Embed Size (px)
Citation preview

lable at ScienceDirect
Tuberculosis 91 (2011) 277e286
Contents lists avai
Tuberculosis
journal homepage: http : / / int l .e lsevierhealth.com/journals / tube
DRUG DISCOVERY AND RESISTANCE
Effective inhibitors of the essential kinase PknB and their potential asanti-mycobacterial agents
Kathryn E.A. Lougheed a,d, Simon A. Osborne b, Barbara Saxty b, David Whalley b, Tim Chapman b,Nathalie Bouloc b, Jasveen Chugh b, Timothy J. Nott c, Dony Patel c, Vicky L. Spivey a,Catherine A. Kettleborough b, Justin S. Bryans b, Debra L. Taylor b, Stephen J. Smerdon c, Roger S. Buxton a,*
aDivision of Mycobacterial Research, MRC National Institute for Medical Research, The Ridgeway, Mill Hill, London, NW7 1AA, United KingdombCentre for Therapeutics Discovery, MRC Technology, 1-3 Burtonhole Lane, Mill Hill, London, NW7 1AD, United KingdomcDivision of Molecular Structure, MRC National Institute for Medical Research, The Ridgeway, Mill Hill, London, NW7 1AA, United Kingdom
a r t i c l e i n f o
Article history:Received 25 October 2010Received in revised form3 March 2011Accepted 15 March 2011
Keywords:Mycobacterium tuberculosisKinasesPknBInhibitors
* Corresponding author. Tel.: þ44 20 8816 2225; faE-mail addresses: [email protected]
[email protected] (S.A. Osborne), [email protected] (D. Whalley),(T. Chapman), [email protected] (N. Boul(J. Chugh), [email protected] (T.J. Nott),(D. Patel), [email protected] (V.L. Spivey), catheac.uk (C.A. Kettleborough), [email protected] (D.L. Taylor), [email protected] (mrc.ac.uk (R.S. Buxton).
d Present address: Department of Life Sciences, SirSouth Kensington Campus, Imperial College LondonKingdom.
1472-9792/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.tube.2011.03.005
s u m m a r y
PknB is an essential serine/threonine kinase of Mycobacterium tuberculosis with possible roles ina number of signalling pathways involved in cell division and metabolism. We screened a library of>50,000 compounds for inhibitors of the in vitro phosphorylation of GarA (Rv1827) by PknB and iden-tified a number of inhibitors. A program of synthetic medicinal chemistry was subsequently conductedaround one class of inhibitors and was successful in generating ATP competitive inhibitors with potencyin the nanomolar range. Compounds in this class showed cross-reactivity with the related M. tuberculosiskinase, PknF, but not with PknG in an in vitro autophosphorylation assay. These synthesised inhibitorswere able to prevent the growth of M. tuberculosis in an Alamar blue assay and in an intracellular modelof infection, but only in the micromolar range. We attempted to determine if cell wall permeability wasan explanation for the discrepancy between the potent in vitro compared with relatively poor in vivoactivity, but found no evidence that the activity of the inhibitors could be improved by weakeningthe cell wall. Despite a number of drug discovery efforts attempting to develop inhibitors against PknB,it is yet to be reported that any such inhibitors prevent mycobacterial growth at submicromolarconcentrations.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Mycobacterium tuberculosis remains one of the world’s mostdevastating pathogens, with more than 13 million people sufferingfrom an active tuberculosis infection and 1.8 million resultingdeaths in 2008 alone.1 The emergence of multi-drug and exten-sively drug resistant strains has highlighted the need for new drugs
x: þ44 20 8906 4477.(K.E.A. Lougheed), [email protected] (B. Saxty),[email protected]), [email protected]@googlemail.com
[email protected] (J.S. Bryans), [email protected]. Smerdon), rbuxton@nimr.
Alexander Fleming Building,, London SW7 2AZ, United
All rights reserved.
to treat tuberculosis. Recent studies have focused on finding newpathways vulnerable to inhibition by small molecules and previ-ously unexploited by drug discovery efforts. The inhibition of sig-nalling pathways both in M. tuberculosis and the host may yieldnew classes of drug targets and a large amount of recent work hasfocused on developing this further.
Target based drug discovery, in which there is in vitro highthroughput screening of a large number of small molecules againsta validated target, has been used on a number of occasions to searchfor new anti-tuberculosis agents. We sought to find inhibitors of anessential M. tuberculosis serine/threonine protein kinase, PknB.Kinases are attractive as drug targets due to the range of crucialcellular processes in which they are involved. There has been muchinterest in developing ATP competitive kinase inhibitors for thetreatment of cancer, a hallmark of which is often aberrant kinaseactivity. A large number of small molecule kinase inhibitors havebeen developed as potential anti-cancer drugs and there is a hugeamount of interest in developing kinase inhibitors to treat a rangeof conditions.2 Kinase-focused libraries of small molecule inhibitorshave been built-up as a result of these studies and a large amount of

K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286278
knowledge has been gained on the action of kinase inhibitors. Theearly success stories from the development of eukaryotic kinaseinhibitors suggested that similar drugs could be developed to treatbacterial infections. The M. tuberculosis serine/threonine proteinkinases (STPKs) are attractive targets partly because of the inferredimportance of serine/threonine phosphorylation in M. tuberculosis:M. tuberculosis is unique within the bacterial world in havinga much higher number of STPKs compared to the more commontwo-component signalling systems.3e6 Mycobacterium leprae,during extensive reductive evolution, has lost all but four of thesekinases: PknA, PknB, PknG and PknL.7 PknA and PknB are essentialinM. tuberculosis,8e10 while PknG has been reported to be involvedin survival in the macrophage, either as a result of a role in glu-tamate metabolism11 or via modulation of host cell traffickingpathways.12
Previous studies have focused on the potential of PknB and PknGas drug targets in M. tuberculosis, although the majority havereported significantly less impressive activity against whole cellsthan against thepurifiedprotein invitro.8,12e14Despite this, there canbenodoubt, fromanumberof lines of evidence, that PknB carries outfunction(s) essential to M. tuberculosis, and possesses a number ofproperties that suggest it is suitable for development of novel drugs:a crystal structure has also been determined for PknB15,16 and thekinase domain exhibits less than 30% similarity with eukaryotickinases, which has positive implications for developing a drugspecific for the bacterial kinases and not those of the host. The pknBgene (Rv0014c) is part of an operon highly conserved among theactinomycetes and also encoding pknA, the other essential kinase;pstP, a phosphoserine/threonine phosphatase; rodA, a cell divisionprotein; pbpA; a protein involved in peptidoglycan biosynthesis; andtwo genes (fhaB and fhaA) coding for forkhead-associated proteins.Over-expression or depletion of this kinase results in gross changesin cell morphology, likely a result of the role of PknB in the phos-phorylation-mediated regulation of a numberof proteins involved incell wall biosynthesis.10,17,18 In addition, both PknB and PknG areknown to phosphorylate a protein involved in the regulation ofglutamate metabolism,19e21 suggesting that an inhibitor of one orboth of these kinases could also impact on the ability ofM. tuberculosis to adequately regulate its centralmetabolicprocesses.Targeting of these bacterial kinases would therefore be a way ofinhibiting evolutionarily-conserved steps in central metabolicprocesses.
