Embed Size (px)
Citation preview

Int. J. Mol. Sci. 2012, 13, 15679-15705; doi:10.3390/ijms131215679
International Journal of
Molecular Sciences ISSN 1422-0067
www.mdpi.com/journal/ijms
Article
Effects of Nickel, Chlorpyrifos and Their Mixture on the Dictyostelium discoideum Proteome
Lara Boatti, Elisa Robotti, Emilio Marengo, Aldo Viarengo and Francesco Marsano *
Department of Science & Technological Innovation (DiSIT), The University of Eastern Piedmont
Amedeo Avogadro, Alessandria, Novara, Vercelli, Viale Teresa Michel, 11-15121 Alessandria, Italy;
E-Mails: [email protected] (L.B.); [email protected] (E.R.);
[email protected] (E.M.); [email protected] (A.V.)
* Author to whom correspondence should be addressed; E-Mail: [email protected];
Tel.: +39-131-360-229; Fax: +39-131-360-243.
Received: 31 October 2012; in revised form: 13 November 2012 / Accepted: 15 November 2012 /
Published: 23 November 2012
Abstract: Mixtures of chemicals can have additive, synergistic or antagonistic interactions.
We investigated the effects of the exposure to nickel, the organophosphate insecticide
chlorpyrifos at effect concentrations (EC) of 25% and 50% and their binary mixture
(Ec25 + EC25) on Dictyostelium discoideum amoebae based on lysosomal membrane
stability (LMS). We treated D. discoideum with these compounds under controlled
laboratory conditions and evaluated the changes in protein levels using a two-dimensional
gel electrophoresis (2DE) proteomic approach. Nickel treatment at EC25 induced changes
in 14 protein spots, 12 of which were down-regulated. Treatment with nickel at EC50
resulted in changes in 15 spots, 10 of which were down-regulated. Treatment with
chlorpyrifos at EC25 induced changes in six spots, all of which were down-regulated;
treatment with chlorpyrifos at EC50 induced changes in 13 spots, five of which were
down-regulated. The mixture corresponding to EC25 of each compound induced changes
in 19 spots, 13 of which were down-regulated. The data together reveal that a different
protein expression signature exists for each treatment, and that only a few proteins are
modulated in multiple different treatments. For a simple binary mixture, the proteomic
response does not allow for the identification of each toxicant. The protein spots that
showed significant differences were identified by mass spectrometry, which revealed
modulations of proteins involved in metal detoxification, stress adaptation, the oxidative
stress response and other cellular processes.
OPEN ACCESS

Int. J. Mol. Sci. 2012, 13 15680
Keywords: Dictyostelium discoideum; toxicity; nickel; chlorpyrifos; proteomics;
mass spectrometry
1. Introduction
Understanding the modes of action of single compounds has been a challenging task. A single
compound can elicit agonistic actions on one biological pathway but antagonistic responses on another
pathway. Therefore, contradictory results can be obtained depending on the endpoint studied [1,2].
Moreover, environmental pollution rarely exists as a single contaminant, resulting in many chances for
multi-compound interactions. These interactions can strongly influence the effects of chemical
stressors on organisms. There are considerable gaps in knowledge about the in vivo effects of toxic
mixtures. Although a number of studies have investigated individual compounds of concern, exposure
studies that focus on chemical mixtures are largely missing from the literature.
A number of toxico-genomic studies have invested large efforts towards a better understanding of
mixture toxicity, but the complex interplay of toxicants among several biological pathways remains
difficult to resolve [3–5]. Nevertheless, single-endpoint mixture studies generate valuable information,
specifically when compound-related biomarkers are used (e.g., the loss of acetylcholinesterase activity
in relation to organophosphate and carbamate pesticide exposure) [6]. Acquiring in-depth information
about the interactions of compounds requires techniques that can provide an overview of several
endpoints and pathways simultaneously, such as proteomic approaches. Currently, only a limited
number of studies have applied proteomics techniques to the evaluation of mixture toxicity responses.
The application of recent “omics”-technologies in (eco)toxicology depends on the assumption that
all toxicologically relevant effects are accompanied by alterations in multiple genes or proteins. This
application is particularly promising because classical chemical analyses can only quantify substances
that are known in advance and are present in relatively high concentrations in the environment, but
they cannot be used to assess environmental and biological conditions with respect to chemical
bioavailability and the effects of toxicants in vivo. Therefore, the best “analyzer” of these complex
effects on organisms is the exposed organism itself [7]. Dictyostelium discoideum has been recognized
as a model organism that can live on humus and decaying leaves, so it is widely exposed to toxicants
in soils and could be a suitable biomonitoring organism for soil pollutants.
Nickel (Ni) is of great environmental concern; moreover, within a cell, its chemical form may be
altered, and it exerts long-term harmful effects [8]. It can be found naturally in metalliferous soils, but
it is often found in the environment as a result of industrial discharges from electroplating, smelting,
mining and refining operations, and other industrial emissions [9]. Compared to other divalent metals,
nickel toxicity has been studied mainly in water species [10–12]. A recent study monitored the
expression of genes involved in the specific molecular response to nickel in a nickel-tolerant fungus [13].
Although several toxicological studies have investigated the effects of Ni in various organisms, the
underlying molecular mechanisms by which Ni causes cellular damage are poorly understood [14].
Moreover, the cellular fate of nickel is interesting for phytoremediation applications, but the core

Int. J. Mol. Sci. 2012, 13 15681
mechanism of the molecules involved and the physiological conditions required, including soil
absorption, neutralization and toxicity, remain elusive [15].
Chlorpyrifos (CHP) is a broad-spectrum organophosphate pesticide with many urban and
agricultural pest control applications. CHP forms the active ingredient in several insecticides and is
among the most widely used insect control products [16]. Poisoning from CHP could affect the central
nervous system, cardiovascular system, and respiratory system; it also acts as a skin and eye irritant.
The major and well-known toxic effects of CHP involve the nervous system, as CHP is an inhibitor of
the enzyme acetylcholine esterase (AChE) [17]; other possible toxic effects on environmentally
important organisms are largely unknown. D. discoideum do not have a nervous system, and given the
evidence of the activity of CHP against AChE [18], this organism could be an appropriate model to
study the underlying effects of CHP and other neurotoxic compounds on the proteome under
conditions where other organisms would suffer from rapid mortality.
Numerous potential molecular targets for CHP (in addition to AChE) have been identified,
including effects on macromolecule synthesis (DNA, RNA, and proteins) and oxidative stress and
effects on microtubules [19–24].
Further information concerning the effects of nickel and CHP and their mixture on the cellular
proteome is crucial for understanding the molecular mechanisms involved in their toxic effects and
allows for further studies in biomonitoring, phytoremediation and toxicological sciences.
The aim of the present study is to characterize the toxic effects and the proteomic responses in the
model organism Dictyostelium discoideum to CHP and nickel. Additionally, we compared the
responses to the individual compounds with those of their binary equitoxic mixture using lysosomal
membrane stability (LMS). The final objective was to evaluate the variation in the protein expression
signature (PES) when cells are exposed to different concentrations of single compounds or a binary
equitoxic mixture.
2. Results
2.1. Experimental Design and Physiological Responses in D. discoideum Cells Exposed to Nickel,
CHP and Their Mixture
2.1.1. Mortality Rate
To evaluate the effect of the toxicants on cells, D. discoideum cells were exposed to increasing
concentrations of NiCl2 (0–10.0 mM) and the organophosphate pesticide CHP (0–50 μM) for 3 h under
laboratory conditions.
The first significant effects on mortality in D. discoideum (LOEC, low effect concentration) were
observed after treatment with 0.75 mM NiCl2 (Figure 1a), while a significant increase in this parameter
was induced after treatment with 10 μM CHP (Figure 1b).
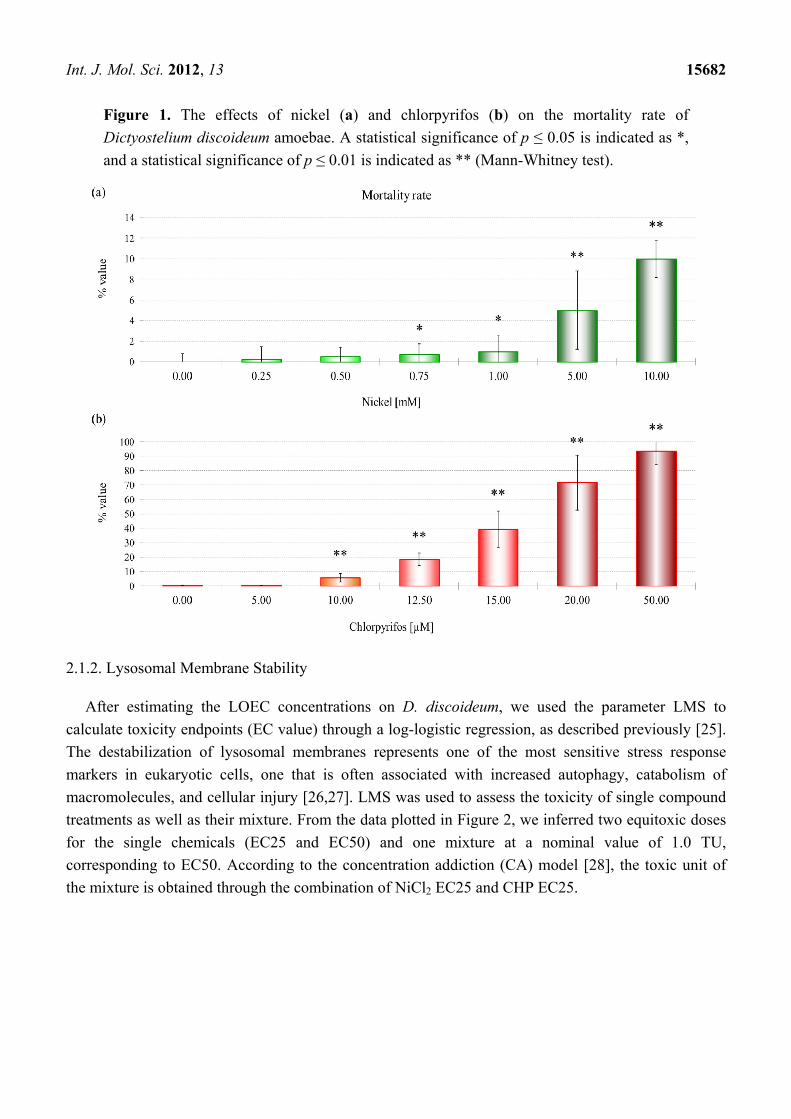
Int. J. Mol. Sci. 2012, 13 15682
Figure 1. The effects of nickel (a) and chlorpyrifos (b) on the mortality rate of
Dictyostelium discoideum amoebae. A statistical significance of p ≤ 0.05 is indicated as *,
and a statistical significance of p ≤ 0.01 is indicated as ** (Mann-Whitney test).
2.1.2. Lysosomal Membrane Stability
After estimating the LOEC concentrations on D. discoideum, we used the parameter LMS to
calculate toxicity endpoints (EC value) through a log-logistic regression, as described previously [25].
The destabilization of lysosomal membranes represents one of the most sensitive stress response
markers in eukaryotic cells, one that is often associated with increased autophagy, catabolism of
macromolecules, and cellular injury [26,27]. LMS was used to assess the toxicity of single compound
treatments as well as their mixture. From the data plotted in Figure 2, we inferred two equitoxic doses
for the single chemicals (EC25 and EC50) and one mixture at a nominal value of 1.0 TU,
corresponding to EC50. According to the concentration addiction (CA) model [28], the toxic unit of
the mixture is obtained through the combination of NiCl2 EC25 and CHP EC25.
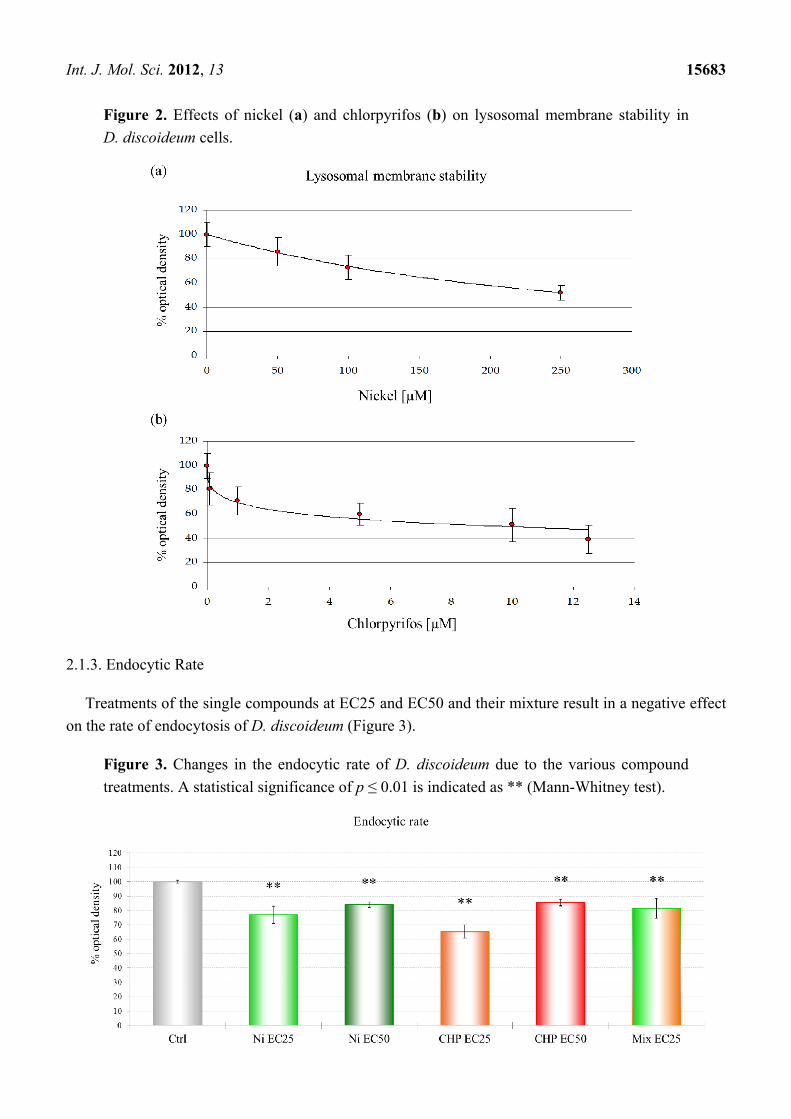
Int. J. Mol. Sci. 2012, 13 15683
Figure 2. Effects of nickel (a) and chlorpyrifos (b) on lysosomal membrane stability in
D. discoideum cells.
2.1.3. Endocytic Rate
Treatments of the single compounds at EC25 and EC50 and their mixture result in a negative effect
on the rate of endocytosis of D. discoideum (Figure 3).
Figure 3. Changes in the endocytic rate of D. discoideum due to the various compound
treatments. A statistical significance of p ≤ 0.01 is indicated as ** (Mann-Whitney test).

