Embed Size (px)
Citation preview

B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
Effect of caffeine on the expression of cytochrome P450 1A2,adenosine A2A receptor and dopamine transporter in controland 1-methyl 4-phenyl 1, 2, 3, 6-tetrahydropyridine treatedmouse striatum
Seema Singha,1, Kavita Singha,1, Satya Prakash Guptaa, Devendra Kumar Patela,Vinod Kumar Singhb, Raj Kumar Singhb, Mahendra Pratap Singha,⁎aIndian Institute of Toxicology Research, (Council of Scientific and Industrial Research), Mahatma Gandhi Marg, Post Box-80,Lucknow-226 001, IndiabChhatrapati Shahuji Maharaj Medical University, Lucknow-226 003, India
A R T I C L E I N F O
⁎ Corresponding author. Fax: +91 522 2628227E-mail address: singhmahendrapratap@r
1 Equal contribution.
0006-8993/$ – see front matter © 2009 Elsevidoi:10.1016/j.brainres.2009.06.002
A B S T R A C T
Article history:Accepted 2 June 2009Available online 9 June 2009
Parkinson's disease (PD) is a progressive neurodegenerative disorder, characterized by theselective loss of dopaminergic neurons of the nigrostriatal pathway. Epidemiological studieshave shown an inverse relationship between coffee consumption and susceptibility to PD.Cytochrome P450 1A2 (CYP1A2) is involved in caffeine metabolism and its clearance.Caffeine, on the other hand, antagonizes adenosine A2A receptor and regulates dopaminesignaling through dopamine transporter (DAT). The present study was undertaken toinvestigate the expression of CYP1A2, adenosine A2A receptor and DAT in mouse striatumand to assess their levels in 1-methyl 4-phenyl 1, 2, 3, 6-tetrahydropryridine (MPTP) treatedmouse striatum with and without caffeine treatment. The animals were treatedintraperitoneally daily with caffeine (20 mg/kg) for 8 weeks, followed by MPTP (20 mg/kg)+caffeine (20 mg/kg) for 4 weeks or vice versa, along with respective controls. Tyrosinehydroxylase immunoreactivity, levels of dopamine and 1-methyl 4-phenylpyridinium ion(MPP+), expressions of CYP1A2, adenosine A2A receptor and DAT and CYP1A2 catalyticactivity were measured in control and treated mouse brain. Caffeine partially protectedMPTP-induced neurodegenerative changes and modulated MPTP-mediated alterations inthe expression and catalytic activity of CYP1A2, expression of adenosine A2A receptor andDAT. The results demonstrate that caffeine alters the striatal CYP1A2, adenosine A2A
receptor and DAT expressions in mice exposed to MPTP.© 2009 Elsevier B.V. All rights reserved.
Keywords:Parkinson's disease1-Methyl 4-phenyl 1, 2, 3, 6-tetrahydropyridineCytochrome P450 1A2CaffeineAdenosine A2A receptorDopamine transporter
1. Introduction
The neurodegeneration in Parkinson's disease (PD) targetsdopaminergic neurons of the nigrostriatal pathway. Dopami-
.ediffmail.com (M.P. Singh
er B.V. All rights reserved
nergic neurodegeneration causes dopamine depletion in thestriatum leading to motor disturbances and onset of restingtremor, rigidity, postural instability and bradykinesia, themajor hallmarks of PD. Aging, genetic factors and environ-
).
.

Fig. 1 – Dopamine content in the striatum (ng/mg of tissue) ofcaffeine treated animals (a) and effect of CYP1A2 inhibitorciprofloxacin on dopamine content in the pre-treatmentgroup (b). The data are expressed as means±S.E.M. (n=3–5separate experiments). Significant changes **(P<0.01),***(P<0.001) are expressed in comparison with control,#(P<0.05), ###(P<0.001) with caffeine, $$(P<0.01),$$$(P<0.001) with MPTP and τ(P<0.05) with caffeine+MPTP.
116 B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
mental exposure to pesticides and heavymetals are themajorcontributors to PD (Tanner et al., 1999; Vieregge et al., 1999;Wirdefeldt et al., 2004). 1-methyl 4-phenyl 1, 2, 3, 6 tetrahy-dropyridine (MPTP) causes selective degeneration of dopami-nergic neurons and is regarded as one of the best rodentmodels to understand the biochemical and molecular eventsleading to PD and also to assess the efficacy of anti-PD drugs.MPTP produces several biochemical, molecular and phenoty-pic symptoms that mimic the sporadic PD (Przedborski et al.,2000; Gerlach and Riederer, 1996). Current therapies alleviatethe symptoms of PD and offer only symptomatic relief to thepatients. Despite delaying neurodegeneration, the availabletherapies exert several adverse effects and therefore attemptsare consistently being made to develop better therapies withminimal side effects.
Caffeine is one of the most widely consumed psychosti-mulants and dietary components, with an average consump-tion of about 200–250 mg/day/person, as a standard cup ofcoffee contains 100 mg of caffeine (Fredholm, 2004). Bothretrospective and prospective epidemiological studies havelinked caffeine consumption to reduced PD risk and postu-lated that the neuroprotective effect of caffeine could be due toits ability to antagonize adenosine A2A receptor (Ascherio etal., 2001). Caffeine (5–30 mg/kg) exposure to animals corre-sponding to the typical human exposure also resists MPTP-induced dopamine depletion in a dose dependent manner(Chen et al., 2001). Although caffeine non-selectively antag-onizes adenosine A1, A2A, A2B and A3 receptors, only A2A
receptor is abundant in the striatum (Svenningsson andFredholm, 2003). Adenosine A2A receptor modulates γ-aminobutyric acid, acetylcholine and glutamate-mediated neuro-transmissions (Kurokawa et al., 1996; Mori et al., 1996;Richardson et al., 1997; Ochi et al., 2000; Fuxe et al., 1998;Canals et al., 2003; Ciruela et al., 2004). Caffeine-mediatedneuroprotection could be partially contributed by its ability toinhibit the blood brain barrier dysfunction in MPTP-treatedmouse (Chen et al., 2008). Caffeine also prevents apoptotic celldeath by the activation of phosphoinositide 3-kinase orserine-threonine protein kinase and Akt pathways (Nakasoet al., 2008). As caffeine alters dopamine signaling andreceptor affinity in the striatum by stimulating dopaminergicresponses, therefore, blockade of adenosine A2A receptorsprobably direct anti-PD effects (Ferre et al., 1992). Caffeinestimulates potassium channel opening, which preventsneurons from depolarization and neurotransmitter releaseand inhibits excitotoxicity by altering neuronal metabolism(Mao et al., 2007; Avshalumov et al., 2005; Jones, 2008). Caffeineis metabolized mainly by CYP1A2 in the liver and increasedCYP1A2 activity is associated with habitual caffeine consump-tion (Carrillo and Benitez, 1996). Dopamine transporter (DAT)is expressed in the dopaminergic neurons and clears thedopamine released into the extra-cellular spaces, therebyregulating the dopamine signaling (Kurosaki et al., 2003).Although CYP1A2, adenosine A2A receptor and dopaminetransporter are critical in PD, their roles in MPTP-mediateddopaminergic neurodegeneration and caffeine-mediated neu-roprotection are not established. The present study wasperformed to investigate the expression of CYP1A2, adenosineA2A receptor and dopamine transporter inmouse striatumandto assess their roles therein.
