Embed Size (px)
Citation preview

Mol. Cells, Vol. 18, No. 1, pp. 107-114
�
Ectopic Expression of a Cold-inducible Transcription Factor, CBF1/DREB1b, in Transgenic Rice (Oryza sativa L.)
Sang-Choon Lee, Kyung-Won Huh, Kyungsook An1, Gynheung An1, and Seong-Ryong Kim* Department of Life Science, Sogang University, Seoul 121-742, Korea; 1 National Research Laboratory of Plant Functional Genomics, Division of Molecular and Life Science, Pohang University of Science and Technology, Pohang 790-784, Korea. (Received April 30, 2004; Accepted May 27, 2004) The gene encoding C-repeat/dehydration-responsive element binding factor 1 (CBF1/DREB1b) of Arabidop-sis was introduced into rice (Oryza sativa L.) under the control of the maize ubiquitin promoter. Its incorpora-tion and expression in transgenic rice plants were con-firmed by DNA and RNA gel-blot analyses. Cold tol-erance in the transgenics was not significantly differ-ent from that of the wild-type plants, as determined by ion leakage, chlorophyll fluorescence, and survival rates. However, the cold-responsive genes lip5, lip9, and OsDhn1 were up-regulated in the transgenic plants, suggesting that the cold signal transduction pathway involving CBF1 is partially conserved in this cold-labile plant. Keywords: CBF1/DREB1b; Cold Stress; Transgenic Rice. Introduction When exposed to low temperatures, plants respond with changes in their pattern of gene expression and protein products (Cattivelli and Bartels, 1992; Guy, 1990). This ability to adapt has an impact on the distribution and sur-vival of the plant, and on crop yields. Many species of tropical or subtropical origin are injured or killed by non-freezing low temperatures, and exhibit various symptoms of chilling injury such as chlorosis, necrosis, or growth retardation (Wang, 1990). In contrast, chilling-tolerant species are able to grow at such cold temperatures (Larcher et al., 1973; Rédei, 1969).
Various low temperature-inducible genes have been * To whom correspondence should be addressed. Tel: 82-2-705-8456; Fax: 82-2-704-3601 E-mail: [email protected]
isolated from plants. Most appear to be involved in toler-ance to cold stress (Crosatti et al., 1996; Guy, 1990; Houde et al., 1992; Hughes and Dunn, 1996; Mohapatra et al., 1989; Thomashow, 1994), and the expression of some of them is regulated by C-repeat/dehydration-responsive element binding (CBF/DREB1) transcription factors. Three CBF/DREB1 genes (CBF3/DREB1a, CBF1/DREB1b, and CBF2/DREB1c) belonging to the AP2/EREBP family of DNA-binding proteins (Riechmann and Meyerowitz, 1998) have been identified in Arabidopsis (Gilmour et al., 1998; Kasuga et al., 1999; Liu et al., 1998; Stockinger et al., 1997). Overexpression of CBF1/DREB1b and CBF3/ DREB1a enhances cold tolerance by inducing COR (cold-regulated) genes (Gilmour et al., 2000; Jaglo-Ottosen et al., 1998; Kasuga et al., 1999; Liu et al., 1998). Further-more, its overexpression causes many biochemical changes, such as the accumulation of sugar and proline (Gilmour et al., 2000). Thus, the CBF/DREB1 genes are thought to be activators that integrate several components of the cold acclimation response by which plants increase their toler-ance to low temperatures after exposure to nonfreezing conditions (Gilmour et al., 2000; Guy, 1990).
In contrast to some crops that show cold tolerance at 0 to 4°C, rice is adversely affected by temperatures ranging from 10 to 20°C (Toenniessen, 1991). Poor seedling vigor, poor fertility, and, consequently, reduced yields are the major responses associated with cold stress. To overcome this problem a transgenic approach to promoting cold tolerance has been widely adopted, with some success. For example, increasing the accumulation of two com-patible solutes, i.e., glycinebetaine and trehalose, in trans-genic rice by overexpressing either E. coli choline oxidase, or trehalose-6-phosphate synthase fused to trehalose-6-phosphate phosphatase, enhanced tolerance to both salt and cold (Jang et al., 2003; Sakamoto et al., 1998). In-creasing photorespiration capacity by introducing chloro-plast glutamine synthetase also increased salt and cold
M olecules and
Cells KSMCB 2004

108 Analysis of Transgenic Rice Expressing CBF1/DREB1b �
tolerance, as shown by a rise in the quantum yield of PSII after exposure to those stresses (Hoshida et al., 2000). Finally, overexpression of a calcium-dependent protein kinase, CDPK7, increased cold and salt/drought tolerance in transgenic rice plants (Saijo et al., 2000).
