Embed Size (px)
Citation preview

Effects of Pseudomonas cepacia and cultural factors on the nodulation of Alnus rubra roots b y Frankia
SUSAN KNOWLTON' A N D JEFFREY 0. DAWSON Foresrr~ Depar~nerlr, Urziversiry qf Illirlois, Urbarlcr, IL. U.S.A. 61801
Received Deccmber I . 1982
KNOWLTON, S., and J . 0. DAWSON. 1983. Effects of Pserrclornorlc~s cepacia and cultural factors on the nodulation of AInlrs rltbra roots by Frankia. Can. J . Bot. 61: 2877-2882.
Infection of Alrllrs rrrbra Bong. roots by Frarlkia isolates was consistently promoted by Pserrdorr~orias cepacia "helper" bacteria under a variety of cultural conditions. Pseuclorr~or~ns cepaccia, while helpful, was ncver necessary in securing nodulation of aseptic A. rubrc~ by Frcrrlkia. Live P. cepacia cells were added with a Frankin isolate to aseptically grown A. rirbr~ secdlings on Hoagland's agar slants. This doubled, on averagc, the number of nodules formed by Frar~kia alonc when Frarikia isolate, age of Frankia inoculum, Frarlkia medium, calcium concentration of seedling substrate, phosphate concentration of seedling substrate, and pH of seedling substrate were varied. There was an apparent interaction between seedling substrate pH and P. cepacia in the promotion of nodulation. At pH levels from 4.0 to 6 .0 the infectivity of Frnnkia isolate Ar13 alone was greatly depressed compared with infectivity at the optimal pH for the growth of ArI3, around 7.0. However, at pH levels of 5 .0 .5 .5 , and 6.0 the ability of P. cepacia to promote nodulation of A. rubra seedlings by ArI3 was greater than at pH 7.0. This interaction may be due to the ability of P. cepcicia to neutralize and grow more rapidly in mildly acidic solutions in our experimental system, combined with the ability of P. cepacia to cause root-hair deformation, which has becn associated with the actinorhizal infection process.
KNOWLTON, S., et J . 0. DAWSON. 1983. Effects of Pseirdon1or~as cepacia and cultural factors on the nodulation of Alrlirs rirbra roots by Frankia. Can. J . Bot. 61: 2877-2882.
L'infection des racines de 1'Alnus rubra Bong. par des isolats de Frankia est systCmatiquement favorisCe par la bactCrie Pseudomonas cepacia, dans diverses conditions de culture. M&me s'il est utile, le P. cepacia n'est jamais nCcessaire pour assurer la nodulation de 1'A. rubra par Frankia dans des conditions aseptiques. Des cellules vivantes du P. cepacia ont CtC ajoutCes, avec un isolat de Frankia, i d e s plantules de I ' A . rirbra croissant de mani&re aseptiquedans des tubes inclines contenant de la gClose de Hoagland. En moyenne, ce traitement double le nombre de nodules formCs par le Frankia seul, lorsqu'on fait varier les facteurs suivants: l'isolat de Frarzkia, l ' ige de I'inoculum de Frarlkia, le milieu de culture du Frankia, la concentration en calcium et en phosphate du substrat de la plantule et le pH du substrat de la plantule. 11 semble y avoir une interaction entre le P. cepacia et le pH du substrat de la plantule dans l'augmentation de la nodulation. Aux pH compris entre 4,O et 6,0, l'isolat Ar13 de Frarlkia, inoculC seul, a une infectivitk beaucoup plus faible qu'au pH optimal pour la croissance de cet isolat, soit environ 7,O. Cependant, aux pH de 5,0, 5,5 et 6,0, le pouvoir du P. cepacia de favoriser la nodulation de plantules de 1'A. ritbra par l'isolat ArI3 est plus grand qu'h pH 7,O. Cette interaction pourrait &tre due h la capacitC du P. cepacia de neutraliser les solutions ICgkrement acides et de croitre plus rapidement dans ces solutions dans notre systkme experimental, de concert avec son pouvoir de provoquer des dCfomations des poils racinaires, lesquelles sont associCes au processus d'infection actinorhizienne.
