Embed Size (px)
Citation preview

Dopamine inhibits pulmonary edema through the VEGF-VEGFR2 axisin a murine model of acute lung injury
Pawan K. Vohra,1 Luke H. Hoeppner,1 Gunisha Sagar,1 Shamit K. Dutta,1 Sanjay Misra,3
Rolf D. Hubmayr,2 and Debabrata Mukhopadhyay1
Departments of 1Biochemistry and Molecular Biology, and 3Radiology, 2Division of Pulmonary and Critical Care Medicine,Mayo Clinic College of Medicine, Rochester, Minnesota
Submitted 16 August 2010; accepted in final form 13 October 2011
Vohra PK, Hoeppner LH, Sagar G, Dutta SK, Misra S, Hub-mayr RD, Mukhopadhyay D. Dopamine inhibits pulmonary edemathrough the VEGF-VEGFR2 axis in a murine model of acute lunginjury. Am J Physiol Lung Cell Mol Physiol 302: L185–L192, 2012.First published October 14, 2011; doi:10.1152/ajplung.00274.2010.—The neurotransmitter dopamine and its dopamine receptor D2 (D2DR)agonists are known to inhibit vascular permeability factor/vascularendothelial growth factor (VEGF)-mediated angiogenesis and vascu-lar permeability. Lung injury is a clinical syndrome associated withincreased microvascular permeability. However, the effects of dopa-mine on pulmonary edema, a phenomenon critical to the pathophys-iology of both acute and chronic lung injuries, have yet to beestablished. Therefore, we sought to determine the potential therapeu-tic effects of dopamine in a murine model of lipopolysaccharide(LPS)-induced acute lung injury (ALI). Compared with sham-treatedcontrols, pretreatment with dopamine (50 mg/kg body wt) amelioratedLPS-mediated edema formation and lowered myeloperoxidase activ-ity, a measure of neutrophil infiltration. Moreover, dopamine signif-icantly increased survival rates of LPS-treated mice, from 0–75%.Mechanistically, we found that dopamine acts through the VEGF-VEGFR2 axis to reduce pulmonary edema, as dopamine pretreatmentin LPS-treated mice resulted in decreased serum VEGF, VEGFR2phosphorylation, and endothelial nitric oxide synthase phosphoryla-tion. We used D2DR knockout mice to confirm that dopamine actsthrough D2DR to block vascular permeability in our lung injurymodel. As expected, a D2DR agonist failed to reduce pulmonaryedema in D2DR�/� mice. Taken together, our results suggest thatdopamine acts through D2DR to inhibit pulmonary edema-associatedvascular permeability, which is mediated through VEGF-VEGFR2signaling and conveys protective effects in an ALI model.
dopamine receptor D2; vascular endothelial growth factor
ACUTE LUNG INJURY (ALI) and its severe form, the acuterespiratory distress syndrome (ARDS), are prevalent causesof morbidity and mortality (51). One of the hallmarks ofALI is the accumulation of protein-rich alveolar edema fluidresulting from impaired vascular barrier properties (29, 33,50, 53). Not only are vascular filtration coefficient andprotein permeability increased, injured lungs are also defec-tive in alveolar fluid clearance (35, 52). Although �-receptoragonists such as albuterol and isoproterenol reliably in-crease alveolar fluid clearance in experimental animals withinjured lungs, their use in patients (42) has met limitedsuccess (32). Dopamine, a neurotransmitter, acts via tworeceptor isoforms (dopamine receptor D1 and D2, D1DR
and D2DR). D1DR has been shown to increase transepithe-lial fluid flux through the trafficking of NaKATPase to thebasolateral membrane of type II alveolar epithelial cells (5).Moreover, activation of D2DR induces NaKATPase geneexpression (17). However, the actions of dopamine onpulmonary vascular barrier properties may not be limited toeffects on transepithelial sodium transport. Activation ofD2DR has also been implicated in the regulation of vascularendothelial growth factor (VEGF)-induced vascular perme-ability as well as tumor angiogenesis (2, 9, 46). Manyexperimental and human studies support the hypothesis thatVEGF plays a critical role in shaping the vascular barrierfunction in ALI (3, 6, 15, 18, 25). In normal human lungs,the VEGF concentration in alveolar lining fluid is greaterthan that in plasma (43). Epithelial injury leads to decom-partmentalization of VEGF, permitting its migration acrosstight junctions from the alveolar space to the vascularcompartment (16, 25, 27, 49), and is therefore thought toplay a pathogenic role in noncardiogenic pulmonary edema(26, 38).
Using a mouse ovarian tumor model, our group previ-ously reported that the endothelium expresses the liganddopamine along with its cognate D2DR, thereby regulatingvascular permeability and endothelial cell (EC) barrier in-tegrity (2, 7). Furthermore, we demonstrated that dopamine,through a D2DR-dependent mechanism, causes the down-regulation of VEGFR2 phosphorylation (47). We now pro-vide evidence that the same mechanisms operate in murineendotoxin-induced ALI and can be targeted to improvemorbidity and mortality in this preclinical model. Sepsis isthe most common cause of ALI in humans (50, 51). Admin-istration of the gram-negative bacterial endotoxin lipopoly-saccharide (LPS) has been widely used as an animal modelof sepsis-related lung injury. LPS is known to cause over-expression of VEGF (11, 12, 22, 54), which in turn upregu-lates nitric oxide (NO) production through phosphorylationof VEGFR2 and endothelial NO synthase (eNOS) (13, 14,40). As postulated, our results suggest that dopamine actsthrough D2DR to inhibit pulmonary edema-associated vas-cular permeability, which is mediated through VEGF-VEGFR2 signaling, and conveys protective effects in anALI model.
