Embed Size (px)
Citation preview

Effect of drought and waterlogging stress on needle monoterpenes of Picea abies
P. KAINULAINEN, J. OKSANEN, V. PALOMAKI, J. K. HOLOPAINEN, AND T. HOLOPAINEN Ecological Lnboratory, Department of Environmental Sciences, University of Kuopio,
P.O. Box 1627, SF-70211 Kuopio, Finland
Received December 12, 1991
KAINULAINEN, P., OKSANEN, J., PALOMAKI, V., HOLOPAINEN, J. K., and HOLOPAINEN, T. 1992. Effect of drought and water- logging stress on needle monoterpenes of Picea abies. Can. J. Bot. 70: 1613- 1616.
Monoterpenes were analysed in needles of clonal Picea abies (L.) Karst. exposed to drought and waterlogging. In drought- stressed trees the amounts of tricyclene, a-pinene, and camphene were significantly (p <0.05) higher than in the control trees. In the drought treatment the total terpene concentration was also significantly ( p <0.05) higher. Waterlogging did not have significant effects on individual terpenes and total terpene concentration. However, multivariate redundancy analysis indicated that the effect of treatments on all the compounds simultaneously was not significant ( p = 0.12). These results are discussed in terms of their potential impact on preferential selection of host plant by insects.
Key words: drought, needles, Picea abies, terpenes, waterlogging.
KAINULA~NEN, P., OKSANEN, J., PALOMAKI, V., HOLOPAINEN, J. K., et HOLOPAINEN, T. 1992. Effect of drought and water- logging stress on needle monoterpenes of Picea abies. Can. J . Bot. 70 : 16 13 - 1616.
Les auteurs ont analysC le contenu en monoterpknes des aiguilles de Picea abies (L.) Karst. clonaux, exposCs B la sCcheresse et B l'excks d'eau. Chez les arbres soumis au stress hydrique, les quantitCs de tricyclbne, d'a-pinbne et de camphbne sont significativement plus ClevCes ( p <0,05). Dans le traitement par la skcheresse, la concentration totale en terpknes est Cgale- ment significativement plus ClevCe ( p <0,05). L'excks d'eau n'a pas d'effets significatifs sur les terpbnes individuels et la concentration totale en terpknes. Cependant, l'analyse multivarite par redondance indique que l'effet des traitements sur tous les composCs pris simultanCment, n'est pas significatif ( p = 0,12). Ces rksultats sont discutCs quant B leur impact potentiel sur la sClection prCfCrentielle de la plante hate par les insectes.
Mots clPs : skcheresse, aiguilles, Picea abies, terpknes, excbs d'eau.
Introduction Monoterpenes are the main components of the essential oils
of coniferous trees and are found in concentrations of several milligrams per gram of needle fresh weight. Monoterpenes mediate many important interactions between trees and their environment. They can control the growth of fungi (Melin and Krupa 197 1 ; Russell and Berrymann 1976; Thibault-Balesdent and Delatour 1985), and they can also repel (Raffa et al. 1985; Hrutfiord and Gara 1989; Bryant et al. 1991) or attract (Francke and Vite 1983; Ishikawa et al. 1986) herbivores.
The composition of monoterpenes in conifers varies widely and is dependent on plant genotype (Schonwitz et al. 1987, 1989, 1990a), environment (Muzika et al. 1989; Heller et al. 1990), and plant-specific factors (Gleizes et al. 1980; Merk et al. 1988; Schonwitz et al. 1990b). Concentrations of needle terpenes are significantly affected by age and position of the whorl, whereas position on the branch is of minor importance (Merk et al. 1988). The hydrocarbon terpenes dominate in spruce needles in the first weeks after bud burst, and the oxygenated terpenes develop later (Schonwitz et al. 1990b). Schonwitz et al. (1990b) suggested that the terpene levels of older needles are influenced by both biosynthetic and catabolic activities.