2. Results
2.1. Dopamine content
MPTP treatment for 2–4 weeks produced a significantreduction in the striatal dopamine content (Fig. 1a). Thedecrease in dopamine level was significantly less both incaffeine pre-and post-treated animals. Treatment with cipro-floxacin, an inhibitor of CYP1A2, enhanced MPTP-mediateddepletion of dopamine contents in the pre-treatment group(Fig. 1b).
2.2. Dopaminergic neurodegeneration
Caffeine treatment did not produce any change in tyrosinehydroxylase (TH) immunoreactivity; however, MPTP

117B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
treatment reduced the TH immunoreactivity. MPTP-mediated reduction was significantly restored in theanimals pre- or post-treated with caffeine (Fig. 2). Cipro-floxacin treatment enhanced MPTP-mediated loss of THimmunoreactivity.
Fig. 2 – The representative photomicrographs showing TH immun(a) and the number of TH+ positive neurons in the substantia nigraP are caffeine, C, H, M and Q are MPTP, D, I, N and R are caffeine+after 2 and 4 weeks of pre- and post-caffeine treatment. The conexperimental values were calculated accordingly for each experiSignificant changes are expressed as **(P<0.01), ***(P<0.001) in coMPTP, τ(P<0.05) in comparison with caffeine+MPTP and $$(P<0
2.3. MPP+ level
Caffeine pre-treatment decreased MPP+ level in the striatum,whereas the level of MPP+ in the post-treatment group wasbelow the detectable limit. Caffeine pre-treated animals that
oreactivity of the dopaminergic neurons at 10×magnificationpars compacta (b). A, F, K andO represent controls, B, G, L andMPTP, E and J are caffeine+MPTP+ciprofloxacin respectivelytrol values were considered 100% in each replicate andment, therefore, there is no error bar in controls in (b).mparison with control; ###(P<0.001) in comparison with.01), $$$(P<0.001) in comparison with caffeine.

Fig. 3 – MPP+ level in the striatum of MPTP, caffeine+MPTPand caffeine+MPTP+ciprofloxacin treated animals in thepre-treatment groups. The data are expressed as means±S.E.M. (n=3–5 separate experiments). Significant changes*(P<0.05), ***(P<0.001) are expressed in comparison withMPTP.
118 B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
were also treated with ciprofloxacin did not produce anychange in MPP+ level (Fig. 3).
2.4. DAT expression
MPTP treatment reduced DAT mRNA expression as comparedwith control. Caffeine treatment in MPTP-treated animalssignificantly restored DAT mRNA level (Figs. 4a and b). Asimilar pattern was observed with DAT protein expression(Figs. 4c and d).
2.5. Adenosine A2A receptor expression
Caffeine treatment significantly attenuated adenosine A2A
receptor mRNA expression. MPTP augmented adenosine A2A
receptor mRNA expression as compared with control (Figs. 5aand b). The adenosine A2A receptor protein expression patternexhibited similar trend as observed with its mRNA expressionunder various treatment conditions (Figs. 5c and d). Both pre-and post-caffeine treatments produced such changes, how-ever, changes were slightly higher in the pre-treated animals.
2.6. CYP1A2 mRNA and protein expression and itscatalytic activity
Caffeine augmented CYP1A2 mRNA expression but it was notstatistically significant, whereas MPTP attenuated its mRNAlevel as compared with control significantly. Both pre- andpost-treatment with caffeine showed restoration in CYP1A2expression in MPTP-treated animals, however, pre-treatmentwith caffeine produced slightly more pronounced effect (Figs.6a and b). In caffeine pre-treated animals, CYP1A2 inhibitor,ciprofloxacin, decreased mRNA expression (Figs. 7a and b). Asimilar pattern was observed at the level of CYP1A2 proteinexpression (Figs. 6c and d). CYP1A2 inhibitor, ciprofloxacin,also decreased CYP1A2 protein expression in caffeine pre-
treated animals (Figs. 7c and d). The pattern of CYP1A2catalytic activity was similar as observed with proteinexpression under both the conditions (Figs. 6e and 7e).
3. Discussion
Caffeine is a well-established neuroprotective agent andreduces the MPTP-induced dopaminergic neurodegeneration(Singh et al., 2008). Although caffeine is mainly metabolized inthe liver, it is expected to partially metabolize in the brainsince it readily crosses the blood brain barrier due to its highdegree of lipid solubility and enters into the brain (McCall etal., 1982). Therefore, it was worthwhile to investigate CYP1A2expression in MPTP-treated mouse brain with and withoutcaffeine exposure. Similarly, adenosine A2A receptor and DATare associatedwith PD pathogenesis, as they participate eitherin caffeine antagonism or neurotransmitter transport. There-fore, their roles in MPTP-induced toxicity and caffeine-mediated neuroprotection were also looked into. Caffeinereduces the incidence of PD inmales rather than in females (ElYacoubi et al., 2000; Lindskog et al., 2002; Svenningsson andFredholm, 2003), the study was therefore conducted in malemice. Since the current treatment paradigms were optimizedin Swiss albino mice (Singh et al., 2008), this strain was usedfor the study. Secondly, Swiss albino mice have been found tobe very sensitive to MPTP as observed by biochemical andelectron microscopic analyses of dopaminergic neurons ofMPTP-treated mice (Rajeswari and Sabesan, 2008).
