Embed Size (px)
Citation preview

University of Veterinary Medicine Hannover, Foundation
Effect of anthropogenic underwater noise on harbour
porpoise hearing in areas of high ecological importance
Einfluss anthropogener Schalleinträge auf das Gehör von Schweinswalen in Gebieten
von hoher ökologischer Bedeutung
INAUGURAL - DISSERTATION
submitted in fulfilment of the requirements for the degree
- Doctor rerum naturalium -
(Dr. rer. nat.)
submitted by
Tobias Schaffeld
Kamp-Lintfort
Büsum 2020

Scientific supervision: Prof. Prof. h. c. Dr. Ursula Siebert
Institute for Terrestrial and Aquatic Wildlife Research,
University of Veterinary Medicine Hannover, Foundation.
Prof. Dr. Felix Felmy
Institute of Zoology,
University of Veterinary Medicine Hannover, Foundation.
First supervisors: Prof. Prof. h. c. Dr. Ursula Siebert
Institute for Terrestrial and Aquatic Wildlife Research,
University of Veterinary Medicine Hannover, Foundation.
Prof. Dr. Felix Felmy
Institute of Zoology,
University of Veterinary Medicine Hannover, Foundation.
Second supervisor: Prof. emeritus Dr. Paul Nachtigall
Institute of Marine Biology,
University of Hawaii, Kailua Hawaii.
Date of oral examination: 13.05.2020

Parts of this study were published in peer-reviewed journals:
Schaffeld, T., Schnitzler, J.G., Ruser, A., Woelfing, B., Baltzer, J., Siebert, U. (2020). Effects
of multiple exposure to pile driving noise on harbor porpoise hearing during
simulated flights – an evaluation tool. Journal of the Acoustical Society of America,
Volume 147, Issue 2, 685-697, doi: 10.1121/10.0000595.
Schaffeld, T., Ruser, A., Woelfing, B., Baltzer, J., Kristensen, J.H., Larsson, J., Schnitzler,
J.G., Siebert, U. (2019). The use of seal scarers as a protective mitigation measure can
induce hearing impairment in harbour porpoises Journal of the Acoustical Society of
America, Volume 146, Issue 6, 4288-4298, doi: 10.1121/1.5135303.
Parts of this study are in preparation for a publication in a peer-reviewed journal:
Schaffeld, T., Schnitzler, J.G., Ruser, A., Woelfing, B., Baltzer, J., Fischer, M., Schuster, M.,
Siebert, U. (2020). Acoustic water velocity measurements in rivers can affect harbour
porpoise hearing.
Results of this study were published in reports:
Ruser, A., Baltzer, J., Schaffeld, T., Woelfing, B., Schnitzler, J.G, Siebert, U. (2019).
Auswirkungen des Unterwasserschalls der Offshore-Windenergieanlagen auf
marine Säugetiere – Unterwasserschall Effekte (UWE). Final Report on behalf of the
Federal Agency for Nature Conservation. Project title: Effects of underwater noise of
offshore-wind farms on marine mammals. Project no. FKZ 3515 82 200.
Schaffeld, T., Ruser, A., Schnitzler, J.G., Baltzer, J., Siebert, U. (2018). Bewertung von
Auswirkungen von Unterwasserschallemissionen auf Schweinswale (Phocoena
phocoena). Report on behalf of the Hamburg Port Authority.
Results of this study were presented on the following conferences:
Schaffeld, T., Ruser, A., Schnitzler, J.G., Woelfing, B., Baltzer, J., Siebert, U. (2018). Hazard
zones for potential auditory damage – Estimated cumulative effects of pile driving
for harbour porpoises in the German North Sea (Poster). In abstract book page 48. 6th
International meeting on the Effects of Sound in the Ocean on Marine Mammals, The
Hague, Netherlands, 9th – 14th of September 2018.
Schaffeld, T., Ruser, A., Schnitzler, J.G., Woelfing, B., Baltzer, J., Siebert, U. (2019). Effects
of multiple exposure to pile driving noise on harbour porpoise hearing during
simulated flights – a risk evaluation tool (Poster and Talk). In abstract book page 163.
The Effects of Noise on Aquatic Life, The Hague, Netherlands, 7th – 12th of July 2019.
Awarded with the 2nd prize of the Rodney Coates award for the best student presentation.
Schaffeld, T., Ruser, A., Schnitzler, J.G., Woelfing, B., Baltzer, J., Siebert, U. (2019). Acoustic
harassment devices (seal scarers) can affect harbour porpoise hearing temporarily
(Talk). In abstract book page 634-635. World Marine Mammal Conference, Barcelona,
Spain, 9th – 12th of December 2019.

Table of Contents
Table of contents
List of abbreviations .......................................................................................................................... I
Introduction ...................................................................................................................................... 1
I. Echolocation ....................................................................................................................................... 1
II. Hearing adaptations ............................................................................................................................ 3
III. Communication ................................................................................................................................... 4
IV. Determination of hearing thresholds ................................................................................................... 6
V. Effects of anthropogenic underwater noise pollution ......................................................................... 7
VI. Protective regulations for harbour porpoises .................................................................................... 10
VII. Motivation ......................................................................................................................................... 12
Effects of multiple exposure to pile driving noise on harbor porpoise hearing during
simulated flights – an evaluation tool. .................................................................................. 14
Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise hearing
during simulated flights – an evaluation tool ....................................................................... 15
Abstract ...................................................................................................................................................... 15
Author contributions .................................................................................................................................. 16
I. Introduction ....................................................................................................................................... 17
II. Material & Methods .......................................................................................................................... 20 A. Study site and pile driving recordings ......................................................................................................... 20 B. Sound propagation modelling and frequency analysis ................................................................................ 21 C. Model assumptions ...................................................................................................................................... 23 D. Estimation of hazard zones .......................................................................................................................... 25
III. Results ............................................................................................................................................... 26 A. Sound propagation in the study area ............................................................................................................ 26 B. Estimation of hazard zones .......................................................................................................................... 28
IV. Discussion ......................................................................................................................................... 33 A. Simulation results ........................................................................................................................................ 33 B. Application of evaluation tool ..................................................................................................................... 36 C. Ecological relevance of disturbance ............................................................................................................ 39 D. Conclusion ................................................................................................................................................... 41 E. Acknowledgement ....................................................................................................................................... 41 F. APPENDIX ................................................................................................................................................. 42
The use of seal scarers as a protective mitigation measure can induce hearing impairment in
harbour porpoises. ................................................................................................................. 44
Chapter 2: The use of seal scarers as a protective mitigation measure can induce hearing
impairment in harbour porpoises ......................................................................................... 45
Abstract ...................................................................................................................................................... 45
Author contributions .................................................................................................................................. 46
I. Introduction ....................................................................................................................................... 47

Table of Contents
II. Material & Methods .......................................................................................................................... 49 A. Study area and animal subject ..................................................................................................................... 49 B. Background noise recordings ...................................................................................................................... 49 C. Experimental procedure to measure hearing thresholds .............................................................................. 50 D. Fatiguing stimulus and experimental procedure .......................................................................................... 52 E. TTS definition ............................................................................................................................................. 55 F. Data analysis ................................................................................................................................................ 56
III. Results ............................................................................................................................................... 58 A. Baseline hearing thresholds ......................................................................................................................... 58 B. Post-exposure thresholds ............................................................................................................................. 58
IV. Discussion ......................................................................................................................................... 60 A. Application of Seal Scarers ......................................................................................................................... 60 B. Effect on harbour porpoise hearing ............................................................................................................. 61 C. Estimated hazard zones ............................................................................................................................... 63 D. Cumulative effects of multiple exposure ..................................................................................................... 64 E. Management approach ................................................................................................................................. 65 F. Conclusion ................................................................................................................................................... 66
V. Acknowledgement ............................................................................................................................ 67
VI. Appendix ........................................................................................................................................... 68
Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing .................................... 72
Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing ................ 73
Abstract ...................................................................................................................................................... 73
Author contributions .................................................................................................................................. 74
I. Introduction ....................................................................................................................................... 75 A. Study area and underwater noise measurements ......................................................................................... 77 B. Sound propagation model ............................................................................................................................ 79 C. Effect of 28 kHz fatiguing sound on harbour porpoise hearing ................................................................... 81 D. Simulation of harbour porpoise migration through acoustic field ............................................................... 82 E. Evaluation of potential to induce a TTS ...................................................................................................... 83
II. Results ............................................................................................................................................... 85 A. Baseline hearing thresholds ......................................................................................................................... 85 B. Post-exposure thresholds ............................................................................................................................. 85 C. TTS potential of flowmeter for single pulses .............................................................................................. 85 D. Received levels on simulated flights ........................................................................................................... 87 E. TTS potential for harbour porpoise which stay around the flowmeters....................................................... 89
III. Discussion ......................................................................................................................................... 90 A. Evaluation of potential effects of the flowmeter on harbour porpoise hearing ............................................ 90 B. Effects on behaviour .................................................................................................................................... 91 C. Management approach ................................................................................................................................. 91
IV. Acknowledgement ............................................................................................................................ 92
Conclusion ....................................................................................................................................... 93
I. Evolution of noise mitigation ............................................................................................................ 93 A. Unintended effects of underwater sound application................................................................................... 95 B. Naive implementation of underwater sounds .............................................................................................. 97 C. Established concept for noise mitigation ..................................................................................................... 98
II. Outlook ........................................................................................................................................... 100

Table of Contents
A. Acoustic flowmeter ................................................................................................................................... 100 B. Acoustic deterrent devices ......................................................................................................................... 101
III. Future directions of TTS research ................................................................................................... 101 A. Biological mitigation as a regulation approach ......................................................................................... 101 B. Understanding of TTS ............................................................................................................................... 102 C. Ecological relevance of temporary threshold shifts ................................................................................... 103
Summary ....................................................................................................................................... 107
Zusammenfassung ........................................................................................................................ 110
Bibliography ................................................................................................................................. 113
List of figures ................................................................................................................................ 133
List of tables .................................................................................................................................. 134
Acknowledgement ........................................................................................................................ 135
Declaration of Originality ............................................................................................................ 136
Curriculum vitae .......................................................................................................................... 137

List of abbreviations
I
List of abbreviations
In addition to the commonly used abbreviations following short forms were used:
ABR Auditory Brainstem Response
ADD Acoustic Deterrent Device
AEP Auditory Evoked Potential
AFM Acoustic Flowmeter
ITAW Institute for Terrestrial and Aquatic Wildlife Research
MDD Minimum Deterrence Distance
MSFD Marine Strategy Framework Directive
OWF Offshore wind farm
PTS Permanent Threshold Shift
SEL Sound Exposure Level (in dB re 1 µPa²s)
SELSS Sound Exposure Level of Single Strike (in dB re 1 µPa²s)
SELcum Cumulative Sound Exposure Level (in dB re 1 µPa²s)
SEL-TTSonset Minimum SEL to induce a significant temporary threshold shift
SHEQ Still Higher than Effective Quiet
SPL Sound Pressure Level (in dB re 1 µPa)
TTS Temporary Threshold Shift

II

Introduction
1
Introduction
I. Echolocation
Among marine mammals, toothed whales (Odontocetes) have extraordinarily adapted to an
environment with limited vision ranges (Fenton et al., 2014). Similar to bats, toothed whales
gain further sensory input in addition to visual and positional sensors by emitting brief acoustic
pulses (clicks) and waiting for returning echoes (Fenton et al., 2014). These echoes carry
information, encoded in the time difference between pulse and echo as same as acoustic
characteristics, to generate an auditory image of their environment (Fenton et al., 2014). This
process is called echolocation. Finding and catching prey items independent from vision
represents a huge evolutionary advantage, which enables toothed whales to occupy habitats
with limited visibility (Fenton et al., 2014). The ability to echolocate characterises the 72
odontocete species, which occur in all marine habitats from deep pelagic zones to rivers (Fenton
et al., 2014; McGowen et al., 2009). The key for this acoustic perception of the environment
was the evolution of a highly specialized nasal sound production system (e.g. Huggenberger et
al., 2009; Madsen et al., 2002). An important development in this context was the resettlement
of the blowhole to the top of the head, which enables to breathe easily at the surface. This was
achieved by a migration of the nasal passages up to a dorsal location, separating from the
esophagus (Fenton et al., 2014). In most tooth whale species, echolocation clicks are generated
by phonic lips, which are located in the upper nasal passages (Cranford et al., 1996; Madsen et
al., 2003). This tissue complex consists of a variety of diverticula and a pair of fatty bursae,
which is embedded in a pair of connective tissue lips (Cranford et al., 1996). This highly
specialized adaptation represents a major advantage compared to echolocating bats, which
spent two to five times more energy when producing calls by the larynx (Dechmann et al.,
2013). Odontocetes however, produce echolocation clicks by forcing pressurised air through
the phonic lips (Ridgway and Carder, 1988). The energetic costs to produce clicks are assumed
to be negligible, since even deep-diving pilot whales require very small amounts of of air per
click (50-100 µL,) which are recycled again between the nasal sacs (Foskolos et al., 2019). The
clicks are emitted in a narrow beam of high intensity after being filtered and collimated by the
concave upper surface of the skull, underlying air sacs and a fatty melon in the forehead (Aroyan
et al., 2000). Among the highly specialised members of the Odontocetes, the sperm whale
evolved an outstanding melon, accounting for about one-third of its total body length (e.g.

Introduction
2
Madsen et al., 2002). Moreover, the clicks travel from the phonic lips positioned at the anterior
end of the melon back through the melon towards the frontal air sacs, where they are reflected
and from where they travel through the junk into the environment (Madsen et al., 2002; Møhl
et al., 2003; Zimmer et al., 2005). This mechanism enables the sperm whale to produce a
narrow, forward-directed and high intense sonar beam, exceeding 236 dB re 1 µPa (Møhl et al.,
2003). This represents the highest biological sound level known.
The ability to produce high amplitude echolocation clicks is substantial to explore the occupied
large water bodies by long-range echolocation (Møhl et al., 2003). The detection range of
toothed whales is positively correlated with body size, as the transmitted acoustic power is
dependent on the size of the vibrating phonic lips and the involved muscles when generating
air pressure (Jensen et al., 2018; Ridgway and Carder, 1988). The vocalization frequency of
terrestrial animals is negatively correlated with body size (Bradbury and Vehrencamp, 2011),
which has been confirmed for toothed whales also (Jensen et al., 2018). The directivity index,
as an indicator for the perceived acoustic field, was shown to be almost the same for a variety
of tested toothed whales with weights between 50 and 30000 kg (Jensen et al., 2018). The
product of melon size as the acoustic transmitter and the frequency of produced echolocation
signals determines directivity (Jakobsen et al., 2013). Accordingly, in order to maintain a
narrow field of view, the harbour porpoise must produce clicks of higher frequency than the
sperm whale, since the melon size is much smaller (Jakobsen et al., 2013; Jensen et al., 2018).
The echolocation clicks of harbour porpoises are emitted in a highly directive beam of 11-13°
(Koblitz et al., 2012), which is half as wide as the receiving system (Kastelein et al., 2005). A
narrow acoustic field of view provides evolutionary benefits such as a larger detection range.
Echolocation clicks can travel further distances, when the acoustic field is focused, allowing
for a detection of prey patches at further distances (Au, 2014). At the same time, unwanted
echoes are also reduced by this spatial filtering, which is highly beneficial when operating in
acoustically complex habitats like a coastal environment (Dukas, 2004; Jensen et al., 2018).
The optimal-localization hypothesis furthermore describes, that a narrow acoustic field is
supportive in localizing prey (Yovel et al., 2010). The detection and localization of prey are
two different mechanisms, which compromise each other. The detection of targets is most
efficiently in the center of the echolocation beam, because returning echoes contain highest
intensity. The localization of targets is more precise, when the target is pointed off axis, at the

Introduction
3
point with the highest slope of the beam (Yovel et al., 2010). Harbour porpoises produce
echolocation clicks with a bandwidth of 6 – 26 kHz, which are centered between 130 and
140 kHz (Møhl and Andersen, 1973; Villadsgaard et al., 2007). These high frequency clicks
have a wavelength of slightly more than 1 cm, which enables to recognize small targets of
similar size (Miller and Wahlberg, 2013). This advantage allows preying upon small fish below
10 cm, which were found to represent an important food source (Wisniewska et al., 2016).
Additionally, the development of a high frequency biosonar represents a selective advantage,
since echolocation clicks are outside the hearing range of killer whales (Orcinus orca)
(Szymanski et al., 1999), which prey upon harbour porpoises (Cosentino, 2015).
II. Hearing adaptations
Mammalian hearing is accomplished by an outer ear, which receives acoustic signals, a middle
ear, which amplifies and filters signals and the inner ear and thus functions as a mechano-
chemical transducer (Ketten, 1997). This land mammal blueprint is generally followed by
cetacean ears; however, adaptations to aquatic life have evolved (Nummela et al., 2004).
Underwater hearing requires further adaptations of the ears, compared to in-air hearing, as these
do not function as a converter from airborne- to fluid-borne sound in the inner ear. In contrast,
an acoustic isolation of the inner ear from the environment is needed, because the density of
water is similar to the tissue, while in air the tissue is much denser (Fenton et al., 2014).
Therefore, evolutionary adaptations are required to compensate for the mismatch in impedance
(Cranford et al., 2010; Fenton et al., 2014). During evolutionary development the external
auditory meatus was eliminated in Odontocetes (Nummela et al., 2004). External auditory
canals lack an observable connection with the temporal bones or tympanic membrane and are
furthermore plugged with dense cerumen and cellular debris (Ketten, 1997). An acoustic
isolation of the inner ears is enhanced by dense tympanic and periotic bones, which are isolated
in sinuses (cited in Fenton et al., 2014). Acoustic signals arrive at the tympanoperiotic complex
through an ‘acoustic window’, a hollow section surrounding the lower jaw, which opens
caudally (Nummela et al., 2007). This section is filled with fat bodies, encapsulated in
connective tissue, with the function to focus incoming sound (reviewed in Koopman, 2018;
Varanasi et al., 1975). The acoustic fat bodies in the lower jaw have a similar density than
seawater, which enables a fast sound transmission. The acoustic fat channels are inserted into
the tympanic plate, which is a particular site of the tympanoperiotric complex, characterized by

Introduction
4
very thin portions of the ventrolateral wall of the tympanic bulla (Nummela et al., 1999). The
process of the malleus receives vibrations of the tympanic bone through a synostosis connection
to the tympanic plate (Cranford et al., 2010; Nummela et al., 1999). Vibrations are conducted
via the osseous chain to the cochlea, where these are transformed into electrical signals
(Cranford et al., 2010; Nummela et al., 1999). The emission of intense outgoing clicks during
echolocation necessitates an isolation from the sending and receiving pathway. This is achieved
by the lower jaw pathway, which allows for an improvement of echolocation performance with
excellent sensitivity and directionality in high frequency hearing of toothed whales (Aroyan,
2001; Au and Moore, 1984; Hemilä et al., 2010; Ketten, 1997; Rauschmann et al., 2006).
Over a period of 60 million years, marine mammals evolved anatomical and physiological
modifications, which enabled the transition from terrestrial to aquatic specialists (cited in
Williams and Worthy, 2002). One challenge which has to be overcome is the locomotion,
impeded by an 800 times higher density and 60 times higher viscousity of water in comparison
to air (Dejours, 1987). A 24 times higher thermal conductivity of water is also challenging for
endothermic mammals, trying to prevent heat loss to the comparatively cold environment of
the ocean (Dejours, 1987; Williams and Worthy, 2002). A concurrent heat loss is mitigated by
a blubber layer, which undergoes cyclical fluctuations to compensate extra costs for
thermoregulation during winter (Rojano-Doñate et al., 2018).
The harbour porpoise (Phocoena phocoena) is the smallest representative amongst the marine
mammals. It shows a high surface-to-volume ratio suggesting to find them in rather warm water
environments, but they exclusively inhabit coastal waters of the northern hemisphere (Gilles et
al., 2016; Hammond et al., 2017). Survival in this cold environment is enabled by elevated field
metabolic rates, which are up to two times higher than for terrestrial mammals of comparable
size (Rojano-Doñate et al., 2018). These findings are in line with determined feeding rates of
wild porpoises, which can reach up to 550 small fish per hour (Wisniewska et al., 2016). There
is scientific evidence, that the high energy demands can only be maintained by almost
continuously foraging, which would demonstrate a low resilience to any disturbance
(Wisniewska et al., 2018a, 2016).
III. Communication
The key to success, when living in an apparently infinite environment, is communication.
Information is exchanged between a sender and a receiver to fulfil multiple kinds of tasks

Introduction
5
(Bradbury and Vehrencamp, 2011; Cholewiak et al., 2018). These messages can contain
information on the position and identity of group members (Lammers and Au, 2003) or food
patches (King and Janik, 2015; Parks et al., 2014), are under suspect to serve essential functions
during mating behaviour (Herman, 2017) or may even elicit anti-predator responses, e.g. when
eavesdropping on killer whale sounds (Curé et al., 2013). The marine environment physically
differs from the terrestrial one. Visibility is strongly limited underwater by the higher absorption
of light by suspended and floating particles (Dejours, 1987). Therefore, even in the photic zone,
underwater vision is barely the key to great hunting success. Although suspended particles limit
vision, these contribute to the four-times higher sound velocity in the water, making acoustic
communication efficient and often the only viable communication channel. With a speed of
sound of approximately 1500 m/s (Dejours, 1987) acoustic communication is the ultimate tool
for communication. Soniferous representatives were found in almost all classes of the animal
kingdom that inhabit the marine environment. Acoustic communication plays a major role for
both, invertebrates (Montgomery et al., 2017; Vermeij et al., 2010) and vertebrates (Clausen et
al., 2010; Ladich, 2019; Ladich and Winkler, 2017; Sørensen et al., 2018). Baseline hearing
abilities could be shown for a variety of fish species from multiple families (reviewed in Ladich
and Fay, 2013). Vocalizations are emitted during courtship or spawning by fishes (Amorim et
al., 2015) or may even mark a territory (Amorim, 2006; Ladich and Myrberg, 2006).
Soundscape ecology, derived from acoustic monitoring, allows for the assessment of
biodiversity, monitoring the health of habitats and the localisation of spawning grounds
(reviewed in Lindseth and Lobel, 2018). While acoustic signalling supports on the one hand the
reproductive success of Atlantic cod (Rowe and Hutchings, 2008), these signals could
concurrently reveal the presence of the fish to predators. Acoustic cues might be used by passive
listening predators, which might be the reason for a large proportion of soniferous fish species
in the diet of dolphins (Burros and Myrberg, 1987). This passive listening theory was validated
through playbacks of prey fish calls, which elicited attraction in bottlenose dolphins (Tursiops
truncatus, Gannon et al., 2005).
Active acoustic communication is the key for marine mammals, which all produce sound in
various contexts (reviewed in Erbe et al., 2016). The emission of species-specific calls has been
associated with mating behaviour in multiple baleen whale species (Croll et al., 2002; Herman,
2017) and seal (Frouin-Mouy et al., 2016; Nikolich et al., 2018; Rogers, 2017) species. The

Introduction
6
mating calls of male bearded seals are just one vivid example for the complexity and highly
developed vocalisations in marine mammals (Frouin-Mouy et al., 2016). The trill calls typically
have a duration of 30 s and consist of numerous repetitions of short sweeps with decreasing
frequency over time (Frouin-Mouy et al., 2016). These stereotypic calls were associated with
the mating period, potentially advertising the breeding condition of males (Frouin-Mouy et al.,
2016; Rogers, 2017). Moreover, single calls can contain sufficient information for recognition
between mother and calves, which is for instance essential for survival in colonial pinnipeds
(Charrier et al., 2010; Insley et al., 2003). Dolphins even have evolved individually distinctive
signature whistles, which share information about the identity of the caller and maintain group
integrity. When isolated from conspecifics, these calls can account for 90% of all vocalisations
(reviewed in Janik and Sayigh, 2013). These whistles account for 38 – 70% in wild animals
(reviewed in Janik and Sayigh, 2013), which develop individual calls during the first year of
life and retain it throughout life (Sayigh et al., 1990). Some species display variations in their
vocal repertoire, which are described as dialects, since these differences occur between groups,
which could potentially interbreed (Conner, 1982). The evolution of different dialects enables
to differentiate between members of different populations living within the same geographic
region (Ford, 1989; Weilgart and Whitehead, 1997).
IV. Determination of hearing thresholds
Auditory thresholds can be measured using two different techniques, which substantially differ
in application possibilities and results. The hearing of marine mammals can either be tested
using psychophysical procedures, which requires extensive training of animals (Kastelein et al.,
2002, 2019b) or by neurophysiological methods with trained (Nachtigall et al., 2016) or
temporarily restrained individuals (Ruser et al., 2014, 2016a). The psychophysical method to
measure audiograms is regarded as the gold standard (Houser et al., 2017), but requires previous
extensive training of the animals by experienced animal trainers. The animals are conditioned
to display, if they heard a hearing test stimulus by the use of positive reinforcement (e.g.
Kastelein et al., 2019; Nachtigall et al., 2017). Hearing thresholds are accordingly determined
testing different sound pressure levels (SPLs). The neurophysiological technique is an indirect
method, because brainstem responses are measured, which are auditory evoked. Auditory
evoked potentials (AEP) are small electrical charges generated by the brain in response to an
acoustic stimulus, which can be measured on the body surface of a harbour porpoise by using

Introduction
7
electrodes imbedded in suction cups. This method allows for a fast audiometric measurement
without the necessity of preliminary conditioning (Supin et al., 2001). The application of the
AEP method to collect audiograms has gained in popularity over the last decade (Houser et al.,
2017). Although the general pattern does not substantially differ between psychophysically and
neurophysiologically derived audiograms, the absolute hearing threshold can differ by up to
31 dB (Mulsow and Reichmuth, 2010). Therefore, comparisons of hearing thresholds gathered
with different techniques have to be made with great caution. However, the AEP method does
not require previous training and is therefore well suited for the application in the field (Ruser
et al., 2016a) or for wild animals (Ruser et al., 2014).
V. Effects of anthropogenic underwater noise pollution
Since toothed whales rely critically on echolocation to find prey and perceive their environment,
they are especially vulnerable to interferences by noise. A significant source of disturbance for
marine life is the substantially increasing anthropogenic underwater noise pollution, which is
now recognized as a global problem (Erbe et al., 2019; Shannon et al., 2016; Williams et al.,
2015). Effects of increased anthropogenic noise pollution are particularly assessed in humans,
with strong evidence for psychological and physiological consequences like cognitive
impairment (Szalma and Hancock, 2011), sleep deprivation (Fyhri and Aasvang, 2010) or a risk
of cardiovascular diseases (Babisch et al., 2005; Hansell et al., 2013). As reviewed by Shannon
et al. (2016), protective legislation for humans was already implemented in the seventies in the
USA (Noise Control Act of 1972, Quiet Communities Act of 1978) and in 2002 in the European
Union (Environmental Noise Directive 2002/49/EC). The assessment of potential effects of
noise on aquatic life is challenging due to limited accessibility to animals. Further complications
in the quantification arise from the fact that the sensitivity to noise varies across taxa (Brumm
and Slabbekoorn, 2005; Shannon et al., 2016; Southall et al., 2019). Furthermore, even intra-
specific sensitivities occur depending on life history, context or sex (reviewed e.g. Ellison et
al., 2012 or Francis and Barber, 2013).
Biological consequences of noise exposure can be reflected on multiple scales. An exemplary
categorization to summarize biological responses towards effects of noise has been proposed
by Shannon et al. (2016) comprising nine different classes. Noise pollution can accordingly
affect the physiology of animals 1) by means of stress, hearing impairment, immunosuppression
or gene expression. The fitness of animals 2) can be directly affected, represented in survival

Introduction
8
rates, fecundity or birth rate or indirectly by effects on 3) mating behaviour in terms of
attraction, mating success, territorial behaviour or pair bonding. These results can be an effect
from masking, which means that biologically relevant signals cannot be recognized, since these
overlap with noise. Potential effects of noise can concern 4) the foraging behaviour in terms of
foraging rate and success. The exposure to noise can influence animals 5) movement
represented by the spatial distribution or could induce flight responses or alterations in diving
behaviour. The vigilance of animals 6) can be affected, which could secondary affect other
behavioural states and could therefore lead to missed foraging opportunities. Effects of noise
on the vocal behaviour 7) can be expressed by changes in the call rate, intensity, frequency,
signal duration, call type and timing of signals. Widespread effects can concern the population
metrics 8) such as abundance, occupancy, settlement or density but also the community level
9), regarding species composition and predator prey interactions (all classes cited from Shannon
et al., 2016).
Ship traffic has become the most pervasive and ubiquitous source of underwater noise pollution
in the recent decades caused by the globalisation of transportation networks, population growth
and expansion of resource extraction (Erbe et al., 2016; Shannon et al., 2016; Williams et al.,
2015). Between 1992 and 2012, the global ship density quadrupled, leading to a steady rise in
ambient noise levels between 10 and 100 Hz a high as 3 dB per decade (Andrew et al., 2002,
2011; Chapman and Price, 2011; Kleinertz et al., 2014; Miksis-Olds et al., 2013; Miksis-Olds
and Nichols, 2016). Vessel noise is considered to be a continuous noise source and has the
potential to elicit strong behavioural responses in harbour porpoises (Akkaya Bas et al., 2017;
Dyndo et al., 2015), which are assumed to indicate disturbance. Wild harbour porpoises in the
Inner Danish waters were exposed to vessel noise in 17 to 89% of the time these animals were
tagged with sound and movement recording tags (Wisniewska et al., 2018b). During the
exposure to vessel noise exceeding 96 dB re 1 µPa (in the 16 kHz third-octave band)
behavioural changes of the tagged harbour porpoises occurred (Wisniewska et al., 2018b). The
animals increased the fluke strike rate, resulting in an increased swim speed and stayed close
to the bottom (Wisniewska et al., 2018b). Additionally, a cessation of echolocation and a
significant decrease of foraging behaviour was determined for these animals (Wisniewska et
al., 2018b), which were found to feed almost continuously to maintain high energy demands
(Wisniewska et al., 2018a, 2016).

Introduction
9
Anthropogenic underwater noise also occurs in brief bursts of sound pressure, which are defined
as impulsive noise (ISO, 2016). Typical sources of anthropogenic impulsive noise are (1)
underwater explosions (e.g. von Benda-Beckmann et al., 2015), which occur when unexploded
ordnance of World War II are cleared, (2) seismic surveys with large arrays containing multiple
air guns in order to locate deposits of fossil fuels in the seafloor (e.g. Sarnocińska et al., 2020)
and (3) pile driving noise, which is emitted when fundaments are driven into the seabed (Brandt
et al., 2018; Graham et al., 2019; Leunissen and Dawson, 2018). These impulsive noise sources
are often characterized by high source levels, leading to higher local effects than vessel noise
(Hildebrand, 2009).
The exposure to impulsive noise with high received levels can be lethal or induce severe injuries
when haemorrhages occur in air filled cavities, around the brain or embolisms arise from
nitrogen bubble transformations in the blood stream (Cox et al., 2006; Fernández et al., 2005;
Siebert et al., 2013). Less severe injuries can affect the hearing of marine mammals (reviewed
in Finneran, 2015), which are not directly lethal, but can have dramatic secondary effects.
Affected hearing can influence the survival rate of single individuals (Mann et al., 2010; Morell
et al., 2017a). Interferences may even result in decreased individual fitness and could lead to
long-term population consequences (King et al., 2015). The exposure to impulsive noise has
the potential to induce physiological changes, such as hearing loss (Lucke et al., 2009). A shift
in hearing ability after the reception of a fatiguing noise may recover, in which case it is defined
as a temporary threshold shift (TTS) or may not resolve, leading to a permanent threshold shift
(PTS) (Finneran, 2015). Noise induced hearing loss can be a result from a very high-level single
impulsive sound event (Lucke et al., 2009), which is typically of short duration or from the
exposure over substantially longer periods to lower-level sounds (Kastelein et al., 2015d, 2016).
Such a TTS affects the hearing threshold for a certain frequency range rather than the complete
hearing curve (Lucke et al., 2009; Popov et al., 2013). The TTS is expressed at the main
frequency of the fatiguing stimulus, but is highest half an octave above the peak frequency for
large hearing shifts (Popov et al., 2013). Moreover, hearing impairment can severely affect
essential life functions (Morell et al., 2017a, 2017b) such as finding and catching prey
(DeRuiter et al., 2009; Verfuß et al., 2009), navigation (Verfuß et al., 2005), communication
with intra-specifics (Clausen et al., 2010; Sørensen et al., 2018) and avoid (Curé et al., 2013)
or find (Gannon et al., 2005) inter-specifics.

Introduction
10
VI. Protective regulations for harbour porpoises
Three marine mammal species are considered to reproduce regularly in German waters. The
harbour seal (Phoca vitulina), the grey seal (Halichoerus grypus) (CWSS, 2017) and the
harbour porpoise (Phocoena phocoena) (Gilles et al., 2016). Although seals display highly
developed hearing abilities in air (Ruser et al., 2014) and underwater (Kastelein et al., 2009a,
2009b), which indicates a necessity for sound perception, this thesis is focused exclusively on
the only Odontocete representative. Among the toothed whales, which are generally vulnerable
to noise disturbance, harbour porpoises are considered to be very sensitive to effects of noise
and are therefore regarded as an indicator species in noise impact evaluations (Southall et al.,
2019; Tougaard et al., 2015). Harbour porpoise distribution extends to coastal waters of the
northern hemisphere, including the Baltic Sea, Black Sea, Northern Atlantic and the North
Pacific (cited in Bjørge and Tolley, 2018). The North- and Baltic Seas are inhabited by three
different harbour porpoise populations, which are distinguished as the North Sea population,
the Inner Danish Waters population and the Baltic proper population (Galatius et al., 2012;
Huggenberger et al., 2002; Wiemann et al., 2010). In a large scale survey in 2016, harbour
porpoise abundance in the North Sea was estimated at 345,373 individuals (CI: 246,526-
495,752), whereas the Kattegat and Belt Seas account for an abundance of 42,324 individuals
(CI: 23,368-76,658, Hammond et al., 2017). Since the population of the Baltic proper was
estimated at 497 animals (CI: 80-1,091, Amundin 2016), this population is considered as
“critically endangered” by the IUCN (Carlén et al., 2018). Harbour porpoises display a sexual
dimorphism with smaller males with a length of about 145 cm and a weight of 50 kg, whereas
females reach average sizes of 160 cm and a weight of 60 kg (reviewed in Bjørge and Tolley,
2018). Nevertheless, maximum female sizes of 200 cm with a weight of 70 kg have been
reported occasionally (cited in Bjørge and Tolley, 2009). Sexual maturity was estimated in
female harbour porpoises of the German North- and Baltic Seas at an age of 4.95 years (±0.6)
(Kesselring et al., 2017). Harbour porpoises are predominantly piscivorous and are considered
as opportunistic predators (Andreasen et al., 2017), which forage nearly continuously
(Wisniewska et al., 2016) to meet high energy demands (Rojano-Doñate et al., 2018). The most
abundant prey species in harbour porpoise diet depends on prey availability within the region
or during season (Andreasen et al., 2017; Benke et al., 1998; Wisniewska et al., 2016).

Introduction
11
In European waters, harbour porpoises are protected amongst others within the framework of
the Habitats Directive (listed in annexes II and IV European Union, 1992) and Council
Regulation 812/2004 (European Union, 2004). This protective regulation implies that special
areas should be established for their conservation with deliberate actions of killing, disturbing,
injuring and habitat deterioration being prohibited throughout its area of distribution (Council
Directive 92 / 43 / EEC, Article 12.1). Furthermore, harbour porpoises are protected by the
agreement of ten European Union member states on the conservation of small cetaceans in the
North and Baltic Seas (ASCOBANS, 2002, 2012), as well as by the Baltic Marine Environment
Protection Commission (Helsinki Commission, HELCOM) in the Baltic Sea and the
Convention for The Protection of the Marine Environment of the North-East Atlantic (the
OSPAR Convention) in the North Sea.
Although the prevention of injuries in marine mammals has been considered globally (e.g. by
the Habitats Directive in Europe or by the U.S. Marine Mammal Protection Act in the United
States), “injury” is defined differently in national policies. According to the Federal Ministry
for the Environment, Nature Conservation, and Nuclear Safety (BMU, 2014) in Germany, a
temporary loss of hearing after exposure to pile driving noise is considered as injury, whereas
most other European countries or the United States only regard a permanent hearing shift (PTS)
as injury. There is no common EU regulation to protect harbour porpoises from hearing
impairment during the construction of offshore wind farms in the EU (Stöber and Thomsen,
2019). However, noise mitigation measures are already obliged in Belgium, Denmark,
Germany and the Netherlands. Belgium and the Netherlands additionally restrict pile driving
activities to designated periods. Germany has enforced the strictest regulations in the EU to
date, by limiting the maximum sound exposure level (SEL) to 160 dB re 1 μPa²s and a
maximum sound pressure level (SPL) of 190 dB re 1 μPa for a single strike at a distance of
750 m (BMU, 2014). Furthermore, wind farm operators are obliged to deter harbour porpoises
prior to pile driving activities and to use a soft start procedure with limited force and longer
pulse intervals in the beginning (BMU, 2014). Despite the regulations of pile driving noise,
there is no further regulation of noise emission.

Introduction
12
VII. Motivation
The aim of this doctoral thesis is to tackle the gap in current protective regulations and identify
present anthropogenic noise sources with the potential to induce a TTS in harbour porpoises.
Since the sources of anthropogenic noise are numerous, this study focuses on a selection of
three, covered in individual chapters:
Chapter 1 is dedicated to the current protective regulations of noise mitigation during the
construction of offshore wind farms in German waters (BMU, 2014). Part of this is the analysis
of underwater noise recordings, which were gathered during the construction of the offshore
wind farm “Amrumbank West”. This wind farm is of particular interest, since it was adjacently
built to the special protection area “Sylt Outer Reef” within the Exclusive Economic Zone
(EEZ) of the German North Sea, which shows high harbour porpoise densities (Gilles et al.,
2011) and serves as an important calving ground (Gilles et al., 2009). The 80 wind turbines of
this wind farm were constructed in compliance with the current noise mitigation concept of the
Federal Ministry for the Environment; Nature Conservation and Nuclear Safety (BMU, 2014).
The output of this study is an evaluation tool, which helps to evaluate the TTS potential for
fleeing harbour porpoises from the reception of multiple pile driving strikes, which is not
considered in the German noise mitigation regulations yet.
Chapter 2 represents the results of a study with a harbour porpoise in human care. Hearing tests
were conducted with an animal held at the Fjord & Bælt Centre in Kerteminde (DK) to test,
whether a seal scarer signal has the potential to induce a TTS in harbour porpoise hearing. These
acoustic deterrent devices (ADDs) are applied in two different situations. ADDs are used to
deter harbour porpoises prior to pile driving activities with the aim to clear the area close to the
construction site (BMU, 2014), where a TTS can be induced by pile driving noise (Lucke et al.,
2009). In this situation ADDs serve as a mitigation method, which wind farm operators are
obliged to follow. ADDs are also applied around marine aquacultures to deter seals in order to
counteract depredation (Dawson et al., 2013). This kind of application represents a significant
chronic noise source over a large spatial range along the Scottish West coast (Findlay et al.,
2018), which overlaps with harbour porpoise occurrence (Embling et al., 2010; Hammond et
al., 2013), as a non-target species. The aim of this study was to determine a minimum sound
energy level, at which a temporary threshold shift can be induced (SEL-TTSonset) by a single
ADD signal. The hearing thresholds were derived from measurements of auditory evoked

Introduction
13
potentials (AEP) prior to and after the exposure to artificial seal scarer signals. The SEL-
TTSonset as an output of this study can serve as an orientation to down regulate ADD source
levels, to enable pursuing application without the risk of a TTS.
Chapter 3 is dedicated to a situation of unintended effects of noise exposure in an area, which
is regularly visited by harbour porpoises. Harbour porpoises seasonally occur in the port of
Hamburg, an area that is highly exposed to ship traffic with vessels of all kinds and sizes.
Despite the high levels of vessel noise they show a seasonal occurrence, which might be driven
by anadromous prey fish, migrating upstream the Elbe River in spring. Within the area of
harbour porpoise occurrence in the port of Hamburg, the flow velocity is measured by acoustic
signals, which are in the frequency range of harbour porpoise hearing. This measuring device
was under suspect to be the cause of death for several harbour porpoises in the Elbe River in
2016. This study aims for a TTS risk assessment for this acoustic flowmeter, by the analysis of
underwater recordings in the port of Hamburg, sound exposure experiments with a harbour
porpoise in human care at the Fjord & Bælt Centre in Kerteminde (DK) and a simulation
approach to estimate the TTS potential for harbour porpoises travelling along the acoustic
flowmeter. The output of this study is an evaluation of the flowmeter to induce a TTS in harbour
porpoises and to provide management suggestions for potentially needed adjustments of the
acoustic flowmeter.
The overarching goal of this thesis is to assess the effect of noise disturbance from different
sources as an antagonist, interfering with acoustic perception of the environment. The aim is to
shed light on gaps in noise regulation and raise attention for the naive application of acoustic
devices, since harbour porpoises are particularly vulnerable to interferences by noise due to
their dependence from acoustically sensing their environment.