Expressing CBF/DREB1 genes in transgenic canola and tomato plants also improved cold tolerance, although freezing tolerance was not increased in the latter species (Hsieh et al., 2002b; Jaglo et al., 2001). The objective of the current study was to introduce a maize ubiquitin pro-moter-CBF1/DREB1b fusion into rice and characterize the effect of its expression at the molecular level. Materials and Methods Plant material and growth conditions Rice (Oryza sativa cv. Dongjin) seeds were surface-sterilized, imbibed for 2 d in tap water, and germinated on water-soaked filter paper in Petri dishes at 30°C in the dark. After 2 d of germination, the seed-lings were transplanted to soil and grown in a greenhouse at 28°C. The light/dark cycle in the greenhouse was 16/8 h. To measure root growth, sterilized seeds were put on paper towels (5 cm from the top). The towels were then folded, covered with aluminum foil, placed vertically in water, and incubated at 30°C for 5 d. Bacterial strains Escherichia coli strain XL-1 Blue MRF′ was used as host for routine cloning experiments. Agrobacterium tumefaciens LBA4404, of the Ach5 chromosomal background, and a disarmed Ti plasmid pAL4404, were used for rice trans-formation (Hoekema et al., 1983). Isolation of the CBF1/DREB1b gene and construction of an expression vector PCR was conducted with forward (5′-AAGCTTAAAGAATCTACCTG-3′) and reverse (5′-GGTAC-CCTCGTTTCTACAAC-3′) primers based on the mRNA se-quence of CBF1/DREB1b (Medina et al., 1999). Genomic DNA from A. thaliana ecotype Columbia was used as template and the amplified fragment was cloned into the pBluescript II KS vector (Stratagene, USA). PCR conditions were 1 min at 94°C, 1 min at 55°C, and 1 min at 72°C, for 35 cycles. Preparation of DNA, sequencing, and computer analyses were performed as described by Hong et al. (2001). A DNA fragment containing the com-plete open reading frame of CBF1/DREB1b was introduced into the binary vector pGA1611 (Kang et al., 1998; Kim et al., 2003) under the maize ubiquitin promoter. Generation of transgenic rice plants Transformation was per-formed by the Agrobacterium-mediated cocultivation method (Hiei et al., 1994; Lee et al., 1999). Plants were regenerated on medium containing 40 mg/L hygromycin B, grown in the green-house and the seeds from individual lines harvested. PCR analy-sis to identify transgenic plants was conducted with the ubiq-uitin primer (5′-CACCCTGTTGTTTGGTG-3′) and the nos
terminator primer (5′-GCGGGACTCTAATCATAAAAACC-3′) located on the binary vector. The PCR conditions were 35 cycles of 1 min at 94°C, 1 min at 55°C, and 1 min at 72°C. DNA and RNA gel-blot analyses Genomic DNA was extracted from rice leaves as described by Chen and Ronald (1999). Five micrograms of DNA, digested with restriction enzymes for 12 h at 37°C, was separated on a 0.8% agarose gel and transferred to a Hybond-N membrane (Amersham, UK) with a vacuum trans-fer system (Hoefer, USA). Total RNA was isolated using Tri reagent (Molecular Research Center, USA), and ten micrograms of total RNA was resolved on a 1.3% formaldehyde agarose gel and blotted onto a nylon membrane (Sambrook et al., 1989). To prepare probes, each DNA was labeled with [α-32P�dCTP, by random priming (Feinberg and Vogelstein, 1983). After hybridi-zation, the membrane was washed with 2× SSC, 0.1% SDS at RT for 15 min; 1× SSC, 0.1% SDS at RT for 15 min; and 0.1× SSC, 0.1% SDS at RT for 15 min. Hybridization signals were detected with a BAS-1500 image analyzer (Fuji, Japan) and exposed on Hyperfilm MP (Amersham, UK). RT-PCR analysis Gene-specific primers for CBF1/DREB1b, rice lip5 (AB011368), lip9 (AB011367), OsDhn1, OsWCOR413-like (AF283006), OsEnolase (U09450), and OsPeroxiredoxin (AF-203879) were used for PCR analysis. Specific primers for the coding region of each gene were as follows: CBF1/DREB1b-forward: 5′-AAGCTTAAAGAATCTACCTG-3′ CBF1/DREB1b-reverse: 5′-GGTACCCTCGTTTCTACAAC-3′ lip5-forward: 5′-CAAATTTCTCTTGAGAC-3′ lip5-reverse: 5′-ACGCCAGGATAATACAC-3′ lip9-forward: 5′-ATGGCCACTCCTGCTCCC-3′ lip9-reverse: 5′-CCAGCCCAAAACCAATACAA-3′ OsDhn1-forward: 5′-AGCTCAAACAAGTCAAGAGC-3′ OsDhn1-reverse: 5′-AAGCACCAAACTAACACACG-3′ OsWCOR413-like protein-forward: 5′-GAGGTGGAGGAGGC-TAG-3′ OsWCOR413-like protein-reverse: 5′-GGGGATAATCACAT-AAGAG-3′ OsEnolase-forward: 5′-GGTGATGAAGGTGGTTTTGC-3′ OsEnolase-reverse: 5′-GACCGGGAAGTCAATTACGG-3′ OsPeroxiredoxin-forward: 5′-CAGGCAGGCAACACAAG-3′ OsPeroxiredoxin-reverse: 5′-CGAAATTTTATTCGCAC-3′