MPTP reduced TH-positive neurons, however, there was nosignificant change in the level of dopaminergic neurons incontrol/caffeine alone treated animals. MPTP-induced loss ofTH-positive neurons in caffeine treated groupswas also less. Asignificant change in TH immunoreactivity in the substantianigra suggested the possible degeneration of dopaminergicnerve terminals in the striatum. An altered level of dopaminein the striatum also confirmed this observation. This is inaccordance with the previous findings, which have shownthat caffeine treatment produces a dose dependent attenua-tion of MPTP-induced dopamine loss in mouse striatum (Xu etal., 2006). Caffeine at doses comparable with the typicalhuman exposure, also produced a dose dependent attenua-tion in the reduction of the striatal dopamine, triggered byMPTP (Chen et al., 2001).
MPTP converts into MPP+, a highly toxic metabolite andenter into the dopaminergic neurons through DAT (Gainetdi-nov et al., 1997). Caffeine-mediated reduction in theMPP+ levelin the pre-treated animals is also in accordance with previousobservations (Singh et al., 2008 Ulanowska et al., 2005). In thepost-treated group, no MPP+ was detected, which could beeither due to its complete removal from the brain, due to itsless stability or due to an outcome of an altered CYP1A2 level,as CYP1A2 is involved in caffeine metabolism. In the post-treatment group, level of MPP+ was analyzed 8 weeks after thelast MPTP treatment. Since the half-life of MPP+ is very short inrodents (Johannessen et al., 1985; Riachi et al., 1988), therefore,the level of MPP+ could not be detected in the MPTP post-treated animals. CYP1A2-mediated demethylation is respon-sible for about 80% of the systemic caffeine clearance (Kalowand Tang, 1993; Gu et al., 1992). Caffeine treatment increased

Fig. 4 – Effect of MPTP and caffeine on DAT mRNA and protein expressions in the pre- and post-treatment groups. DAT mRNAexpression is shown in panel (a), band density ratios (DAT /GAPDH mRNA) in (b), Western blot in (c) and relative proteinexpression in (d). Lanes 1, 5, 9, and 13—controls, lanes 2, 6, 10, and 14—caffeine, lanes 3, 7, 11, and 15—MPTP, lanes 4, 8, 12, and16—caffeine+MPTP, respectively in the pre-treated and post-treated groups. The data are expressed as means±S.E.M. (n=3–5separate experiments). Significant changes are expressed as *(P<0.05), **(P<0.01), ***(P<0.001) in comparison with control,#(P<0.05), ##(P<0.01), ###(P<0.001) with caffeine and $(P<0.05), $$(P<0.01) with MPTP.
119B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
CYP1A2 expression and activity, whereas MPTP treatmentattenuated the same. As CYP1A2 contributes to the detoxifica-tion of MPTP, therefore, caffeine was expected to alter MPTPdetoxification (Xu et al., 2002). The MPP+ level measured in thepresence of CYP1A2 inhibitor, ciprofloxacin, did not show anyalteration. The results indicate that an altered MPTP metabo-lism as a result of caffeine treatment could be via some otherroutes (Singh et al., 2008) and was not mediated by CYP1A2.
Caffeine pre- and post-treatment prevented the MPTP-mediated reduction in CYP1A2 enzymatic activity and its
expression. Caffeine is known to increase its own metabolismin a dose dependent manner through induction of CYP1A2 inthe liver (Goasduff et al., 1996). Altered CYP1A2 activity isexpected to be a reason for neuroprotection offered by caffeinein this study, as CYP1A2 get induced in vivo by chroniccaffeine exposure (Xu et al., 2002). Ciprofloxacin, a moderatelypotent inhibitor of CYP1A2, attenuated the expression andactivity of CYP1A2. This is in accordance with the previousreport showing 70% decrease in the activity of CYP1A2 in vitroby ciprofloxacin (Fuhr et al., 1992).

Fig. 5 – Effect of MPTP and caffeine on adenosine A2A mRNA and protein expression in the pre- and post-treatment groups.Adenosine A2A mRNA expression is shown in panel (a), band density ratios (A2A/GAPDH mRNA) in (b), Western blot in (c) andrelative protein expression in (d). Lanes 1, 5, 9, and 13—controls, lanes 2, 6, 10, and 14—caffeine, lanes 3, 7, 11, and 15—MPTP,lanes 4, 8, 12, and 16—caffeine+MPTP, respectively in the pre-treated and post-treated groups (a and c). The data are expressedas means±S.E.M. (n=3–5 separate experiments). Significant changes *(P<0.05), **(P<0.01), ***(P<0.001) are expressed incomparison with control, #(P<0.05), ##(P<0.01), ###(P<0.001) with caffeine and $(P<0.05), $$$(P<0.001) with MPTP.
120 B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
Caffeine, a non-selective antagonist of adenosine A2A
receptor attenuated its expression at transcriptional andtranslational levels, whereas MPTP treatment augmented itsexpression. This is supported by the fact that the neuropro-tective effect of caffeine is mediated by adenosine receptorantagonism (Fredholm et al., 1999), and the inactivation orinhibition of A2A receptor inhibits MPTP-induced dopaminer-gic damage (Pierri et al., 2005). Blockade of A2A receptors viacaffeine alters the release of GABA, acetylcholine and gluta-
mate neurotransmitters. A2A receptor antagonism has beenreported to play a critical neuroprotective role in brain injury(Jones, 2008; Monopoli et al., 1998; Ravina et al., 2003). Theactivation of the striatal adenosine A2A receptors decreasesthe affinity of D2 receptors for dopamine and has opposingeffect on cyclic AMP (cAMP) formation, which ultimately leadsto the activation of the striatopallidal/indirect pathway.Blockade of adenosine A2A receptor activity inhibits theindirect pathway/inhibitory pathway leading to facilitated

Fig. 6 – Effect of MPTP and caffeine on CYP1A2 mRNA and protein expression and its catalytic activity in the pre- andpost-treatment groups. CYP1A2 mRNA expression is shown in panel (a), band density ratios (CYP1A2/GAPDH mRNA) in (b),Western blot in (c), relative protein expression in (d) and catalytic activity (MROD) in (e). Lanes 1, 5, 9, and 13—control, lanes 2, 6,10, and 14—caffeine, lanes 3, 7, 11, and 15—MPTP, lanes 4, 8, 12, and 16—caffeine+MPTP, respectively in the pre- andpost-treatment groups (a and c). The data are expressed as means±S.E.M. (n=3–5 separate experiments). Significant changes*(P<0.05), **(P<0.01), ***(P<0.001) are expressed in comparison with control, #(P<0.05), ##(P<0.01), ###(P<0.001) with caffeine,$(P<0.05), $$(P<0.01), $$$(P<0.001) with MPTP.