Effects of multiple exposure to pile driving noise on harbor porpoise hearing during
simulated flights – an evaluation tool.
14
Published:
Reproduced from,
Schaffeld, T., Schnitzler, J.G., Ruser, A., Woelfing, B., Baltzer, J., Siebert, U. (2020). Effects of multiple exposure to pile driving noise on harbor porpoise hearing during
simulated flights – an evaluation tool. Journal of the Acoustical Society of America,
147 (2), 685-697, https://doi.org/10.1121/10.0000595
with the permission of the Acoustical Society of America.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
15
Chapter 1: Effects of multiple exposure to pile driving noise on harbor
porpoise hearing during simulated flights – an evaluation tool
T. Schaffeld, J.G. Schnitzler, A. Ruser, B. Woelfing, J. Baltzer, U. Siebert.
Institute for Terrestrial and Aquatic Wildlife Research, University of Veterinary Medicine
Hannover, Foundation, Werftstraße 6, 25761 Büsum, Germany
This paper is part of a special issue on The Effects of Noise on Aquatic Life.
Abstract
Exploitation of renewable energy from offshore wind farms is substantially increasing
worldwide. The majority of wind turbines are bottom mounted, causing high levels of impulsive
noise during construction. To prevent temporary threshold shifts (TTS) in harbor porpoise
hearing, single strike sound exposure levels (SELSS) are restricted in Germany by law to a
maximum of 160 dB re 1µPa²s at a distance of 750 m from the sound source. Underwater
recordings of pile driving strikes, recorded during the construction of an offshore wind farm in
the German North Sea, were analyzed. Using a simulation approach, it was tested whether a
TTS can still be induced under current protective regulations by multiple exposures. The
evaluation tool presented here can be easily adjusted for different sound propagation, acoustic
signals or species and enables to calculate a minimum deterrence distance. Based on this
simulation approach, only the combination of SELSS regulation, previous deterrence and soft
start allow harbor porpoises to avoid a TTS from multiple exposures. However, deterrence
efficiency has to be monitored.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
16
Author contributions
J.B., T.S. and A.R. were involved in the underwater noise recordings around the Offshore Wind
Farm. T.S. and A.R. conceived the original theory for this simulation approach, which was
further developed with support from J.S. and B.W.. The sound propagation modelling was
performed by T.S. and J.S. T.S. performed the computations of the simulations and figures.
Mathematical equations were derived by B.W. T.S. wrote the manuscript. T.S. was supervised
by A.R., J.S. and U.S. All authors provided critical feedback and helped shape the research,
analysis and manuscript.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
17
I. Introduction
The marine environment provides an almost infinite source of offshore renewables, which may
be exploited with limited negative environmental impacts if guidelines are followed and the
planning and scaling of projects are suitable (Pelc and Fujita, 2002). Increasing efforts were
made worldwide to exploit offshore renewables. In Germany, plans to refrain from extracting
energy from fossil fuels or to close down nuclear power plants have been made over the last
years and became even more ambitious after the disaster in 2011 at the nuclear power plant in
Fukushima (Japan). Eighty percent of energy demands in Germany should be covered by
renewable forms of energy by 2050. Since 2009 the German Federal Government’s goal has
been to reach 25 Gigawatt by 2030 from offshore wind capacity by undertaking intensive
building of offshore wind farms (OWF) in German waters (BMWI, 2012). In total, 92 offshore
wind farms have been constructed to date in eleven European countries including sites with
partial grid connection, accounting for 4149 connected wind turbines (Remy and Mbistrova,
2018). This increased human encroachment overlaps with protected areas like the Sylt Outer
Reef and could have negative effects on health, distribution and behavior of key ecological
species inhabiting German offshore areas.
The inconspicuous and only resident cetacean in the German North Sea is the harbor porpoise
(Phocoena phocoena). It inhabits coastal waters and is therefore subject to anthropogenic
pressures, e.g. accidental bycatch (ASCOBANS, 2002), continuous shipping noise (Akkaya
Bas et al., 2017; Dyndo et al., 2015; Wisniewska et al., 2018b) and impulsive noise from pile
driving (Brandt et al., 2018; Tougaard et al., 2009), seismic surveys (Pirotta et al., 2014) or
underwater explosions (von Benda-Beckmann et al., 2015). In European waters, harbor
porpoises are protected among others within the framework of the Habitats Directive (listed in
annexes II and IV (European Union, 1992)) and Council Regulation 812/2004 (European
Union, 2004), implying that special areas should be established for their conservation with
deliberate actions of killing, disturbing, injuring and habitat deterioration being prohibited
throughout its range (Council Directive 92 / 43 / EEC, Article 12.1).
Although the prevention of injury in marine mammals has been considered globally (e.g. by the
Habitats Directive in Europe or by the U.S. Marine Mammal Protection Act in the United
States), injury is defined differently in national policies. According to the Federal Ministry for
the Environment, Nature Conservation, and Nuclear Safety (BMU (2014)) in Germany, a

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
18
temporary loss of hearing after exposure to pile driving noise (temporary threshold shifts, TTS)
is considered as an injury, whereas most other European countries or the United States regard
a permanent hearing shift (PTS) as an injury. Lucke et al. (2009) derived data on TTS induced
by single impulsive airgun stimuli, and defined an onset at a sound exposure level (SEL) of
164 dB re 1 µPa²s at a hearing frequency of 4 kHz, showing that harbor porpoises are more
sensitive to impulsive noise than other high frequency cetaceans (reviewed in Southall et al.,
2019). Consequently, the BMU published a regulation, which restricts the maximum SEL to
160 dB re 1 µPa²s for single impulsive noise at a distance of 750 m from the source (BMU,
2014) in reference to the findings of Lucke et al. (2009). To keep noise levels below this
threshold, wind farm operators are obliged to use most innovative noise mitigation systems
(NMS) like bubble curtains (Dähne et al., 2017; Lucke et al., 2011; Würsig et al., 2000) and
hydro sound dampers (HSD, Elmer et al. 2012). Additionally, acoustic deterrent devices (pinger
and seal scarer) are deployed before pile driving, to deter animals from the area, where noise
levels can exceed the threshold for an SEL of a single strike of 160 dB re 1 µPa²s.
Recent studies showed that besides the danger from a single pulse with high energy, the
reception of multiple pile driving strikes with single strike sound exposure levels (SELSS) well
below the legal threshold can also induce a TTS because of the total received energy. Indeed,
playbacks of pile driving sounds at an SELSS of 146 dB re 1 µPa²s induced a TTS in harbor
porpoises. A significant TTS at 4 and 8 kHz occurred after the playback of 2760 strikes within
60 minutes (Kastelein et al., 2015a). A TTSonset at 8 kHz hearing frequency was determined at
a cumulative SEL (SELcum) of 175 dB re 1 µPa²s, corresponding to 1385 pile driving strikes of
145 dB re 1 µPa²s (SELSS) in 30 minutes (Kastelein et al., 2016). Although regulations to
protect harbor porpoises from TTS by single impulsive sounds have already been established,
it was shown that the multiple reception of pile driving strikes can still induce a TTS.
In our present study, potential auditory hazard zones were estimated, within which hearing
impairment is theoretically possible for harbor porpoises. Several behavioral scenarios were
considered, which simulate effects for harbor porpoises that stay within the area of noise
exposure and those that show a flight response at different literature-based swim speeds. The
accumulation of sound energy from multiple pile driving strikes for harbor porpoises is
determined by using real underwater recordings from recent pile driving activities in the
German North Sea and its sound propagation. We estimated the potential to induce a TTS based

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
19
on the distance to the pile driving site, and identified the minimum distance a harbor porpoise
must be away from the pile driving site at the moment of the first strike to avoid auditory
impairment.
The aim of this study is to provide a tool to evaluate effects of multiple pile driving events on
harbor porpoise hearing. Equations presented here are adjustable for areas with differing sound
propagation or further species with different TTS onsets, and allow the necessary minimum
deterrent distances to be estimated. The outcomes of this study will highlight the efficiency of
current protective measures in force in Germany to prevent temporary hearing shifts in harbor
porpoises from pile driving noise. Currently, these measures restrict the maximum single strike
exposure levels to 160 dB re 1 µPa²s at a distance of 750 m and the use of acoustic deterrent
and harassment devices which are deployed before pile driving.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
20
II. Material & Methods
A. Study site and pile driving recordings
Underwater recordings during the construction of 50 monopiles were conducted between
August 27th 2014 and March 18th 2015 in order to determine underwater noise. Therefore,
Autonomous Multichannel Acoustic Recorders (AMAR, JASCO Applied Sciences, , Canada)
were bottom mounted at a depth of 20 m at seven measuring positions in the surrounding area
of the German offshore Amrumbank West wind farm at distances between 2.4 and 36.8 km to
pile driving sites (see FIG. 1 for further details on measuring positions and piles). Distances
between measuring positions and pile driving sites could be determined by logging the position
of measuring stations and reported pile positions by the wind farm operator. Background
underwater noise was continuously recorded in 30 min files sampling at 32 ksamples s-1 and a
16-bit resolution. All AMAR recorders were equipped with omnidirectional GTI-M8E
hydrophones (GeoSpectrum Technologies Inc., Dartmouth, Canada) with nominal sensitivities
of -160 and -200 dB re 1VµPa-1. In order to compensate for different received levels of pile
driving strikes, less sensitive hydrophones were deployed at closer distances. The recording
system was protected from bottom trawling by a trawl shield, meaning a glass fiber reinforced
plastic housing (270×1250×1000 mm, H×W×D, 8 mm material thickness) built by DW-
ShipConsult GmbH, Schwentinental Germany (see picture of trawl shield in Gerdes and Görler
(2016), Fig. 3 (right picture)). The trawl shield contained cropped circles on every side for a
better sound transmission. The hydrophone was fixed inside the trawl shield below a cropped
circle in a vertical position, pointing in the direction of the sea surface. External battery supplies
were also employed inside the trawl shield to enable continuous recording over an
approximately three-month period. The applied recording system fulfilled the requirements of
ISO 18406 (2017) and the German guidelines (Müller and Zerbs, 2011) in terms of sampling
frequency, data format, self-noise of the mooring system and electronic components,
hydrophone sensitivity (<2dB over the frequency range from 0.02 to 16 kHz) and interval of
calibration (two years).
The investigated monopiles were deployed by two different wind farm installation vessels (MPI
Discovery (Flag: the Netherlands) & HLV Svanen (Flag: the Bahamas)). Further vessels
applied big bubble curtains (BBC) and hydro sound dampers (HSD) for noise mitigation.
Further information about the application of these NMS during the construction is not publicly

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
21
accessible. Therefore, we cannot distinguish which NMS or combination of different NMS was
utilized.
FIG. 1: Research area in the German North Sea with marked positions of underwater recorders
(triangle) and positions where pile driving activities were conducted (points). Offshore
wind farms (OWF) which are active to date are marked as solid areas, while OWF, which
are planned or under construction are shown as dashed areas. Dashed lines represent
FFH protected areas (European Union, 1992). A zoom in on the research area (square)
is shown in the top-right figure.
B. Sound propagation modelling and frequency analysis
Single strike sound exposure levels (SELSS) for the full frequency spectrum were calculated for
each measuring position and constructed pile foundation in accordance with German
measurement guidelines (Müller and Zerbs, 2011). All pile driving strikes, which were detected
in the underwater noise recordings over the entire construction period, were analyzed. The
median SELSS of all pile driving strikes per hour (SEL50 h-1) was determined for each station
and pile, describing the accumulated sound energy of these impulsive noise events related to
1 s and the reference pressure of 1 µPa (ISO 18406, 2017).

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
22
The sound propagation, based on the determined median SELSS per hour and distances to the
pile driving site, was estimated by a non-linear regression. The intercept and the logarithmic
regression factor were estimated by a non-linear least squares approach, using the nls function
in R (R Core Team, 2019). Furthermore we estimated the decay factor A in dB per meter within
the nls approach. A weighting was applied to the model regarding the number of pile driving
strikes within the analyzed hour. The received level (RL) was estimated as
𝑅𝐿(𝑅𝐾) = 𝐼𝑛𝑡𝑒𝑟𝑐𝑒𝑝𝑡 − 𝑠𝑙𝑜𝑝𝑒 × 𝑙𝑜𝑔10(𝑅𝑘) − 𝐴(𝑅𝐾) (1),
where Rk is the distance to the pile driving site, the intercept is the intercept of the regression,
the slope is the slope of the regression, which is expected in the range of 10-20 and the decay
factor A, which is a result from multiple reflections from the surface and seabed (Ainslie et al.,
2014; Lippert et al., 2018; Martin and Barclay, 2019; Zampolli et al., 2013). The estimated
propagation parameters slope and A depend on the surface and bottom roughness, the sediment
type and the speed profile within the water column. This sound propagation model was
empirical based, but has to be considered as a broad estimate, because it does not consider any
variations in bottom composition or bathymetry. Although sound propagation is much more
complex and local variability may occur, this simple model enables a conceptual understanding
of the propagation of pile driving noise (Ainslie et al., 2014; Lippert et al., 2018; Martin and
Barclay, 2019).
For comparative reasons, we also modeled a theoretical transmission loss (TL) over distance
(Rk), which would be expected if guidelines are followed. The radiation characteristic of pile
driving noise is considered to be more similar to a line than a point source, since the pile as a
resonating body covers the entire water column in most of the cases. The proposed damped
cylindrical spreading decay formula (Zampolli et al., 2013) considers this sound propagation
and has been shown to be applicable for pile driving noise propagation within the North Sea up
to a distance of approximately 15 km (Lippert et al., 2018). Based on the current German
regulation, we assumed an SELSS of 160 dB re 1µPa²s at a distance of 750 m. The theoretical
received level (RL) at the distance Rk was obtained from equation (1), under the assumption
that pile driving noise follows a damped cylindrical spreading, with Rk as the reference range
of 750 m, where the SELSS is equal to 160 dB re 1 µPa²s. The decay factor was estimated by
an empirical fit within a non-linear model, based on our empirical data of SEL50 h-1 over

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
23
distance for data up to 15 km and the number of pile driving strikes per hour as a
weighting factor. Ambient noise was determined for a 30 s fraction of underwater sound
recordings prior to pile driving activities, when no pile driving strikes occurred. Third octave
spectra (2-base) were calculated for each 10 Hz high pass filtered 1 s window within the time
window, for each pile at each measuring position for center frequencies ranging from 62.5 Hz
to 12.7 kHz. In total, background recordings prior to six pile driving events at two measuring
positions were analyzed, forming a database of eleven 30 s windows.
C. Model assumptions
The calculation of the potential hazard zone for auditory damages was based on several
assumptions, which are described in the following:
A TTSonset at 8 kHz hearing frequency was determined at a cumulative SEL (SELcum) of 175
dB re 1 µPa²s, corresponding to 1385 pile driving strikes of 145 dB re 1 µPa²s (SEL) in
30 minutes (Kastelein et al., 2016).
Single events below a certain SELSS never induce a TTS or affect recovery and are therefore
regarded as ‘effective quiet’ (Finneran, 2015; Ward et al., 1976). To date, no study has
determined an effective quiet threshold for harbor porpoises but the best estimate can be derived
from the lowest SELSS with the potential to induce a TTS, regardless of frequency or duration
(Finneran, 2015). The lowest determined SELSS with the potential to induce a TTS was
determined at 145 dB re 1 µPa²s (Kastelein et al., 2016). In the absence of an empirically
derived effective quiet threshold, we defined instead a threshold which is ‘still higher than
effective quiet threshold’ (SHEQ) as a proxy. A single strike sound exposure level of
145 dB re 1 µPa²s was defined as the SHEQ with the motivation to estimate a potential to cause
a TTS by means of an SELSS, which proved to induce a TTS after multiple reception, instead
of being conservative by considering all exposures. The implication of this is further discussed
in chapter A of the discussion. Pile driving strikes with SELSS below this SHEQ were therefore
excluded from the calculation of cumulative received levels. The SHEQ was used in
combination with the modeled sound propagation, to determine a ‘safe distance’, where SELSS
are below the SHEQ, and will not affect harbor porpoise hearing. The concept of the safe
distance should be understood as a novel approach to estimate hazard zones instead of a fixed

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
24
threshold. Further investigations are critically needed to determine an effective quiet threshold,
in order to replace the SHEQ, which is currently suggested as the best guess.
Data on the swim speed of harbor porpoises are rare due to their inconspicuous lifestyle and
poor accessibility. Only a few studies focused on the analysis of swim speed of harbor
porpoises. Maximum swim speed for animals in human care were determined at 4.3 m s-1 (Otani
et al., 2001) and in the wild at 4.3 and 6.1 m s-1 (Gaskin et al., 1974; Otani et al., 2000). A
maximum swim speed, derived from surfacing positions, was measured at 3.3 m s-1 for free-
ranging animals (Brandt et al., 2013a; Linnenschmidt et al., 2013). Mean swim speed of a free-
ranging harbor porpoise was determined at 0.9 m s-1 (Otani et al., 2000). An estimate of
maximum swim speed endurance has not been published to date. Assumptions on swim speed
are based on these available studies to cover a broad range of possible flight situations.
Accordingly, harbor porpoise flights were simulated at a swim speed of 0.9, 3.3, 4.3 and 6.1 m s-
1.
We selected a pulse interval of 1.3 s which was used in the study by Kastelein et al. (2015,
2016), where the TTSonset for multiple pile driving strikes was taken from and which also fits
well to the analyzed pile driving events. We assumed equal source levels for all pile driving
strikes and SELSS for a certain distance were treated as equal for the whole water column, since
the acoustic field was measured from bottom-mounted sound recorders only. The implication
of this is further discussed in chapter A of the discussion. All model assumptions are shown in
TABLE I.
TABLE I. Summary of variables and values used for the model.
Variable Assumption Value
TTSonset SELcum 175 dB re 1 µPa²s
Effective quiet SELSS 145 dB re 1 µPa²s
Safe distance Distance where SELSS is below effective quiet 5384 m
Swim speed Assumed to be constant 6.1, 4.3, 3.3, 0.9 m s-1
Pulse interval Time between pile driving strikes 1.3 s

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
25
D. Estimation of hazard zones
To estimate hazard zones, where a TTS can be induced by the reception of multiple pile driving
strikes, we defined a ‘safe distance’. At distances larger than the safe distance, SELSS are below
the SHEQ (Finneran, 2015; Ward et al., 1976) and will never induce a TTS or affect recovery,
no matter how many signals will be received. A safe distance was determined using the slope,
intercept and decay factor of the modeled sound propagation.
The received cumulative sound exposure level was calculated as the sum of all received single
strikes a harbor porpoise would receive on a simulated flight track, when swimming straight
away from the sound source up to the determined safe distance. Harbor porpoise positions on
the track were determined by steps with a length according to the given pulse interval of 1.3 s
and the swim speed of the porpoise, straight away from the sound source. The pulse interval
was derived from the analysis of underwater recordings. The expected porpoise position during
the kth pile driving strike, as a distance to the pile driving site (Rk) is thus
𝑅𝑘 = 𝑘𝑝𝑖𝑙𝑒 𝑠𝑡𝑟𝑖𝑘𝑒 × 𝑝𝑢𝑙𝑠𝑒 𝑖𝑛𝑡𝑒𝑟𝑣𝑎𝑙 × 𝑠𝑤𝑖𝑚 𝑠𝑝𝑒𝑒𝑑 + 𝑠𝑡𝑎𝑟𝑡𝑖𝑛𝑔 𝑑𝑖𝑠𝑡𝑎𝑛𝑐𝑒 (2).
SELSS values were calculated as a function of distance to the pile driving position and the
determined sound propagation for all simulated harbor porpoise positions on the flight track
using equation (1).
The received SELcum for the entire flight from a simulated start position up to a distance, where
SELSS is below the effective quiet threshold, can be obtained by
𝑆𝐸𝐿𝑐𝑢𝑚 = 10 × 𝑙𝑜𝑔10 ∑ 10(𝑆𝐸𝐿𝑆𝑆𝑘
10)
𝑛=𝑡𝑜𝑡𝑎𝑙 𝑛𝑜. 𝑜𝑓 𝑠𝑡𝑟𝑖𝑘𝑒𝑠
𝑘=1
[𝑑𝐵 𝑟𝑒 1 µ𝑃𝑎2𝑠] (3)
Based on the aforementioned equations we derived a closed-form solution for the received
SELcum for a fleeing porpoise (see results section). We verified this analytical solution by
simulating fleeing porpoises in which we iteratively summed up received levels of single
strikes. All analyses were performed and figures created using R (R Core Team, 2019).

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
26
III. Results
A. Sound propagation in the study area
A total of 912820 pile driving strikes were detected and analyzed in recordings with pile driving
activity. Inter-pulse intervals varied between 0.8 and 2.2 s. The most distinct inter-pulse interval
was found at 1.0 s, followed by 1.6 s (Ruser et al., 2016b), indicating that the fixed inter-pulse
interval in the simulation approach of 1.3 s was representative for actual pile driving activities.
Energy spectra of 76448 pile driving strikes from six monopiles at two measuring positions
were further analyzed. Most of the energy was found to be below 2 kHz with a peak around
160 Hz (FIG. 2). A further lower peak for frequencies around 5 kHz was also found for some
piles (ABW48, ABW59 in FIG. 2).
FIG. 2: Third octave level spectrum of the analyzed pile driving events, color and symbol coded
for measuring position and pile. Each mark on the line represents the determined median
SELSS for the corresponding center frequency of the third octave band. The median
ambient noise level of all analyzed 30 s recordings prior to pile driving activities is shown
as the lowest line with vertical lines representing the 1.5 interquartile range.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
27
Received levels of single strikes (SELSS) ranged from 101.9 to 165.2 dB re 1 µPa²s at distances
between 2.4 and 36.8 km (see FIG. 3).
FIG. 3: Determined SELSS in dB (re 1 µPa²s) of pile driving activities during the construction
of 50 monopiles in the Amrumbank West OWF.Sound recordings were conducted at seven
static measuring positions. The median SELSS of all detected strikes per hour is marked by
points. Sound propagation was modeled by a non-linear regression (solid line), estimating
the intercept (200.5), slope (-13.64) and decay factor A (0.00078). The theoretical sound
propagation, assuming a damped cylindrical spreading with a received level of
160 dB re 1 µPa²s at a distance of 750 m is presented by the dashed line. The empirical
based best fit for the decay factor A was estimated at 0.0012 dB per meter, which is in line
with the empirical derived decay factors reported by Lippert et al. (2018) The number of
strikes within an hour is shown by color-coded points and its distribution is shown in the
histogram in the top right corner.
The received level (RL) was estimated by a non-linear logarithmic regression, estimating the
intercept, slope and decay factor A in dB per meter based on the determined median SELSS per
hour. We found
𝑅𝐿(𝑅𝑘) = 200.5 − 13.64 × 𝑙𝑜𝑔10(𝑅𝑘) + 0.00078 (𝑅𝑘) [𝑑𝐵 𝑟𝑒 1 µ𝑃𝑎2𝑠] (4) as the
best fit for the analyzed pile driving strikes over distance. The modeled sound propagation is

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
28
shown in FIG. 3 as a solid line. We used the number of pile driving strikes as a weighting factor
for the model. A median of 772 pile driving strikes per hour was found (sd=753, range 1-3586).
The SELSS at a distance of 750 m was estimated at 160.8 dB re 1 µPa²s.The number of strikes
within an hour is shown by color-coded points and its distribution is shown in the histogram in
the top right corner (FIG. 3).
The theoretical sound propagation, assuming a damped cylindrical spreading with a received
level of 160 dB re 1 µPa²s at a distance of 750 m is presented by the dashed line. The best fit
for the decay factor A was estimated at 0.0012 dB per meter, which is in line with the empirical
derived decay factors reported by Lippert et al. (2018). For distances up to 10 km the predicted
sound propagation and the theoretical sound propagation (Lippert et al., 2018; Zampolli et al.,
2013) showed high similarity. The theoretical sound propagation is not reliable for distances
further than 15 km.
B. Estimation of hazard zones
The calculation of the potential hazard zone for auditory damages was based on several
assumptions, which are described in Sec. II.
The potential to induce a TTS in harbor porpoises, which do not show a flight response, was
presented by showing the maximum radius where a TTS can be induced from multiple pile
driving strikes above the SHEQ. The hazard radius corresponds therefore to the determined safe
distance using the slope, intercept and attenuation factor of the modeled sound propagation
along with the effective quiet threshold. Therefore, we re-arranged equation (4) in order to
calculate a safe distance with a given received level, corresponding to the effective quiet
threshold. The rearrangement required the usage of the Lambert W function (Corless et al.,
1996) to solve the equation in which the unknown distance appears both outside and inside a
logarithmic function, leading to
𝑠𝑎𝑓𝑒 𝑑𝑖𝑠𝑡𝑎𝑛𝑐𝑒 =
− {𝑠𝑙𝑜𝑝𝑒 × 𝑊 [−10
−𝑖𝑛𝑡𝑒𝑟𝑐𝑒𝑝𝑡+𝑒𝑞𝑡𝑠𝑙𝑜𝑝𝑒 × 𝐴 × ln (10)
𝑠𝑙𝑜𝑝𝑒]}
𝐴 × ln (10) [𝑚] (5).
To calculate the safe distance, defined as a distance where the single strike level is
145 dB re 1 µPa²s, we inserted the intercept (200.5 dB), slope (-13.64 dB) and the attenuation

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
29
factor A (0.00078 dB m-1) of the determined logarithmic regression of the received levels into
this equation. The safe distance was determined at 5604 m.
According to an assumed pulse interval of 1.3 s, the SELcum would exceed the TTSonset of
175 dB re 1 µPa²s in 21.7 minutes within the area up to the safe distance. Animals located
within the area up to the determined safe distance at 5.6 km are estimated to suffer from a TTS
within a maximum of 21.7 minutes.
By a simulation approach, we determined the minimum distance a harbor porpoise must be
deterred prior to pile driving activities, to escape hearing impairment by a continuous flight up
to the assumed safe distance. Therefore, we modeled the total received SELcum for a complete
flight track from various start positions up to the safe distance at which the received SELSS was
below 145 dB re 1 µPa²s (SHEQ). Results were obtained for all possible start positions ranging
from zero to the safe distance. This simulation was conducted using the variables and their
specified values presented in TABLE I.
Thus, the received SELcum for the entire track of a harbor porpoise swimming straight away
from the sound source from an assumed position (start distance) up to the safe distance
(determined at 5604 m) can be obtained by
𝑆𝐸𝐿𝑐𝑢𝑚 = 10 × 𝑙𝑜𝑔 {10
𝑖𝑛𝑡𝑒𝑟𝑐𝑒𝑝𝑡10
𝑝𝑢𝑙𝑠𝑒 × 𝑠𝑝𝑒𝑒𝑑×
𝐴 × 𝑙𝑛(10)
10
𝑠𝑙𝑜𝑝𝑒10
−1
× [𝛤 (1 −𝑠𝑙𝑜𝑝𝑒
10,𝐴 × 𝑙𝑛(10) × 𝑑𝑖𝑠𝑡. 𝑠𝑡𝑎𝑟𝑡𝑘
10)
− 𝛤 (1 −𝑠𝑙𝑜𝑝𝑒
10,𝐴 × 𝑙𝑛(10) × safe distance
10, )]} [𝑑𝐵 𝑟𝑒 1 µ𝑃𝑎2𝑠] (6).
For the received SELcum we find for 𝛤(𝑎, 𝑥) = ∫ 𝑡𝑎−1∞
𝑥 𝑒−𝑡 𝑑𝑡 as the upper incomplete gamma
function, giving an estimate for any transmission loss factor (slope) between cylindrical
(10×log10) and spherical (20×log10) spreading. To integrate the decay factor A, the incomplete
gamma function had to be included, which arises as a solution for certain integrals. We
calculated the received SELcum for all tested swim speeds with varying start distances
(dist.startk) ranging from 750 m up to the safe distance with equation (6) (see the R-script in
Appendix A).

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
30
Afterwards, we determined the minimum distance at which the TTSonset of 175 dB re 1 µPa²s
(Kastelein et al., 2016) was not exceeded. This distance was defined as the minimum distance
a harbor porpoise must be away from the pile driving site at the moment of the first strike to
reach the safe distance by a continuous flight before the cumulative energy can induce a TTS.
This distance was called the minimum deterrence distance.
Minimum deterrence distances are shown in TABLE II for simulated harbor porpoises
swimming away at four different speed levels. Assuming an immediate flight with 6.1 m s-1
after the first pile driving strike, harbor porpoises have to be further away than 2399 m from
the construction site to successfully prevent a TTS (FIG. 4, TABLE II).
TABLE II. Simulation results for the maximum distance as a start position for a flight, where a
TTS could still be induced before reaching the safe distance. Moreover, the number of
received strikes on the track from the minimum deterrence distance up to the safe distance
are given as same as the time of travelling in minutes. Simulation results are presented for
four swim speed levels.
Swim speed
in m s-1
Min. deterrence
in m
Strikes to safe
distance
Travel time to safe distance
in min
0.9 4659 807 17.5
3.3 3255 547 11.9
4.3 2897 484 10.5
6.1 2399 404 8.8
A harbor porpoise starting a flight at a distance of 2432 m with a constant speed of 6.1 m s-1
can still receive cumulative sound energy exceeding the TTSonset after 8.8 minutes of a
continuous flight and the reception of 405 pile driving strikes in that period. Harbor porpoises,
which are capable of fleeing at a speed of 4.3 m s-1, have to be more than 2897 m away from
the construction site to reach the safe distance before the received SELcum exceeds the TTSonset.
Slowly swimming harbor porpoises, traveling at a speed of 0.9 m s-1 can still suffer from a TTS
when starting a flight at the beginning of pile driving activities at a distance of 4659 m. Within
17.5 minutes, a harbor porpoise fleeing at 0.9 m s-1 could receive a cumulative SEL exceeding
the TTSonset after 808 strikes before reaching the safe distance.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
31
FIG. 4: Simulation harbor porpoise fleeing from a certain start position (x-axis) up to the safe
distance (5604 m). The received SELcum for the complete flight track (y-axis) is color-
coded for each tested swim speed. The horizontal dashed line at 175 dB re 1 µPa²s
indicates the assumed TTSonset..
Single pile driving strikes are limited by legislation to a maximum of 160 dB re 1 µPa²s at a
distance of 750 m in Germany (FIG. 5, green circle). Underwater recordings revealed that up
to a distance of 5604 m, SELSS are high enough, to induce a TTS by multiple exposures (FIG.
5 red circle). We could show by means of a simulation that even harbor porpoises fleeing at a
constant speed of 6.1 m s-1 have to be deterred 2.4 km prior to the first pile driving strike to
successfully reach the safe distance before the TTSonset is exceeded by multiple exposures and
a TTS is likely.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
32
FIG. 5: Radii of hazard zones around a pile driving site, where the TTSonset of 175 dB re 1 µPa²s
can be exceeded by multiple events above the SHEQ of 145 dB re 1 µPa²s for single strikes.
A TTS could be induced in harbor porpoises within 21.7 minutes at a distance up to 5.6 km,
if animals stayed within (outer circle). Harbor porpoises closer than 2.4 km could still
suffer from a TTS, even if fleeing immediately after the 1st pile driving strike with a swim
speed of 6.1 m s-1 (dashed middle circle). The distance of 750 m, where SELSS were
restricted to 160 dB re 1 µPa²s as a protective measure, is shown as the inner circle.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
33
IV. Discussion
In this study, underwater sound recordings of real pile driving events were used in combination
with modeled sound propagation, swim speed and pulse intervals to estimate potential TTS
hazard zones for harbor porpoises. The transmission loss is strongly dependent on site-specific
parameters like frequency of sound, water temperature, salinity, depth, acidity, bottom type and
sea state (Marsh and Schulkin, 1962) and can often, even for single paths, not be extrapolated
to larger distances (Madsen et al., 2006). While the analysis of 912820 analyzed pile driving
strikes enabled us to model an accurate sound propagation within the research area, the
extrapolation of the results of this study to other areas should be made with great caution.
Source levels of pile driving strikes, applied noise mitigation systems and numerous physical
parameters of the acoustic field influence the sound propagation and frequency spectra of these
strikes, which are the driving factors for predicting potential hazard zones. We present a
comprehensive but also parsimonious model, which can easily be adjusted to other areas with
different sound propagation conditions or integrating updated variables.
As a novel approach, we considered an effective quiet threshold to classify areas where a TTS
can be induced by multiple exposures. Since we assume the same pulse interval and only
consider SELSS above the playbacks used in the study by Kastelein et al. (2016), we are
confident that the TTSonset is reliable or could be even lower when exposed to SELSS of higher
energy. Our simulation approach is, however, limited for predictions with pulse intervals, which
are higher than the assumed 1.3 s. The assumption that equal cumulative energy induces an
equal hearing shift is only valid for continuous fatiguing noises or for exposures with similar
duty cycles (e.g. Finneran et al., 2010a, 2010b, Kastelein et al., 2015b, 2014; Mooney et al.,
2009; Popov et al., 2014). Based on the analysis of underwater recordings from 50 monopiles
over a range from 2.4 and 36.8 km, we could show that noise levels are high enough to induce
a TTS by multiple exposures up to a distance of 5604 m.
A. Simulation results
In contrast to direct effects on hearing, behavioral reactions of free-ranging harbor porpoises to
pile driving strikes are not fully understood yet. Tagging of harbor porpoises with high
resolution sound and movement recording tags (DTAG, Johnson et al. 2009) allows for detailed
analysis of behavioral and physiological reactions on an individual basis. Tag-based studies
could show that vessel noise evokes clear behavioral responses in harbor porpoises, coinciding

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
34
with deeper dives, disturbance of foraging, increased fluke strike rates and cessation of
echolocation (Wisniewska et al., 2018b). Since we do not know how harbor porpoises could
react to pile driving noise, we simulated two different scenarios from the perspective of a wild
harbor porpoise exposed to pile driving noise: However, future research is needed to ascertain
their feasibility.
If the porpoise did not show a flight response and stayed inside the area where the noise levels
are high enough to induce a TTS by multiple exposures up to a distance of 5604 m, the received
SELcum would exceed the TTSonset anywhere within this area after 21.7 minutes at a pulse rate
of 1.3 s at the latest. Ecological reasons for staying within hazardous areas could be due to
strong inter-individual variability with animals not responding to that disturbance or tolerating
the noise if staying in an area is beneficial, for instance, because of high quality food. Staying
within a hazardous area could be either a result of natural decision-making or also caused by a
lack of information in which direction to swim. Sound source localization ability of harbor
porpoises has been found to be better for longer signals but has been tested for frequencies
above 16 kHz only (Kastelein et al., 2007). It has not been described whether harbor porpoises
are capable of localizing sound sources at such low frequencies and short duration like pile
driving strikes. The spectral content of sounds determines the ability to localize its origin
(Branstetter and Mercado III, 2006; Kastelein et al., 2005). Therefore, it is reasonable to assume
that localizing signals with higher frequency content is easier for harbor porpoises, like shown
for harbor seals, which are also underwater hearing specialists (Bodson et al., 2007).
Consequently, mitigated pile driving strikes could be even harder to localize, because bubble
curtains are more effective in mitigating frequency content above 1 kHz (Dähne et al. 2017,
FIG. 3).
In order to estimate potential hazard zones for harbor porpoises, which immediately flee after
the first pile driving event, we simulated the received SELcum for a complete flight track up to
the safe distance. We wanted to determine a minimum distance by this approach, a harbor
porpoise must be deterred prior to pile driving activities for in order to prevent a TTS, before
the TTSonset is exceeded. We could demonstrate that a harbor porpoise, which is within a radius
of 2.4 km to the pile driving site, cannot reach the safe distance before the TTSonset is exceeded
by the reception of multiple strikes. The simulated fleeing harbor porpoise received pile driving
strikes from behind. Although hearing in harbor porpoises is directional with better abilities for

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
35
signals ahead (Kastelein et al., 2005), our simulation is assumed to be valid. The receiving beam
of harbor porpoises is wider for lower frequencies (Kastelein et al., 2005) and the TTSonset was
derived from a study where harbor porpoises were exposed from varying positions while
swimming freely in a pool (Kastelein et al. 2015, 2016). Our simulation approach cumulates
multiple received pile driving strikes, which are equal to or higher than the SELSS used in a
playback experiment with animals in human care, in which a TTSonset was determined at 175 dB
re 1 µPa²s (Kastelein et al., 2016). Within that playback study, SELSS were the same throughout
the whole exposure. A fleeing harbor porpoise would, however, receive pile driving strikes with
decreasing SELSS with increasing distance to the sound source. In contrast to Kastelein et al.
(2016), the harbor porpoise would receive multiple strikes with variable SELSS, which are all
above the SELSS in that playback experiment. It is reasonable to assume that this could lead to
a lower TTSonset, because the TTSonset is dependent on the duty cycle, SPL and received SELcum
(Kastelein et al., 2015b, 2014b). However, a TTSonset for varying or decreasing SELSS over time
has not been determined yet.
Although effects of pile driving noise on harbor porpoise sightings and acoustic detections have
been described (Brandt et al., 2018; Dähne et al., 2013), behavioral reactions of harbor
porpoises regarding swim speed, echolocation behavior, diving depth and duration remain
unknown to date. In order to protect its hearing, harbor porpoises could flee close to the surface
or bottom, to benefit from interferences, mitigating received SELSS by reflections (Lloyd mirror
effect). This could be the case for described reactions of a harbor porpoise towards high levels
of vessel noise, which remained close to the bottom during highest levels of exposure
(Wisniewska et al., 2018b). The actual swimming depth of harbor porpoises within the hazard
area can highly change the received exposure level, which accordingly affects estimated hazard
zones. The modeled sound propagation has been derived from recordings of bottom mounted
underwater sound recorders, limiting the validity for predictions for the entire water column.
There are only few data available for the variability of received levels of pile driving strikes in
the water column. Received levels of pile driving strikes at distances between 3.8 and 14.6 km
from the pile driving source showed a depth dependency with a variability of up to 5.8 dB
between received levels at 1 and 13 m from the bottom (total depth 23 m, Gerdes et al. (2016)).
The received levels were highest at the bottom, decreased with distance to the surface and
displayed also a slight tendency for the level difference to decrease with increasing distance to