We screened for smallmolecule inhibitors of PknBand, as a resultof a medicinal chemistry program (manuscript in preparation), ourlead compounds were able to inhibit PknB activity in vitro in thenanomolar range. However, the potency of our compounds againstwhole cells in culture or in amacrophagemodel of infectionwas twoorders of magnitude lower than expected from the in vitro potency.An often suggested explanation for low anti-tuberculosis activity isthe problem of cell wall permeability. Since the M. tuberculosis cellwall presents an extremely hydrophobic barrier which can impedethe entry of drugs into the cell, we sought to determine if cell wallpermeabilitymight explain the difficulties in improving the potencyof our PknB inhibitors. In addition, we investigated the role of effluxpumps, protein binding in the assay media and inhibitor specificityas alternative explanations.
2. Materials and methods
2.1. Compounds
Compounds for the high throughput screening included theMRCT compound collection comprising 45,000 diverse templatesfrom commercially available collections, as well as 6400 kinase-focused templates (Biofocus DPI, Cambridge, UK) selected on the
basis of bio-informatics provided by the crystal structure of PknBand other serine/threonine kinases. Focused libraries, based aroundcompounds identified from the initial screen, were subsequentlygenerated by synthetic medicinal chemistry.
2.2. Protein expression and purification
GarA was expressed in Escherichia coli and purified as des-cribed.22 The 279 residue kinase domain of PknB and the 292residue kinase domain of PknF were expressed as 3C-proteasecleavable GST-fusions in E. coli Rosetta 2 (DE3) pLysS cells. Theproteins were purified with glutathione Sepharose 4B resin(GE Healthcare) and the GST-tag cleaved from the protein prior toelution using 3C protease (GE Healthcare).
2.3. High throughput protein kinase assays
A non-radioactive PknB kinase assay was developed to followATP depletion during the PknB-catalyzed phosphorylation of GarA(Rv1827). The Promega ‘Kinase Glo’ assay kit was used to measurelevels of phosphorylation via detection of the remaining ATP afterthe completion of the kinase reaction. Briefly, luciferase and beetleluciferin were introduced to the reaction mix and, in the presenceof Mg2þ, ATP and oxygen, the oxygenation of luciferin generatesone photon of light per turnover. PknB activity is inverselyproportional to the intensity of the luminescence signal. Using thisassay format the Km of ATP was determined to be 1.5 mM and ATPwas used at this concentration for screening.
The following reaction mix was prepared: 0.15 mM PknB and5 mM Rv1827 in reaction buffer (50 mM TriseHCl, 150 mM NaCl,1 mM DTT and 0.01% Triton X-100). The assay was carried out in22 ml volumes in 384-well white plates (Matrix, 4316). A BeckmanBiomek Fx liquid handling robot was used to dispense premixedreaction mix into the wells and the 2 ml of the compound/DMSOadded to a final concentration of 10 mM/1% DMSO. The plates wereincubated for 30 min and the reaction initiated with the addition of1 mM MnCl2/1.5 mM ATP using the Perkin Elmer Flexdrop non-contact liquid dispenser. The reaction was stopped after 20 minwith the addition of 22 ml Kinase Glo reagent and luminescenceread with a BMG Polarstar reader. Control wells with and withoutPknB were included on every plate in columns 1, 2 and 23, 24 andwere used to calculate Z0 values.23 Data from any plates with Z0<0.5were rejected and were repeated.
2.4. Strains and growth conditions
M. tuberculosis H37Rv, Mycobacterium smegmatis mc2155,M. smegmatis DfbpA (gift from C. Thompson), and M. tuberculosisDompA (gift from P. Draper) cultures were grown at 37 �C in Dubosbroth supplemented with 0.05% (vol/vol) Tween 80, 0.2% (vol/vol)glycerol, and 4% (vol/vol) Dubos medium albumin (Becton Dick-inson). M. smegmatis strains were grown either in 5 ml volumes in20ml universals or in 50ml volumes in 250mlflasks, incubated in anorbital shaker at 150 rpmat 37 �C.M. tuberculosiswas growneither in100 ml volumes in a Bellco roll-in incubator (2 rpm) or in 10 mlvolumes in static universals. Kanamycin was used at a final concen-tration of 25 mg ml�1. M. tuberculosis was grown on 7H11 supple-mentedwith10%OADCenrichment (Middlebrook) and 0.5% (vol/vol)glycerol.
2.5. M. tuberculosis minimum inhibitory concentration (MIC)determination
MICs were determined for M. tuberculosis using a 96-well platemethod with Alamar blue, as described previously.24,25 A modified

K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286 279
version of the assay usedM. smegmatiswith a 3 day incubation. Thecell wall weakening and efflux pump inhibition assays were mod-ifications of the Alamar blue assay, as described in the results.Z0 values were used as a measure of assay quality, using thefollowing formula: Z0 ¼ 1e3 � SSD/R, where SSD is the sum of theStandard Deviation of the negative controls and Standard Deviationof the positive controls, and R is the mean of the maximum signalcontrol minus the mean of the negative signal control.23
2.6. Intracellular activity of PknB inhibitors in a macrophageinfection by M. tuberculosis
Intracellular inhibition was tested in murine bone marrow-derived macrophages at a range of concentrations as previouslydescribed.25 Briefly, monocytes were isolated from Balb/c bonemarrow and differentiated with L-cell supernatant for 5e7 days.Themacrophages were infected withM. tuberculosiswith anMOI of0.5 bacteria: 1 macrophage for 4 h, and the infection incubated inthe presence of various concentrations of the inhibitors for 5 days.At this point, the macrophages were lysed with dH20 þ 0.05%Tween 80 and plated for CFU enumeration. Prior to the infection,the compounds were tested to rule out any effect of toxicity againstthe macrophages. Alamar blue was used to determine macrophageviability after incubation with the inhibitors (not shown).