121B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
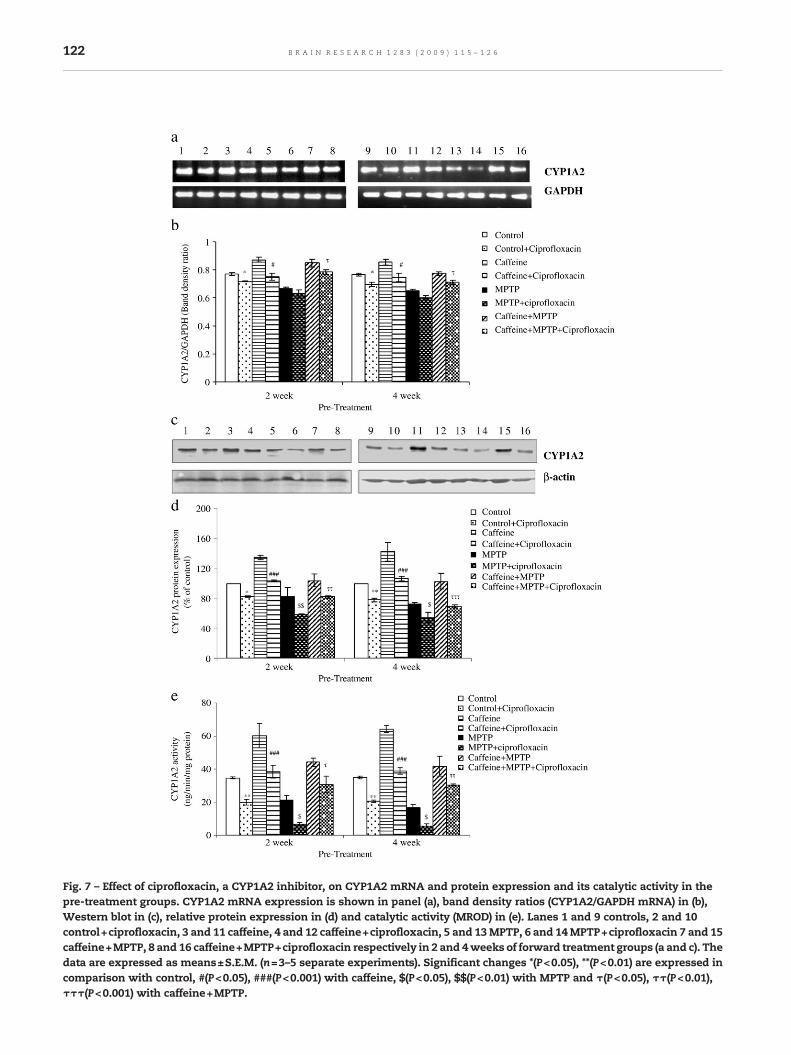
Fig. 7 – Effect of ciprofloxacin, a CYP1A2 inhibitor, on CYP1A2 mRNA and protein expression and its catalytic activity in thepre-treatment groups. CYP1A2 mRNA expression is shown in panel (a), band density ratios (CYP1A2/GAPDH mRNA) in (b),Western blot in (c), relative protein expression in (d) and catalytic activity (MROD) in (e). Lanes 1 and 9 controls, 2 and 10control+ciprofloxacin, 3 and 11 caffeine, 4 and 12 caffeine+ciprofloxacin, 5 and 13MPTP, 6 and 14MPTP+ciprofloxacin 7 and 15caffeine+MPTP, 8 and 16 caffeine+MPTP+ciprofloxacin respectively in 2 and 4weeks of forward treatment groups (a and c). Thedata are expressed as means±S.E.M. (n=3–5 separate experiments). Significant changes *(P<0.05), **(P<0.01) are expressed incomparison with control, #(P<0.05), ###(P<0.001) with caffeine, $(P<0.05), $$(P<0.01) with MPTP and τ(P<0.05), ττ(P<0.01),τττ(P<0.001) with caffeine+MPTP.
122 B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6

123B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
movements (Ochi et al., 2000). VMAT-2 expression is downregulated by cAMP production (Nakanishi et al., 1995), whichmay alter the toxicity response, thus protecting dopaminergicneurons.
In this study, MPTP treatment significantly attenuatedDAT mRNA and protein expression. This is not an unusualphenomenon, as reduced DAT mRNA expression followingMPTP treatment has been reported in a number of studies(Bousquet et al., 2008; Thiriet et al., 2008; Xu et al., 2005). Liuet al. (2008a; 2008b) have also reported a significant decreasein DAT expression in the midbrain of MPTP-treated mice intwo independent studies. The change in DAT level is inaccordance with the fact that MPTP targets DA neuron andDAT acts as a molecular gateway for MPP+ entry (Kurosaki etal., 2003; Gainetdinov et al., 1997). Caffeine restored DATexpression in both pre- and post-treated groups possibly byprotecting DA neurons from degeneration. The present studydemonstrates that caffeine-mediated neuroprotective effectcould be partially contributed by CYP1A2, adenosine A2A
receptor and DAT.
