be intense in the mammalian (Gen- eser-Jensen et al., 1974) mossy fibre system arising from the granule cells of the dentate gyrus. However, mossy fibres have not been verified in the avian HP (Montagnese et al., 1995).

HIPPOCAMPAL PROJECTIONS IN THE SONGBIRD 211
Fig. 11. A-D: Schematic diagram of the efferent connections from HV in the zebra finch telencephalon. Both injections resulted in a similar pattern of projections, one of which is shown in the drawing. Passing fibres are represented by dashed lines, and axon terminals are labelled by dots. The telencephalic white matter laminae are named only in A. For abbreviations, see list.
3) The V subdivision (Fig. 13C) corresponds to the area termed Hp (Fig. 13A) by Karten and Hodos (1967) and occupies the same position as areas 1, 2, and the medial aspect of area 3 (Fig. 13B) of Erichsen et al. (1991). In the terminology of Montagnese et al. (19951, V is named as the medial and lateral branches of the v-shaped neuronal plate (HCm and HC1, respectively), and it also contains the ventromedial part of their APH.
It is worth mentioning that our three subregions of HP can also be demonstrated by Calbindin immunocytochemis- try (Montagnese et al., 199313). However, the precise distri-
bution of immunopositive elements varies between differ- ent passeriform species.
Comparisons to other vertebrates The projections arising from the songbird HP, as de-
scribed in the present study, show substantial resemblance to the efferent connectivity of the mammalian hippocampal complex. Most importantly, there is a medial-to-lateral topographical organisation of hippocampal efferents in relation to the septa1 complex both in songbirds and in mammals. Namely, axons arising from the ventral portion

212 A.D. SZEKELY AND J.R. KREBS
Fig. 12. Low-power photomicrograph of a Nissl-stained section cut in the frontal plane from the zebra finch telencephalon. The lateral boundary of HP (arrowheads) can be distinguished by an increased cell density in the substance P field (SPD of the neighbouring hyperstriatal region, and the ventral, or septohippocampal, junction (arrowheads) is
marked clearly by a sharp decrease of cell density in the septum. The cell density is rather uniform in the DL, DM, and V subregions, the borders of which (dashed lines) were defined by the commonality of output rather than by visible cytoarchitectural differences. For abbre- viations, see list. Scale bar = 1 mm.
(i.e., towards the septohippocampal junction) enmesh the medial septum, whereas fibres from a more dorsal and lateral region (i.e., towards the hippocampal-hyperstriatal junction) innervate the lateral septum. Also, the individual subdivisions (DL, DM, and V) of the avian HP might be compared to portions of the mammalian HP. The DL region could be paralleled to the subiculum of mammals (Witter, 1989; Amaral, 19931, with its main projections to the basal ganglia, to the limbic archistriatum, to FDB and SL of the septa1 complex, and to the paraxial mesodiencephalic cen- tres. These findings support previous mapping studies of the pigeon hippocampal complex (Benowitz and Karten, 1976; Casini et al., 19861, although our DL region corre- sponds only to the lateral portion of their APH. In lizards, a corresponding region is found in the dorsal cortex (Martinez- Guijarro et al., 1990).
The V subdivision (or hippocampus proper) is likely to be the avian homologue of the Ammon’s horn of mammals (Benowitz and Karten, 1976) on the basis of its commis- sural projections upon the contralateral HP. This is also confirmed by Montagnese et al. (1995), who described hippocampal neurones of pyramidal morphology in this area using Golgi impregnation. The dorsomedial cortex of lizards, which contains bipyramidal neurones and receives zinc-positive fibres and commissural input, is considered to be homologous to the Ammon’s horn (Martinez-Guijarro et al., 1990).
It is particularly difficult to parallel the subdivision termed DM with a certain subregion of the mammalian hippocampal complex. In this respect, it is of crucial importance that our tracer injections were restricted to the DM field and that the efferent projections of this field
proved to be clearly distinct from those arising from the neighbouring regions. Previous connectivity studies (Krayn- iak and Siegel, 1978; Casini et al., 1986) have included the DM subdivision in their more extensive tracer injections; therefore, the significance of this subdivision has not been specifically pointed out. Based on its purely intrinsic connec- tivity, the DM region is a good candidate for an equivalent of the dentate gyrus (DG), and it partially coincides with the areas 4 and 5 of Erichsen et al. (19911, for which a possible homology with DG has been suggested. The medial cortex of reptiles, which is the site of origin for zinc-positive fibres (Olucha et al., 1988; Martinez-Guijarro et al., 19901, also occupies the dorsomedial corner of the pallium. However, its position, relative to the dorsomedial cortex, differs from the relative positions of DM and V in the bird, probably due to different trends in pallial development.