2.7. Kinase autophosphorylation assay
In vitro autophosphorylation of PknA, PknB, PknG and PknF(1 mg) was carried out in 20 ml reaction mixture containing 2.5 mlbuffer P (25mMTriseHCl, pH 7.0,1 mMdithiothreitol, 5mMMgCl2,1 mM EDTA). The reaction was initiated with the additional of0.08 mCi [g-32P] ATP and incubated at 37 �C for 30 min. The reactionwas terminated with the addition of SDS-PAGE sample buffer andheating themixture at 100 �C for 5min. The reactionmixtures wereanalyzed by SDS-PAGE. The gels were soaked in 20% tricholoro-acetic acid for 10 min at 90 �C and dried. Radioactive proteins werevisualized by autoradiography and quantified by Phosphorimageanalysis using a STORM 840 Optical Scanner System and Image-Quant (version 5.2, Molecular Dynamics, Sunnyvale, CA). Imageswere adjusted for contrast and brightness using Adobe Photoshop.
2.8. Radioactive competition assay
Reactions were set up in a volume of 17 ml kinase buffer (50 mMTriseHCl pH7.2, 1 mM DTT, 0.01% TX-100) containing 100 nMpurified PknB and 3 mM purified Rv1827 and increasing amounts ofeach inhibitor diluted in neat DMSO (from 0.03 mM to 100 mM) inpolypropylene U-bottomed 96 well plates. The reactions werestarted by the addition of 5 ml of solutions containing MnCl2 andATP at Km or 50xKm, to achieve final concentrations of 1 mMMnCl2with 1.5 mM ATP and 0.1 mCi 33P or 75 mM ATP and 1 mCi 33P.
After incubating for 70min at room temperature, reactions wereterminated by the addition of 20 ml 50% orthophosphoric acid.Reactions were then transferred to a 96 well glass filter captureplate (GF/C) and unbound 33P was washed away with 200 ml 1.5%orthophosphoric acid, using a Tomtec Harvester 9600. After dryingovernight at room temperature, 25 ml scintillant was added to eachwell and quantification of Rv1827 phosphorylation achieved bydetecting incorporated 33P with the PE Topcount.
2.9. Kinase profiling against a panel of mammalian kinases
This was carried out using a 33P-ATP filter-binding assay26 ona panel of 70 mammalian kinases at the National Centre for Protein
Kinase Profiling in the MRC Protein Phosphorylation Unit at theUniversity of Dundee.
2.10. Docking studies with PknB inhibitors
The mitoxantrone bound crystal structure (PDB:2FUM) wasselected for docking studies owing to its greater suitability in termsof ligand-induced binding site architecture as compared to the ATPbound structure (PDB:1MRU). Protein preparation, grid generationand docking (GlideSP) were all carried out using the Schrodingermolecular modelling suite (Schrodinger LLC: New York. http://www.schrodinger.com).
3. Results
3.1. High throughput screening for PknB inhibitors
PknB has been previously validated as a suitable target for drugdiscovery and we sought to identify small molecule inhibitorstargeted against this kinase. A number of potential substrates forPknB were tested, with GarA (Rv1827) being chosen as the moststrongly phosphorylated protein in our assay. GarA has beendescribed as an in vivo target for both PknB and PknG19,20 andinteraction with this target is known in some detail.22,27 A non-radioactive PknB kinase assay was developed to follow ATPdepletion during the PknB-catalyzed phosphorylation of GarA.Substrate Kms were determined for both ATP (1.5 mM) and Rv1827(>20 mM). The Km ATP concentrations and an excess of peptide(5 mM) were selected to achieve a robust assay window >0.5within the linear phase of the reaction, maximizing sensitivity toATP competitive inhibitors.
In total w54,000 compounds were screened against PknB. Thecompound collection was sourced by MRCT from commercialsuppliers. Drug-like filters to assess both physicochemical propertiesand known toxicophores were applied to the selection process. TheMRCT library comprised w44,000 diverse compound collection andw9000 kinase-focused collection (targeting ATP binding site). Inaddition, w1000 natural products (Phytoquest) were screened. Theaverage Z0 was 0.76 and a confirmed hit rate of 0.14%was observed. Intotal, 76 compounds confirmed IC50 in the range 1e200 mM.
Limitations of the Kinase glo assay format did not permitdetailed enzyme kinetics to be performed. However using a radio-active assay, a number of compounds were assessed for ATPcompetitiveness by measuring IC50 at a high (75 mM) and a low(1.5 mM, Km) ATP concentration. From the ChengePrussoff equa-tion28 we would predict that for an ATP competitive inhibitor,a 25-fold shift in IC50 would be seen between ATP at Km (1.5 mM)and 50XKm (75 mM). Five compounds from the series, including theoriginal screen hit, with a range of potencies 50e1000 nM in theprimary assay were selected. Fold shifts in the range 15.5e32 wereobserved for all compounds, consistent with an ATP competitivemechanism. It was expected that, due to the higher ATP concen-tration in the cells compared to the in vitro assay, a shift in MICcompared to IC50 enzyme activity would be observed. However,a consistent shift was not observed.
Docking studies (Figure 1) indicate that the compounds shouldbe ATP competitive. They bind to the hinge region via two H-bondsbetween the ligand core and the peptide backbone of Val95 alongwith an additional H-bond via the ligand pyrazole NH to thepeptide carbonyl of Glu93. Numerous Van Der Waals contactscontribute to binding, most notably between the ligand cyclopropylgroup in the hydrophobic pocket towards the Met92 gatekeeperand also via extensive contact along the glycine rich loop at theentrance to the binding site.

Figure 2. Compounds inactive against M. tuberculosis had a range of IC50s against thepurified protein. A similar spread was observed for the compounds able to inhibitM. tuberculosis. Starred section indicated the area of the graph where compoundshighly active against both the purified protein and whole cells would appear.
Figure 3. Intracellular activity of PknB inhibitors in a bone marrow derived macro-phage model. Macrophages were infected with an MOI of 0.5:1 and incubated in thepresence of a dilution series of the inhibitors. After 5 days, the macrophages were lysedand the intracellular bacteria enumerated by CFU platings. Data is expressed aspercentage survival compared to the untreated control. Inhibitor structures and cor-responding in vitro activity against purified protein are shown in Table 1.
Figure 1. Docking of MRCT67127 into PknB active site. Green ribbon represents PknB(Code: 1MRU) with ATP co-crystallised (shown as green structure). Grey ribbonrepresents PknB (Code: 2FUM) with mitoxantrone co-crystallised. The mitoxantronestructure was removed and docking studies conducted with this PknB conformation:owing to the flat nature of the docked compounds (making them similar to mitox-antrone), this was considered a better way to obtain reliable results. MRT67127 isshown (grey structure) docked into PknB.