Terpene composition appears to be under genetic control, implying that individual trees will have distinctive leaf oil compositions. A study of leaf monoterpenes in an 80-year-old Norway spruce population indicated significant variations in the concentrations of different terpenes among the investigated trees (Schonwitz et al. 1987). However, variation of terpenes within clones of Norway spruce has been found to be fairly small (Merk et al. 1988).
The effects of season (Schonwitz et al. 1990b), temperature and light (Lincoln and Langenheim 1978), mineral nutrition (Muzika et al. 1989), and air pollutants (Heller et al. 1990) on Printed in Canada / lrnprtmC nu Canada
the terpene content of conifer needles have been investigated, but information on the influence of water availability on mono- terpenes of needles of coniferous trees is limited (Major 1990). The present study was conducted to assess the influence of drought and waterlogging stress on the quantitative terpene composition of the Norway spruce needles.
Material and methods Plant material
One clone of Norway spruce (Picea abies (L.) Karst.) originating from central Finland (provenance E2089 Langelmaki x K953 Piek- samaki) was obtained from the Pieksamaki Nursery of the Foundation for Tree Breeding in Finland. Five-year-old cuttings were planted in the previous spring in 18-L plastic containers, two cuttings per con- tainer, in a 1:2 mixture of sand and fertilized peat (liming 3 kg/m3, fertilization 1.25 kg/m3 of 15:5: I1 (N -P-K) fertilizer with micro- nutrients). During the previous summer and autumn the plants were grown in the botanic garden of the Kuopio University, Finland, receiving natural rainfall. They were moved into the greenhouse in early January and kept at a temperature of +5"C for 1 week. The plants were then transferred to a greenhouse with a 22 h light : 2 h dark cycle at a temperature of 23:16"C (light:dark), which cor- responds to the natural light and temperature cycle in June in central Finland. The supplemental light (about 60 pmol m-2 s-') was given with multimetal lamps. The relative humidity was about 40% in the light and 70% in the dark.
The plants were given sufficient water (three times a week) during the 7-week growing period and then they were randomized in three treatments (control, drought, waterlogging). Waterlogging stress was induced by placing the containers in a water bath, and drought- stressed plants were not watered at all for 8 weeks. The development of drought and waterlogging was monitored by measuring the water content of the needles during the experiment. The changes in the ultrastructure of needles were followed during the experiment. In the drought treatment 50- 100% of the mesophyll and transfusion paren- chyma cells were plasmolysed, but in the waterlogging treatment
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
nive
rsity
of
Ten
ness
ee o
n 08
/24/
13Fo
r pe
rson
al u
se o
nly.

1614 CAN. I. BOT. VOL. 70, 1992
TABLE 1. Elution times (min) and relative reten- tion times (RRT) of the identified terpenes on a
30-m long DP-5 capillary column
Compound Elution time RRT
Tricyclene a-Pinene Camphene Sabinene /3-Pinene Myrcene a-Phellandrene 3-Carene a-Terpinene p-Cymene Limonene 1,8-Cineole Unknown CIOHI6 1-Chlorooctane Unknown C,,H,, Linalool Camphor Camphene hydrate Borneo1 Terpinen-4-01 a-Terpineol Piperitone Bornyl acetate a-Copaene Unknown CI5H2, Longifolene /3-Caryophy Ilene a-Humulene Unknown CI5H2, Unknown C,,H,, Unknown C15H2,
plasmolysed cells were only found occasionally. The detailed results of the ultrastructural study will be published separately.
Sarnpling procedure Shoots from the previous year were collected from six plants of
each treatment 8 weeks after the beginning of the treatment. In the laboratory, the needles (300 mg) were immediately cut into small pieces (about 1 mm long) in 2 mL of n-hexane and extracted for 15 min. 1-Chlorooctane was used as an internal standard. The extrac- tion method was tested, and the coefficient variation of individual compounds in replicate extractions was an average 5 .7%. The ana- lytical error was less than 5 % for most of the analysed terpenes. This method gave the same terpene pattern as the analysis of pulverized frozen needles, although terpene concentrations were lower. Without further purification, the extracts were analysed by gas chromatog- raphy - mass spectrometry (GC -MS).