Int. J. Mol. Sci. 2012, 13 15684
2.2. Effects of the Toxicants on the D. discoideum Proteome
2.2.1. Nickel
Treatment with 114.6 μM NiCl2 (EC25) induced changes in 14 protein spots, two of which were
up-regulated (Figure 4, Table 1). One down-regulated spot contained a mixture of dihydropteridine
reductase, peroxiredoxin and an unknown protein, which prevents any quantitative evaluation and will
not be discussed further.
Figure 4. Two-dimensional gel electrophoresis (2-DE) gel portion of protein spots that are
differentially expressed after treatment with 114.6 μM NiCl2 (EC25). The graph shows the
ratio (% value) obtained by the PDQuest software (p ≤ 0.05, derived from the Mann-Whitney
U-test). Protein identification details can be found in Table 1.

Int. J. Mol. Sci. 2012, 13 15685
Table 1. Nickel EC25—identities of proteins with altered levels. Additional data can be
found in Table S1.
Spot n° Protein ID gi DDB Ratio
(% value)
0013 60S acidic ribosomal protein P2 gi|133059 DDB0191484 −45.3 0106 LIM-type zinc finger-containing protein gi|7107412 DDB0201567 −55.7 1108 Rho GDP-dissociation inhibitor gi|66803106 DDB0216235 −55.5 1115 Actin-related protein 2/3 complex, subunit 5 gi|66806101 DDB0191138 −41.5 3711 Glutamate-ammonia ligase gi|60469746 DDB0231551 −35.6 4106 Peroxiredoxin gi|28828367 DDB0231647 −31.2
5108 Dihydropteridine reductase gi|66823097 DDB0237752
−48.5 Hypothetical protein DDBDRAFT_0206195 gi|66815899 DDB0206195 Peroxiredoxin gi|66821043 DDB0231647
5412 S-adenosylmethionine synthetase gi|66803080 DDB0230070 83.8 5718 Succinate dehydrogenase gi|66813780 DDB0214886 61.2 6106 Similar to ribosomal protein gi|66821505 DDB0167610 −62.4 6414 S-adenosyl-methionine synthetase gi|60463691 DDB0230070 −35.4 7112 Hypothetical protein DDB_0233902 gi|60475381 DDB0233902 −47.7 7414 Elongation factor 1 β gi|10801150 DDB0191174 −24.0 7609 Cyclase-associated protein gi|66805581 DDB0191139 −49.5
NiCl2 treatment at EC50 (249.5 μM) showed a completely different PES with respect to the PES
observed after the EC25 treatment, and the proteins with altered levels after the EC25 treatment
returned to control levels after the EC50 treatment, showing a non-linear dose-response in protein
levels. After treatment with NiCl2 at EC50, we observed the differential expression of 15 spots, five of
which were up-regulated (Table 2, Figure 5).
Table 2. Nickel EC50—identities of proteins with altered levels. Additional data can be
found in Table S1.
Spot n° Protein ID gi DDB Ratio
(% value)
0124 Putative polypeptide-associated complex alpha subunit
gi|66812950 DDB0233328 −52.8
0125 Cytosolic glycoprotein FP21 gi|66826197 DDB0191107 −52.0
0526 RepC-binding protein A gi|4336714 DDB0191177
−50.1 Calreticulin gi|66810606 DDB0191384
0721 Glucose-regulated protein 94 gi|66814268 DDB0215015 −42.0
1125 Nascent polypeptide-associated complex alpha subunit
gi|66812950 DDB0233328 −18.1
2334 N-acyl-L-amino-acid amidohydrolase gi|66825457 DDB0201767
−61.1 26S proteasome non-ATPase regulatory subunit 6 gi|66826503 DDB0232985
2831 Hypothetical protein DDBDRAFT_0192196 gi|66799895 DDB0192196 −44.8 3423 Phosphoglycerate kinase gi|22711882 DDB0191349 −40.4
3721 Glutamate-ammonia ligase gi|66815105 DDB0231551
42.3 Prolyl oligopeptidase gi|4584573 DDB0185041