[Traduit par le journal]
Introduction 7.0. It has been suggested that Frarzkia is more inhibited A number of environmental factors have been shown
to influence nodule initiation by Frankia, including pH, combined nitrogen, and presence of rhizosphere micro- organisms. Even though most actinorhizal plants are able to grow in a wide pH range, there is some indication that nodule formation is inhibited by low pH. Bond et a l . (1954) found that successful nodulation was achieved in only half of the inoculated Alnus and Myrica plants grown in water cultures maintained at pH 4.2. At the same pH, Hippophae plants did not nodulate, even though growth of nonnodulated plants supplied with nitrogen was unaffected by the acidic conditions. Nodule formation in these plants was maximum at pH
'present address: E. I. DuPont de Nemours & Co., Central Research and Development Department, Experimental Station, Wilmington, DE, U.S.A. 19898. .
at low pH than some host plants (Stewart 1966; Caiiizo et al . 1978).
In addition to the pH of the medium, high levels of combined nitrogen (10, 50, 100, 150mg NH4-N per litre) can suppress nodule formation in Alnus glutinosa (MacConnell and Bond 1957; Stewart and Bond 1971). Calcium and phosphate have been found to influence nodulation in leguminous plants (Lie 1974; Andrew 1978), although such influences have not been estab- lished for the actinorhizal association.
Work by Knowlton e t a l . (1980) indicated that microorganisms in the rhizosphere play a significant role in the infection of actinorhizal plants by Frankia. Under most conditions, aseptic Alrzus rubra seedlings inocu- lated with Frankia did not become nodulated unless other microorganisms were added to the seedling environment. Pseudomonas cepacia strains, in addition
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y T
exas
A&
M U
nive
rsity
on
11/1
2/14
For
pers
onal
use
onl
y.

2878 CAN. J. BOT. VOL. 61. 1983
to several rhizosphere isolates, promoted nodulation and elicited massive root-hair deformation typical of that observed in early stages of infection. It was suggested that "helpers" promote infection by causing root-hair deformation which is associated with Frarzkia infection. Such deformation could entrap the nonmotile actino- mycete ensuring intimate contact with the root hair and, thus, promote infection.
Although infection is promoted by Pseuclornonas strains, Frarzkia is, by itself, able to infect aseptic host plants. However, infection of aseptic plants in the past has sometimes been difficult to achieve (Knowlton et al . 1980). It has been suggested that cultural conditions under which the host plant (Lalonde et al. 1980) or the Frarzkia isolate (Knowlton et al . 1980) is grown may affect the infectivity of Frankia in the presence or absence of "helpers."
The objective of this research was to determine how a variety of cultural conditions influenced the ability of Pse~idomonas cepacia strain 85 to promote the infection of aseptic Alrzus rubra Bong. seedlings by Frankia isolates. By varying the cultural conditions, we hoped to elucidate specific ways in which "helpers" promote nodule formation in our experimental system.
Materials and methods Preparntiorl of rnicroorgnr~isrns
Pure cultures of the Frnnkin isolate ArI3 (Berry and Torrey 1979) were used to inoculate most test seedlings. In one experiment, Frnrlkia isolates CpIl (Callaham et nl. 1978), AvcI 1 (Baker et nl. 1979), and AcN 1 AG (Lalonde and Calvert 1979) were tested to determine whether differences occur among Frarzkia strains with respect to the effect of "helper" bacteria.
All Frnrlkia cultures were grown in the dark in 10 mL of a liquid medium in 18-mm-diameter culture tubes. For routine maintenance, Frnnkin was grown in a yeast extract medium described by Baker and Torrey (1979). When specified, Frnnkin was grown for experimental purposes in Q-Mod (Lalonde and Calvert 1979), a Tween 80 - casamino acids medium (Blom et al. 1980), or a defined succinate medium (Tjepkema etnl. 1980). In most cases, the cultures were grown for 30 days and then used to inoculate aseptic seedlings. In some experiments, various ages of ArI3 were tested to determine their infective capacity. These included cultures of ages 2, 4, and 6 weeks.
To prepare inocula, Frankin cultures were centrifuged at 2000rpm for 15 min and the pellet was washed twice and resuspended in sterile distilled water. The bacterial suspension was applied at a concentration of 0.005 mL packed cell volume per plant.
Growth rates of ArI3 were determined by measuring packed cell volumes using Kimax-brand, Bauer-Schenck centrifuge tubes (VWR Scientific Inc., Cat. No. 21074-005) calibrated in units of 0.004 mL. Each mean growth determination was made by centrifuging four samples separately for 15 min at 2000 rpm.