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
36
the pile (Gerdes et al. 2016). However, this limitation within the simulation approach has to be
reconsidered if more information on harbor porpoise reactions to pile driving noise is reported.
B. Application of evaluation tool
This study presents a novel approach to evaluating the impact of anthropogenic impulsive noise
on harbor porpoise hearing, by considering a distance where noise exposure does not affect
hearing anymore. Equation (6) can be applied to calculate the received cumulative SEL on a
flight up to the safe distance, and can be easily adjusted to different sound propagation
measurements. Furthermore, frequency weighting functions can be easily applied to the
TTSonset and the effective quiet threshold, if perceived loudness proves to be the best predictor
for auditory impairment (Houser et al., 2017; Kastelein et al., 2017; Southall et al., 2019;
Tougaard and Dähne, 2017). All presented simulations involve simplifications and assumptions
leading to non-negligible uncertainty in the estimated hazard zones.
As an example, we applied the formula to estimate hazard zones on reported SELSS of further
offshore wind farms in the German North Sea. Based on underwater recordings during the
construction of seven offshore wind farms in the German North Sea, average SELSS of 168 dB
re 1 µPa²s could be determined for noise mitigated pile driving events at a distance of 750 m
(Brandt et al., 2018). The SELSS of unmitigated pile driving strikes at a distance of 750 m was
determined at 175 dB re 1 µPa²s in the same study. The sound propagation determined by
Brandt et al. (2018) was very similar to the sound propagation modeled in this study. Using the
SELSS results of Brandt et al. (2018) for mitigated pile driving strikes and assuming the same
sound propagation as in this study, the safe distance would be at 10.4 km. The minimum
deterrence distance, derived from our simulation approach, would be between 6.6 and 9.4 km
for noise mitigated pile driving events, depending on swim speed. The safe distance for
unmitigated pile driving strikes would be at 15.8 km and minimum needed deterrent distances
ranged between 11.7 and 14.8 km, depending on swim speed. A clear deterrence effect was
found for SELSS above 143 dB re 1 µPa²s, reaching up to distances of 17 km (Brandt et al.,
2018). These findings are in line with the determined threshold at which harbor porpoises in
human care began to respond to pile driving playbacks with porpoising behavior (Kastelein et
al., 2013). These behavioral thresholds of about 145 dB re 1 µPa²s correspond to the assumed
SHEQ, which determines the safe distance and the lowest level of single strikes, which were
taken into account. The safe distance was determined at 15.8 km, whereas the deterrence range

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
37
was measurable up to 17 km (Brandt et al., 2018). Due to a high similarity in results, we assume
that the evaluation tool allows for a reliable prediction of potential hazard zones where a
temporary threshold shift can be induced by multiple pile driving strikes.
Although attempts are made to protect marine mammals from injuries worldwide, national
policies disagree in the definition of injury. While Germany considers a temporary threshold
shift as an injury, most other states define injury in the context of hearing as a permanent
threshold shift (e.g., U.S. Marine Mammal Protection Act in the United States). Recently,
updated noise exposure criteria have been proposed to predict the onset of auditory effects in
marine mammals (Southall et al., 2019). The SEL onsets are presented as weighted levels,
accounting for the frequency dependent effects in order to better meet perceived loudness of
the animal. The harbor porpoise belongs to the group of the very high frequency cetaceans
(VHF) in the recommended marine mammal noise exposure criteria (Southall et al., 2019). The
weighted onset was determined at 155 dB re 1 µPa²s for PTS and at 140 dB re 1 µPa²s for TTS.
In order to adopt these suggested PTS- and TTSonset thresholds in our simulation approach, the
SHEQ also has to be adjusted to the frequency weighting. The SHEQ is based on the playback
study by Kastelein et al. (2016), which were found to be 40 dB lower with applied frequency
weighting (Tougaard and Dähne, 2017, see table 1). Accordingly, the weighted threshold was
also decreased by 40 dB, down to 105 dB re 1 µPa²s.
Within the analysis of pile driving recordings in this study, frequency weighting was not
included in the project scope. Alternatively, published acoustic properties of pile driving strikes
recorded in the offshore DanTysk wind farm can be consulted (Dähne et al., 2017) to estimate
the potential to induce a PTS or TTS for a frequency-weighted sound propagation. A frequency-
weighted source level of 170 dB re 1 µPa²s was estimated from pile driving activities of 80
piles, measured at distances between 1 and 31 km, by fitting a simple transmission loss model
with 15×log10(R) and no absorption (Dähne et al., 2017). Accordingly, the calculated safe
distance extends to 21.5 km for this data-set, consisting of unmitigated, single- and double
bubble curtain-mitigated strikes. To escape a PTS, a minimum deterrence distance between 57
and 1155 m (TABLE III) would be needed for swim speeds between 6.1 and 0.9 m s-1, following
the criteria of Southall et al. (2019). The hazard zone where a PTS can be induced for fleeing
harbor porpoises extends to 1.2 km in the worst case scenario with a swim speed of 0.9 m s-1.
Regarding the reported effectiveness of previous deterrence (Brandt et al., 2013b, 2013a), a
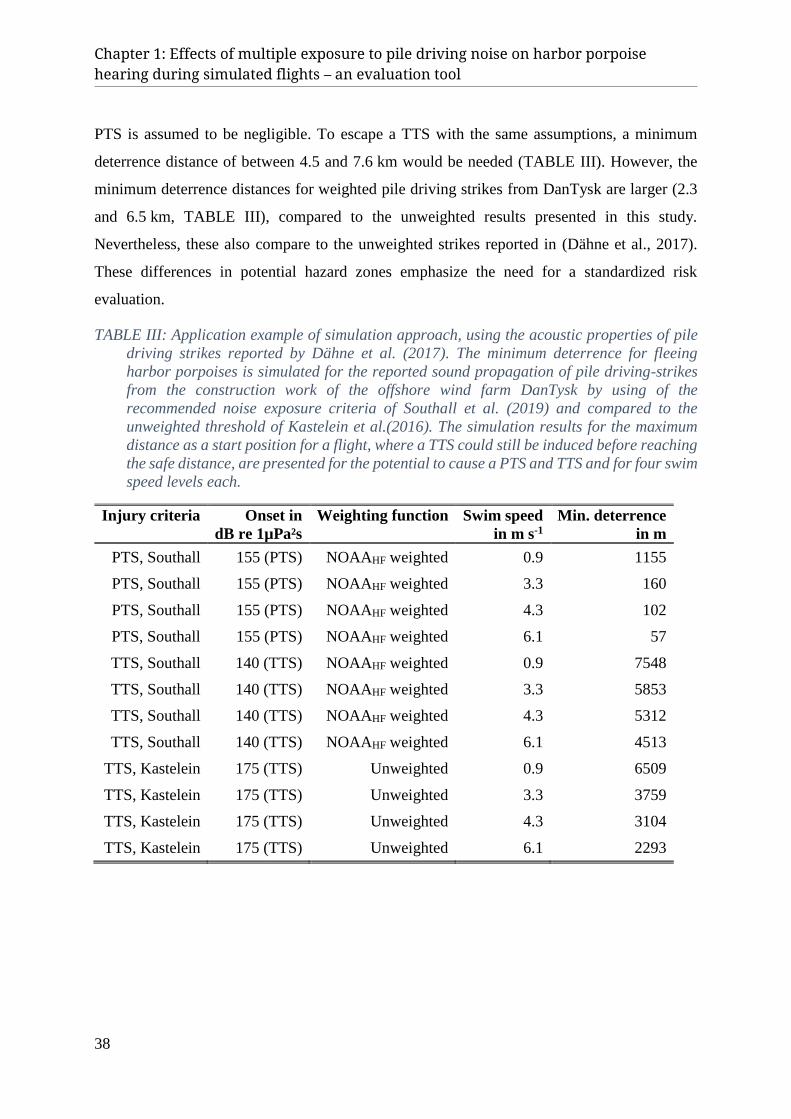
Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
38
PTS is assumed to be negligible. To escape a TTS with the same assumptions, a minimum
deterrence distance of between 4.5 and 7.6 km would be needed (TABLE III). However, the
minimum deterrence distances for weighted pile driving strikes from DanTysk are larger (2.3
and 6.5 km, TABLE III), compared to the unweighted results presented in this study.
Nevertheless, these also compare to the unweighted strikes reported in (Dähne et al., 2017).
These differences in potential hazard zones emphasize the need for a standardized risk
evaluation.
TABLE III: Application example of simulation approach, using the acoustic properties of pile
driving strikes reported by Dähne et al. (2017). The minimum deterrence for fleeing
harbor porpoises is simulated for the reported sound propagation of pile driving-strikes
from the construction work of the offshore wind farm DanTysk by using of the
recommended noise exposure criteria of Southall et al. (2019) and compared to the
unweighted threshold of Kastelein et al.(2016). The simulation results for the maximum
distance as a start position for a flight, where a TTS could still be induced before reaching
the safe distance, are presented for the potential to cause a PTS and TTS and for four swim
speed levels each.
Injury criteria Onset in
dB re 1µPa²s
Weighting function Swim speed
in m s-1
Min. deterrence
in m
PTS, Southall 155 (PTS) NOAAHF weighted 0.9 1155
PTS, Southall 155 (PTS) NOAAHF weighted 3.3 160
PTS, Southall 155 (PTS) NOAAHF weighted 4.3 102
PTS, Southall 155 (PTS) NOAAHF weighted 6.1 57
TTS, Southall 140 (TTS) NOAAHF weighted 0.9 7548
TTS, Southall 140 (TTS) NOAAHF weighted 3.3 5853
TTS, Southall 140 (TTS) NOAAHF weighted 4.3 5312
TTS, Southall 140 (TTS) NOAAHF weighted 6.1 4513
TTS, Kastelein 175 (TTS) Unweighted 0.9 6509
TTS, Kastelein 175 (TTS) Unweighted 3.3 3759
TTS, Kastelein 175 (TTS) Unweighted 4.3 3104
TTS, Kastelein 175 (TTS) Unweighted 6.1 2293

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
39
C. Ecological relevance of disturbance
The TTSonset derives from a study, which determined a statistically significant TTS after
multiple exposure to pile driving playbacks, which was small and could only be measured due
to quiet experimental pools and low variability (Kastelein et al., 2016). However, a statistically
significant TTS does not inevitably mean it is ecologically significant. To date, it is still
unknown what the ecological effects of TTS are. Nevertheless, it is assumed that these are
related to the duration, affected frequency range and magnitude (Kastelein et al., 2017).
Potential consequences could be reflected in difficulties to hear in noisy environments. The
acoustical perception of the environment is of key importance for harbor porpoises when
navigating (Villadsgaard et al., 2007), finding and catching prey (DeRuiter et al., 2009;
Wisniewska et al., 2016) and for intra-specific communication (Clausen et al., 2010; Sørensen
et al., 2018) and any impairment could potentially negatively affect individual fitness,
reproduction or survival. Anthropogenic noise does not inevitably lead to individual mortality
but can affect the behavior of individuals causing sublethal effects (Pirotta et al., 2015). Noise
exposure can also lead to indirect mortality caused by stress responses, affecting physiology
(Aguilar de Soto et al., 2016; Wright et al., 2007). Effects can occur in various forms and can
therefore be interacting or cumulative (Kunc et al., 2016).
The here tested maximum swim speed of 6.1 m s-1 (Gaskin et al., 1974) probably overestimates
the average speed of harbor porpoises, determined as being between 0.7 and 2.2 m s-1
(Linnenschmidt et al., 2013; Otani et al., 2000) or between 1.3 and 3.2 m s-1 during flight
responses to seal scarer signals (Brandt et al., 2013a). Harbor porpoises in human care were
capable of maintaining an increased swim speed of 2 m s-1 throughout 30 minutes, when
exposed to pile driving playbacks with an SELSS of 145 dB re 1 µPa²s (Kastelein et al., 2018b).
The extent to which swim speed levels increase in wild animals is essential for predicting
energetic costs. The resulting drag from moving in a medium increases with the square of swim
speed and likewise the needed costs of locomotion for propulsion against the drag (Gallagher
et al., 2018; van der Hoop et al., 2014). Harbor porpoises live on an energetic knife edge, which
makes them particularly vulnerable to anthropogenic disturbance (Wisniewska et al., 2016).
Therefore, every extra needed food intake due to increased energetic costs and missed time for
foraging during a disturbance could have a severe impact.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
40
Although there is no common EU regulation concerning noise mitigation during the
construction of offshore wind farms, wind farm industries are obliged in several European
countries to actively mitigate noise emission (Belgium, Denmark and Germany and the
Netherlands) or to restrict piling activities to designated time periods (Belgium, the
Netherlands). German legislation has enforced strictest regulations in the EU to date, by
determining a maximum single strike SEL of 160 dB re 1 µPa²s and a maximum SPL of
190 dB re 1 µPa at a distance of 750 m, obligatory deterrence of harbor porpoises prior to piling
activities and using a soft start procedure with limited force and longer pulse intervals. Our
analysis of the recorded pile driving strikes show that the protective measure of restricting
SELSS to 160 dB re 1 µPa²s in 750 m, in order to protect harbor porpoises from TTS by single
strikes, was respected. Additionally, this measure is very effective in reducing the potential
hazard zones, where a TTS can be induced by multiple exposures. We could show that
deterrence of at least 2.4 km prior to pile driving strikes is necessary to allow a flight of harbor
porpoises up to the safe distance before the TTSonset is exceeded. In order to deter harbor
porpoises, seal scarers are deployed prior to piling activities. The effective deterrence range of
a commercial seal scarer (Lofitech, Leknes, Norway) was measured as being 1.9 km by visually
observing surfacing positions (Brandt et al., 2013a). The available data within the piling
protocols concerning the soft start was incomplete. Neither the duration of the soft start nor the
pulse interval or used strike energy is regulated in Germany. In combination with the soft start
in the beginning of pile driving activities with restricted force and longer pulse intervals, this
previous deterrence could be sufficient to allow for a continuous flight up to the safe distance
at 5604 m before TTSonset is exceeded. However, effectiveness of deterrence effort has to be
monitored if all harbor porpoises should be protected. Only combining noise mitigation and
deterrence efforts within current regulations in Germany could be sufficient to enable harbor
porpoises to flee in time, but this has to be monitored for effectiveness. Deterrence should be
great enough prior to pile driving activities (minimum deterrence distance) to give harbor
porpoises sufficient time to flee to areas where no TTS can be induced. However, on the other
hand, it has to be as low as possible, to reduce temporary habitat loss.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
41
D. Conclusion
Our approach to evaluating the potential for causing a TTS from multiple pile driving events
can be easily adjusted to different areas with other sound propagation characteristics.
Furthermore, it can be adjusted for other sound signals or species with different TTSonsets or for
an updated TTSonset if further studies are conducted, using variable SELSS for example. Our
simulations show that implemented measures during the construction of OWF in Germany
represent a valuable tool for protecting harbor porpoises not only from single but also from
multiple pile driving strikes. The deterrence prior to pile driving events is particularly important
and has to be monitored to give harbor porpoises sufficient time to leave hazardous areas at
moderate speeds. Based on our simulation approach, only the combination of restricting the
maximum SELSS to 160 dB re 1 µPa²s at a distance of 750 m, a previous deterrence and a soft
start with reduced energy and longer pulse intervals allow harbor porpoises to avoid a TTS from
multiple exposures. However, deterrence efficiency has to be monitored.
E. Acknowledgement
The underwater sound recordings for this study were obtained as part of the research carried
out during the projects “Impacts of underwater noise on marine vertebrates - Cluster 7” (Z 1.2–
53302/2010/14) and “Impact of underwater noise on marine mammals (UWE)” FKZ 3515 82
2000), funded by the Federal Agency for Nature Conservation (BfN). We would like to thank
Matthias Fischer and Max Schuster from DW-ShipConsult GmbH (Germany) for making and
analyzing underwater recordings and Dominik Nachtsheim for his support in generating
figures. We are very thankful for the proofreading of Frances Sherwood-Brock and her valuable
comments. We wish to express our gratitude to the legacy of Rodney Coates for awarding the
first author for the presentation of this study at the Effects of Noise on Aquatic Life Conference
in 2019 in The Hague (the Netherlands). Also we would like to thank the “Society of Friends
of the University of Veterinary Medicine Hannover (Gesellschaft der Freunde der
Tierärztlichen Hochschule Hannover e.V.” for funding the attendance of the first author at this
conference.

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
42
F. APPENDIX
R commands to calculate the safe distance and received SELcum on flight track
# Define base parameters
sheq <- 145 # effective quiet threshold
pulse <- 1.3 # interpulse interval
TTS_onset <- 175 # TTS onset for multiple exposure with pile driving strikes
slope <- -13.64 # slope of regression
intercept <- 200.5 # intercept of regression, estimated SL
A <- 0.00078 # attenuation factor in dB/m, absorption integrated
# calculate safe distance, where SELss is 145 dB, corresponds to equation (6) in manuscript
library(lamW)
safe_distance <-- (slope*lamW::lambertW0(-(10^((-intercept+sheq)/slope)
*A*log(10))/slope))/(A*log(10))
print(safe_distance) # 5604m
Rk <- safe_distance # calculate SELss for safe distance to cross check
SELss <- intercept+slope*log10(Rk)-A*Rk
print(SELss) #must be equal to sheq
# Calculate cumulative SEL for all received pile driving strikes on flight track up to the safe distance
# corresponds to equation (6) in the manuscript and results presented in table II
library(pracma)
dist.start <- 2399 #corresponds to minimum deterrence distance, SELcum must be equal to TTS onset
speed <- 6.1 #m/s, highest reported swim speed
max_sel_cum <- 10*log10(10^(1/10*intercept)/(pulse*speed)*(-1)*(A/10*log(10))^(-slope/10-
1)*(incgam(x=A/10*log(10)*safe_distance,a=1+slope/10)-
incgam(x=A/10*log(10)*dist.start,a=1+slope/10)))
print(max_sel_cum) # 175 dB re 1 µPa²s, TTS onset, minimum deterrence distance

Chapter 1: Effects of multiple exposure to pile driving noise on harbor porpoise
hearing during simulated flights – an evaluation tool
43
# simulation with multiple swim speeds, generate results for table II
speed_all <- c(6.1,4.3,3.3,.9) # all simulated speed levels
for(s in 1:length(speed_all)){
speed <- speed_all[s]
hp.pos <- seq(from=750, by=speed*pulse, to = safe_distance) # harbor porpoise positions at
time of strikes
max_SEL_cum<-c()
for (i in 1:length(hp.pos)){
max_SEL_cum[i]<- 10*log10(as.numeric(sum((10^( (intercept+slope*log10(hp.pos)-
A*hp.pos) /10))[length(hp.pos):i])))
}
det <- (hp.pos[max(which(max_SEL_cum>TTS_onset))]) #minimum deterrence distance to
avoid TTS
strikes <-((safe_distance-det)/speed)/pulse #number of strikes
time <-((safe_distance-det)/speed) #time in s
print(c(det,strikes,time/60)) #results of table II
}

The use of seal scarers as a protective mitigation measure can induce hearing
impairment in harbour porpoises.
44
Published:
Reproduced from,
Schaffeld, T., Ruser, A., Woelfing, B., Baltzer, J., Kristensen, J.H., Larsson, J., Schnitzler,
J.G., Siebert, U. (2019). The use of seal scarers as a protective mitigation measure
can induce hearing impairment in harbour porpoises. Journal of the Acoustical
Society of America, 146 (6), 4288-4298, https://doi.org/10.1121/1.5135303
with the permission of the Acoustical Society of America.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
45
Chapter 2: The use of seal scarers as a protective mitigation measure can
induce hearing impairment in harbour porpoises
T. Schaffeld1, A. Ruser1, B. Woelfing1, J. Baltzer1, J.H. Kristensen2, J. Larsson2, J.G.
Schnitzler1, U. Siebert1
1 Institute for Terrestrial and Aquatic Wildlife Research, University of Veterinary Medicine
Hannover, Foundation. Werftstr. 6, 25761 Büsum, Germany
2 Fjord&Bælt, 5300 Kerteminde, Denmark
This paper is part of a special issue on The Effects of Noise on Aquatic Life.
Abstract
Acoustic deterrent devices (ADDs) are used to deter seals from aquacultures but exposure of
harbour porpoises (Phocoena phocoena) occurs as a side-effect. At construction sites, by
contrast, ADDs are used to deter harbour porpoises from the zone in which pile driving noise
can induce temporary threshold shifts (TTSs). ADDs emit such high pressure levels that there
is concern that ADDs themselves may induce a TTS. A harbour porpoise in human care was
exposed to an artificial ADD signal with a peak frequency of 14 kHz. A significant TTS was
found, measured by auditory evoked potentials, with an onset of 142 dB re 1 lPa2s at 20 kHz
and 147 dB re 1 lPa2s at 28 kHz. The authors therefore strongly recommend to gradually
increase and down regulate source levels of ADDs to the desired deterrence range. However,
further research is needed to develop a reliable relationship between received levels and
deterrence.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
46
Author contributions
A.R. and U.S. were involved in designing the project and grant acquisition. The experimental
design was developed by A.R. with support of T.S. The experiments with the harbour porpoise
in human care were conducted by T.S. and A.R.. The animal was trained by J.H.K and J.L.,
who also monitored and instructed the animal during the experiments. T.S. analysed the data
with support from A.R. and J.S. T.S. was supervised by A.R., J.S. and U.S. The manuscript was
written by T.S.. All authors provided critical feedback and helped to shape the research, analysis
and contributed to the final version of the manuscript.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
47
I. Introduction
Acoustic deterrent devices (ADDs, e.g. seal scarers) are applied in two different scenarios.
These devices are mainly applied to deter seals from fish farms with the aim to prevent
economic loss, due to seal depredation and damage to fishing gear. In European areas with
extensive fish farms (e.g. along the west coast of Scotland), ADDs are a chronic source of
anthropogenic noise pollution (Findlay et al., 2018). Coastal areas where aquacultures are
located, are a typical habitat of harbour porpoises (Phocoena phocoena) (Brandt et al., 2018;
Findlay et al., 2018; Gilles et al., 2016; Hammond et al., 2017; Peschko et al., 2016; Viquerat
et al., 2014). Harbour porpoises have a very wide hearing range and are capable of hearing seal
scarer signals (Kastelein et al., 2002; Ruser et al., 2016a). While ADDs show highly varying
success in seal deterrence (Götz and Janik, 2013) and could even attract animals, harbour
porpoises exhibit strong avoidance reactions (Mikkelsen et al., 2017). The much further
deterrence of harbour porpoises occurs as an unwanted side effect in cases where ADDs are
deployed to deter seals from fish farms (Brandt et al., 2013a).
The second scenario of ADD application is the use as a deterrent device prior to pile driving
activities in offshore wind farms with the aim to deter harbour porpoises as a target species.
Offshore wind farm construction is substantially increasing in Europe, providing a promising
alternative to fossil fuels and nuclear power. In total, 92 offshore wind farms have been
constructed to date in 11 European countries including sites with partial grid connection,
amounting to 4,149 connected wind turbines (Remy and Mbistrova, 2018). The majority of
wind turbines are bottom mounted. High levels of impulsive noise arise, when piles are driven
into the seabed (Bailey et al., 2010; Brandt et al., 2018; Tougaard et al., 2009). The increasing
human encroachment which accompanies the construction of offshore wind farms has negative
effects on key ecological species such as the harbour porpoise (Brandt et al., 2013b, 2018;
Dähne et al., 2013; Tougaard et al., 2009), which inhabits coastal areas.
Due to its high sensitivity towards anthropogenic noise, the harbour porpoise can be regarded
as an indicator species in noise impact evaluations (Southall et al., 2007; Tougaard et al., 2015).
Harbour porpoises face the risk of a temporary hearing impairment, if they stay in areas close
to the pile driving site, since impulsive noise has the potential to induce a temporary threshold
shift from single (Lucke et al., 2009) or multiple exposure (Kastelein et al., 2015a, 2016).
Implementation of noise exposure criteria into national legislation differs between countries

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
48
(Stöber and Thomsen, 2019). In particular, injury is defined differently between national
policies (e.g. by the Habitats Directive in Europe or by the U.S. Marine Mammal Protection
Act in the United States). In Germany, a temporary loss of hearing after the exposure to pile
driving noise (temporary threshold shift, TTS) is considered as an injury (BMU 2014), whereas
most other states in Europe or in the United States only regard a permanent threshold shift
(PTS) as an injury. Harbour porpoises are protected throughout Europe and are listed in Annex
II and IV of the European Union Habitats Directive (Council Directive 92/43/EEC on the
Conservation of natural habitats and of wild fauna and flora).
Although several countries established criteria frameworks, which define limits for hearing
impairments, regulations pertaining to the use of deterrent devices differ between countries
(reviewed for Germany, Denmark and United States in Stöber and Thomsen (2019). In German
waters single strike sound exposure level (SELSS) of 160 dB re 1 µPa²s and a peak pressure
level (Lp) of 190 dB re 1 µPa must not be exceeded at a distance of 750 m to the pile driving
site (BMU, 2014). In order to prevent physical damage harbour porpoises must be deterred from
the near-field, where sound exposure thresholds can exceed the threshold (BMU, 2014).
Following German legislation, permissions for offshore wind farm constructions generally
include the condition to deter harbour porpoises prior to pile driving activities (BMU, 2014).
ADDs are regularly applied as a tool to deter harbour porpoises prior to pile driving activities
(Brandt et al., 2018). ADDs emit signals between 10 to 40 kHz, corresponding to the range of
best under water hearing in seals (Kastak and Schusterman, 1998; Kastelein et al., 2009b,
2018a; Reichmuth et al., 2013), at very high source levels up to 193 dB re 1 µPa (Lepper et al.,
2004). Seal scarer signals leads to deterrence effects in harbour porpoises, measured by
surfacing distance to sound source (Brandt et al., 2013a; Mikkelsen et al., 2017), aerial surveys
(Brandt et al., 2013b) or by echolocation activity (Brandt et al., 2013b).
The aim of this study was to test, whether a seal scarer has the potential to induce a TTS in
harbour porpoise hearing. Additionally, we determined the threshold distance, at which single
exposures to seal scarer signals can induce a TTS, in order to estimate hazard zones for seal
scarers. We provide critically needed information to develop protective measures, which ensure
both, sufficient deterrence and the avoidance of hearing impairment inflicted by ADD signals.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
49
II. Material & Methods
A. Study area and animal subject
All measurements were conducted at the Fjord & Bælt centre in Kerteminde (Denmark). One
harbour porpoise was kept in the 36×15 m semi-natural outdoor enclosure during this study.
The enclosure was constructed of nets with a mesh size of 10 cm², allowing for a natural flow
of seawater from the Kerteminde Fjord and the Great Belt. Water depth within the enclosure
varied between three and four meters, depending on position and tide. Measurements were
conducted in a 6×4×1.9 m floating holding pool. A gate on one side was always open, allowing
the animal to leave whenever it wanted. The study subject was a female harbour porpoise born
in 1995, which has been kept in human care at Fjord & Bælt since 1997, after it was rescued
from a pound net.
The harbour porpoise is kept under human care by the Fjord&Bælt in Kerteminde (DK) under
permit no. SVANA-610-00084 from the Danish Ministry of Food, Agriculture and Fisheries.
All trials were conducted adhering to the respective ethical principles as well as to the relevant
international and national guidelines for animal experiments and under constant supervision of
experienced biologists animal trainers. Experienced animal trainers monitored animal condition
and signs of stress of the animal throughout all experiments. The harbour porpoise has been
trained two times a day through standard operant conditioning and positive reinforcement
techniques by a team of professional animal trainers. Trials were only conducted, if visibility
allowed for observing the harbour porpoise underwater at the bite plate throughout the whole
experiment. During all exposure sessions, the animal was visually observed by the trainer from
above and additionally recorded by an underwater camera. In case of an observed stress
response the experiments would have been stopped immediately.
B. Background noise recordings
Vessels passing close to the enclosure increased background noise levels, since the semi-natural
enclosure was solely separated from the harbour of Kerteminde by nets. Therefore, background
noise was continuously monitored during measurements. Experiments were only conducted, if
no vessel was passing by or no other unwanted noise source was present. Background noise
was recorded using a custom-made software application (LabVIEW, USA) with a hydrophone
(TC4032, Reson Teledyne, DK), pre-amplified by 20 dB (ETEC amplifier, DK) and band pass
filtered (100 Hz-180 kHz, ETEC 1501, DK). Recordings were digitized with a data acquisition

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
50
card (NI USB 6251, National Instruments, USA) sampling at 400 ksamples s-1 and at a 16 bit
resolution.
C. Experimental procedure to measure hearing thresholds
Hearing thresholds were measured by monitoring the auditory evoked potential (AEP)
response, a commonly used non-invasive electrophysiological technique (Finneran, 2018;
Nachtigall et al., 2017; Ruser et al., 2016a). In principle, a hearing stimulus (1) is presented to
the test individual. If the acoustic stimulus is above threshold levels, the neurons within the
acoustic pathway are stimulated and the neuronal discharges can be detected by electrodes
placed on the head (2).
1. Hearing test stimulus
Hearing thresholds of the animal were determined at 20 and 28 kHz. Short tones centred at
these frequencies were emitted, while auditory evoked potentials of the brainstem were
simultaneously recorded. A rugged notebook (Panasonic Toughbook CF30) computer was used
to digitally generate stimuli, which were converted to analogue by a USB multifunction data
acquisition card (NI USB 6251, National Instruments, USA). The stimuli were updated at a
1 MHz rate with a 16 bit resolution and band pass filtered (100 Hz-250 kHz, 24 dB/octave,
Krohn Hite, USA) before emission by the transducer. The generated stimuli were emitted by a
TC4033 transducer (Teledyne Reson, DK), placed at depth of 0.8 m and a distance of 1 m in
front of the animal. Generated stimuli consist of 1024 tone pips for each tested sound intensity.
Each pip consists of two sine rise, one sines steady and two sines fall, with an epoch length of
17 m s (FIG. 6). Accordingly, the exposure per tested sound pressure level (SPL) had a duration
of 17.4 s.
The targeted maximum dive duration of 40 s allowed for testing two different SPL values per
dive. All playbacks of hearing test stimuli and recordings of auditory evoked potentials were
conducted with a custom written software (Evoked Response Study Tool, EVREST, (Finneran,
2009; Finneran et al., 2008). Received SPL of test stimuli were calibrated by averaging 1024
stimuli prior to each trial at an SPL of 111 dB re 1 µPa with a TC4013 hydrophone (Teledyne
Reson, DK) placed at the bite plate, close to the position of the porpoise during stimulus
presentation. Recorded stimuli signals were pre-amplified by 60 dB (ETEC amplifier, DK),
band pass filtered from 1 to 180 kHz (ETEC B 1501, DK) and then digitized at 500 kHz with a
16 bit DAQ-card (NI USB 6251, National Instruments, USA).

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
51
FIG. 6: Tone pips, used as hearing stimuli. A) Section with ten tone pips out of a complete train
consisting of 1024 repetitions. B) Detail view of one pip (five cycles of 20 kHz carrier)
within the pip train. C) Frequency spectra of the 20 and 28 kHz tone pips with -3 dB
bandwidth levels (grey horizontal lines).
2. Brain potential acquisition and hearing sensitivity assessment
Auditory evoked potentials were measured using 10 mm silver-plated electrodes, imbedded in
suction cups, which were gently attached to the body surface along with standard conductive
gel, which is regularly used in human electroencephalography (EEG) recordings. Small
electrical charges, generated by the brain in response to the acoustic stimuli could be measured
by placing the active (+) electrode behind the blowhole, the inverting electrode (-) along the
dorsal midline of the porpoise between the blowhole and dorsal fin and the ground electrode
(⏚) on top of that suction cup. Impedance between electrodes was tested with an impedance

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
52
meter (Temec Instruments BV, NL) to ensure that impedance was <3 kOhm during all
measurements. Electrodes were connected with 10 m long shielded cables to a bio-potential
amplifier (CP511, GRASS Technologies, USA) in order to amplify (+100 dB) and filter (0.3-
3 kHz bandpass filter) the measured voltage between the inverting and non-inverting electrodes.
The signal was digitized at 50 kHz and 16 bit resolution by the data acquisition card, connected
to the computer. AEPs were recorded with the custom-made software (EVREST) that was also
used to emit stimuli.
3. Experimental set up
First, the animal had to get accustomed to wear suction cups on the skin. Suction cups contained
silver-plated electrodes, which were attached via cables to the computer. Second, the animal
was trained to dive to a bite plate at a depth of 0.8 m and be stationary.
Two separate stations, approximately 4 m apart at the right and left corner of the holding pool
were used for the trials. Both bite plates were positioned at a fixed distance of 1 m to the
projecting transducer. At the left bite plate, the animal could be exposed to the fatiguing sound,
hearing tests were conducted solely at the right bite plate. Each trial to determine a hearing
threshold included four dives to the bite plate, each lasting 40 s. During a dive, two different
sound amplitudes could be tested, allowing for testing eight intensities in four dives. Sound
amplitude started at 80 dB re 1 µPa [root-mean square (rms)] for baseline measurements and
was increased by the EVREST software internally in steps of 5 dB up to 120 dB re 1 µPa (rms).
D. Fatiguing stimulus and experimental procedure
We aimed at testing, if seal scarer signals can induce a temporary hearing shift in harbour
porpoises. Therefore, we exposed the animal to artificial seal scarer signals at a range of sound
exposure levels and tested, if post-exposure hearing thresholds differed from baseline hearing
thresholds. One complete session consisted of three trials, which were conducted on the same
day. Three trials were needed to determine 1) the pre-exposure baseline hearing at experimental
day, 2) the post-exposure hearing and 3) the recovery. In trials 1 and 3 a hearing threshold was
determined as described, whereas prior to trial 2, the animal was exposed to a fatiguing
stimulus. During all exposure sessions, the animal was visually observed by the trainer from
above and additionally recorded by an underwater camera.
In all trials, the harbour porpoise was first sent to the exposure station, at the left bite plate. The
animal stayed there upon receiving an acoustic signal from the trainer. For the baseline hearing

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
53
or recovery measurements, the animals was not exposed, but prior to exposure trials, the animal
received a fatiguing stimulus, while staying at the bite plate. To allow for a complete recovery
of the hearing system before the next trial the animal was exposed only once a day. Recovery
measurements of the hearing thresholds took place two hours after exposure to the fatiguing
stimulus.
We exposed the animal to an artificial seal scarer signal, comparable to a Lofitech (Lofitech
AS, Leknes, Norway) seal scarer as fatiguing stimuli. This artificial seal scarer signal was built
following Lofitech seal scarer signals, recorded at a distance of 130 m (Brandt et al., 2013a).
The main frequency component of the 0.5 s signal was set to 14 kHz and four harmonics (28,
42, 56 and 70 kHz) with gradually decreasing sound levels were added. The artificial signal
was generated in R (R Core Team, 2019), using the package ‘seewave’ (Sueur et al., 2008). The
stimuli level of each frequency component was calibrated before the first exposure of this study,
meeting the gradually decreasing components in Brandt et al. (2013). The same signal was used
for all exposures within this study.
Initially the fatiguing sound were presented at very low exposure levels and then subsequently
increased by a maximum of 3 dB between days. The signals were transmitted by an ITC-1001
transducer (International Transducer Corporation, USA). A power amplifier (PA1001, ETEC,
Dk) was used to increase sound energy emission in steps of 3 dB. Signals were amplified
(PA1001, ETEC, DK) and recorded at the bite plate with a TC4013 hydrophone (Reson
Teledyne, DK), pre-amplified by 40 dB (ETEC-B, 1501 amplifier, DK) and bandpass filtered
with a passband from 1 to 180 kHz. Fatiguing stimuli were played back by a custom written
software (LabVIEW, USA), which was also used for background recordings. The sound
exposure level (SEL) was determined using this software. The frequency spectrum of the
artificial seal scarer signal, used as a fatiguing stimulus on October 18th, is presented in FIG. 7.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
54
FIG. 7: Recorded frequency spectrum of one generated artificial seal scarer signal (top), which
was used for animal exposure. The STFT of the recorded signal (bottom) shows the peak
frequency of the seal scarer signal at 14 kHz with 4 octave overtones. Porpoise clicks
around 130 kHz were recorded prior and after exposure. Overlap=87.5%, window
length= 1048 samples.
In further analysis, recordings of seal scarer exposures were additionally frequency weighted,
adjusting the signal to perceived loudness, based on the harbour porpoise hearing spectra. We
used the proposed NOAAHF (National Oceanic and Atmospheric Administration) frequency
weighting, which has been recommended to evaluate effects of underwater noise on the hearing
of marine mammals (National Marine Fisheries Service, 2018; Southall et al., 2019). Frequency
weighting of raw wav recordings has been conducted by the recently published MATLAB
(MathWorks, USA) function ‘NOAAweighted’ (Tougaard and Beedholm, 2018). Within the
recommended Marine Mammal Noise Exposure Criteria Southall et al. (2019) refer to this same
weighting function as ‘very high frequency cetaceans’. The SEL was determined afterwards by
the same software used for the playback of fatiguing sounds.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
55
E. TTS definition
All hearing thresholds with no prior exposure were used to determine the baseline. Due to the
small sample size of baseline measurements at 28 kHz, we included hearing thresholds, which
have been measured 2 h post-exposure, in recovery trials. These two events have been tested
by a Welch two sample t-test for unpaired samples for differences compared to the 8 baseline
measurements. The thresholds measured in recovery trials did not show significant differences
from the baseline trials. Therefore hearing thresholds at 28 kHz measured in the recovery trials
were pooled with the baseline measurements. This pooling is assumed to be conservative in the
detection of a TTS, since only higher and no lower hearing thresholds are expected in the post-
exposure trials. The baseline-hearing threshold was defined as the mean of all these trials. Post
exposure threshold measurement procedure was the same as for baseline hearing trials. Hearing
thresholds were measured immediately after exposure and were gathered in an interval of 1 to
8 minutes after exposure. Using the mean hearing threshold and its standard deviation, we
obtained the threshold for a significant hearing shift (p=0.05). A temporary threshold shift was
defined as a hearing threshold exceeding the mean threshold by 1.65 × the standard deviation
(p=0.05, one-sided test). A typical TTS definition is a hearing thresholds 6 dB above the
baseline (Southall et al., 2019). However, this threshold is based on the smallest shift, which
was clearly distinguishable due to the variability of 3-4 dB in baseline measurements in the
study of Schlundt et al.( 2000). Although we follow the TTS definition, as a significant shift
from baseline hearing (see e.g. Finneran et al. (2005); Kastelein et al. (2014b, 2016)), the 6 dB
criterion onset can be obtained from the provided TTS regression.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
56
F. Data analysis
Data recorded by the electrodes was first checked for quality, in order to exclude obvious
disturbances, which can be produced by environmental effects (waves, rain) or technical
problems (crosstalk between cables). Recorded signals with peak voltages above 20 µV were
rejected, in order to exclude myogenic artefacts with large amplitudes, originating from
movements or respiration of the animal. Data were post-processed with a digital Butterworth
bandpass filter of 8th order, to get a clean signal between 0.3 and 3 kHz. To objectively
determine, if stimuli at a certain SPL could be heard by the animal, we tested if the recorded
brainstem signal differed from background noise within the recordings by a single point F-test
(Don et al., 1984; Elberling and Don, 1984). This method uses a variance analysis in
determining the ratio of the magnitude of the ABR to the estimated averaged background noise.
Afterwards, all responses were verified by two trained assessors, searching for stereotypic
patterns manually. The stereotypic wave V in an auditory evoked potential has been proven to
have a significant relationship to loudness (Serpanos et al., 1997) and therefore served as an
indicator for sound perception. A signal was determined as perceived (“hit”), if wave V was
determined in a time window of 3.8 to 5.2 ms after exposure. If no wave V was found the signal
was determined as “miss”. Hearing thresholds were defined as the mean SPL of the lowest “hit”
and the highest “miss” (FIG. 8). All analyses were conducted and figures created with R
Studio(R Core Team, 2019).