Identijkation and quantijkation of terpenes A Hewlett Packard 5890 gas chromatograph equipped with a mass
selective detector (type 5970) was used. Separations were carried out on a 30-m DP-5 (i.d. 0.25 mm; film thickness 0.25 pm; J & W Scien- tific, Folsom, Calif.) capillary column. Helium was used as carrier gas, and the linear velocity was about 30 cm s-I. The splitless (purge time off 0.5 min) sampling technique was used and 1 pL was injected. The column temperature was programmed from 50 to 250°C at 5"CImin. The electron ionizing energy was 70 eV. Mass numbers from m/z 30 to 300 were recorded, and the peak areas were used to quantify the individual substances.
The individual terpene compounds were identified by their mass spectra and retention times (Table 1). The retention times were compared with those of the pure substances and with the values given
in the literature (Jiittner 1988). For quantification, 20 available standards (purchased from Fluka AG, Sigma Chemical Co., St. Louis, Mo., and Aldrich-Chemie GmbH & Co. KG, Steinheim, Germany) were used. Quantification was based on injecting known amounts of authentic compounds under identical conditions and determining response factors for each terpene relative to known amounts of the internal standard, 1-chlorooctane. The results are expressed as micro- grams per gram of needle fresh weight.
Statistical analyses The effect of treatments on quantities of terpenes was assessed
separately for each compound by analysis of variance. Untrans- formed concentrations per ffesh weight were used in all analyses. The significance of differences against the control was assessed by Duncan's multiple range test. Analysis of the effect of treatments on all the components simultaneously was studied by redundancy analy- sis (Jongman et al. 1987), which was performed by the program CANOCO (ter Braak 1987). To give equal weight to all the compounds, redundancy analysis was based on correlations between the com- pounds. Statistical significance of the separation was estimated by a Monte Carlo procedure (ter Braak 1987) using 999 replicates.
Results
The most abundant compounds were a-pinene, camphene, myrcene, limonene, and bornyl acetate. The rest consisted mainly of tricyclene, sabinene, P-pinene, 1,8-cineole, camphor, camphene hydrate, borneol, and a-terpineol. Small amounts of compounds such as a-phellandrene, 3-carene, a-terpinene, p-cymene, linalool, terpinen4-01, a-terpineol, and some uniden- tified monoterpenes were also found. The most abundant sesquiterpenes were P-caryophyllene and a-humulene. Small amounts of a-copaene, longifolene, and some unidentified sesquiterpenes were also found.