It has to be noted that our ventral injections also covered areas 3 and 4 of Erichsen et al. (1991), which are thought to correspond to the hilar and granule fields of DG on the basis of the chemical nature of cellular elements. Therefore, no conclusion can be drawn concerning the separate connectiv- ity of the latter regions; nevertheless, the pattern of efferent connections reported in the present study would suggest that a part of the DG-equivalent area may lie dorsomedial to its previously suggested site. This problem is further complicated by the uncertainty in the localisation of avian “entorhinal cortex,” which is the major source of the perforant pathway input in mammals. However, a quest for cover-to-cover homologies may not be essential for the understanding of the structural organisation of the avian hippocampal complex.

HIPPOCAMPAL PROJECTIONS IN THE SONGBIRD 213
thank Dr. Andras Csillag and Dr. Jonathan Erichsen for helpful discussions. The excellent technical assistance of Mrs. Jenny Corrigan and Mr. Alan Woodington is also acknowledged. This study was supported by the Royal Society, U.K.
A
C
Fig. 13. Schematic drawings of the hippocampal subdivisions as described earlier and in the present study. A The subdivisions of HP (redrawn after the pigeon atlas by Karten and Hodos, 1967). Anteropos- terior coordinate, 6.00. These authors did not define a lateral boundary of the complex towards HA or the exact separation between Hp and AF’H. B: The subdivisions of the pigeon H P established on the basis of the distribution of neurotransmitters and neuropeptides (redrawn after Erichsen e t al., 1991). Anteroposterior coordinate, 7.50. C: The three subdivisions of the zebra finch hippocampus as defined in the present study on the basis of projection patterns following PHAL iontophoresis. The drawing represents the central portion of H P at 1 m m anterior from lambda. For abbreviations, see list.
ACKNOWLEDGMENTS We thank Professor David Smith and Ms. Sarah Bacon
for providing laboratory and surgery facilities. We also
LITERATURE CITED Amaral, D.G. (1993) Emerging principles of hippocampal organization.
Current Opin. Neurobiol. 3:225-229. Ariens-Kappers, C.U., G.C. Huber, and E.C. Crosby (1936) The Comparative
Anatomy of the Nervous System of the Vertebrates, Including Man. New York: Macmillan.
Benowitz, L.I., and H.J. Karten (1976) The tractus infundibuli and other afferents to the parahippocampal region of the pigeon. Brain Res. 102,174-180.
Bingman, V.P., and G. Yates (1992) Hippocampal lesions impair navigational learning in experienced homing pigeons. Behav. Neurosci. 106:229-232.
Bingman, V.P., P. Ioale, G. Casini, and P. Bagnoli (1988) Hippocampal ablated homing pigeons show a persistent impairment in the time taken to return home. J. Comp. Physiol. A 163:559-563.
Campbell, C.B.G., and W. Hodos (1970) The concept of homology and the evolution of the nervous system. Brain Behav. Evol. 3:353-367.
Casini, G., V.P. Bingman, and P. Bagnoli (1986) Connections of the pigeon dorsomedial forebrain studied with WGA-HRP and 3H-proline. J. Comp. Neurol. 245:454470.
Craigie, E.H. (1930) Studies on the brain of the kiwi (Apteryx australis). J. Comp. Neurol. 56:223-357.
Csillag, A,, A.D. Szekely, and D.C. Davies (1994) The termination pattern of hyperstriatum ventrale efferents in the domestic chick archistriatum. An anterograde tracing study combined with Calbindin immunocytochemis- try and Golgi impregnation. J. Comp. Neurol. 348:394402.
Erichsen, J.T., V.P. Bingman, and J.R. Krebs (1991) The distribution of neuropeptides in the dorsomedial forebrain of the pigeon (Columba liuia): A basis for regional subdivisions. J. Comp. Neurol. 314:478492.
Geneser-Jensen, F.A., F.-M.S. Haug, and G. Danscher (1974) Distribution of heavy metals in the hippocampal region of the guinea pig: A light microscope study with Timm’s sulphid silver method. Z. Zellforsch. 147:441478.
Gerfen, C.R., and P.E. Sawchenko (1984) An anterograde neuroanatomical tracing method that shows the detailed morphology of neurons, their axons and terminals: Immunohistochemical localization of an axonally transported plant lectin, Phaseolus vulgaris leucoagglutinin (PHA-L). Brain Res. 290:219-238.
Giintiirkiin, O., and H.J. Karten (1991) An immunocytochemical analysis of the lateral geniculate complex in the pigeon (Columba liuia). J. Comp. Neurol. 31 4:72 1-749.
Hamassaki, D.E., and L.R.G. Britto (1990) Thalamic origin of neuropeptide y innervation of the accessory optic nucleus of the pigeon (Columbia liuia). Visual Neurosci. 5:149-159.