Int. J. Mol. Sci. 2012, 13 15686
Table 2. Cont.
Spot n° Protein ID gi DDB Ratio
(% value)
3826 Major vault protein gi|66825911 DDB0191259 −47.3 4529 Actin binding protein gi|66827977 DDB0191115 112.9 4732 AAA ATPase domain-containing protein gi|66805423 DDB0231600 127.5 6132 Peroxiredoxin gi|66808689 DDB0238212 109.2 6218 Putative epimerase gi|66820416 DDB0167407 41.9 6223 Phosphoribosyl pyrophosphate synthetase gi|66809487 DDB0237882 −32.1
Figure 5. 2-DE gel portion of protein spots that were differentially expressed after
treatment with 249.5 μM NiCl2 (EC50). The graph shows the ratio (% value) obtained
using the PDQuest software (p ≤ 0.05, derived from the Mann-Whitney U-test). Protein
identification details can be found in Table 2.
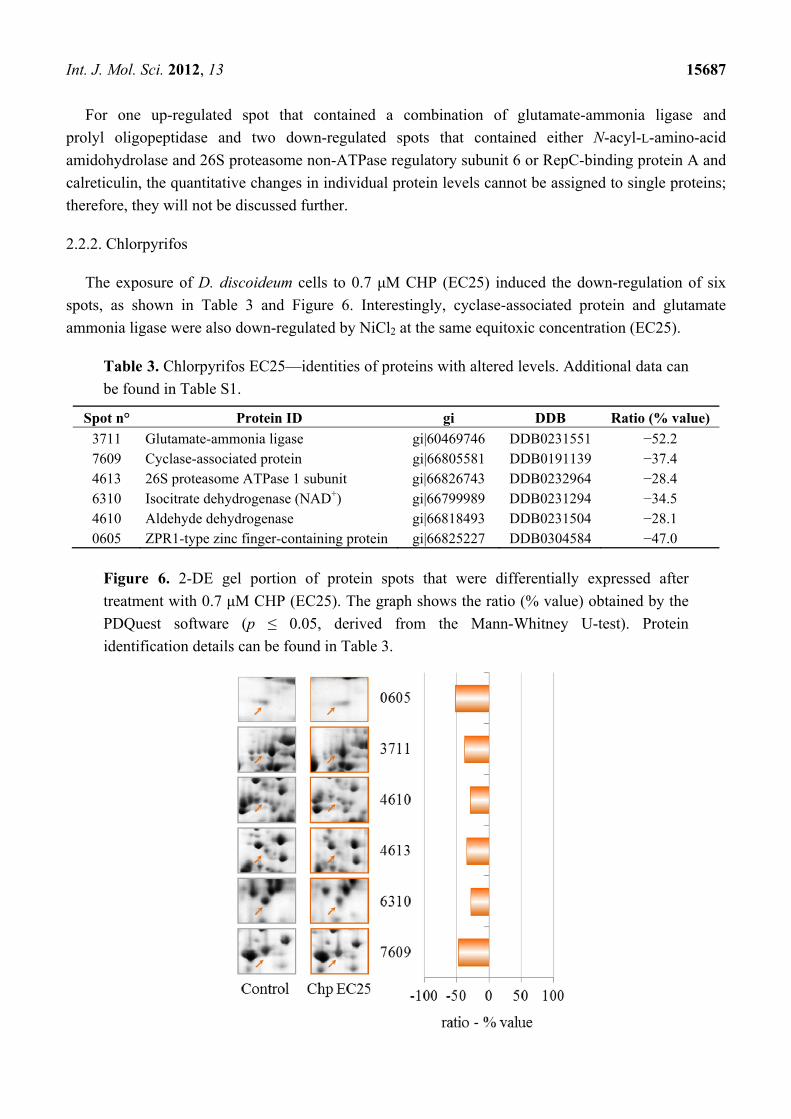
Int. J. Mol. Sci. 2012, 13 15687
For one up-regulated spot that contained a combination of glutamate-ammonia ligase and
prolyl oligopeptidase and two down-regulated spots that contained either N-acyl-L-amino-acid
amidohydrolase and 26S proteasome non-ATPase regulatory subunit 6 or RepC-binding protein A and
calreticulin, the quantitative changes in individual protein levels cannot be assigned to single proteins;
therefore, they will not be discussed further.
2.2.2. Chlorpyrifos
The exposure of D. discoideum cells to 0.7 μM CHP (EC25) induced the down-regulation of six
spots, as shown in Table 3 and Figure 6. Interestingly, cyclase-associated protein and glutamate
ammonia ligase were also down-regulated by NiCl2 at the same equitoxic concentration (EC25).
Table 3. Chlorpyrifos EC25—identities of proteins with altered levels. Additional data can
be found in Table S1.
Spot n° Protein ID gi DDB Ratio (% value)
3711 Glutamate-ammonia ligase gi|60469746 DDB0231551 −52.2 7609 Cyclase-associated protein gi|66805581 DDB0191139 −37.4 4613 26S proteasome ATPase 1 subunit gi|66826743 DDB0232964 −28.4 6310 Isocitrate dehydrogenase (NAD+) gi|66799989 DDB0231294 −34.5 4610 Aldehyde dehydrogenase gi|66818493 DDB0231504 −28.1 0605 ZPR1-type zinc finger-containing protein gi|66825227 DDB0304584 −47.0
Figure 6. 2-DE gel portion of protein spots that were differentially expressed after
treatment with 0.7 μM CHP (EC25). The graph shows the ratio (% value) obtained by the
PDQuest software (p ≤ 0.05, derived from the Mann-Whitney U-test). Protein
identification details can be found in Table 3.
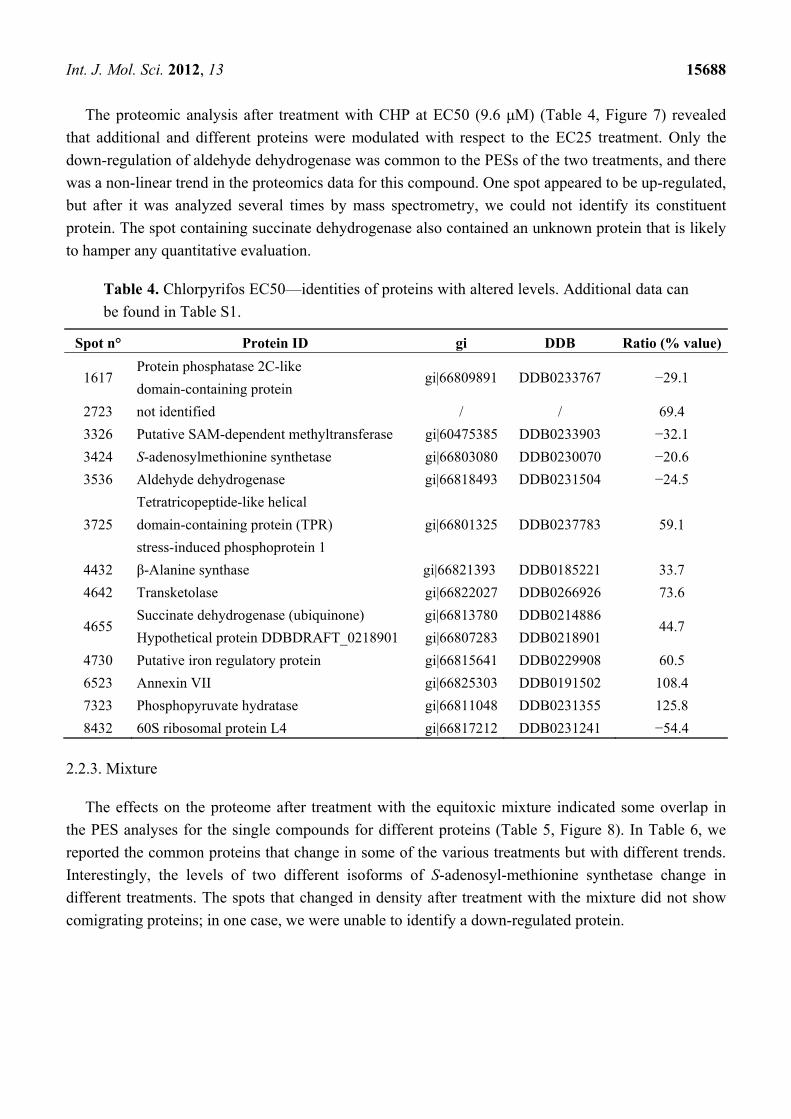
Int. J. Mol. Sci. 2012, 13 15688
The proteomic analysis after treatment with CHP at EC50 (9.6 μM) (Table 4, Figure 7) revealed
that additional and different proteins were modulated with respect to the EC25 treatment. Only the
down-regulation of aldehyde dehydrogenase was common to the PESs of the two treatments, and there
was a non-linear trend in the proteomics data for this compound. One spot appeared to be up-regulated,
but after it was analyzed several times by mass spectrometry, we could not identify its constituent
protein. The spot containing succinate dehydrogenase also contained an unknown protein that is likely
to hamper any quantitative evaluation.
Table 4. Chlorpyrifos EC50—identities of proteins with altered levels. Additional data can
be found in Table S1.
Spot n° Protein ID gi DDB Ratio (% value)
1617 Protein phosphatase 2C-like
domain-containing protein gi|66809891 DDB0233767 −29.1
2723 not identified / / 69.4
3326 Putative SAM-dependent methyltransferase gi|60475385 DDB0233903 −32.1
3424 S-adenosylmethionine synthetase gi|66803080 DDB0230070 −20.6
3536 Aldehyde dehydrogenase gi|66818493 DDB0231504 −24.5
3725
Tetratricopeptide-like helical
domain-containing protein (TPR)
stress-induced phosphoprotein 1
gi|66801325 DDB0237783 59.1
4432 β-Alanine synthase gi|66821393 DDB0185221 33.7
4642 Transketolase gi|66822027 DDB0266926 73.6
4655 Succinate dehydrogenase (ubiquinone) gi|66813780 DDB0214886
44.7 Hypothetical protein DDBDRAFT_0218901 gi|66807283 DDB0218901
4730 Putative iron regulatory protein gi|66815641 DDB0229908 60.5
6523 Annexin VII gi|66825303 DDB0191502 108.4
7323 Phosphopyruvate hydratase gi|66811048 DDB0231355 125.8
8432 60S ribosomal protein L4 gi|66817212 DDB0231241 −54.4
2.2.3. Mixture
The effects on the proteome after treatment with the equitoxic mixture indicated some overlap in
the PES analyses for the single compounds for different proteins (Table 5, Figure 8). In Table 6, we
reported the common proteins that change in some of the various treatments but with different trends.
Interestingly, the levels of two different isoforms of S-adenosyl-methionine synthetase change in
different treatments. The spots that changed in density after treatment with the mixture did not show
comigrating proteins; in one case, we were unable to identify a down-regulated protein.

Int. J. Mol. Sci. 2012, 13 15689
Figure 7. 2-DE gel portion of protein spots that were differentially expressed after
treatment with 9.6 μM CHP (EC50). The graph shows the ratio (% value) obtained by the
PDQuest software (p ≤ 0.05, derived from the Mann-Whitney U-test). Protein
identification details can be found in Table 4.
Int. J. Mol. Sci. 2012, 13 15690
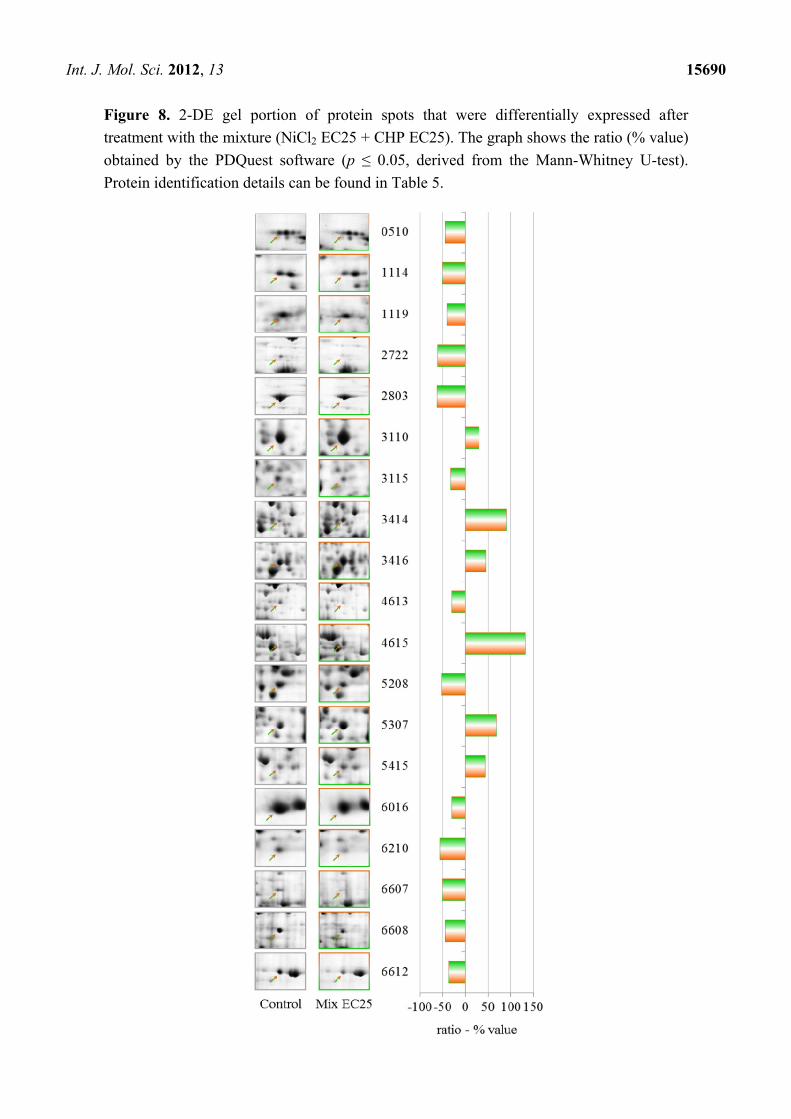
Figure 8. 2-DE gel portion of protein spots that were differentially expressed after
treatment with the mixture (NiCl2 EC25 + CHP EC25). The graph shows the ratio (% value)
obtained by the PDQuest software (p ≤ 0.05, derived from the Mann-Whitney U-test).
Protein identification details can be found in Table 5.