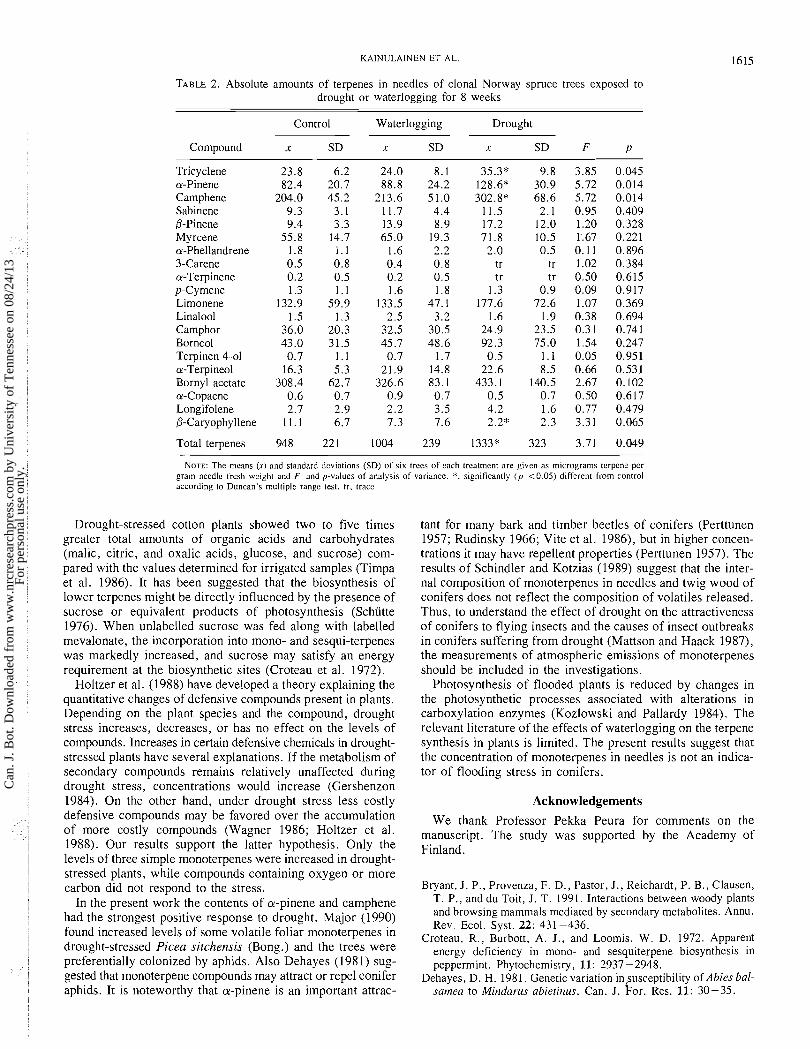
Two-way ANOVA indicated that drought treatment had a sig- nificant ( p = 0.024) effect on total terpene content, but the effect of waterlogging was not significant ( p = 0.721). Under drought stress the amounts of tricyclene, a-pinene, and cam- phene were significantly higher ( p <0.05) than in control trees (Table 2). The water contents of the needles were 5 1.7 % (SD 3.1), 52.7% (SD 4.0), 46.4% (SD 7.5) in the control, waterlogging, and drought treatments, respectively, but these values did not differ from each other significantly.
The compounds that were indicated by univariate ANOVA to have significantly different levels in treatments were also the most extreme ones in the multivariate redundancy analysis. The effect of treatments was masked by the within-clone varia- tion and the effect was not significant ( p = 0.12) for indi- vidual compounds simultaneously. However, the variation seemed to be mainly unidimensional, since the first axis was by far the most important.
Discussion
Few studies have been made on the effects of water avail- ability on monoterpene composition of plants. Gershenzon et al. (1978) observed a small decrease in the percentage of monoterpenoids with a p-menthane carbon skeleton and a reduction of leaf weight and monoterpene yield per leaf in the herb Satureja douglasii under water shortage. Increased a - and P-pinene (Hodges and Lorio 1975) and increased a-pinene associated with decreased P-pinene, myrcene, and limonene (Gilmore 1977) are the most typical changes in monoterpene composition in xylem oleoresin of moisture-stressed Pinus taeda. Camphene increased with water shortage in one test year but decreased in stressed tr,ees during the other test year (Gilmore 1977).
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
nive
rsity
of
Ten
ness
ee o
n 08
/24/
13Fo
r pe
rson
al u
se o
nly.
KAINULAINEN ET AL.
TABLE 2. Absolute amounts of terpenes in needles of clonal Norway spruce trees exposed to drought or waterlogging for 8 weeks
Control Waterlogging Drought
Compound x SD x SD x SD F P
Tricyclene a-Pinene Camphene Sabinene /3-Pinene Myrcene a-Phellandrene 3-Carene a-Terpinene p-Cy mene Limonene Linalool Camphor Borneo1 Terpinen-4-01 a-Terpineol Bornyl acetate a-Copaene Longifolene /3-Caryophy llene
Total terpenes
NOTE: The means (1.) and standard deviations (SD) of six trees of each treatment are given as micrograms terpcnc per gram needle fresh weight and F- and p-values of analysis of variance. *, significantly ( p <0.05) different from control according to Duncan's multiple range test. tr. trace.