MATERIALS AND METHODS
Animals
Pathogen-free male Balb/C mice purchased from the NationalCancer Institute (Frederick, MD) and D2DR knockout C57Bl/6 micefrom Jackson Laboratory (Bar Harbor, ME) were used in LPS-induced
Address for reprint requests and other correspondence: D. Mukhopadhyay,Dept. of Biochemistry and Molecular Biology, Mayo Clinic College ofMedicine, Guggenheim 1334, 200 First St. S.W., Rochester, MN 55905(e-mail: [email protected]).
Am J Physiol Lung Cell Mol Physiol 302: L185–L192, 2012.First published October 14, 2011; doi:10.1152/ajplung.00274.2010.
1040-0605/12 Copyright © 2012 the American Physiological Societyhttp://www.ajplung.org L185
by 10.220.33.4 on April 19, 2017
http://ajplung.physiology.org/D
ownloaded from

ALI. The animals were housed in separate cages in a temperature-controlled room with alternating 12-h:12-h light/dark cycles and wereallowed 1 wk to acclimate to their surroundings. The animals werealso fed a standard diet.
Reagents
LPS (Salmonella enterica), FITC-albumin, hexadecyl-trimethyl-ammonium bromide, hydrogen peroxide, quinpirole, ectilopride, andO-dianisidine hydrochloride were purchased from Sigma-Aldrich (St.Louis, MO). The antibodies used were phospho-eNOS (S1177) andtotal eNOS (BD Biosciences, Franklin Lakes, NJ), VEGFR-2 (SC-504, Santa Cruz Biotechnology, Santa Cruz, CA), and phospho-tyrosine 4G10 clone (Millipore, Billerica, MA).
Dopamine Preparation
A pharmaceutical-grade dopamine solution (40 mg/ml) was pur-chased from the Mayo Clinic Pharmacy. Dopamine (50 mg/kg bodywt), quinpirole (10 mg/kg body wt), and eticlopride (10 mg/kg bodywt) doses were selected on the basis of prior studies demonstratingtheir regulatory effects on vascular permeability (2).
Experimental Procedure
The wild-type animals were divided into six treatment groups ascontrol (sham treated, n � 20), dopamine alone (n � 20), LPSalone (n � 20), dopamine � LPS (n � 20), quinpirole (D2DRspecific agonist) � LPS (n � 20), and dopamine � LPS �eticlopride (D2DR antagonist, n � 20). In D2DR knockout study,animals were divided into six treatment groups as control (sham-treated D2�/�, n � 14), LPS alone (D2�/�, n � 14), quinpirole �LPS (�/�, n � 14), quinpirole � LPS (�/�, n � 14), quinpirole � LPS(D2�/�, n � 14), and dopamine � LPS (D2�/�, n � 14). The MayoClinic Institutional Animal Care and Use Committee approved allexperimental procedures. LPS (400 �g/mouse) was injected intraperi-toneally (IP) to induce lung edema (54). The control and treatedanimals were euthanized by spinal break. One group of animals wastreated 30 min before the onset of LPS administration with an IPinjection of dopamine (50 mg/kg body wt). One dose of dopamine wasgiven the day of LPS treatment, and an additional dose was given thefollowing day. The same procedure was repeated with quinpirole (10mg/kg body wt).
Lung Injury Analysis
Lung injury is a broad term that can be applied to conditionsranging from mild interstitial edema without cellularity to massiveand fatal destruction of the lung. Here, we used physical and bio-chemical methods to investigate the effect of dopamine on LPS-induced lung injury. The LPS-challenged animals become very sickand started dying within 2–3 h at this stage. We selected 24 h as thetime point in all groups for histological comparison except for bron-choalveolar lavage (BAL) analysis, which was done at 6 h as well as24 h after LPS challenge.
Lung Water Content
After completion of the animal experiment, mice were eutha-nized by spinal break. The left lung was harvested for the wet-to-dry weight ratio. The dry weight was determined after incubatingthe lungs at 80°C for 72 h, and the wet-to-dry weight ratio wascalculated (56).
Histology
The right lung was inflated with 10% (vol/vol) formaldehydeembedded in paraffin, and cut into 5-�m-thick sections. Sections werestained with hematoxylin and eosin, and images were taken with aNikon Eclipse E800 microscope with a �20 objective. The extent of
lung injury was determined by a blinded observer using a semiquan-titative score based on congestion, interstitial edema, neutrophil infil-tration, and air space hemorrhage, as follows: 0 � no change; 1� �focal, mild, subtle change, 2� � multifocal mild changes; 3� �multifocal prominent changes; and 4� � extensive prominentchanges (1, 48). Histological comparison was done at 24 h in allgroups.
