Embed Size (px)
Citation preview

1
ECOLOGY AND BIOLOGY OF GYRFALCONS IN GREENLAND
KURT K. BURNHAM1,2,3 AND WILLIAM A. BURNHAM2,^
1Edward Grey Institute of Field Ornithology, Department of Zoology, University of Oxford,South Parks Road, Oxford, OX1 3PS, UK
2The Peregrine Fund, 5668 West Flying Hawk Lane, Boise, ID 83709, USA
3High Arctic Institute, 603 10th Avenue, Orion, IL 61273, USA (current address). E-mail: [email protected]
^Deceased
ABSTRACT.—We studied two breeding populations of Gyrfalcons (Falco rusticolus) approximately1,200 km apart in Greenland. During 1998–2006, we collected data from a population in the cen-tral-west (Kangerlussuaq), and during 1994–2006 and 2008–2010 from a population in the north-west (Thule). Gyrfalcons in Kangerlussuaq bred approximately 18 days earlier than those in Thule,with egg laying, on average, starting on 17 April and 5 May, respectively. Reproduction was sim-ilar between areas, with no significant difference between the number of young produced at occu-pied or successful nests. Years in which the Kangerlussuaq population bred later showed fewersuccessful nests, while the lay-date advanced significantly for the Thule population during thisstudy. Diet varied between areas, with the more coastal population in Thule feeding primarily onDovekies (Alle alle) and the more inland Kangerlussuaq population feeding primarily upon RockPtarmigan (Lagopus muta) and passerines. Plumage color in Thule was > 99% white, while nearlyequal numbers of white and grey adults were observed in Kangerlussuaq. No changes in plumagecolor were observed in Kangerlussuaq from the 1950s onward; however, an unexplained differ-ence in plumage color between sexes was documented during this study. The Gyrfalcon populationin Thule appeared relatively stable, whereas the Kangerlussuaq breeding population was likelycyclical, and possibly declining since the early 1990s. Based on the relatively long-term data setsavailable, these populations are ideal for continued monitoring to determine what effects futureclimate change may have on Gyrfalcon populations. Received 6 April 2011, accepted 20 July 2011.
BURNHAM, K. K., AND W. A. BURNHAM. 2011. Ecology and biology of Gyrfalcons in Greenland.Pages 1–20 in R. T. Watson, T. J. Cade, M. Fuller, G. Hunt, and E. Potapov (Eds.). Gyrfalconsand Ptarmigan in a Changing World, Volume II. The Peregrine Fund, Boise, Idaho, USA.http://dx.doi.org/ 10.4080/gpcw.2011.0209
Key words: Gyrfalcon, Greenland, Peregrine Falcon, nesting chronology, cyclic population, prey,reproduction.
1

GYRFALCONS (Falco rusticolus) have histori-cally been reported breeding along much ofGreenland's ice-free coastlines from PearyLand (83° N: Manniche 1910, Koch 1925,Johnsen 1953) to Kap Farvel (60° N: Rafn1933, Salomonsen 1950) at the southern tip(Figure 1). Most records were of sightingsmade and specimens collected prior to 1950.In the Zoological Museum, Copenhagen, Den-mark, there are more than 429 Gyrfalcon spec-imens that were collected in Greenland alone,primarily to investigate variation in plumagecolor. Gyrfalcons were commonly shot and/ornoted during polar expeditions and collectingtrips (e.g., Sabine 1819, Walker 1860, Hall1876, Hagerup 1891, Stone 1895, Chapman1899). Explorers and collectors documentedlocations where falcons were observed breed-ing or migrating and helped to establish therange and migration timing of the species inGreenland.
Beyond these records from early explorers, lit-tle information exists on Gyrfalcons in Green-land other than the occasional anecdotalmention in the literature and observations col-lected on individual nests or over small por-tions of a breeding season (e.g., Fletcher andWebby 1977, Fowles 1981, Cabot et al. 1988,Booms and Fuller 2003). Not until the early1970s did investigations of Gyrfalcon ecologyin Greenland begin in the Low Arctic, andeven then, the Gyrfalcon was of secondaryinterest to the Peregrine Falcon (Falco pere-grinus) (Burnham and Mattox 1984). Onlyduring one four-year period (1990–93) didsystematic surveys for nesting Gyrfalcons takeplace (central-west Greenland); however,results from these surveys were difficult tointerpret (W. Mattox, unpubl. data, M. Yatesunpubl. data). In High Arctic Greenland (~70°N or more, see Bliss 1979), summer informa-tion was restricted to notes made by earlyexplorers, birding surveys/expeditions, or sin-gle seasons spent observing particular nests(e.g., Stone 1895, Cabot et al. 1988). In areview article about the Gyrfalcon, Cade et al.(1998) commented that “fresh data from
Greenland and Russia lag far behind” thatfrom other locations.
To address this lack of knowledge, we collectedlong-term data from both Low and High Arcticbreeding populations in central-west (Kanger-lussuaq) and northwest (Thule) Greenland,respectively (Figure 1). We compared databetween areas and considered historical datawhere available. We analyzed changes inbreeding biology over time to look for potentialshifts possibly associated with climate change.
METHODS
We studied populations of breeding Gyrfalconsin central-west (Kangerlussuaq, 66.5–67.5° N)Greenland from 1994–2006 and northwest(Thule, 75.9–77.6° N) Greenland from 1994–2005 and 2006–2010 (Figure 1). These loca-tions are geographically separated byapproximately 1,170 km with limited or noice-free land existing between the two sites.
The Kangerlussuaq study area is consideredLow Arctic and is located in one of the largestice-free areas within Greenland. The area isbisected by numerous lakes, rivers, streams,and rolling valleys (Figure 2). Vegetation isprimarily sedges and grasses, with willow(Salix) up to 2 m in height. Small passerines,for example, Lapland Longspurs (Calcariuslapponicus), Snow Buntings (Plectrophenaxnivalis), Northern Wheatear (Oenantheoenanthe), and others, occur in abundance,with Rock Ptarmigan (Lagopus muta), water-fowl, and shorebirds also present. For addi-tional information on the Kangerlussuaqstudy area, see Burnham and Mattox (1984)and Burnham (2008).
Further north, the Thule study area is almostentirely coastal, with the largest ice-free marginextending approximately 26 km from the sea.The area is considered to be High Arctic, withsparse, prostrate vegetation. Cliffs are almostalways sea-facing (Figure 2). The most numer-ous bird species in the Thule area is the
2
– BURNHAM AND BURNHAM –

3
– GYRFALCON BIOLOGY IN GREENLAND –
Dovekie (Alle alle), which has an estimatedbreeding population of 33 million pairs in theThule area (Egevang et al. 2003). Additionally,Thick-billed Murres (Uria lomvia), Black-legged Kittiwakes (Rissa tridactyla), and BlackGuillemots (Cepphus grille) occur in largenumbers, with lesser numbers of passerines andRock Ptarmigan. For further information on theThule study area, see Burnham (2008).
In the Kangerlussuaq area, most Gyrfalconnest sites were previously located by Burnhamand Mattox (1984), M. Yates (unpubl. data),and W. Mattox (unpubl. data), with most dis-covered incidentally while surveying for Pere-grines. New nest sites were also located duringthis study while traveling between known nest-ing sites. In addition to an historic literature
search, individuals with first-hand knowledgeof birds in Thule were contacted. Locationsidentified by local hunters and those found inthe literature were first surveyed, with addi-tional nesting locations found during system-atic surveys of the coastline.
Occupied sites were those where one or moreterritorial adults and/or young were observed.To be considered successful, one or morechicks had to be observed. If only adults werepresent, they had to be seen on multiple visitsto the site or to act territorially towardresearchers to be considered as occupants. Inthe case of breeding, nests were rappelled orclimbed into, prey remains were identified, andyoung were banded. Age of young was deter-mined by counting backwards from the esti-
KangerlussuaqStudy Area
Thule Study Area
DiskoBay
Uummannaq Area
Peary Land
Kap Farvel
BaffinBay
N
0 km 600 kmFigure 1. Map of Greenland showing study areasand locations mentioned in the text.
Figure 2. Photos of the Kangerlussuaq (A) andThule (B) study areas showing typical habitat.