K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286280
3.2. Anti-tuberculosis activity of lead compounds
Compounds that showed potent activity against PknB in vitro, nocytotoxicity and good selectivity against a panel of mammaliankinases, were tested in an M. tuberculosis Alamar blue MIC assay.Briefly, 2-fold dilutions of the inhibitors were incubated at 37 �Cwith approximately 1�105 cells per well in 96-well microplates for7 days. Untreated M. tuberculosis and media only controls wereincluded on every plate, and the control drugs isoniazid, strepto-mycin and ethambutol were tested in parallel with each batch ofPknB inhibitors to ensure that the assay was reproducible. TheAlamar blue reagent was added on the final day of the incubation,and the fluorescence measured at the completion of the experi-ment. A 90% inhibition of fluorescence compared to the untreatedcontrol was chosen as the cutoff for the determination of inhibitoryconcentrations. The statistical parameter, Z0-factor, was used asa measure of assay quality, with a score greater than 0.75 taken asan indication of good data.
MICs were determined for more than 100 compounds, witha range of MICs being observed varying between inactive at 250 mMto inhibitory at 8e16 mM. The compounds were also tested againstM. smegmatis (data not shown) with a generally good agreementbetween the two assays. This confirms previously published exper-iments (e.g.29) which have suggested that testing compounds againstM. smegmatis is a goodmodel for TB drug discovery. Therewas a poorcorrelation between IC50 against the purified recombinant proteinand anti-M. tuberculosis MIC, with the majority of compoundsproving to be inactive against the whole cells. The lack of correlationbetween protein and whole cell activity is illustrated in Figure 2.Despite extensive rounds of chemistry, unfortunately it was notpossible to reduce the M. tuberculosis MICs below the 10 mM level.
3.3. Intracellular activity of PknB inhibitors against M. tuberculosisin a bone marrow derived macrophage model of infection
A subset of compounds was chosen for further investigation.The key criteria in this selection included reasonable activityagainst M. tuberculosis in vitro, a low MIC to IC50 ratio as a possible
indication of good cell wall permeability, good selectivity againsta panel of human kinases and good physicochemical parameters,includingmoderate logD and low hydrogen bond donor count, bothof which are requirements for good membrane permeability. Theintracellular activity of these compounds against M. tuberculosis ina bone marrow derived macrophage model of infection was tested.
The compounds showed a dose dependant inhibition of intra-cellularM. tuberculosis, although a number of the compounds werenot able to inhibit 99% growth compared to the untreated control(chosen as the cutoff for determining intracellular inhibition).Figure 3 shows the inhibition of intracellular growth by the 12 PknBinhibitors chosen for further investigation. At the lowest concen-tration tested, 2.5 mM, none of the compounds were able to inhibitgrowthbymore than99%of the control. Anumberof the compoundswere active at the higher concentrations of 10 and 20 mM. Whileconcentrations up to 100 mMwere tested, these are not showndue toa number of the compounds proving to have toxic effects against themacrophages, possibly due to inhibition of host kinases.

K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286 281
A summary of the data is shown in Table 1. Little correlationbetween intracellular activity and whole cell inhibition was found.
3.4. Role of cell wall permeability and efflux on inhibitor MIC
Poor anti-tuberculosis activity by compounds that are potent inthe in vitro protein assay is often attributed to impermeability of theM. tuberculosis cell wall. M. tuberculosis has a thick and ‘waxy’ cellwall, which limits the ability of compounds to penetrate and reachthe cytoplasm.30 Much of the impermeability stems from theunique mycolic acid layer. While little is known about the specificproperties of small molecules that enable them to diffuse across theM. tuberculosis cell wall, it is possible to modify certain functionalgroups of the compounds inways that will hopefully improve entryinto the cytoplasm. For example, modifications were made to ourmore active compounds in an attempt to vary the compounds’ LogPvalues and increase their hydrophobicity (to be discussed ina separate medicinal chemistry paper). In one case, it was possibleto improve the MIC/IC50 ratio using this approach. However, theresulting activity of the compounds againstM. tuberculosiswas stillpoor. Before embarking on further chemistry with the hope ofimproving cell penetration, we attempted to determine whethercell wall permeability truly was a major contribution to low MICvalues through the use of several cell wall-weakened models withthe subset of inhibitors shown in Figure 3.
A number of chemicals and antibiotics were tested, includingTriton X-100, glycine, lysozyme, isoniazid and ethambutol. Therehave been previous reports of sub-lethal concentrations of antibi-otics targeted to the cell wall being able to enhance permeabilityof certain compounds.31,32 In particular, ethambutol has beendemonstrated to enhance the activity of rifampicin, streptomycinand isoniazid against drug resistant strains.33 In our experiments,the cell wall weakening agent was used at 0.5 MIC in conjunctionwith the PknB inhibitors to determine whether the inhibitors werebetter able to inhibit M. tuberculosis in the presence of a weakenedcell wall. As a control, we demonstrated that the MIC of rifampicinwas reduced by 4-fold in the presence of sub-inhibitory concen-trations of ethambutol, confirming published data that reportedthat accumulation of and sensitivity to rifampicin is enhanced byethambutol.34 We tested a number of compounds, including thesubset of 12 active compounds and a number that were previouslyinactive against M. tuberculosis. Several compounds were able toinhibitM. tuberculosis in the presence of ethambutol when they hadbeen previously inactive. This implied that cell wall permeabilitywas, indeed, a problem for some structural families. However, theMICs of the most efficacious compounds were not improved byethambutol treatment.
In addition to the ethambutol cell-wall weakened model, wewanted to use a more defined model. AnM. smegmatis fbpAmutanthas been shown to possess increased sensitivity to a number ofantibiotics due to alterations in its cell wall.35 FbpA is involved inmycolic acid biosynthesis and the mutant has a loss of cell wallhydrophobicity compared with the wild type. We tested thesensitivity of this mutant to the PknB inhibitors and, in agreementwith the ethambutol data, there was no improvement in inhibitionwith a weakened cell wall. In comparison, vancomycin MIC wasimproved by 16-fold and erythromycin by 8-fold. This effect wasspecific to certain antibiotics as the MICs and streptomycin andethambutol were not improved in the assay.
After testing the role of the mycolic acid layer in drug perme-ability, we sought also to investigate other possible paths of entryinto the cell. Antibiotics can either diffuse across the cell wall,a possibility investigated in the experiments detailed above, oralternatively may enter through hydrophilic channels. Porins in themycobacterial outer membrane are required for the diffusion of
hydrophilic solutes and are the route of entry for small and hydro-philic antibiotics such as isoniazid and ethambutol.36 It has beendemonstrated that an mspA porin deletion mutant of M. smegmatishas a multi-drug resistant phenotype to antibiotics includingampicillin.37 Although M. tuberculosis does not encode Msp-likeproteins, expression of M. smegmatis MspA in M. tuberculosis enh-ances antibiotic sensitivity38 and anM. tuberculosis outermembranechannel-forming protein has been demonstrated to restore antibi-otic sensitivity to a M. smegmatis mspA mutant.39 One of the bestunderstoodporins inM. tuberculosis is OmpA, thathas been linked tothe diffusion of pyrazinamide across the cell wall.40 M. tuberculosisompAmutants have not been reported to have an antibiotic resistantphenotype similar to M. smegmatis mspA. Therefore, we tested thesensitivity of the mutant to a number of antibiotics compared to thewild type strain. UnlikeM. smegmatismspA,M. tuberculosis ompAdidnot have increased sensitivity to the small, hydrophilic antibioticstested. However, it did show a slightly increased resistance to thelarge antibiotics vancomycin and erythromycin. This result was alsoobserved for M. smegmatis mspA, despite both of these antibioticsbeing too large to enter the cell via porins. It is likely that alterationsin the cell wall have a knock-on effect on entry of large antibioticsinto the cell. This experiment does not provide any evidence thatsmall antibiotics enter M. tuberculosis via OmpA channels, but wecannot rule out that OmpA is involved in the uptake of other anti-biotics not tested. We also tested the ompA mutant with the PknBinhibitors and observed that the majority of them were equallyactive against the wild type and mutant.