As a control, we used the following primers specific for the rice actin gene RAc1 (McElroy et al., 1990): 5′-CATGCTAT-CCCTCGTCTCGACCT-3′ (forward) and 5′-CGCACTTCATG-ATGGAGTTGTAT-3′ (reverse). Ten micrograms of total RNA were used for RT-PCR analysis according to Takakura et al. (2000). PCR cycles numbered 30 for CBF1/DREB1b, 22 for lip5, 28 for lip9, 24 for OsDhn1, 35 for OsWCOR413-like, 25 for OsEnolase, 25 for OsPeroxiredoxin, and 22 for RAc1. Determination of chlorophyll fluorescence Chlorophyll fluo-rescence was measured with a Plant Efficiency Analyzer (Hansatech, UK) according to Jeong et al. (2002) with minor

Sang-Choon Lee et al. 109 �
modification. Approx. 5 cm-long segments of the youngest ex-tended leaves of 2-week-old or 2-month-old plants were either floated on distilled water at 4°C for 24 h or air-dried in a PEA clip in a 28°C incubator for 8 h. During stress, illumination was provided by white fluorescent lamps, at 250 µmol/m/s. After-wards the samples were returned to normal growth conditions for recovery. Fluorescence signals from each sample, which had been dark-adapted for 30 min, were measured at the indicated times. The ratio of Fv to Fm (Fv/Fm), representing the activity of photosystem II, was used to assess functional damage to the plants (Genty et al., 1989). Determination of electrolyte leakage Relative electrolyte leak-age after various periods of cold-stress treatment was measured as described by Lee et al. (1993). Briefly, leaves from 2-week-old and 2-month-old plants were cut into 3-cm segments and washed with distilled water. They were then floated on 20 ml of distilled water and placed under continuous light of 250 µmol/ m/s at 4°C. Solutions containing plant materials were autoclaved for 15 min at 120°C, frozen at −20°C for 30 min, then thawed in a 100°C water bath for 10 min. Total conductivity was deter-mined with a conductivity meter (VWR, UK). Relative electro-lyte leakage in the chilled leaves was calculated as the ratio of leakage conductivity to total conductivity. Determination of survival rate Plants were grown for about 2 weeks in the growth chamber (30°C, with light/dark cycle of 16/8 h) and then used to measure survival rate after stress treat-ment. For the cold-survival test, plants were transferred to a 4°C chamber for 4 d, and for the drought-survival test, they received no water for 4 d. Afterwards they were returned to normal growth conditions for 7 d and numbers of wilted and healthy plants were counted. Results Construction of the CBF1/DREB1b expression vector We isolated the Arabidopsis CBF1/DREB1b gene by PCR, using genomic DNA and gene-specific primers located in the UTR regions. A PCR fragment of 0.9 kb was cloned, and sequenced to confirm that it was the CBF1/DREB1b gene (Medina et al., 1999). This fragment was inserted into the KpnI/HindIII site of the binary vector pGA1611, placing the gene under the maize ubi promoter. The plas-mid was designated pSK146 (Fig. 1). Introduction of ubi-CBF1/DREB1b into rice The ubi-CBF1/DREB1b fusion construct was introduced into rice by co-cultivating scutellum calli with Agrobacterium that harbored pSK146. Among the 30 hygromycin-resistant plants that were regenerated, 17 grew to maturity and 13 died. DNA gel-blot analysis demonstrated that the chi- meric molecule was incorporated into the rice genome in nine plants (Fig. 2), with each carrying one to several
Fig. 1. Construction of pSK146. CBF1/DREB1b genomic DNA (0.9 kb) was cloned into the pBluescript II KS vector to produce pSK145, then subcloned into the HindIII/KpnI site of pGA1611, generating pSK146. PUbi, the maize ubiquitin promoter; P35S, the CaMV 35S promoter; Tnos, terminator sequence of the nopaline synthase gene; T7, terminator sequence of transcript 7; Hph, hygromycin phosphotransferase gene; BR and BL, right and left border sequences, respectively, of the Ti plasmid from A. tumefaciens. Fig. 2. DNA gel-blot analysis of CBF1/DREB1b in transgenic plants. Ten micrograms of genomic DNA from mature leaves was digested with HindIII and hybridized with radiolabeled CBF1/DREB1b probe. Positions and sizes of HindIII-digested lambda DNA are indicated on the left as size markers. copies of the transgene. Six of the transgenic plants had the dwarf phenotype, and four of them (C6, C8, C11, and C15) were sterile and did not set seeds. However, the re-maining two dwarf plants (C16 and C18) and three nor-mal-sized ones (C3, C4, and C13) produced fertile seeds.
Seeds of the transgenic plants were germinated on pa-per towels in the dark. After 5 d, the roots on transgenic plant C16 were 6.7 ± 2.0 cm long and those on plant C18 were 6.7 ± 1.6 cm long. The roots of neither plant grew straight downwards (Fig. 3). These dwarf lines also had shallow rooting at the harvest stage. In contrast, the non-
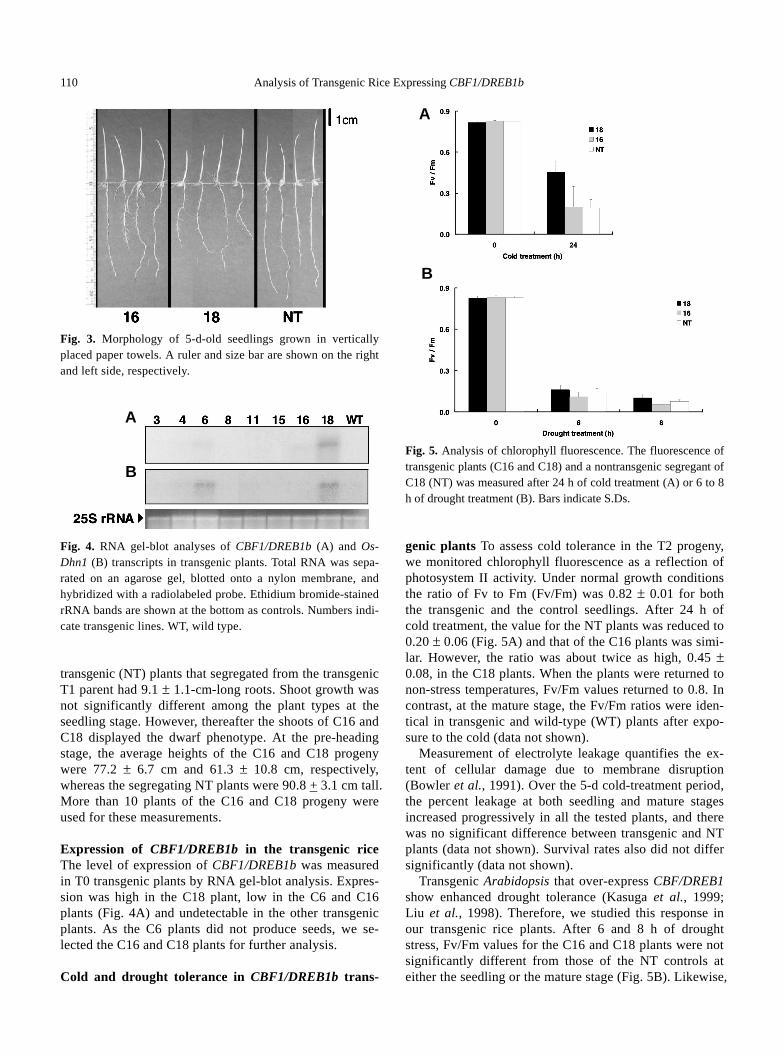
110 Analysis of Transgenic Rice Expressing CBF1/DREB1b �
Fig. 3. Morphology of 5-d-old seedlings grown in vertically placed paper towels. A ruler and size bar are shown on the right and left side, respectively.