4. Experimental procedures
4.1. Chemicals
The following chemicals were procured from Sigma-Aldrich,USA—acetic acid, acetone, agarose, acetonitrile, bovine serumalbumin, Bradford reagent, bromophenol blue, caffeine,chloroform, disodium hydrogen orthophosphate (Na2HPO4),dithiothreitol (DTT), ethidium bromide, ethanol, ethylene-diamine-tetra-acetic acid (EDTA), goat anti-rabbit IgG horse-radish peroxidase (HRP) conjugated secondary antibody,heptane sulphonic acid, hydrogen peroxide, magnesiumchloride, magnesium sulphate, methanol, MPTP hydro-chloride, rabbit monoclonal-HRP conjugate, monoclonal anti-tyrosine hydroxylase antibody, monoclonal anti-β-actinantibody, mouse monoclonal-HRP conjugate, methanol,nicotinamide adenine dinucleotide phosphate (NADPH),perchloric acid, phenyl methylsulphonyl fluoride (PMSF),p-nitrophenol, potassium phosphate, 4-nitrocatechol, potas-sium chloride (KCl), sodium chloride (NaCl), sodium dihydro-gen phosphate (NaH2PO4), sodium dodecyl sulphate (SDS),sodium hydroxide, sodium pyrophosphate, sucrose, tris-base,Triton-X100 and Tween-20. RT-PCR kit were procured fromMBI Fermentas, USA and forward and reverse primers for A2A,CYP1A2, DAT and GAPDH, Taq DNA polymerase, dNTPs and100 bp ladder were purchased from Bangalore Genei, India.Polyvinylidene difluoride (PVDF) membrane was procuredfrom GE Healthcare, Europe, mouse monoclonal anti-A2A
antibody from Santa Cruz Biotechnology, Incorporation; rabbitanti-DAT, rabbit anti-rat 1A2 antibody from Chemicon Inter-national, USA and Vectastain Universal Quick kit was pro-cured from Vector Laboratory, USA.
4.2. Animal treatment
Male Swiss albinomice (20–25 g)were obtained from the animalcolony of the Indian Institute of Toxicology Research (IITR),Lucknow. The animals were kept under standard conditions
(temperature—22 °C±2 °C, humidity—45–55% and light inten-sity 300–400 lx). The animals were fed standard pellet diet andwater ad libitum (Singh et al., 2008). The Institutional EthicsCommittee for the Use of Laboratory Animals approved thestudy. In the pre-treatment group, the animals were treateddaily intraperitoneally with caffeine (20 mg/kg) for 8 weeksfollowed by MPTP hydrochloride (20 mg/kg) co-treatment oncein a day for 2–4 weeks (Singh et al., 2008). In the post-treatmentgroup, the animals were treated daily intraperitoneally withMPTP hydrochloride (20 mg/kg) once in a day for 2–4 weeks,followedby 2–4weeks caffeine (20mg/kg) and finallywith eithernormal saline (0.9% NaCl) or caffeine for 8 weeks. Control/vehicle animals were treatedwith the corresponding volume ofnormal saline. After the final treatment, the animals weresacrificed via cervical dislocation; the striatum was dissectedout and frozen immediately in liquid nitrogen till further use.
4.3. Immunohistochemistry
TH immunoreactivity was performed using standard proce-dure (Gorbatyuk et al., 2008). Animals were anesthetized andintra-cardiac perfusionwas donewith normal saline, followedby paraformaldehyde (4%) in phosphate buffered saline. Afterperfusion, the brain was dissected coronally through medianeminence. The caudal block was post-fixed, cryoprotected andsections were cut using a cryostat. The non-specific labelingwas blocked by incubating the sections in blocking buffer (1.5%normal horse serum 0.1% triton X-100 in phosphate bufferedsaline) for 2 h. The sections were incubated in monoclonalanti-TH antibody (1:250) at 4 °C for 12 h. The sections werewashed thrice with phosphate buffered saline for 15 min eachand incubated with secondary antibody for 1 h, followed bystreptavidin peroxidase complex for 30 min. The color wasdeveloped with 3, 3 diaminobenzidine and the sections weredehydrated in graded ethanol and permanentlymountedwithDPX. The mounted sections were visualized under themicroscope. The images were captured at 10× magnification.The TH+ cells were counted using a method describedelsewhere (Mochizuki et al., 2001).
4.4. High performance liquid chromatography (HPLC)analysis of dopamine and 1-methyl 4-phenylpyridiniumion (MPP+)
The striatal dopamine and MPP+ content were measured, asdescribed previously (Singh et al. 2008). The dopamine level isexpressed in ng/mg of tissue and MPP+ level is expressed asng/mg of protein. The limit of detection of MPP+ was 5 ng/ml.
4.5. Preparation of microsomes
The perfused striatum was homogenized in potassiumphosphate buffer (10% w/v; 0.1 M, pH 7.4), containing EDTA(1 mM), pepstatin (0.1 mM), PMSF (0.1 mM) 0.135 M KCl andglycerol (20%) in ice-cold condition. The supernatant obtained(900 ×g, 10 min) was re-centrifuged (100,000 ×g, 1 h). The pelletwas suspended in phosphate buffer (67 mM), containingsucrose (0.25 M), EDTA (1 mM), PMSF (0.1 mM), proteaseinhibitor cocktail (20 g/ml) and glycerol (20%) andwas stored at−80 °C till further use.

124 B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
4.6. Estimation of the protein content
The protein content was measured using standard curve ofbovine serum albumin (Bradford 1976).
4.7. Western blotting
Protein (100 μg) was denatured with SDS buffer, separated on12% polyacrylamide gel and transferred to PVDF membrane(Sigma). The blots were blocked with tris buffered saline (TBS)containing 5% non-fat dry milk for 2 h to arrest non-specificbinding and incubated with primary antibodies againstCYP1A2, A2A receptor and DAT (1:200) in TBS at 4 °C overnight.Rabbit monoclonal-HRP conjugate, mouse monoclonal-HRPconjugate and goat anti-rabbit IgG HRP conjugated secondaryantibody (1:500) respectively in TBS were used to detectCYP1A2, A2A and DAT. The blots were visualized using DAB/H2O2 as substrates. Relative band density was calculated withrespect to β-actin and the data are expressed as % of control.