Int. J. Mol. Sci. 2012, 13 15691
Table 5. Mixture EC25—identities of proteins with altered levels. Additional data can be
found in Supplemental Table 1.
Spot n° Protein ID gi DDB Ratio (% value)
0510 Hypothetical protein DDBDRAFT_0187751 gi|66806459 DDB0187751 −44.4 1114 Pyruvate dehydrogenase E1 beta subunit gi|66818919 DDB0229442 −50.8 1119 Putative delta-24-sterol methyltransferase gi|60464861 DDB0237965 −41.5 2722 Clathrin heavy chain gi|66818048 DDB0185029 −60.9 2803 Myosin II heavy chain gi|134047850 DDB0191444 −62.2 3110 Aldehyde reductase gi|38637654 DDB0215363 30.1 3115 not identified / / −32.9 3414 Aldehyde dehydrogenase gi|66818493 DDB0231504 90.3 3416 Phosphoglycerate kinase gi|22711882 DDB0191349 44.8 4613 Glucosamine-6-phosphate isomerase gi|66808101 DDB0234127 −30.6 4615 Putative transport protein gi|166240434 DDB0235163 132.4 5208 Isocitrate dehydrogenase (NAD+) gi|60462281 DDB0231294 −52.7 5307 S-adenosyl-methionine synthetase gi|60463691 DDB0230070 68.0 5415 3-phosphoglycerate dehydrogenase gi|66813238 DDB0230052 43.3 6016 Actin binding protein gi|60468343 DDB0215335 −30.0 6210 G β-like protein gi|66820452 DDB0185122 −56.7 6607 Cyclase-associated protein gi|66805581 DDB0191139 −50.8 6608 ATP citrate synthase gi|66816585 DDB0235360 −44.7 6612 Aconitase, mitochondrial gi|60470010 DDB0230168 −37.5
Table 6. Common proteins with altered expression levels after the various treatments. The
variations in protein levels are expressed as percentages.
gi Protein ID Chlorpyrifos Nickel
MixtureEC25 EC50 EC25 EC50
gi|66805581 cyclase-associated protein −37.4 / −49.5 / −50.8 gi|60469746 glutamate-ammonia ligase −52.2 / −35.6 / / gi|66818493 aldehyde dehydrogenase −28.1 −24.5 / / 90.3 gi|60463691 S-adenosyl-methionine synthetase / / −35.4 / 68.0 gi|66803080 S-adenosylmethionine synthetase / −20.6 83.8 / / gi|22711882 phosphoglycerate kinase / / / −40.4 44.8 gi|60462281 isocitrate dehydrogenase (NAD+) −34.5 / / / −52.7
3. Discussion
The mortality and lysosomal membrane destabilization caused by CHP and NiCl2 treatments in
amoebae were evaluated to determine appropriate concentrations of both toxicants for further analyses.
To evaluate the effects on mortality, various concentrations of CHP (0–50 μM) and NiCl2 (0–10 mM)
were tested. LMS data were evaluated for CHP from 0 to 12.5 μM and for NiCl2 form 0 to 250 μM,
which allowed us to evaluate the sublethal treatments that induce toxic or adaptive effects. For the
single compounds, two experimental levels based on LMS data were tested and represented realistic
(NiCl2 114.6 μM; CHP 0.7 μM) and extreme (NiCl2 249.5 μM; CHP 9.6 μM) scenarios. Moreover, a

Int. J. Mol. Sci. 2012, 13 15692
mixture of the two substances at equitoxic concentrations was assessed to obtain information about
their interaction at the physiological and molecular levels.
The differentially expressed proteins were analyzed by Blast2GO for molecular processes and
biological functions and the results can be found as Supplemental File (Blast2GO Analysis).
3.1. Nickel
NiCl2 is not extremely toxic to D. discoideum, probably because of an energy-dependent nickel
export that exists in wild-type and mutant strains, but its role in heavy metal resistance has not been
confirmed [29]. The mortality rate and lysosomal membrane destabilization data (Figures 1 and 3)
revealed that the cytotoxicity increased with an increase in the NiCl2 concentration. Although the first
effects on mortality were observed only after treatment with 0.75 mM NiCl2, at the subcellular level,
LMS and proteomic data show effects at 10% of that concentration.
A concentration of 114.6 μM NiCl2, which corresponds to an EC25 for lysosomal membrane
destabilization, was chosen for subsequent experiments. The sublethal effects at this concentration
were evident, which might indicate that differential protein expression already occurs at this
concentration. Moreover, after the sublethal treatments, amoebae could maintain a normal phenotype,
which allowed us to extract sufficient amounts of proteins for 2DE experiments. A more toxic
concentration of 249.5 μM of NiCl2, corresponding to EC50 for LSM, was chosen for comparison.
The data in Table 1 show that clear effects on the proteome can be observed after treatment with
NiCl2 at EC25. Two proteins were up-regulated, succinate dehydrogenase and an isoform of
S-adenosylmethionine synthetase (gi|66803080). The activity of succinate dehydrogenase has been
previously observed to be inhibited by cadmium, with the possibility of interfering with energy
transport [30]. S-Adenosylmethionine synthetase (SAM-S) catalyzes the biosynthesis of
S-adenosylmethionine from Met and ATP. In plants, the expression of SAM-S is significantly altered
in response to salt and drought [31,32]. In yeast, the activity of this enzyme is strongly inhibited by
heavy metal ions (Cu2+ and Ag2+) and ethylenediaminetetraacetic acid (EDTA). Moreover, the
expression levels of SAM-S were markedly up-regulated in response to arsenate in rice roots [33].
Unexpectedly, a different isoform of S-adenosyl-methionine synthetase (gi|60463691) was
down-regulated.
Cyclase-associated proteins (CAPs) are widely distributed and highly conserved proteins that
regulate actin remodeling, vesicle trafficking and development in response to cellular signals [34]. In
D. discoideum, CAPs are essential for the functions of the endo-lysosomal system [35]. This protein
was down-regulated after treatment with CHP at EC25, and both NiCl2 and CHP EC25 treatments had
the clearest effects on endocytosis. The effects on endocytosis are probably linked to the
down-regulation of a subunit of the actin-related protein 2/3 (ARP2/3) complex, which plays a major
role in actin cytoskeletal regulation.
Glutamate-ammonia ligase was recently identified as a nickel-binding protein by mass
spectrometry, and its activity is inhibited by nickel [36].
Rho GDP dissociation inhibitor (RhoGDI) plays an essential role in the control of a variety of
cellular functions through its interactions with Rho family GTPases. RhoGDI is frequently

Int. J. Mol. Sci. 2012, 13 15693
over-expressed in human tumors and chemo-resistant cancer cell lines [37]. The observed
down-regulation due to NiCl2 could be of great interest and should be studied further.
The down-regulation of LIM-type zinc finger-containing protein after NiCl2 treatment is not
surprising because low concentrations of transition metal have been shown to interfere with DNA
transcription and repair. Various effects of heavy metals on zinc finger-containing proteins have been
identified, including the displacement of zinc by cadmium or cobalt, the formation of mixed
complexes, incomplete coordination of toxic metal ions, and the oxidation of cysteine residues within
the metal-binding domain [38].
The down-regulation of 60S acidic ribosomal protein P2, a ribosomal protein-like and elongation
factor 1β, suggests a modulation in protein synthesis caused by the presence of nickel.
After comparing the results of the cytotoxicity tests and protein expression data, it can be seen that
when the cells were treated with NiCl2 concentrations ranging from EC25 to EC50, the cytotoxic
effects were linear, but different altered proteins were identified (Tables 1 and 2). These data indicate
that NiCl2 toxicity cannot be quantitatively assessed by following the expression of single proteins in
such a wide concentration range.
NiCl2 at EC50 induces different patterns of up-regulation, including an increase in the level of
peroxiredoxin. The enzymes in this family consist of sulfhydryl-dependent peroxidases that exhibit
high reactivity with hydrogen peroxide and play major roles in peroxide defense, redox cell signaling
and metal responses [39,40]. Peroxiredoxin has recently been found to have an increased expression in
nickel-exposed workers [41]. Nickel causes increased levels of endogenous cellular hydrogen peroxide
and its short-lived reactive oxygen species [42–44].
Up-regulation of actin binding protein along with a modulation of the ARP2/3 complex and CAPs
substantiates the involvement of the cytoskeleton in the cellular response to nickel. This up-regulation
of cytoskeletal proteins in the EC50 treatment could explain the partial recovery of endocytic function.
Accordingly, the up-regulation of AAA-ATPase domain-containing protein and a putative epimerase
should be investigated further. There is little information about these two proteins, but a member of the
AAA-ATPase family has recently been described as a novel anti-apoptotic factor in lung
adenocarcinoma cells [45].
We observed the down-regulation of different proteins after treatment with NiCl2 at EC50. Major
vault protein (MVP), which is the main component of the vault complex, is over-expressed in many
multidrug-resistant cancer cell lines, suggesting a possible role for MVP in cell signaling and
survival [46,47]. Vault complexes are implicated in the cellular response to environmental toxins, but
we observed a reduction in the amount of this protein; thus, it is possible that NiCl2 does not activate
this response pathway.
Glucose-regulated protein 94 is a member of the endoplasmic reticulum (ER) chaperone family. In
D. discoideum, it is a constitutively expressed protein that is over-expressed in certain abnormal
cellular conditions, including the depletion of glucose and calcium, low oxygen levels and low
pH [48]. We observed down-regulation of this protein after treatment with NiCl2 at EC50.
We observed the down-regulation of phosphoglycerate kinase after treatment with NiCl2 at EC50.
The role of phosphoglycerate kinase in glycolysis is very important for the production of ATP through
substrate level phosphorylation. Regulation of this protein is thereby controlled by the energy level of
the cell.