A
B
Fig. 4. RNA gel-blot analyses of CBF1/DREB1b (A) and Os-Dhn1 (B) transcripts in transgenic plants. Total RNA was sepa-rated on an agarose gel, blotted onto a nylon membrane, and hybridized with a radiolabeled probe. Ethidium bromide-stained rRNA bands are shown at the bottom as controls. Numbers indi-cate transgenic lines. WT, wild type. transgenic (NT) plants that segregated from the transgenic T1 parent had 9.1 ± 1.1-cm-long roots. Shoot growth was not significantly different among the plant types at the seedling stage. However, thereafter the shoots of C16 and C18 displayed the dwarf phenotype. At the pre-heading stage, the average heights of the C16 and C18 progeny were 77.2 ± 6.7 cm and 61.3 ± 10.8 cm, respectively, whereas the segregating NT plants were 90.8 + 3.1 cm tall. More than 10 plants of the C16 and C18 progeny were used for these measurements. Expression of CBF1/DREB1b in the transgenic rice The level of expression of CBF1/DREB1b was measured in T0 transgenic plants by RNA gel-blot analysis. Expres-sion was high in the C18 plant, low in the C6 and C16 plants (Fig. 4A) and undetectable in the other transgenic plants. As the C6 plants did not produce seeds, we se-lected the C16 and C18 plants for further analysis. �
Cold and drought tolerance in CBF1/DREB1b trans-�
A �
�
�
�
�
�
�
�
�
B �
�
�
�
�
�
�
�
Fig. 5. Analysis of chlorophyll fluorescence. The fluorescence of transgenic plants (C16 and C18) and a nontransgenic segregant of C18 (NT) was measured after 24 h of cold treatment (A) or 6 to 8 h of drought treatment (B). Bars indicate S.Ds. genic plants To assess cold tolerance in the T2 progeny, we monitored chlorophyll fluorescence as a reflection of photosystem II activity. Under normal growth conditions the ratio of Fv to Fm (Fv/Fm) was 0.82 ± 0.01 for both the transgenic and the control seedlings. After 24 h of cold treatment, the value for the NT plants was reduced to 0.20 ± 0.06 (Fig. 5A) and that of the C16 plants was simi-lar. However, the ratio was about twice as high, 0.45 ± 0.08, in the C18 plants. When the plants were returned to non-stress temperatures, Fv/Fm values returned to 0.8. In contrast, at the mature stage, the Fv/Fm ratios were iden-tical in transgenic and wild-type (WT) plants after expo-sure to the cold (data not shown).
Measurement of electrolyte leakage quantifies the ex-tent of cellular damage due to membrane disruption (Bowler et al., 1991). Over the 5-d cold-treatment period, the percent leakage at both seedling and mature stages increased progressively in all the tested plants, and there was no significant difference between transgenic and NT plants (data not shown). Survival rates also did not differ significantly (data not shown).