mated average age of nestlings or fledglings.We assumed a four-egg clutch with an egg laidevery other day, with incubation beginning onthe third egg and a 35-day incubation period(C. Sandfort pers. comm., Platt 1977). Pooleand Bromley (1988) reported that malesfledged at 45–47 days and females at 47–50days. We considered 47 days as an averagefledging age.
While multiple nests on the same section ofcliff (up to 0.5 km long) were considered to bethe same nest site it is apparent that some Gyr-falcon nesting territories have alternate nestsites. Seven territories in Kangerlussuaq hadmultiple adjacent cliffs with nest sites (one tofour) that were never used simultaneously dur-ing the entire study. This makes a strong casethey were alternative nest sites within the sameterritories, although this cannot be proven withcertainty as the adults were not banded. If true,the total number of known nesting territoriesavailable for breeding would be greatlyreduced and nest sites could be groupedtogether by territory for analyses.
Depending upon weather conditions, we vis-ited nests with breeding Gyrfalcons more thanonce per season to record the number ofnestlings/fledglings. However, remote loca-tions and inclement weather frequently pre-cluded follow-up visits. We banded Gyrfalconswith a standard Danish numbered lock-onband on one leg and an alpha-numeric band(ACRAFT, Edmonton, Canada) on the other.
RESULTS
Surveys and Occupancy.—Surveys for breed-ing Gyrfalcons took place from 1998–2006 inKangerlussuaq and from 1993–2006 and2008–2010 in Thule. Results from Kangerlus-suaq and Thule surveys should be consideredminimal estimates, with some nests likely fail-ing before they were checked the first time,resulting in a bias towards successful nests.Surveys in Kangerlussuaq were primarily con-ducted by helicopter and occurred in early-to-
mid-June when young were usually around 20days old. Surveys in Thule typically occurredfrom mid-to-late July through mid-August,when young were frequently at least 30 daysold and in some cases had already left the nest.Surveys in Thule were usually conducted byboat, and due to the late break-up of oceanpack-ice, earlier surveys were not possible.
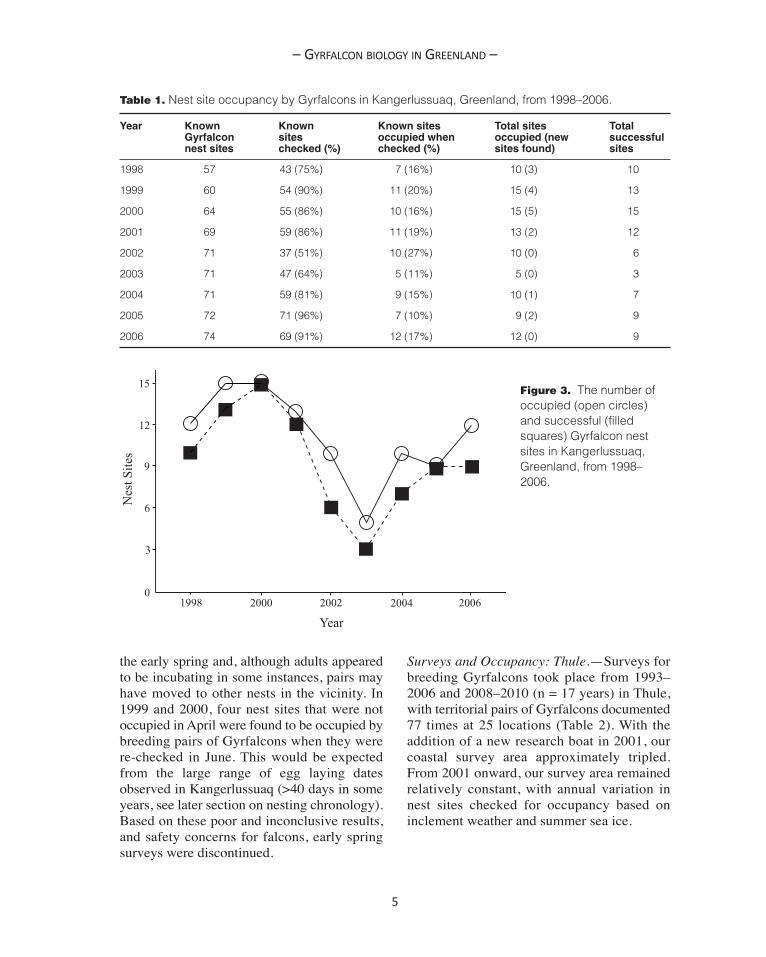
Surveys and Occupancy: Kangerlussuaq.—From 1998–2006 (n = 9), 51–96% (mean =80%) of known Gyrfalcon nesting locationswere checked for occupancy in Kangerlussuaq(Table 1). During this period territorial Gyrfal-cons were documented on 99 occasions and anadditional 17 new nesting sites were located,giving a total of 74 known sites. The occu-pancy rate at previously known nest sitesranged from 10–27% (mean = 17%). Whenincluding new nest sites, a high of 15 siteswere occupied in 1999 and 2000, and a low offive in 2003 (Table 1, Figure 3).
While it is possible that sites with single, non-breeding pairs, or failed pairs of Gyrfalconswere missed, most locations where reproduc-tion occurred were probably discovered. From1998–2000, a limited number of nest sites sur-veyed by helicopter were also surveyed byother falcon researchers on foot during thesame period. In total, 51 known Gyrfalconnests were surveyed by both methods, withresults between helicopter and foot surveysbeing the same.
Little information is available on the failure rateof Gyrfalcon nests in Kangerlussuaq during thisstudy. However, in 1999 and 2000, surveys ofknown Gyrfalcon nest sites were conducted inthe early spring (April, when egg layingoccurred) in an attempt to determine the pro-portion later failing, with eight and six nestsfound occupied in the two years, respectively.Of those nests found occupied in the earlyspring, four were no longer occupied at a repeatsurvey in June 1999, and one was no longeroccupied at a repeat survey in June 2000. Eggswere not observed at any of these nest sites in
4
– BURNHAM AND BURNHAM –
5
– GYRFALCON BIOLOGY IN GREENLAND –
the early spring and, although adults appearedto be incubating in some instances, pairs mayhave moved to other nests in the vicinity. In1999 and 2000, four nest sites that were notoccupied in April were found to be occupied bybreeding pairs of Gyrfalcons when they werere-checked in June. This would be expectedfrom the large range of egg laying datesobserved in Kangerlussuaq (>40 days in someyears, see later section on nesting chronology).Based on these poor and inconclusive results,and safety concerns for falcons, early springsurveys were discontinued.
Surveys and Occupancy: Thule.—Surveys forbreeding Gyrfalcons took place from 1993–2006 and 2008–2010 (n = 17 years) in Thule,with territorial pairs of Gyrfalcons documented77 times at 25 locations (Table 2). With theaddition of a new research boat in 2001, ourcoastal survey area approximately tripled.From 2001 onward, our survey area remainedrelatively constant, with annual variation innest sites checked for occupancy based oninclement weather and summer sea ice.
Table 1. Nest site occupancy by Gyrfalcons in Kangerlussuaq, Greenland, from 1998–2006.
Year Known Known Known sites Total sites Total Gyrfalcon sites occupied when occupied (new successful nest sites checked (%) checked (%) sites found) sites
1998 57 43 (75%) 7 (16%) 10 (3) 10
1999 60 54 (90%) 11 (20%) 15 (4) 13
2000 64 55 (86%) 10 (16%) 15 (5) 15
2001 69 59 (86%) 11 (19%) 13 (2) 12
2002 71 37 (51%) 10 (27%) 10 (0) 6
2003 71 47 (64%) 5 (11%) 5 (0) 3
2004 71 59 (81%) 9 (15%) 10 (1) 7
2005 72 71 (96%) 7 (10%) 9 (2) 9
2006 74 69 (91%) 12 (17%) 12 (0) 9
15
12
9
6
3
01998 2000 2002 2004 2006
Year
Nes
t Site
s
Figure 3. The number ofoccupied (open circles)and successful (filledsquares) Gyrfalcon nestsites in Kangerlussuaq,Greenland, from 1998–2006.