One-half of all experimental plants were also inoculated with Pseudomor~as cepacia strain 85 (PC85) described by
Stanier et al. (1966). The bacteria were grown in 10 mL of a liquid medium which contained (in grams per litre) yeast extract, 5.0; dextrose, 10.0: and casamino acids (acid hydrolyzed), 5.0. The cultures were grown in shaken culture for 16 hat room temperature and then centrifuged for 15 min at 2000rpm. The pellet was washed twice and resuspended in sterile distilled water. Two drops of a visibly turbid suspension were applied to each seedling root with a sterile Pasteur pipette. For those seedlings which did not receive the Pseudomor~as inoculum, two drops of sterile distilled water were applied instead.
Prepamtior1 of seedlings Seeds of Alnus rubrn were soaked in distilled water for
several hours and surface sterilized for 30 min in 30% hydro- gen peroxide with Ivory liquid detergent added as a surfactant. They were then rinsed five times in sterile distilled water and planted aseptically in Petri dishes which contained approxi- mately 20 mL of a 114-strength Hoagland's nitrogenfree solution (Hoagland and Arnon 1950) with 1% Noble agar, 1% dextrose, and 5 ppm nitrogen (NH4N03). Dextrose was added to reveal any contaminants on sterilized seeds to ensure the asepsis of seedlings used in experiments. Petri dishes sealed with Parafilm were placed in a growth chamber and maintained at 25°C for a 16-h day and at 20°C for an 8-h night. Light in the chamber was provided by both incandescent bulbs and cool-white fluorescent tubes.
After germination, uncontaminated seedlings were asepti- cally transferred to agar slants in sterile 25-mm-diameter culture tubes which contained 15 mL of a 1/4-strength Hoagland's nitrogenfree solution with 0.85% Noble agar and 5 ppm nitrogen. In some experiments, the calcium and phosphorous concentrations as well as the pH of the ~ o a g l a n d ' s solution were modified to determine their effect on nodule formation. Seedlings in tubes were capped with plastic test-tube caps (Bellco Kaputs). At the first true-leaf stage, seedlings were inoculated with a Frankin isolate alone or with Frnrlkin plus an additional inoculum of Pseudornonas cepncia strain 85.
All manipulations were made under a laminar-Row transfer hood to minimize contamination, and periodic, random tests were made to confirm the presence of only ArI3 or ArI3 plus PC85. This was done by culturing samples from seedling tubes in nutrient broth or agar. No contamination was revealed.
The individual growth rates of PC85 and ArI3 and their effects on substrate pH were determined on nutrient media and on 114-strength Hoagland's agar slants across a range of initial pH levels from 4.0 to 8.0. The nutrient media were a yeast extract broth (Baker and Torrey 1979) for Frnrlkin ArI3 and a broth containing 5 g / L Bacto-peptone and 3 g / L of Bacto-beef extract for PC85. The effects of seedlings alone on the pH of Hoagland's agar slants at initial pH's from 4.0 to 8.0 were also determined. The pH's were determined before and after organismal influences, using a combination pH electrode (Orion 91-35) inserted into nutrient broth, directly in root rhizospheres, or on agar surfaces, as well as 1 cm beneath the surface of agar slants.
Results A total of 1700 seedlings grown under a variety of
cultural conditions were tested for infection by Frankia
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y T
exas
A&
M U
nive
rsity
on
11/1
2/14
For
pers
onal
use
onl
y.

KNOWLTON AND DAWSON
TABLE I . A comparison of A. rubra nodulation patterns of four different Fratlkir~ isolates with or without "helper" bacteria
No. Total no. % seedlings Seedling inoculum plants nodules nodulated
Ar13 only Ar13 plus P. cepacia
Ar13 plus dead cells of P. cepacin
AvcIl only AvcIl plus P. cepacia
A C N I ~ ~ only A C N I ~ ~ plus P. cepacia
CpIl only CpIl plus P. cepacia
in this study. Half of these seedlings (850) were inoculated with Frankia isolate Ar13 and Pseudo- monas cepacia strain 85, while the other half received only an inoculum containing ArI3. Of those inoculated with ArI3 alone, only 304 (36%) formed nodules in a 4-week assay period. However, when the seedlings were inoculated with Ar13 plus PC85, 652 (77%) formed nodules in the same amount of time. The mean number of nodules formed per plant rose from 0.6 to 1.7 with the addition of the PC85 inoculum. The mean number of nodules formed per nodulated plant rose from 1.6 to 2.2 when "helpers" were added to seedling roots. The "helper" bacteria, in the presence of ArI3, consistently increased both the number of nodulated plants and the number of nodules formed on each seedling. There were no instances in which P. cepacia decreased nodulation by Frankia ArI3.