Survival
A survival study was carried out separately with initial interven-tion. Animals (n � 8, each group) were observed daily and survivalcalculated 158 h after the initial intervention. D2DR knockout animals(n � 8, each group) were monitored up to 32 h.
Biochemical Assays
Pulmonary microvascular permeability. FITC-labeled albumin, amacromolecular marker, is widely used to evaluate pulmonarymicrovascular permeability (56). Two hours before euthanasia,FITC-labeled albumin (5 mg/kg body wt) was administered via atail-vein injection at 6 h (n � 3) and at 24 h (n � 3). Immediatelyafter euthanasia, the lungs were lavaged three times with phos-phate-buffered saline (0.5 ml per lavage) and the samples com-bined. Fluid recovery was roughly 95%. The BAL samples werecentrifuged at 3,000 revolution/min for 10 min. FITC fluorescencein the BAL fluid was measured using a fluorescence spectropho-tometer with excitation at 484 nm and emission at 510 nm (56).
Myeloperoxidase assay. Tissue myeloperoxidase (MPO) activityis a quantitative marker for the presence of neutrophils in the lungs(55). MPO activity was measured in tissue homogenates at 24 hafter LPS injection in placebo and dopamine treated animals (n �6). The MPO activity was expressed as absorbance per gram oftissue (39).
VEGF-A Levels and VEGFR-2 Phosphorylation
Serum was collected from the different experimental groups at 24h after the LPS challenge. Mouse VEGF-164 levels in serum weremeasured by an ELISA assay (R&D Systems, Minneapolis, MN) witha specific mouse VEGF primary antibody that has no cross reactivitywith human VEGF. Phosphorylation of VEGFR2 was assessed byWestern blot analysis. Frozen whole-lung tissues were homogenizedin 400 �l of RIPA lysis buffer supplemented with protease andphosphatase inhibitors. The homogenates were centrifuged at 14,000revolution/min for 15 min at 4°C, and the supernatants were collected.VEGFR2 was immunoprecipitated with a VEGFR2 antibody andresolved on a 10% SDS-PAGE. The samples were analyzed forVEGFR2 phosphorylation using the antiphosphotyrosine clone 4G10.A mouse anti-IgG-horseradish peroxidase conjugate was used todetect the band with the enhanced chemiluminescence reagent (GEHealthcare Life Sciences, Piscataway, NJ). The intensities of thebands were analyzed using Image J from the National Institutes ofHealth.
Measurement of eNOS
Histology slides were stained for eNOS using a blood vesselstaining kit (ECM590, Millipore) per the manufacturer’s instructions.Immunohistological staining of slides from all treatment groups wasperformed with phospho-eNOS (Serine 1177) and total eNOS anti-bodies (BD Biosciences). A quantitative analysis of immunostainedslides was done with Metamorph software (greater than three slidesper group).
Statistics
The data in the figures are represented as means and SD. The errorbar was based on SD values. Differences between the treated groupsvs. the injured group (LPS/saline) were assessed using the unpaired
L186 DOPAMINE INHIBITS PULMONARY EDEMA IN LUNG INJURY
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00274.2010 • www.ajplung.org
by 10.220.33.4 on April 19, 2017
http://ajplung.physiology.org/D
ownloaded from
Student’s t-tests for unpaired observation, and ANOVA and/orDunn’s test. Statistical significance was assigned when the P valuewas �0.05.
RESULTS
Given the well-established role of dopamine in preventingvascular permeability, we sought to determine the potentialtherapeutic effects of dopamine in a murine model of ALI. Tothis end, we challenged mice with LPS to induce pathophysi-ological features of pulmonary edema such as alveolar floodingand collapse, tissue infiltration with inflammatory cells, andpulmonary hemorrhage.