Int. J. Mol. Sci. 2012, 13 15694
Phosphoribosyl pyrophosphate synthetase (PRPS), which was also down-regulated, plays a key role
in the biosynthesis of several amino acids and pyrimidines. PRPS is required to convert ribose
5-phosphate into phosphoribosyl pyrophosphate. The latter is a central compound for cellular
metabolism and may be considered as a link between carbon and nitrogen metabolism.
NiCl2 induces the down-regulation of the cytosolic glycoprotein FP21 of D. discoideum, which is
an orthologue of human and mouse S-phase kinase-associated protein 1. This protein is an essential
component of the SCF (SKP1-CUL1-F-box protein) ubiquitin ligase complex, which mediates the
ubiquitination of proteins involved in cell cycle progression, signal transduction and transcription.
We observed the down-regulation of the nascent polypeptide-associated complex alpha subunit.
This protein prevents short, recently synthesized ribosome-associated polypeptides from inappropriate
interactions with cytosolic proteins.
3.2. Chlorpyrifos
The data show a toxic effect of CHP in D. discoideum, an organism that possesses no nervous
system. CHP at 9.6 μM already induces the first lethal effects and 50% lysosomal membrane
destabilization. Moreover, this compound exerts negative effects on endocytosis.
The proteomic data reveal that CHP at EC25 induces the down-regulation but not up-regulation of
proteins. Cyclase-associated proteins are multifunctional proteins that contain several structural
domains. Recent studies have suggested important roles for these proteins in linking cell signaling with
actin polymerization and highlight their roles in vesicle trafficking and development, as discussed for
NiCl2 at EC25.
We observed the down-regulation of the 26S proteasome ATPase 1 subunit. The ubiquitin/26S
proteasome pathway is implicated in numerous diseases, including cancer and neurodegenerative
diseases [49]. The 26S proteasome regulatory complex has intrinsic ATPase activity that seems to play
an essential role in its function. Presumably, one role of the ATPase is to supply energy continuously
for the selective degradation of target proteins by the active 26S proteasome.
In bacteria, yeast and plants, glutamate-ammonia ligase (glutamine synthetase) expression is tightly
regulated by the metabolic status of the cell [50]. A previous study in freshwater crabs showed that
glutamine synthetase activity increased after long-term organophosphate exposure [51]. We observed
the down-regulation of this enzyme, indicating that this protein is involved in the response to acute
exposure to CHP in D. discoideum.
Isocitrate dehydrogenase (ICDH) catalyzes the oxidative decarboxylation of isocitrate. Eukaryotes
possess two distinct types of ICDH: an NAD-specific enzyme (EC 1.1.1.41), which is present only in
the mitochondria, and an NADP-specific enzyme (EC 1.1.1.42), which is found in both the cytoplasm
and the mitochondria. The reduced activity of liver mitochondrial ICDH has been correlated with
organophosphate toxicity [52,53]. There is also evidence correlating the activities of ICDH and the
maintenance of the cellular redox state, suggesting that ICDH plays an important role in cellular
defenses against oxidative stress [54].
Aldehyde dehydrogenase catalyzes the oxidation of various aldehydes, using NAD+ as a cofactor [55].
The enzyme is inhibited by dithiocarbamate fungicides and has previously been described as a suitable

Int. J. Mol. Sci. 2012, 13 15695
detector for these pesticides in the environment [56,57]. This enzyme is down-regulated by treatment
with CHP at EC25.
Increased transcription of ZPR1-type zinc finger-containing protein has been observed in potato
tuber periderm after heat stress [58]. The deficiency of the zinc finger protein ZPR1 causes
neuro-degeneration and defects in transcription in the mouse [59] and cell cycle progression in HeLa
cells [60]. After treatment with CHP at EC25, we observed the down-regulation of this protein.
The EC50 treatment induces variations in the levels of different protein spots; two of these spots,
which were up-regulated, contained two comigrating proteins. In one spot, we identified a
tetratricopeptide-like helical domain-containing protein and stress-induced phosphoprotein 1. The
other spot contained succinate dehydrogenase and an unknown protein. These spots, which contained
comigrating proteins, give no quantitative information because it is impossible to establish which
individual protein has altered levels.
The only protein that was down-regulated in both CHP treatments was aldehyde dehydrogenase,
which is expected because this protein was previously found to act as a detector of pesticides in the
environment [56,57].
Among the up-regulated proteins, there is a putative iron regulatory protein (IRP) that acts as a key
regulator of cellular iron homoeostasis as a result of the translational control of the expression of a
number of iron metabolism-related genes. It is likely that various agents and conditions affect IRP
activity, thereby modulating iron and oxygen radical levels in different circumstances [61].
Previous studies have demonstrated that the up-regulation of transketolase (TKT) might play a
critical role in maintaining a reducing environment in the mouse cornea [62]. Moreover, in the
cyanobacterium Anabaena, UV-B upregulated transketolase and protein down-regulation were
observed with Cd treatment compared to a control treatment [63]. The up-regulation that was observed
in the present study suggests a role for this protein in the response to CHP.
Another protein that was up-regulated by CHP at EC 50 is annexin VII. Annexins belong to a large
family of glycoproteins that bind both Ca2+ and negatively charged phospholipids and are considered
important components of calcium signaling pathways [64,65]. Various studies have shown that
annexins play important roles in the cell during the response to oxidative stress [66–68]. The
peroxidase activity that is exhibited by annexins is further exemplified by studies in plants [69,70].
β-alanine synthase is an enzyme that catalyzes the final step of pyrimidine catabolism and is
required for the biosynthesis of β-alanine in animals. In mammals, β-alanine is a natural antioxidant
that is produced by the degradation of uracil or carnosin, which is a polypeptide that is formed by
β-alanine and L-lysine. Moreover, β-alanine is necessary for the production of CoA, which is required
for fatty acid metabolism. The treatment of D. discoideum cells with 9.6 μM CHP induced an increase
in ROS production (Figure S1) with a simultaneous up-regulation of β-alanine synthase, which is the
enzyme required for the production of β-alanine. Moreover, as described previously, we found the
down-regulation of aldehyde dehydrogenase and S-adenosylmethionine synthetase (SAMS), both of
which are needed in other mechanisms of β-alanine metabolism. We could hypothesize that the
attempt by D. discoideum to modulate a specific pathway for β-alanine production is probably an
alternative defense mechanism in response to the oxidative stress that is induced by pesticide treatment.
The expression levels of S-adenosylmethionine synthetase (SAMS) and a SAM-dependent
methyltransferase are markedly up-regulated in response to arsenate in plants [33]. Conversely, we

Int. J. Mol. Sci. 2012, 13 15696
observed the down-regulation of these proteins after CHP treatment at EC50. This difference requires
further investigation.
We observed an increase in phosphopyruvate hydratase (also known as enolase), which is a
metalloenzyme that is required for the penultimate step of glycolysis. Increased expression of enolase
has recently been reported in stressed plants [71].
The 60S ribosomal protein L4 is a component of the large ribosomal subunit. Yang et al. (2005)
have show that this protein is required for rRNA processing in mammalian cells, and that decreasing
the level of this peptide inhibits the production of several rRNAs [72].
The exact physiological role of protein phosphatase 2C-like domain containing protein, which we
found to be down-regulated after CHP at EC50 treatment, is unclear.
3.3. Mixture
The results have demonstrated that each treatment results in different PESs, and the mixture gives a
unique PES, making it difficult to apply PES to the toxicological evaluation of complex environmental
matrixes. However, the data show that some proteins that have altered levels after the single toxicant
treatments are involved in the cellular response to the mixture. The prominent protein in this class is
cyclase-associated protein, which is down-regulated in both single treatments at EC25. Aldehyde
dehydrogenase is down-regulated after treatment with CHP at EC25 but up-regulated by treatment
with the mixture, along with aldehyde reductase, indicating an attempt by the cell to counteract the
formation of lipid peroxide-derived aldehydes that are produced downstream of ROS [73].
The down-regulation of G β-like protein indicates an involvement of G proteins in the response to
the mixture. G proteins represent one of the most prevalent signaling systems in mammalian cells,
regulating systems as diverse as sensory perception, cell growth and hormonal regulation.
Phosphoglycerate kinase catalyzes the reversible transfer of a phosphate group from ATP to
3-phosphoglycerate, producing ADP and 1,3-bisphosphoglycerate. This protein is down-regulated by
CHP at EC50 but is up-regulated by the mixture.
S-adenosyl-methionine synthetase levels are altered by both NiCl2 EC25 and CHP EC50, and the
protein was strongly up-regulated after treatment with the mixture; a putative δ-24-sterol
methyltransferase that uses S-adenosyl-methionine as substrate was also up-regulated by the mixture.
Different cytoskeletal proteins, including actin binding protein, myosin II heavy chain, and clathrin
heavy chain, were down-regulated by the mixtures, indicating the involvement of the cytoskeleton in
the response to toxic substances, despite the fact that treatment with the mixture showed a partial
recovery of the endocytic rate.
Isocitrate dehydrogenase (NAD+) levels were down-regulated by the mixture and by CHP at EC25.
Aconitase (mitochondrial) has been established as a sensitive target of ROS, and the levels of this
enzyme were down-regulated after treatment with the mixture.
ATP citrate synthase catalyzes the first reaction in the Krebs cycle, the conversion of oxaloacetate
and acetyl-coenzyme A into citrate and coenzyme A. This reaction is important for energy generation
and for carbon assimilation and was down-regulated by the mixture.
We observed the up-regulation of 3-phosphoglycerate dehydrogenase, which catalyzes the
transition of 3-phosphoglycerate to 3-phosphohydroxypyruvate. This reaction is the first and