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
57
FIG. 8: Example of measured AEP for 8 sound pressure levels (colour coded) between 87 and
122 dB re 1 µPa. Data was recorded after an exposure to a seal scarer signal with an SEL
of 147.3 dB re 1 µPa²s. The dashed black vertical lines indicate the time window, where
the wave V was searched. The post-analysis with the single point F-test and visual
screening of two experienced assessors determined the lowest “hit” at a SPL of
97 dB re 1 µPa and the highest “miss” at 92 dB re 1 µPa. The hearing threshold was
determined in the middle of these values and corrected for the results of the SPL
calibration (-5.3 dB), resulting in a threshold of 89.2 dB re 1µPa. The vertical black line
in the top-left corner represents the voltage scale.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
58
III. Results
A. Baseline hearing thresholds
Baseline hearing at 20 kHz was measured on 25 days (see TABLE V in APPENDIX), resulting
in a mean baseline hearing threshold of 90.0 dB re 1 µPa ± 3.7 dB. An inspection of quantile-
quantile plots verified, that baseline hearing thresholds were normally distributed and a critical
value (p<0.05) of 96.1 dB re 1 µPa was obtained. Therefore, a hearing shift higher than 6.1 dB
was regarded as a significant hearing elevation.
Baseline measurements for stimulation at 28 kHz were conducted on 6 days, resulting in 10
measurements (appendix A). Since the measurements of baseline and recovery sessions were
not significantly different (p=0.9713), pre- and post-exposure hearing thresholds were pooled
to determine a baseline hearing threshold for 28 kHz. The mean baseline hearing threshold was
85.4 dB re 1 µPa ± 2.9 dB. An inspection of quantile-quantile plots verified, that baseline
hearing thresholds were normally distributed and a critical value (p<0.05) of 90.4 dB re 1 µPa
was obtained. Therefore, a hearing shift higher than 4.8 dB was regarded as a significant hearing
elevation.
B. Post-exposure thresholds
The harbour porpoise showed an aversive reaction to the presentation of the fatiguing stimulus
in almost all cases with an exposure exceeding 143 dB re 1 µPa²s. It consisted of a short
backward movement without leaving the bite plate. This reaction could represent the acoustic
startle reflex, which mirrors the audiogram 80-90 dB above the hearing threshold (Pilz et al.,
1987). Acoustic signals with a short rise time can elicit an oligo-synaptic reflex arc in the
brainstem, provoking the contraction of refractor muscles (Koch and Schnitzler, 1997). This
reaction did not occur during all trials. The harbour porpoise stayed at its position until it was
called back by the trainer during all exposure sessions.
For hearing tests at 20 kHz, exposure trials were conducted on 9 days with sound exposure
levels between 137.6 and 157.5 dB re 1 µPa²s. In 5 trials a significant temporary hearing shift
was measured at 20 kHz, as the post-exposure threshold was above 96.1 dB re 1 µPa, which
was determined as the critical value for a TTS at 20 kHz. Calculating linear regression for these
determined hearing shifts, we obtained a TTSonset at a sound exposure level of
141.8 dB re 1 µPa²s (FIG. 9).

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
59
Post-exposure hearing at 28 kHz was tested in 5 trials with sound exposure levels from 146.9
to 155.6 dB re 1 µPa²s. In 4 trials a significant TTS, defined as a hearing threshold above
90.2 dB re 1 µPa, could be measured at 28 kHz. We inferred a TTSonset at a SEL of
146.9 dB re 1 µPa²s , using linear regression (FIG. 9).
We obtained a SPL-TTSonset for each SEL-TTSonset by a linear regression of measured SPL and
SEL (see APPENDIXFIG. 11Appendix C). NOAAHF weighted TTSonsets were derived from the
calculated regression of frequency weighted exposure recordings and the determined TTS (see
FIG. 12 in Appendix D). All results are summarized in TABLE IV.
FIG. 9: Effects of seal scarer exposure (at 14 kHz, with four harmonics) on hearing thresholds
at 20 (black circles) and 28 kHz (grey triangles). Filled symbols indicate a significant shift
from baseline hearing, which was determined at >6.1 dB for 20 and >4.8 dB for 28 kHz.
Unfilled symbols indicate hearing thresholds, which were not significantly shifted after
exposure. A linear regression for each hearing frequency, using only exposures which led
to a TTS, was calculated in order to estimate the TTSonset. The 95 % confidence intervals
are shown by dashed lines. The TTSonset at 20 kHz was determined at 141.8 and at 28 kHz
at 146.9 dB re 1 µPa²s.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
60
TABLE IV. Summary of the determined TTSonset after exposure to artificial seal scarer signals.
The signal consisted of a 14 kHz tone, with four harmonics. Hearing thresholds were
measured at 20 and 28 kHz. TTSonsets were determined for unweighted and auditory based
NOAAHF (National Oceanic and Atmospheric Administration; National Marine Fisheries
Service, 2016) weighted exposures.
Frequency
kHz
Weighting Intercept slope SEL-TTSonset
dB re 1 µPa²s
SPL-TTSonset
dB re 1 µPa
20 - -81.7 0.6 141.8 155.2
20 NOAAHF -78.9 0.6 138.4 152.9
28 - -103.9 0.8 146.9 160.3
28 NOAAHF -136,6 1.0 143.6 157.4
IV. Discussion
A. Application of Seal Scarers
Acoustic deterrent devices are widely used, for instance to counteract the economic loss by seal
depredation in aquacultures. Within the past 30 years the worldwide farming of finfish species
has substantially increased (FAO, 2018; Findlay et al., 2018) and so has the economic loss by
seal depredation. These sites represent an easily accessible food source with a high profitability,
if no anti-predator control methods are applied. Although ADDs are extensively applied where
fish farms exist in Europe, e.g. along the west coast of Scotland, no official statistics exist on
the number, types and duration of ADD usage (Findlay et al., 2018). Furthermore, its deterrence
efficiency is highly variable and may also decrease over time, due to habituation (reviewed in
Götz and Janik, 2013).
In contrast to seals, harbour porpoises show strong avoidance behaviour to ADDs (Brandt et
al., 2013a; Mikkelsen et al., 2017; Olesiuk et al., 2002). Seal scarer operators should be aware,
that the behavioural manipulation of seals as a target species, might also affect non-target
species, which is widely overlooked, when ADDs are applied around aquacultures. The rather
unspecific signals of seal scarers are not specifically tuned to the auditory abilities of seals.
Areas where aquacultures are located, regularly overlap with harbour porpoise occurrence
(Findlay et al., 2018). Harbour porpoises are especially sensitive for underwater noise (Southall
et al., 2019). For the case of fish farms, harbour porpoises (Phocoena phocoena) are regarded
as a non-target species because there is no evidence that these animals feed on farmed fish or
damage fishing gear (Götz and Janik, 2013). The strong reactions of harbour porpoises to ADDs

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
61
have been exploited in other applications, such as a deterrent device prior to pile driving
activities with the aim to prevent hearing impairment from the high noise levels of pile driving
strikes. Indeed, a far reaching avoidance behaviour occurred up to distances of 7.5 (Brandt et
al., 2013b) or even 12 km (Dähne et al., 2017). Although the exploitation of renewable energy
represents an important component towards a more environmentally friendly power production,
impacts on marine fauna, caused by anthropogenic noise during and prior to the construction
have to be considered.
B. Effect on harbour porpoise hearing
Caution is required when using deterrent devices with high source levels for both, the
application around fish farms and as deterrent devices to prohibit TTS from pile driving strikes.
We could show that these signals itself have the potential to induce a significant temporary
hearing shift in a harbour porpoise both at 20 and 28 kHz hearing frequency. Harbour porpoises
critically rely on hearing to navigate (Villadsgaard et al., 2007), find and catch prey items
(DeRuiter et al., 2009; Wisniewska et al., 2016) and communicate (Clausen et al., 2010;
Sørensen et al., 2018). Disturbance effects arise from numerous sources in addition to noise
pollution (Andreasen et al., 2017; ASCOBANS, 2002, 2012; Beineke et al., 2005; Jepson et al.,
2016; Knoop and Müller, 2009; Mahfouz et al., 2014; Reeves et al., 2013). Affected hearing
can influence the survival rate of single individuals (Mann et al., 2010; Morell et al., 2017a).
Disturbance effects may even result in decreased individual fitness and could lead to long-term
population consequences (King et al., 2015).
Although the experiments were conducted in a semi-natural enclosure with a sound field that
we could not control, the measured TTS was most likely induced by the exposure of the seal
scarer signal only. While passing vessels can lead to behavioural reactions of harbour porpoises
(Dyndo et al., 2015; Wisniewska et al., 2018b), there is no evidence for the potential to induce
a TTS by this continuous noise source. Experiments were only conducted in the absence of
anthropogenic noise. Therefore, we can exclude the possibility that the induced TTS derived
from another noise source than the seal scarer signal.
The pre-exposure baseline hearing threshold at 20 kHz is about 40 dB higher than measured by
Kastelein et al. (2010). This difference is substantially caused by differences in the applied
methods to measure hearing thresholds. Using psychophysical techniques instead of AEP
measurements can lead to thresholds, which are 1 – 31 dB lower (Mulsow and Reichmuth,

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
62
2010). Hearing thresholds of wild harbour porpoises from the Inner Danish Waters, which have
been determined by AEP measurements are about 10 dB lower (Ruser et al., 2016a) than the
baseline hearing of this study at 20 kHz. Our data derives from one animal, at a senior age of
23 years, in a semi-natural enclosure restricted by nets from the harbour solely, but is still within
the 1.5 interquartile range of wild harbour porpoise hearing thresholds (Ruser et al., 2016a).
We tested the effect of the artificial sear scarer signal on harbour porpoise hearing at 20 kHz,
although the main frequency component of the fatiguing stimulus was at 14 kHz, which is
~0.5 octaves below. This was due to technical difficulties in measuring hearing thresholds
below 20 kHz in the semi-natural environment and with this certain animal. The hearing
threshold at 28 kHz is at the frequency of the first harmonic of the artificial seal scarer signal.
Conclusively, this limits the comparability between the estimated onsets for 20 and 28 kHz
because a TTS is expected to be greatest 0.5 octaves above the fatiguing stimulus (Kastelein et
al., 2014c; McFadden and Plattsmier, 1983; Popov et al., 2011).
In one case for each 20 and 28 kHz trials, we did not measure a TTS after the exposure, even if
the SEL exceeded the TTSonset (FIG. 9, unfilled circle, unfilled triangle). Although we took care
in counteracting any potential conditioning behaviour, the unaffected hearing threshold after
exposure could be a result of a self-protective mechanism, which actively dampens hearing
sensitivity, in expectation of an impending loud noise event. This self-mitigation has previously
been shown for four Odontocete species (Nachtigall et al., 2017). The harbour porpoise in this
study could have reduced its hearing abilities in order to prevent a TTS during exposure. Since
this effect seems to be of short duration (Finneran, 2018; Nachtigall et al., 2016), we could not
find a TTS when we measured the hearing in the period after the exposure, although expected
for these exposure levels, because either a TTS was not induced, or the self-mitigation was not
active anymore.
The study design was adjusted to counteract any conditioning behaviour by the porpoise, to
reduce hearing sensitivity in expectation of sound exposure. This was done by keeping all
experimental processes stable in exposure and non-exposure trials, besides sound exposure.
The measured hearing shifts are rather a result of a fatigued hearing after exposure, than a self-
mitigation. This phenomenon only occurred after conditioning the animals to expect an

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
63
unpleasant signal after a preceding warning stimulus (Nachtigall et al., 2017). In this study, we
tested hearing after exposure, so there was no expectation of an upcoming unpleasant event.
C. Estimated hazard zones
Based on the determined SPL-TTSonset of 155.2 dB re 1 µPa (p-p, TABLE IV), we estimated a
hazard zone, where a TTS could be induced in harbour porpoise hearing after the reception of
a single seal scarer signal. To estimate hazard zones, a theoretical sound propagation of seal
scarer signals was modelled. Specified by the manufacturer, the source level of the Lofitech
seal scarer is between 189 and 193 dB re 1 µPa (http://www.lofitech.co.uk/). To consider the
most precautionary approach, we assumed the highest reported source level of 193 dB re 1 µPa.
Based on this source level, a theoretical sound propagation was modelled as a simple
logarithmic regression. A similar approach with a simple logarithmic regression close to
spherical spreading proved to be valid for seal scarer sound propagation in the German North
Sea (Brandt et al., 2013b). The theoretical propagation was modelled for deep (spherical
spreading with a transmission loss of 20× log10 (r)) and shallow water (cylindrical spreading
with a transmission loss of 10× log10 (r)). A practical spreading with 15× log10 (r) in between
deep and shallow spreading was additionally modelled, resulting in three tested factors for the
slope of the transmission loss. The absorption coefficient α was estimated for typical North Sea
parameters (assuming 15°C water temperature, salinity of 35 ppt, a depth of 20 m, acidity of
pH=8) and a peak frequency of 14 kHz, at 1.5 dB km-1 (Ainslie and McColm, 1998).
Accordingly, for the received level we obtained 𝑅𝐿 = 𝑆𝐿 − 𝑠𝑙𝑜𝑝𝑒 × 𝑙𝑜𝑔10(r) − α × r.
Since the source level of 193 dB re 1 µPa corresponds to a rms value, the SPL-TTSonset (p-p)
was corrected. For the SPL-TTSonset of 155.2 dB re 1 µPa (p-p,) we obtain a rms SPL-TTSonset
of 146.2 dB re 1µPa (FIG. 10, horizontal dashed line), by subtracting 9 dB (Madsen, 2005).
Single seal scarer signals with a source level of 193 dB re 1 µPa are assumed to induce a TTS
in harbour porpoises up to distances between 211 m (spherical spreading in deep water) and
5.9 km (cylindrical spreading in shallow water), depending on theoretical sound propagation
(FIG. 10).
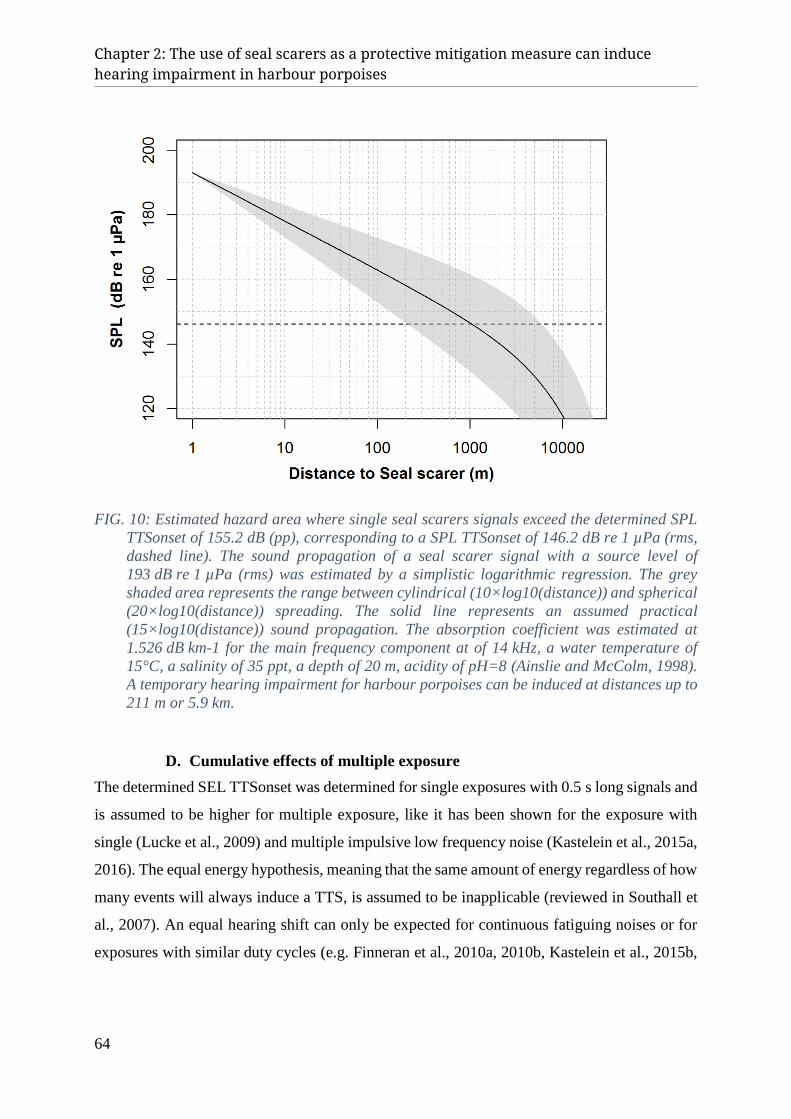
Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
64
FIG. 10: Estimated hazard area where single seal scarers signals exceed the determined SPL
TTSonset of 155.2 dB (pp), corresponding to a SPL TTSonset of 146.2 dB re 1 µPa (rms,
dashed line). The sound propagation of a seal scarer signal with a source level of
193 dB re 1 µPa (rms) was estimated by a simplistic logarithmic regression. The grey
shaded area represents the range between cylindrical (10×log10(distance)) and spherical
(20×log10(distance)) spreading. The solid line represents an assumed practical
(15×log10(distance)) sound propagation. The absorption coefficient was estimated at
1.526 dB km-1 for the main frequency component at of 14 kHz, a water temperature of
15°C, a salinity of 35 ppt, a depth of 20 m, acidity of pH=8 (Ainslie and McColm, 1998).
A temporary hearing impairment for harbour porpoises can be induced at distances up to
211 m or 5.9 km.
D. Cumulative effects of multiple exposure
The determined SEL TTSonset was determined for single exposures with 0.5 s long signals and
is assumed to be higher for multiple exposure, like it has been shown for the exposure with
single (Lucke et al., 2009) and multiple impulsive low frequency noise (Kastelein et al., 2015a,
2016). The equal energy hypothesis, meaning that the same amount of energy regardless of how
many events will always induce a TTS, is assumed to be inapplicable (reviewed in Southall et
al., 2007). An equal hearing shift can only be expected for continuous fatiguing noises or for
exposures with similar duty cycles (e.g. Finneran et al., 2010a, 2010b, Kastelein et al., 2015b,

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
65
2014; Mooney et al., 2009; Popov et al., 2014). In fact, the TTSonset decreases with increasing
SPL and duty cycle (Kastelein et al., 2015b, 2014b).
The inter pulse intervals of seal scarers are usually randomized at intervals between 0.6 and
90 s to counteract potential habituation effects (e.g. Lofitech used by Brandt et al. (2013)).
Within these varying inter pulse intervals the hearing can recover. Since higher duty cycles
induce a TTS at a lower SELcum, a TTS is more likely for multiple pulses with short inter pulse
intervals. However, further experiments are needed, to determine the TTSonset for varying sound
exposure levels and inter pulse intervals. This is critically needed to reliably predict a TTS
potential for fleeing harbour porpoises from multiple exposures.
E. Management approach
We could demonstrate that a single exposure to seal scarer signals can lead to hearing
impairment of non-target species at hundreds of meters. Negative effects could be even more
dramatic, if multiple ADDs are applied simultaneously on multiple cages within a single
aquaculture or at further adjacent sites. We obtained a frequency weighted TTSonset of
138 dB re 1 µPa²s, which is surprisingly 15 dB lower than the updated recommended marine
mammal noise exposure criteria for continuous noise (Southall et al., 2019). Following these
recommendations, a PTSonset is estimated 20 dB above the TTSonset level, which corresponds to
158 dB re 1 µPa²s, according to our results. Regarding the high source level of commercially
available seal scarers, even a PTS can be induced at very close distances. Given the evidence
that seal scarers can induce an injury in harbour porpoises, no matter if defined as TTS (e.g.
German regulation, BMU, 2014) or PTS (most other states in Europe and in the United States,
e.g. (Southall et al., 2019)), there is a clear requirement to manage the application of seal
scarers.
For the application of seal scarers to deter seals around aquacultures, a shift to lower frequencies
could decrease the risk for hearing impairment for the harbour porpoise as a non-target species.
A shift to lower frequencies, as also proposed by Götz and Janik (2013), would be beneficial,
since the hearing abilities for lower frequencies are better for seals than for porpoises (Kastelein
et al., 2002; Ruser et al., 2016a). Deterring signals for seals, should not contain much energy
above 5 kHz, if Odontocetes use habitats around the fish farm (Götz and Janik, 2013). On the
contrary, using lower frequencies could be worse for baleen whales. Therefore it has to be
considered, if baleen whales inhabit areas around the site, where ADDs are applied. In case of

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
66
further developments of acoustic deterrent devices, the target specificity must be validated by
independent studies prior deployment around aquacultures.
Although ADDs are a significant and chronic source of underwater noise pollution along the
Scottish west coast around aquacultures with a steady increase in ADD usage and substantial
geographic expansion (Findlay et al., 2018), neither licence is required to deploy these devices,
nor any statistics on usage exist (Coram et al., 2014). An unwanted deterrence of harbour
porpoises has been regarded as a comparatively benign side effect, while ADDs were deployed
to counteract economic loss by seals. While behavioural responses and exclusion from key
habitats of harbour porpoises were evaluated as insufficient arguments to regulate the use of
ADDs, the evidence that a TTS or even PTS can be induced, must lead to a regulation system.
Additionally, an adjustment of the source level of ADDs has to be considered, to reduce the
potential impact on harbour porpoise hearing. This adjustment should be taken into account for
the use around aquacultures, but also as a deterrent device for harbour porpoises, prior to pile
driving activities. From a conservation point of view, this deterrence should be adjusted to the
expected TTS hazard zone from pile driving strikes. While received levels above the presented
TTSonset are assumed to induce increased hearing thresholds, the deterrence efficiency would
be decreased accordingly, which is a problem from a commercial perspective (Götz and Janik,
2013). We therefore advice to down regulate source levels as a protective measure and use an
amplitude ramp up, giving harbour porpoises sufficient time to leave hazardous areas.
F. Conclusion
Seal scarer signals have the potential to impair harbour porpoise hearing. The TTSonset was
determined at an SEL of 141.8 dB re 1 μPa²s at the hearing threshold of 20 kHz and at a 146.9
dB re 1 μPa²s for 28 kHz hearing frequency. The frequency weighted TTSonset was 15 dB below
the recommended Marine Mammal Noise Exposure Criteria (Southall et al., 2019). Hazard
zones, where a TTS can be induced by a single exposure are dependent on sound propagation,
but are expected to be between 211 m and 5.9 km for reported source levels of up to 193 dB re
1 µPa. Based on our findings, effects of multiple exposure cannot be predicted due to the
random inter pulse intervals. In order to use seal scarers to deter harbour porpoises instead of
impairing its hearing, we suggest to down regulate source levels to the desired deterrence range
and to slowly increase the source level, giving harbour porpoises time to flee.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
67
V. Acknowledgement
Experiments within this study were conducted within the project “Impact of underwater noise
on marine mammals (UWE)” FKZ 3515 82 2000), funded by the Federal Agency for Nature
Conservation (BfN). We would like to express our gratitude to the team of animal trainers of
the Fjord & Bælt Centre, especially to Fredrik Johansson, for the huge effort in training and
handling the harbour porpoises prior to and during experiments.

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
68
VI. Appendix
A: Measured hearing thresholds for baseline hearing measurements
TABLE V shows the results for the hearing tests which were conducted without any prior
exposure. The measured hearing thresholds were used to determine the baseline hearing
threshold.
TABLE V: Measured hearing thresholds, which have been used to calculate mean hearing
thresholds
Date Frequency
in kHz
Hearing threshold
in dB re 1 µPa
13.07.2017 20 87.6
13.07.2017 20 93.0
04.09.2017 20 85.8
04.09.2017 20 86.2
22.09.2017 20 86.5
27.06.2018 20 91.8
28.06.2018 20 82.1
28.06.2018 20 86.5
06.07.2018 20 95.7
06.07.2018 20 88.2
06.07.2018 20 92.0
17.07.2018 20 87.2
19.07.2018 20 89.6
20.07.2018 20 96.4
22.08.2018 20 93.6
23.08.2018 20 92.6
04.09.2018 20 92.6
13.09.2018 20 92.2
18.09.2018 20 87.9
19.09.2018 20 93.0
29.06.2017 28 84.0
10.10.2018 28 87.2
10.10.2018 28 84.7
10.10.2018 28 89.9
11.10.2018 28 89.0
17.10.2018 28 82.4
17.10.2018 28 83.7
18.10.2018 28 86.1
18.10.2018 28 87.3
25.10.2018 28 80.0

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
69
Appendix B: Measured hearing thresholds after exposure
TABLE VI shows the measured hearing thresholds after the exposure to the artificial seal scarer
signal
TABLE VI. Measured hearing thresholds after exposure, which resulted in a significant hearing
shift, relative to the baseline. Asterisks indicate levels of significance (*: p<0.05; **:
p<0.01; ***: p<0.001).
Date Frequency
kHz
Threshold
dB re 1 µPa
Exposure
dB re 1 µPa²s
z-value p-value
17.07.2018 20 93.73 138.4 1.01 0.1570
19.07.2018 20 89.75 137.6 -0.08 0.5300
20.07.2018 20 100.7 148.4 2.90 0.0019**
22.08.2018 20 89.2 147.2 -0.22 0.5889
31.08.2018 20 108 157.5 4.89 0.0000***
04.09.2018 20 103.7 154.2 3.72 0.0001***
13.09.2018 20 102.2 155.7 3.31 0.0005***
18.09.2018 20 93.3 143.1 0.89 0.1867
19.09.2018 20 98.3 145.3 2.25 0.0122*
11.10.2018 28 91 146.9 1.92 0.0273*
17.10.2018 28 92.6 153.1 2.47 0.0067**
18.10.2018 28 101 155.6 5.37 0.0000***
24.10.2018 28 96.7 152.5 3.89 0.0001***
25.10.2018 28 79.7 148.4 -1.98 0.9760

Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
70
Appendix C: Relationship between SEL and SPL of exposures
FIG. 11 shows the modeled correlation between sound exposure and sound pressure levels of
the recorded artificial seal scarer sounds. This linear regression allowed for the transfer of the
determined SEL-TTSonset to the SPL-TTSonset.
FIG. 11. Linear regression of SEL–SPL of all played back seal scarer signals during hearing
tests at 20 and 28 kHz. Black points represent the unweighted SEL of all exposures. The
circles represent the NOAAHF frequency weighted SEL of all exposures. The regression
was used to estimate the corresponding SPL-TTSonset from the SEL-TTSonset.
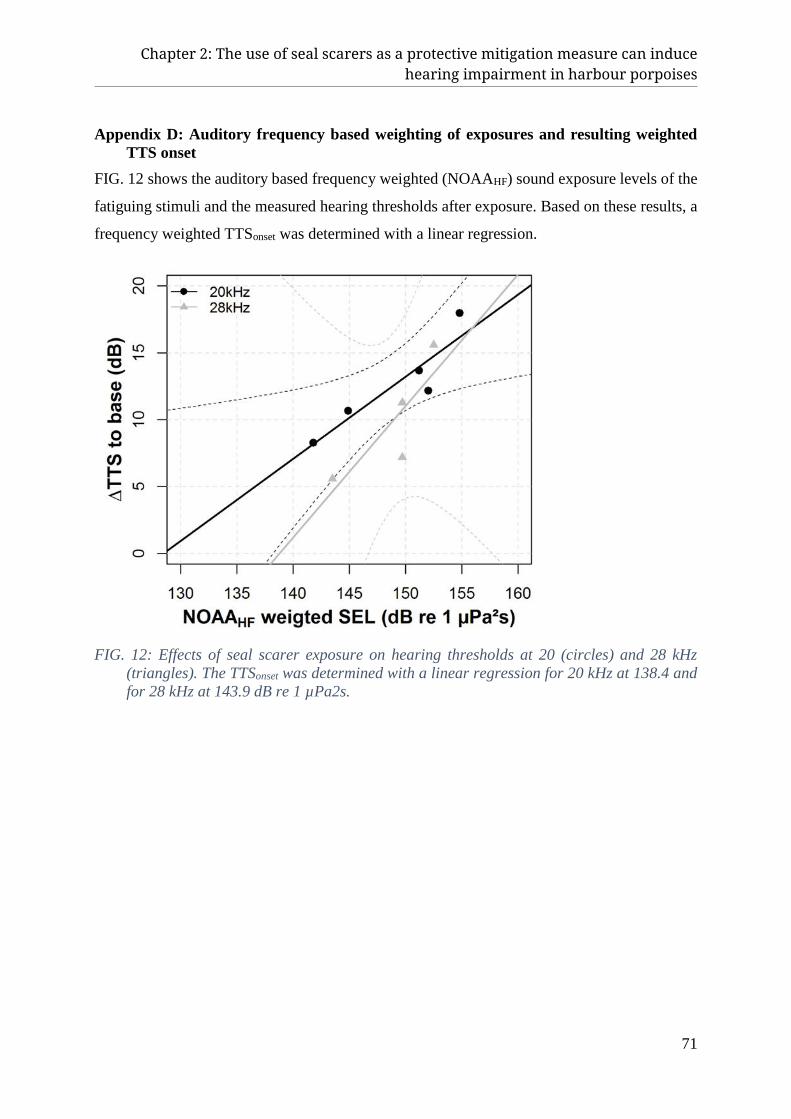
Chapter 2: The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises
71
Appendix D: Auditory frequency based weighting of exposures and resulting weighted
TTS onset
FIG. 12 shows the auditory based frequency weighted (NOAAHF) sound exposure levels of the
fatiguing stimuli and the measured hearing thresholds after exposure. Based on these results, a
frequency weighted TTSonset was determined with a linear regression.
FIG. 12: Effects of seal scarer exposure on hearing thresholds at 20 (circles) and 28 kHz
(triangles). The TTSonset was determined with a linear regression for 20 kHz at 138.4 and
for 28 kHz at 143.9 dB re 1 µPa2s.

Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
72
Manuscript:
T. Schaffeld, J.G. Schnitzler, B.J. Baltzer, M. Fischer, M. Schuster, A. Ruser, U. Siebert (2020).
Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
73
Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise
hearing
T. Schaffelda, J.G. Schnitzlera, J. Baltzera, M. Fischerb, M. Schusterb, A. Rusera, U. Sieberta
a Institute for Terrestrial and Aquatic Wildlife Research, University of Veterinary Medicine
Hannover, Foundation, Werftstraße 6, 25761 Büsum, Germany
b DW-ShipConsult GmbH, Lise-Meitner-Straße 9, 24223 Schwentinental, Germany
Abstract
After decades of absence, reports of harbour porpoise sightings in the Elbe or Weser River
increased again over the last years. Harbour porpoises annually occur, with a peak in spring in
the port of Hamburg, despite the high number of present vessels. This seasonal occurrence
might be related to anadromous prey species, which could represent an important food source
when travelling upstream for spawning. After numerous strandings of dead animals in 2016
close to the port of Hamburg, an acoustic flowmeter in the port was suspected as a cause. By
an initiative of the Hamburg Port Authority, which operates the flowmeter, underwater noise
was recorded in this area. The flowmeter emits short pulses with a length of 0.2 s and a
frequency of 28 kHz, which can be heard by harbour porpoises. These signals are transmitted
from the northern and southern banks of the Elbe River, which are about 400 m away from each
other. The transmission alternates every 2 s between the two sides , with a pulse rate of 4.1 s
for each of the flowmeters. Within this chapter, we evaluated the potential of these signals to
induce a temporary threshold shift (TTS) in harbour porpoises by a single or multiple exposure.
The modelled sound field demonstrates, that the acoustic flowmeters emit pulses in a highly
directional beam. The pulses contain sufficient energy to induce a TTS in harbour porpoises by
the exposure of single signals up to a distance of approximately 72 m from the sound source.
Although the area with pulses of highest amplitude are comparably small, total received sound
exposure levels of up to 165 dB re 1 µPa²s were estimated for travelling harbour porpoises The
total received cumulative energy for harbour porpoises travelling along the flowmeters are
mainly dependent on the timing and position of the animals. Accordingly, a close approach to
the flowmeter at the time of transmission should be prevented. This could be the case, if vessels
force harbour porpoises to elude closer to the flowmeters. Following the recommendations of
this chapter, the HPA adjusted the source level of the flowmeter.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
74
Author contributions
A.R., M.S. and U.S. were involved in designing the project and applying for the project grant.
A.R. and T.S. prepared the underwater noise measurements, which were conducted by A.R.,
T.S. and M.S. The underwater noise recordings were analysed by M.F. and M.S. who provided
the results as a modelled sound field to T.S.. Hearing tests with a harbour porpoise in human
care were performed by A.R.. All analyses to evaluate the TTS potential for a harbour porpoise
in the sound field of the acoustic flowmeters were performed by T.S. under supervision of A.R.,
J.S. and U.S. All authors provided critical feedback and helped to shape the research and
analysis. The manuscript was written by T.S. with supportive contributions to the final version
of A.R., J.S., J.B. and U.S..

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
75
I. Introduction
Harbour porpoises (Phocoena phocoena) inhabit coastal waters of the northern hemisphere
(Gilles et al., 2016; Hammond et al., 2017). It is the only cetacean representative, with a regular
reproduction in German waters (Gilles et al., 2016). Historic reports prove that harbour
porpoises commonly inhabited lower stretches of large rivers in the 19th century as shown by
sightings or strandings in the Weser River south of Bremen (Häpke, 1880) or even up to
Magdeburg, which is upstream the Elbe River at a distance of 400 km from the coast (Kölmel,
1998). Following decades of absence, harbour porpoise sightings increased again in rivers
entering the German North Sea (Wenger et al., 2016). In 2013, a reporting system was
established, allowing collecting harbour porpoise sightings (Wenger et al., 2016). This resulted
in 233 reports of harbour porpoises in the lower Elbe River between 27th of February and 9th of
July in 2013. A large extent of the sightings were reported close to the port of Hamburg. In
addition to the reporting system, a static acoustic monitoring with porpoise click detectors (C-
POD, Chelonia Limited, UK) was conducted, which could confirm a clear seasonality in
detections with a peak in spring (Wenger et al., 2016). The acoustic monitoring revealed the
continuous presence of harbour porpoises in the port of Hamburg over the entire monitoring
period from February to June, reaching a maximum of 90 % detections per daytime in the
middle of April. According to newspaper reports, 80-100 harbour porpoises were
simultaneously present in this area, occurring in groups of up to 10 animals, which were seen
foraging together (https://www.abendblatt.de/hamburg/article207486167/Schon-100-Wale-
im-Hafen-Gefahr-durch-Hafengeburtstag.html, 26.04.2016 and pers. comm. Denise Wenger).
In the lack of an established systematic monitoring of harbour porpoises, these counts have to
be considered with great caution, since these are not corrected for repeated sightings. Due to
the determined strong seasonality in harbour porpoise presence and the determined foraging
activity by sightings and acoustic monitoring, it is assumed that harbour porpoises enter large
rivers because they follow migrating anadromous fish species (Wenger et al., 2016). Spatial
and temporal occurrence of harbour porpoises in this area correlates with the annual migration
of anadromous fish species like smelt (Osmerus eperlanus) and twaite shad (Allosa fallax),
which are known to spawn in this area (Thiel, 2015).
In 2013, 26 dead harbour porpoises were found around the area of the harbour in Hamburg
(Wenger et al., 2016). Since no necropsies were conducted, no cause of death could be

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
76
determined. Harbour porpoises critically rely on a well-functioning hearing system. Harbour
porpoises produce short high frequency clicks to acoustically sense their environment by the
reception of returning echoes (e.g. Verfuß et al., 2005). This ability to echolocate is shared with
bats (reviewed in Fenton et al., 2014) and enables navigation (Verfuß et al., 2005) as same as
finding and catching prey (DeRuiter et al., 2009; Verfuß et al., 2009) in complete darkness.
Among the toothed whales, which are generally vulnerable to noise disturbance, harbour
porpoises are considered to be very sensitive to effects of noise and are therefore regarded as
an indicator species in noise impact evaluations (Southall et al., 2019; Tougaard et al., 2015).
In European waters, harbour porpoises are protected amongst others within the framework of
the Habitats Directive (listed in annexes II and IV European Union, 1992) and Council
Regulation 812/2004 (European Union, 2004). This protective regulation implies that special
areas should be established for their conservation with deliberate actions of killing, disturbing,
injuring and habitat deterioration being prohibited throughout its area of distribution (Council
Directive 92 / 43 / EEC, Article 12.1). Sources of anthropogenic disturbance are numerous for
harbour porpoises in German waters. The largest impact is caused by unintended bycatch in
set-net fisheries (Korpinen and Braeger, 2013; Wehrmeister et al., 2013), chemical pollution
(Huber et al., 2012; Mahfouz et al., 2014) and the introduction of underwater noise (Akkaya
Bas et al., 2017; Brandt et al., 2018; Dyndo et al., 2015; Sarnocińska et al., 2020; Wisniewska
et al., 2018b). The exposure to underwater noise can induce a temporary threshold shift (TTS)
in harbour porpoise hearing (Kastelein et al., 2016; Lucke et al., 2009). Since a hearing shift is
regarded as an injury in Germany, noise regulation aims at preventing a TTS. Accordingly, the
evidence of a TTS potential from impulsive noise (Lucke et al., 2009) was adopted very quickly
in noise regulation measures during the construction of Offshore Wind farms in Germany
(BMU, 2014).
Since strandings of cetaceans have been associated with anthropogenic sonar signals in the past
(reviewed in Simmonds et al., 2014) and the high sensitivity of harbour porpoises towards noise
are commonly known (Southall et al., 2019), an acoustic device in the port of Hamburg was
suspected as the cause for the reported strandings. This device is applied by the Hamburg Port
Authority (HPA) in the Elbe River in the area of Finkenwerder in order to measure the water
flow velocity. The flow speed is obtained from the measured travel time of ultrasonic acoustic
signals, which are transmitted from both sides of the river. To evaluate the potential of this

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
77
flowmeter, the HPA funded a project themselves, including underwater noise measurements
and a biological evaluation, because they wanted to exclude, that the flowmeter has negative
impacts on harbour porpoises. The overarching goal of chapter 3 is an estimate of the TTS
potential for harbour porpoises from this underwater flowmeter. This was achieved by
underwater noise measurements in the port of Hamburg and the application of data from
previous hearing tests with a harbour porpoise in human care.
A. Study area and underwater noise measurements
The flowmeter of the HPA is located in Hamburg Finkenwerder, which is in the West of
Hamburg in close proximity to the Airbus airport and adjacent to the Natura 2000 conservation
area “Neßsand”. This system consists of two measuring devices, which are located at the north
and south riverbank. The water depth at this site is almost constant 18 m with a very steep slope
in the area close to the riverbanks.
The ultrasonic aqua flowmeter transmits acoustic signals from both sides of the Elbe River to
obtain the flow speed from the measured travel time. The underwater noise recordings were
conducted on two consecutive days (1st and 2nd of June in 2017) on board of the “Reinhard
Woltman” (GER) of the Hamburg Port Authority (HPA). Recordings were made at three depth
levels (2, 5, 8 m). This linear vertical hydrophone array was attached to a rope, which was fixed
with snap hooks to a steel guidance cable. The steel cable was weighted by a 500 kg concrete
anchor and lowered by a winch. At each water depth, underwater noise was recorded by two
hydrophones with different sensitivities, to cover a wide dynamic range (see Fischer and
Schuster 2017 for more details). Underwater noise was recorded using the Avisoft Sound
Recording Software (Avisoft Bioacoustics, Germany). Signals were pre-amplified with an
eight-channel power amplifier (B2008APC – SN1212000, ETEC, DK) with a passband from 1
to 300 kHz. Recordings were digitized with a data acquisition card (NI USB 6251, National
Instruments, USA) sampling at 750 ksamples s-1 and saved as wav files with a 24-bit resolution.
Acoustic signals from the flowmeter on both sides of the Elbe River were recorded on transects
along the river. Therefore the vessel was steered upstream the flowmeter, where the engine was
stopped. The vessel drifted on transects at different distances to the flowmeters downstream,
while the hydrophone array was in the water. Additionally, underwater noise was recorded
during one transect from one flowmeter to the other by diagonally crossing the Elbe River.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
78
During this transect the engine was running. A mobile GPS device (Garmin 72H, CH) on top
of the hydrophone array recorded simultaneously GPS positions at a sample rate of 1 Hz.
The underwater noise recordings, which were gathered on the different transects were analysed
with custom written manuscripts in MATLAB (MathWorks, USA) by DW-ShipConsult (see
Fischer and Schuster 2017 for more details). The sound pressure level (SPL, peak-to-peak) and
sound energy level (SEL) were calculated for each detected signal and assigned to the
corresponding GPS position on the track. The positions of the flowmeters in the North and
South of the Elbe River, the recorded GPS track of the vessel and the positions, where an
acoustic signal could be recorded are shown in (FIG. 13). The sound pressure levels (SPL peak-
to-peak in dB re 1 µPa) are given by colour coded points on the tracks.
FIG. 13: Study area in the Elbe River in the West of Hamburg. The positions of the underwater
flowmeters are shown by red triangles. The GPS tracks of the vessel are presented as lines,
which are separated by colour for different days. The positions where signals of the
flowmeter could be detected in the acoustic recordings are given by points, which show
the determined sound pressure level by a colour coding.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
79
B. Sound propagation model
A sound propagation model was run by DW-ShipConsult based on the determined sound
exposure levels for single acoustic signals and the corresponding locations. The aim of the
sound propagation model was to generate a high-resolution array with modelled SELs of single
signals for the entire area of investigation. The sound transmission of the flowmeter signals is
dependent on the propagation loss but also on the directivity of the sound source. Therefore,
the propagation loss was estimated based on the recordings from the transect between the two
flowmeters only, as this was assumed to be in the center of the beam. The source level was
estimated from these results by the use of a logarithmic regression. The further determined
signals were used to determine the directional characteristic of the transmission beam. As a
result, the source level was determined at a sound pressure level of 210 dB re 1 µPa for both
flowmeters at the north and south river bank. The signal consisted of a 0.2 s tone at a frequency
of 28 kHz with a pulse rate of 4.2 s. The emission of signals alternates between the north and
south flowmeter, leading to an effective pulse rate of 2 s.
The sound propagation was modelled for a three-dimensional area reaching 1000 m (x-axis)
down- and upstream the acoustic flowmeter, a width of 391 m between the two flowmeters (y-
axis) and a depth of 18 m (z-axis). The received level was calculated for each cube within the
three-dimensional array with a resolution of 1×1×0.1 m (x×y×z). This calculation was based on
a simplified ray tracing model and the passive sonar equation. Broadly speaking, the model
assumes a spherical propagation loss of 20×log10(distance) and considers a three-way-
propagation, taking the direct path and the first reflection from the surface and the bottom into
account. This propagation model is described in more detail in Fischer and Schuster (2017).
The estimated source level, propagation loss over distance and the directivity index were taken
into account. The estimated energy for each cube within the array corresponds to the sum of the
direct path and the first reflection from the surface and the bottom. Absorption was not
considered, as this was assumed negligible in freshwater and for short distances.
The results of the sound propagation modelling were delivered as MATLAB files, containing
single sound exposure levels in a three-dimensional array. These files were loaded in R, using
the function read.mat() of the “rmatio” package (Widgren and Hulbert, 2019). The sound field
was visualized as a 2D plot in R (R Core Team, 2019). A top view on the acoustic field for two