4.8. Isolation of RNA and reversetranscription-polymerase chain reaction
RNA was extracted from the striatum using Trizol reagentfollowingmanufacturer's instructions. Revert aid™ minusMu-LV reverse transcriptase was used to synthesize cDNA. Theprimers were synthesized and PCR amplification conditionswere used as reported elsewhere. In brief, primers used forCYP1A2 (Choudhary et al., 2003) were 5′ 5′GACGTCAG-CATCCTCTTGCT3 ′ ( f o rward ) and 5 ′GGCACTTG-TGATGTCTTGGA3′ (reverse), annealing temperature was65.5 °C and the product size was 400-base pair. Similarly, theprimers for DAT (Patel et al., 2008) were 5′CGGTGGCAGC-TCACAGC3′ (forward) and 5′TGGAGAAGGCGATCAGCAC3′(reverse), annealing temperature was 60 °C and the productsize was 292-base pair. The primers used for A2A (Tarditi et al.,2006) were 5′TGTCCTGGTCCTCACGCAGAG3′ (forward) and 5′CGGATCCTGTAGGCGTAGATGAAGG3′ (reverse), annealingtemperature was 55 °C and the product size was 600-basepair. The primer sequences for GAPDH (Singh et al., 2008) were5′CTCATGACCACAGTCCATGC3′ (forward) and 5′CACATTG-GGGGTAGGAACAC3′ (reverse). The GAPDH was amplifiedconcurrently with the respective genes. PCR products werevisualized in 2% agarose gel using ethidium bromide underUV. The band density was calculated by computerizeddensitometry system and normalized with GAPDH.
4.9. CYP1A2 catalytic activity
Catalytic activity of CYP1A2 (7-methoxyresorufin O-demethy-lase, MROD) was measured by mixing 50–250 μg microsomalprotein with 0.1 M phosphate buffer (pH 7.4) containing 5 mMglucose–6-phosphate, 2 U of glucose–6-phosphate dehydro-genase, 5 mM magnesium sulphate, 1.6 mg/ml BSA, 1.5 μM 7-methoxy resorufin (Upadhyay et al., 2007). In brief, NADPH(0.6 nmol) was added into the reaction mixture to initiate thereaction. Themixed content was incubated at 37 °C for 20 minand the reaction was stopped by adding 2.5 ml methanol andkeeping the mixture in ice. Reaction mixture was centrifugedat 825 ×g for 10 min and supernatant was collected. Fluores-
cencewasmeasuredat 550nmexcitationand585nmemissionwavelengths. The enzymatic activity was calculated in pmol/min/mg protein.
4.10. Statistical analysis
The data are expressed as means±standard error of means(S.E.M.). Two-way analysis of variance (ANOVA) was used forcomparison between different groups with Bonferroni post-test. The difference was considered statistically significantwhen “P” value was less than 0.05.
Acknowledgments
The authors thank the Department of Biotechnology (DBT) forthe financial support. Authors also acknowledge the Univer-sity Grant Commission (UGC), New Delhi, India for providingresearch fellowship to Seema Singh. The IITR communicationnumber of this article is 2728.
R E F E R E N C E S
Ascherio, A., Zhang, S.M., Hernan, M.A., Kawachi, I., Colditz, G.A.,Speizer, F.E., Willett, W.C., 2001. Prospective study of caffeineconsumption and risk of Parkinson's disease in men andwomen. Ann. Neurol. 50, 56–63.
Avshalumov, M.V., Chen, B.T., Koos, T., Tepper, J.M., Rice, M.E.,2005. Endogenous hydrogen peroxide regulates the excitabilityof midbrain dopamine neurons via ATP-sensitive potassiumchannels. J. Neurosci. 25, 4222–4231.
Bousquet, M., Saint-Pierre, M., Julien, C., Salem, N.Jr., Cicchetti, F.,Calon, F., 2008. Beneficial effects of dietary omega-3polyunsaturated fatty acid on toxin-induced neuronaldegeneration in an animal model of Parkinson's disease.FASEB J. 22, 1213–1225.
Bradford, M.M., 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizingthe principle of protein-dye binding. Anal. Biochem. 72,248–254.
Canals, M., Marcellino, D., Fanelli, F., Ciruela, F., de Benedetti, P.,Goldberg, S.R., Neve, K., Fuxe, K., Agnati, L.F., Woods, A.S.,Ferre, S, Lluis, C., Bouvier, M., Franco, R., 2003. AdenosineA2A-dopamine D2 receptor–receptor heteromerization:qualitative and quantitative assessment by fluorescence andbioluminescence energy transfer. J. Biol. Chem. 278,46741–46749.
Carrillo, J.A., Benitez, J., 1996. CYP1A2 activity, gender andsmoking, as variables influencing the toxicity of caffeine. Br. J.Clin. Pharmacol. 41, 605–608.
Chen, J.F., Xu, K., Petzer, J.P., Staal, R., Xu, Y.H., Beilstein, M.,Sonsalla, P.K., Castangoli, K., Castangoli, N.J., Scharzchild, M.A.,2001. Neuroprotection by caffeine and A(2A) adenosinereceptor inactivation in a model of Parkinson's disease.J. Neurosci. 21, RC143.
Chen, X., Lan, X., Roche, I., Liu, R., Geiger, J.D., 2008. Caffeineprotects against MPTP-induced blood–brain barrierdysfunction in mouse striatum. J Neurochem. 107, 1147–1157.
Choudhary, D., Jansson, I., Schenkman, J.B., Sarfarazi, M., Stoilov,I., 2003. Comparative expression profiling of 40 mousecytochrome P450 genes in embryonic and adult tissues. Arch.Biochem. Biophys. 414, 91–100.
Ciruela, F., Burgueno, J., Casado, V., Canals, M., Marcellino, D.,Goldberg, S.R., Bader, M., Fuxe, K., Agnati, L.F., Lluis, C., Franco,

125B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
R., Ferre, S., Woods, A.S., 2004. Combining mass spectrometryand pull-down techniques for the study of receptorheteromerization. Direct epitope–epitope electrostaticinteractions between adenosine A2A and dopamine D2receptors. Anal. Chem. 76, 5354–5363.
El Yacoubi, M., Ledent, C., Menard, J.F., Parmentier, M., Costentin,J., Vaugeois, J.M., 2000. The stimulant effects of caffeine onlocomotor behaviour in mice are mediated through itsblockade of adenosine A(2A) receptors. Br. J. Pharmacol. 129,1465–1473.