Int. J. Mol. Sci. 2012, 13 15697
rate-limiting step in the phosphorylated pathway of serine biosynthesis; serine is a constituent of
proteins and a precursor of several metabolites, including cysteine, glycine and choline.
We observed the down-regulation of the pyruvate dehydrogenase E1 beta subunit. The pyruvate
dehydrogenase complex is a nuclear-encoded mitochondrial multi-enzyme complex that catalyzes the
overall conversion of pyruvate to acetyl-CoA and CO2 and provides a primary link between glycolysis
and the tricarboxylic acid (TCA) cycle.
Another down-regulated protein was glucosamine-6-phosphate isomerase, which catalyzes the
conversion of D-glucosamine 6-phosphate (GlcN6P) to D-fructose 6-phosphate and ammonia. This
reaction is the last step in the pathway for N-acetylglucosamine utilization in bacteria or fungi.
Another interesting protein that was strongly up-regulated by the mixture was a putative transport
protein that should be studied further to clarify its involvement in the response to the toxic mixture.
Based on these results, it is possible to investigate the molecular mechanisms of cytotoxicity on a
proteomic scale by using a proteomic approach, despite the fact that each treatment has a different
PES. In particular, this approach, beyond the simultaneous quantitative observation of proteins that are
known to be stress responsive in a single experiment, allows us to identify novel proteins that have
never previously been observed to change in level after toxic treatments.
4. Experimental Section
4.1. Dictyostelium Growth and Treatments
The social amoeba Dictyostelium discoideum (strain AX-2) was grown axenically in AX-2 medium
as described previously [74]. The cells were grown under laboratory conditions on a Gallenkamp
orbital incubator (Sanyo Gallenkamp, Plc. Loughborough, UK) at 150 rpm and at 21 °C [75]. To
synchronize the cell cycle, the amoebae in logarithmic phase (2–4 × 106 cells/mL) were stored
overnight at 4 °C [76]. The cell treatment was performed in diluted medium for 6 h as described
previously [77]. Chemicals of analytical grade were purchased from Sigma-Aldrich if not
stated otherwise.
Nickel was administered as a chloride salt (NiCl2) from a concentrated stock solution (1000×), CHP
was diluted in dimethylsulphoxide (DMSO) and added at the desired concentration from a
concentrated stock solution (1000×). To exclude any effects of the DMSO vehicle, the same solvent
volume was added to reference cells and NiCl2-exposed cells at the same dilution.
To evaluate the first significant effects on mortality in D. discoideum (LOEC), the cells were
exposed to increasing doses of NiCl2 (0–10.0 mM) and CHP (0–50 μM).
After estimating the LOEC concentrations on the D. discoideum cells, we exposed the cells to
increasing doses of NiCl2 (0–250 μM) and CHP (0–12.5 μM) to calculate toxicity endpoints (EC)
using the parameter of lysosomal membrane stability (LMS).
For the proteomics studies and endocytic rate studies, D. discoideum cells were treated with two
different equitoxic doses of the single chemicals—0.5 (EC25) and 1.0 (EC50) TU, and one mixture at
a nominal 1.0 TU. According to the CA model [28], these toxic levels are obtained through the
combination of 2 × 0.5 TU. The cells were exposed to 114.6 and 249.5 μM NiCl2 and 0.7 and 9.6 μM
CHP, which correspond to EC25 and EC50, respectively.

Int. J. Mol. Sci. 2012, 13 15698
4.2. Mortality Rate, Lysosomal Membrane Stability and Endocytic Rate
Biomarker assays were performed as described previously [77]. Briefly, the mortality rate was
assessed using the DNA-binding dye SYBR Green™ (Sigma-Aldrich, Milan, Italy), and observations
of the cells were performed at 460× magnification on a Zeiss Axiovert 100M. For the evaluation of
LMS, the retention times of neutral red (NRRT) dye in the lysosomes after exposure to several doses
of pollutants were monitored.
Slides were observed at 1449× magnification in an inverted camera-equipped microscope
(Zeiss Axiovert 100 M) that was controlled remotely via AxioVision software (Zeiss, Germany).
For the endocytic rate, an aliquot of 40 μL cell suspension was incubated for 15 min with
Fluorescent Bioparticles (Escherichia coli K-12 strain; Abs/Em maxim = 505/513 nm; Molecular
Probes, Eugene, US). After incubation, D. discoideum cells were fixed in 3.7% paraformaldehyde and
observed with a microscope at 1.449× magnification.
The images that were obtained from LMS and endocytosis were analyzed using an image analysis
system (Scion Image freeware v 1.62) (Scion Corp., Frederick, MD, USA) that allowed the
quantification of lysosomal Neutral Red (NR) and the quantification of fluorescent bacteria in the cells.
The data were expressed as a percentage of the optical density with respect to the control condition.
Data analysis was conducted using Winks SDA (ver. 6.0.5) statistics software (Texasoft, Duncanville,
TX, USA). A non-parametric Mann-Whitney test was used to compare the control and treated samples.
A statistical significance of p ≤ 0.05 is indicated as *, and a statistical significance of p ≤ 0.01 is
indicated as **.
4.3. Cell Lysis and Protein Extraction
After treatments with several doses of nickel, CHP and their mixture, D. discoideum at a culture
concentration of 45 × 106 cells were washed twice with PAS, centrifuged at 500× g, precipitated and
lysed in buffer (0.3% SDS, 200 mM DTT, 30 mM Tris, Protease Inhibitor Cocktail, P2714,
Sigma-Aldrich, St Louis, MO, USAS) as described previously [74]. Protein precipitation was
performed in 80% (v/v) cold acetone in MilliQ water and centrifuged at 5000× g for 10 min.
After discarding the supernatant, the protein pellet was solubilized in a typical buffer for
first-dimension isoelectric focusing (IEF) (7 M urea, 2 M thiourea, 4% w/v CHAPS, and 50 mM DTT).
4.4. 2-DE Gel Electrophoresis
The proteins were extracted from seven biological replicates for each treatment and were separated
by 2-DE gel electrophoresis (2-DE) according to a method described previously [74].
For IEF, immobilized strip gels (pH 3–10 linear, 13 cm) (GE Healthcare Europe GmbH) were used
to separate 800 μg of proteins. IEF was conducted at 20 °C on an IPGphor unit (GE Healthcare
Europe GmbH).
For the second dimension, the focused proteins on the strips were reduced in an SDS equilibration
buffer (50 mM Tris-HCl pH 8.8, 6 M Urea, 30% (v/v) glycerol, 2% (w/v) SDS) containing
10 mg/mL DTT for 15 min and subsequently alkylated with 25 mg/mL iodoacetamide for 10 min.

Int. J. Mol. Sci. 2012, 13 15699
SDS-PAGE on 12% gels was performed in a Protean II XL 2-D cell (Bio-Rad Laboratories). The
gels were fixed in a solution containing 40% (v/v) methanol and 10% (v/v) acetic acid in MilliQ water
and stained with Colloidal Coomassie brilliant blue G250 (Bio-Rad Laboratories) according to
Neuhoff et al. [78]. Excess stain was removed by rinsing the gels with a solution containing methanol
and acetic acid in MilliQ water.
4.5. Image Analysis
The gels (3–10 pH) were scanned using a densitometer GS-710 (Bio-Rad Laboratories) and
analyzed using PDQuestTM Software version 7.3.1 (Bio-Rad Laboratories), following the typical
analysis workflow. The PDQuest matching summary is provided in Supplemental Table 2, and a
typical 2DE map is presented in Supplemental Figure 2.
The relative abundance of each spot in the control and treated gels, obtained from seven biological
replicates for each treatment, was performed by a software program using the Mann-Whitney U-test,
and the differences were considered significant at p ≤ 0.05.
4.6. Automated ProteinSspot Picking and In-Gel Digestion
The differentially expressed protein spots were transferred from gels to 96-well plates using a
Proteome Works Spot Cutter (Bio-Rad Laboratories) (Richmond, CA, USA) that was equipped with a
1 mm diameter needle. For high-throughput identification by nanoHPLC-ESI-QTOF, the proteins were
digested by trypsin (Roche Diagnostics, Monza, Italy) and extracted with Multiprobe II (Perkin Elmer,
Wellesley, MA, USA) as described previously [74].
4.7. Protein Identification by Nano-LC/Q-TOF MS/MS
The peptides samples that were obtained by in-gel digestion were vacuum dried and stored at −20°C
until nano-high performance liquid chromatography (HPLC) ESI-Q-TOF MS/MS analysis.
Mascot.dll v 1.4804.0.22 (Matrix Science/AB Sciex) was used to generate Mascot (.mgf) files with
peak lists from the Analyst QS 2.0 (.wiff) files; the default parameters were used.
The MS/MS spectra that were obtained by spot samples were analyzed as Mascot generic files
against all entries in the public NCBInr database using the online Mascot program [79] without a
taxonomy filter.
The principal parameter settings for the Mascot search were as follows: database NCBInr
(version 2012.04.07; 17751536 sequences) [80]; enzyme trypsin; allow up to one missed cleavage;
variable modifications carbamidomethyl (C), oxidation (M), deamidated (NQ); tolerances
peptide ± 0.25 Da and MS/MS ± 0.25 Da; instrument ESI-QUAD-TOF; default charge state was set to
2+, 3+, and 4+. Only proteins with at least two peptides identified with significant ion scores (p ≤ 0.05)
were considered as identified. Protein identification details are shown in Table S1.
5. Conclusions
A large number of studies have focused on acute pollutant toxicity [81]. The ability of
D. discoideum to eliminate copper ions has been described previously [82]. D. discoideum has been

Int. J. Mol. Sci. 2012, 13 15700
proposed as a model organism in toxicology [83]. However, there is little information about the effects
of toxic mixtures on the D. discoideum proteome.
We elucidated the proteomic responses of D. discoideum that were exposed to NiCl2, CHP and their
mixture. It is likely that the direct use of PES requires much more data and more sensitive techniques
to resolve the molecular mechanisms of the cellular responses to be routinely applicable to toxicity
evaluations in environmental matrixes. There are obvious difficulties in understanding the biological
complexity of the molecular mechanisms of the cell, and the technical procedures of the present study
could be used to study the interactions between other mixtures and in other organisms. They could also
be used to develop new methods to evaluate the toxicity of individual toxicants and their mixtures. The
comparisons of the protein fingerprints of an equitoxic binary mixture and its single constituent
compounds allows us to individuate additional molecular mechanisms that are switched on, which can
be studied in the future with more specific approaches (e.g., the involvement of the cytoskeleton in
toxic responses). Our study not only detected known pollutant-modulated proteins such as
peroxiredoxins, aldehyde reductase and dehydrogenase, and S-adenosyl-methionine synthetase, which
were observed previously [84–86], but also revealed significant changes in the levels of unknown
proteins, such as a putative transport protein (gi|166240434), that were strongly induced by the mixture
but were never previously studied during a toxic response.
Acknowledgments
This study was supported by a grant from the UE 6th FW Program, NoMiracle, IP Contract number
003956.
References
1. Cedergreen, N.; Streibig, J.C. Can the choice of endpoint lead to contradictory results of
mixture-toxicity experiments. Environ. Toxicol. Chem. 2005, 24, 1676–1683.
2. Amorim, M.J.B.; Pereira, C.; Menezes-Oliveira, V.B.; Campos, B.; Soares, A.M.V.M.; Loureiro, S.
Assessing single and joint effects of chemicals on the survival and reproduction of
Folsomia candida (Collembola) in soil. Environ. Pollut. 2012, 160, 145–152.
3. Altenburger, R.; Scholz, S.; Schmitt-Jansen, M.; Busch, W.; Escher, B.I. Mixture toxicity
revisited from a toxicogenomic perspective. Environ. Sci. Technol. 2012, 46, 2508–2522.
4. Rager, J.E.; Lichtveld, K.; Ebersviller, S.; Smeester, L.; Jaspers, I.; Sexton, K.G.; Fry, R.C.
A toxicogenomic comparison of primary and photochemically altered air pollutant mixtures.
Environ. Health Persp. 2011, 119, 1583–1589.
5. Finne, E.F.; Cooper, G.A.; Koop, B.F.; Hylland, K.; Tollefsen, K.E. Toxicogenomic responses in
rainbow trout (Oncorhynchus mykiss) hepatocytes exposed to model chemicals and a synthetic
mixture. Aquat. Toxicol. 2007, 81, 293–303.
6. Quandt, S.A.; Chen, H.; Grzywacz, J.G.; Vallejos, Q.M.; Galvan, L.; Arcury, T.A. Cholinesterase
depression and its association with pesticide exposure across the agricultural season among Latino
farmworkers in North Carolina. Environ. Health Persp. 2010, 118, 635–639.