Nodules were observed approximately 10 days after inoculation with ArI3. Often they formed on the main root, either below or in rows between the regions of lateral roots.
Effective nitrogen fixation was evidenced by green foliar color and increased growth of nodulated seedlings compared with control seedlings. Control seedlings became yellow and grew little within the 4-week period prior to assay for nodulation. Seedlings inoculated with only PC85 and uninoculated controls never formed nodules.
In addition to ArI3, several other Frankia isolates were tested to determine if nodule formation was similarly increased with the addition of the Pseudo- monas strain (Table 1). In each case, nodule formation was increased when seedlings were inoculated with Pseudomonas cepacia plus CpI 1, AvcI 1, or AcN 1 Thus, no differences exist between these Frankia isolates with respect to effect of the "helper" bacteria. In addition, an inoculum was prepared by washing 3-month-old cells of Pseudomonas which had exhausted their medium and could not be recultured. When applied
to seedling roots, these cells did not cause an increase in nodulation as did the live cells.
Frankin isolate ArI3 exhibited various growth rates and morphologies when grown in different media. Frankia grown in the Tween 80 based medium grew most rapidly with a doubling time for packed cell volume of approximately 2 days, confirming results by Blom et al. (1980). The yeast extract medium produced the second fastest growth, while growth in Q-Mod ceased after 2-3 weeks.
Sporulation of ArI3 was immediate and greatest in the defined succinate medium. When grown in the yeast extract medium, Ar13 did not produce nearly as many sporangia as were formed in Q-Mod, defined succinate, or the Tween 80 - casamino acids medium. Even after 6 weeks, most colonies still contained primarily immature sporangia. In contrast, Ar13 grown in the Tween 80 - casamino acids medium produced very few sporangia after 2 weeks but produced many mature sporangia by the 4th week. In Q-Mod, ArI3 developed sporangia by the 2nd week and, after 4 weeks, many sporangia were broken apart releasing free-floating spores. This be- havior continued into the 6th week.
Because of these growth differences, we hypothe- sized that ArI3 infectivity might vary when Frankia was grown in different media or at various ages of subculture. In particular, we thought that sporulating cultures might be more infective than those with only hyphal growth. Frankia-induced nodule formation, however, with or without "helpers," could not be correlated with sporulation. The number of nodules that formed under our experimental conditions seemed to remain the same or decrease slightly when a Frankia inoculum in which the sporangia had matured and many spores had been dispersed was used.
The plant growth medium was also modified to determine the effect of different medium components on root nodule formation. Modifications included four phosphorus and three calcium concentrations in
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y T
exas
A&
M U
nive
rsity
on
11/1
2/14
For
pers
onal
use
onl
y.