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
80
exemplary water depths (5 and 12 m) is shown in FIG. 14 each for the flowmeter in the north
and south.
FIG. 14: Top view on the modelled sound field of the acoustic flowmeter on the north (top row)
and south (bottom row) riverbank. The sound field is shown for a depth of 5 m (A, B) and
12 m (C, D). The sound exposure level of a single pulse is shown by colour coding in
dB re 1 µPa²s.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
81
The single arrays of modelled sound fields for the two flowmeters were combined in a shared
array (FIG. 15). This was done by transferring the arrays to the linear scale, summing up the
values of both arrays and re-transferring the values to the logarithmic scale. This combined
array was used in all further simulations.
FIG. 15: Top view on the combined sound field of the two flowmeters at a depth of 5 m. This
combination derives from summing up the single arrays for the two flow meters. The
flowmeters alternately emit pulses with a pulse rate of 2 s. The pulse rate for each
flowmeter is 4.1 s. The colour coded sound exposure level represents a double pulse,
meaning a pulse of each flowmeter.
C. Effect of 28 kHz fatiguing sound on harbour porpoise hearing
The hearing tests with the harbour porpoise in human care were conducted at the Fjord & Bælt
Centre in Kerteminde (DK). Hearing thresholds were derived from measurements of auditory
evoked potentials (AEP) in the same manner as described in chapter 2. The database derives
from measurements of auditory evoked potentials in 2014, which remain unpublished to data.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
82
The hearing tests were conducted in the project „Cluster 7“ funded by the Federal Agency for
Nature Conservation (FKZ 3514824000). The animal was exposed to a fatiguing stimulus using
the same procedure as described in chapter 2. The fatiguing stimulus was a tone at 28 kHz with
a duration of 3.5 s. The harbour porpoise was exposed once a day with varying sound exposure
levels. Following the same procedure as described in chapter 2, the SEL-TTSonset was defined
as a significant shift from baseline hearing. This TTSonset was estimated by a linear regression
of sound exposure levels (dB re 1 µPa²s) on the delta in the determined hearing threshold (in
dB), compared to the baseline.
D. Simulation of harbour porpoise migration through acoustic field
In order to evaluate the effect of this acoustic underwater flowmeter on the hearing of harbour
porpoises, we simulated swimming tracks through the area. We defined transects in the
direction of the river with a constant y-axis position, passing the flowmeters. We simulated
such transects for each of the 391 m possible y-axis positions over the complete width of the
river. For each of these 391 simulated tracks along the x-axis, we calculated harbour porpoise
positions at the time of the reception of the acoustic flowmeter signals. We calculated the
positions of the harbour porpoise on each track from 1000 m before up to the distance of 1000 m
behind the flowmeters. The first reception was assumed at the starting position at a distance of
1000 m to the flowmeter. Each following reception was obtained from the pulse rate×swim
speed. We used three literature based swim speed levels, which should represent a maximum,
mean and slow swim speed scenario. This broad range was chosen to represent harbour
porpoises at all life stages and to regard also slowly swimming individuals, like mother and calf
pairs. Data on the swim speed of harbour porpoises are rare due to their inconspicuous lifestyle
and poor accessibility. Only a few studies focused on the analysis of swim speed of harbour
porpoises. Maximum swim speed for animals in human care were determined at 4.3 m s-1 (Otani
et al., 2001) and in the wild at 4.3 and 6.1 m s-1 (Gaskin et al., 1974; Otani et al., 2000). A mean
swim speed of a free-ranging harbour porpoise was determined at 0.9 m s-1 (Otani et al., 2000).
Since the authors are not aware of any publication of swim speed levels of harbour porpoises
in river systems, the effect of flow velocity on swim speed was estimated as the sum of the
maximum swim speed and assumed maximum flow velocity. The maximum flow velocity was
estimated at 1.6 m s-1 (pers. comm. Nino Ohle in 2017, HPA). Accordingly, harbour porpoise
flights were simulated at swim speed levels of 0.9, 4.3, 6.1 and 7.7 m s-1. Simulated harbour
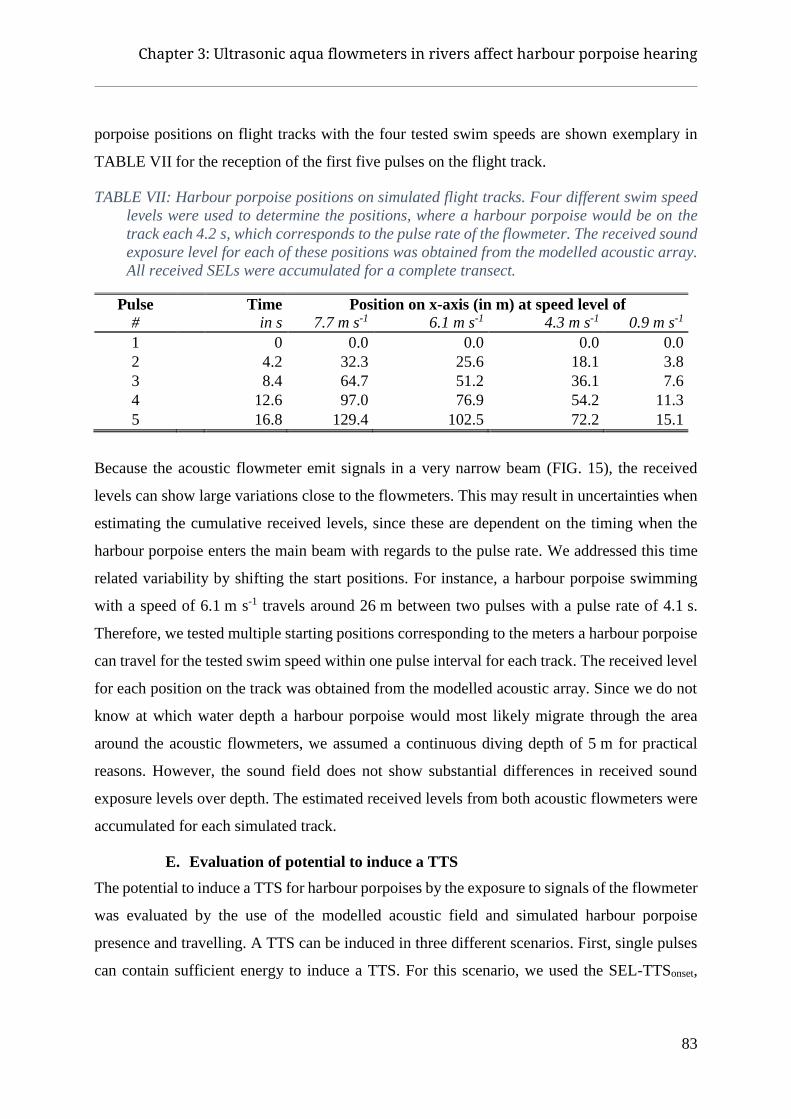
Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
83
porpoise positions on flight tracks with the four tested swim speeds are shown exemplary in
TABLE VII for the reception of the first five pulses on the flight track.
TABLE VII: Harbour porpoise positions on simulated flight tracks. Four different swim speed
levels were used to determine the positions, where a harbour porpoise would be on the
track each 4.2 s, which corresponds to the pulse rate of the flowmeter. The received sound
exposure level for each of these positions was obtained from the modelled acoustic array.
All received SELs were accumulated for a complete transect.
Pulse Time Position on x-axis (in m) at speed level of
# in s 7.7 m s-1 6.1 m s-1 4.3 m s-1 0.9 m s-1
1 0 0.0 0.0 0.0 0.0
2 4.2 32.3 25.6 18.1 3.8
3 8.4 64.7 51.2 36.1 7.6
4 12.6 97.0 76.9 54.2 11.3
5 16.8 129.4 102.5 72.2 15.1
Because the acoustic flowmeter emit signals in a very narrow beam (FIG. 15), the received
levels can show large variations close to the flowmeters. This may result in uncertainties when
estimating the cumulative received levels, since these are dependent on the timing when the
harbour porpoise enters the main beam with regards to the pulse rate. We addressed this time
related variability by shifting the start positions. For instance, a harbour porpoise swimming
with a speed of 6.1 m s-1 travels around 26 m between two pulses with a pulse rate of 4.1 s.
Therefore, we tested multiple starting positions corresponding to the meters a harbour porpoise
can travel for the tested swim speed within one pulse interval for each track. The received level
for each position on the track was obtained from the modelled acoustic array. Since we do not
know at which water depth a harbour porpoise would most likely migrate through the area
around the acoustic flowmeters, we assumed a continuous diving depth of 5 m for practical
reasons. However, the sound field does not show substantial differences in received sound
exposure levels over depth. The estimated received levels from both acoustic flowmeters were
accumulated for each simulated track.
E. Evaluation of potential to induce a TTS
The potential to induce a TTS for harbour porpoises by the exposure to signals of the flowmeter
was evaluated by the use of the modelled acoustic field and simulated harbour porpoise
presence and travelling. A TTS can be induced in three different scenarios. First, single pulses
can contain sufficient energy to induce a TTS. For this scenario, we used the SEL-TTSonset,

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
84
which derives from the hearing tests with a harbour porpoise in human care exposed to a 28 kHz
tone. A TTS can be induced also by multiple exposures. The single SEL-TTSonset cannot be
directly transferred to a multiple exposure, as the same energy of exposure does not
automatically induce the same shift, if the energy is received over a longer duration (as
discussed in chapter 2). The assumption that equal cumulative energy induces an equal hearing
shift is only valid for continuous fatiguing noises or for exposures with similar duty cycles (e.g.
Finneran et al., 2010a, 2010b, Kastelein et al., 2015b, 2014; Mooney et al., 2009; Popov et al.,
2014). We followed instead the suggested noise exposure criteria of Southall et al. (2019, table
6), who suggested a frequency weighted cumulative SEL-TTSonset of 153 dB re 1 µPa²s. Since
this SEL-TTSonset is frequency weighted, based on the audiogram of harbour porpoises
(National Oceanic and Atmospheric Administration; National Marine Fisheries Service, 2016;
Southall et al., 2019), this onset cannot be transferred directly to the single pulse exposure levels
of the modelled sound field. According to the proposed frequency weighting, the amplitude of
a 28 kHz signal decreases by 0.3 dB. This means, that the SEL-TTSonset for multiple pulses
accounts for 153.3 dB re 1 µPa²s. Flight transects were simulated in order to estimate the TTS
potential for a multiple exposure. For this case, we tested, if the cumulative received SEL for
the entire track exceeds the cumulative SEL-TTSonset suggested by Southall et al. (2019). As a
third approach, we evaluated the potential to induce a TTS for harbour porpoises, which do not
show a flight response and rather stay in the area. Based on the sound field of the flowmeter
and the determined pulse rate, we calculated the time for each position, needed to exceed the
cumulative SEL-TTSonset of 153 dB re 1 µPa²s (frequency weighted).

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
85
II. Results
A. Baseline hearing thresholds
Baseline hearing at 28 kHz was measured in 14 trials resulting in a mean hearing threshold of
82.0 dB re 1 µPa ± 2.4 dB (range 78.2-88.1 dB re 1 µPa). We followed the same approach to
define a significant TTS as described for the measurements in chapter 2. With regards to the
determined hearing threshold and the standard deviation, we obtained a critical value (p≤0.05)
of 85.7 dB re 1 µPa as the threshold for a significant hearing elevation. Accordingly, any
hearing shift larger than 3.8 dB was regarded as a TTS.
B. Post-exposure thresholds
Hearing thresholds after an exposure to the 28 kHz fatiguing tone were conducted in eleven
trials. The animal was exposed to SELs between 139.7 and 156.3 dB re 1 µPa²s. Post-exposure
hearing thresholds were measured between 86.7 and 97.6 dB re 1 µPa, accounting for elevated
thresholds between 4.8 and 15.7 dB. Accordingly, all measures post-exposure thresholds
exceeded the critical value of 3.8 dB. The TTSonset was estimated by a linear regression of
threshold shifts on sound exposure levels. According to the linear regression (intercept=-83.95,
slope=0.64), the critical value of 3.8 dB was exceeded at an SEL of 137.4 dB re 1 µPa²s (FIG.
16). Conclusively, this SEL corresponds to the TTSonset for a single exposure to a 28 kHz tone.
C. TTS potential of flowmeter for single pulses
The SEL-TTSonset for a single exposure was determined at 137.4 dB re 1 µPa²s. The underwater
noise measurements and the following sound propagation model could reveal that the source
level of the underwater flowmeter exceeds this TTSonset. This means, that single pulses contain
sufficient energy to induce a TTS in a harbour porpoise. The flowmeters are highly directional
leading to a narrow beam, in which amplitudes are highest. Within the beam of highest
amplitude, the modelled sound fields at a depth of 5 m show single SELs above the TTSonset up
to a distance of approximately 72 m from the flowmeter in the north and 74 m from the
flowmeter in the south. Since the flowmeters are not aligned in a 90° angle to the x-axis these
distances in the beam do not correspond to the distance in the y-axis, meaning the position in
the width of the river. Harbour porpoises which travel along a transect need to keep a distance
of 40 m to the flowmeter in the north and 60 m to the flowmeter in the south in order to avoid
a TTS from a single exposure.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
86
FIG. 16: Effects of 28 kHz tone exposure on hearing thresholds at 28 kHz (black circles). The
horizontal red dashed line represents the critical value of 3.8 dB shift, which indicates the
threshold of significance. A linear regression was calculated in order to estimate the
TTSonset, using only exposures, which led to a TTS. The TTSonset at 28 kHz was determined
at 137.4 dB re 1 µPa²s.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
87
D. Received levels on simulated flights
The received energy for harbour porpoises travelling along the acoustic flow meters were
simulated, assuming swim speed levels between 0.9 and 7.7 m s-1. We accumulated the received
cumulative sound exposure levels for each complete transect line. The results of this simulation
are presented in FIG. 17. The position of the simulated transect line within the width of the river
is shown on the x-axis in m. The received cumulative SEL in dB re 1 µPa²s for the complete
transect line is shown on the y-axis. The received cumulative SEL is given by a black line,
assuming the reception of the first pulse at a distance of -1000m. The grey area indicates the
possible received minimum and maximum received levels. This deviation arises from the pulse
rate and narrow beam of the flowmeter. We tried to address this problem by varying the position
of the first received pulse. The number of variations is dependent on the swim speed and the
pulse rate. For the swim speed of 7.7 m s-1 33 positions of the first reception were simulated
(7.7m s-1×4.2s=32.3 m). The numbers of possible starting positon are given in the top right
corner. This simulation approach indicates, that travelling harbour porpoises with swim speed
levels of 7.7, 6.1 and 4.3 m s-1 would not receive sufficient energy from multiple pulses to
expect a TTS, according to the SEL-TTSonset of 153.3 dB re 1 µPa²s. The TTS-potential is
comparably low. However, harbour porpoises swimming with a speed of 0.9 m s-1 could receive
cumulative sound exposure levels exceeding the SEL-TTSonset up to distances of about 20 m.
The received SELcum is strongly dependent on the timing of pulses. If the harbour porpoises is
within the narrow beam of highest amplitude at the moment, when a pulse is emitted, the
received cumulative SEL can be as high as 165 dB re 1 µPa²s for all swim speed levels and a
TTS must be assumed.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
88
FIG. 17: Received cumulative sound exposure levels (SELcum) on simulated transects along
acoustic flowmeters.The simulation was performed assuming four different swim speed
levels (0.9-7.7 m s-1) and a continuous depth of 4.7 m. The received SELcum (black line)
represents the cumulative SEL a harbour porpoise would receive, when travelling along
a constant position within the width of the river (y-axis). The red horizontal line represents
the cumulative SEL-TTSonset of 153.3 dB re 1 µPa²s, estimated by (Southall et al., 2019).
The variability in the received SELcum, which derives from the time of entering the beam
of highest amplitude, is shown by a blue area, which represents the minimum and
maximum values. The number of simulated starting positions for the reception of the first
pulse is shown in the top-right corner.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
89
E. TTS potential for harbour porpoise which stay around the flowmeters
In order to estimate the potential to induce a TTS in harbour porpoises, which do not show a
flight response and rather stay within the area, we calculated the needed time to exceed the
cumulative SEL-TTSonset. This was done for each position within the modelled sound field and
the pulse rate of 4.1 s. FIG. 18 displays a top-view on the sound field at a depth of 5 m. The
needed time to exceed the cumulative SEL-TTSonset in minutes is presented by colour coding.
Only the area where a TTS can be induced by multiple pulses within 60 minutes is marked. The
area of highest potential to induce a TTS is between the two flowmeters. If the harbour porpoise
stays within this area a TTS can be induced within 1 and 30 minutes. The risk for a TTS strongly
decreases, when leaving the narrow beam of highest amplitude.
FIG. 18: Required time within sound field to exceed cumulative SEL-TTSonset.The time in
minutes is presented (colour coded), which is needed to exceed the cumulative SEL-
TTSonset of 153.3 dB re 1 µPa²s. Only the areas, where a TTS can be induced within
60 minutes for a pulse rate of 4.1 s are marked. The flowmeter positions are marked by
black triangles.

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
90
III. Discussion
The distribution of harbour porpoises largely overlaps with areas where anthropogenic activities
take place. A perfect management of these activities would prevent any negative impact on
harbour porpoise health or behaviour but simultaneously maintain anthropogenic activities. The
results of chapter 3 could show that the acoustic flowmeter has the potential to induce a
temporary threshold shift in harbour porpoises by the exposure to single, but also to multiple
signals.
A. Evaluation of potential effects of the flowmeter on harbour porpoise
hearing
The hearing tests with a harbour porpoise in human care could show, that a 28 kHz signal with
a length of 3.5 s can induce a TTS, if sound exposure levels exceed 137.4 dB re 1 µPa²s.
Transferred to the modelled sound field of the acoustic flowmeters in the Elbe River, a TTS can
be induced by the exposure to single signals at close distances up to 74 m. Since harbour
porpoises have good hearing abilities at 28 kHz (e.g. Ruser et al., 2016a) it is very likely, that
signals can be heard from larger distances and animals might not come so close to the
flowmeter. The simulation of harbour porpoises travelling along the acoustic flow meter could
reveal a high variability in the received cumulative SEL. This large variability is due to the
pulse rate of 4.1 s and the narrow beam of the flowmeter. Although harbour porpoises
swimming at lower speed levels would receive more pulses on the complete track, the highest
possible received levels, due to the timing variability, do not differ significantly between
different swim speed levels. This clearly demonstrates that the timing and the position of signal
reception are the most important factors for the received cumulative SEL. The simulation could
show, that received cumulative SELs of up to 165 dB re 1 µPa²s are possible for travelling
transect at close distances (FIG. 17). Following the recommend noise exposure criteria for
marine mammals (Southall et al., 2019), a permanent threshold shift is expected at a cumulative
SEL of 173 dB re 1 µPa²s. The highest SEL within the acoustic field was estimated at a SEL of
165 dB re 1 µPa²s. To exceed the PTS-SELonset of 173 dB re 1 µPa²s, seven pulses with a SEL
of 165 dB 1 µPa²s would be needed. Accordingly, a harbour porpoise would need to stay at
close distance to the flowmeter for 7×4.1 s=28.7 s. Due to this relatively long time, a harbour
porpoise must stay at close distance to the flowmeter, it is very unlikely, that a PTS will be
induced for this assumed TTSonset. The presence of harbour porpoises in the port of Hamburg is

Chapter 3: Ultrasonic aqua flowmeters in rivers affect harbour porpoise hearing
91
believed to be driven by the migration of prey species (Wenger et al., 2016). Since many fish
species do not show good hearing abilities at frequencies as high as the 28 kHz flowmeter
signals (Ladich and Fay, 2013), it is reasonable to assume, that the flowmeter does not have a
large effect on their distribution. If prey species of harbour porpoises aggregate close to the
flowmeter, it could be that harbour porpoises try to get access to this important food source
despite high sound exposure levels. If the energetic benefit is high enough, harbour porpoises
could tolerate the exposure to these signals. Our simulation in FIG. 18 could show, that along
the entire width of the Elbe River, a period of 60 minutes is sufficient to induce a TTS and even
decreases with decreasing distance to the flow meters.
B. Effects on behaviour
Effects of anthropogenic noise are numerous and must not be restricted to injuries and death.
The introduction of underwater noise within the hearing range of harbour porpoises can also
elicit behavioural changes. This could be shown for example by Kastelein et al. (2014a), who
found behavioural changes, when a harbour porpoise in human care was exposed to pinger
signals. These devices are similar in the frequency range to the acoustic flowmeter. Harbour
porpoises reacted to the introduction of pinger signals by an increased swim speed and breathing
rate, jumping out of the water and increased distances to the transducer (Kastelein et al., 2014a).
Accordingly, it is likely that harbour porpoises are likewise disturbed by the flowmeters in the
port of Hamburg and try to avoid close distances. Such disturbance effects may even result in
decreased individual fitness and could lead to long-term population consequences (King et al.,
2015).
C. Management approach
The underwater measurements in the port of Hamburg and the following evaluation of potential
effects on harbour porpoises were conducted on behalf of the Hamburg Port Authority. The
outcomes of the project were presented to the responsible employees of HPA (Schaffeld et al.,
2018). Although the area of highest impact was limited to close distances of below 100 m and
in a very narrow beam, the HPA targeted at an improvement of the current situation. Due to the
lack of an established monitoring of harbour porpoises in the Elbe River, we do not have any
information if harbour porpoises regularly occur close to the acoustic flowmeters. Although
harbour porpoises were sighted up- and downstream the acoustic flowmeters (Wenger et al.,
2016) it could be the case, that passing the sound field affects harbour porpoises. That risk could

Conclusion
92
be taken, if the expected benefit is high. We do not have detailed information on the overall
application of these devices in large river systems. In addition to this described flowmeter in
the port of Hamburg, the HPA runs two more flowmeters upstream the Elbe River (pers. comm.
HPA). The presence of harbour porpoise in this area is assumed lower, but the current database
is insufficient for reliable estimates. It could be also the case, that further flowmeters are applied
along the Elbe River or other river systems, which we are not aware of. It is reasonable to
assume, that harbour porpoises have to pass further acoustic barriers during their annual
upstream travel. Our results demonstrate that the risk for a TTS is highest in a comparably small
area up to 74 m from the source. This leads to the conclusion that travelling harbour porpoises,
which want to pass this acoustic field should rather travel in the middle of the Elbe River.
However, this outcome raises the need for an adjustment of the acoustic flowmeter. Since the
Elbe River is highly exposed to ship traffic with vessels of all kinds, which travel in the middle
of the river, harbour porpoises could be forced to elude closer to the riverbanks, where the
flowmeters are. Unfortunately, this is the area with highest levels of the acoustic flowmeter and
the highest potential to induce a TTS. Since this combination of simultaneous harbour porpoise
and vessel presence is a problem, the HPA decided to adjust the source levels of the flowmeters.
Following the provided evaluation, the HPA instructed a reduction of the flowmeter source
levels.
IV. Acknowledgement
We would like to thank the Hamburg Port Authority for funding this project. We would also
like to express our gratitude to the team of the vessel “Reinhard Woltman” who supported us
during the measurements of underwater noise.

Conclusion
93
Conclusion
I. Evolution of noise mitigation
Anthropogenic noise pollution has been recognized as one of the most hazardous forms of
pollution, which is omnipresent in all terrestrial and aquatic ecosystems (Andrew et al., 2002;
World Health Organization, 2017) leading to environmental changes, which are considered as
a key threat to biodiversity at an unpredictable scale (Kunc et al., 2016). The introduction of
underwater noise arises from the industry, e.g. during the construction and operation of offshore
wind farms (OWFs) (Brandt et al., 2018; Dähne et al., 2013; Graham et al., 2019) or shipping
(e.g. reviewed in Erbe et al., 2019) as an unintended by-product. The clearance of unexploded
ordnance from World War II represents a further unintended but regularly occurring noise
source (von Benda-Beckmann et al., 2015). Anthropogenic noise pollution occurs, however,
also on purpose for seismic explorations of the seafloor (e.g. Sarnocińska et al., 2020), military
sonar (Curé et al., 2016; Filadelfo et al., 2009; Isojunno et al., 2016) or to acoustically deter
animals (Brandt et al., 2013b, 2013a; Götz and Janik, 2013).
The understanding of effects of noise on marine life evolved over the last decades and is still
scarce. Only 14 years ago an intensely discussed research project ended, investigating the large-
scale thermal fields in the North Pacific Ocean by sending intense acoustic low frequency
signals over large distances without any prior risk assessments for marine life (Dushaw et al.,
2009). This study examined thermal trends by measuring the travel time of sounds between
measuring positions thousands of kilometres away, taking advantage of the correlation between
speed of sound and temperature (Dushaw et al., 2009). The acoustic thermometry of ocean
climate (ATOC) is just one example, which vividly displays the naive emission of underwater
noise without considering potential effects on the marine environment. Only recently, potential
effects of noise pollution awakened awareness. A relationship between the emission of
underwater noise and biological implications was first reported in 1974 after the stranding of
four Cuvier’s Beaked Whales (Ziphius cavirostris) in the Caribbean, which was suspected to
be a result from some kind of underwater explosion (van Bree and Kristensen, 1974). Various
mass stranding events of beaked whales e.g. in Greece in 1996, in the Caribbean in 1998 or the
Bahamas in 2000 led the international Whaling Commission’s Scientific Committee explain
that “there is now compelling evidence implicating military sonar as a direct impact on beaked
whales in particular” (reviewed in Simmonds et al., 2014). A great step forward in noise

Conclusion
94
regulation was achieved by the identification of the mechanism of the “gas and fat embolic
syndrome” as the underlying mechanism of noise-related mortalities in beaked whales (Cox et
al., 2006). This mechanism causes haemorrhaging around the brain and lesions in vital organ
tissues, which were identified in several cases of stranded cetaceans and were accordingly under
suspect to be caused by the exposure to intense acoustic signals (Fernández et al., 2005; Jepson
et al., 2003; Siebert et al., 2013).
Despite military activities, further anthropogenic noise sources contribute to the noise pollution,
including impulsive noise from seismic surveys (Bain and Williams, 2006; Sarnocińska et al.,
2020) and pile driving strikes, generated during the construction of offshore wind farms (Brandt
et al., 2018; Dähne et al., 2013), together representing the most prominent contributors. The
effect of impulsive noise on harbour porpoise hearing was first investigated by Lucke et al.
(2009) after it was suggested to define noise exposure criteria for marine mammals (Southall et
al., 2007). These were based on data from bottlenose dolphins (Tursiops truncatus) and belugas
(Delphinapterus leucas), which were assumed to be questionable in transferability for high
frequency hearing cetaceans like the harbour porpoise (Southall et al., 2007). The exposure to
airgun pulses induced a TTS in a harbour porpoise in human care, if received levels exceeded
164.3 dB re 1 µPa²s (Lucke et al., 2009), showing that harbour porpoises are more sensitive to
impulsive noise than assumed (Southall et al., 2007). The measured TTSonset was subsequently
adopted in a regulation of noise mitigation during the construction of offshore wind farms in
Germany by the Federal Ministry for the Environment, Nature Conservation, and Nuclear
Safety (BMU, 2014). This comparably fast implementation of the TTSonset in noise mitigation
regulations clearly demonstrates the willingness to regulate underwater noise, but it has to be
taken into account that therefore reliable thresholds are neccessary. Although this is a good
example for a regulation of underwater noise, it might still be insufficient to prevent a TTS
from pile driving noise in harbour porpoises (Kastelein et al., 2015a, 2016). Moreover, to date
further anthropogenic noise sources remain unregulated.
This thesis aimed to assess the effect of anthropogenic noise pollution from different sources
on harbour porpoise hearing. The three chapters represent an evolution of the understanding of
noise effects. The introduction of underwater noise occurs in different scenarios, which can be
on purpose or unintended, some are already regulated or are tried to be regulated or even

Conclusion
95
replaced by alternatives. The contributions of this thesis demonstrate that improvements in
noise mitigation are required in all of these exemplified scenarios of noise pollution.
A. Unintended effects of underwater sound application
The field of research regarding potential effects of anthropogenic underwater noise is relatively
new. This could be the reason why acoustic devices are still in use without considering potential
biological effects, which is exemplary shown in chapter 3). The results of this thesis illustrate
that the acoustic flowmeters in the port of Hamburg emit signals with sufficient energy to
induce a temporary threshold shift (TTS) in harbour porpoises. The onset of a TTS (TTSonset)
was determined at a received sound exposure level (SEL) of 138 dB re 1 µPa²s for a tonal
signals with a peak frequency of 28 kHz. The cumulative sound energy levels an animal could
receive from the acoustic flowmeter sound signals can be as high as a maximum of
165 dB re 1 µPa²s. Accordingly, up to a distance of 72 m from the sound source (which
corresponds to a distance to the border of 60 m) received SELs exceeded the TTSonset. Results
from the simulation approach demonstrate that received cumulative sound exposure levels for
harbour porpoises travelling along the flowmeter can be as high as 165 dB re 1 µPa²s. The
received levels for harbour porpoises, which travel along the flowmeter are found to be highly
variable. This is due to the high directionality of the flowmeters and the pulse interval of 4.2 s.
Results demonstrate that the risk for a TTS is highest in a comparably small area up to 72 m
from the source. This leads to the conclusion that travelling harbour porpoises, which want to
pass this acoustic field are well-advised to rather travel in the middle of the Elbe River.
However, this outcome raises the need for an adjustment of the acoustic flowmeter. Since the
Elbe River is highly exposed to ship traffic with a high variety of different vessels types, which
travel in the middle of the river, harbour porpoises could be forced to elude closer to the
riverbanks. Unfortunately, this is the area with highest levels of the acoustic flowmeter and the
highest potential to induce a TTS. Since this combination of simultaneous harbour porpoise and
vessel presence is a problem, switching off the flowmeter in the presence of vessels could be
an option. A good solution could be a dynamic switch, which recognizes increased background
noise caused by vessels and then turns off the flowmeter.
Furthermore, the necessity for emitting signals at such high pulse rates of 4.2 s from each side
is doubtful. Therefore, the pulse rate could be adjusted to a temporal resolution needed for
determination of flow speed trends. In this case, the source levels need to be increasing

Conclusion
96
gradually, if the pulse rate decreases. This is necessary to prevent, that harbour porpoises are
surprised by a high intense signal at close ranges. The advantage of the currently high source
levels and short pulse intervals is that travelling harbour porpoises can hear the signal at
sufficient distances and are warned previously. In theory, this reduces the probability that
harbour porpoises enter the area of highest energy. A possible adjustment could be that the flow
speed is measured on an hourly basis. A measurement could start with signals of reduced
energy, which are gradually increased at steps of 3 dB, giving the harbour porpoise time to flee.
For instance, the sound exposure level could start at 102 dB re 1 µPa²s and will be increased at
steps of 3 dB each 4.2 s. After the emission of 13 signals, corresponding to 50 s, the TTSonset
will be exceeded. This would give the harbour porpoise sufficient time to leave the area of
highest intensity, until the actual flow velocity measurement starts. To my understanding, such
an adjustment represents an effective noise regulation and should be considered, if technically
feasible. The underwater measurements revealed that the acoustic flowmeters on the northern
and southern bank of the Elbe River were misaligned. Accordingly, an adjustment of the
orientation could allow for lower source levels, without any loss in quality of the results.
The outcomes of chapter 3 cast a new light on the thoughtless application of acoustic devices.
This raises the question, if this is just a rare example of thoughtless noise pollution or if this
happens also in other situations. From the results of chapter 2, it is clear, that this is not the only
example. Seal scarers were likewise naively introduced to prevent a TTS from pile driving
events (BMU, 2014), without a careful evaluation of further effects. It could be demonstrate,
however, that these devices have the potential itself to induce a hearing shift. This needs to be
considered in any future applications of seal scarers. As a further example, there is a demand
for an alternative to seismic airguns to locate deposits of fossil fuels in the seafloor, since
airguns can impair harbour porpoise hearing (Lucke et al., 2009) or affect behaviour of marine
mammals (Sarnocińska et al., 2020). Recent developments have yielded at alternative marine
seismic sources with expected reduced environmental impacts while maintaining efficiency
(Duncan et al., 2017; Pramik, 2013). The marine vibroseis could represent such an alternative
to airgun arrays. The emitted signals are of lower amplitude, longer duration and smaller
bandwidth than airgun pulses (Pramik, 2013; Tenghamn, 2006). Since most of the energy is at
frequencies below 200 Hz and amplitudes are lower, the potential to induce a TTS is assumed
to be lower than for airguns (Duncan et al., 2017; Pramik, 2013). However, the acoustic signals

Conclusion
97
are of longer duration than airgun signals and the sound exposure levels could be similar. In
addition, these signals consist of tone sweeps at frequencies below 100 Hz (Duncan et al., 2017;
Tenghamn, 2006). The combination of longer durations and tones at certain frequencies
increases the potential to mask biologically relevant signals (reviewed in Erbe et al., 2016).
These frequencies overlap with the typical frequency range of blue or fin whale communication
(Širović et al., 2007). Before such devices are applied on a wide scale, potential effects have to
be investigated to exclude that marine life is affected. This is done, for instance, in an ongoing
project of the Institute for Terrestrial and Aquatic Wildlife Research with blue whale sound
exposure experiments in Iceland.
The outcomes of this dissertation emphasize the need for a general previous authorization
process prior to the use of underwater sound for any purpose with potential effects on aquatic
life. It has to be considered that potential effects for target and non-target species are carefully
reviewed.
B. Naive implementation of underwater sounds
The example described in chapter 2) demonstrates an approach to deter marine mammals out
of a certain area by introducing of special underwater sounds. Acoustic deterrent devices are
widely used to counteract the economic loss by seal depredation (Dawson et al., 2013; Götz
and Janik, 2013) around aquacultures or to deter harbour porpoises out of the vicinity prior to
pile driving events (Brandt et al., 2013a). The outcomes of chapter 1 clearly confirm that prior
deterrence is substantially needed to protect harbour porpoises from a TTS appearing after the
reception of multiple pile driving strikes. However, this thesis also confirms the potential of a
single seal scarer signal to induce a TTS in harbour porpoises at estimated distances between
211 m and 5.9 km. These contradictory outcomes indicate the need for a thorough regulation
of underwater noise emissions during the construction of offshore wind farms, which needs to
be adopted in noise protection concepts (e.g. BMU, 2014). Therefore, source levels need to be
down regulated and slowly increased, giving harbour porpoises enough time to flee, when
applied as a regulation to prevent TTS. In order to minimize temporary habitat loss and potential
food sources, the source levels of ADDs must be adjusted to the desired deterrence range. One
limitation of our suggested implementation is the gap of knowledge concerning a reliable
correlation between deterrence range and received levels.

Conclusion
98
In addition to the application as a mitigation tool, the results raise the need for a regulation in
form of an authorization process and monitoring for the application of seal scarers around
aquacultures. Although ADDs are a significant and chronic source of underwater noise
pollution along the Scottish west coast around aquacultures with a steady increase in ADD
usage and substantial geographic expansion (Findlay et al., 2018), neither a license is required
to deploy these devices nor any statistics on the usage exist (Coram et al., 2014). ADDs were
deployed to counteract economic loss by seals. The observed deterrence of harbour porpoises,
which accompanies their usage, has been regarded as a comparatively benign side effect
(reviewed in Götz and Janik 2013). Nevertheless, the introduction of ADD signals significantly
influences the quality of a habitat and should be therefore considered in the evaluation of the
environmental status in the Marine Strategy Framework Directives (MSFD) on a European
level (Tasker et al., 2010). The extensive usage of ADDs around marine aquacultures represents
a local major source of chronic anthropogenic noise pollution, which has to be addressed in
national marine registries to meet international protective agreements, e.g. MSFD and Oslo and
Paris Convention (OSPAR) (cited from Findlay et al., 2018). Regarding the previously reported
behavioural responses of harbour porpoises to seal scarer signals, such as increased swim speed,
porpoising behaviour, increased breathing rates (Kastelein et al., 2015c), but also the exclusion
from habitats (Brandt et al., 2013a) can be assumed to pose a high risk for chronic stress and
potential cumulative effects on general health. Nevertheless, these effects are evaluated to date
as insufficient arguments to regulate the use of ADDs (Coram et al., 2014; Findlay et al., 2018).
The evidence provided in chapter 2 of this thesis that a TTS or even PTS can be induced should
lead to the development of a regulation system.
C. Established concept for noise mitigation
Chapter 1) is dedicated to the mitigation of pile driving noise, which was recognized as a threat
for harbour porpoise hearing (Lucke et al., 2009) and a concept for regulation was already
introduced six years ago (BMU, 2014). Nevertheless, this thesis could show that further
measures are urgently needed to consider the TTS potential from multiple exposure.
Regarding the protection of harbour porpoises, Germany has enforced the strictest regulations
in the EU to date. Single pile driving strikes must not exceed a SEL of 160 dB re 1 μPa²s at a
distance of 750 m, in order to prevent a temporary threshold shift. Regarding a TTS as an injury
is outstanding, since most other European countries and the United States of America consider

Conclusion
99
a PTS as an injury and regulate noise accordingly (reviewed in Stöber and Thomsen, 2019).
Furthermore, wind farm operators are obliged to deter harbour porpoises prior to pile driving
activities and to use a soft start procedure with limited hammer force and longer pulse intervals
in the beginning of pile driving, following German regulations (BMU, 2014). However, current
regulations do not consider the evident TTS potential from the exposure to multiple pile driving
strikes, which previously has been scientifically proven (Kastelein et al., 2015a, 2016). This
thesis provides a novel evaluation tool, which allows estimating the TTS potential for harbour
porpoises, fleeing during pile driving activities, representing a significant contribution to an
effective protective noise regulation. This tool considers current knowledge on TTSonset for
multiple pile driving strikes, harbour porpoise swim speed capacity and sound propagation of
pile driving strikes in a shallow region in the German North Sea. One of the key benefits of this
evaluation tool is that all applied variables are easy to adjust for different soundscapes or for
different TTSonsets. The simulation approach of chapter 1 supports current noise mitigation
regulations during the construction of OWF in Germany as a valuable measure for protecting
harbour porpoises not only from single but also from multiple pile driving strikes.
Based on this simulation approach, only the combination of restricting the maximum single
strike sound exposure level (SELSS) to 160 dB re 1 µPa²s at a distance of 750 m, a previous
deterrence and a soft start with reduced energy and longer pulse intervals allow harbour
porpoises to avoid a TTS from multiple exposures. To my understanding, there is no reason to
question the current single strike regulation. This simulation approach highlights the need for
considering also the introduced minimum deterrence distance (MDD). The MDD corresponds
to a distance, a harbour porpoise must be at least deterred prior to any pile driving activity for
being able to escape from a TTS by a continuous flight. The MDD is mostly depending on the
possible swim speed of harbour porpoises, the source level and sound propagation. A MDD of
2.4 km was determined for harbour porpoises, which are capable of swimming at a speed of
6.1 m s-1 and even of 4.7 km for harbour porpoises fleeing at a speed of 0.9 m s-1. Any tolerated
increase in maximum levels of single pile driving strikes exceeding the 160 dB re 1 µPa²s at a
distance of 750 m would substantially affect the already large minimum deterrence distance.
This dissertation aims for raising the attention for further critically needed studies, which tackle
the gap of an empirical based effective quiet threshold (EQT). In the absence of such a
threshold, a proxy was considered, named “still higher than effective quiet threshold” (SHEQ).