During surveys for nesting Gyrfalcons inThule, few if any occupied nests were likely tohave been missed. Adults and juveniles werealmost exclusively bright white in plumage andin general were obvious to the unaided eyewhen observed on cliffs, even at long distances.From 1993 to 2003, no obvious use of alterna-tive nest sites occurred, and all breeding tookplace on ledges or in potholes. Beginning in2004 and continuing to 2010, it appeared likelythat pairs began using alternate nesting siteswithin breeding territories; however, adultswere not ringed and it was not possible to deter-mine the specific movements of individuals.During this period, the use of Northern Raven(Corvus corax) nests by Gyrfalcons in theThule area was observed for the first time, withmultiple raven nests used in most years after2004. The increase of new nesting sites after2004 was primarily a result of these raven nests
being used for nesting by Gyrfalcons, and inalmost all instances, previously used ledges orpotholes were within several kilometers.
Results from the Thule area provide the firstdata on the local Gyrfalcon population, butdespite 17 years of research, only limited con-clusions can be drawn on occupancy. Not until2001 did surveys begin on a large enough scaleto provide data on a number of nests each year.It appears that the Thule population was rela-tively stable, apart from 2003 and 2006, whenonly two and five occupied sites were found,respectively. The indicated low number ofoccupied nests in 2003 resulted from fewerpreviously known nest sites surveyed (seeTable 2), and it does not appear that the popu-lation was peaking or crashing at any pointduring this study.
6
– BURNHAM AND BURNHAM –
Table 2. Nest site occupancy by Gyrfalcons in Thule, Greenland, from 1993–2006 and from 2008–2010.
Year Known Known sites Known sites Sites observed Observed Gyrfalcon checked occupied when occupied (new successfulnest sites (%) checked (%) sites found) sites
1993 – – – – 0
1994 – – – (1) 1
1995 1 1 (100%) 1 (100%) 2 (1) 2
1996 2 1 (50%) 1 (100%) 1 (0) 0
1997 2 2 (100%) 2 (100%) 2 (0) 2
1998 2 2 (100%) 2 (100%) 2 (0) 1
1999 2 2 (100%) 1 (50%) 3 (2) 2
2000 4 3 (75%) 3 (100%) 3 (0) 2
2001 4 4 (100%) 1 (25%) 4 (3) 4
2002 7 7 (100%) 6 (86%) 8 (2) 8
2003 9 4 (44%) 1 (25%) 2 (1) 2
2004 10 10 (100%) 7 (70%) 9 (2) 9
2005 12 12 (100%) 6 (50%) 11 (5) 8
2006 17 16 (94%) 5 (31%) 5 (1) 5
2008 18 16 (89%) 6 (38%) 8 (2) 7
2009 20 16 (80%) 5 (31%) 7 (2) 7
2010 22 14 (64%) 5 (36%) 8 (3) 8

7
– GYRFALCON BIOLOGY IN GREENLAND –
Occupancy of nest sites is difficult to comparebetween Kangerlussuaq and Thule. Within theKangerlussuaq area, the majority of Gyrfalconnest sites were documented by others prior tothis study, which provided us with significantoverall knowledge of the study area. In Thule,we had no previous knowledge of nest sitelocations, and each year required new andextended surveys.
Reproduction: Kangerlussuaq.—During 1998–2006, a minimum of 236 young were producedat the 99 occupied Gyrfalcon nest sites locatedin Kangerlussuaq (2.38 young per occupiedsite). At successful sites (n = 84), where at leastone chick was observed, an average of 2.81young were produced, with the number ofyoung per nest ranging between one and five.
The numbers presented here are minima as itwas frequently impossible to ascertain the totalnumber of young. In some cases, chicks weretoo young, too near fledging, or cliffs were toooverhung for the nest to be entered safely.When young were small, less than one weekold, adult females frequently did not leave thenest when approached by helicopter. Observa-tions were thus limited to a mass of nestlingdown under the front of the female’s breast, orin some cases, nothing was visible under thefemale, in which case, we noted the female asincubating/brooding but did not count the nestas successful. To some extent, this explains thedifference between occupied (n = 99) and suc-cessful (n = 84) nests, with some nests havingnot actually failed, but reproduction beingunknown. Given the wide range of nestingdates (some years up to 42 days between nests,
Table 3. Nesting chronology of Gyrfalcons in the Kangerlussuaq (1998–2006) and Thule (1994–2010)study areas, Greenland. Hatch-dates were calculated from estimated age of young observed in thenest (see text), with data included from 66 nests in Kangerlussuaq and 40 in Thule.
Kangerlussuaq mean (min–max, median) Thule mean (min–max, median)
Lay-date 17 Apr (28 Mar–9 May, 18 Apr) 5 May (24 Apr–24 May, 4 May)
Hatch-date 26 May (6 May–17 Jun, 27 May) 13 Jun (2 Jun–2 Jul, 12 Jun)
Fledge-date 12 Jul (22 Jun–3 Aug, 13 Jul) 30 Jul (19 Jul–18 Aug, 29 Jul)
201020082006200420022000199819961994
136
131
126
121
116
Julia
n da
te200620052004200320022001200019991998
121
116
111
106
101
96
Year
Julia
n da
te
A
B
Figure 4. The mean (±SE) of hatching datesrecorded in Kangerlussuaq (A) from 1998 to2006 and in Thule (B) from 1994–2006 and2007–2010. No data available from Thule for1996, 1998, and 2007.

see section below on nesting chronology), thesenumbers should be viewed with caution, as asizeable portion of chicks were very youngwhen their age was noted and some proportionlikely did not survive to fledging. Numbersgiven here for productivity should not be inter-preted as numbers of chicks fledged.
Reproduction: Thule.—Using the same criteriawe used for sites being occupied and success-ful in the Kangerlussuaq area, we located 77occupied territories in Thule that producedover 179 chicks, for an average of at least 2.38young per occupied nest. At the 69 successfulsites, an average of 2.63 young were produced.The number of young produced at individualnests ranged from one to five, with young overhalf-grown at all but one of the successful nestsites when first visited. Young in Thule wererarely banded, with young being too nearfledging to safely enter the nest, or already fly-ing in many cases. No females were observedincubating during surveys in Thule.
Nesting Chronology.—Gyrfalcons in Kanger-lussuaq began nesting approximately 18 daysearlier than those in Thule, with the first egglaid on 17 April (SD ± 9.3) and 5 May (SD ±7.0), respectively (Table 3, Figure 4). Calculat-ing forward, young departed the nest around 12July in Kangerlussuaq and 30 July in Thule(Table 3). We were unable to determine the ageof young at 22 nests in Thule as they hadalready fledged. The majority of these nests(77%) were surveyed from early Augustonward (after the mean fledging date), but datafrom these nests would have likely had littleimpact on calculated nesting chronology. Basedon research in Canada and Iceland (Cramp andSimmons 1980, Nielsen and Cade 1990,respectively), showing that young Gyrfalconsbecome independent of adult care at an averageof ≥4 weeks after they left the nest, independ-ence would be reached in mid-August inKangerlussuaq and early September in Thule.
In Kangerlussuaq, a significant relationship (P< 0.05, r2 = 0.48) existed between the number
of successful nests found each year and theaverage date that breeding was initiated; fewerpairs of Gyrfalcons produced young in years oflate breeding (Figure 5). No significance wasfound between the total number of occupiednests and timing of breeding (P > 0.05, r2 =0.27). In Thule, from 1994–2010 (no data for1996, 1998, and 2007), the average lay-datefor the Gyrfalcon population significantlyadvanced (P < 0.05, r2 = 0.32, Figure 6), withthe average lay-date for the final year of thestudy being six days earlier than for the entire17-year study period as a whole (29 April vs.5 May).
In general, Gyrfalcons in Kangerlussuaqshowed a much wider range of dates duringwhich reproduction occurred. In Kangerlus-suaq, fledging dates spanned a 43-day period(22 June–3 August) compared to a 31-dayperiod in the more northern Thule area (19July–18 August) (Table 3). In one instance inKangerlussuaq, two nests were visited on thesame day (within 20 km of each other), withone nest containing pipped/hatching eggs andthe other nest having young that were almostfully feathered and ready to depart the nest(approximate 42-day difference).
Prey.—Jenkins (1978), Burnham and Mattox(1984), and Booms and Fuller (2003) havedocumented prey species delivered to Gyrfal-con nest sites in Kangerlussuaq. It was deter-mined that Gyrfalcons prey on virtually allsmall- to medium-size birds and mammals inthe area. During this study, ptarmigan remainswere the most common prey item in Gyrfalconnests, e.g., one 2005 nest, with four, approxi-mately 38-day-old nestling Gyrfalcons, con-tained 55 ptarmigan and 2 Long-tailed Duck(Clangula hyemalis) sterna. Arctic Hare(Lepus arcticus), Snow Bunting, and LaplandLongspur remains were also found, but inmuch less frequency than ptarmigan remains.Burnham and Mattox (1984) emphasized theapparent importance of passerines (from 1972to 1981) as food to supplement ptarmigan andhare when their numbers were few.
8
– BURNHAM AND BURNHAM –