Drought-stressed cotton plants showed two to five times greater total amounts of organic acids and carbohydrates (malic, citric, and oxalic acids, glucose, and sucrose) com-
, pared with the values determined for irrigated samples (Timpa I et al. 1986). It has been suggested that the biosynthesis of
lower terpenes might be directly influenced by the presence of sucrose or equivalent products of photosynthesis (Schiitte 1976). When unlabelled sucrose was fed along with labelled mevalonate, the incorporation into mono- and sesqui-terpenes was markedly increased, and sucrose may satisfy an energy requirement at the biosynthetic sites (Croteau et al. 1972).
Holtzer et al. (1988) have developed a theory explaining the quantitative changes of defensive compounds present in plants. Depending on the plant species and the compound, drought stress increases, decreases, or has no effect on the levels of compounds. Increases in certain defensive chemicals in drought- stressed plants have several explanations. If the metabolism of secondary compounds remains relatively unaffected during drought stress, concentrations would increase (Gershenzon 1984). On the other hand, under drought stress less costly defensive compounds may be favored over the accumulation
' of more costly compounds (Wagner 1986; Holtzer et al. I 1988). Our results support the latter hypothesis. Only the
levels of three simple monoterpenes were increased in drought- stressed plants, while compounds containing oxygen or more
I carbon did not respond to the stress.
I In the present work the contents of a-pinene and camphene had the strongest positive response to drought. Major (1990) found increased levels of some volatile foliar monoterDenes in drought-stressed Picea sitchensis (Bong.) and the trees were preferentially colonized by aphids. Also Dehayes (1981) sug- gested that monoterpene compounds may attract or repel conifer aphids. It is noteworthy that a-pinene is an important attrac-
tant for many bark and timber beetles of conifers (Perttunen 1957; Rudinsky 1966; Vite et al. 1986), but in higher concen- trations it may have repellent properties (Perttunen 1957). The results of Schindler and Kotzias (1989) suggest that the inter- nal composition of monoterpenes in needles and twig wood of conifers does not reflect the composition of volatiles released. Thus, to understand the effect of drought on the attractiveness of conifers to flying insects and the causes of insect outbreaks in conifers suffering from drought (Mattson and Haack 1987), the measurements of atmospheric emissions of monoterpenes should be included in the investigations.
Photosynthesis of flooded plants is reduced by changes in the photosynthetic processes associated with alterations in carboxylation enzymes (Kozlowski and Pallardy 1984). The relevant literature of the effects of waterlogging on the terpene synthesis in plants is limited. The present results suggest that the concentration of monoterpenes in needles is not an indica- tor of flooding stress in conifers.
Acknowledgements We thank Professor Pekka Peura for comments on the
manuscript. The study was supported by the Academy of Finland.
Bryant, J . P., Provenza, F. D., Pastor, J. , Reichardt, P. B., Clausen, T. P., and du Toit, J. T. 1991. Interactions between woody plants and browsing mammals mediated by secondary metabolites. Annu. Rev. Ecol. Syst. 22: 43 1 -436.
Croteau, R., Burbott, A. J. , and Loomis, W. D. 1972. Apparent energy deficiency in mono- and sesquiterpene biosynthesis in peppermint. Phytochemistry , 11: 2937 -2948.
Dehayes, D. H. 1981. Genetic variation ingusceptibility of Abies bal- sarnea to Mindarus abietirlus. Can. J . For. Res. 11: 30-35.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
nive
rsity
of
Ten
ness
ee o
n 08
/24/
13Fo
r pe
rson
al u
se o
nly.