Conclusion
100
As a best guess, the lowest SELSS, which has been proven to induce a TTS, was chosen as the
SHEQ. This emphasizes the need for a real effective quiet threshold, which determines the safe
distance for harbour porpoises, where a TTS cannot be induced. The absence of an EQT
constraints the ability to regulate mandatory minimum deterrence distances.
A challenging problem of this simulation approach is that equal energy does not automatically
have to generate the same amount of TTS, because the TTSonset is depends on the duty cycle,
the SPL and the received SELcum (Kastelein et al., 2015b, 2014b). Regarding our simulation
approach we assume, that the TTSonset for fleeing harbour porpoises could be lower than the
proposed value of 175 dB re 1 µPa²s, which derives from a study with constant exposure to
single strikes with an SEL of 145 dB re 1 µPa²s (Kastelein et al., 2016). However, a fleeing
harbour porpoise would receive pile driving strikes with decreasing SELSS and increasing
distance to the sound source. In contrast to Kastelein et al. (2016), the harbour porpoise would
receive multiple strikes with variable SELSS, which are all above the SELSS in that playback
experiment. To my knowledge, no study has yet yielded at a TTSonset for varying or decreasing
SELSS over time.
II. Outlook
The analyses in this thesis raise further research questions, which should be addressed in future
projects:
A. Acoustic flowmeter
The Elbe River is highly exposed to ship traffic, which contribute significantly to the
underwater soundscape. However, this area is most likely of high ecological importance for
harbour porpoises and provide a great benefit, because harbour porpoises regularly occur in this
noisy area. Future studies should investigate the habitat usage of harbour porpoises in this area
to determine its ecological importance. Currently no systematic monitoring of harbour
porpoises in large river systems exists, although regularly sightings are reported (Wenger et al.,
2016; Wenger and Koschinski, 2012). A monitoring is substantially needed for harbour
porpoises, which are protected throughout their range, for better understanding the driving
factors for entering rivers. In addition to potential hearing impairments, these devices can also
induce behavioural changes or affect harbour porpoise migration. It is reasonable to assume,
that entering the sound field elicits discomfort or stress in the animals. In this case, the acoustic
flowmeter could create a barrier effect. Future research should focus on a potential barrier effect

Conclusion
101
and habitat exclusion. Furthermore, this acoustic flowmeter in the port of Hamburg is most
certainly not the only position, where the water flow is measured. It is reasonable to assume
that multiple, more similar devices emit acoustic signals in larger river systems like the Elbe or
Weser River. Information on the existence of additional acoustic flowmeters should be
collected in future studies, since these could represent a source of potential injury for present
harbour porpoises (Wenger and Koschinski, 2012). To the best of my knowledge, there is no
prior approval procedure needed to apply these acoustic devices. A legal obligation for the
introduction of underwater noise with the potential to induce hearing impairment for harbour
porpoises should be carefully considered. This possible barrier with a TTS potential should be
also addressed for harbour seals, since these animals also occur in this area and are capable of
hearing the acoustic flowmeter signal (Kastelein et al., 2009a, 2019a). Habitat exclusion is most
likely for seals, since the frequency of the flowmeter signal is represented also in signals of
acoustic deterrent devices and should therefore be considered in future research.
B. Acoustic deterrent devices
A large selection of different acoustic deterrent devices exists with an increasing field of
application possibilities. Available devices differ in signal amplitude, frequency and duration
or duty cycle. This increases the necessity to examine their potential to induce a TTS in harbour
porpoises or other marine mammals. In order to prevent side effects on non-target species,
hearing abilities of these species should be considered in the development of ADD signals. This
will become even more important, if the field of application increases, for example as a
deterrent device prior to the clearance of unexploded ordnance of World War II in German
waters. Additionally, dose responses of received levels and resulting deterrence ranges, should
be investigated to adjust the source levels of ADDs to the deterrence range desired. Reported
deterrence ranges are highly variable and do not allow for reliable estimates of dose responses
to date (reviewed in Götz and Janik 2013).
III. Future directions of TTS research
A. Biological mitigation as a regulation approach
Another novel finding is the ability of at least four toothed whales to actively attenuate auditory
brainstem responses, which is believed to serve as a protective mechanism (Finneran, 2018;
Nachtigall et al., 2016, 2017; Nachtigall and Supin, 2014). This conditioned “self-mitigation”
in the expectancy of an intense sound exposure, led to elevated hearing thresholds up to 40 dB

Conclusion
102
(Finneran, 2018). This conditioned mechanism is believed to protect from outgoing
echolocation clicks, which contain very high source levels, but also from an expected exposure
to underwater noise (Finneran, 2018; Nachtigall et al., 2017). This conditioned attenuation is
most efficient, if the animals expected a following intense exposure. This raises the question, if
the ability of self-mitigation can serve, to protect from anthropogenic noise effects. Based on
the current knowledge, a previous signal would be needed to warn animals, for example before
pile driving activities start. To date it is not possible to reliable estimate, how many conditioning
trials are needed to develop a protection against an impending sound. Furthermore, harbour
porpoises are believed to use large spatial areas, so animals in the surrounding area could be
“conditioned” or are still naive. The main limitation is the lack of knowledge about the
underlying mechanism of this conditioned attenuation, which does not allow for reliable
estimates of effectiveness and duration. It also remains unsolved to date, if such a mechanism
derives from middle ear (conductive) mechanisms or from a neural mechanism involving
efferent inhibitory projections (Finneran, 2018; Nachtigall et al., 2017). Further research is
needed to assess these limitations and to determine, if any impairment still occurs, which cannot
be seen by measuring the hearing threshold. Current knowledge is insufficient to consider this
described self-mitigation as a tool to prevent TTS.
B. Understanding of TTS
This thesis provides scientific evidence for a TTS potential from a single exposure to seal scarer
signals. However, this evaluation cannot reliably estimate a TTS potential for the reception of
multiple signals. Regarding the randomization of inter pulse intervals of seal scarers at intervals
between 0.6 and 90 s (cited in Mikkelsen et al., 2017), a TTSonset cannot be estimated without
the understanding of the relationship between TTS growth and exposure variables like SPL,
duty cycle and cumulative SEL. This represents also a limitation for the simulation approach
of chapter 1. The TTSonset, which was applied in the simulation approach (Kastelein et al.,
2016), derives from an experiment with a repeated exposure to signals of the same source level.
In the case of a fleeing harbour porpoise, the received levels are expected to decrease over time,
when the distance to the sound source increases. Based on current knowledge about TTS in
harbour porpoises, it is reasonable to assume that a TTSonset from a multiple exposure could be
lower, if the received levels start higher and decrease over time. The relationship between

Conclusion
103
received levels, duty cycles and corresponding TTS growth in cumulative exposures should be
addressed in future studies. This would represent a huge step in estimates of TTS potential.
C. Ecological relevance of temporary threshold shifts
This study investigated the potential of different anthropogenic noise sources to induce a TTS
in harbour porpoises, which is a common approach to assess the effect of noise pollution
(reviewed in Finneran, 2015). However, the implications of a TTS remain unknown to date.
Future studies are critically needed to determine the ecological relevance of a temporary hearing
shift. To the best of the author’s knowledge, no one was able to determine ecological
consequences of hearing impairment to date, although no one would question that harbour
porpoises critically rely on hearing. Echolocation is a very high-evolved specialisation towards
living in a niche environment (Fenton et al., 2014). Any interference in acoustic perception can
reduce the small advantage of predators and could have strong effects on their survival.
Assuming that harbour porpoises show a low resilience to any disturbance, which could be
represented by their continuous foraging to maintain energetic demands, any disturbance might
have severe effects (Wisniewska et al., 2016). This situation is aggravated by the health status
of harbour porpoises in the German North- and Baltic Sea, which show a high prevalence for
parasites in lungs and ears as well as associated bronchopneumonia (Lehnert et al., 2005; Morell
et al., 2017b; Siebert et al., 2001). Most of the risk assessment studies assume a healthy
population, which is not the case. This gap of knowledge strengthens the need for further
research on infectious agents and other pathological findings, which might affect hearing
abilities (Morell et al., 2017b; Prahl et al., 2011; Wohlsein et al., 2019) and potentially the
ability to recognize possible threats, such as set-nets (Kastelein et al., 2000; Villadsgaard et al.,
2007). Further insight into ecological effects of TTS, stress responses and the health status of
harbour porpoises could help to shed light on the dramatic situation of harbour porpoises, which
often die before females are sexually mature (Kesselring et al., 2019).
To date, the consequences of a temporal hearing shift at 4 (Lucke et al., 2009), 8 (Kastelein et
al., 2016), 14 or 28 kHz (Schaffeld et al., 2019) for harbour porpoises are unknown. Thus,
assumptions about ecological relevance would be highly speculative. These comparably low
frequencies are all in the hearing range of harbour porpoises but far away from the range of best
hearing, which is around 130 kHz (Kastelein et al., 2002, 2010; Ruser et al., 2016a),
corresponding to the frequency of echolocation clicks (Møhl and Andersen, 1973; Villadsgaard

Conclusion
104
et al., 2007). However, it is reasonable to assume, that hearing lower frequencies is of
highimportance for harbour porpoises, because it was preserved in evolution, when
echolocation ability and high frequency hearing evolved. The actual cause for this ability
remains unknown, giving the chance to hypothesize and develop further research ideas. A
possible explanation could be, that acoustic signals of predators should be recognized in order
to elicit antipredator behaviour, like it has been shown for multiple cetaceans exposed to
predator signals (Aguilar de Soto et al., 2020; Bowers et al., 2018; Curé et al., 2013; Jacobson
et al., 2015; Rankin et al., 2012). Actually, in the absence of regularly occurring killer whales
or bottlenose dolphins in German waters as typical predators of harbour porpoises, the predation
of grey seals on harbour porpoises has been revealed recently (Bouveroux et al., 2014; Heers et
al., 2018; Jauniaux et al., 2014; Leopold et al., 2015). This marine top predator of German
waters is highly vocal underwater, especially within the breeding season (Asselin et al., 1993),
and harbour porpoises could take advantage of acoustic cues to avoid them.
The ability to hear lower frequencies can also support harbour porpoises in their ability to find
prey by passive listening. Likewise, bottlenose dolphins elicited attraction to playbacks of
soniferous fish signals (Gannon et al., 2005), which represent a large amount of their diet
(Burros and Myrberg, 1987). The ability to localize prey by passive listening could be of high
importance for harbour porpoises since their echolocation beam is highly directional and
adapted for short distance localization (Koblitz et al., 2012). Eavesdropping on fish, which were
found to acoustically communicate to a large extent (reviewed in Ladich, 2019), could help to
find prey over larger distances. This could be very advantageous in a seemingly endless
environment with patchy distributed prey at decreasing densities. Future research should
investigate, if harbour porpoises take advantage of passive listening to find prey. If it turns out
that harbour porpoises use low frequencies to find prey, the effect of masking becomes more
important than previously assumed. Masking is believed to be of less importance for harbour
porpoises, since echolocation signals are at much higher frequencies than anthropogenic noise
pollution occurs. However, if a dependence on low frequent fish sounds exist, these biological
relevant signals are likely overheard in the presence of high background noise levels.
In addition to potential effects of masked low frequency signals, harbour porpoises could be
distracted by the presence of a TTS. Likewise, hearing impaired human listeners perform worse
in distinguishing two different simultaneous heard messages, if ambient noise increases,

Conclusion
105
although no differences to the normal hearing group can be seen in situations of low ambient
noise (Best et al., 2010). If this is also the case for harbour porpoises, the ecological relevance
of a TTS might dramatically increase due to the high levels of noise pollution in the areas
inhabited by harbour porpoises.
The prevention of a TTS is usually proclaimed as a management goal in underwater noise
regulation (e.g. in Germany, BMU 2014). From a conservation perspective, this precautionary
approach is preferred above trying to prevent permanent shifts like in different countries (e.g.
U.S. Marine Mammal Protection Act in the United States). However, it is not clear, if this
protection is sufficient. Conventional reporting of hearing impairment is limited to
measurements of hearing thresholds (reviewed in Finneran 2015) or hair cells (Morell et al.,
2017a). Nonetheless, a hearing impairment in humans can also be reflected in decreased
auditory evoked potentials, which remain undetected when measuring hearing thresholds and
are therefore referred to as a “hidden hearing loss” (Kujawa and Liberman, 2015; Lobarinas et
al., 2017). This can be a result of a permanently decreased number of synaptic connections
between the inner hair cells and auditory nerve fibres (Le Prell, 2019). Noise exposures,
however, which induce reversible thresholds shifts can lead to permanent cochlear
synaptopathy, which can have compounding effects for hearing in situations with elevated noise
levels (Kujawa and Liberman, 2015). These findings raise the concern, that a TTSonset could be
an insufficient indicator for protective regulations. Further research is needed to firstly
determine, if temporary threshold shifts can also lead to decreasing auditory evoked potentials
in harbour porpoises, and secondly to investigate its effect on acoustic perception.
A threshold for hearing impairment, either temporary or permanent, serves as a template for
noise regulation. However, the ecological consequences of such a TTS are not understood yet.
Further experiments are critically needed to determine effects of a TTS on the echolocation
performance of harbour porpoises. This information is critically needed to estimate
consequences on the energy budget or health of individuals and on the long run for the
population. As an experimental approach, harbour porpoises could be trained to perform
working tasks like object discrimination, catching fish or navigating blindfolded, while a TTS
is simulated. Such a simulation could be achieved by the development of an acoustic filter,
designed to damp certain frequencies, which could be attached to the animal around the jaw.

Conclusion
106
Animals could then be trained to perform certain tasks, which are evaluated in efficiency
compared to trials with normal hearing.
This study represents an important contribution for the implementation of protective directives
but also for the improvement of existing directives.

Summary
107
Summary
Tobias Schaffeld - Effect of anthropogenic underwater noise on harbour porpoise hearing in
areas of high ecological importance
Although aquatic noise pollution has been recognized as a key threat to biodiversity at an
unpredictable scale, the scientific discussion about effects on marine life has just evolved over
the last decades and is ongoing. Anthropogenic underwater noise occurs as an unintended by-
product, e.g. from shipping or construction work, but can also be on purpose, e.g. to deter
marine mammals or for acoustic localisation with sonars. Especially the evaluation of the
impacts on marine wildlife is in focus in modern science.
The species of interest in this thesis is the harbour porpoise (Phocoena phocoena), as the only
cetacean representative in German waters. Beyond that, these animals are particularly
vulnerable to interferences by noise due to their dependence from acoustically sensing their
environment. They demonstrate a very low resilience to disturbance, as they need to forage
almost continuously to maintain their high energy demands. The main goal of this thesis is to
assess the effect of noise disturbance from different sources as an antagonist, interfering with
acoustic perception of the habitat. This helps to shed light on gaps in noise regulation and raises
attention to the naive application of acoustic devices. This thesis comprises evaluations of three
different noise sources with potential effects on harbour porpoise hearing.
Firstly, to reveal the potential of multiple pile driving strikes to induce a temporary threshold
shift (TTS) under current German noise regulations, simulation approaches were conducted.
During the construction of an offshore wind farm underwater noise was recorded and sound
exposure levels (SEL) of single strikes were determined. This served for modelling the sound
propagation. Current noise regulations target single strikes up to a distance of 750 m for
preventing a TTS. However, the modelled sound field demonstrates that pile driving strikes at
distances up to 5.4 km contain sufficient energy, to induce a TTS after a repeated reception.
The “minimum deterrence distance” (MDD) was introduced as a novel protective unit, which
was determined between 2.3 and 4.7 km for four tested swim speed levels. The MDD marks
the minimum distance to the pile driving site, at which an animal must located, to be able to
escape a TTS by a flight. The simulation approach supports the current noise regulation in
Germany with a single strike SEL, a soft start and prior deterrence with seal scarers.

Summary
108
Secondly, a seal scarer was tested for its potential to induce a TTS in harbour porpoises. Hearing
tests with a harbour porpoise in human care prior to and after the exposure to artificial seal
scarer signals were conducted. The fatiguing signal consisted of a 0.5 s pure tone at 14 kHz
with four octave overtones with decreasing amplitudes. Hearing thresholds were measured by
monitoring the auditory evoked potential (AEP) response. Exposures exceeding a sound
exposure level of 142 dB re 1 µPa²s induced a significant TTS at a hearing frequency of 20 kHz
and at 28 kHz for SELs above 147 dB re 1 µPa²s. Assuming a theoretical transmission loss
between cylindrical and spherical spreading, a TTS could be induced at distances between
211 m and 5.9 km by a single exposure. Based on these findings a down-regulation of seal
scarer source levels was recommended.
Thirdly, an acoustic flowmeter (AFM) was evaluated in the port of Hamburg for its potential
to induce a TTS in harbour porpoises. The Hamburg Port Authority initiated this project after
several dead harbour porpoises stranded in this area of the Elbe River. These animals could be
forced in hazard areas by passing vessels, which occur in high numbers in this area. Therefore,
AFM signals were recorded underwater on board of a vessel for modelling the acoustic field.
The AFM transmits short signals with a peak frequency at 28 kHz in a highly directional beam
and at a pulse rate of 4.1 s from two positions at opposite sides of the river. A TTSonset for a
single exposure was derived from sound exposure experiments with an animal in human care
and for multiple exposures suggested from literature. A maximum SEL of 165 dB re 1 µPa²s
was determined. Single pulses exceeded the SEL-TTSonset of 138 dB re 1 µPa²s up to distances
of 72 m. The reception of multiple pulses was simulated for travelling harbour porpoises. The
received levels are highly variable due to the timing and high directionality of the AFM.
Although hazard zones are comparably small and signals are assumed to be heard at large
distances, a down-regulation of the AFM source level was suggested.
Within the three presented chapters of this thesis, an evolution of the understanding noise
effects could be shown. The introduction of underwater noise can be on purpose or unintended,
is already regulated or its regulation is currently in progress as well as possibly replaced by
alternatives. The contributions of this thesis demonstrate that improvements in noise mitigation
are required in all of these exemplified scenarios. This thesis complements studies assessing
the potential to induce a TTS. Far more important it will help to find the right starting point,

Summary
109
where adjustments in regulations are most effective. Moreover, the presented studies indicate
gaps in the understanding of TTS, which should be addressed in future research.

Zusammenfassung
110
Zusammenfassung
Tobias Schaffeld - Einfluss anthropogener Schalleinträge auf das Gehör von Schweinswalen
in Gebieten von hoher ökologischer Bedeutung
Die durch den Menschen verursachte Lärmverschmutzung in den Meeren stellt eine erhebliche
Bedrohung unvorhersehbaren Ausmaßes für die Biodiversität dar. Die wissenschaftliche
Auseinandersetzung mit möglichen Auswirkungen von Lärm auf marine Organismen findet
erst seit kurzem statt und ist noch lange nicht abgeschlossen. Anthropogener Unterwasserlärms
entsteht unter anderem als unbeabsichtigtes Nebenprodukt der Schiffverkehr oder durch
Bauarbeiten. Akustische Signale werden aber auch gezielt verwendet um Meeressäuger zu
vergrämen oder zur Navigation mit Sonaren.
Die in dieser Dissertation behandelte Zielart ist der Schweinswal (Phocoena phocoena) als
einziger Vertreter der Wale in deutschen Gewässern. Als Vertreter der Zahnwale erfasst der
Schweinswal seine Umgebung mittels Echoortung akustisch, sodass eine große Abhängigkeit
von einem intakten Gehör besteht. Schweinswale reagieren dementsprechend besonders
sensibel auf Störungen durch Lärm. Da diese fast ununterbrochen jagen müssen, um ihren
hohen Energiebedarf aufrechtzuerhalten, wird von einer sehr geringen Resilienz gegenüber
Störungen ausgegangen. Die vorliegende Studie bewertet unterschiedliche Lärmquellen
hinsichtlich ihrer Funktion als Antagonisten der akustischen Wahrnehmung für Schweinswale.
Das Ziel richtet sich hierbei auf das Auffinden von Lücken in der aktuellen Schallregulierung
und der Schaffung eines Bewusstseins bezüglich der naiven Anwendung akustischer Geräte.
Zu diesem Zweck werden die möglichen Auswirkungen von drei unterschiedlichen
Lärmquellen auf das Gehör von Schweinswalen behandelt.
Als erstes wurde durch einen Simulationsansatz ermittelt, ob das aktuelle Schallschutzkonzept
für Rammarbeiten in Offshore Windparks einen ausreichenden Schutz bietet, um eine
temporäre Hörschwellenverschiebung (TTS), das heißt reversible Verschlechterung des
Gehörs, durch mehrere Rammschläge zu verhindern. Hierfür wurde während des Baus eines
Offshore Windparks (OWP) Unterwasserschall aufgenommen und die Schallenergiepegel
(SEL) von Einzelschlägen ermittelt, mit denen die Schallausbreitung modelliert werden konnte.
Die aktuellen Lärmschutzbestimmungen zielen darauf ab, eine TTS durch Einzelschläge bis in
eine Entfernung von 750 m zu verhindern. Das modellierte Schallfeld zeigt jedoch, dass

Zusammenfassung
111
Rammschläge in Entfernungen von bis zu 5,4 km ausreichend Energie besitzen, um eine TTS
nach einer Mehrfachbeschallung auszulösen. Als neue Einheit zur Bewertung des
Gefährdungspotentials wurde die „minimale Vergrämungsdistanz“ (MVD) eingeführt, die für
vier getestete Schwimmgeschwindigkeitsstufen zwischen 2,3 und 4,7 km bestimmt wurde. Die
MVD markiert die Mindestentfernung zur Schallquelle, in der sich Schweinswale bei Beginn
der Rammarbeiten befinden müssen, um einer TTS durch Flucht zu entkommen. Der
Simulationsansatz unterstützt die derzeitig angewandte Lärmregulierung in Deutschland mit
einer begrenzten Maximalenergie für Einzelschläge, einem „soft start“ und einer vorherigen
Vergrämung mit Seal Scarern.
Im zweiten Teil wurde getestet, ob ein Seal Scarer das Potential besitzt, ein TTS bei
Schweinswalen zu induzieren. Hierfür wurden Hörtests mit einem Schweinswal in
menschlicher Fürsorge vor und nach der Exposition mit künstlichen Seal Scarer Signalen
durchgeführt. Das ermüdende Signal bestand aus einem 0,5 s langen 14 kHz Sinuston mit vier
Oktav-Obertönen mit abnehmenden Amplituden. Die Hörschwellen wurden anhand auditorisch
evozierter Potentiale (AEP) bestimmt. Schallexpositionen mit mehr als 142 dB re 1 µPa²s
induzierten eine signifikante TTS bei einer Hörfrequenz von 20 kHz und ab 147 dB re 1 µPa²s
bei 28 kHz. Unter der Annahme eines theoretischen Ausbreitungsverlusts zwischen
zylindrischer und kugelförmiger Ausbreitung könnte ein TTS in Entfernungen zwischen 211 m
und 5,9 km durch eine einzelne Exposition induziert werden. Basierend auf diesen Befunden
wurde eine Senkung des Quellschallpegels von Seal Scarern empfohlen.
Als Drittes wurde eine akustische Strömungsmessanlage im Hamburger Hafen hinsichtlich
ihres Potentials untersucht, eine TTS bei Schweinswalen hervorzurufen. Dieses Projekt wurde
von der Hamburg Port Authority initiiert, nachdem mehrere tote Schweinswale in diesem
Gebiet der Elbe strandeten. Der Schiffverkehr ist in diesem Bereich sehr hoch und
Schweinswale könnten durch vorbeifahrende Schiffe in den Gefahrenbereichen abgedrängt
werden. Die Unterwasserschallaufnahmen der akustischen Signale der Strömungsmessanlage
wurden an Bord eines Schiffes durchgeführt und das Schallfeld anschließend modelliert. Die
Strömungsmessanlage verwendet kurze Signale mit einer Frequenz von 28 kHz, die in einem
stark gerichteten Kegel und einer Pulsfrequenz von 4,1 s von zwei Positionen auf
gegenüberliegenden Seiten des Flusses emittiert werden. Ein TTSonset für eine
Einfachbeschallung wurde mittels Hörversuchen mit einem Tier in menschlicher Fürsorge

Zusammenfassung
112
ermittelt, während der Schwellenwert für eine Mehrfachbeschallung Literaturbasiert ist. Ein
maximaler SEL von 165 dB re 1 µPa²s wurde bestimmt. Einzelne Signale überschritten den
SEL-TTSonset von 138 dB re1 µPa²s bis in eine Entfernung von 72 m. Der Empfang mehrerer
Schallsignale wurde für Schweinswale simuliert, die an der Strömungsmessanlage entlang
wandern. Die empfangenen Pegel sind aufgrund des Timings und der hohen Richtwirkung der
Strömungsmessanlage sehr variabel. Obwohl die Gefahrenzonen vergleichsweise klein sind
und Schweinswale höchstwahrscheinlich die Signale in großen Entfernungen wahrnehmen
können, wurde empfohlen den Quellschallpegel zu senken.
In den drei vorgestellten Kapiteln dieser Arbeit konnte eine Entwicklung des Verständnisses
von Unterwasserlärmeffekten gezeigt werden. Der Eintrag von Unterwasserlärm kann
absichtlich oder unbeabsichtigt erfolgen, kann bereits reguliert sein oder es werden Versuche
zu deren Regulierung unternommen. Teilweise werden sogar Alternativen gesucht um
Lärmquellen zu ersetzen. Die Arbeiten in dieser Dissertation zeigen, dass in all den
vorgestellten Szenarien Verbesserungspotential in der Schallregulierung besteht. Diese Arbeit
erweitert den aktuellen Kenntnisstand in der Bewertung des Potenzials zur Auslösung einer
TTS, wird jedoch vor allem dazu beitragen, den richtigen Ansatzpunkt zu finden, an dem
Regulierungen am effektivsten sind. Die vorgestellten Studien zeigen Wissenslücken im
Verständnis von Hörschwellenverschiebungen bei Schweinswalen. Zukünftige
Forschungsvorhaben sollten sich diesen widmen.

Bibliography
113
Bibliography
Aguilar de Soto, N., Gkikopoulou, K., Hooker, S., Isojunno, S., Johnson, M., Miller, P., Tyack,
P., et al. (2016). “From physiology to policy: A review of physiological noise effects on
marine fauna with implications for mitigation,” Proc. Meet. Acoust., Dublin, 040008.
doi:10.1121/2.0000299
Aguilar de Soto, N., Visser, F., Tyack, P. L., Alcazar, J., Ruxton, G., Arranz, P., Madsen, P. T.,
et al. (2020). “Fear of Killer Whales Drives Extreme Synchrony in Deep Diving Beaked
Whales,” Sci. Rep., 10, s41598-019-55911–3. doi:10.1038/s41598-019-55911-3
Ainslie, M. A., Dahl, P. H., de Jong, C. A. F., and Laws, R. M. (2014). “Practical Spreading
Laws : the Snakes and Ladders,” UA2014 - 2nd Int. Conf. Exhib. Underw. Acoust. (isl.
Rhodes, Greece),.
Ainslie, M. A., and McColm, J. G. (1998). “A simplified formula for viscous and chemical
absorption in sea water,” J. Acoust. Soc. Am., 103, 1671–1672. doi:10.1121/1.421258
Akkaya Bas, A., Christiansen, F., Amaha Öztürk, A., Öztürk, B., and McIntosh, C. (2017). “The
effects of marine traffic on the behaviour of Black Sea harbour porpoises (Phocoena
phocoena relicta) within the Istanbul Strait, Turkey,” (S. Li, Ed.) PLoS One, 12, e0172970.
doi:10.1371/journal.pone.0172970
Amorim, M. C. P. (2006). “Communication in Fishes: Diversity of Sound Production in Fish,”
Commun. Fishes, 1, 71–105. Retrieved from
http://nova.wh.whoi.edu/palit/Amorim_2006_Communication in Fishes_Diversity of
sound production in fish
Amorim, M. C. P., Vasconcelos, R. O., and Fonseca, P. J. (2015). “Fish Sounds and Mate
Choice,” In F. Ladich (Ed.), Sound Commun. fishes, Springer, Vienna, Vol. 4, pp. 1–33.
doi:10.1007/978-3-7091-1846-7
Amundin, M. (2016). SAMBAH - Static Acoustic Monitoring of the Baltic Sea Harbour
Porpoise Kolmården, Sweden, 1–77 pages. Retrieved from www.sambah.org
Andreasen, H., Ross, S. D., Siebert, U., Andersen, N. G., Ronnenberg, K., and Gilles, A. (2017).
“Diet composition and food consumption rate of harbor porpoises (Phocoena phocoena)
in the western Baltic Sea,” Mar. Mammal Sci., 33, 1053–1079. doi:10.1111/mms.12421
Andrew, R. K., Howe, B. M., and Mercer, J. A. (2011). “Long-time trends in ship traffic noise
for four sites off the North American West Coast,” J. Acoust. Soc. Am., 129, 642–651.
doi:10.1121/1.3518770
Andrew, R. K., Howe, B. M., Mercer, J. A., and Dzieciuch, M. A. (2002). “Ocean ambient
sound: comparing the 1960s with the 1990s for a receiver off the California coast,” Acoust.
Res. Lett. Online, 3, 65–70. doi:10.1121/1.1461915
Aroyan, J. L. (2001). “Three-dimensional modeling of hearing in Delphinus delphis,” J. Acoust.
Soc. Am., 110, 3305. doi:10.1121/1.1401757
Aroyan, J. L., McDonald, M. A., Webb, S. C., Hildebrand, J. A., Clark, D., Laitman, J. T., and
Reidenberg, J. S. (2000). “Acoustic Models of Sound Production and Propagation,” Hear.
by whales dolphins, Springer, New York, NY, pp. 409–469. doi:10.1007/978-1-4612-
1150-1_10
ASCOBANS (2002). Recovery plan for Baltic harbour porpoises (Jastarnia Plan) Bonn, July
2002: ASCOBANS.
ASCOBANS (2012). Conservation of Harbour Porpoises and Adoption of a Conservation Plan
for the Western Baltic, the Belt Sea and Kattegat Brighton, UK 22-24 Oct 2012:
ASCOBANS, 46 pages.

Bibliography
114
Asselin, S., Hammill, M. O., and Barrette, C. (1993). “Underwater vocalizations of ice breeding
grey seals,” Can. J. Zool., 71, 2211–2219. doi:10.1139/z93-310
Au, W. W. L. (2014). “Dolphin biosonar target detection in noise: Wrap up of a past
experiment,” J. Acoust. Soc. Am., 136, 9–12. doi:10.1121/1.4883476
Au, W. W. L., and Moore, P. W. B. (1984). “Receiving beam patterns and directivity indices
of the Atlantic bottlenose dolphin Tursiops truncatus,” J. Acoust. Soc. Am., 75, 255.
doi:10.1121/1.390403
Babisch, W., Beule, B., Schust, M., Kersten, N., and Ising, H. (2005). “Traffic noise and risk
of myocardial infarction,” Epidemiology, 16, 33–40.
doi:10.1097/01.ede.0000147104.84424.24
Bailey, H., Senior, B., Simmons, D., Rusin, J., Picken, G., and Thompson, P. M. (2010).
“Assessing underwater noise levels during pile-driving at an offshore windfarm and its
potential effects on marine mammals.,” Mar. Pollut. Bull., 60, 888–897.
doi:10.1016/j.marpolbul.2010.01.003
Bain, D. E., and Williams, R. (2006). Long-range effects of airgun noise on marine mammals:
responses as a function of received sound level and distance Meet. Int. Whal. Comm. 26
May–6 June 2006, St. Kitts,SC58/E35,. Retrieved from
https://tethys.pnnl.gov/sites/default/files/publications/Bain-and-Williams-2006.pdf
Beineke, A., Siebert, U., McLachlan, M., Bruhn, R., Thron, K., Failing, K., Müller, G., et al.
(2005). “Investigations of the potential influence of environmental contaminants on the
thymus and spleen of harbor porpoises (Phocoena phocoena).,” Environ. Sci. Technol.,
39, 3933–3938. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/15984767
von Benda-Beckmann, A. M., Aarts, G., Sertlek, H. Ö., Lucke, K., Verboom, W. C., Kastelein,
R. A., Ketten, D. R., et al. (2015). “Assessing the Impact of Underwater Clearance of
Unexploded Ordnance on Harbour Porpoises (Phocoena phocoena) in the Southern North
Sea,” Aquat. Mamm., 41, 503–523. doi:10.1578/AM.41.4.2015.503
Benke, H., Siebert, U., Lick, R., Bandomir, B., and Weiss, R. (1998). “The current status of
harbour porpoises (Phocoena phocoena) in German waters,” Arch. Fish. Mar. Res., 46,
97–123.
Best, V., Gallun, F. J., Mason, C. R., Kidd, G., Shinn-Cunningham, B. G., and Shinn-
Cunningham, B. G. (2010). “The impact of noise and hearing loss on the processing of
simultaneous sentences.,” Ear Hear., 31, 213–20. doi:10.1097/AUD.0b013e3181c34ba6
Bjørge, A., and Tolley, K. A. (2009). “Harbor Porpoise: Phocoena phocoena,” In W. F. Perrin,
B. Würsig, and J. G. M. Thewissen (Eds.), Encycl. Mar. Mamm., 2nd editio., pp. 530–533.
doi:10.1016/B978-0-12-373553-9.00125-5
Bjørge, A., and Tolley, K. A. (2018). “Harbor Porpoise: Phocoena phocoena,” In W. F. Perrin,
B. Würsig, and J. G. M. Thewissen (Eds.), Encycl. Mar. Mamm., Elsevier, Amsterdam,
3nd ed., pp. 448–451. doi:10.1016/B978-0-12-804327-1.00144-8
BMU (2014). Concept for the Protection of Harbour Porpoises from Sound Exposures during
the Construction of Offshore Wind Farms in the German North Sea (Sound Protection
Concept) 33 pages.
BMWI (2012). “Research for an environmentally sound, reliable and affordable energy
supply,” 6th Energy Res. Program. Fed. Gov.,. Retrieved from
https://www.bmwi.de/Redaktion/EN/Publikationen/research-for-an-environmentally-
sound-reliable-and-affordable-energy-supply.pdf?__blob=publicationFile&v=3
Bodson, A., Miersch, L., and Dehnhardt, G. (2007). “Underwater localization of pure tones by
harbor seals (Phoca vitulina),” J. Acoust. Soc. Am., 122, 2263–2269.