Dopamine Modulates the Pathophysiological Features ofLPS-Induced ALI
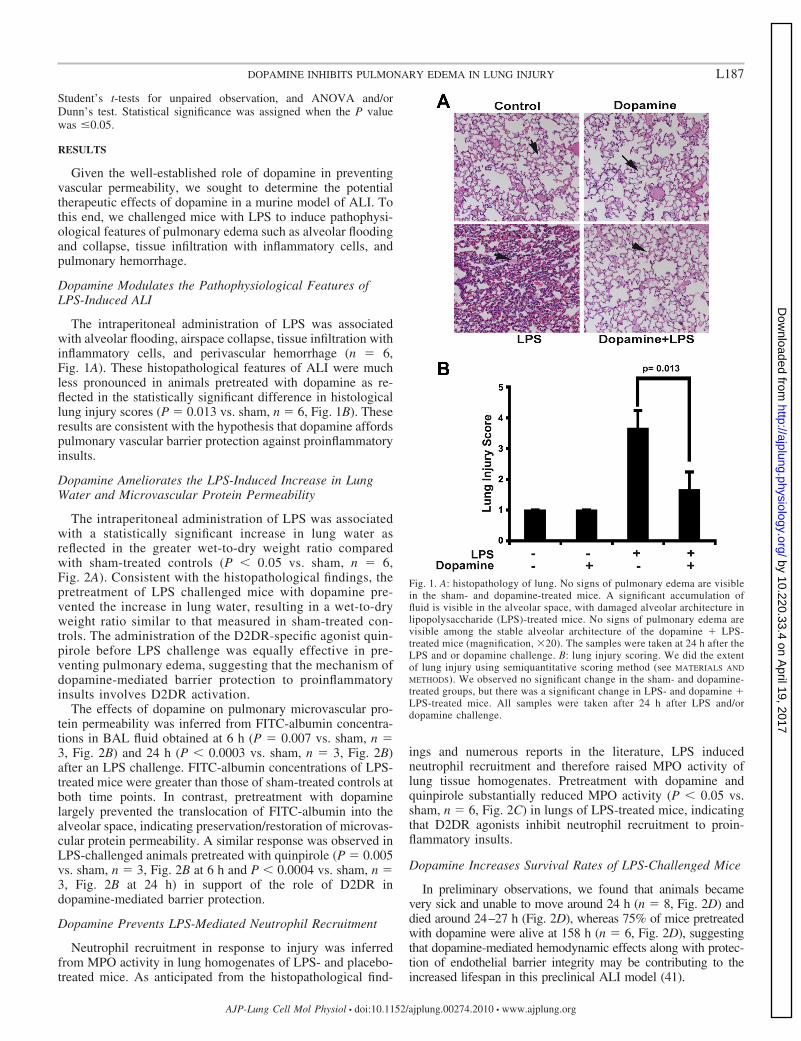
The intraperitoneal administration of LPS was associatedwith alveolar flooding, airspace collapse, tissue infiltration withinflammatory cells, and perivascular hemorrhage (n � 6,Fig. 1A). These histopathological features of ALI were muchless pronounced in animals pretreated with dopamine as re-flected in the statistically significant difference in histologicallung injury scores (P � 0.013 vs. sham, n � 6, Fig. 1B). Theseresults are consistent with the hypothesis that dopamine affordspulmonary vascular barrier protection against proinflammatoryinsults.
Dopamine Ameliorates the LPS-Induced Increase in LungWater and Microvascular Protein Permeability
The intraperitoneal administration of LPS was associatedwith a statistically significant increase in lung water asreflected in the greater wet-to-dry weight ratio comparedwith sham-treated controls (P � 0.05 vs. sham, n � 6,Fig. 2A). Consistent with the histopathological findings, thepretreatment of LPS challenged mice with dopamine pre-vented the increase in lung water, resulting in a wet-to-dryweight ratio similar to that measured in sham-treated con-trols. The administration of the D2DR-specific agonist quin-pirole before LPS challenge was equally effective in pre-venting pulmonary edema, suggesting that the mechanism ofdopamine-mediated barrier protection to proinflammatoryinsults involves D2DR activation.
The effects of dopamine on pulmonary microvascular pro-tein permeability was inferred from FITC-albumin concentra-tions in BAL fluid obtained at 6 h (P � 0.007 vs. sham, n �3, Fig. 2B) and 24 h (P � 0.0003 vs. sham, n � 3, Fig. 2B)after an LPS challenge. FITC-albumin concentrations of LPS-treated mice were greater than those of sham-treated controls atboth time points. In contrast, pretreatment with dopaminelargely prevented the translocation of FITC-albumin into thealveolar space, indicating preservation/restoration of microvas-cular protein permeability. A similar response was observed inLPS-challenged animals pretreated with quinpirole (P � 0.005vs. sham, n � 3, Fig. 2B at 6 h and P � 0.0004 vs. sham, n �3, Fig. 2B at 24 h) in support of the role of D2DR indopamine-mediated barrier protection.
Dopamine Prevents LPS-Mediated Neutrophil Recruitment
Neutrophil recruitment in response to injury was inferredfrom MPO activity in lung homogenates of LPS- and placebo-treated mice. As anticipated from the histopathological find-
ings and numerous reports in the literature, LPS inducedneutrophil recruitment and therefore raised MPO activity oflung tissue homogenates. Pretreatment with dopamine andquinpirole substantially reduced MPO activity (P � 0.05 vs.sham, n � 6, Fig. 2C) in lungs of LPS-treated mice, indicatingthat D2DR agonists inhibit neutrophil recruitment to proin-flammatory insults.
Dopamine Increases Survival Rates of LPS-Challenged Mice
In preliminary observations, we found that animals becamevery sick and unable to move around 24 h (n � 8, Fig. 2D) anddied around 24–27 h (Fig. 2D), whereas 75% of mice pretreatedwith dopamine were alive at 158 h (n � 6, Fig. 2D), suggestingthat dopamine-mediated hemodynamic effects along with protec-tion of endothelial barrier integrity may be contributing to theincreased lifespan in this preclinical ALI model (41).
Fig. 1. A: histopathology of lung. No signs of pulmonary edema are visiblein the sham- and dopamine-treated mice. A significant accumulation offluid is visible in the alveolar space, with damaged alveolar architecture inlipopolysaccharide (LPS)-treated mice. No signs of pulmonary edema arevisible among the stable alveolar architecture of the dopamine � LPS-treated mice (magnification, �20). The samples were taken at 24 h after theLPS and or dopamine challenge. B: lung injury scoring. We did the extentof lung injury using semiquantitative scoring method (see MATERIALS AND
METHODS). We observed no significant change in the sham- and dopamine-treated groups, but there was a significant change in LPS- and dopamine �LPS-treated mice. All samples were taken after 24 h after LPS and/ordopamine challenge.