9
– GYRFALCON BIOLOGY IN GREENLAND –
In Thule, prey remains were collected at 40Gyrfalcon nest sites during the 17-year study.Dovekie remains were present in 75% of thenests. Fourteen nests had exclusively Dovekieremains, the only prey species for which thatwas the case. Rock Ptarmigan remains werefound at 12 nests (30%), Black Guillemot at 10(25%), Arctic Hares at 6 (15%), Long-tailedDucks at 5 (13%), Black-legged Kittiwakes at3 (8%), passerines at 2 (5%), and CommonEiders (Somateria mollissima), Snow Geese(Chen caerulescens), and Thick-billed Murresat 1 (3%). Although prey transects were notconducted, there was an obvious positive cor-relation between potential prey seen in theareas of nest sites and prey species remains in
nests. For example, one nest site locatedapproximately 60 km up a long fjord and fur-ther from any breeding seabird colony yieldedmainly Arctic Hare leverets, Rock Ptarmigan,and passerine remains.
Plumage Color Variants.—During 1998–2006in Kangerlussuaq, the plumage color of 137adults was recorded as 71 (52%) “white,” 16(12%) “silver,” and 50 (36%) “grey.” Observa-tions of adult females at nests were more fre-quent (80 vs. 57) than males, as might beexpected (Table 4). The ratio of white and greyfemales was nearly even (43% vs. 41%), but65% of adult males were white compared toonly 30% grey.
0
4
8
12
16
141 143 145 147 149 151 153 155
Julian date
Prod
uctiv
e ne
sts
Figure 5. Significantrelationship (P > 0.05, r2 =0.48) between the averagedate of breeding (Julianhatch-date shown here) andthe number of successfulnests. This suggests that late-breeding Gyrfalcons producefewer broods and that overallproduction is lower in yearswhen the population as awhole nests later.
201020072004200119981995
176
172
168
164
160
Year
Julia
n da
te
Figure 6. Negative trend overtime showing that the averagedate breeding was initiated inThule significantly (P < 0.05, r2
= 0.32) advanced (becameearlier) during the 17-yearstudy period. Julian hatch-date shown here.

More than 285 juvenile and adult Gyrfalconswere observed during 1993–2010 in Thule, ofwhich only two had grey plumage (an adultfemale and her daughter), and one silver. Theremainder of the Gyrfalcons had very whiteplumages and many showed no visible dark pig-mentation on their tails, even as juveniles.Repeat sightings may have occurred from oneyear to the next in both study areas as few adultswere ringed and it was not possible to tell indi-viduals apart. Color phases of individuals wereexpected to remain consistent throughout theirlives, as evident from captive birds.
DISCUSSION
The world population of Gyrfalcons was esti-mated by Cade (1960, 1982) at 15,000–17,000pairs. More recently Potapov and Sale (2005)estimated the world population at 7,880–10,900 pairs by totaling country estimates. ForGreenland alone, Mattox (1970) estimated 850pairs, Cade (1982) 500–1,000 pairs, and mostrecently Boertmann (2008) estimated the pop-ulation to be no more than 500 pairs. Based onthese estimates, Greenland contains a possible5–10% of the world Gyrfalcon population.
One of the most important influences on thebreeding density of birds is food supply (Lack1954, 1966, Newton 1979, 1998). Ratcliffe(1969) showed that for three inland PeregrineFalcon populations in the United Kingdomthe average breeding home range per pair foreach area was correlated with primary-pro-ductivity and food. Numerous other raptorspecies have shown significant relationships
between prey abundance and nesting density(see Newton 1979).
Rock Ptarmigan have been shown to havecyclic population fluctuations throughout theArctic (Gudmundsson 1960, Weeden and The-berge 1972, Watson et al. 1998, Moss and Wat-son 2001), and specifically in Greenland(Salomonsen 1950). Based on hunter kills,Vibe (1967) found ptarmigan populations tofluctuate in Greenland on approximately 11-year cycles from 1886 to 1954. Nielsen (1999)reported a 10-year cycle for ptarmigan in Ice-land and found that Gyrfalcon populationslagged behind by 3 years. Mossop and Hays(1982, 1994) reported that, in the Yukon Terri-tory, the Gyrfalcon breeding population fluc-tuated on a 10-year cycle, probably followingpopulation fluctuations of Willow Grouse(Lagopus lagopus). Additionally, Nyström etal. (2005) showed a significant relationshipbetween the number of Gyrfalcon chicksfledged each year and ptarmigan populationlevels. Alternatively, Mindell et al. (1987) andMindell and White (1988) found no relation-ship between Gyrfalcon and ptarmigan popu-lations in Alaska, and Shank and Poole (1994)found the same for Canada’s Northwest Terri-tories (but see Mossop 2011, Barichello andMossop 2011).
In Kangerlussuaq, where ptarmigan remainswere found most frequently in Gyrfalcon nestsfrom 1998–2006, it is difficult to knowwhether a response relationship between Gyr-falcons and ptarmigan exist. However, basedon our data, it appears a peak in occupied Gyr-falcon nest sites occurred in 1999 and 2000,followed by a steady decline and then increase(Figure 3). This pattern would be expected ifindeed the Gyrfalcon population was fluctuat-ing with a cyclic ptarmigan population. Unfor-tunately, no recent information is available onthe annual fluctuations of the ptarmigan popu-lation in Greenland, with historic data based oncasual observations, local inhabitants, andhunter kills (Salomonsen 1950, Vibe 1967).
10
– BURNHAM AND BURNHAM –
Table 4. Plumage color of adult Gyrfalconsobserved in Kangerlussuaq from 1998–2006.
Adult females Adult males
White 34 (43%) 37 (65%)
Silver 13 (16%) 3 (5%)
Grey 33 (41%) 17 (30%)
Total 80 57