Bibliography
115
doi:10.1121/1.2775424
Bouveroux, T., Kiszka, J. J., Heithaus, M. R., Jauniaux, T., and Pezeril, S. (2014). “Direct
evidence for gray seal (Halichoerus grypus) predation and scavenging on harbor porpoises
(Phocoena phocoena),” Mar. Mammal Sci., 30, 1542–1548. doi:10.1111/mms.12111
Bowers, M. T., Friedlaender, A. S., Janik, V. M., Nowacek, D. P., Quick, N. J., Southall, B. L.,
and Read, A. J. (2018). “Selective reactions to different killer whale call categories in two
delphinid species.,” J. Exp. Biol., 221, jeb162479. doi:10.1242/jeb.162479
Bradbury, J. W., and Vehrencamp, S. L. (2011). Principles of Animal Communication, Sinauer
Associates, Sunderland, MA, 2nd ed., 697 pages pages.
Brandt, M. J., Dragon, A. C., Diederichs, A., Bellmann, M. A., Wahl, V., Piper, W., Nabe-
Nielsen, J., et al. (2018). “Disturbance of harbour porpoises during construction of the first
seven offshore wind farms in Germany,” Mar. Ecol. Prog. Ser., 596, 213–232.
doi:10.3354/meps12560
Brandt, M. J., Höschle, C., Diederichs, A., Betke, K., Matuschek, R., and Nehls, G. (2013a).
“Seal scarers as a tool to deter harbour porpoises from offshore construction sites,” Mar.
Ecol. Prog. Ser., 475, 291–302. doi:10.3354/meps10100
Brandt, M. J., Höschle, C., Diederichs, A., Betke, K., Matuschek, R., Witte, S., and Nehls, G.
(2013b). “Far-reaching effects of a seal scarer on harbour porpoises , Phocoena phocoena,”
Aquat. Conserv. Mar. Freshw. Ecosyst., 23, 222–232. doi:10.1002/aqc.2311
Branstetter, B. K., and Mercado III, E. (2006). “Sound localization by cetaceans,” Int. J. Comp.
Psychol., 19, 26–61. Retrieved from http://escholarship.org/uc/item/28c0q755.pdf
van Bree, P. J. H., and Kristensen, I. (1974). “On the Intriguing Stranding of Four Cuvier’s
Beaked Whales, Ziphius Cavirostris G. Cuvier, 1823, on the Lesser Antillean Island of
Bonaire,” Bijdr. tot Dierkd., 44, 235–238. Retrieved from
https://brill.com/view/journals/btd/44/2/article-p235_3.xml
Brumm, H., and Slabbekoorn, H. (2005). “Acoustic Communication in Noise,” Adv. Study
Behav.,. doi:10.1016/S0065-3454(05)35004-2
Burros, N. B., and Myrberg, A. A. (1987). “Prey detection by means of passive listening in
bottlenose dolphins ( Tursiops truncatus ),” J. Acoust. Soc. Am., 82, S65–S65.
doi:10.1121/1.2024924
Carlén, I., Thomas, L., Carlström, J., Amundin, M., Teilmann, J., Tregenza, N., Tougaard, J.,
et al. (2018). “Basin-scale distribution of harbour porpoises in the Baltic Sea provides basis
for effective conservation actions,” Biol. Conserv., 226, 42–53.
doi:10.1016/j.biocon.2018.06.031
Chapman, N. R., and Price, A. (2011). “Low frequency deep ocean ambient noise trend in the
Northeast Pacific Ocean,” J. Acoust. Soc. Am., 129, EL161–EL165.
doi:10.1121/1.3567084
Charrier, I., Aubin, T., and Mathevon, N. (2010). “Mother–Calf vocal communication in
Atlantic walrus: a first field experimental study,” Anim. Cogn., 13, 471–482.
doi:10.1007/s10071-009-0298-9
Cholewiak, D., Clark, C., Ponirakis, D., Frankel, A., Hatch, L., Risch, D., Stanistreet, J., et al.
(2018). “Communicating amidst the noise: modeling the aggregate influence of ambient
and vessel noise on baleen whale communication space in a national marine sanctuary,”
Endanger. Species Res., 36, 59–75. doi:10.3354/esr00875
Clausen, K. T., Wahlberg, M., Beedholm, K., DeRuiter, S., and Madsen, P. T. (2010). “Click
communication in harbour porpoises Phocoena phocoena,” Bioacoustics, 20, 1–28.
doi:10.1080/09524622.2011.9753630

Bibliography
116
Conner, D. A. (1982). “Dialects versus geographic variation in mammalian vocalizations,”
Anim. Behav., 30, 297–298. doi:10.1016/S0003-3472(82)80269-8
Coram, A., Gordon, J., Thompson, D., and Northridge, S. (2014). Evaluating and assessing the
relative effectiveness of Acoustic Deterrent Devices and other non-lethal measures on
marine mammals St Andrews, Scotland.
Corless, R. M., Gonnet, G. H., Hare, D. E. G., Jeffrey, D. J., and Knuth, D. E. (1996). “On the
Lambert W function,” Adv. Comput. Math., 5, 329–359.
Cosentino, a. M. (2015). “First record of Norwegian killer whales attacking and feeding on a
harbour porpoise,” Mar. Biodivers. Rec., 8, e108. doi:10.1017/S1755267215000895
Cox, T. M., Ragen, T. J., Read, A. J., Vos, E., Baird, R. W., Balcomb, K., Barlow, J., et al.
(2006). “Understanding the impacts of anthropogenic sound on beaked whales,” J.
Cetacean Res. Manag., 7, 177–187. Retrieved from
http://cascadiaresearch.org/Robin/Coxetal2006.pdf
Cranford, T. W., Amundin, M., and Norris, K. S. (1996). “Functional morphology and
homology in the Odontoceti nasal complex implications for sound generation,” J.
Morphol., 228, 223–285. doi:10.1002/(SICI)1097-4687(199606)228:3<223::AID-
JMOR1>3.0.CO;2-3
Cranford, T. W., Krysl, P., and Amundin, M. (2010). “A New Acoustic Portal into the
Odontocete Ear and Vibrational Analysis of the Tympanoperiotic Complex,” (A. A. Farke,
Ed.) PLoS One, 5, e11927. doi:10.1371/journal.pone.0011927
Croll, D. A., Clark, C. W., Acevedo, A., Tershy, B., Flores, S., Gedamke, J., and Urban, J.
(2002). “Only male fin whales sing loud songs.,” Nature, 417, 809. doi:10.1038/417809a
Curé, C., Antunes, R., Alves, A. C., Visser, F., Kvadsheim, P. H., and Miller, P. J. O. (2013).
“Responses of male sperm whales (Physeter macrocephalus) to killer whale sounds:
implications for anti-predator strategies,” Sci. Rep., 3, 1579. doi:10.1038/srep01579
Curé, C., Isojunno, S., Visser, F., Wensveen, P., Sivle, L., Kvadsheim, P., Lam, F., et al. (2016).
“Biological significance of sperm whale responses to sonar: comparison with anti-predator
responses,” Endanger. Species Res., 31, 89–102. doi:10.3354/esr00748
CWSS (2017). “Introduction,” Wadden Sea Qual. Status Rep. 2017,.
Dähne, M., Gilles, A., Lucke, K., Peschko, V., Adler, S., Krügel, K., Sundermeyer, J., et al.
(2013). “Effects of pile-driving on harbour porpoises (Phocoena phocoena) at the first
offshore wind farm in Germany,” Environ. Res. Lett., 8, 025002. doi:10.1088/1748-
9326/8/2/025002
Dähne, M., Tougaard, J., Carstensen, J., Rose, A., and Nabe-Nielsen, J. (2017). “Bubble
curtains attenuate noise from offshore wind farm construction and reduce temporary
habitat loss for harbour porpoises,” Mar. Ecol. Prog. Ser., 580, 221–237.
doi:10.3354/meps12257
Dawson, S. M., Northridge, S., Waples, D., and Read, A. J. (2013). “To ping or not to ping: the
use of active acoustic devices in mitigating interactions between small cetaceans and
gillnet fisheries,” Endanger. Species Res., 19, 201–221. doi:10.3354/esr00464
Dechmann, D. K. N., Wikelski, M., van Noordwijk, H. J., Voigt, C. C., and Voigt-Heucke, S.
L. (2013). “Metabolic costs of bat echolocation in a non-foraging context support a role in
communication,” Front. Physiol., 4, 1–6. doi:10.3389/fphys.2013.00066
Dejours, P. (1987). “Water and air physical characteristics and their physiological
consequences,” In P. Dejours, L. Bolis, C. R. Taylor, and E. R. Weibel (Eds.), Comp.
Physiol. Life water L., Fidia Research Series, Springer, New York, pp. 3–11.
DeRuiter, S. L., Bahr, A., Blanchet, M.-A., Hansen, S. F., Kristensen, J. H., Madsen, P. T.,

Bibliography
117
Tyack, P. L., et al. (2009). “Acoustic behaviour of echolocating porpoises during prey
capture.,” J. Exp. Biol., 212, 3100–3107. doi:10.1242/jeb.030825
Don, M., Elberling, C., and Waring, M. (1984). “Objective detection of averaged auditory
brainstem responses,” Int. J. Audiol., 13, 219–228. doi:10.3109/14992028409042130
Dukas, R. (2004). “Causes and consequences of limited attention,” Brain. Behav. Evol., 63,
197–210. doi:10.1159/000076781
Duncan, A. J., Weilgart, L. S., Leaper, R., Jasny, M., and Livermore, S. (2017). “A modelling
comparison between received sound levels produced by a marine Vibroseis array and those
from an airgun array for some typical seismic survey scenarios,” Mar. Pollut. Bull., 119,
277–288. doi:10.1016/J.MARPOLBUL.2017.04.001
Dushaw, B. D., Worcester, P. F., Munk, W. H., Spindel, R. C., Mercer, J. A., Howe, B. M.,
Metzger, K., et al. (2009). “A decade of acoustic thermometry in the North Pacific Ocean,”
J. Geophys. Res., 114, C07021. doi:10.1029/2008JC005124
Dyndo, M., Wiśniewska, D. M., Rojano-Doñate, L., and Madsen, P. T. (2015). “Harbour
porpoises react to low levels of high frequency vessel noise,” Sci. Rep., 5, 11083.
doi:10.1038/srep11083
Elberling, C., and Don, M. (1984). “Quality Estimation of Averaged Auditory Brainstem
Responses,” Scand. Audiol., 13, 187–197. doi:10.3109/01050398409043059
Ellison, W. T., Southall, B. L., Clark, C. W., and Frankel, A. S. (2012). “A New Context-Based
Approach to Assess Marine Mammal Behavioral Responses to Anthropogenic Sounds,”
Conserv. Biol., 26, 21–28. doi:10.1111/j.1523-1739.2011.01803.x
Elmer, K.-H., Gattermann, J., Kuhn, C., and Bruns, B. (2012). “Mitigation of underwater piling
noise by air filled balloons and PE-foam elements as hydro sound dampers,” J. Acoust.
Soc. Am., 132, 2056–2056. doi:10.1121/1.4755571
Embling, C. B., Gillibrand, P. A., Gordon, J., Shrimpton, J., Stevick, P. T., and Hammond, P.
S. (2010). “Using habitat models to identify suitable sites for marine protected areas for
harbour porpoises (Phocoena phocoena),” Biol. Conserv., 143, 267–279.
doi:10.1016/j.biocon.2009.09.005
Erbe, C., Marley, S. A., Schoeman, R. P., Smith, J. N., Trigg, L. E., Embling, C. B., and
Radford, C. A. (2019). “The Effects of Ship Noise on Marine Mammals — A Review,”
Front. Mar. Sci., , doi: 10.3389/fmars.2019.00606. doi:10.3389/fmars.2019.00606
Erbe, C., Reichmuth, C., Cunningham, K., Lucke, K., and Dooling, R. (2016). “Communication
masking in marine mammals: A review and research strategy,” Mar. Pollut. Bull., , doi:
10.1016/j.marpolbul.2015.12.007. doi:10.1016/j.marpolbul.2015.12.007
European Union (1992). “Council Directive 92/43/EEC of 21 May 1992,” Off. J. Eur.
Communities, 35, 7–51.
European Union (2004). Council Regulation (EC) No 812/2004 of 26.4.2004 laying down
measures concerning incidental catches of cetaceans in fisheries and amending
Regulation (EC) No 88/89 Off. J. Eur. Union, 1–20 pages. Retrieved from https://eur-
lex.europa.eu/legal-content/DE/TXT/?uri=CELEX:32004R0812
FAO (2018). The State of World Fisheries and Aquacultures. Meeting the sustainable
development goals. Rome. Licence: CC BY-NC-SA 3.0 IGO Rome, 1–227 pages.
Fenton, B., Jensen, F. H., Kalko, E. K. V., and Tyack, P. L. (2014). “Sonar Signals of Bats and
Toothed Whales,” In A. Surlykke, P. E. Nachtigall, R. R. Fay, and A. N. Popper (Eds.),
Biosonar. Springer Handb. Audit. Res. vol. 51, Springer, New York, NY, pp. 11–59.
doi:10.1007/978-1-4614-9146-0_2
Fernández, A., Edwards, J. F., Rodríguez, F., Espinosa de los Monteros, A., Herráez, P., Castro,

Bibliography
118
P., Jaber, J. R., et al. (2005). “‘Gas and fat embolic syndrome’ involving a mass stranding
of beaked whales (family Ziphiidae) exposed to anthropogenic sonar signals.,” Vet.
Pathol., 42, 446–457. doi:10.1354/vp.42-4-446
Filadelfo, R., Mintz, J., Michlovich, E., D’Amico, A., Tyack, P. L., and Ketten, D. R. (2009).
“Correlating Military Sonar Use with Beaked Whale Mass Strandings: What Do the
Historical Data Show?,” Aquat. Mamm., 35, 435–444. doi:10.1578/AM.35.4.2009.435
Findlay, C. R., Ripple, H. D., Coomber, F., Froud, K., Harries, O., van Geel, N. C. F., Calderan,
S. V., et al. (2018). “Mapping widespread and increasing underwater noise pollution from
acoustic deterrent devices,” Mar. Pollut. Bull., 135, 1042–1050.
doi:10.1016/j.marpolbul.2018.08.042
Finneran, J. J. (2009). “Evoked response study tool: a portable, rugged system for single and
multiple auditory evoked potential measurements.,” J. Acoust. Soc. Am., 126, 491–500.
doi:10.1121/1.3148214
Finneran, J. J. (2015). “Noise-induced hearing loss in marine mammals : A review of temporary
threshold shift studies from 1996 to 2015,” J. Acoust. Soc. Am., 138, 1702–1726.
doi:10.1121/1.4927418
Finneran, J. J. (2018). “Conditioned attenuation of auditory brainstem responses in dolphins
warned of an intense noise exposure: Temporal and spectral patterns,” J. Acoust. Soc. Am.,
143, 795–810. doi:10.1121/1.5022784
Finneran, J. J., Carder, D. A., Schlundt, C. E., and Dear, R. L. (2010a). “Growth and recovery
of temporary threshold shift at 3 kHz in bottlenose dolphins: experimental data and
mathematical models.,” J. Acoust. Soc. Am., 127, 3256–3266. doi:10.1121/1.3372710
Finneran, J. J., Carder, D. A., Schlundt, C. E., and Dear, R. L. (2010b). “Temporary threshold
shift in a bottlenose dolphin (Tursiops truncatus) exposed to intermittent tones.,” J. Acoust.
Soc. Am., 127, 3267–3272. doi:10.1121/1.3377052
Finneran, J. J., Carder, D. A., Schlundt, C. E., and Ridgway, S. H. (2005). “Temporary threshold
shift in bottlenose dolphins (Tursiops truncatus) exposed to mid-frequency tones,” J.
Acoust. Soc. Am., 118, 2696–2705. doi:10.1121/1.2032087
Finneran, J. J., Houser, D. S., Blasko, D., Hicks, C., Hudson, J., and Osborn, M. (2008).
“Estimating bottlenose dolphin (Tursiops truncatus) hearing thresholds from single and
multiple simultaneous auditory evoked potentials,” J. Acoust. Soc. Am., 123, 542–551.
doi:10.1121/1.2812595
Fischer, M., and Schuster, M. (2017). “Vermessung hydroakustischer Quellen im Hafen
Hamburg,” DW-ShipConsult GmbH,.
Ford, J. K. B. (1989). “Acoustic behaviour of resident killer whales (Orcinus orca) off
Vancouver Island, British Columbia,” Can. J. Zool., 67, 727–745.
Foskolos, I., Aguilar de Soto, N., Madsen, P. T., and Johnson, M. (2019). “Deep-diving pilot
whales make cheap, but powerful, echolocation clicks with 50 µL of air,” Sci. Rep., 9,
15720. doi:10.1038/s41598-019-51619-6
Francis, C. D., and Barber, J. R. (2013). “A framework for understanding noise impacts on
wildlife: an urgent conservation priority,” Front. Ecol. Environ., 11, 305–313.
doi:10.1890/120183
Frouin-Mouy, H., Mouy, X., Martin, B., and Hannay, D. (2016). “Underwater acoustic behavior
of bearded seals ( Erignathus barbatus ) in the northeastern Chukchi Sea, 2007-2010,” Mar.
Mammal Sci., 32, 141–160. doi:10.1111/mms.12246
Fyhri, A., and Aasvang, G. M. (2010). “Noise, sleep and poor health: Modeling the relationship
between road traffic noise and cardiovascular problems,” Sci. Total Environ., 408, 4935–

Bibliography
119
4942. doi:10.1016/j.scitotenv.2010.06.057
Galatius, A., Kinze, C. C., and Teilmann, J. (2012). “Population structure of harbour porpoises
in the Baltic region: evidence of separation based on geometric morphometric
comparisons,” J. Mar. Biol. Assoc. UK, 92, 1669–1676.
doi:10.1017/S0025315412000513
Gallagher, C. A., Stern, S. J., and Hines, E. (2018). “The metabolic cost of swimming and
reproduction in harbor porpoises ( Phocoena phocoena ) as predicted by a bioenergetic
model,” Mar. Mammal Sci., 34, 875–900. doi:10.1111/mms.12487
Gannon, D. P., Barros, N. B., Nowacek, D. P., Read, A. J., Waples, D. M., and Wells, R. S.
(2005). “Prey detection by bottlenose dolphins, Tursiops truncatus: An experimental test
of the passive listening hypothesis,” Anim. Behav., 69, 709–720.
doi:10.1016/j.anbehav.2004.06.020
Gaskin, D. E., Arnold, P. W., and Blair, B. A. (1974). “Phocoena phocoena,” Mamm. Species,
Retrieved from https://www.jstor.org/stable/3504041. Retrieved from
https://www.jstor.org/stable/3504041
Gerdes, F., and Görler, M. (2016). “Untersuchungen der Variabilität der
Unterwasserschallausbreitung in der Nordsee (Investigations on variability of underwater
sound propagation in the North Sea),” Fortschritte der Akust., 143–146. Retrieved from
https://www.fino3.de/files/forschung/WTD/2016_ConferencePaper_DAGA2016_Gerdes
Goerler.pdf
Gerdes, F., Wildemann, M., Müller, A., and Zerbs, C. (2016). “Measurements of pile driving
noise at the research platform FINO3,” INTER-NOISE NOISE-CON Congr. Conf. Proc.,
Institute of Noise Control Engineering, 2844–2851.
Gilles, A., Adler, S., Kaschner, K., Scheidat, M., and Siebert, U. (2011). “Modelling harbour
porpoise seasonal density as a function of the German Bight environment: Implications
for management,” Endanger. Species Res., 14, 157–169. doi:10.3354/esr00344
Gilles, A., Scheidat, M., and Siebert, U. (2009). “Seasonal distribution of harbour porpoises
and possible interference of offshore wind farms in the German North Sea,” Mar. Ecol.
Prog. Ser., 383, 295–307. doi:10.3354/meps08020
Gilles, A., Viquerat, S., Becker, E. A., Forney, K. A., Geelhoed, S. C. V, Haelters, J., Nabe-
Nielsen, J., et al. (2016). “Seasonal habitat-based density models for a marine top predator,
the harbor porpoise, in a dynamic environment,” Ecosphere, 7, e01367.
doi:10.1002/ecs2.1367
Götz, T., and Janik, V. (2013). “Acoustic deterrent devices to prevent pinniped depredation:
efficiency, conservation concerns and possible solutions,” Mar. Ecol. Prog. Ser., 492, 285–
302. doi:10.3354/meps10482
Graham, I. M., Merchant, N. D., Farcas, A., Barton, T. R., Cheney, B., Bono, S., and Thompson,
P. M. (2019). “Harbour porpoise responses to pile-driving diminish over time,” R. Soc.
Open Sci., 6, 190335. doi:10.1098/rsos.190335
Hammond, P. S., Lacey, C., Gilles, A., Viquerat, S., Börjesson, P., Herr, H., Macleod, K., et al.
(2017). Estimates of cetacean abundance in European Atlantic waters in summer 2016
from the SCANS-III aerial and shipboard surveys 1–39 pages. Retrieved from
https://synergy.st-andrews.ac.uk/scans3/files/2017/04/SCANS-III-design-based-
estimates-2017-04-28-final.pdf
Hammond, P. S., Macleod, K., Berggren, P., Borchers, D. L., Burt, L., Cañadas, A., Desportes,
G., et al. (2013). “Cetacean abundance and distribution in European Atlantic shelf waters
to inform conservation and management,” Biol. Conserv., 164, 107–122.

Bibliography
120
doi:10.1016/j.biocon.2013.04.010
Hansell, A. L., Blangiardo, M., Fortunato, L., Floud, S., De Hoogh, K., Fecht, D., Ghosh, R.
E., et al. (2013). “Aircraft noise and cardiovascular disease near Heathrow airport in
London: Small area study,” BMJ, 347, f5432. doi:10.1136/bmj.f5432
Häpke, L. (1880). “Fische und Fischerei im Wesergebiete,” Abhandlungen des
Naturwissenschaftlichen Vereins zu Bremen, 6, 577–616. Retrieved from
http://www.nwv-bremen.de/publik/abhandlungen-
nwv.html?option=com_biblio&view=biblio&Itemid=109&bandthe=06%2F3&jahrthe=1
880&suchaut=&suchtit=&fachthe=&gebithe=&geogthe=&spamsch=afrika&suchen=suc
hen
Heers, T., van Neer, A., Becker, A., Gross, S., Hansen, K. A., Siebert, U., and Abdulmawjood,
A. (2018). “Loop-mediated isothermal amplification (LAMP) as a confirmatory and rapid
DNA detection method for grey seal (Halichoerus grypus) predation on harbour porpoises
(Phocoena phocoena),” J. Sea Res., 140, 32–39. doi:10.1016/j.seares.2018.07.008
Hemilä, S., Nummela, S., and Reuter, T. (2010). “Anatomy and physics of the exceptional
sensitivity of dolphin hearing (Odontoceti: Cetacea).,” J. Comp. Physiol. A, 196, 165–179.
doi:10.1007/s00359-010-0504-x
Herman, L. M. (2017). “The multiple functions of male song within the humpback whale (
Megaptera novaeangliae ) mating system: review, evaluation, and synthesis,” Biol. Rev.,
92, 1795–1818. doi:10.1111/brv.12309
Hildebrand, J. A. (2009). “Anthropogenic and natural sources of ambient noise in the ocean,”
Mar. Ecol. Prog. Ser., 395, 5–20. doi:10.3354/meps08353
van der Hoop, J. M., Fahlman, A., Hurst, T., Rocho-Levine, J., Shorter, K. a., Petrov, V., and
Moore, M. J. (2014). “Bottlenose dolphins modify behavior to reduce metabolic effect of
tag attachment,” J. Exp. Biol., 217, 4229–4236. doi:10.1242/jeb.108225
Houser, D. S., Yost, W., Burkard, R., Finneran, J. J., Reichmuth, C., and Mulsow, J. (2017). “A
review of the history, development and application of auditory weighting functions in
humans and marine mammals,” J. Acoust. Soc. Am., 141, 1371–1413.
doi:10.1121/1.4976086
Huber, S., Ahrens, L., Bårdsen, B. J., Siebert, U., Bustnes, J. O., Víkingsson, G. A., Ebinghaus,
R., et al. (2012). “Temporal trends and spatial differences of perfluoroalkylated substances
in livers of harbor porpoise (Phocoena phocoena) populations from Northern Europe,
1991-2008,” Sci. Total Environ., 419, 216–224. doi:10.1016/j.scitotenv.2011.12.050
Huggenberger, S., Benke, H., and Kinze, C. C. (2002). “Geographical variation in harbour
porpoise (Phocoena phocoena) skulls: support for a separate non-migratory population in
the Baltic proper,” Ophelia, 56, 1–12. Retrieved from
http://cat.inist.fr/?aModele=afficheN&cpsidt=13752446
Huggenberger, S., Rauschmann, M. A., Vogl, T. J., and Oelschläger, H. H. A. (2009).
“Functional morphology of the nasal complex in the harbor porpoise (Phocoena phocoena
L.),” Anat. Rec., 292, 902–920. doi:10.1002/ar.20854
Insley, S. J., Phillips, A. V, and Charrier, I. (2003). “A review of social recognition in
pinnipeds,” Aquat. Mamm., 29, 181–201. doi:10.1578/016754203101024149
ISO (2016). “Acoustics - Description, measurement and assessment of environmental noise.
Part 1: Basic quantities and assessment procedures (ISO 1996-1:2016).,” Retrieved from
https://www.iso.org/obp/ui/#iso:std:iso:1996:-1:ed-3:v1:en
ISO 18406 (2017). Underwater acoustics — Measurement of radiated underwater sound from
percussive pile driving,. Retrieved from

Bibliography
121
https://www.iso.org/obp/ui/#iso:std:iso:18406:ed-1:v1:en
Isojunno, S., Curé, C., Kvadsheim, P. H., Lam, F. P. A., Tyack, P. L., Wensveen, P. J., and
Miller, P. J. O. M. (2016). “Sperm whales reduce foraging effort during exposure to 1-2
kH z sonar and killer whale sounds,” Ecol. Appl., 26, 77–93. doi:10.1890/15-
0040.1/suppinfo
Jacobson, E. K., Forney, K. a., and Harvey, J. T. (2015). “Acoustic evidence that harbor
porpoises ( Phocoena phocoena ) avoid bottlenose dolphins ( Tursiops truncatus ),” Mar.
Mammal Sci., 31, 386–397. doi:10.1111/mms.12154
Jakobsen, L., Ratcliffe, J. M., and Surlykke, A. (2013). “Convergent acoustic field of view in
echolocating bats,” Nature, 493, 93–96. doi:10.1038/nature11664
Janik, V. M., and Sayigh, L. S. (2013). “Communication in bottlenose dolphins: 50 years of
signature whistle research.,” J. Comp. Physiol. A, 199, 479–489. doi:10.1007/s00359-013-
0817-7
Jauniaux, T., Garigliany, M.-M., Loos, P., Bourgain, J.-L., Bouveroux, T., Coignoul, F.,
Haelters, J., et al. (2014). “Bite injuries of grey seals (Halichoerus grypus) on harbour
porpoises (Phocoena phocoena),” PLoS One, 9, e108993.
doi:10.1371/journal.pone.0108993
Jensen, F. H., Johnson, M., Ladegaard, M., Wisniewska, D. M., and Madsen, P. T. (2018).
“Narrow Acoustic Field of View Drives Frequency Scaling in Toothed Whale Biosonar,”
Curr. Biol., , doi: 10.1016/j.cub.2018.10.037. doi:10.1016/j.cub.2018.10.037
Jepson, P. D., Arbelo, M., Deaville, R., Patterson, I. A. P., Castro, P., Baker, J. R., Degollada,
E., et al. (2003). “Gas-bubble lesions in stranded cetaceans,” Nature, 425, 575–576.
Jepson, P. D., Deaville, R., Barber, J. L., Aguilar, À., Borrell, A., Murphy, S., Barry, J., et al.
(2016). “PCB pollution continues to impact populations of orcas and other dolphins in
European waters,” Sci. Rep., 6, 18573. doi:10.1038/srep18573
Johnson, M. P., Aguilar de Soto, N., and Madsen, P. T. (2009). “Studying the behaviour and
sensory ecology of marine mammals using acoustic recording tags: a review,” Mar. Ecol.
Prog. Ser., 395, 55–73. doi:10.3354/meps08255
Kastak, D., and Schusterman, R. J. (1998). “Low-frequency amphibious hearing in pinnipeds:
methods, measurements, noise, and ecology,” J. Acoust. Soc. Am., 103, 2216–2228.
Kastelein, R. A., Au, W. W. L., and Haan, D. De (2000). “Detection distances of bottom-set
gillnets by harbour porpoises (Phocoena phocoena) and bottlenose dolphins (Tursiops
truncatus),” Mar. Environ. Res., 49, 359–375.
Kastelein, R. A., Bunskoek, P., Hagedoorn, M., Au, W. W. L., and de Haan, D. (2002).
“Audiogram of a harbor porpoise (Phocoena phocoena) measured with narrow-band
frequency-modulated signals,” J. Acoust. Soc. Am., 112, 334–344.
doi:10.1121/1.1480835
Kastelein, R. A., Gransier, R., Marijt, M. A. T., and Hoek, L. (2015a). “Hearing frequency
thresholds of harbor porpoises (Phocoena phocoena) temporarily affected by played back
offshore pile driving sounds,” J. Acoust. Soc. Am., 137, 556–564. doi:10.1121/1.4906261
Kastelein, R. a., Gransier, R., Schop, J., and Hoek, L. (2015b). “Effects of exposure to
intermittent and continuous 6–7 kHz sonar sweeps on harbor porpoise (Phocoena
phocoena) hearing,” J. Acoust. Soc. Am., 137, 1623–1633. doi:10.1121/1.4916590
Kastelein, R. A., Haan, D. De, and Verboom, W. C. (2007). “The influence of signal parameters
on the sound source localization ability of a harbor porpoise (Phocoena phocoena),” J.
Acoust. Soc. Am., 122, 1238–1248. doi:10.1121/1.2747202
Kastelein, R. A., van Heerden, D., Gransier, R., and Hoek, L. (2013). “Behavioral responses of

Bibliography
122
a harbor porpoise (Phocoena phocoena) to playbacks of broadband pile driving sounds.,”
Mar. Environ. Res., 92, 206–214. doi:10.1016/j.marenvres.2013.09.020
Kastelein, R. A., Helder-Hoek, L., Covi, J., and Gransier, R. (2016). “Pile driving playback
sounds and temporary threshold shift in harbor porpoises (Phocoena phocoena): Effect of
exposure duration,” J. Acoust. Soc. Am., 139, 2842–2851. doi:10.1121/1.4948571
Kastelein, R. A., Helder-Hoek, L., and Gransier, R. (2019a). “Frequency of greatest temporary
hearing threshold shift in harbor seals (Phoca vitulina) depends on fatiguing sound level,”
J. Acoust. Soc. Am., 145, 1353–1362. doi:10.1121/1.5092608
Kastelein, R. a., Helder-Hoek, L., Gransier, R., Terhune, J. M., Jennings, N., and de Jong, C. a.
F. (2015c). “Hearing thresholds of harbor seals (Phoca vitulina) for playbacks of seal
scarer signals, and effects of the signals on behavior,” Hydrobiologia, 756, 75–88.
doi:10.1007/s10750-014-2152-6
Kastelein, R. A., Helder-Hoek, L., van Kester, R., Huisman, R., and Gransier, R. (2019b).
“Temporary Hearing Threshold Shift in Harbor Porpoises (Phocoena phocoena) Due to
One-Sixth Octave Noise Band at 16 kHz,” Aquat. Mamm., 45, 280–292.
doi:10.1578/AM.45.3.2019.280
Kastelein, R. A., Helder-Hoek, L., and Terhune, J. M. (2018a). “Hearing thresholds, for
underwater sounds, of harbor seals (Phoca vitulina) at the water surface,” J. Acoust. Soc.
Am., 143, 2554–2563. doi:10.1121/1.5034173
Kastelein, R. A., Helder-Hoek, L., Van de Voorde, S., von Benda-Beckmann, A. M., Lam, F.-
P. A., Jansen, E., de Jong, C. A. F., et al. (2017). “Temporary hearing threshold shift in a
harbor porpoise (Phocoena phocoena) after exposure to multiple airgun sounds,” J.
Acoust. Soc. Am., 142, 2430–2442. doi:10.1121/1.5007720
Kastelein, R. a., Hoek, L., Gransier, R., de Jong, C. a. F., Terhune, J. M., and Jennings, N.
(2014a). “Hearing thresholds of a harbor porpoise (Phocoena phocoena) for playbacks of
seal scarer signals, and effects of the signals on behavior,” Hydrobiologia, 756, 89–103.
doi:10.1007/s10750-014-2035-x
Kastelein, R. A., Hoek, L., Gransier, R., Rambags, M., and Claeys, N. (2014b). “Effect of level,
duration, and inter-pulse interval of 1-2 kHz sonar signal exposures on harbor porpoise
hearing.,” J. Acoust. Soc. Am., 136, 412–422. doi:10.1121/1.4883596
Kastelein, R. A., Hoek, L., de Jong, C. A. F., and Wensveen, P. J. (2010). “The effect of signal
duration on the underwater detection thresholds of a harbor porpoise (Phocoena phocoena)
for single frequency-modulated tonal signals between 0.25 and 160 kHz.,” J. Acoust. Soc.
Am., 128, 3211–3222. doi:10.1121/1.3493435
Kastelein, R. A., Janssen, M., Verboom, W. C., and de Haan, D. (2005). “Receiving beam
patterns in the horizontal plane of a harbor porpoise (Phocoena phocoena),” J. Acoust.
Soc. Am., 118, 1172–1179. doi:10.1121/1.1945565
Kastelein, R. A., Schop, J., Hoek, L., and Covi, J. (2015d). “Hearing thresholds of a harbor
porpoise (Phocoena phocoena) for narrow-band sweeps,” J. Acoust. Soc. Am., 138, 2508–
2512. doi:10.1121/1.4932024
Kastelein, R. A., Van de Voorde, S., and Jennings, N. (2018b). “Swimming Speed of a Harbor
Porpoise (Phocoena phocoena) During Playbacks of Offshore Pile Driving Sounds Title,”
Aquat. Mamm., Retrieved from https://doi.org/10.1578/AM.44.1.2018.92. Retrieved from
https://doi.org/10.1578/AM.44.1.2018.92
Kastelein, R. A., Wensveen, P. J., Hoek, L., and Terhune, J. M. (2009a). “Underwater hearing
sensitivity of harbor seals (Phoca vitulina) for narrow noise bands between 0.2 and 80
kHz.,” J. Acoust. Soc. Am., 126, 476–483. doi:10.1121/1.3132522

Bibliography
123
Kastelein, R. A., Wensveen, P. J., Hoek, L., Verboom, W. C., and Terhune, J. M. (2009b).
“Underwater detection of tonal signals between 0 . 125 and 100 kHz by harbor seals
(Phoca vitulina),” J. Acoust. Soc. Am., 125, 1222–1229. doi:10.1121/1.3050283
Kastelein, R. a, Schop, J., Gransier, R., and Hoek, L. (2014c). “Frequency of greatest temporary
hearing threshold shift in harbor porpoises (Phocoena phocoena) depends on the noise
level,” J. Acoust. Soc. Am., 136, 1410–1418. doi:10.1121/1.4892794
Kesselring, T., Viquerat, S., Brehm, R., and Siebert, U. (2017). “Coming of age : - Do female
harbour porpoises (Phocoena phocoena) from the North Sea and Baltic Sea have sufficient
time to reproduce in a human influenced environment ?,” PLoS One, 12, e0186951.
doi:10.1371/ journal.pone.0186951
Kesselring, T., Viquerat, S., IJsseldijk, L. L., Langeheine, M., Wohlsein, P., Gröne, A.,
Bergmann, M., et al. (2019). “Testicular morphology and spermatogenesis in harbour
porpoises (Phocoena phocoena),” Theriogenology, 126, 177–186.
doi:10.1016/j.theriogenology.2018.11.031
Ketten, D. R. (1997). “Structure and function in whale ears,” Bioacoustics, 8, 103–135.
King, S. L., and Janik, V. M. (2015). “Come dine with me: food-associated social signalling in
wild bottlenose dolphins (Tursiops truncatus),” Anim. Cogn., , doi: 10.1007/s10071-015-
0851-7. doi:10.1007/s10071-015-0851-7
King, S. L., Schick, R. S., Donovan, C., Booth, C. G., Burgman, M., Thomas, L., and Harwood,
J. (2015). “An interim framework for assessing the population consequences of
disturbance,” (C. Kurle, Ed.) Methods Ecol. Evol., 6, 1150–1158. doi:10.1111/2041-
210X.12411
Kleinertz, S., Hermosilla, C., Ziltener, A., Kreicker, S., Hirzmann, J., Abdel-Ghaffar, F., and
Taubert, A. (2014). “Gastrointestinal parasites of free-living Indo-Pacific bottlenose
dolphins (Tursiops aduncus) in the Northern Red Sea, Egypt,” Parasitol. Res., 113, 1405–
1415. doi:10.1007/s00436-014-3781-4
Knoop, V., and Müller, K. (2009). “Gene und Stammbäume Ein Handbuch zur molekularen
Phylogenetik,” , doi: 10.1007/978-3-8274-2230-9. doi:10.1007/978-3-8274-2230-9
Koblitz, J. C., Wahlberg, M., Stilz, P., Madsen, P. T., Beedholm, K., and Schnitzler, H.-U.
(2012). “Asymmetry and dynamics of a narrow sonar beam in an echolocating harbor
porpoise.,” J. Acoust. Soc. Am., 131, 2315–2324. doi:10.1121/1.3683254
Koch, M., and Schnitzler, H.-U. (1997). “The acoustic startle response in rats—circuits
mediating evocation, inhibition and potentiation,” Behav. Brain Res., 89, 35–49.
Kölmel, R. (1998). “Wale aus der Elbe, 340 Jahre Funde und Fang,” Schriftreihen Natureum
Niederelbe, 2, 48–49.
Koopman, H. N. (2018). “Function and evolution of specialized endogenous lipids in toothed
whales.,” J. Exp. Biol., , doi: 10.1242/jeb.161471. doi:10.1242/jeb.161471
Korpinen, S., and Braeger, S. (2013). Number of drowned mammals and waterbirds in fishing
gear HELCOM Core Indic. Biodivers., 1–22 pages. Retrieved from
http://www.helcom.fi/Core Indicators/HELCOM-CoreIndicator-
Number_of_drowned_mammals_and_waterbirds_in_fishing_gear.pdf
Kujawa, S. G., and Liberman, M. C. (2015). “Synaptopathy in the noise-exposed and aging
cochlea: Primary neural degeneration in acquired sensorineural hearing loss,” Hear. Res.,
330, 191–199. doi:10.1016/j.heares.2015.02.009
Kunc, H. P., McLaughlin, K. E., and Schmidt, R. (2016). “Aquatic noise pollution: Implications
for individuals, populations, and ecosystems,” Proc. R. Soc. B Biol. Sci., 283, 20160839.
doi:10.1098/rspb.2016.0839

Bibliography
124
Ladich, F. (2019). “Ecology of sound communication in fishes,” Fish Fish., 20, 552–563.
doi:10.1111/faf.12368
Ladich, F., and Fay, R. R. (2013). Auditory evoked potential audiometry in fish, Rev. Fish Biol.
Fish., Vol. 23, 317–364 pages. doi:10.1007/s11160-012-9297-z
Ladich, F., and Myrberg, A. A. (2006). “Agonistic Behavior and Acoustic Communication,” In
F. Ladich, S. P. Collin, P. Moller, and B. G. Kapoor (Eds.), Commun. Fishes, pp. 122–
148. Retrieved from https://homepage.univie.ac.at/friedrich.ladich/Ladich & Myrberg
2006.pdf
Ladich, F., and Winkler, H. (2017). “Acoustic communication in terrestrial and aquatic
vertebrates.,” J. Exp. Biol., 220, 2306–2317. doi:10.1242/jeb.132944
Lammers, M. O., and Au, W. W. L. (2003). “Directionality in the whistles of Hawaiian Spinner
Dolphins (Stenella Longirostris): A signal feature to cue direction of movement,” Mar.
Mammal Sci., 19, 249–264. doi:10.1111/j.1748-7692.2003.tb01107.x
Lehnert, K., Raga, J., and Siebert, U. (2005). “Macroparasites in stranded and bycaught harbour
porpoises from German and Norwegian waters,” Dis. Aquat. Organ., 64, 265–269.
doi:10.3354/dao064265
Leopold, M. F., Begeman, L., van Bleijswijk, J. D. L., IJsseldijk, L. L., Witte, H. J., and Gröne,
A. (2015). “Exposing the grey seal as a major predator of harbour porpoises.,” Proc. R.
Soc. London B Biol. Sci., 282, 20142429. doi:10.1098/rspb.2014.2429
Lepper, P. a, Goodson, a D., and Black, K. D. (2004). “Source levels and spectra emitted by
three commercial aquaculture anti-predation devices,” Proc. 7th Eur. Conf. Underw.
Acoust. ECUA 5-8 July 2004, 7, 6.
Leunissen, E. M., and Dawson, S. M. (2018). “Underwater noise levels of pile-driving in a New
Zealand harbour, and the potential impacts on endangered Hector’s dolphins,” Mar. Pollut.
Bull., 135, 195–204. doi:10.1016/j.marpolbul.2018.07.024
Lindseth, A., and Lobel, P. (2018). “Underwater Soundscape Monitoring and Fish
Bioacoustics: A Review,” Fishes, 3, 36. doi:10.3390/fishes3030036
Linnenschmidt, M., Teilmann, J., Akamatsu, T., Dietz, R., and Miller, L. A. (2013). “Biosonar,
dive, and foraging activity of satellite tracked harbor porpoises ( Phocoena phocoena ),”
Mar. Mammal Sci., 29, E77–E97. doi:10.1111/j.1748-7692.2012.00592.x
Lippert, T., Ainslie, M. A., and von Estorff, O. (2018). “Pile driving acoustics made simple:
Damped cylindrical spreading model,” J. Acoust. Soc. Am., 143, 310–317.
doi:10.1121/1.5011158
Lobarinas, E., Spankovich, C., and Le Prell, C. G. (2017). “Evidence of ‘hidden hearing loss’
following noise exposures that produce robust TTS and ABR wave-I amplitude
reductions,” Hear. Res., 349, 155–163. doi:10.1016/J.HEARES.2016.12.009
Lucke, K., Lepper, P. A., Blanchet, M.-A., and Siebert, U. (2011). “The use of an air bubble
curtain to reduce the received sound levels for harbor porpoises (Phocoena phocoena),” J.
Acoust. Soc. Am., 130, 3406–3412. doi:10.1121/1.3626123
Lucke, K., Siebert, U., Lepper, P. A., and Blanchet, M.-A. (2009). “Temporary shift in masked
hearing thresholds in a harbor porpoise (Phocoena phocoena) after exposure to seismic
airgun stimuli.,” J. Acoust. Soc. Am., 125, 4060–4070. doi:10.1121/1.3117443
Madsen, P. T. (2005). “Marine mammals and noise: Problems with root mean square sound
pressure levels for transients,” J. Acoust. Soc. Am., 117, 3952. doi:10.1121/1.1921508
Madsen, P. T., Carder, D. A., Au, W. W. L., Nachtigall, P. E., Møhl, B., and Ridgway, S. H.
(2003). “Sound production in neonate sperm whales (L),” J. Acoust. Soc. Am., 113, 2988–
2991. doi:10.1121/1.1572137

Bibliography
125
Madsen, P. T., Payne, R., Kristiansen, N. U., Wahlberg, M., Kerr, I., and Møhl, B. (2002).
“Sperm whale sound production studied with ultrasound time/depth-recording tags.,” J.
Exp. Biol., 205, 1899–1906. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/12077166
Madsen, P. T., Wahlberg, M., Tougaard, J., Lucke, K., and Tyack, P. (2006). “Wind turbine
underwater noise and marine mammals: implications of current knowledge and data
needs,” Mar. Ecol. Prog. Ser., 309, 279–295. doi:10.3354/meps309279
Mahfouz, C., Henry, F., Courcot, L., Pezeril, S., Bouveroux, T., Dabin, W., Jauniaux, T., et al.
(2014). “Harbour porpoises (Phocoena phocoena) stranded along the southern North Sea:
An assessment through metallic contamination,” Environ. Res., 133, 266–273.
doi:10.1016/j.envres.2014.06.006
Mann, D., Hill-Cook, M., Manire, C., Greenhow, D., Montie, E., Powell, J., Wells, R., et al.
(2010). “Hearing loss in stranded odontocete dolphins and whales.,” PLoS One, 5, e13824.
doi:10.1371/journal.pone.0013824
Marsh, H. W., and Schulkin, M. (1962). “Shallow-water transmission,” J. Acoust. Soc. Am.,
34, 863–864. doi:http://dx.doi.org/10.1121/1.1918212
Martin, S. B., and Barclay, D. R. (2019). “Determining the dependence of marine pile driving
sound levels on strike energy, pile penetration, and propagation effects using a linear
mixed model based on damped cylindrical spreading,” J. Acoust. Soc. Am., 146, 109–121.
doi:10.1121/1.5114797
McFadden, D., and Plattsmier, H. S. (1983). “Frequency patterns of TTS for different exposure
intensities,” J. Acoust. Soc. Am., 74, 1178–1184. Retrieved from
http://link.aip.org/link/?JASMAN/74/1178/1
McGowen, M. R., Spaulding, M., and Gatesy, J. (2009). “Divergence date estimation and a
comprehensive molecular tree of extant cetaceans,” Mol. Phylogenet. Evol., 53, 891–906.
doi:10.1016/j.ympev.2009.08.018
Mikkelsen, L., Hermannsen, L., Beedholm, K., Madsen, P. T., and Tougaard, J. (2017).
“Simulated seal scarer sounds scare porpoises, but not seals: species-specific responses to
12 kHz deterrence sounds,” R. Soc. Open Sci., 4, 170286. doi:10.1098/rsos.170286
Miksis-Olds, J. L., Bradley, D. L., and Maggie Niu, X. (2013). “Decadal trends in Indian Ocean
ambient sound,” J. Acoust. Soc. Am., 134, 3464–3475. doi:10.1121/1.4821537
Miksis-Olds, J. L., and Nichols, S. M. (2016). “Is low frequency ocean sound increasing
globally?,” J. Acoust. Soc. Am., 139, 501–511. doi:10.1121/1.4938237
Miller, L. A., and Wahlberg, M. (2013). “Echolocation by the harbour porpoise: life in coastal
waters,” Front. Physiol., 4, 52. doi:10.3389/fphys.2013.00052
Møhl, B., and Andersen, S. (1973). “Echolocation: high-frequency component in the click of
the Harbour Porpoise (Phocoena ph. L.),” J. Acoust. Soc. …, 54, 1368–1372. Retrieved
from http://scitation.aip.org/content/asa/journal/jasa/54/5/10.1121/1.1914435
Møhl, B., Wahlberg, M., Madsen, P. T., Heerfordt, A., and Lund, A. (2003). “The monopulsed
nature of sperm whale clicks,” J. Acoust. Soc. Am., 114, 1143. doi:10.1121/1.1586258
Montgomery, J. C., Jeffs, A., Simpson, S. D., Meekan, M., and Tindle, C. (2017). “Sound as an
Orientation Cue for the Pelagic Larvae of Reef Fishes and Decapod Crustaceans,” Adv.
Mar. Biol., 51, 143–196. doi:10.1016/S0065-2881(06)51003-X
Mooney, T. A., Nachtigall, P. E., Breese, M., Vlachos, S., and Au, W. W. L. (2009). “Predicting
temporary threshold shifts in a bottlenose dolphin (Tursiops truncatus): The effects of
noise level and duration,” J. Acoust. Soc. Am., 125, 1816–1826. doi:10.1121/1.3068456
Morell, M., Brownlow, A., McGovern, B., Raverty, S. A., Shadwick, R. E., and André, M.