2880 CAN. J . BOT. VOL. 61. 1983
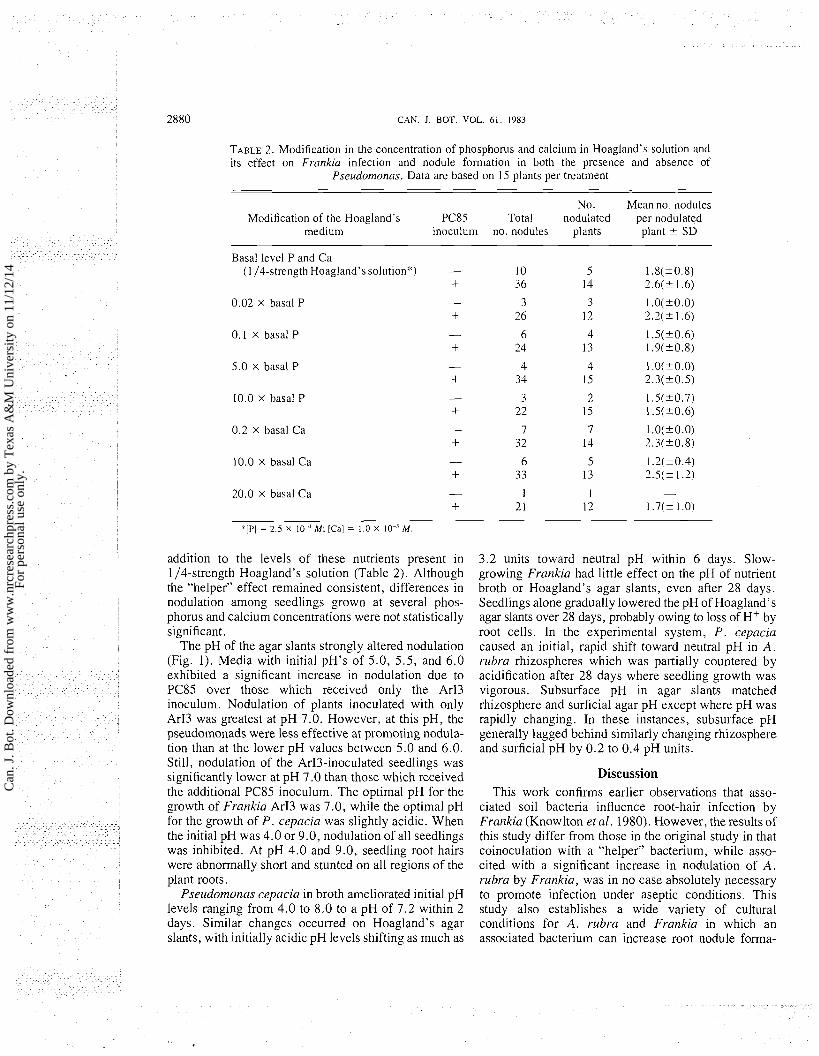
TABLE 2. Modification in the concentration of phosphorus and calcium in Hoagland's solution and its effect on Frankin infection and nodule formation in both the presence and absence of
Pseurlornonns. Data are based on 15 plants per treatment
No. Mean no. nodules Modification of the Hoagland's PC85 Total nodulated per nodulated
medium inoculum no. nodules plants plant * SD
Basal level P and Ca (I 14-strength Hoagland's solution:&) - I0 5 1.8(*0.8)
+ 36 14 2.6(* 1.6)
0.02 X basal P
0 . I x basal P
5.0 x basal P
10.0 x basal P
0.2 X basal Ca
10.0 X basal Ca
20.0 x basal Ca
'[PI = 2.5 x lo1 M; [Ca] = 1.0 X lo-%.
addition to the levels of these nutrients present in 114-strength Hoagland's solution (Table 2). Although the "helper" effect remained consistent, differences in nodulation among seedlings grown at several phos- phorus and calcium concentrations were not statistically significant.
The pH of the agar slants strongly altered nodulation (Fig. 1). Media with initial pH's of 5.0, 5.5, and 6.0 exhibited a significant increase in nodulation due to PC85 over those which received only the Ar13 inoculum. Nodulation of plants inoculated with only Ar13 was greatest at pH 7.0. However, at this pH, the pseudomonads were less effective at promoting nodula- tion than at the lower pH values between 5.0 and 6.0. Still, nodulation of the ArI3-inoculated seedlings was significantly lower at pH 7.0 than those which received the additional PC85 inoculum. The optimal pH for the growth of Frankia Ar13 was 7.0, while the optimal pH for the growth of P. cepacia was slightly acidic. When the initial pH was 4.0 or 9.0, nodulation of all seedlings was inhibited. At pH 4.0 and 9.0, seedling root hairs were abnormally short and stunted on all regions of the plant roots.
Pseudomonas cepacia in broth ameliorated initial pH levels ranging from 4.0 to 8.0 to a pH of 7.2 within 2 days. Similar changes occurred on Hoagland's agar slants, with initially acidic pH levels shifting as much as
3.2 units toward neutral pH within 6 days. Slow- growing Frnizkia had little effect on the pH of nutrient broth or Hoagland's agar slants, even after 28 days. Seedlings alone gradually lowered the pH of Hoagland's agar slants over 28 days, probably owing to loss of H+ by root cells. In the experimental system, P. cepacin caused an initial, rapid shift toward neutral pH in A. mbra rhizospheres which was partially countered by acidification after 28 days where seedling growth was vigorous. Subsurface pH in agar slants matched rhizosphere and surficial agar pH except where pH was rapidly changing. In these instances, subsurface pH generally lagged behind similarly changing rhizosphere and surficial pH by 0.2 to 0.4 pH units.