Int. J. Mol. Sci. 2012, 13 15701
7. Schröder, P.; Collins, C.D.; Markert, B.; Wünschmann, S. Bioindicators and Biomonitors: Use of
Organisms to Observe the Influence of Chemicals on the Environment. In Organic Xenobiotics
and Plants; Springer Netherlands: Dordrecht, The Netherlands, 2011; Volume 8, pp. 217–236.
8. Coogan, T.P.; Latta, D.M.; Snow, E.T.; Costa, M. Toxicity and carcinogenicity of nickel
compounds. Crit. Rev. Toxicol. 1989, 19, 341–384.
9. US Environmental Protection Agency. Ambient Water Quality Criteria for Nickel; Office of Water
Regulations and Standards: Washington, DC, USA, 1986. EPA/440/5-86/004.
10. Brix, K.V.; Keithly, J.T.; DeForest, D.K.; Laughlin, T. Acute and chronic toxicity of nickel to
rainbow trout (Oncorhynchus mykiss). Environ. Toxicol. Chem. 2004, 23, 2221–2228.
11. Banni, M.; Jebali, J.; Guerbej, H.; Dondero, F.; Boussetta, H.; Viarengo, A. Mixture toxicity
assessment of nickel and chlorpyrifos in the sea bass Dicentrarchus labrax. Arch. Environ.
Contam. Toxicol. 2011, 60, 124–131.
12. Keithly, J.; Brooker, J.A.; DeForest, D.K.; Wu, B.K.; Brix, K.V. Acute and chronic toxicity
of nickel to a cladoceran (Ceriodaphnia dubia) and an amphipod (Hyalella azteca).
Environ. Toxicol. Chem. 2004, 23, 691–696.
13. Majorel, C.; Hannibal, L.; Soupe, M.-E.; Carriconde, F.; Ducousso, M.; Lebrun, M.; Jourand, P.
Tracking nickel-adaptive biomarkers in Pisolithus albus from New Caledonia using a
transcriptomic approach. Mol. Ecol. 2012, 21, 2208–2223.
14. Macomber, L.; Hausinger, R.P. Mechanisms of nickel toxicity in microorganisms. Metallomics
2011, 3, 1153–1162.
15. Yusuf, M.; Fariduddin, Q.; Hayat, S.; Ahmad, A. Nickel: An overview of uptake, essentiality and
toxicity in plants. Bull. Environ. Contam. Toxicol. 2011, 86, 1–17.
16. Lemus, R.; Abdelghani, A. Chlorpyrifos: An unwelcome pesticide in our homes. Rev. Environ.
Health 2000, 15, 421–433.
17. Eaton, D.L.; Daroff, R.B.; Autrup, H.; Bridges, J.; Buffler, P.; Costa, L.G.; Coyle, J.; McKhann, G.;
Mobley, W.C.; Nadel, L.; et al. Review of the toxicology of chlorpyrifos with an emphasis on
human exposure and neurodevelopment. Crit. Rev. Toxicol. 2008, 38, 1–125.
18. Amaroli, A.; Trielli, F.; Bianco, B.; Giordano, S.; Moggia, E.; Corrado, M.U.D. Effects of
time-variant extremely low-frequency (ELF) electromagnetic fields (EMF) on cholinesterase
activity in Dictyostelium discoideum (Protista). Chem. Biol. Interact. 2005, 157, 355–356.
19. Qiao, D.; Seidler, F.J.; Slotkin, T.A. Developmental neurotoxicity of chlorpyrifos modeled
in vitro: Comparative effects of metabolites and other cholinesterase inhibitors on DNA synthesis
in PC12 and C6 cells. Environ. Health Persp. 2001, 109, 909–913.
20. Richards, S.M.; Kendall, R.J. Biochemical effects of chlorpyrifos on two developmental stages of
Xenopus laevis. Environ. Toxicol. Chem. 2002, 21, 1826–1835.
21. Whitney, K.D.; Seidler, F.J.; Slotkin, T.A. Developmental neurotoxicity of chlorpyrifos—Cellular
mechanisms. Toxicol. Appl. Pharmacol. 1995, 134, 53–62.
22. Narra, M.R.; Begum, G.; Rajender, K.; Rao, J.V. Sub-lethal effect of chlorpyrifos on protein
metabolism of the food fish Clarias batrachus and monitoring of recovery. Toxicol. Environ. Chem.
2011, 93, 1650–1658.

Int. J. Mol. Sci. 2012, 13 15702
23. Demir, F.; Uzun, F.G.; Durak, D.; Kalender, Y. Subacute chlorpyrifos-induced oxidative stress in
rat erythrocytes and the protective effects of catechin and quercetin. Pestic. Biochem. Physiol.
2011, 99, 77–81.
24. Jiang, W.; Duysen, E.G.; Hansen, H.; Shlyakhtenko, L.; Schopfer, L.M.; Lockridge, O. Mice
treated with chlorpyrifos or chlorpyrifos oxon have organophosphorylated tubulin in the brain and
disrupted microtubule structures, suggesting a role for tubulin in neurotoxicity associated with
exposure to organophosphorus agents. Toxicol. Sci. 2010, 115, 183–193.
25. Dondero, F.; Banni, M.; Negri, A.; Boatti, L.; Dagnino, A.; Viarengo, A. Interactions of a
pesticide/heavy metal mixture in marine bivalves: A transcriptomic assessment. BMC Genomics
2011, 12, 195.
26. Moore, M.N.; Farrar, S.V. Effects of polynuclear aromatic-hydrocarbons on lysosomal
membranes in mollusks. Mar. Environ. Res. 1985, 17, 222–225.
27. Viarengo, A.; Nott, J.A. Mechanisms of heavy-metal cation homeostasis in marine-invertebrates.
Comp. Biochem. Physiol. C 1993, 104, 355–372.
28. Loewe, S.; Muischnek, H. Effect of combinations: Mathematical basis of problem. Arch. Exp.
Pathol. Pharmacol. 1926, 114, 313–326.
29. Hughes, J.E.; DeLange, K.L.; Welker, D.L. Dictyostelium discoideum cobB mutants show
reduced heavy metal accumulation associated with gene amplification. Mol. Gen. Genet. 1996,
253, 65–73.
30. Karthikeyan, J.; Bavani, G. Effect of cadmium on lactate dehyrogenase isoenzyme, succinate
dehydrogenase and NA(+)-K+-ATPase in liver tissue of rat. J. Environ. Biol. 2009, 30, 895–898.
31. Espartero, J.; Pintortoro, J.A.; Pardo, J.M. Differential accumulation of S-adenosylmethionine
synthetase transcripts in response to salt stress. Plant Mol. Biol. 1994, 25, 217–227.
32. Mayne, M.B.; Coleman, J.R.; Blumwald, E. Differential expression during drought conditioning
of a root-specific S-adenosylmethionine synthetase from jack pine (Pinus banksiana Lamb)
seedlings. Plant Cell Environ. 1996, 19, 958–966.
33. Ahsan, N.; Lee, D.-G.; Alam, I.; Kim, P.J.; Lee, J.J.; Ahn, Y.-O.; Kwak, S.-S.; Lee, I.-J.; Bahk, J.D.;
Kang, K.Y.; et al. Comparative proteomic study of arsenic-induced differentially expressed
proteins in rice roots reveals glutathione plays a central role during As stress. Proteomics 2008, 8,
3561–3576.
34. Hubberstey, A.V.; Mottillo, E.P. Cyclase-associated proteins: CAPacity for linking signal
transduction and actin polymerization. FASEB J. 2002, 16, 487–499.
35. Sultana, H.; Rivero, F.; Blau-Wasser, R.; Schwager, S.; Balbo, A.; Bozzaro, S.; Schleicher, M.;
Noegel, A.A. Cyclase-associated protein is essential for the functioning of the endo-lysosomal
system and provides a link to the actin cytoskeleton. Traffic 2005, 6, 930–946.
36. Sun, Y.; Ou, Y.; Cheng, M.; Ruan, Y.; van der Hoorn, F.A. Binding of nickel to testicular
glutamate-ammonia ligase inhibits Its enzymatic activity. Mol. Reprod. Dev. 2011, 78, 104–115.
37. Zhang, B.L.; Zhang, Y.Q.; Dagher, M.C.; Shacter, E. Rho GDP dissociation inhibitor protects
cancer cells against drug-induced apoptosis. Cancer Res. 2005, 65, 6054–6062.
38. Hartwig, A. Zinc finger proteins as potential targets for toxic metal ions: Differential effects on
structure and function. Antioxid. Redox Signal. 2001, 3, 625–634.