1616 CAN. J . BOT. VOL. 70. 1992
Francke, W., and Vite, J. 1983. Oxygenated terpenes in pheromone systems of bark beetles. Z. Angew. Entomol. 96: 146-156.
Gershenzon, J. 1984. Changes in the levels of plant secondary metabolites under water and nutrient stress. Recent Adv. Phytochem. 18: 273 -320.
Gershenzon, J., Lincoln, D. E., and Langenheim, J. H. 1978. The effect of moisture stress on monoterpenoid yield and composition in Satureja douglasii. Biochem. Syst. Ecol. 6: 33 -43.
Gilmore, A. R. 1977. Effects of soil moisture stress on monoterpenes in loblolly pine. J. Chem. Ecol. 3: 667 -676.
Gleizes, M., Pauly, G., Bernard-Dagan, C., and Jaqueus, R. 1980. Effects of light on terpene synthesis in Pinus pinaster. Physiol. Plant. 50: 16-20.
Heller, W., Rosemann, D., Osswald, W. F., Benz, B., Schonwitz, R., Lohwasser, K., Kloos, M., and Sandermann, H., Jr. 1990. Biochemical response of Norway spruce (Picea abies (L.) Karst.) towards 14-month exposure to ozone and acid mist. I . Effects of polyphenol and monoterpene metabolism. Environ. Pollut. 64: 353 -366.
Hodges, J. D., and Lorio, P. L. 1975. Moisture stress and composi- tion of xylem oleoresin in loblolly pine. For. Sci. 21: 283-290.
Holtzer, T. O., Archer, T. L., and Norman, J . M. 1988. Host plant suitability in relation to water stress. In Plant stress-interactions. Edited by E. A. Heinrichs. Wiley-Interscience, New York. pp. 111-137.
Hrutfiord, B., and Gara, R. 1989. The terpene complement of slow and fast growing Sitka spruce terminals as related to Pissodes strobi (Peck) (Col., Curculionidae) host selection. J. Appl. Entomol. 108: 21 -23.
Ishikawa, M., Shito, Y., and Watanabe, H. 1986. 0-Myrcene, a potent attractant component of pine wood for the pine wood nematode, Bursaphelench~ts xylophilus. Agric. Biol. Chem. 50: 1863 - 1866.
Jongman, R. H. G., ter Braak, C. J . F., and van Tongeren, 0. F. R. 1987. Data analysis in community and landscape ecology. Pudoc, Wageningen.
Juttner, F. 1988. Changes of monoterpene concentrations in needles of pollution-injured Picea abies exhibiting montane yellowing. Physiol. Plant. 72: 48-56.
Kozlowski, T. T., and Pallardy, S. G. 1984. Effect of flooding on water, carbohydrate and mineral relations. In Flooding and plant growth. Edited by T. T. Kozlowski. Academic Press, Orlando. pp. 165-193.
Lincoln, D. E., and Langenheim, J. H. 1978. Effect of light and tem- perature on monoterpenoid yield and composition in Satureja douglasii. Biochem. System. Ecol. 6: 21 -32.
Major, E. J. 1990. Water stress in Sitka spruce and its effect on the green spruce aphid Elatobiurn abietinum. In Population dynamics of forest insects. Edited by A. D. Watt, S. R. Leather, M. D. Hunter, and N. A. C. Kidd. Intercept Ltd., Dorset, U.K. pp. 85-93.
Mattson, W. J., and Haack, R. A. 1987. The role of drought stress in provoking outbreaks of phytophagous insects. In Insect out- breaks. Edited by P. Barbosa and J. C. Schultz. Academic Press, San Diego.
Melin, E., and Krupa, S. 1971. Studies on ectomycorrhizae on pine. 11. Growth inhibition of mycorrhizal fungi by volatile organic con- stituents of Pinus silvestris (Scots pine) roots. Physiol. Plant. 25: 337-340.