Transgenic Arabidopsis that over-express CBF/DREB1 show enhanced drought tolerance (Kasuga et al., 1999; Liu et al., 1998). Therefore, we studied this response in our transgenic rice plants. After 6 and 8 h of drought stress, Fv/Fm values for the C16 and C18 plants were not significantly different from those of the NT controls at either the seedling or the mature stage (Fig. 5B). Likewise,

Sang-Choon Lee et al. 111 �
Fig. 6. Semiquantitative RT-PCR analyses of putative cold-responsive genes. Total RNAs from transgenic plants C16 and C18, and nontransgenic segregants (NT) were used as templates for RT-PCR. CBF1/DREB1b, Arabidopsis CRT/DRE binding factor1; lip5, lip9, and OsWCOR413-like, cold-responsive rice genes; OsEnolase and OsPeroxiredoxin, rice homologues of putative Arabidopsis CBF3/DREB1a target genes. Arrow indi-cates rice actin gene, RAc1. survival rates after drought stress did not differ (data not shown). Expression of cold-responsive genes in CBF1/DREB1b transgenic rice The CBF/DREB1 gene family is involved in the development of cold acclimation, during which dozens of cold-responsive genes are induced (Gilmour et al., 2000; Jaglo-Ottosen et al., 1998; Kasuga et al., 1999; Liu et al., 1998). We therefore tested whether the CBF1/ DREB1b transgene had any influence on the expression of cold-responsive genes in the rice. We have recently iden-tified a LEA (late-embryogenesis-abundant) class SK3 type-dehydrin, OsDhn1 that is inducible by cold and drought stresses (unpublished data). OsDhn1 is a homo-logue of wheat Wcor410 (Danyluk et al., 1994) and Arabidopsis cor47 (Welin et al., 1995), which are both known to be expressed during cold acclimation. We hy-bridized de-probed RNA blots (Fig. 4A) to radiolabeled OsDhn1 cDNA and found that the OsDhn1 transcript was present in the transgenic plants that expressed CBF1/ DREB1b (Fig. 4B), indicating that its expression is regu-lated by CBF1/DREB1b.
We also examined other cold-responsive genes in the CBF1/DREB1b transgenic plants. Expression of lip5 and dehydrin-homologous lip9 genes (Aguan et al., 1991) was induced in C18 but not in the control NT (Fig. 6). How-ever, the expression of another cold-inducible gene, OsWCOR413-like (de los Reyes et al., 2003), was un-changed in the transgenics. We also examined two EST sequences, OsPeroxiredoxin (AF203879) and OsEnolase (U09450) that are homologous to the Arabidopsis DRE-B1A target genes (Seki et al., 2001). Each was induced by 300 mM NaCl but not by cold, (data not shown) and their expression was not noticeably altered in the transgenic plants (Fig. 6). Based on these results we suggest that CBF1/DREB1b specifically induces OsDhn1, lip5, and
Fig. 7. RNA gel-blot analysis of lip5 expression. Ten micro-grams of total RNA from C16 and C18 transgenic plants, non-transgenic segregants (NT), and WT cold (12°C)-treated for the indicated times was separated, blotted, and hybridized with ra-dio-labeled lip5 probe. Ethidium bromide-stained rRNA bands are shown at the bottom as controls. lip9 in the transgenic rice.
In another experiment we examined whether expression of the cold-responsive gene lip5 was different in the transgenic plants than in cold-stressed WT plants. As shown in Fig. 7, lip5 expression was reduced in the trans-genic plants indicating that ectopic expression of CBF1/DREB1b is not sufficient to promote the expression of lip5 in rice, even though the latter gene is partially de-pendent on CBF1/DREB1b expression. Therefore, there must be some other mechanism for inducing lip5 in rice. Discussion The CBF/DREB1 transcription factors are major compo-nents of the ABA-independent cold signaling pathway in Arabidopsis (Shinozaki and Yamaguchi-Shinozaki, 2000). Plants with improved cold tolerance have been generated in Arabidopsis, tomato, and canola by ectopic expression of some of these genes (Gilmour et al., 2000; Hsieh et al., 2002b; Jaglo et al., 2001; Jaglo-Ottosen et al., 1998; Ka-suga et al., 1999; Liu et al., 1998).
To our knowledge, the research reported here is the first to describe transgenic expression of CBF genes in a monocot species. Our main aim was to investigate whether expression of the Arabidopsis CBF1/DREB1b gene could confer stress tolerance in rice. Transformation frequency was low and 13 out of 30 regenerated plants died before maturity. Among the 17 plants that grew to the reproductive stage, only 9 were true transgenics, an efficiency much lower than the 80 to 90% otherwise rou-tinely obtained in our laboratory. Most of the transfor-mants that died during regeneration probably succumbed to a toxic effect of the transgene, which is presumably detrimental in rice.
Six out of those nine transformants were dwarf; and four of the six were sterile. Expression of CBF/DREB1 also produces dwarfism in Arabidopsis and tomato (Gil-mour et al., 2000; Hsieh et al., 2002b; Kasuga et al., 1999; Liu et al., 1998). Liu et al. (1998) have suggested

112 Analysis of Transgenic Rice Expressing CBF1/DREB1b �
that this is due to constitutive overexpression of stress-inducible genes such as CORs. The introduction of a wheat DREB2A-homologue into rice plants also results in dwarfism, although the stress tolerance of such transfor-mants has not been examined (Shen et al., 2003). Evi-dently strong expression of CBF1/DREB1b is also detri-mental to growth in rice, as also manifested in the low frequency of transformation that we observed. Moreover, fewer regenerants were obtained than with vector alone in the control experiment.
We observed no obvious differences in cold or drought tolerance between the transgenic plants that expressed CBF1/DREB1b and the controls. This result differs from the reports of Hsieh et al. (2002a; 2002b) showing that when CBF1/DREB1b was introduced into cold-sensitive tomato plants, the transgenics were both chilling- and drought-tolerant. Therefore, the cold tolerance signaling pathway apparently differs between the dicot species and rice.