L187DOPAMINE INHIBITS PULMONARY EDEMA IN LUNG INJURY
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00274.2010 • www.ajplung.org
by 10.220.33.4 on April 19, 2017
http://ajplung.physiology.org/D
ownloaded from

Dopamine Inhibits Pulmonary Edema by Decreasing SerumVEGF Levels and Phosphorylation of VEGFR2
We previously demonstrated in an ovarian tumor model thatdopamine acts through D2DRs to induce endocytosis ofVEGFR2, preventing VEGF binding and receptor activation,thereby preserving vascular barrier properties. Because serumVEGF levels are increased in noncardiogenic pulmonary
edema, we tested whether dopamine altered the lung responseto an LPS challenge by a similar mechanism. As postulated, weobserved a significant reduction in serum VEGF levels (P �0.0001 vs. sham, n � 6, Fig. 3A) and VEGFR2 (P � 0.005 vs.sham, n � 6, Fig. 3, B and C) phosphorylation in dopamine-pretreated animals challenged with an LPS-induced lung injurycompared with sham-treated groups. Our findings are consis-
Fig. 2. Effects of LPS and dopamine on lungs.The water content of lungs was determined bycalculating the wet-to-dry weight (see MATERI-ALS AND METHODS) at 24 h in sham (n � 6)-,dopamine- (n � 6), LPS- (n � 6), dopamine �LPS- (n � 6), and quinpirole � LPS-treated(n � 6) mice (A). The measurement of FITC-albumin release is indicative of vascular leak-iness in the bronchioalveolar lavage (BAL) ofsham- (n � 3), dopamine- (n � 3), LPS- (n �3), dopamine � LPS-, and quinpirole � LPS-treated mice at 6 h (gray bar) and 24 h (solidblack bar) (B). Pulmonary sequestration ofneutrophils was assessed by lung myeloperox-idase (MPO) activity in sham- (n � 6), LPS-(n � 6), dopamine- (n � 6), dopamine � LPS-(n � 6), and quinpirole � LPS- (n � 6) treatedmice at 24 h (C). Animal survival was moni-tored up to 158 h in sham- (n � 8), dopamine-(n � 8), LPS-(n � 8), and LPS � dopamine-(n � 8) treated groups based on animal activ-ity, movement, and physical examination (D).
Fig. 3. Effect of dopamine on serum vascularendothelial growth factor (VEGF) levels andVEGFR2 receptor phosphorylation. SerumVEGF levels were assessed with ELISA aswell as the phosphorylation of VEGFR2 byWestern blotting at 10% SDS-PAGE on lunghomogenate in sham- (n � 6), LPS- (n � 6),dopamine- (n � 6), dopamine � LPS- (n � 6),quinpirole � LPS- (n � 6), dopamine � ec-tilopride (dopamine receptor D2, D2DR, an-tagonist)� LPS-treated (n � 6) mice. We ob-served a significant reduction in serum VEGFlevel in dopamine- and quinpirole-pretreatedmice (n � 6; A). We also assessed VEGFR2phosphorylation in sham-, LPS-, dopamine-,and dopamine � LPS-pretreated animals (B).Quantitative analysis of VEGFR2 phosphory-lation (n � 6) was done with National Insti-tutes of Health imaging software (C). All sam-ples were taken at 24 h after LPS challenge.
L188 DOPAMINE INHIBITS PULMONARY EDEMA IN LUNG INJURY
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00274.2010 • www.ajplung.org
by 10.220.33.4 on April 19, 2017
http://ajplung.physiology.org/D
ownloaded from
tent with, but not definitive for, the effect of dopamine onsystemic VEGF levels. Similar results were obtained using theD2DR-specific agonist quinpirole (P � 0.0001 vs. sham, n �6, Fig. 3A). Eticlopride, (an antagonist of D2DR) abrogateddopamine-mediated decrease in VEGF (P � 0.1 vs. sham, n �6, Fig. 3A) levels. These results provided a further support thatD2DR, but not D1DR, plays a role in preventing pulmonaryedema by inhibiting VEGF-VEGFR2 signaling.