Merk, L., Kloos, M., Schonwitz, R., and Ziegler, H. 1988. Influence of various factors on quantitative composition of leaf monoterpenes of Picea abies (L.) Karst. Trees, 2: 45 -51.
Muzika, R. M., Pregitzer, K. S., and Hanover, J. W. 1989. Changes in terpene production following nitrogen fertilization of grand fir (Abies grandis (Dougl.) Lindl.) seedlings. Oecologia, 80: 485 -489.
Perttunen, V. 1957. Reactions of two bark beetle species, Hylurgops palliatus Gyll. and Hylastes ater payk. (Col. Scotylidae) to the terpene a-pinene. Ann. Entomol. Fenn. 23: 101 -1 10.
Raffa, K. F., Berryman, A. A., Simasko, J., Teal, W., and Wong, B. L. 1985. Effects of grand fir monoterpenes on fir engaver, Scolytus ventralis (Coleoptera: Scolytidae), and its symbiotic fungus. Environ. Entomol. 14: 552-556.
Rudinsky, J. A. 1966. Scotylid beetles associated with Douglas fir: response to terpenes. Science (Washington, D.C.), 152: 218-219.
Russell, C. E., and Berryman, A. 1976. Host resistance to the fir engaver beetle. 1. Monoterpene composition of Abies grandis pitch blisters and fungus-infected wounds. Can. J . Bot. 54: 14-18.
Schindler, T., and Kotzias, D. 1989. Comparison monoterpene volatilization and leaf-oil composition of conifers. Naturwissen- schaften, 76: 475 -476.
Schutte, H.-R. 1976. Secondary plant substances. Monoterpenes. Prog. Bot. 38: 129-147.
Schonwitz, R., Merk, L., and Ziegler, H. 1987. Naturally occurring monoterpenoids in needles of Picea abies (L.) Karst. Trees, 1: 88-93.
Schonwitz, R., Merk, L., and Ziegler, H. 1989. Variability in the monoterpenes of needles of Picea abies (L.) Karst. Flavour Fragrance J. 4: 149 - 153.
Schonwitz, R., Kloos, M., Merk, L., and Ziegler, H. 1990a. Patterns of monoterpenes stored in the needles of Picea abies (L.) Karst. from several locations in mountainous regions of southern Germany. Trees, 4: 27-33.
Schonwitz, R., Lohwasser, K., Kloos, M., and Ziegler, H. 19906. Seasonal variation in the monoterpenes in needles of Picea abies (L.) Karst. Trees, 4: 34-40.
ter Braak, C. J. F. 1987. CANOCO-a FORTRAN program for canonical community ordinations by [partial] [detrended] [canonical] cor- respondence analysis, principal components analysis and redun- dancy analysis (version 2.1). TNO Institute of Applied Computer Science, Wageningen.
Thibault-Balesdent, M., and Delatour, C. 1985. Variability of Heterobasidium annosurn behaviour to three monoterpenes. Eur. J. For. Pathol. 15: 301 -302.
Timpa, J. D., Burke, J. J., Quisenberry, J. E., and Wendt, C. W. 1986. Effects of water stress on the organic acid and carbohydrate compositions of cotton plants. Plant Physiol. 82: 724-728.
Vite, J. P., Volz, H. A,, Paiva, M. R. , and Bakke, A. 1986. Semio- chemicals in host selection and colonization of pine trees by the pine shoot beetle Totlzicus piniperda. Naturwissenschaften, 73: 39-40.
Wagner, M. R. 1986. Influence of moisture stress and induced resis- tance in ponderosa pine, Pinusponderosa Dougl. ex. Laws, on the pine sawly, Neodiprion autumnalis Smith. For. Ecol. Manage. 15: 43-53.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
nive
rsity
of
Ten
ness
ee o
n 08
/24/
13Fo
r pe
rson
al u
se o
nly.