If the CBF/DREB1-mediated cold signaling pathway is indeed present in rice, we would expect to find its target genes in that species. However, there have been no re-ports of rice COR genes controlled by CBF/DREB1. Ho-mology searches in the public genome databases failed to identify homologues of RD29A/COR78, KIN1, KIN2/ COR6.6, and COR15A (20% cutoff value of homology at the amino acid level). Nevertheless, LIP9 and OsDHN1, which share > 37% homology with RD17/COR47 and EDR10, are present in the rice genome (unpublished data). Both rd17/cor47 and erd10 encode hydrophilic LEA pro-teins, or dehydrins, and their products are thought to func-tion during dehydration and cold stresses (Close, 1997; Thomashow, 1999). In addition, putative CBF3/DREB1a-target genes (Seki et al., 2001) have been searched in rice. Rice OsEnolase (U09450), OsWCOR413-like (AF283006), and OsPeroxiredoxin (AF203879) have been retrieved as homologues of enolase (X58107), the cold acclimation protein (AB044404), and peroxiredoxin TPX1 (AF121355), respectively. In our transgenic plants that expressed the CBF/DREB1 gene, transcripts of lip5, lip9, and OsDhn1 increased, but those of OsWCOR413-like, OsEnolase and OsPeroxiredoxin did not. These observations indicate that Arabidopsis CBF1/DREB1b only activates certain COR genes in rice.
CBF/DREB1-target genes contain the DRE/CRT ele-ment (A/GCCGAC) in their promoter regions (Seki et al., 2001). The rice lip5, lip9, and OsDhn1 genes possess one (-861), two (-331, -257), and two (-279, -205) DRE/CRT elements, respectively, in their 1-kb promoter regions. However, the promoter regions of OsEnolase, OsW-COR413-like, and OsPeroxiredoxin do not contain such elements. These latter genes probably fail to be induced in the CBF/DREB1-expressing plants because they lack DRE/CRT elements.
Although the mRNA level of lip5 increased when our
transgenic plants were grown under normal conditions, it was lower than in cold-treated WT plants. This suggests that, although the CBF/DREB1 gene functions in rice, it is not effective enough to confer stress tolerance. Dubouzet et al. (2003) have reported that rice CBF/DREB1 genes are functionally similar to Arabidopsis CBF/DREB1, al-though the specificity of their binding to the CRT/DRE element is somewhat different. Interestingly, transgenic Arabidopsis expressing 35S:OsDREB1A did not signifi-cantly up-regulate such COR genes as erd10 and kin2/ cor6.6. These results, together with our data, demonstrate that the CBF/DREB1 genes of Arabidopsis and rice are functionally different.
The fact that rice is much more sensitive to cold than Arabidopsis is probably due to the lack of rice homo-logues of some Arabidopsis CBF/DREB1 target genes. For example, the COR15a protein previously identified as cryoprotective (Steponkus et al., 1998) is not found in the rice database. Fowler and Thomashow (2002) have shown that more than 40 Arabidopsis genes are up-regulated by the constitutive expression of CBF/DREB1, and Rabbani et al. (2003) have reported 36 cold-induced genes in rice. However, most of the rice genes do not match the 40 up-regulated Arabidopsis genes. Future research is needed to determine whether this difference is responsible for the chilling-labile nature of rice. Cold tolerance is the result of complex mechanisms, including changes in membrane lipid composition, accumulation of compatible solutes such as sucrose and proline, and the expression of COR genes (Thomashow, 1999). The identification and analy-sis of each of these rice genes and comparative genomics involving other Gramineae species should greatly expand our understanding of the cold-response mechanism in rice. Acknowledgments This work was supported in part by grants from the Korea Research Foundation (KRF-1999-015-DP0283) and the Crop Functional Genomic Center of the 21st Century Frontier Program (CG1113 and CG1111). We thank Hong-Seok Choi, Yong-Souck Kang, Ji-Youn Kim, Kang Lee, and Min-Kyung Kim for technical assistance. References Aguan, K., Sugawara, K., Suzuki, N., and Kusano, T. (1991)
Isolation of genes for low-temperature-induced proteins in rice by a simple subtractive method. Plant Cell Physiol. 32, 1285−1289.
Bowler, C., Slooten, L., Vandenbranden, S., de Rycke, R., Bot-terman, J., Sybesma, C., van Montagu, M., and Inze, D. (1991) Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J. 10, 1723−1732.
Cattivelli, L. and Bartels, D. (1992) Biochemistry and molecular biology of cold-inducible enzymes and proteins in higher

Sang-Choon Lee et al. 113 �
plants. Soc. Exp. Biol. Semin. Ser. 49, 267−288. Chen, D. H. and Roland, P. C. (1999) A rapid DNA miniprepa-
ration method suitable for AFLP and other PCR applications. Plant Mol. Biol. Rep. 17, 53−57.
Close, T. J. (1997) Dehydrins: a commonality in the response of plants to dehydration and low temperature. Physiol. Plant. 100, 291−296.
Crosatti, C., Nevo, E., Stanca, A. M., and Cattivelli, L. (1996) Genetic analysis of the accumulation of COR14 proteins in wild (Hordeum spontaneum) and cultivated (Hordeum vul-gare) barley. Theor. Appl. Genet. 93, 975−981.
Danyluk, J., Houde, M., Rassart, É., and Sarhan, F. (1994) Dif-ferential expression of a gene encoding an acidic dehydrin in chilling sensitive and freezing tolerance gramineae species. FEBS Lett. 344, 20−24.
de los Reyes, B. G., Morsy, M., Gibbons, J., Varma, T. S., An-toine, W., McGrath, J. M., Halgren, R., and Redus, M. (2003) A snapshot of the low temperature stress transcrip-tome of developing rice seedlings (Oryza sativa L.) via ESTs from subtracted cDNA library. Theor. Appl. Genet. 107, 1071−1082.