11
– GYRFALCON BIOLOGY IN GREENLAND –
Prey differences between Kangerlussuaq andThule were apparent primarily in the largenumbers of songbirds in Kangerlussuaq frommid-May to mid-September, and in the tens ofmillions of seabirds present in northwestGreenland from mid-May to August–Septem-ber. In general, Gyrfalcons appeared to preyupon all suitable species present locally in theirbreeding areas.
In both Kangerlussuaq and Thule, it seemsprobable that Gyrfalcon numbers, or nestingdensity, were regulated to prey, and specificallyptarmigan. Of primary importance is the preyavailability when the Gyrfalcons first arrive onterritory in spring; Rock Ptarmigan, SnowBuntings, and adult Arctic Hares are present inKangerlussuaq, and Rock Ptarmigan and adultArctic Hares in Thule. It is likely that RockPtarmigan are the single most important preyspecies early in the season, as shown by Nielsenand Cade (1990) in Iceland, and are thus essen-tial to Gyrfalcon breeding in Greenland.
Population Changes.—Although there areinsufficient data to state accurately whether apopulation change in Gyrfalcons has occurredin Greenland, some regional decreases mayhave occurred, particularly from the Uumman-naq District south to the southwestern tip ofGreenland. For this region, comparing thenumber of Gyrfalcons reported by Bertelsen(1921) in the early 1900s with that of Burnhamet al. (2005) in 2000, almost 100 years later,fewer pairs were located during the later sur-vey. Additionally, in the Disko Bay area, nobreeding Gyrfalcons were located by Burnhamand Sherrod in 1974 (Burnham and Mattox1984), despite Salomonsen’s (1950) statementthat the Gyrfalcon breeds “rather locally inS.W. Greenland but in the Disko Bay region itincreases considerably in number,” referencingBertelsen (1932) and Fencker (1929).
Only four years of previous surveys were donein the Kangerlussuaq area specific to Gyrfal-cons, from 1990 to 1993 (M. Yates unpubl.data, W. Mattox unpubl. data). During this
period, Gyrfalcons were observed at 13, 22, 7,and 6 nest sites, respectively, and all these siteswere included in our study. However, betweenthe earlier study period and our study period,the overall size of the study area increased,with a new and very productive valley to thenorth of the original study area included in thesurveys from 1998 onward. The data from theoriginal study area in both periods are given inTable 5. Data in the “initial study area” encom-passed all nest sites known from 1990 to 1993,in addition to other nest sites found later, butwithin the same area/boundaries.
Only from 1990–1993 and from 1998–2006,when surveys were conducted specifically forGyrfalcons, were occupancy records compara-ble. Using occupied and successful nest siteslocated within the initial study area (Table 5),there appeared to be two possible Gyrfalconpopulation peaks: in 1991 and 1999–2001. Thehigh in 1991 was followed by a large and imme-diate drop, while the high from 1999 to 2001was followed by a similar rapid drop. The timeinterval between population peaks and lowsappeared to be 8 and 10 years, respectively, andfollowed a cyclical pattern similar to that asdescribed by Nielsen (1999) and Mossop andHays (1982, 1994) for Gyrfalcon populationsheavily dependent upon ptarmigan. Althoughthe initial peak and decline from 1991–1993was slightly more severe than for the period1999–2003, this may simply be a function ofpeak height and trough depth varying from onecycle to another, as has been observed in otherspecies which fluctuate cyclically.
Despite a doubling in the number of nest sitesknown and surveyed, and improved surveytechniques (foot, small planes, and some heli-copters in the early 1990s versus primarilysmall helicopter during this study), the numberof occupied nests has decreased in the Kanger-lussuaq area from the earlier to latter studyperiod (Table 5, Figure 7).
Interactions Between Gyrfalcons and Pere-grines.—The Peregrine Falcon population in

12
– BURNHAM AND BURNHAM –
Table 5. Nest site occupancy by Gyrfalcons in Kangerlussuaq, Greenland, from 1990 to 1993 and from1998 to 2006 in the “initial study area.” (M. Yates unpubl. data for 1990–1993, Mattox unpubl. data for1990–1993).
Occupied (% of Known Gyr sites Known Year checked occupied) Successful checked (%) Gyrfalcon sites
1990 13 (52%) 7 25 (100%) 25
1991 22 (69%) 12 32 (100%) 32
1992 7 (18%) 3 38 (100%) 38
1993 6 (15%) 0 40 (100%) 40
1998 8 (18%) 8 45 (78%) 58
1999 11 (20%) 10 54 (92%) 59
2000 11 (20%) 11 55 (90%) 61
2001 11 (20%) 10 55 (90%) 62
2002 8 (25%) 4 32 (52%) 62
2003 5 (12%) 2 42 (68%) 62
2004 7 (13%) 6 53 (85%) 62
2005 6 (10%) 6 60 (97%) 62
2006 8 (14%) 7 58 (94%) 62
200620042002200019981996199419921990
60
50
40
30
20
10
0
Year
Nes
t site
s
Figure 7.Changes in thenumber ofsurveyed (filledboxes), occupied(open circles), andsuccessful (opentriangles)Gyrfalcon nests inKangerlussuaqfrom 1990–1993and 1998–2006.

13
– GYRFALCON BIOLOGY IN GREENLAND –
the Kangerlussuaq area has dramaticallyincreased over time, from 34 known nest sitesfound from 1972 to 1981 (Burnham and Mat-tox 1984) to over 170 known nests sites in2006 (Burnham 2008). There is little doubtthat, at the same time the ratio of nesting Pere-grines to Gyrfalcons has also changed from areported ratio of nearly 1:1 during 1972–1981(Burnham and Mattox 1984) to between 4 and8:1 by the early 1990s (calculated using Mat-tox 1998). Using information on occupancycollected by Mattox (1998) during 1988–1997,the last 10 years of large scale surveys forPeregrines made in the Kangerlussuaq area, anaverage of 84.7% (median = 85, SD ± 6.7) ofchecked sites were occupied. If occupancyremained similar during this study, thenapproximately 144 Peregrine nests were likelyoccupied each season, making the ratio ofPeregrines to Gyrfalcons in the Kangerlussuaqarea between 10 and 29:1, depending upon thenumber of occupied Gyrfalcon nests each year.
Although perhaps ultimately driven by climatechange, might the probable Gyrfalcon popula-tion decline be associated with the increase inPeregrines? Salomonsen (1950) states, “Thenumber of Gyr-Falcons in the low-arcticregion is probably to some extent checked bythe Peregrine…” He made the statementbecause of overlap between the species in foodand nest sites. In Kangerlussuaq, territorialPeregrines and Gyrfalcons were observed nest-ing on the same cliff ≥25 times from 1983–2006 (W. Mattox unpubl. data and this study).During 1972–1982, Burnham and Mattox(1984) never observed the two species nestingon the same cliff. At a cliff where a pair ofGyrfalcons and Peregrines nested simultane-ously in 2001, the adult female Gyrfalcon wasfound below the cliff unable to fly, evidentlybecause of concussion. After being held in cap-tivity for a short time she was released at thenest site, but disappeared within days, hersmall young dying shortly after. The Pere-grines were very aggressive, and successfullyreared their own young. Aerial encountersbetween the species were seen several times,
and we were never convinced that Gyrfalconswere dominant over Peregrines. Rather, onseveral occasions, Peregrines were observeddriving adult Gyrfalcons away from nestsand/or fledged young, on occasion even strik-ing the Gyrfalcon and locking feet. Similarobservations were made by Fowles (1981)who commented that “The large falcon tendedto look clumsy and confused as it attempted toevade the agile stoops of the…Peregrines.”The Peregrine is typically very aggressivetoward other species, including other Pere-grines (Tordoff and Redig 1997).
Other than by direct aggression and displace-ment, how might Peregrines influence breed-ing Gyrfalcons? Direct displacement isprobably difficult for Peregrines because theGyrfalcons begin breeding earlier in thespring. But indirect displacement may occurthrough the Peregrine’s influence on theptarmigan population. As shown by Rosenfieldet al. (1995), fledgling Rock Ptarmigan canmake up a significant portion of the diet atPeregrine nests in Kangerlussuaq. When Gyr-falcons arrive at nest sites, and throughoutmuch of their incubation, the prey speciesavailable are primarily ptarmigan, adult hare,and Snow Buntings. Of the three, ptarmigan isprobably the most important. As Peregrinenumbers have increased in Kangerlussuaq, sohas their probable impact on ptarmigan, reduc-ing the number surviving the summer andautumn seasons, and reducing the number ofbirds available in the spring.
A similar situation with Peregrines and HenHarriers (Circus cyaneus) holding Red Grouse(Lagopus l. scoticus) populations at low levelshas been described in Scotland (Redpath andThirgood 1999). Again, both raptors were sup-ported at high density mainly by other prey,but had their major impact on the more vulner-able grouse. Neither raptor in Scotland was asheavily dependent on grouse as GreenlandGyrfalcons were on ptarmigan. Likewise, inNorway, Selås (1998) studied the effect of RedFox (Vulpes vulpes) density on Goshawks