Kdlen, B. (1962) Embryogenesis of brain nuclei in the chick telencephalon. Ergebn. Anat. Enwickl-Gesch. 36.62-82.
Karten, H.J., and W. Hodos (1967) A Stereotaxic Atlas of the Brain of the Pigeon (Colurnba liuia). Baltimore: Johns Hopkins Press.
Krayniak, P.F., and A. Siege1 (1978) Efferent connections of the hippocam- pus and adjacent regions in the pigeon. Brain Behav. Evol. 15:372-388.
Krebs, J.R., J.T. Erichsen, and V.P. Bingman (1991) The distribution of neurotransmitters and neurotransmitter-related enzymes in the dorso- medial telencephalon of the pigeon (Columba liuia). J. Comp. Neurol. 314:467477.
Krebs, J.R., D.F. Sherry, S.D. Healy. V.H. Perry, and A.L. Vaccarino (1989) Hippocampal specialisation of food-storing birds. Proc. Natl. Acad. Sci. USA 86:1388-1392.
Kiihlenbeck, H. (1938) The ontogenetic development and phylogenetic significance of the cortex telencephali in the chick. J. Comp. Neurol. 69:273-301.
Martinez-Guijarro, F.J., E. Desfilis, and C. Lopez-Garcia (1990) Organiza- tion of the dorsomedial cortex in the lizard Podarcis hispanica. In W.K. Schwerdtferger and P. Germroth (eds): The Forebrain in Nonmammals. New Aspects of Structure and Development, Exp. Brain Res., Series 19. Berlin: Springer Verlag, pp. 77-92.
Miceli, D., J. Reperant, J. Villalobos, and L. Dionne (1987) Extratelence- phalic projections of the avian visual wulst. A quantitative autoradio- graphic study in the pigeon (Columba liuza). J. Hirnforsch. 28:45-57.

214 A.D. SZEKELY AND J.R. KREBS
Montagnese, C.M., F.A. Geneser, and J.R. Krebs (1993a) Histochemical distribution of zinc in the brain of the zebra finch (Taenopygia guttata). Anat. Embryol. 188:173-187.
Montagnese, C.M., J.R. Krebs, A.D. Szekely, and A. Csillag (199313) A subpopulation of large calbindin-like immunopositive neurones is pres- ent in the hippocampal formation in food-storing but not in nonstoring species of bird. Brain Res. 614t291-300.
Montagnese, C.M., J.R. Krebs, and G. Meyer (1996) The dorsomedial and dorsolateral forebrain of the zebra finch, Taenopygiaguttata: A Golgi study. Cell Tissue Res. 286:263-282.
Morris, R.G.M., R.F. Halliwell, and N. Bowery (1989) Synaptic plasticity and learning. 11. Do different kinds of plasticity underline different kinds of learning? Neuropsychologia 27:4 1-59.
O’Keefe, J., and L. Nadel (1978) The Hippocampus As a Cognitive Map. Oxford: Clarendon Press.
Olucha, F., F. Martinez-Garcia, L. Poch, W.K. Schwerdtferger, and C. Lopez-Garcia (1988) Projections from the medial cortex in the brain of lizards: Correlation of anterograde and retrograde transport of horserad- ish peroxidase with Timm staining. J. Comp. Neurol. 276:469-480.
Rehkamper, G., H.D. Frahm, and K. Zilles (1991) Quantitative development of brain and brain structures in birds (Galliformes and Passerifor- mes)compared to that in mammals (Insectivores and Primates). Brain Behav. Evol. 37:125-143.
Squire, L.R. (1992) Memoryand the hippocampus: A synthesis from findings with rats, monkeys and humans. Psychol. Rev. 99:195-231.
Stokes, T.M., C.M. Leonard, and F. Nottebohm (1974) The telencephalon, diencephalon and mesencephalon of the canary, Serinus canaria, in stereotaxic coordinates. J. Comp. Neurol. 156:337-374.
Szekely, A.D., A. Csillag, and T. Gores (1992) Neuropeptide Y innervation of retinorecipient layers of chick optic tectum. J. Neurocytol. 21:148-156.
Wieraszko, A., and G.F. Ball (19911 Long-term enhancement of synaptic responses in the songbird hippocampus. Brain Res. 538: 102-106.
Wieraszko, A., and G.F. Ball (1993) Long-term potentiation in the avian hippocampus does not require activation of the N-methyl-D-aspartate (NMDA) receptor. Synapse 13t173-178.
Witter, M.P. (1989) Connectivity of the rat hippocampus. In V. Chan-Palay and C. Kohler (eds): The Hippocampus-New Vistas. Neurology and Neurobiology, Vol. 52. New York: Alan R. Liss, Inc., pp. 53-69.





