Dubouzet, J. G., Sakuma, Y., Ito, Y., Kasuga, M., Dubouzet, E. G., Miura, S., Seki, M., Shinozaki, K., and Yamaguchi-Shinozaki, K. (2003) OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J. 33, 751−763.
Feinberg, A. P. and Vogelstein, B. (1983) A technique for radio-labeling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 132, 6−13.
Fowler, S. and Thomashow, M. F. (2002) Arabidopsis transcrip-tome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 14, 1675−1690.
Genty, B., Briantais, J.-M., and Baker, N. R. (1989) The rela-tionship between the quantum yield of photosynthetic elec-tron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 990, 87−92.
Gilmour, S. J., Zarka, D. G., Stockinger, E. J., Salazar, M. P., Houghton, J. M., and Thomashow, M. F. (1998) Low tem-perature regulation of the Arabidopsis CBF family of AP2 transcriptional activator as an early step in cold-induced COR gene expression. Plant J. 16, 433−442.
Gilmour, S. J., Sebolt, A. M., Salazar, M. P., Everard, J. D., and Thomashow, M. F. (2000) Overexpression of the Arabidopsis CBF3 transcriptional activator mimics multiple biochemical changes associated with cold acclimation. Plant Physiol. 124, 1854−1865.
Guy, C. L. (1990) Cold acclimation and freezing stress toler-ance: role of protein metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 41, 187−223.
Hiei, Y., Ohta, S., Komari, T., and Kumashiro, T. (1994) Effi-cient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 6, 271−282.
Hoekema, A., Hirsch, P. R., Hooykaas, P. J. J., and Schilperoort, R. A. (1983) A binary plant vector strategy based on separa-tion of vir- and T-region of the Agrobacterium tumefaciens Ti-plasmid. Nature 303, 179−180.
Hong, S. T., Lee, M. Y., and Kim, S. R. (2001) Molecular clon-
ing of the anther-preferential nonspecific lipid transfer pro-tein cDNA in hot pepper by mRNA differential display. Mol. Cells 11, 241−249.
Hoshida, H., Tanaka, Y., Hibino, T., Hayashi, Y., Tanaka, A., Takabe, T., and Takabe, T. (2000) Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast gluta-mine synthetase. Plant Mol. Biol. 43, 103−111.
Houde, M., Dhindsa, R. S., and Sarhan, F. (1992) A molecular marker to select for freezing tolerance in Gramineae. Mol. Gen. Genet. 234, 43−48.
Hsieh, T. H., Lee, J. T., Charng, Y. Y., and Chan, M. T. (2002a) Tomato plants ectopically expressing Arabidopsis CBF1 show enhanced resistance to water deficit stress. Plant Physiol. 130, 618−626.
Hsieh, T. H., Lee, J. T., Yang, P. T., Chiu, L. H., Charng, T. Y., Wang, T. C., and Chan, M. T. (2002b) Heterology expression of the Arabidopsis C-repeat/Dehydration Response Element Binding Factor 1 gene confers elevated tolerance to chilling and oxidative stresses in transgenic tomato. Plant Physiol. 129, 1086−1094.
Hughes, M. A. and Dunn, M. A. (1996) The molecular biology of plant acclimation to low temperature. J. Exp. Bot. 47, 291−305.
Jaglo, K. R., Kleff, S., Amundsen, K. L., Zhang, X., Haake, V., Zhang, J. Z., Deits, T., and Thomashow, M. F. (2001) Com-ponents of the Arabidopsis C-repeat/dehydration-responsive element binding factor cold-response pathway are conserved in Brassica napus and other plant species. Plant Physiol. 127, 910−917.
Jaglo-Ottosen, K. R., Gilmour, S. J., Zarka, D. G., Schabenber-ger, O., and Thomashow, M. F. (1998) Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 280, 104−106.
Jang, I. C., Oh, S. J., Seo, J. S., Choi, W. B., Song, S. I., Kim, C. H., Kim, Y. S., Seo, H. S., Choi, Y. D., Nahm, B. H., and Kim, J. K. (2003) Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth. Plant Physiol. 131, 516− 524.
Jeong, S. W., Choi, S. M., Lee, D. S., Ahn, S. N., Hur, Y., Chow, W. S., and Park, Y. I. (2002) Differential susceptibil-ity of photosynthesis to light-chilling stress in rice (Oryza sa-tiva L.) depends on the capacity for photochemical dissipa-tion of light. Mol. Cells 13, 419−428.
Kang, H.-G., Jeon, J.-S., Lee, S., and An, G. (1998) Identifica-tion of class B and class C floral organ identity genes from rice plants. Plant Mol. Biol. 38, 1021−1029.
Kasuga, M., Liu, Q., Miura, S., Yamaguchi-Shinozaki, K., and Shinozaki, K. (1999) Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-in-ducible transcription factor. Nat. Biotechnol. 17, 287−291.
Kim, S.-R., Lee, S., Kang, H.-G., Jeon, J.-S., Kim, K.-M., and An, G. (2003) A complete sequence of the pGA1611 binary vector. J. Plant Biol. 46, 211−214.
Larcher, W., Heber, U., and Santarius, K. A. (1973) Limiting temperatures for life functions; in Temperature and Life, Precht, H., Christopherson, J., Hensel, H., and Larcher, W. (eds.), pp. 195−291, Academic Press, New York.