The activity of efflux pump inhibitors has been shown to playa role in the acquired drug resistance ofM. tuberculosis for a numberof antibiotics,41e44 due to the upregulation of efflux pumps in drugresistant strains. It has been suggested that the use of efflux pumpinhibitors in conjunction with anti-tuberculosis therapy could beused to reverse the efflux pump-mediated drug resistance in someinfections.45 There is also some evidence that efflux plays a role inthe intrinsic drug resistance of mycobacteria.46e48 Therefore, wetested whether the addition of sub-inhibitory concentrations of theefflux pump inhibitors reserpine and verapamil to our assay wouldimprove the PknB inhibitor MICS. No change was observed in MICat either 20 or 40 mg/ml of either efflux pump inhibitor, while theMIC of the ethidium bromide control was decreased by 4-fold. Thisis perhaps not surprising due to the observation that the effect ofefflux inhibitors is most dramatic when there is overexpressionor upregulation of efflux pumps, rather than against wild typelevels.34,45
3.5. Specificity of PknB inhibitors
Attempts to select resistant mutants of the 12 most potent PknBinhibitors were unsuccessful, possibly due to either the high MICsof the compounds, or non-specific activity of the inhibitors. Weplated 5 � 109 M. tuberculosis directly on to media containing 2, 4and 8�MIC inhibitors as well as pre-growing the cells on 0.5 and1�MIC before replating on the above concentrations. Neithermethod generated any resistant colonies. This, combined with thelack of correlation between MIC and activity in the in vitro proteinassay led us to consider whether the inhibitors are truly targetingPknB. It is possible that some of the anti-tuberculosis activity of theinhibitors is due to off-target activity, meaning that further PknB-focused chemistry based around these structural families is un-likely to lead to an improvement in MIC. Alternatively, despite goodselectivity overmammalian kinases, the inhibitors could be bindingto a number of the other more closely related M. tuberculosiskinases, and it is possible that the inhibitor is being sequesteredaway from PknB by its activity against any of the eight non-essential kinases in M. tuberculosis.

Table 1Structures and inhibitor activity against purified PknB andM. tuberculosis in an Alamar blue and an intracellular assay. IC50 is shown as the mean of three experimental repeats,Alamar blue and intracellular MIC is representative of three or more repeats.
Inhibitor Structure IC50 PknB mM MIC M. tuberculosis mM* MIC Intracellular mMy
1.MRT67127N
N
NH
NH
N
N
NR
0.053 32 5
2.MRT67153
N
N
NH
NH
N
ON
0.056 32 20
3.MRT68667
N
N
Br
NH NH
N
NH
O
N
N
0.09 64 20
4.MRT68606N
N
NH
N
F
F
F
N
N
Br
R
0.519 16 >20
5.MRT68572N
N
NH
N F
F
F
N
NR
16.554 16 >20
6.MRT68634N
N
NH
N
O
NH
N
NR
1.499 64 >20
K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286282

Table 1 (continued)
Inhibitor Structure IC50 PknB mM MIC M. tuberculosis mM* MIC Intracellular mMy
7.MRT67131
N
N
NH
NH
N
NH
NH
N
0.345 125 >20
8.MRT67150
N
N
NH
NH
N
ON
0.055 32 5
9.MRT67155
N
N
NH
NH
N
ON
N 0.096 64 10
10.MRT67156
N
N
NH
NH
N
ON
0.092 16 5
11.MRT67319N
N
NH
N
N
NR
6.194 32 10
12.MRT68664
N
N
Br
NH NH
N
NH
O
0.065 64 5
* 90% inhibition fluorescence compared to untreated control.y 99% inhibition compared to untreated control.
K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286 283
K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286284
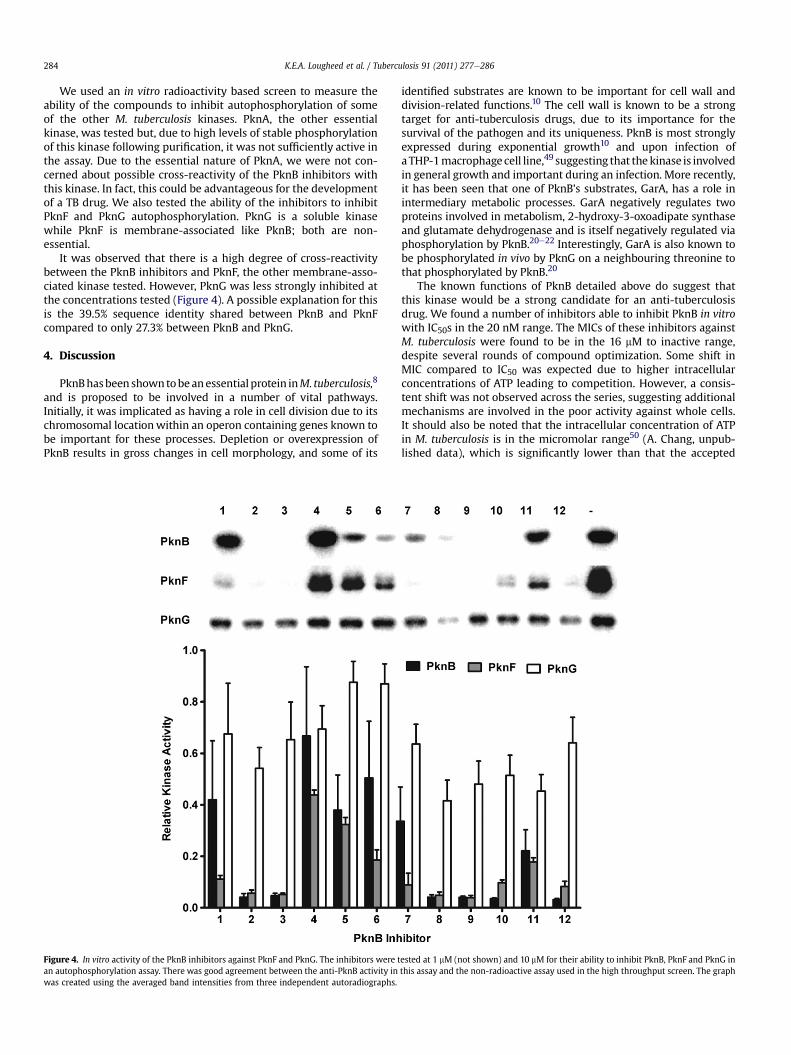
We used an in vitro radioactivity based screen to measure theability of the compounds to inhibit autophosphorylation of someof the other M. tuberculosis kinases. PknA, the other essentialkinase, was tested but, due to high levels of stable phosphorylationof this kinase following purification, it was not sufficiently active inthe assay. Due to the essential nature of PknA, we were not con-cerned about possible cross-reactivity of the PknB inhibitors withthis kinase. In fact, this could be advantageous for the developmentof a TB drug. We also tested the ability of the inhibitors to inhibitPknF and PknG autophosphorylation. PknG is a soluble kinasewhile PknF is membrane-associated like PknB; both are non-essential.