Int. J. Mol. Sci. 2012, 13 15703
39. Ahsan, N.; Lee, D.-G.; Lee, S.-H.; Kang, K.Y.; Lee, J.J.; Kim, P.J.; Yoon, H.-S.; Kim, J.-S.;
Lee, B.-H. Excess copper induced physiological and proteomic changes in germinating rice seeds.
Chemosphere 2007, 67, 1182–1193.
40. Bhatt, I.; Tripathi, B.N. Plant peroxiredoxins: Catalytic mechanisms, functional significance and
future perspectives. Biotechnol. Adv. 2011, 29, 850–859.
41. Bonin, S.; Larese, F.F.; Trevisan, G.; Avian, A.; Rui, F.; Stanta, G.; Bovenzi, M. Gene expression
changes in peripheral blood mononuclear cells in occupational exposure to nickel. Exp. Dermatol.
2011, 20, 147–148.
42. Kubrak, O.I.; Husak, V.V.; Rovenko, B.M.; Poigner, H.; Mazepa, M.A.; Kriews, M.; Abele, D.;
Lushchak, V.I. Tissue specificity in nickel uptake and induction of oxidative stress in kidney and
spleen of goldfish Carassius auratus, exposed to waterborne nickel. Aquat. Toxicol. 2012, 118, 88–96.
43. Lynn, S.; Yew, F.H.; Chen, K.S.; Jan, K.Y. Reactive oxygen species are involved in nickel
inhibition of DNA repair. Environ. Mol. Mutagen. 1997, 29, 208–216.
44. Randhawa, V.K.; Zhou, F.Z.; Jin, X.L.; Nalewajko, C.; Kushner, D.J. Role of oxidative stress and
thiol antioxidant enzymes in nickel toxicity and resistance in strains of the green alga
Scenedesmus acutus f. alternans. Can. J. Microbiol. 2001, 47, 987–993.
45. Fang, H.-Y.; Chang, C.-L.; Hsu, S.-H.; Huang, C.-Y.; Chiang, S.-F.; Chiou, S.-H.; Huang, C.-H.;
Hsiao, Y.-T.; Lin, T.-Y.; Chiang, I.P.; et al. ATPase family AAA domain-containing 3A is a
novel anti-apoptotic factor in lung adenocarcinoma cells. J .Cell Sci. 2010, 123, 1171–1180.
46. Berger, W.; Steiner, E.; Grusch, M.; Elbling, L.; Micksche, M. Vaults and the major vault protein:
Novel roles in signal pathway regulation and immunity. Cell Mol. Life Sci. 2009, 66, 43–61.
47. Ryu, S.J.; Park, S.C. Targeting major vault protein in senescence-associated apoptosis resistance.
Expert Opin. Ther. Targets 2009, 13, 479–484.
48. Baviskar, S.N.; Shields, M.S. RNAi silenced Dd-grp94 (Dictyostelium discoideum
glucose-regulated protein 94 kDa) cell lines in Dictyostelium exhibit marked reduction in growth
rate and delay in development. Gene Expr. 2010, 15, 75–87.
49. Mani, A.; Gelmann, E.P. The ubiquitin-proteasome pathway and its role in cancer. J. Clin. Oncol.
2005, 23, 4776–4789.
50. Oliveira, I.C.; Coruzzi, G.M. Carbon and amino acids reciprocally modulate the expression of
glutamine synthetase in arabidopsis. Plant Physiol. 1999, 121, 301–309.
51. Reddy, P.S.; Bhagyalakshmi, A. Fenitrothion-induced alterations in ammonia metabolism in the
crab, Oziotelphusa-senex-senex Fabricius. Pestic. Sci. 1992, 35, 249–254.
52. Lukaszewicz-Hussain, A.; Moniuszko-Jakoniuk, J. Chlorfenvinphos, an organophosphate
insecticide, affects liver mitochondria antioxidative enzymes, glutathione and hydrogen peroxide
concentration. Pol. J. Environ. Stud. 2004, 13, 397–401.
53. Almeida, J.R.; Oliveira, C.; Gravato, C.; Guilhermino, L. Linking behavioural alterations with
biomarkers responses in the European seabass Dicentrarchus labrax L. exposed to the
organophosphate pesticide fenitrothion. Ecotoxicology 2010, 19, 1369–1381.
54. Lee, S.M.; Koh, H.J.; Park, D.C.; Song, B.J.; Huh, T.L.; Park, J.W. Cytosolic
NADP(+)-dependent isocitrate dehydrogenase status modulates oxidative damage to cells.
Free Radic. Biol. Med. 2002, 32, 1185–1196.

Int. J. Mol. Sci. 2012, 13 15704
55. Noguer, T.; Marty, J.L. High sensitive bienzymic sensor for the detection of dithiocarbamate
fungicides. Anal. Chim. Acta 1997, 347, 63–70.
56. Marty, J.L.; Noguer, T. Bienzyme amperometric sensor for the detection of dithiocarbamate
fungicides. Analysis 1993, 21, 231–233.
57. Marty, J.L.; Mionetto, N.; Noguer, T.; Ortega, F.; Roux, C. Enzyme sensors for the detection of
pesticides. Biosens. Bioelectron. 1993, 8, 273–280.
58. Ginzberg, I.; Barel, G.; Ophir, R.; Tzin, E.; Tanami, Z.; Muddarangappa, T.; de Jong, W.;
Fogelman, E. Transcriptomic profiling of heat-stress response in potato periderm. J. Exp. Bot.
2009, 60, 4411–4421.
59. Doran, B.; Gherbesi, N.; Hendricks, G.; Flavell, R.A.; Davis, R.J.; Gangwani, L. Deficiency of
the zinc finger protein ZPR1 causes neurodegeneration. Proc. Natl. Acad. Sci. USA 2006, 103,
7471–7475.
60. Gangwani, L. Deficiency of the zinc finger protein ZPR1 causes defects in transcription and cell
cycle progression. J. Biol. Chem. 2006, 281, 40330–40340.
61. Cairo, G.; Pietrangelo, A. Iron regulatory proteins in pathobiology. Biochem. J. 2000, 352, 241–250.
62. Sax, C.M.; Salamon, C.; Kays, W.T.; Guo, J.; Yu, F.S. X.; Cuthbertson, R.A.; Piatigorsky, J.
Transketolase is a major protein in the mouse cornea. J. Biol. Chem. 1996, 271, 33568–33574.
63. Singh, P.K.; Rai, S.; Pandey, S.; Agrawal, C.; Shrivastava, A.K.; Kumar, S.; Rai, L.C. Cadmium
and UV-B induced changes in proteome and some biochemical attributes of Anabaena sp.
PCC7120. Phykos 2012, 42, 39–50.
64. Gerke, V.; Moss, S.E. Annexins and membrane dynamics. BBA-Mol. Cell Res. 1997, 1357, 129–154.
65. Gerke, V.; Creutz, C.E.; Moss, S.E. Annexins: Linking Ca2+ signalling to membrane dynamics.
Nat. Rev. Mol. Cell Biol. 2005, 6, 449–461.
66. Rhee, H.J.; Kim, G.Y.; Huh, J.W.; Kim, S.W.; Na, D.S. Annexin I is a stress protein induced by
heat, oxidative stress and a sulfhydryl-reactive agent. Eur. J. Biochem. 2000, 267, 3220–3225.
67. Sacre, S.M.; Moss, S.E. Intracellular localization of endothelial cell annexins is differentially
regulated by oxidative stress. Exp. Cell Res. 2002, 274, 254–263.
68. Tanaka, T.; Akatsuka, S.; Ozeki, M.; Shirase, T.; Hiai, H.; Toyokuni, S. Redox regulation of
annexin 2 and its implications for oxidative stress-induced renal carcinogenesis and metastasis.
Oncogene 2004, 23, 3980–3989.
69. Gorecka, K.M.; Konopka-Postupolska, D.; Hennig, J.; Buchet, R.; Pikula, S. Peroxidase activity
of annexin 1 from Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2005, 336, 868–875.
70. Jami, S.K.; Clark, G.B.; Turlapati, S.A.; Handley, C.; Roux, S.J.; Kirti, P.B. Ectopic expression of
an annexin from Brassica juncea confers tolerance to abiotic and biotic stress treatments in
transgenic tobacco. Plant Physiol. Biochem. 2008, 46, 1019–1030.
71. Forsthoefel, N.R.; Cushman, M.A.F.; Cushman, J.C. Posttranscriptional and Posttranslational
Control of Enolase Expression in the Facultative Crassulacean Acid Metabolism Plant
Mesembryanthemum crystallinum L. Plant Physiol. 1995, 108, 1185–1195.
72. Yang, H.S.; Henning, D.; Valdez, B.C. Functional interaction between RNA helicase II Gu alpha
and ribosomal protein L4. FEBS J. 2005, 272, 3788–3802.

Int. J. Mol. Sci. 2012, 13 15705
73. Spycher, S.E.; TabatabaVakili, S.; Odonnell, V.B.; Palomba, L.; Azzi, A. Aldose reductase
induction: A novel response to oxidative stress of smooth muscle cells. FASEB J. 1997, 11,
181–188.
74. Marsano, F.; Boatti, L.; Ranzato, E.; Cavaletto, M.; Magnelli, V.; Dondero, F.; Viarengo, A.
Effects of mercury on Dictyostelium discoideum: Proteomics reveals the molecular mechanisms
of physiological adaptation and toxicity. J. Proteome Res. 2010, 9, 2839–2854.
75. Watts, D.J.; Ashworth, J.M. Growth of myxameobae of the cellular slime mould
Dictyostelium discoideum in axenic culture. Biochem. J. 1970, 119, 171–174.
76. Sussman, M. Cultivation and synchronous morphogenesis of Dictyostelium under controlled
experimental conditions. Methods Cell Biol. 1987, 28, 9–29.
77. Dondero, F.; Jonsson, H.; Rebelo, M.; Pesce, G.; Berti, E.; Pons, G.; Viarengo, A. Cellular
responses to environmental contaminants in amoebic cells of the slime mould
Dictyostelium discoideum. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 150–157.
78. Neuhoff, V.; Arold, N.; Taube, D.; Ehrhardt, W. Improved staining of proteins in polyacrylamide
gels including isoelectric focusing gels with clear background at nanogram sensitivity using
Coomassie Brilliant Blue G-250 and R-250. Electrophoresis 1988, 9, 255–262.
79. Matrix Science. Available online: http://www.matrixscience.com/ (accessed on 1 June 2012).
80. National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/
(accessed on 1 July 2012).
81. Powlesland, C.; George, J. Acute and chronic toxicity of nickel to larvae of Chironomus riparis
(Meigen). Environ. Pollut. A 1986, 42, 47–64.
82. Burlando, B.; Evangelisti, V.; Dondero, F.; Pons, G.; Camakaris, J.; Viarengo, A. Occurrence of
Cu-ATPase in Dictyostelium: Possible role in resistance to copper. Biochem. Biophys. Res.
Commun. 2002, 291, 476–483.
83. Gayatri, R.; Chatterjee, S. Phagocytic-activity of dictyostelium amoebae treated with an
organochlorine pesticide. Cell Biol. Internat. 1993, 17, 349–352.
84. Yin, L.; Mano, J.; Wang, S.; Tsuji, W.; Tanaka, K. The involvement of lipid peroxide-derived
aldehydes in aluminum toxicity of tobacco roots. Plant Physiol. 2010, 152, 1406–1417.
85. Costa, P.M.; Chicano-Galvez, E.; Barea, J.L.; DelValls, T.A.; Costa, M.H. Alterations to
proteome and tissue recovery responses in fish liver caused by a short-term combination treatment
with cadmium and benzo a pyrene. Environ. Pollut. 2010, 158, 3338–3346.
86. Zhou, Q.; Wu, C.G.; Dong, B.; Li, F.H.; Liu, F.Q.; Xiang, J.H. Proteomic analysis of acute
responses to copper sulfate stress in larvae of the brine shrimp, Artemia sinica. Chin. J. Oceanol.
Limnol. 2010, 28, 224–232.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article
distributed under the terms and conditions of the Creative Commons Attribution license
(http://creativecommons.org/licenses/by/3.0/).





