114 Analysis of Transgenic Rice Expressing CBF1/DREB1b �
Lee, T. M., Lur, H. S., and Chu, C. (1993) Role of abscisic acid in chilling tolerance of rice (Oryza sativa L.) seedlings. I. Endogenous abscisic acid levels. Plant Cell Environ. 16, 481−490.
Lee, S., Jeon, J.-S., Jung, K.-H., and An, G. (1999) Binary vec-tors for efficient transformation of rice. J. Plant Biol. 42, 310−316.
Liu, Q., Kasuga, M., Sakuma, Y., Abe, H., Miura, S., Yamagu-chi-Shinozaki, K., and Shinozaki, K. (1998) Two transcrip-tion factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 10, 1391−1406.
McElroy, D., Rothernberg, M. M., and Wu, R. (1990) Structural characterization of a rice actin gene. Plant Mol. Biol. 14, 163−171.
Medina, J., Bargues, M., Terol, J., Perez-Alonso, M., and Salinas, J. (1999) The Arabidopsis CBF gene family is com-posed of three genes encoding AP2 domain-containing pro-teins whose expression is regulated by low temperature but not by abscisic acid or dehydration. Plant Physiol. 119, 463− 470.
Mohapatra, S. S., Wolfraim, L., Poole, R. J., and Dhindsa, R. S. (1989) Molecular cloning and relationship to freezing toler-ance of cold-acclimation-specific genes of alfalfa. Plant Physiol. 89, 375−380.
Rabbani, M. A., Maruyama, K., Abe, H., Khan, M. A., Katsura, K., Ito, Y., Yoshiwara, K., Seki, M., Shinozaki, K., and Ya-maguchi-Shinozaki, K. (2003) Monitoring expression pro-files of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA microarray and RNA gel-blot analyses. Plant Physiol. 133, 1755−1767.
Rédei, G. P. (1969) Arabidopsis thaliana (L.) Heynh. A review of the genetics and biology. Bibliograph. Genet. 21, 1−151.
Riechmann, J. L. and Meyerowitz, E. M. (1998) The AP2/ EREBP family of plant transcription factors. Biol. Chem. 379, 633−646.
Saijo, Y., Hata, S., Kyozuka, J., Shimamoto, K., and Izui, K. (2000) Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J. 23, 319−327.
Sakamoto, A., Alia, Murata, N., and Murata, A. (1998) Meta-bolic engineering of rice leading to biosynthesis of glycine-betaine and tolerance to salt and cold. Plant Mol. Biol. 38, 1011−1019.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor
Laboratory Press, Cold Spring Harbor, New York. Seki, M., Narusaka, M., Abe, H., Kasuga, M., Yamaguchi-
Shinozaki, K., Carninci, P., Hayashizaki, Y., and Shinozaki, K. (2001) Monitoring the expression pattern of 1300 Arabi-dopsis genes under drought and cold stresses by using a full-length cDNA microarray. Plant Cell 13, 61−72.
Shen, Y. G., Zhang, W. K., He, S. J., Zhang, J. S., Liu, Q., and Chen, S. Y. (2003) An EREBP/AP2-type protein in Triticum aestivum was a DRE-binding transcription factor induced by cold, dehydration and ABA stress. Theor. Appl. Genet. 106, 923−930.
Shinozaki, K. and Yamaguchi-Shinozaki, K. (2000) Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 3, 217−223.
Steponkus, P. L., Uemura, M., Joseph, R. A., Gilmour, S. J., and Thomashow, M. F. (1998) Mode of action of the COR15a gene on the freezing tolerance of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 95, 14570−14575.
Stockinger, E. J., Gilmour, S. J., and Thomashow, M. F. (1997) Arabidopsis thaliana CBF1 encodes an AP2 domain-contain-ing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcrip-tion in response to low temperature and water deficit. Proc. Natl. Acad. Sci. USA 94, 1035−1040.
Takakura, Y., Ito, T., Saito, H., Inoue, T., Komari, T., and Ku-wata, S. (2000) Flower-predominant expression of a gene en-coding a novel class I chitinase in rice (Oryza sativa L.). Plant Mol. Biol. 42, 883−897.
Thomashow, M. F. (1994) Arabidopsis thaliana as a model for studying mechanisms of plant cold tolerance; in Arabidopsis, Meyerowitz, E. and Somerville, C. (eds.), pp. 807−834, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Thomashow, M. F. (1999) Plant cold acclimation: freezing tol-erance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50, 571−599.
Toenniessen, G. H. (1991) Potentially useful genes for rice ge-netic engineering; in Rice Biotechnology, Khush, G. S. and Toenniessen, G. H. (eds.), pp. 253−280, International Rice Research Institute and Wallingford (United Kingdom), CAB International, Manila.
Wang, C. Y. (1990) Chilling Injury of Horticultural Crops, CRC Press, Boca Raton, FL, USA.
Welin, B. V., Olson, A., and Palva, E. T. (1995) Structure and organization of two closely related low-temperature-induced dhn/lea/rab-like genes in Arabidopsis thaliana L. Heynh. Plant Mol. Biol. 29, 391−395.















![A Drought-Inducible Transcription FactorA Drought-Inducible Transcription Factor Delays Reproductive Timing in Rice1[OPEN] Chunyu Zhang2, Jun Liu2,TaoZhao2, Adam Gomez, Cong Li, Chunsheng](https://img.dokumen.tips/doc/110x75/5f0576657e708231d4131579/a-drought-inducible-transcription-a-drought-inducible-transcription-factor-delays.jpg)