It was observed that there is a high degree of cross-reactivitybetween the PknB inhibitors and PknF, the other membrane-asso-ciated kinase tested. However, PknG was less strongly inhibited atthe concentrations tested (Figure 4). A possible explanation for thisis the 39.5% sequence identity shared between PknB and PknFcompared to only 27.3% between PknB and PknG.
4. Discussion
PknBhasbeen showntobeanessential protein inM. tuberculosis,8
and is proposed to be involved in a number of vital pathways.Initially, it was implicated as having a role in cell division due to itschromosomal locationwithin an operon containing genes known tobe important for these processes. Depletion or overexpression ofPknB results in gross changes in cell morphology, and some of its
Figure 4. In vitro activity of the PknB inhibitors against PknF and PknG. The inhibitors were tan autophosphorylation assay. There was good agreement between the anti-PknB activity inwas created using the averaged band intensities from three independent autoradiographs.
identified substrates are known to be important for cell wall anddivision-related functions.10 The cell wall is known to be a strongtarget for anti-tuberculosis drugs, due to its importance for thesurvival of the pathogen and its uniqueness. PknB is most stronglyexpressed during exponential growth10 and upon infection ofaTHP-1macrophage cell line,49 suggesting that thekinase is involvedin general growth and important during an infection. More recently,it has been seen that one of PknB’s substrates, GarA, has a role inintermediary metabolic processes. GarA negatively regulates twoproteins involved in metabolism, 2-hydroxy-3-oxoadipate synthaseand glutamate dehydrogenase and is itself negatively regulated viaphosphorylation by PknB.20e22 Interestingly, GarA is also known tobe phosphorylated in vivo by PknG on a neighbouring threonine tothat phosphorylated by PknB.20
The known functions of PknB detailed above do suggest thatthis kinase would be a strong candidate for an anti-tuberculosisdrug. We found a number of inhibitors able to inhibit PknB in vitrowith IC50s in the 20 nM range. The MICs of these inhibitors againstM. tuberculosis were found to be in the 16 mM to inactive range,despite several rounds of compound optimization. Some shift inMIC compared to IC50 was expected due to higher intracellularconcentrations of ATP leading to competition. However, a consis-tent shift was not observed across the series, suggesting additionalmechanisms are involved in the poor activity against whole cells.It should also be noted that the intracellular concentration of ATPin M. tuberculosis is in the micromolar range50 (A. Chang, unpub-lished data), which is significantly lower than that the accepted
ested at 1 mM (not shown) and 10 mM for their ability to inhibit PknB, PknF and PknG inthis assay and the non-radioactive assay used in the high throughput screen. The graph

K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286 285
millimolar concentrations for mammalian cells. The poor MIC toIC50 ratios appear to be a similar result to the published data ofother groups who have also attempted to develop PknB or PknGinhibitors.8,12e14 This poor activity was evident even in an intra-cellular assay.
We investigated the role of cell wall permeability, either diffu-sion through the hydrophobic cell wall or diffusion through hydr-ophilic channels, and found no evidence that thiswas a limiting stepin the action of the inhibitors. We also investigated the role of effluxand observed that inhibitor activity was not improved in an effluxpump inhibited model.
A possibility is that the more potent inhibitors, which werechosen for compound optimization, were not targeting PknB in thecells. A similar problem was described by Payne et al.51 in theirreviewofGlaxoSmithKline’s 7 years of antibacterial high throughputscreen (HTS) research. They attributed the antibacterial activity ofsome of their lead compounds to non-specific membrane interac-tions that led to cell lysis. We did attempt to determine the in vivotarget of the inhibitors by generating resistant mutants, howeverthis proved unsuccessful. In a previous study, Fernandez et al. usedM. smegmatis overexpressing PknB to demonstrate that an increasein PknB levels was able to increase resistance to their PknB inhibi-tors.8 However, they were only able to see a modest 2-fold increasein resistance compared to the control strain. Our attempts toperform a similar experiment in M. tuberculosis proved difficult,perhaps becauseM. tuberculosis is more sensitive to overexpressionof PknB than M. smegmatis. Even a modest increase in PknB exp-ressionwas sufficient to slow growth and alter cell morphology, andthe effect of overexpression on inhibitor sensitivity was not signifi-cant (K.E.A. Lougheed, unpublished data).
It is also possible that PknB may not be bound by the inhibitorswhen it is in a physiologically relevant configuration. In other drugdiscovery efforts, it has been shown that small molecule inhibitorsare ineffective when their targets are involved in proteineproteininteractions.52 Such signalling complexes may behave differently incells than in solution perhaps because the binding site is masked byinteracting proteins or because of differences in the in vivo proteinconformation compared to the purified protein against which theinhibitors were screened.
Although pknB is an essential gene, very low levels of activitymay be sufficient for survival, especially under the conditions weused to test our compounds. The M. tuberculosis kinases do displaysome redundancy, at least in the in vitro screens which have beenused to investigate the substrates of the kinases. It is also possiblethat the cross-reactivity of the inhibitors against other kinases orthe binding of alternative proteins results in a sequestering of theavailable inhibitor away from PknB. In fact, our results havedemonstrated that themajority of the inhibitors tested are also ableto inhibit PknF in vitro, although PknGwas not as strongly inhibited.Ideally, a more specific inhibitor would need to be developed thatcould completely inhibit PknB in vivo without its effects beingdiluted by off-target binding.