Discussion This work confirms earlier observations that asso-
ciated soil bacteria influence root-hair infection by Frankia (Knowlton et al . 1980). However, the results of this study differ from those in the original study in that coinoculation with a "helper" bacterium, while asso- cited with a significant increase in nodulation of A. rubra by Frankia, was in no case absolutely necessary to promote infection under aseptic conditions. This study also establishes a wide variety of cultural conditions for A. rubra and Frankia in which an associated bacterium can increase root nodule forma-
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y T
exas
A&
M U
nive
rsity
on
11/1
2/14
For
pers
onal
use
onl
y.

KNOWLTON AND DAWSON
1 ' 1 I I I 1
4.0 511 6.0 7.0 8.0 9.0
Initial pH of Hoagland's solution
FIG. I. The effect of initial pH on the infection of A . r1d1rn by Frnrzkin ArI3 in both the presence and absence of Pseudomorzns. Nodule number is the mean total for two replications of 30 plants per treatment bracketed by bars I SD in length.
tion. "Helpers" consistently increased the number of nodules formed through Frarzkia infection. Further- more, ArI3 was not the only Frankia isolate in which infection was increased in the presence of other rhizosphere bacteria. Infection by AvcI 1, CpI 1, and AcN lAG was similarly increased.
Although different media elicited different predomi- nant morphologies in Frankia, the medium in which Frankia was grown did not influence its infective capacity under the conditions of our experiments. Frankia spore formation has been associated with increased infectivity in A. gli(tirzosa (van Dijk 1978, 1979). H e found that crushed nodule suspensions prepared from spore-positive nodules were 100 to 1000 times more infective than suspensions prepared from spore-negative nodules. However, in this study, an increase in nodulation could not be correlated with Frankia spore formation in vitro.
In each medium tested, sporangia formation and maturation increased with the age of the culture, while infectivity declined after 4 weeks. Quispel and Burg- graaf (1980) found a similar age phenomenon with their cultures of an Alnus glutinosa endophyte unless supplementary lipids were added to the culture.
Substrate pH seems to play an important role in actinorhizal associations. At pH 4.0 and 9 .0 all root hairs were abnorn~al ly short and stunted and nodulation was inhibited in plants inoculated either with ArI3 or with ArI3 plus PC85. However, seedlings inoculated with only ArI3 and those with ArI3 plus Pse~(r1omorzas did not respond similarly at pH values between 5 . 0 and 8.0. Nodulation of seedlings inoculated with only ArI3 was highest around pH 7 .0 , which was also optimal for the growth of ArI3. This is in agreement with an earlier study (Bond et al. 1954) in which nodulation of Alnus, Myrica, and Hippophae species was highest at pH 7.0. However, at the same pH, pseudomonads were less effective at promoting nodulation than at the lower pH values of 5 .0 to 6.0. Pse~(dornorzas cepacia was most effective in promoting A. rubra infection by ArI3 in mildly acidic conditions, where it grew most rapidly and raised the pH of A. rubra rhizospheres to levels more conducive to Frankia growth and infection (pH 7.0). This suggests that the lower pH may inhibit infection by Frankia in our cultural system when other microorga- nisms which can neutralize acidic solutions are not present in the environment.
It is important that the total number of nodules produced on seedlings at pH 7 .0 was still lower than the number produced in the presence of Pseurlomorzns. Thus, pH may not be the only factor influencing infection. It is more likely that a combination of factors, in particular the neutralization of substrate pH combined with the massive root-hair deformation elicited by the rhizosphere bacteria, promote nodulation in our experimental system. Such interactions may play a role in natural ecosystems, although this remains to be demonstrated.
ANDREW, C. S. 1978. Legumes and acid soils. In Limitations and potentials for biological nitrogen fixation in the tropics. Edited by J. Dobereiner, R. H. Burris, and A. Hollaender. Plenum Publishing Corp., New York. pp. 135-160.
BAKER, D., and J . G. TORREY. 1979. The isolation and cultivation of actinomycetous root nodule endophytes. In Symbiotic nitrogen fixation in the management of temperate forests. Edited by J . Gordon. Oregon State University, Corvallis, OR. pp. 38-56.