(Accipter gentilis). Through a natural experi-ment where Goshawk, fox, and grouse popula-tions were monitored over several decades ina 650-km2 area, it was determined that RedFox influenced Goshawk breeding densitiesnegatively by limiting the number of grouse.We suggest that this type of relationship isoccurring between Peregrine, Gyrfalcon, andptarmigan populations in Kangerlussuaq.
Reproduction.—Reproduction may have variedregionally and annually, but we do not haveenough information to show this in Greenland.As mentioned in the results, in both theKangerlussuaq and Thule areas, it was fre-quently difficult to determine the total numberof young at Gyrfalcon nest sites. Therefore,most numbers provided are minima. Addition-ally, the difference in reproduction betweenoccupied and successful nests was likely under-reported, because surveys took place late in thebreeding season; at nest sites that had failed,adults may have already left the area, biasingthe results towards successful nests. Further,although Gyrfalcon reproduction has beenshown elsewhere to fluctuate between years inresponse to prey densities (White and Cade1971, Nyström et al. 2005), it was not possibleto determine if this was the case in Greenland.
Potapov and Sale (2005) reported a meanbrood size of 2.6 young (SD ± 0.91, n = 364)across the entire range of the Gyrfalcon. InKangerlussuaq, Burnham and Mattox (1984)reported an average of 2.6 young per success-ful nest (n = 20) from 1972–1981 for neststhey could observe. During this study, slightlymore young per successful nest were producedin Kangerlussuaq (2.81) compared to datafrom Burnham and Mattox (1984), while analmost identical number of young per success-ful nest were found in Thule (2.63). Chicks inThule appeared to be 2–3 weeks older on aver-age when surveys occurred, with many sites inThule having young near to fledging or alreadyfledged. Accordingly, the observed differencein nest success would likely have been moresimilar had surveys occurred when young were
approximately the same age in both areas. Onseveral occasions in Kangerlussuaq, we foundthe remains of dead young from the previousyear, in some cases fully feathered, below thenest site.
Breeding Chronology.—As expected, the morenorthern Thule population bred seasonallylater than the Kangerlussuaq population to thesouth, with similar differences observedbetween Peregrine populations in both areas(Burnham 2008, Burnham et al. in press).Greater nesting synchrony was also observedin the starting dates for Gyrfalcons breeding inthe north versus south, as described for otherraptors by Newton (1979).
For Kangerlussuaq, fewer successful nestswere found in years when breeding was initi-ated later in the season (Figure 5). Similartrends have been observed in numerous otherraptor species, including Sparrowhawks(Accipiter nisus), Peregrines, and Bald Eagles(Haliaeetus leucocephalus) (Newton and Mar-quiss 1984, Herbert and Herbert 1965, Broley1947, respectively). Why the Gyrfalcon popu-lation in Kangerlussuaq bred later in someyears is unknown. The availability of prey andinclement weather have been shown to be themost common causes of variation in nesting inmost birds (Lack 1954, 1966, Newton 1979,1998), but without more information on eitherof these variables, particularly ptarmigan den-sity, no conclusions can be drawn.
During this study, the lay-date of Gyrfalcons inThule significantly advanced (Figure 6).Advancement of lay-date has also beenobserved in numerous other species of birds(e.g., Crick et al. 1997, Crick and Sparks 1999,Dunn and Winkler 1999, Both et al. 2004) andis considered to be one of the most likelyeffects of climate change on bird species overtime (IPCC 2007). From north Greenland,including the Thule area, temperatures havebeen increasing since the late 1900s (Whitfieldet al. 2004). In particular, Mountain et al.(2001) and Serreze et al. (2000) found spring
14
– BURNHAM AND BURNHAM –

15
– GYRFALCON BIOLOGY IN GREENLAND –
temperatures to be increasing throughout thisarea, allowing for a milder and earlier spring.Already this change in temperature has beenaccompanied by an increasing Peregrine pop-ulation in the Thule area (Burnham 2008,Burnham et al. in press) and is likely alsoenabling Gyrfalcons to begin nesting earlier.
Plumage Color Variants.—The observedplumage color variation in Kangerlussuaq andThule is comparable with findings reportedelsewhere for Gyrfalcons, with a higher propor-tion of white falcons in the north. Slightly to thesouth of the Kangerlussuaq area, Salomonsen(1950) reported a 50:50 ratio of white and dark-plumage Gyrfalcons. In Kangerlussuaq during1972–1977, the plumage of 29 adult Gyrfal-cons was recorded as 16 (55%) white and 13(45%) grey (W. Burnham unpubl. data). During1998–2006, we documented plumage color of137 adults as 71 (52%) white, 16 (12%) silverand 50 (36%) grey. During the 1970s, silverwas included with grey, and if we lump silverand grey Gyrfalcons together our ratio is nearlyidentical. Based on these results it appears noshift in plumage color has occurred in the past50–60 years.
The observed difference in plumage colorbetween breeding females and males inKangerlussuaq was unexpected, with 43% offemales and 65% of males being white. To ourknowledge this is the first instance in Gyrfal-cons where such a large difference wasobserved between sexes within the same area.More research is needed to determine why thisis occurring.
CONCLUSION
These are the first long-term studies ever con-ducted on Gyrfalcon populations withinGreenland. Based on information collected byothers and data from this study it appears thatthe population in Kangerlussuaq is at best sta-ble, and may be slowly declining. Populationpeaks were observed in both 1991 and in 1999and 2000 in Kangerlussuaq, with approxi-mately one-third fewer occupied sites found
during the latter period and 69% of surveyedsites occupied vs. 20% and 16%, respectively,with similar numbers of successful sitesobserved between periods. However, morepreviously-used nest sites were known andsurveyed during the latter period, with morethorough and improved surveys not findingmore nesting falcons. The population in Thuleappears to be stable, with only minor fluctua-tions occurring during this study, but withbreeding pairs increasingly moving throughoutthe study area and using alternate nest sites.
The Gyrfalcon population in Kangerlussuaqbegan nesting approximately 18 days earlierthan the population in Thule. No large differ-ences were observed in reproduction, with theKangerlussuaq sample producing slightly moreyoung per successful nest but equal numbersof young per occupied nest. In years whenbreeding began later in Kangerlussuaq, fewersuccessful nests were found. The initiation ofbreeding advanced in Thule during our studyperiod, likely in association with earlier andwarmer springs.
It is difficult to determine with certainty whateffect climate change is having on Gyrfalcons,although it is clear that it is the driving forcebehind large-scale ecosystem change. Currentobserved climate change and future predictionssuggest that Peregrine densities in Greenlandwill continue to increase, while at the same timetheir range will continue to move north intoareas previously occupied only by Gyrfalcons.In contrast, Gyrfalcons, adapted to live in thecold Arctic conditions, may well retreat frommany areas where they now occur. Continuedresearch is needed to determine what effects, ifany, both a warming climate and an expandingPeregrine population will have on Gyrfalcons inGreenland and throughout the Arctic.
ACKNOWLEDGMENTS
We thank I. Newton and J. Burnham for com-ments and suggestions. Additional thanks tothe Greenland Home Rule Government forproviding permits for this research. Logistical