Ferre, S., Fuxe, K., von Euler, G., Johansson, B., Fredholm, B.B., 1992.Adenosine–dopamine interactions in the brain. Neuroscience51, 501–512.
Fredholm, B.B., 2004. Connection between caffeine, adenosinereceptors and dopamine. Coffee reduces the risk of Parkinsondisease. Lakartidningen 101, 2552–2555.
Fredholm, B.B., Battig, K., Holmen, J., Nehlig, A., Zvartau, E.E., 1999.Actions of caffeine in the brain with special reference to factorsthat contribute to its widespread use. Pharmacol. Rev. 51,83–133.
Fuhr, U., Anders, E.M., Mahr, G., Sorgel, F., Staib, A.H., 1992.Inhibitory potency of quinolone antibacterial agents againstcytochrome P450IA2 activity in vivo and in vitro. Antimicrob.Agents Chemother. 36, 942–948.
Fuxe, K., Ferre, S., Zoli, M., Agnati, L.F., 1998. Integratedevents in central dopamine transmission as analyzed atmultiple levels. Evidence for intramembrane adenosineA2A/dopamine D2 and adenosine A1/dopamine D1 receptorinteractions in the basal ganglia. Brain Res. Brain Res. Rev. 26,258–273.
Gainetdinov, R.R., Fumagalli, F., Jones, S.R., Caron, M.G., 1997.Dopamine transporter is required for in vivo MPTPneurotoxicity: evidence from mice lacking the transporter.J. Neurochem. 69, 1322–1325.
Gerlach, M., Riederer, P., 1996. Animal models of Parkinson'sdisease: an empirical comparison with the phenomenologyof the disease in man. J. Neural. Transm. 103, 987–1041.
Goasduff, T., Dreano, Y., Guillois, B., Menez, J.F., Berthou, F., 1996.Induction of liver and kidney CYP1A1/1A2 by caffeine in rat.Biochem. Pharmacol. 52, 1915–1919.
Gorbatyuk, O.S., Li, S., Sullivan, L.F., Chen, W., Kondrikova, G.,Manfredsson, F.P., Mandel, R.J., Muzyczka, N., 2008. Thephosphorylation state of Ser-129 in human α-synucleindetermines neurodegeneration in a rat model ofParkinson disease. Proc. Natl. Acad. Sci. U. S. A. 105,763–768.
Gu, L., Gonzalez, F.J., Kalow, W., Tang, B.K., 1992.Biotransformation of caffeine, paraxanthine, theophylline,and theobromine by cDNA expressed human CYP1A2 andCYP2E1. Pharmacogenetics 2, 73–77.
Johannessen, J.N., Chiueh, C.C., Burns, R.S., Markey, S.P., 1985.Differences in the metabolism of MPTP in the rodent andprimate parallel differences in sensitivity to its neurotoxiceffects. Life Sci. 36, 219–224.
Jones, G., 2008. Caffeine and other sympathomimetic stimulants:modes of action and effects on sports performance. EssaysBiochem. 44, 109–123.
Kalow, W., Tang, B.K., 1993. The use of caffeine for enzyme assays;a critical appraisal. Clin. Pharmacol. Ther. 53, 503–514.
Kurokawa, M., Koga, K., Kase, H., Nakamura, J., Kuwana, Y., 1996.Adenosine A2a receptor-mediated modulation of striatalacetylcholine release in vivo. J. Neurochem. 66, 1882–1888.
Kurosaki, R., Muramatsu, Y., Watanabe, H., Michimata,M., Matsubara, M., Imai, Y., Araki, T., 2003. Role ofdopamine transporter against MPTP(1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) neurotoxicityin mice. Metab. Brain Dis. 18, 139–146.
Lindskog, M., Svenningsson, P., Pozzi, L., Kim, Y., Fienberg, A.A.,Bibb, J.A., Fredholm, B.B., Nairn, A.C., Greengard, P., Fisone, G.,
2002. Involvement of DARPP-32 phosphorylation in thestimulant action of caffeine. Nature 418, 774–778.
Liu, B., Xie, J.X., Tsang, L.L., Rowlands, D.K., Ho, L.S., Gou, Y.L.,Chung, Y.W., Chan, H.C., 2008a. Bak Foong protectsdopaminergic neurons against MPTP-inducedneurotoxicity by its anti-apoptotic activity. Cell Biol. Int.32, 86–92.
Liu, L.X., Chen, W.F., Xie, J.X., Wong, M.S., 2008b.Neuroprotective effects of genistein on dopaminergicneurons in the mice model of Parkinson's disease. Neurosci.Res. 60, 156–161.
Mao, X., Chai, Y., Lin, Y.F., 2007. Dual regulation of theATP-sensitive potassium channel by caffeine. Am. J. Physiol.Cell Physiol. 292, C2239–2258.
McCall, A.L, Millington, W.R., Wurtman, R.J., 1982. Blood–brainbarrier transport of caffeine: dose-related restriction ofadenine transport. Life Sci. 31, 2709–2715.
Mochizuki, H., Hayakawa, H., Migita, M., Shibata, M., Tanaka, R.,Suzuki, A., Shimo-Nakanishi, Y., Urabe, T., Yamada, M.,Tamayose, K., Shimada, T., Miura, M., Mizuno, Y., 2001.An AAV-derived Apaf-1 dominant negative inhibitorprevents MPTP toxicity as antiapoptotic gene therapy forParkinson's disease. Proc. Natl. Acad. Sci. U. S. A. 98,10918–10923.
Monopoli, A., Lozza, G., Forlani, A., Mattavelli, A., Ongini, E., 1998.Blockade of adenosine A2A receptors by SCH 58261 resultsin neuroprotective effects in cerebral ischaemia in rats.NeuroReport 9, 3955–3959.
Mori, A., Shindou, T., Ichimura, M., Nonaka, H., Kase, H., 1996.The role of adenosine A2a receptors in regulating GABAergicsynaptic transmission in striatal medium spiny neurons.J. Neurosci. 16, 605–611.
Nakanishi, N., Onozawa, S., Matsumoto, R., Kurihara, K., Ueha, T.,Hasegawa, H., Minami, N., 1995. Effects of protein kinaseinhibitors and protein phosphatase inhibitors on cyclicAMP-dependent down-regulation of vesicular monoaminetransport in pheochromocytoma PC12 cells. FEBS Lett. 368,411–414.