BAKER, D., J . G. TORREY, and G. H. KIDD. 1979. Isolation by sucrose-density fractionation and cultivation in vitro of actinomycetes from nitrogen-fixing root nodules. Nature (London), 281: 76-78.
BERRY, A., and J . G. TORREY. 1979. Isolation and characterization in vivo and in vitro of an actinomycetous endophyte from Alnus rubra Bong. In Symbiotic nitrogen fixation in the management of temperate forests. Edited by J . Gordon. Oregon State University, Corvallis, OR. pp. 69-83.
BLOM, J . , W. ROELOFSEN, and A. D. L. AKKERMANS. 1980. Growth of Frankin AvcI 1 on media containing Tween 80 as C-source. FEMS Microbiol. Lett. 9: 13 1- 135.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y T
exas
A&
M U
nive
rsity
on
11/1
2/14
For
pers
onal
use
onl
y.

2882 CAN. J. BOT. VOL. 61. 1983
BOND, G. , W. W. FLETCHER, and T. P. FERGUSON. 1954. The development and function of the root nodules of Alnus, Myrica, and Hippophae. Plant Soil, 5: 309-323.
CALLAHAM, D. , P. DEL TREDICI, and J . G. TORREY. 1978. Isolation and cultivation in vitro of the actinomycete causing root nodulation in Comptonia. Science (Washing- ton, D.C.), 199: 899-902.
CARIZO, A, , C. MIGUEL, and C. RODRIGUEZ-BARRUECO. 1978. The effect of pH on nodulation and growth of Coriaria tnyrtifolia L. Plant Soil, 94: 195- 198.
HOAGLAND, D. R . , and D. I. ARNON. 1950. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. No. 347.
KNOWLTON, S. , A. BERRY, and J. G. TORREY. 1980. Evidence that associated soil bacteria may influence root hair infection of actinorhizal plants by Frankia. Can. J . Microbiol. 26: 97 1-977.
LALONDE, M., and H. E. CALVERT. 1979. Production of Frankia hyphae and spores as an infective inoculant for Alnus species. In Symbiotic nitrogen fixation in the management of temperate forests. Edited by J. Gordon. Oregon State University, Corvallis, OR. pp. 95-1 10.
LALONDE, M., H. E. CALVERT, and S. PINE. 1980. Isolation and use of Frankia strains in actinorhizae formation. In Current perspectives in nitrogen fixation. Australian Acad- emy of Science, Canberra.
LIE, T. A. 1974. Environmental effects on nodulation and symbiotic nitrogen fixation. In The biology of nitrogen fixation. Edited by A. Quispel. North Holland Publishing Co., Amsterdam. pp. 555-582.
MACCONNELL, J . T. , and G. BOND. 1957. A comparison of the effect of combined nitrogen on nodulation in non- legumes and legumes. Plant Soil, 8: 378-388.
QUISPEL, A., and A. J . P. BURGGRAAF. 1980. Frankia, the diazotrophic endophyte from actinorhiza's. In Current perspectives in nitrogen fixation. Australian Academy of Science, Canberra.
STANIER, R. Y., N. J . PALLERONI, andM. DOUDOROFF. 1966. The aerobic pseudomonads: a taxonomic study. J . Gen. Microbiol. 43: 159-27 1.
STEWART, W. D. P. 1966. Symbiotic nitrogen fixation. 111. Physiology. In Nitrogen fixation in plants. The Athlone Press, London. pp. 39- 155.
STEWART, W. D. P., and G. BOND. 1961. Effect of ammonium nitrogen on fixation of elemental nitrogen in Alnus and Myrica. Plant Soil, 14: 347-359.
TJEPKEMA, J . D., W. OMEROD, and J . G. TORREY. 1980. Vesicle formation and acetylene reduction activity in Frankia sp. Cpll cultured in defined nutrient media. Nature (London), 287: 633-635.
VAN DIJK, C. 1978. Spore formation and endophyte diversity in root nodules of Alnus glrctinosa (L.) Vill. New Phytol. 81: 601-615.
1979. Endophyte distribution in the soil. In Symbiotic nitrogen fixation in the management of temperate forests. Edited by J. Gordon. Oregon State University, Corvallis, OR. pp. 84-94.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y T
exas
A&
M U
nive
rsity
on
11/1
2/14
For
pers
onal
use
onl
y.





