support was provided by Polar Field Services,the United States Air Force, KangerlussuaqInternational Science Support, VECO (nowCH2M HILL), 109th Air National Guard, andthe US Department of the Interior Bureau ofLand Management. For many years of criticalhelp collecting data in Greenland we give ourthanks to B. and R. Mutch, J. Stephens, J. Caf-ferty, C. Offield, J. Johnson, and B. Konkel.Thanks also go to R. Abbott, E. Vaughn, E.Stockard, S. Zager, B. Vængtoft, B. Brodersen,K. Derry, and all the residents of both ThuleAir Base and Kangerlussuaq for their friend-ship and assistance. Financial support was pro-vided by The Peregrine Fund, Calen Offieldand the Offield Family Foundation, Ruth O.Mutch, the Wolf Creek Charitable Foundation,Peter Pfendler, Comer Science and EducationFoundation, The G. Unger Vetlesen Founda-tion, and Joe and Flinda Terteling.
LITERATURE CITED
BARICHELLO, N., AND MOSSOP, D. 2011. Theoverwhelming influence of ptarmiganabundance on Gyrfalcon reproductive suc-cess in the central Yukon, Canada. In R. T.Watson, T. J. Cade, M. Fuller, G. Hunt, andE. Potapov (Eds.). Gyrfalcons and Ptarmi-gan in a Changing World. The PeregrineFund, Boise, Idaho, USA. http://dx.doi.org/10.4080/gpcw.2011.0206
BERTELSEN, A. 1921. The Birds of UmanaqDistrict. (From Danish by W. G. Mattox).Meddelelser om Grønland 62:144–214.
BERTELSEN, A. 1932. Reports about some ofthe more commonly occurring birds inWest Greenland’s Districts between 60degrees and 77 degrees N.—particularly ontheir distribution, breeding areas, andmigration. (From Danish by W. G. Mattox).Meddelelser om Grønland 91:28–31.
BLISS, L. C. 1979. Arctic heathlands. Pages415–424 in R. L. Specht (Ed.). Heathlandsand Related Shrublands of the World. ADescriptive Studies. Elsevier ScientificPublishing Company, Netherlands.
BOERTMANN, D. 2008. Grønlands Rødliste–2007. (From Danish by D. Boertmann).Danmarks Miljøundersøgelser, AarhusUniversitet, og Grønlands Hjemmestyre.
BOOMS, T. L., AND M. R. FULLER. 2003. Gyr-falcon diet in central west Greenland dur-ing the nesting period. Condor105:528–537.
BOTH, C., A. V. ARTEMYEV, B. BLAAUW, R. J.COWIE, A. J. DEKHUIJZEN, T. EEVA, A. ENE-MAR, L. GUSTAFSSON, E. V. IVANKINA, A.JÄRVINEN, N. B. METCALFE, E. I. NYHOLM,J. POTTI, P. RAVUSSIN, J. J. SANZ, B. SIL-VERIN, F. M. SLATER, L. V. SOKOLOV, J.TÖRÖK, W. WINKEL, J. WRIGHT, H. ZANG,AND M. E. VISSER. 2004. Large-scale geo-graphical variation confirms that climatechange causes birds to lay earlier. Proceed-ings of the Royal Society B: Biological Sci-ences 271:1657–1662.
BROLEY, C. L. 1947. Migration and nesting ofFlorida Bald Eagles. Wilson Bulletin 59:3–20.
BURNHAM, K. K. 2008. Inter- and intraspecificvariation of breeding biology, movements,and genotype in Peregrine Falcon Falcoperegrinus and Gyrfalcon F. rusticolus pop-ulations in Greenland. D.Phil. dissertation,University of Oxford, Oxford, UK.
BURNHAM, K. K., W. A. BURNHAM, I. NEWTON,J. A. JOHNSON, AND A. G. GOSLER. (inpress) The History and Range Expansion ofPeregrine Falcons in the Thule Area, North-west Greenland. Meddelelser om Grønland,Bioscience.
BURNHAM, W. A., K. K. BURNHAM, AND T. J.CADE. 2005. Past and present assessmentsof bird life in Uummannaq District, WestGreenland. Dansk Ornithologisk ForeningsTidsskrift 99:196–208.
BURNHAM, W. A., AND W. G. MATTOX. 1984.Biology of the Peregrine and Gyrfalcon inGreenland. Meddelelser om Grønland, Bio-science 14:1–28.
CABOT, D., R. GOODWILLIE, AND M. VINEY.1988. Irish Expedition of North-east Green-land 1987. Barnacle Books, Dublin, Ire-land.
16
– BURNHAM AND BURNHAM –

CADE, T. J. 1960. Ecology of the Peregrine andGyrfalcon populations in Alaska. Univer-sity of California Publications in Zoology63:151–290.
CADE, T. J. 1982. The Falcons of the World.Comstock/Cornell University Press, Ithaca,New York, USA.
CADE, T. J., P. KOSKIMIES, AND O. K. NIELSEN.1998. Falco rusticolus Gyrfalcon. BWPUpdate, vol. 2, no. 1. Oxford UniversityPress, Oxford, UK.
CHAPMAN, F. M. 1899. Report on birdsreceived through the Peary Expedition toGreenland. Bulletin of the AmericanMuseum of Natural History 12:219–244.
CRAMP, S., AND K. E. L. SIMMONS (EDS.). 1980.Birds of the Western Palearctic, vol. 2.Oxford University Press, Oxford, UK.
CRICK, H. Q. P., C. DUDLEY, D. E. GLUE, AND
D. L. THOMSON. 1997. UK birds are layingeggs earlier. Nature 388:526.
CRICK, H. Q. P., AND T. H. SPARKS. 1999. Cli-mate change related to egg-laying trends.Nature 399:423–424.
DUNN, P. O., AND D. W. WINKLER. 1999. Cli-mate change has affected the breeding dateof Tree Swallows throughout North Amer-ica. Proceedings of the Royal Society B:Biological Sciences 266:2,487–2,490.
EGEVANG, C., D. BOERTMANN, A. MOSBECH,AND M. P. TAMSTORF. 2003. Estimatingcolony area and population size of LittleAuks Alle alle at Northumberland Islandusing aerial images. Polar Biology 26:8–13.
FENCKER, E. C. 1929. Nogle efterladte opteg-nelser om Nordgrønlands Fugle. Ved H.Fencker and H. Scheel. Dansk Ornitholo-gisk Forenings Tidsskrift 23:33–50.
FLETCHER, D. J., AND K. WEBBY. 1977. Obser-vations on Gyrfalcons Falco rusticolus innortheast Greenland. Dansk OrnithologiskForenings Tidsskrift 71:29–35.
FOWLES, A. P. 1981. Gyrfalcon. Pages 169–170 in A. D. Fox and D. A. Stroud (Eds.).Greenland White-fronted Goose Study:Report of the 1979 Expedition to Eqalung-miut Nunât, West Greenland. Greenland
White-fronted Goose Study, Aberystwyth,Wales, UK.
GUDMUNDSSON, F. 1960. Some reflections onptarmigan cycles in Iceland. Proceedings ofthe International Ornithological Congress12:259–265.
HAGERUP, A. T. 1891. The Birds of Greenland.Little, Brown, and Company, Boston, Mas-sachusetts, USA.
HALL, C. F. 1876. Narrative of the North PolarExpedition. US ship Polaris, CaptainCharles Francis Hall commanding. Gov-ernment Printing Office, Washington, DC,USA.
HERBERT, R. A., AND K. G. S. HERBERT. 1965.Behavior of Peregrine Falcons in the NewYork City Region. Auk 82:62–94.
IPCC. 2007. Climate Change 2007: The Phys-ical Science Basis. Contribution of Work-ing Group I to the Fourth AssessmentReport of the Intergovernmental Panel onClimate Change. S. Solomon, D. Qin, M.Manning, Z. Chen, M. Marquis, K. B.Averyt, M. Tignor, and H. L. Miller (Eds.).Cambridge University Press, Cambridge,UK and New York, New York, USA.
JENKINS, M. A. 1978. Gyrfalcon nesting behav-ior from hatching to fledging. Auk 95:122–127.
JOHNSEN, P. 1953. Birds and mammals ofPeary Land in north Greenland. Med-delelser om Grønland 128:1–135.
KOCH, L. 1925. Nord om Grønland. Køben-havn, Denmark.
LACK, D. 1954. The Natural Regulation ofAnimal Numbers. University Press,Oxford, UK.
LACK, D. 1966. Population Studies of Birds.University Press, Oxford, UK.
MANNICHE, A. L. 1910. The terrestrial mam-mals and birds of north-east Greenland.(From Danish by W. G. Mattox). Medde-leser om Grønland 45:183–187.
MATTOX, W. G. 1970. Banding Gyrfalcons(Falco rusticolus) in Greenland, 1967. BirdBanding 41:31–37.
MATTOX, W. G. 1998. High Arctic Institute andGreenland Peregrine Falcon Survey
17
– GYRFALCON BIOLOGY IN GREENLAND –