Dopamine Inhibits VEGFR2 Signaling
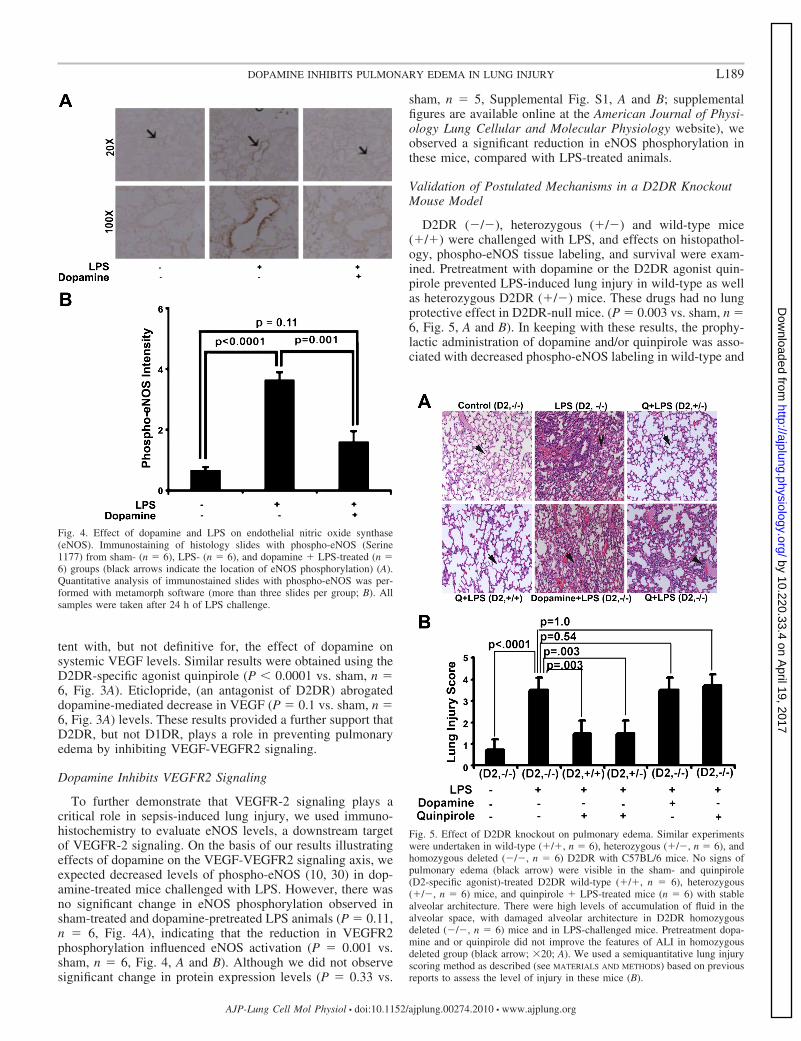
To further demonstrate that VEGFR-2 signaling plays acritical role in sepsis-induced lung injury, we used immuno-histochemistry to evaluate eNOS levels, a downstream targetof VEGFR-2 signaling. On the basis of our results illustratingeffects of dopamine on the VEGF-VEGFR2 signaling axis, weexpected decreased levels of phospho-eNOS (10, 30) in dop-amine-treated mice challenged with LPS. However, there wasno significant change in eNOS phosphorylation observed insham-treated and dopamine-pretreated LPS animals (P � 0.11,n � 6, Fig. 4A), indicating that the reduction in VEGFR2phosphorylation influenced eNOS activation (P � 0.001 vs.sham, n � 6, Fig. 4, A and B). Although we did not observesignificant change in protein expression levels (P � 0.33 vs.
sham, n � 5, Supplemental Fig. S1, A and B; supplementalfigures are available online at the American Journal of Physi-ology Lung Cellular and Molecular Physiology website), weobserved a significant reduction in eNOS phosphorylation inthese mice, compared with LPS-treated animals.
Validation of Postulated Mechanisms in a D2DR KnockoutMouse Model
D2DR (�/�), heterozygous (�/�) and wild-type mice(�/�) were challenged with LPS, and effects on histopathol-ogy, phospho-eNOS tissue labeling, and survival were exam-ined. Pretreatment with dopamine or the D2DR agonist quin-pirole prevented LPS-induced lung injury in wild-type as wellas heterozygous D2DR (�/�) mice. These drugs had no lungprotective effect in D2DR-null mice. (P � 0.003 vs. sham, n �6, Fig. 5, A and B). In keeping with these results, the prophy-lactic administration of dopamine and/or quinpirole was asso-ciated with decreased phospho-eNOS labeling in wild-type and
Fig. 5. Effect of D2DR knockout on pulmonary edema. Similar experimentswere undertaken in wild-type (�/�, n � 6), heterozygous (�/�, n � 6), andhomozygous deleted (�/�, n � 6) D2DR with C57BL/6 mice. No signs ofpulmonary edema (black arrow) were visible in the sham- and quinpirole(D2-specific agonist)-treated D2DR wild-type (�/�, n � 6), heterozygous(�/�, n � 6) mice, and quinpirole � LPS-treated mice (n � 6) with stablealveolar architecture. There were high levels of accumulation of fluid in thealveolar space, with damaged alveolar architecture in D2DR homozygousdeleted (�/�, n � 6) mice and in LPS-challenged mice. Pretreatment dopa-mine and or quinpirole did not improve the features of ALI in homozygousdeleted group (black arrow; �20; A). We used a semiquantitative lung injuryscoring method as described (see MATERIALS AND METHODS) based on previousreports to assess the level of injury in these mice (B).
Fig. 4. Effect of dopamine and LPS on endothelial nitric oxide synthase(eNOS). Immunostaining of histology slides with phospho-eNOS (Serine1177) from sham- (n � 6), LPS- (n � 6), and dopamine � LPS-treated (n �6) groups (black arrows indicate the location of eNOS phosphorylation) (A).Quantitative analysis of immunostained slides with phospho-eNOS was per-formed with metamorph software (more than three slides per group; B). Allsamples were taken after 24 h of LPS challenge.