Nakaso, K., Ito, S., Nakashima, K., 2008. Caffeine activates thePI3K/Akt pathway and prevents apoptotic cell death in aParkinson's disease model of SH-SY5Y cells. Neurosci. Lett.432, 146–150.
Ochi, M., Koga, K., Kurokawa, M., Kase, H., Nakamura, J., Kuwana,Y., 2000. Systemic administration of adenosine A(2A) receptorantagonist reverses increased GABA release in the globuspallidus of unilateral 6-hydroxydopamine-lesioned rats: amicrodialysis study. Neuroscience 100, 53–62.
Patel, S., Singh, K., Singh, S., Singh, M.P., 2008. Geneexpression profiles of mouse striatum in control andmaneb-paraquat induced Parkinson's disease phenotype:validation of differentially expressed energy metabolizingtranscripts. Mol. Biotechnol. 40, 59–68.
Pierri, M., Vaudano, E., Sager, T., Englund, U., 2005. KW-6002protects from MPTP induced dopaminergic toxicity in themouse. Neuropharmacology 48, 517–524.
Przedborski, S., Jackson Lewis, V., Djaldetti, R., Liberatore,G., Vila, M., Vukosavic, S., Almer, G., 2000. TheParkinsonian-toxin MPTP: action and mechanism. Restor.Neurol. Neurosci. 16, 135–142.
Rajeswari, A., Sabesan, M., 2008. Neuropathologicalchanges induced by neurotoxin1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in male Swissalbino mice. Toxicol. Ind. Health 24, 189–194.
Ravina, B.M., Fagan, S.C., Hart, R.G., Hovinga, C.A., Murphy, D.D.,Dawson, T.M., Marler, J.R., 2003. Neuroprotective agents forclinical trials in Parkinson-s disease: a systematic assessment.Neurology 60, 1234–1240.
Riachi, N.J., Harik, S.I., Kalaria, R.N., Sayre, L.M., 1988.On the mechanisms underlying

126 B R A I N R E S E A R C H 1 2 8 3 ( 2 0 0 9 ) 1 1 5 – 1 2 6
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine neurotoxicity.II. Susceptibility among mammalian species correlates withthe toxin's metabolic patterns in brain microvessels and liver.J. Pharmacol. Exp. Ther. 244, 443–448.
Richardson, P.J., Kase, H., Jenner, P.G., 1997. Adenosine A2Areceptor antagonists as new agents for the treatment ofParkinson-s disease. Trends Pharmacol. Sci. 18, 338–344.
Singh, S., Singh, K., Patel, S., Patel, D.K., Singh, C., Nath, C.,Singh, M.P., 2008. Nicotine and caffeine-mediatedmodulation in the expression of toxicant responsivegenes and vesicular monoamine transporter-2 in1-methyl 4-phenyl-1,2,3,6-tetrahydropyridine'inducedParkinson's disease phenotype in mouse. Brain Res. 1207,193–207.
Svenningsson, P., Fredholm, B.B., 2003. Exciting news about A2Areceptors. Neurology 61, S10–S11.
Tanner, C.M., Ottman, R., Goldman, S.M., Ellenberg, J., Chan, P.,Mayeux, R., Langston, J.W., 1999. Parkinson disease in twins: anetiologic study. JAMA 281, 341–346.
Tarditi, A., Camurri, A., Varani, K., Borea, P.A., Woodman, B., Bates,G., Cattaneo, E., Abbracchio, M.P., 2006. Early and transientalteration of adenosine A2A receptor signaling in a mousemodel of Huntington disease. Neurobiol. Dis. 23, 44–53.
Thiriet, N., Amar, L., Toussay, X., Lardeux, V., Ladenheim, B.,Becker, K.G., Cadet, J.L., Solinas, M., Jaber, M., 2008.Environmental enrichment during adolescence regulatesgene expression in the striatum of mice. Brain Res. 1222,31–41.
Ulanowska, K., Piosik, J., Gwizdek-Wisniewska, A., We Grzyn, G.,2005. Formation of stacking complexes between caffeine(1,2,3-trimethylxanthine) and1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mayattenuate biological effects of this neurotoxin.Bioorg. Chem. 33, 402–413.
Upadhyay, G., Kumar, A., Singh, M.P., 2007. Effect of silymarin onpyrogallol-and rifampicin-induced hepatotoxicity in mouse.Eur. J. Pharmacol. 565, 190–201.
Vieregge, P., Hagenah, J., Heberlein, I., Klein, C., Ludin, H.P., 1999.Parkinson's disease in twins: a follow-up study. Neurology 53,566–572.
Wirdefeldt, K., Gatz, M., Schalling, M., Pedersen, N.L., 2004. Noevidence for heritability of Parkinson disease in Swedish twins.Neurology 63, 305–311.
Xu, K., Xu, Y., Brown-Jermyn, D., Chen, J.F., Ascherio, A.,Dluzen, D.E., Schwarzschild, M.A., 2006. Estrogen preventsneuroprotection by caffeine in the mouse1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model ofParkinson's disease. J. Neurosci. 26, 535–541.
Xu, K., Xu, Y.H., Chen, J.F., Schwarzschild, M.A., 2002.Caffeine-s neuro-protection against1-methyl-4-phenyl-1,2,3,6tetrahydropyridine toxicity showsno tolerance to chronic caffeine administration in mice.Neurosci. Lett. 322, 13–16.
Xu, Z., Cawthon, D., McCastlain, K.A., Slikker Jr., W., Ali, S.F., 2005.Selective alterations of gene expression in mice induced byMPTP. Synapse 55, 45–51.




![[3104] Samhällskunskap, SAM - Skolverket...Social studies 1a2 SAMSAM01a2 Social studies 1a2 The course social studies 1a2 covers points 1–5 under the heading Aim of the subject,](https://img.dokumen.tips/doc/110x75/5f3860bd8cb5b531b06e290f/3104-samhllskunskap-sam-skolverket-social-studies-1a2-samsam01a2-social.jpg)
