Progress Report 1998. The Peregrine Fund,Boise, Idaho, USA.
MINDELL, D. P., J. L. B. ALBUQUERQUE, AND C.M. WHITE. 1987. Breeding population fluc-tuations in some raptors. Oecologia72:328–388.
MINDELL, D. P., AND C. M. WHITE. 1988. Fluc-tuations of observed breeding Rough-legged Hawks and Gyrfalcons: Regularityreconsidered. Oecologia 77:14–18.
MOSS, R., AND A. WATSON. 2001. Populationcycles in birds of the grouse family(Tetraonidae). In Advances in EcologicalResearch 32:53–111. Academic Press, Lon-don, UK.
MOSSOP, D. H. 2011. Long-term studies ofWillow Ptarmigan and Gyrfalcon in theYukon Territory: A collapsing 10-year cycleand its apparent effect on the top predator.In R. T. Watson, T. J. Cade, M. Fuller, G.Hunt, and E. Potapov (Eds.). Gyrfalconsand Ptarmigan in a Changing World. ThePeregrine Fund, Boise, Idaho, USA.http://dx.doi.org/10.4080/gpcw.2011.0206
MOSSOP, D. H., AND R. HAYES. 1982. TheYukon Territory Gyrfalcon harvest experi-ment (1974–80). Pages 263–280 in W. N.Ladd and P. F. Schempf (Eds.). Proceed-ings, Symposium and Workshop: RaptorManagement and Biology in Alaska andWestern Canada. US Fish and WildlifeService, Anchorage, Alaska, USA.
MOSSOP, D. H., AND R. HAYES. 1994. Longterm trends in the breeding density and pro-ductivity of Gyrfalcon Falco rusticolus inthe Yukon Territory, Canada. Pages 403–413 in B. -U. Meyburg and R. D. Chancel-lor (Eds.). Raptor Conservation Today.Helm Information Inc, Mountfield, EastSussex, UK.
MOUNTAIN, K. R., D. A. HOWARTH, AND S. R.JOULE. 2001. Spatial and temporal temper-ature change in the arctic basin since 1994:Assessment of localized short-term datasets. Polar Geography 25:1–21.
NEWTON, I. 1979. Population Ecology of Rap-tors. T. & A. D. Poyser, London, UK.
NEWTON, I. 1998. Population Limitations inBirds. Academic Press, London, UK.
NEWTON, I., AND M. MARQUISS. 1984. Seasonaltrend in the breeding Performance of Spar-rowhawks. Journal of Animal Ecology53:809–829.
NIELSEN, Ó. K. 1999. Gyrfalcon predation onptarmigan: Numerical and functionalresponses. Journal of Animal Ecology68:1034–1050.
NIELSEN, Ó. K., AND T. J. CADE. 1990. Sea-sonal changes in food habits of Gyrfalconsin NE-Iceland. Ornis Scandinavica 21:202–211.
NYSTRÖM, J., J. EKENSTEDT, J. ENGSTRÖM, AND
A. ANGERBJÖRN. 2005. Gyr Falcons,ptarmigan and microtine rodents in north-ern Sweden. Ibis 147:587–597.
PLATT, J. B. 1977. The breeding behavior ofwild and captive Gyrfalcons in relation totheir environment and human disturbance.Ph.D. dissertation, Cornell University,Ithaca, New York, USA.
POOLE, K. G., AND R. G. BROMLEY. 1988. Nat-ural history of the Gyrfalcon in the centralCanadian Arctic. Arctic 41:31–38.
POTAPOV, E., AND R. SALE. 2005. The Gyrfal-con. Yale University Press, New Haven,Connecticut, USA.
RAFN, A. 1933. Lidt om fuglene I og omkringJulianehaab. Dansk Ornithologisk Foren-ings Tidsskrift 27:19–25.
RATCLIFFE, D. A. 1969. Population trends ofthe Peregrine Falcon in Great Britain.Pages 239–269 in J. J. Hickey (Ed.). Pere-grine Falcon Populations: Their Biologyand Decline. University Press, Madison,Wisconsin, USA.
REDPATH, S. M., AND S. J. THIRGOOD. 1999.Numerical and functional responses of gen-eralist predators: Harriers and Peregrineson grouse moors. Journal of Animal Ecol-ogy 68:879–892.
ROSENFIELD, R. N., J. W. SCHNEIDER, J. M.PAPP, AND W. S. SEEGAR. 1995. Prey ofPeregrine Falcons breeding in West Green-land. Condor 97:763–770.
18
– BURNHAM AND BURNHAM –

SABINE, E. 1819. A memoir on the birds ofGreenland; with descriptions and notes onthe species observed in the late voyage ofdiscovery in Davis’s Straits and Baffin’sBay. Transactions of the Linnean Society12:527–559.
SALOMONSEN, F. 1950. Grønlands Fugle/TheBirds of Greenland. Ejnar Munksgaard,Copenhagen, Denmark.
SELÅS, V. 1998. Does food competition fromRed Fox (Vulpes vulpes) influence thebreeding density of Goshawk (Accipitergentilis)? Evidence from a natural experi-ment. Journal of Zoology 246:325–335.
SERREZE, M. C., J. E. WALSH, F. S. CHAPIN III,T. OSTERKAMP, M. DYURGEROV, V.ROMANOVSKY, W. C. OECHEL, J. MORISON,T. ZHANG, AND G. BARRY. 2000. Observa-tional evidence of recent change in thenorthern high-latitude environment. Cli-matic Change 46:59–207.
SHANK, C. C., AND K. G. POOLE. 1994. Statusof Gyrfalcon Falco rusticolus populationsin the Northwest Territories, Canada. Pages421–436 in B.-U. Meyburg and R. D.Chancellor (Eds.). Raptor ConservationToday. Helm Information Inc, Mountfield,East Sussex, UK.
STONE, W. 1895. List of birds collected innorth Greenland by the Peary expedition of1891–92 and the relief expedition of 1892.
Proceedings of the Academy of NaturalSciences of Philadelphia 47:502–505.
TORDOFF, H. B., AND P. T. REDIG. 1997. Mid-west Peregrine Falcon demography, 1982–1995. Journal of Raptor Research31:339–346.
VIBE, C. 1967. Arctic animals in relation to cli-matic fluctuations. The Danish Zoogeo-graphical Investigations in Greenland.Meddelelser om Grønland 170:1–227.
WALKER, D. 1860. The last arctic expeditionunder Captain Sir F. L. M’Clintock, R. N.Journal of the Royal Dublin Society 3:61–77.
WATSON, A., R. MOSS, AND S. RAE. 1998. Pop-ulation dynamics of Scottish Rock Ptarmi-gan cycles. Ecology 79:1174–1192.
WEEDEN, R. B., AND J. B. THEBERGE. 1972.The dynamics of a fluctuating populationof ptarmigan in Alaska. Proceedings of theInternational Ornithological Congress15:90–106.
WHITE, C. M., AND T. J. CADE. 1971. Cliff-nesting raptors and ravens along theColville river in arctic Alaska. Living Bird10:107–150.
WHITFIELD, P. H., A. W. HALL, AND A. J. CAN-NON. 2004. Changes in the seasonal cyclein the circumpolar Arctic, 1976–95: Tem-perature and precipitation. Arctic 57:80–93.
19
– GYRFALCON BIOLOGY IN GREENLAND –

20
– BURNHAM AND BURNHAM –





