L189DOPAMINE INHIBITS PULMONARY EDEMA IN LUNG INJURY
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00274.2010 • www.ajplung.org
by 10.220.33.4 on April 19, 2017
http://ajplung.physiology.org/D
ownloaded from
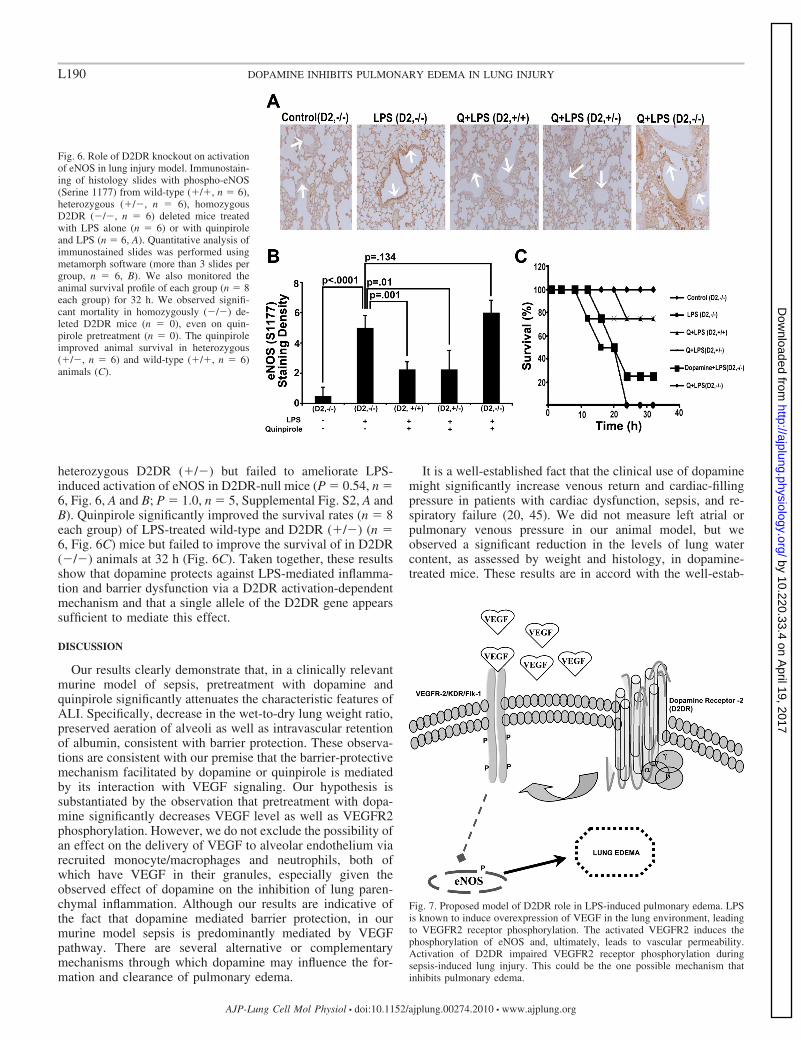
heterozygous D2DR (�/�) but failed to ameliorate LPS-induced activation of eNOS in D2DR-null mice (P � 0.54, n �6, Fig. 6, A and B; P � 1.0, n � 5, Supplemental Fig. S2, A andB). Quinpirole significantly improved the survival rates (n � 8each group) of LPS-treated wild-type and D2DR (�/�) (n �6, Fig. 6C) mice but failed to improve the survival of in D2DR(�/�) animals at 32 h (Fig. 6C). Taken together, these resultsshow that dopamine protects against LPS-mediated inflamma-tion and barrier dysfunction via a D2DR activation-dependentmechanism and that a single allele of the D2DR gene appearssufficient to mediate this effect.
DISCUSSION
Our results clearly demonstrate that, in a clinically relevantmurine model of sepsis, pretreatment with dopamine andquinpirole significantly attenuates the characteristic features ofALI. Specifically, decrease in the wet-to-dry lung weight ratio,preserved aeration of alveoli as well as intravascular retentionof albumin, consistent with barrier protection. These observa-tions are consistent with our premise that the barrier-protectivemechanism facilitated by dopamine or quinpirole is mediatedby its interaction with VEGF signaling. Our hypothesis issubstantiated by the observation that pretreatment with dopa-mine significantly decreases VEGF level as well as VEGFR2phosphorylation. However, we do not exclude the possibility ofan effect on the delivery of VEGF to alveolar endothelium viarecruited monocyte/macrophages and neutrophils, both ofwhich have VEGF in their granules, especially given theobserved effect of dopamine on the inhibition of lung paren-chymal inflammation. Although our results are indicative ofthe fact that dopamine mediated barrier protection, in ourmurine model sepsis is predominantly mediated by VEGFpathway. There are several alternative or complementarymechanisms through which dopamine may influence the for-mation and clearance of pulmonary edema.