Our work appears to have highlighted one of the difficultiesassociated with target-based drug discovery: namely, that scre-ening a relatively small library of inhibitors in vitro regularly fails toyield compounds with potent in vivo activity. However, the suit-ability of PknB as a drug target cannot yet be refuted. Our discoveryof inhibitors with good in vitro activity and modest in vivo activitysuggests that further screening on a larger scale may yet yieldpotent inhibitors able to completely inhibit the activity of PknB invivo. To take into account the possibility of redundancy among thekinases, we believe further attempts to develop M. tuberculosiskinase inhibitors might usefully target a broad specificity inhibitortargeting multiple kinases as suggested by Hegymegi-Barakonyiet al.13
Funding: This workwas supported byMRC Technology througha grant from the MRCT Development Gap Fund (A853-0058) and bythe Medical Research Council (U117585867 and U117584228).
Competing interests: None declared.
Ethical approval: Not required.
Acknowledgements
We thank Kristian Birchall for providing Figure 1, CharlesThompson for the gift of the fbpA mutant, Philip Draper for theompA mutant, and Elaine Davis for helpful discussions.
References
1. Global Alliance for TB Drug Development. Tuberculosis. Scientific blueprint fortuberculosis drug development. Tuberculosis (Edinb) 2001;81(Suppl. 1):1e52.
2. Lapenna S, Giordano A. Cell cycle kinases as therapeutic targets for cancer. NatRev Drug Discov 2009;8:547e66.
3. Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, et al. Deciphering thebiology of Mycobacterium tuberculosis from the complete genome sequence.Nature 1998;393:537e44.
4. Av-Gay Y, Everett M. The eukaryotic-like Ser/Thr protein kinases of Mycobac-terium tuberculosis. Trends Microbiol 2000;8:238e44.
5. Narayan A, Sachdeva P, Sharma K, Saini AK, Tyagi AK, Singh Y. Serine threonineprotein kinases of mycobacterial genus: phylogeny to function. Physiol Geno-mics 2007;29:66e75.
6. Molle V, Kremer L. Division and cell envelope regulation by Ser/Thr phos-phorylation: Mycobacterium shows the way. Mol Microbiol 2010;75:1064e77.
7. Eiglmeier K, Parkhill J, Honore N, Garnier T, Tekaia F, Telenti A, et al. Thedecaying genome of Mycobacterium leprae. Lepr Rev 2001;72:387e98.
8. Fernandez P, Saint-Joanis B, Barilone N, Jackson M, Gicquel B, Cole ST, et al. TheSer/Thr protein kinase PknB is essential for sustaining mycobacterial growth.J Bacteriol 2006;188:7778e84.
9. Sassetti CM, Boyd DH, Rubin EJ. Genes required for mycobacterial growthdefined by high density mutagenesis. Mol Microbiol 2003;48:77e84.
10. Kang CM, Abbott DW, Park ST, Dascher CC, Cantley LC, Husson RN. TheMycobacterium tuberculosis serine/threonine kinases PknA and PknB:substrate identification and regulation of cell shape. Genes Dev 2005;19:1692e704.
11. Cowley S, Ko M, Pick N, Chow R, Downing KJ, Gordhan BG, et al. The Myco-bacterium tuberculosis protein serine/threonine kinase PknG is linked tocellular glutamate/glutamine levels and is important for growth in vivo. MolMicrobiol 2004;52:1691e702.
12. Walburger A, Koul A, Ferrari G, Nguyen L, Prescianotto-Baschong C, Huygen K,et al. Protein kinase G from pathogenic mycobacteria promotes survival withinmacrophages. Science 2004;304:1800e4.
13. Hegymegi-Barakonyi B, Szekely R, Varga Z, Kiss R, Borbely G, Nemeth G, et al.Signalling inhibitors against Mycobacterium tuberculosis - early days of a newtherapeutic concept in tuberculosis. Curr Med Chem 2008;15:2760e70.
14. Szekely R, Waczek F, Szabadkai I, Nemeth G, Hegymegi-Barakonyi B, Eros D,et al. A novel drug discovery concept for tuberculosis: inhibition of bacterialand host cell signalling. Immunol Lett 2008;116:225e31.
15. Young TA, Delagoutte B, Endrizzi JA, Falick AM, Alber T. Structure of Myco-bacterium tuberculosis PknB supports a universal activation mechanism for Ser/Thr protein kinases. Nat Struct Biol 2003;10:168e74.
16. Ortiz-Lombardia M, Pompeo F, Boitel B, Alzari PM. Crystal structure of thecatalytic domain of the PknB serine/threonine kinase from Mycobacteriumtuberculosis. J Biol Chem 2003;278:13094e100.
17. Veyron-Churlet R, Zanella-Cleon I, Cohen-Gonsaud M, Molle V, Kremer L.Phosphorylation of the Mycobacterium tuberculosis beta-ketoacyl-acyl carrierprotein reductase MabA regulates mycolic acid biosynthesis. J Biol Chem2010;285:12714e25.
18. Parikh A, Verma SK, Khan S, Prakash B, Nandicoori VK. PknB-mediated phos-phorylation of a novel substrate, N-acetylglucosamine-1-phosphate uridyl-transferase,modulates its acetyltransferase activity. JMol Biol2009;386:451e64.
19. Villarino A, Duran R, Wehenkel A, Fernandez P, England P, Brodin P, et al.Proteomic identification of M. tuberculosis protein kinase substrates: PknBrecruits GarA, a FHA domain-containing protein, through activation loop-mediated interactions. J Mol Biol 2005;350:953e63.
20. O’Hare HM, Duran R, Cervenansky C, Bellinzoni M, Wehenkel AM, Pritsch O,et al. Regulation of glutamate metabolism by protein kinases in mycobacteria.Mol Microbiol 2008;70:1408e23.
21. de Carvalho LP, Zhao H, Dickinson CE, Arango NM, Lima CD, Fischer SM, et al.Activity-based metabolomic profiling of enzymatic function: identification ofRv1248c as a mycobacterial 2-hydroxy-3-oxoadipate synthase. Chem Biol2010;17:323e32.
22. Nott TJ, Kelly G, Stach L, Li J, Westcott S, Patel D, et al. An intramolecular switchregulates phosphoindependent FHA domain interactions in Mycobacteriumtuberculosis. Sci Signal 2009;2. ra12.

K.E.A. Lougheed et al. / Tuberculosis 91 (2011) 277e286286
23. Zhang JH, Chung TD, Oldenburg KR. A simple statistical parameter for use inevaluation and validation of high throughput screening assays. J Biomol Screen1999;4:67e73.
24. Collins L, Franzblau SG. Microplate alamar blue assay versus BACTEC 460system for high-throughput screening of compounds against Mycobacteriumtuberculosis and Mycobacterium avium. Antimicrob Agents Chemother 1997;41:1004e9.
25. Lougheed KE, Taylor DL, Osborne SA, Bryans JS, Buxton RS. New anti-tuber-culosis agents amongst known drugs. Tuberculosis (Edinb) 2009;89:364e70.