Bibliography
126
(2017a). “Implementation of a method to visualize noise-induced hearing loss in mass
stranded cetaceans,” Sci. Rep., 7, 41848. doi:10.1038/srep41848
Morell, M., Lehnert, K., IJsseldijk, L., Raverty, S., Wohlsein, P., Gröne, A., André, M., et al.
(2017b). “Parasites in the inner ear of harbour porpoise: cases from the North and Baltic
Seas,” Dis. Aquat. Organ., 127, 57–63. doi:10.3354/dao03178
Müller, A., and Zerbs, C. (2011). Offshore-Windparks Messvorschrift für
Unterwasserschallmessungen (Offshore-windfarm regulation for underwater sound
measurements) Hamburg: Müller-BBM und BSH, 35 pages. Retrieved from
https://doi.org/10.2314/GBV:771747691
Mulsow, J., and Reichmuth, C. (2010). “Psychophysical and electrophysiological aerial
audiograms of a Steller sea lion (<i>Eumetopias jubatus<i> ),” J. Acoust. Soc. Am., 127,
2692–2701. doi:10.1121/1.3327662
Nachtigall, P. E., and Supin, A. Y. (2014). “Conditioned hearing sensitivity reduction in a
bottlenose dolphin (Tursiops truncatus).,” J. Exp. Biol., 217, 2806–13.
doi:10.1242/jeb.104091
Nachtigall, P. E., Supin, A. Y., Pacini, A. F., and Kastelein, R. A. (2016). “Conditioned hearing
sensitivity change in the harbor porpoise (Phocoena phocoena),” J. Acoust. Soc. Am., 140,
960–967. doi:10.1121/1.4960783
Nachtigall, P. E., Supin, A. Y., Pacini, A. F., and Kastelein, R. A. (2017). “Four Odontocete
Species Change Hearing Levels When Warned of Impending Loud Sound,” Integr. Zool.,
, doi: 10.1111/1749-4877.12286. doi:10.1111/1749-4877.12286
National Marine Fisheries Service (2018). 2018 Revisions to: Technical Guidance for assessing
the effects of anthropogenic sound on marine mammal hearing (Version 2.0): Underwater
thresholds for onset of permanent and temporary threshold shifts 167 pages. Retrieved
from https://www.fisheries.noaa.gov/action/2018-revision-technical-guidance-assessing-
effects-anthropogenic-sound-marine-mammal-hearing
National Oceanic and Atmospheric Administration; National Marine Fisheries Service (2016).
Technical Guidance for Assessing the Effects of Anthropogenic Sound on Marine Mammal
Hearing: Underwater Acoustic Thresholds for Onset of Permanent and Temporary
Threshold Shifts Silver Spring, 178 pages. Retrieved from
http://www.nmfs.noaa.gov/pr/acoustics/Acoustic Guidance Files/opr-
55_acoustic_guidance_tech_memo.pdf
Nikolich, K., Frouin-Mouy, H., and Acevedo-Gutiérrez, A. (2018). “Clear diel patterns in
breeding calls of harbor seals ( Phoca vitulina ) at Hornby Island, British Columbia,
Canada,” Can. J. Zool., 96, 1236–1243. doi:10.1139/cjz-2018-0018
Nummela, S., Reuter, T., Hemilä, S., Holmberg, P., and Paukku, P. (1999). “The anatomy of
the killer whale middle ear (Orcinus orca),” Hear. Res., 133, 61–70. doi:10.1016/S0378-
5955(99)00053-2
Nummela, S., Thewissen, J. G. M., Bajpai, S., Hussain, S. T., and Kumar, K. (2004). “Eocene
evolution of whale hearing.,” Nature, 430, 776–778. doi:10.1038/nature02720
Nummela, S., Thewissen, J. G. M., Bajpai, S., Hussain, T., and Kumar, K. (2007). “Sound
transmission in archaic and modern whales: Anatomical adaptations for underwater
hearing,” Anat. Rec., 290, 716–733. doi:10.1002/ar.20528
Olesiuk, P. F., Nichol, L. M., Sowden, M. J., and Ford, J. K. B. (2002). “Effect of the sound
generated by an acoustic harassment device on the relative abundance and distribution of
harbor porpoises (Phocoena phocoena) in Retreat Passage, British Columbia,” Mar.
Mammal Sci., 18, 843–862. doi:10.1111/j.1748-7692.2002.tb01077.x

Bibliography
127
Otani, S., Naito, Y., Kato, a, and Kawamura, a (2000). “Diving behavior and swimming speed
of a free-ranging harbor porpoise, Phocoena phocoena,” Mar. Mammal Sci., 16, 811–814.
Otani, S., Naito, Y., Kato, A., and Kawamura, A. (2001). “Oxygen consumption and swim
speed of the harbor porpoise Phocoena phocoena,” Fish. Sci., 67, 894–898.
doi:10.1046/j.1444-2906.2001.00338.x
Parks, S. E., Cusano, D. A., Stimpert, A. K., Weinrich, M. T., Friedlaender, A. S., and Wiley,
D. N. (2014). “Evidence for acoustic communication among bottom foraging humpback
whales,” Sci. Rep., 4, 1–7. doi:10.1038/srep07508
Pelc, R., and Fujita, R. M. (2002). “Renewable energy from the ocean,” Mar. Policy, 26, 471–
479. doi:10.1016/S0308-597X(02)00045-3
Peschko, V., Ronnenberg, K., Siebert, U., and Gilles, A. (2016). “Trends of harbour porpoise
(Phocoena phocoena) density in the southern North Sea,” Ecol. Indic., 60, 174–183.
doi:10.1016/j.ecolind.2015.06.030
Pilz, P. K. D., Schnitzler, H.-U., and Menne, D. (1987). “Acoustic startle threshold of the albino
rat (<i>Rattus norvegicus<i\>),” J. Comp. Psychol., 101, 67.
Pirotta, E., Brookes, K. L., Graham, I. M., and Thompson, P. M. (2014). “Variation in harbour
porpoise activity in response to seismic survey noise,” Biol. Lett., 10, 5.
doi:10.1098/rsbl.2013.1090
Pirotta, E., Harwood, J., Thompson, P. M., New, L. F., Cheney, B., Arso, M., Hammond, P. S.,
et al. (2015). “Predicting the effects of human developments on individual dolphins to
understand potential long-term population consequences,” Proc. R. Soc. B Biol. Sci., 282,
20152109. doi:10.1098/rspb.2015.2109
Popov, V. V, Supin, A. Y., Rozhnov, V. V, Nechaev, D. I., and Sysueva, E. V (2014). “The
limits of applicability of the sound exposure level (SEL) metric to temporal threshold shifts
(TTS) in beluga whales, Delphinapterus leucas.,” J. Exp. Biol., 217, 1804–1810.
doi:10.1242/jeb.098814
Popov, V. V, Supin, A. Y., Rozhnov, V. V, Nechaev, D. I., Sysuyeva, E. V, Klishin, V. O.,
Pletenko, M. G., et al. (2013). “Hearing threshold shifts and recovery after noise exposure
in beluga whales, Delphinapterus leucas,” J. Exp. Biol., 216, 1587–1596.
doi:10.1242/jeb.078345
Popov, V. V, Supin A Ya, Wang, D., Wang, K., Dong, L., and Wang, S. (2011). “Noise-induced
temporary threshold shift and recovery in Yangtze finless porpoises Neophocaena
phocaenoides asiaeorientalis.,” J. Acoust. Soc. Am., 130, 574–584.
doi:10.1121/1.3596470
Prahl, S., Jepson, P. D., Sanchez-Hanke, M., Deaville, R., and Siebert, U. (2011). “Aspergillosis
in the middle ear of a harbour porpoise (Phocoena phocoena): A case report,” Mycoses,
54, 260–264. doi:10.1111/j.1439-0507.2010.01863.x
Pramik, B. (2013). “Marine Vibroseis: shaking up the industry,” First Break, 31, 67–72.
Le Prell, C. G. (2019). “Effects of noise exposure on auditory brainstem response and speech-
in-noise tasks: a review of the literature,” Int. J. Audiol., 58, S3–S32.
doi:10.1080/14992027.2018.1534010
R Core Team (2019). “R: A language and environment for statistical computing. R Foundation
for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.,”
Rankin, S., Archer, F., and Barlow, J. (2012). “Vocal activity of tropical dolphins is inhibited
by the presence of killer whales, Orcinus orca,” Mar. Mammal Sci., , doi: 10.1111/j.1748-
7692.2012.00613.x. doi:10.1111/j.1748-7692.2012.00613.x
Rauschmann, M. A., Huggenberger, S., Kossatz, L. S., and Oelschläger, H. H. A. (2006). “Head

Bibliography
128
morphology in perinatal dolphins: a window into phylogeny and ontogeny,” J. Morphol.,
267, 1295–1315. doi:10.1002/jmor.10477
Reeves, R. R., McClellan, K., and Werner, T. B. (2013). “Marine mammal bycatch in gillnet
and other entangling net fisheries, 1990 to 2011,” Endanger. Species Res., 20, 71–97.
doi:10.3354/esr00481
Reichmuth, C., Holt, M. M., Mulsow, J., Sills, J. M., and Southall, B. L. (2013). “Comparative
assessment of amphibious hearing in pinnipeds,” J. Comp. Physiol. A Neuroethol.
Sensory, Neural, Behav. Physiol., 199, 491–507. doi:10.1007/s00359-013-0813-y
Remy, T., and Mbistrova, A. (2018). Offshore wind in Europe - Key trends and statistics 2017
Brussels, 1–37 pages. doi:10.1016/S1471-0846(02)80021-X
Ridgway, S. H., and Carder, D. A. (1988). “Nasal Pressure and Sound Production in an
Echolocating White Whale, Delphinapterus leucas,” Anim. Sonar, Springer US, Boston,
MA, pp. 53–60. doi:10.1007/978-1-4684-7493-0_5
Rogers, T. L. (2017). “Calling underwater is a costly signal: size-related differences in the call
rates of Antarctic leopard seals,” Curr. Zool., 63, 433–443. doi:10.1093/cz/zox028
Rojano-Doñate, L., McDonald, B. I., Wisniewska, D. M., Johnson, M., Teilmann, J., Wahlberg,
M., Højer-Kristensen, J., et al. (2018). “High field metabolic rates of wild harbour
porpoises,” J. Exp. Biol., 221, jeb185827. doi:10.1242/jeb.185827
Rowe, S., and Hutchings, J. A. (2008). “A link between sound producing musculature and
mating success in Atlantic cod,” J. Fish Biol., 72, 500–511. doi:10.1111/j.1095-
8649.2007.01713.x
Ruser, A., Dähne, M., van Neer, A., Lucke, K., Sundermeyer, J., Siebert, U., Houser, D. S., et
al. (2016a). “Assessing auditory evoked potentials of wild harbor porpoises (Phocoena
phocoena),” J. Acoust. Soc. Am., 140, 442–452. doi:10.1121/1.4955306
Ruser, A., Dähne, M., Sundermeyer, J., Lucke, K., Houser, D. S., Finneran, J. J., Driver, J., et
al. (2014). “In-air evoked potential audiometry of grey seals (Halichoerus grypus) from
the North and Baltic Seas,” (B. Sokolowski, Ed.) PLoS One, 9, e90824.
doi:10.1371/journal.pone.0090824
Ruser, A., Woelfing, B., Baltzer, J., Schaffeld, T., Fischer, M., Schuster, M., Findeisen, H., et
al. (2016b). Ermittlung des Einflusses der Bauphase des Windparks Amrumbank West im
SCi Sylter Außenriff auf Schweinswale (Determination of Impact of the Construction
Phase of the Wind Farms “Butendiek” and “Amrumbank West” in SCI “Sylt Outer Reef”
on Harbor Porpoises Büsum, 75 pages.
Sarnocińska, J., Teilmann, J., Balle, J. D., van Beest, F. M., Delefosse, M., and Tougaard, J.
(2020). “Harbor Porpoise (Phocoena phocoena) Reaction to a 3D Seismic Airgun Survey
in the North Sea,” Front. Mar. Sci., 6, 1–10. doi:10.3389/fmars.2019.00824
Sayigh, L. S., Tyack, P. L., Wells, R. S., and Scott, M. D. (1990). “Signature whistles of free-
ranging bottlenose dolphins Tursiops truncatus: stability and mother-offspring
comparisons,” Behav. Ecol. Sociobiol., 26, 247–260. doi:10.1007/BF00178318
Schaffeld, T., Ruser, A., Schnitzler, J. G., Baltzer, J., and Siebert, U. (2018). Bewertung von
Auswirkungen von Unterwasserschallemissionen auf Schweinswale (Phocoena phocoena)
Büsum, 55 pages.
Schaffeld, T., Ruser, A., Woelfing, B., Baltzer, J., Kristensen, J. H., Larsson, J., Schnitzler, J.
G., et al. (2019). “The use of seal scarers as a protective mitigation measure can induce
hearing impairment in harbour porpoises,” J. Acoust. Soc. Am., 146, 4288–4298.
Schlundt, C. E., Finneran, J. J., Carder, D. A., and Ridgway, S. H. (2000). “Temporary shift in
masked hearing thresholds of bottlenose dolphins, Tursiops truncatus, and white whales,

Bibliography
129
Delphinapterus leucas, after exposure to intense tones,” J. Acoust. Soc. Am., 107, 3496–
3508.
Serpanos, Y. C., O’malley, H., and Gravel, J. S. (1997). “The relationship between loudness
intensity functions and the click-ABR wave V latency,” Ear Hear., 18, 409–419.
Shannon, G., McKenna, M. F., Angeloni, L. M., Crooks, K. R., Fristrup, K. M., Brown, E.,
Warner, K. A., et al. (2016). “A synthesis of two decades of research documenting the
effects of noise on wildlife,” Biol. Rev., 91, 982–1005. doi:10.1111/brv.12207
Siebert, U., Jepson, P. D., and Wohlsein, P. (2013). “First indication of gas embolism in a
harbour porpoise (Phocoena phocoena) from German waters,” Eur. J. Wildl. Res., 59,
441–444. doi:10.1007/s10344-013-0700-4
Siebert, U., Wünschmann, A., Weiss, R., Frank, H., Benke, H., and Frese, K. (2001). “Post-
mortem Findings in Harbour Porpoises (Phocoena phocoena) from the German North and
Baltic Seas,” J. Comp. Pathol., 124, 102–114. doi:10.1053/jcpa.2000.0436
Simmonds, M., Dolman, S., Jasny, M., Parsons, E., Weilgart, L., Wright, A., and Leaper, R.
(2014). “Marine Noise Pollution - Increasing Recognition But Need for More Practical
Action,” J. Ocean Technol., 9, 71–90. Retrieved from
https://animalstudiesrepository.org/acwp_ehlm/9
Širović, A., Hildebrand, J. A., and Wiggins, S. M. (2007). “Blue and fin whale call source levels
and propagation range in the Southern Ocean.,” J. Acoust. Soc. Am., 122, 1208–1215.
doi:10.1121/1.2749452
Sørensen, P. M., Wisniewska, D. M., Jensen, F. H., Johnson, M., Teilmann, J., and Madsen, P.
T. (2018). “Click communication in wild harbour porpoises (Phocoena phocoena),” Sci.
Rep., 8, 9702. doi:10.1038/s41598-018-28022-8
Southall, B. L., Bowles, A. E., Ellison, W. T., Finneran, J. J., Gentry, R. L., Greene Jr, C. R.,
Kastak, D., et al. (2007). “Marine mammal noise exposure criteria: Initial scientifc
recommendations,” Aquat. Mamm., 33, 411–522. doi:10.1080/09524622.2008.9753846
Southall, B. L., Finneran, J. J., Reichmuth, C., Nachtigall, P. E., Ketten, D. R., Bowles, A. E.,
Ellison, W. T., et al. (2019). “Marine Mammal Noise Exposure Criteria: Updated
Scientific Recommendations for Residual Hearing Effects,” Aquat. Mamm., 45, 125–232.
doi:10.1578/am.45.2.2019.125
Stöber, U., and Thomsen, F. (2019). “Effect of impact pile driving noise on marine mammals:
A comparison of different noise exposure criteria,” J. Acoust. Soc. Am., 145, 3252–3259.
doi:10.1121/1.5109387
Sueur, J., Aubin, T., and Simonis, C. (2008). “Seewave: a free modular tool for sound analysis
and synthesis,” Bioacoustics, 18, 213–226. doi:10.1080/09524622.2008.9753600
Supin, A. Y., Popov, V. V, and Mass, A. M. (2001). The sensory physiology of aquatic
mammals, Kluwer Academic Publishers, Boston/Dordrecht/London.
Szalma, J. L., and Hancock, P. A. (2011). “Noise effects on human performance: a meta-
analytic synthesis.,” Psychol. Bull., 137, 682–707. doi:10.1037/a0023987 682
Szymanski, M. D., Bain, D. E., Kiehl, K., Pennington, S., Wong, S., and Henry, K. R. (1999).
“Killer whale (Orcinus orca) hearing: Auditory brainstem response and behavioral
audiograms,” J. Acoust. Soc. Am., 106, 1134–1141.
Tasker, M. L., Amundin, M., Andre, M., Hawkins, a, Lang, W., and Merck, T. (2010). MARINE
STRATEGY FRAMEWORK Task Group 11 Report and other forms of energy Underwater
noise, Group, Vol. 245, 516–519 pages. doi:10.2788/87079
Tenghamn, R. (2006). “An electrical marine vibrator with a flextensional shell,” Explor.
Geophys., 37, 286–291. doi:10.1071/EG06286

Bibliography
130
Thiel, R. (2015). Atlas der Fische und Neunaugen Hamburgs - Arteninventar, Ökologie,
Verbreitung, Bestand, Rote Liste, Gefährdung und Schutz, Freie und Hansestadt Hamburg,
Behörde für Stadtentwicklung und Umwelt, Amt für Natur- und Ressourcenschutz,
Abteilung Naturschutz, Hamburg, 170 pages.
Tougaard, J., and Beedholm, K. (2018). “Practical implementation of auditory time and
frequency weighting in marine bioacoustics,” Appl. Acoust., 145, 137–143.
doi:10.1016/j.apacoust.2018.09.022
Tougaard, J., Carstensen, J., Teilmann, J., Skov, H., and Rasmussen, P. (2009). “Pile driving
zone of responsiveness extends beyond 20 km for harbor porpoises (Phocoena phocoena
(L.)).,” J. Acoust. Soc. Am., 126, 11–14. doi:10.1121/1.3132523
Tougaard, J., and Dähne, M. (2017). “Why is auditory frequency weighting so important in
regulation of underwater noise?,” J. Acoust. Soc. Am., 142, EL415–EL420.
doi:10.1121/1.5008901
Tougaard, J., Wright, A. J., and Madsen, P. T. (2015). “Cetacean noise criteria revisited in the
light of proposed exposure limits for harbour porpoises,” Mar. Pollut. Bull., 90, 196–208.
doi:10.1016/j.marpolbul.2014.10.051
Varanasi, U., Feldman, H. R., and Malins, D. C. (1975). “Molecular basis for formation of lipid
sound lens in echolocating cetaceans,” Nature, 255, 340–343. doi:10.1038/255340a0
Verfuß, U. K., Miller, L. A., Pilz, P. K. D., and Schnitzler, H.-U. (2009). “Echolocation by two
foraging harbour porpoises (Phocoena phocoena),” J. Exp. Biol., 212, 823–834.
doi:10.1242/jeb.022137
Verfuß, U. K., Miller, L. A., and Schnitzler, H.-U. (2005). “Spatial orientation in echolocating
harbour porpoises (Phocoena phocoena),” J. Exp. Biol., 208, 3385–3394.
doi:10.1242/jeb.01786
Vermeij, M. J. A., Marhaver, K. L., Huijbers, C. M., Nagelkerken, I., and Simpson, S. D.
(2010). “Coral Larvae Move toward Reef Sounds,” (S. Vollmer, Ed.) PLoS One, 5,
e10660. doi:10.1371/journal.pone.0010660
Villadsgaard, A., Wahlberg, M., and Tougaard, J. (2007). “Echolocation signals of wild harbour
porpoises, Phocoena phocoena,” J. Exp. Biol., 210, 56–64. doi:10.1242/jeb.02618
Viquerat, S., Herr, H., Gilles, A., Peschko, V., Siebert, U., Sveegaard, S., and Teilmann, J.
(2014). “Abundance of harbour porpoises (Phocoena phocoena) in the western Baltic, Belt
Seas and Kattegat,” Mar. Biol., 161, 745–754. doi:10.1007/s00227-013-2374-6
Ward, W. D., Cushing, E. M., and Burns, E. M. (1976). “Effective quiet and moderate TTS:
Implications for noise exposure standards,” J. Acoust. Soc. Am., 59, 160–165.
doi:10.1121/1.380835
Wehrmeister, E., Ulrich, A., Danehl, S., Lehnert, K., Schmidt, K., Hillmann, M., Siebert, U., et
al. (2013). Lebensraumverbesserung des Ostseeschweinswales Büsum, Germany:
Naturschutzstiftung Deutsche Ostsee, 1–33 pages.
Weilgart, L., and Whitehead, H. (1997). “Group-specific dialects and geographical variation in
coda repertoire in South Pacific sperm whales,” Behav. Ecol. Sociobiol., 40, 277–285.
doi:10.1007/s002650050343
Wenger, D., and Koschinski, S. (2012). “Harbour porpoise (Phocoena phocoena Linnaeus,
1758) entering the Weser river after decades of absence,” Mar. Biol. Res., 8, 737–745.
Wenger, D., Siebert, U., and Hennig, V. (2016). “The return of harbour porpoises (<i>Phocoena
phocoena) to the lower Elbe and Weser rivers , following anadromous fish shoals ,
foraging in Hamburg harbour,” Conf. Eur. Cetacean Soc. Poster Present.,.
doi:10.13140/RG.2.2.35932.69766

Bibliography
131
Widgren, S., and Hulbert, C. (2019). “rmatio: Read and write ‘Matlab’ Files,.”
Wiemann, A., Andersen, L. W., Berggren, P., Siebert, U., Benke, H., Teilmann, J., Lockyer,
C., et al. (2010). “Mitochondrial control region and microsatellite analyses on harbour
porpoise (Phocoena phocoena) unravel population differentiation in the Baltic Sea and
adjacent waters,” Conserv. Genet., 11, 195–211. doi:10.1007/s10592-009-0023-x
Williams, R., Wright, a. J., Ashe, E., Blight, L. K., Bruintjes, R., Canessa, R., Clark, C. W., et
al. (2015). “Impacts of anthropogenic noise on marine life: Publication patterns, new
discoveries, and future directions in research and management,” Ocean Coast. Manag.,
115, 17–24. doi:10.1016/j.ocecoaman.2015.05.021
Williams, T. M., and Worthy, G. A. J. (2002). “Anatomy and Physiology: the Challenge of
Aquatic Living,” In A. R. Hoelzel (Ed.), Mar. mammal Biol. an Evol. approach, Blackwell
Science, pp. 73–97. Retrieved from
https://books.google.de/books?hl=de&lr=&id=OwF05MfTagsC&oi=fnd&pg=PA73&dq
=marine+mammal%3B+energy%3B+challenge%3B+breathing%3B+cold+environment
&ots=myrE5xACDi&sig=NvJKNnhICwF_Fio6mAcji0gjydc#v=onepage&q&f=true
Wisniewska, D. M., Johnson, M., Teilmann, J., Rojano-Doñate, L., Shearer, J., Sveegaard, S.,
Miller, L. A., et al. (2018a). “Response to ‘Resilience of harbor porpoises to anthropogenic
disturbance: Must they really feed continuously?,’” Mar. Mammal Sci., 34, 265–270.
doi:10.1111/mms.12463
Wisniewska, D. M., Johnson, M., Teilmann, J., Siebert, U., Galatius, A., Dietz, R., and Madsen,
P. T. (2018b). “High rates of vessel noise disrupt foraging in wild harbour porpoises (
Phocoena phocoena ),” Proc. R. Soc. B Biol. Sci., 285, 20172314.
doi:10.1098/rspb.2017.2314
Wisniewska, D. M. M., Johnson, M., Teilmann, J., Rojano-Doñate, L., Shearer, J., Sveegaard,
S., Miller, L. A. A., et al. (2016). “Ultra-High Foraging Rates of Harbor Porpoises Make
Them Vulnerable to Anthropogenic Disturbance,” Curr. Biol., 26, 1441–1446.
doi:10.1016/j.cub.2016.03.069
Wohlsein, P., Seibel, H., Beineke, A., Baumgärtner, W., and Siebert, U. (2019).
“Morphological and Pathological Findings in the Middle and Inner Ears of Harbour
Porpoises (Phocoena phocoena),” J. Comp. Pathol., 172, 93–106.
doi:10.1016/j.jcpa.2019.09.005
World Health Organization (2017). “Burden of disease from environmental noise:
Quantification of healthy life years lost in Europe,” Geneva, Switzerland: World Health
Organization.
Wright, A. J., Soto, N. A., Baldwin, A. L., Bateson, M., Beale, C. M., Clark, C., Deak, T., et al.
(2007). “Do Marine Mammals Experience Stress Related to Anthropogenic Noise?,” Int.
J. Comp. Psychol., 20, 274–316.
Würsig, B., Greene Jr, C. R., and Jefferson, T. A. (2000). “Development of an air bubble curtain
to reduce underwater noise of percussive piling,” Mar. Environ. Res., 49, 79–93.
doi:10.1016/S0141-1136(99)00050-1
Yovel, Y., Falk, B., Moss, C. F., and Ulanovsky, N. (2010). “Optimal localization by pointing
off axis,” Science (80-. )., 327, 701–704. doi:10.1126/science.1183310
Zampolli, M., Nijhof, M. J. J., de Jong, C. A. F., Ainslie, M. A., Jansen, E. H. W., and Quesson,
B. A. J. (2013). “Validation of finite element computations for the quantitative prediction
of underwater noise from impact pile driving,” J. Acoust. Soc. Am., 133, 72–81.
doi:10.1121/1.4768886
Zimmer, W. M. X., Tyack, P. L., Johnson, M. P., and Madsen, P. T. (2005). “Three-dimensional

Bibliography
132
beam pattern of regular sperm whale clicks confirms bent-horn hypothesis,” J. Acoust.
Soc. Am., 117, 1473. doi:10.1121/1.1828501

List of figures
133
List of figures
FIG. 1: Research area in the German North Sea with marked positions of underwater
recorders (triangle) and positions where pile driving activities were conducted (points).
.......................................................................................................................................... 21
FIG. 2: Third octave level spectrum of the analyzed pile driving events, color and symbol
coded for measuring position and pile. ............................................................................ 26
FIG. 3: Determined SELSS in dB (re 1 µPa²s) of pile driving activities during the construction
of 50 monopiles in the Amrumbank West OWF.............................................................. 27
FIG. 4: Simulation harbor porpoise fleeing from a certain start position (x-axis) up to the safe
distance (5604 m). ............................................................................................................ 31
FIG. 5: Radii of hazard zones around a pile driving site, where the TTSonset of 175 dB re 1
µPa²s can be exceeded by multiple events above the SHEQ of 145 dB re 1 µPa²s for
single strikes. .................................................................................................................... 32
FIG. 6: Tone pips, used as hearing stimuli. .............................................................................. 51
FIG. 7: Recorded frequency spectrum of one generated artificial seal scarer signal (top),
which was used for animal exposure................................................................................ 54
FIG. 8: Example of measured AEP for 8 sound pressure levels (colour coded) between 87 and
122 dB re 1 µPa. ............................................................................................................... 57
FIG. 9: Effects of seal scarer exposure (at 14 kHz, with four harmonics) on hearing thresholds
at 20 (black circles) and 28 kHz (grey triangles). ............................................................ 59
FIG. 10: Estimated hazard area where single seal scarers signals exceed the determined SPL
TTSonset of 155.2 dB (pp), corresponding to a SPL TTSonset of 146.2 dB re 1 µPa
(rms, dashed line). ............................................................................................................ 64
FIG. 11. Linear regression of SEL–SPL of all played back seal scarer signals during hearing
tests at 20 and 28 kHz. ..................................................................................................... 70
FIG. 12: Effects of seal scarer exposure on hearing thresholds at 20 (circles) and 28 kHz
(triangles). ........................................................................................................................ 71
FIG. 13: Study area in the Elbe River in the West of Hamburg. The positions of the
underwater flowmeters are shown by red triangles. ......................................................... 78
FIG. 14: Top view on the modelled sound field of the acoustic flowmeter on the north (top
row) and south (bottom row) riverbank. .......................................................................... 80
FIG. 15: Top view on the combined sound field of the two flowmeters at a depth of 5 m. .... 81
FIG. 16: Effects of 28 kHz tone exposure on hearing thresholds at 28 kHz (black circles). ... 86
FIG. 17: Received cumulative sound exposure levels (SELcum) on simulated transects along
acoustic flowmeters. ......................................................................................................... 88
FIG. 18: Required time within sound field to exceed cumulative SEL-TTSonset. .................... 89

List of tables
134
List of tables
TABLE I. Summary of variables and values used for the model. ........................................... 24
TABLE II. Simulation results for the maximum distance as a start position for a flight, where
a TTS could still be induced before reaching the safe distance. ...................................... 30
TABLE III: Application example of simulation approach, using the acoustic properties of pile
driving strikes reported by Dähne et al. (2017). ............................................................... 38
TABLE IV. Summary of the determined TTSonset after exposure to artificial seal scarer
signals. .............................................................................................................................. 60
TABLE V: Measured hearing thresholds, which have been used to calculate mean hearing
thresholds ......................................................................................................................... 68
TABLE VI. Measured hearing thresholds after exposure, which resulted in a significant
hearing shift, relative to the baseline. ............................................................................... 69
TABLE VII: Harbour porpoise positions on simulated flight tracks. ...................................... 83

Acknowledgement
135
Acknowledgement
I wish to acknowledge my supervisors and the external reviewer allowing me to work on this
thesis. I would like to express my very great appreciation to Ursula Siebert for giving me the
opportunity to focus on this highly interesting topic and her continuous trust and support.
I am particularly grateful for the assistance given by the entire ITAW-crew in Büsum, who are
creating a family-like atmosphere. As very important representatives of this second family. I
would like to thank Bianca Unger, Abbo van Neer and Dominik Nachtsheim for inspiring
discussions, calming down and sharing excitement in changing roles. But also to Susanne
Carstens-Michaelis for keeping the team together.
I am especially grateful for my bioacoustics group with the best colleagues to imagine, who
always supported me without any doubts. Johannes Baltzer, you are my most favourite
roommate and I really appreciated your calm manner, lending both ears and providing
distraction or attention, when critically needed. Thank you Fiete, you are a great mentor and
taught me countless theoretical and practical things for experiments and life. My deepest
appreciations goes to my second mentor Joseph Schnitzler who pushed me a lot and taught me
so much about efficiency and the importance of broaden one’s mind. You both shaped the
scientist I am today.
I would like to express my greatest appreciation to my whole family: Mom, Dad, Gerrit and
Marvin, thank you for your continuous support and trust. You were always there for me and
supported every step I made. I hope I can give things back to my parents and will be some day
half a good role model as they were for me.
I owe my deepest gratitude to my lovely wife Claudia. All your love and trust was the most
important support of all. You made it possible that I could do this all. Thanks for your patience
and understanding. You took care that I could live in my bubble while you managed
overwhelming projects and simultaneously took care that I left the bubble from time to time to
breathe. Thank you for all the long evenings and weekends with experiments, in which you
were multiple exposed to my obstacles of the thesis with varying sound exposure levels. You
never left the experiments and always had my back.
A last thank goes to my kind logarithms, you are true friends.

Declaration of Originality
136
Declaration of Originality
I hereby declare that this dissertation, entitled “Effect of anthropogenic underwater noise on
harbour porpoise hearing in areas of high ecological importance” is my own original work.
Further, I have acknowledged all sources used and have cited these in the reference section.
No additional help from third parties was used for preparing this thesis.
Neither did I consult any services, such as consulting agencies, for completing this thesis nor
did anybody receive direct or indirect payment for carrying out any tasks related to this thesis.
The dissertation was drawn up in the following institution:
Institute for Terrestrial and Aquatic Wildlife Research, University of Veterinary Medicine
Hannover, Foundation, Werftstraße 6, 25761 Büsum, Germany
The dissertation has not been submitted previously to any institution for assessment purposes.
I hereby confirm that, to the best of my knowledge, all the above and attached declarations are
complete and correct.
………………………………………………………………………………
Date, Signature

Curriculum vitae
137
Curriculum vitae
Personal Details
Name: Tobias Schaffeld
Birthday: 20.01.1987
Birthplace: Kamp-Lintfort
Nationality: German
University Education
2011 – 2016: Master of Science
Wildlife Ecology and Wildlife Management
University of Natural Resources and Life Sciences
Vienna, Austria
Thesis title: “Reduction of accidental bycatches in set-net
fisheries by the use of acoustic signals”
Final grade: 2.0
2007 – 2011 Bachelor of Science
Biology
Heinrich-Heine University,
Düsseldorf, Germany
Thesis title: “Analysis of the foraging behavior of harbour
porpoises (Phocoena phocoena) in selected areas of the Baltic
Sea by static-passive-acoustic monitoring”
Final grade: 2.4
School Education
1997 – 2006: Abitur (A-levels, higher education entrance qualification)
Amplonius-Gymnasium, Rheinberg
1993 – 1997: Primary school
Gemeinschaftsgrundschule Millingen, Rheinberg

Curriculum vitae
138
Work experience
Since 2017 Doctoral candidate
Institute for Terrestrial and Aquatic Wildlife Research,
University of Veterinary Medicine Hannover, Foundation
Since 2016 Research associate in different projects of external funds
Institute for Terrestrial and Aquatic Wildlife Research,
University of Veterinary Medicine Hannover, Foundation
2015 – 2016 Student research assistant in different projects of external funds
Institute for Terrestrial and Aquatic Wildlife Research,
University of Veterinary Medicine Hannover, Foundation
2012 – 2014 Student research assistant in project “Testing new acoustic
harbour porpoise anti-bycatch devices in the German set-net
fisheries.
Von Thünen Institute of Baltic Sea Fisheries
Rostock, Germany
2011 – 2012 Student research assistant in different projects of external funds
German Oceanographic Museum
Stralsund, Germany
2010 Student research assistant, tutor in the course „Biology for
physicians”
Heinrich-Heine University
Düsseldorf, Germany
Scholarships
Grant of the “Gesellschaft der Freunde der TiHo“ for giving a talk and presenting a poster
entitled: “Effects of multiple exposures to pile driving noise on harbour porpoise hearing
during simulated flights a risk evaluation tool” at the „Effects of Noise on Aquatic Life”
conference from 07.-12.07.2019 in the Hague, Netherlands.
Grant for presenting a poster entitled: “Living on the bright side – Harbour porpoises don’t like
rain” and attending the “BioAcoustic Summer School (SeaBASS)” from 5.-10.06.2016 in
Syracuse, USA.

Curriculum vitae
139
Publications
Schaffeld, T., Schnitzler, J. G., Ruser, A., Woelfing, B., Baltzer, J., and Siebert, U. (2020).
“Effects of multiple exposures to pile driving noise on harbor porpoise hearing during
simulated flights—An evaluation tool,” J. Acoust. Soc. Am., 147, 685–697.
doi:10.1121/10.0000595
Schaffeld, T., Ruser, A., Woelfing, B., Baltzer, J., Kristensen, J. H., Larsson, J., Schnitzler, J.
G., Siebert, U. (2019). “The use of seal scarers as a protective mitigation measure can
induce hearing impairment in harbour porpoises,” J. Acoust. Soc. Am., 146, 4288–4298.
Zein, B., Woelfing, B., Dähne, M., Schaffeld, T., Ludwig, S., Rye, J. H., Baltzer, J., Ruser,
A., Siebert, U. (2019). “Time and tide: Seasonal, diel and tidal rhythms in Wadden Sea
Harbour porpoises (Phocoena phocoena),” (S. Li, Ed.) PLoS One, 14, e0213348.
doi:10.1371/journal.pone.0213348
Schnitzler, J. G., Pinzone, M., Autenrieth, M., van Neer, A., IJsseldijk, L. L., Barber, J. L.,
Deaville, R., Jepson, P., Brownlow, A., Schaffeld, T., Thomé, J.P., Tiedemann, R., Das,
K. (2018). “Inter-individual differences in contamination profiles as tracer of social
group association in stranded sperm whales,” Sci. Rep., 8, 10958. doi:10.1038/s41598-
018-29186-z
Schnitzler, J. G., Frédérich, B., Früchtnicht, S., Schaffeld, T., Baltzer, J., Ruser, A., and
Siebert, U. (2017). “Size and shape variations of the bony components of sperm whale
cochleae,” Sci. Rep., 7, 46734. doi:10.1038/srep46734
Schaffeld, T., Bräger, S., Gallus, A., Dähne, M., Krügel, K., Herrmann, A., Jabbusch, M.,
Ruf, T., Verfuß, U.K., Benke, H., Koblitz, J.C. (2016). “Diel and seasonal patterns in
acoustic presence and foraging behaviour of free-ranging harbour porpoises,” Mar. Ecol.
Prog. Ser., 547, 257–272. doi:10.3354/meps11627





