26. Bain J, Plater L, Elliott M, Shpiro N, Hastie CJ, McLauchlan H, et al. The selectivityof protein kinase inhibitors: a further update. Biochem J 2007;408:297e315.
27. England P, Wehenkel A, Martins S, Hoos S, Andre-Leroux G, Villarino A, et al.The FHA-containing protein GarA acts as a phosphorylation-dependentmolecular switch in mycobacterial signaling. FEBS Lett 2009;583:301e7.
28. Cheng Y, Prusoff WH. Relationship between the inhibition constant (K1) andthe concentration of inhibitor which causes 50 per cent inhibition (I50) of anenzymatic reaction. Biochem Pharmacol 1973;22:3099e108.
29. Andries K, Verhasselt P, Guillemont J, Gohlmann HW, Neefs JM, Winkler H,et al. A diarylquinoline drug active on the ATP synthase of Mycobacteriumtuberculosis. Science 2005;307:223e7.
30. Brennan PJ, Nikaido H. The envelope of mycobacteria. Annu Rev Biochem1995;64:29e63.
31. Malaviya A, Gomes J. Enhanced biotransformation of sitosterol to androste-nedione by Mycobacterium sp. using cell wall permeabilizing antibiotics. J IndMicrobiol Biotechnol 2008;35:1235e9.
32. Rastogi N, Goh KS, David HL. Enhancement of drug susceptibility of Mycobac-terium avium by inhibitors of cell envelope synthesis. Antimicrob Agents Che-mother 1990;34:759e64.
33. Jagannath C, Reddy VM, Gangadharam PR. Enhancement of drug susceptibilityof multi-drug resistant strains of Mycobacterium tuberculosis by ethambutoland dimethyl sulphoxide. J Antimicrob Chemother 1995;35:381e90.
34. Piddock LJ, Williams KJ, Ricci V. Accumulation of rifampicin by Mycobacteriumaurum, Mycobacterium smegmatis and Mycobacterium tuberculosis. J AntimicrobChemother 2000;45:159e65.
35. Nguyen L, Chinnapapagari S, Thompson CJ. FbpA-Dependent biosynthesis oftrehalose dimycolate is required for the intrinsic multidrug resistance, cell wallstructure, and colonial morphology of Mycobacterium smegmatis. J Bacteriol2005;187:6603e11.
36. LambertPA.Cellular impermeabilityanduptakeofbiocidesandantibiotics inGram-positive bacteria and mycobacteria. J Appl Microbiol 2002;92(Suppl.):46Se54S.
37. Stephan J, Mailaender C, Etienne G, Daffe M, Niederweis M. Multidrug resis-tance of a porin deletion mutant of Mycobacterium smegmatis. AntimicrobAgents Chemother 2004;48:4163e70.
38. Mailaender C, Reiling N, Engelhardt H, Bossmann S, Ehlers S, Niederweis M. TheMspA porin promotes growth and increases antibiotic susceptibility of both
Mycobacterium bovis BCG and Mycobacterium tuberculosis. Microbiology2004;150:853e64.
39. Siroy A, Mailaender C, Harder D, Koerber S, Wolschendorf F, Danilchanka O,et al. Rv1698 of Mycobacterium tuberculosis represents a new class of channel-forming outer membrane proteins. J Biol Chem 2008;283:17827e37.
40. Raynaud C, Laneelle MA, Senaratne RH, Draper P, Laneelle G, Daffe M. Mech-anisms of pyrazinamide resistance in mycobacteria: importance of lack ofuptake in addition to lack of pyrazinamidase activity. Microbiology 1999;145:1359e67.
41. Takiff HE, Cimino M, Musso MC, Weisbrod T, Martinez R, Delgado MB, et al.Efflux pump of the proton antiporter family confers low-level fluoroquinoloneresistance in Mycobacterium smegmatis. Proc Natl Acad Sci USA 1996;93:362e6.
42. Gupta AK, Chauhan DS, Srivastava K, Das R, Batra S, Mittal M, et al. Estimation ofeffluxmediated multi-drug resistance and its correlation with expression levelsof two major efflux pumps in mycobacteria. J Commun Dis 2006;38:246e54.
43. Escribano I, Rodriguez JC, Llorca B, Garcia-Pachon E, Ruiz M, Royo G. Impor-tance of the efflux pump systems in the resistance of Mycobacterium tubercu-losis to fluoroquinolones and linezolid. Chemotherapy 2007;53:397e401.
44. Srivastava S, Musuka S, Sherman C, Meek C, Leff R, Gumbo T. Efflux-pump-derived multiple drug resistance to ethambutol monotherapy in Mycobacte-rium tuberculosis and the pharmacokinetics and pharmacodynamics ofethambutol. J Infect Dis 2010;201:1225e31.
45. Amaral L, Martins M, Viveiros M. Enhanced killing of intracellular multidrug-resistant Mycobacterium tuberculosis by compounds that affect the activity ofefflux pumps. J Antimicrob Chemother 2007;59:1237e46.
46. Ramon-Garcia S, Martin C, De Rossi E, Ainsa JA. Contribution of the Rv2333cefflux pump (the Stp protein) from Mycobacterium tuberculosis to intrinsicantibiotic resistance in Mycobacterium bovis BCG. J Antimicrob Chemother2007;59:544e7.
47. Ramon-Garcia S, Martin C, Thompson CJ, Ainsa JA. Role of the Mycobacteriumtuberculosis P55 efflux pump in intrinsic drug resistance, oxidative stressresponses, and growth. Antimicrob Agents Chemother 2009;53:3675e82.
48. Li XZ, Zhang L, Nikaido H. Efflux pump-mediated intrinsic drug resistance inMycobacterium smegmatis. Antimicrob Agents Chemother 2004;48:2415e23.
49. Singh A, Singh Y, Pine R, Shi L, Chandra R, Drlica K. Protein kinase I of Myco-bacterium tuberculosis: cellular localization and expression during infection ofmacrophage-like cells. Tuberculosis (Edinb) 2006;86:28e33.
50. Koul A, Vranckx L, Dendouga N, Balemans W, Van den Wyngaert I,Vergauwen K, et al. Diarylquinolines are bactericidal for dormant mycobacteriaas a result of disturbed ATP homeostasis. J Biol Chem 2008;283:25273e80.
51. Payne DJ, Gwynn MN, Holmes DJ, Pompliano DL. Drugs for bad bugs: con-fronting the challenges of antibacterial discovery. Nat Rev Drug Discov2007;6:29e40.
52. Hoshi N, Langeberg LK, Gould CM, Newton AC, Scott JD. Interaction withAKAP79 modifies the cellular pharmacology of PKC. Mol Cell 2010;37:541e50.





