It is a well-established fact that the clinical use of dopaminemight significantly increase venous return and cardiac-fillingpressure in patients with cardiac dysfunction, sepsis, and re-spiratory failure (20, 45). We did not measure left atrial orpulmonary venous pressure in our animal model, but weobserved a significant reduction in the levels of lung watercontent, as assessed by weight and histology, in dopamine-treated mice. These results are in accord with the well-estab-
Fig. 7. Proposed model of D2DR role in LPS-induced pulmonary edema. LPSis known to induce overexpression of VEGF in the lung environment, leadingto VEGFR2 receptor phosphorylation. The activated VEGFR2 induces thephosphorylation of eNOS and, ultimately, leads to vascular permeability.Activation of D2DR impaired VEGFR2 receptor phosphorylation duringsepsis-induced lung injury. This could be the one possible mechanism thatinhibits pulmonary edema.
Fig. 6. Role of D2DR knockout on activationof eNOS in lung injury model. Immunostain-ing of histology slides with phospho-eNOS(Serine 1177) from wild-type (�/�, n � 6),heterozygous (�/�, n � 6), homozygousD2DR (�/�, n � 6) deleted mice treatedwith LPS alone (n � 6) or with quinpiroleand LPS (n � 6, A). Quantitative analysis ofimmunostained slides was performed usingmetamorph software (more than 3 slides pergroup, n � 6, B). We also monitored theanimal survival profile of each group (n � 8each group) for 32 h. We observed signifi-cant mortality in homozygously (�/�) de-leted D2DR mice (n � 0), even on quin-pirole pretreatment (n � 0). The quinpiroleimproved animal survival in heterozygous(�/�, n � 6) and wild-type (�/�, n � 6)animals (C).
L190 DOPAMINE INHIBITS PULMONARY EDEMA IN LUNG INJURY
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00274.2010 • www.ajplung.org
by 10.220.33.4 on April 19, 2017
http://ajplung.physiology.org/D
ownloaded from

lished effects of dopamine on alveolar fluid clearance in ALI(31, 34–37). Although we did not measure alveolar fluidclearance independent of lung water or protein permeability,we concluded that a significantly decreased concentration ofFITC-albumin in the BAL fluid of dopamine- and LPS-treatedmice would not be consistent with a clearance-dominatedmechanism of action. In contrast, with capillary fluid egress,alveolar fluid clearance remains the most viable and activeenergy-dependent process, mediated by coordinated interac-tions between apical sodium channels and basolateral Na/KATPase. Both type 1 and type 2 alveolar epithelial cellspossess the machinery for transepithelial ion transport, thoughwater simply moves along osmotic gradients (4). Typicalalveolar fluid clearance-measurement methods infer water fluxfrom changes in alveolar concentration relative to cell-imper-meable molecules such as albumin. Therefore, we anticipateda higher rather than lower FITC-albumin concentration in theBAL fluid of dopamine-treated mice if enhanced clearance hadbeen the dominant mechanism. The preponderance of our datasupports the fact that observed dopamine-induced change inalbumin extravasation likely arises from the changes in thehydraulic conductance. However, the conclusion that vasocon-striction prevents alveolar edema formation cannot be made ondata obtained from albumin extravasation ratio alone.
The numerous therapeutic interventions include simvastatin,adenosine triphosphate (ATP), sphingosine 1-phosphate (S1P)and activated protein C (APC), currently (23) being proposedto protect and maintain endothelial barrier integrity in ALI. Aprevious report from our group clearly enunciated the role ofD2DR in protection of endothelial barrier integrity and mod-ulation of vascular permeability (47). The above-mentionedreport became the premise for testing the role of a directmodulator of EC barrier function in our murine model of ALI(23). Excessive production of NO has also been implicated inpathophysiology of many diseases including sepsis. Previousreports have suggested that increased phosphorylation of eNOSupon lung injury plays a pivotal role in vascular permeability(8, 19, 21); however, a consensus has not been reached on themechanism of activation, as the signaling cues correlatingVEGF-NOS pathways has not been ascertained until now (24).In this context, a previous report from Knoll and colleaguessuggests that indeed VEGFR2 activation upregulates eNOSactivation and NO production (28). Another possibility is thatthere is intracellular cross talk between the D2DR signalingpathway and the phosphorylation of eNOS (44). None of thesepossibilities can be definitively excluded in the current find-ings. These observations are in sync with the earlier observa-tions pertaining to dopamine-mediated inhibition of eNOSphosphorylation in vivo and correspond to its role in vascularpermeability. Here, we have conclusively demonstrated for thefirst time that dopamine, acting through its D2 receptors, couldsignificantly attenuate pulmonary edema, suggesting that inex-pensive drugs such as dopamine or its specific D2DR agonistscould have potential use as an agonist in the treatment ofsepsis-induced lung injury (Fig. 7).
Although there is a large body of literature documenting thepathophysiology and treatment of severe sepsis, there is still anunacceptably high rate of mortality, emphasizing the need todevelop novel cost-effective, clinically effective therapeuticstrategies. We anticipate that the observations made in thecurrent study will form the rationale for novel cost-effective
therapeutic strategies. Further explorative studies are requiredto investigate the effects of other D2DR agonists and tissue-specific knockdown of D2DR to delineate the role in lunginjury. In summary, this study not only reveals a novel linkbetween D2DR and pulmonary edema but also indicates thatdopamine and or its D2DR agonists may have a therapeuticvalue in sepsis-induced lung injury.
Dopamine acts through D2DR to inhibit pulmonary edema-associated vascular permeability mediated through VEGF-VEGFR2 signaling, conveying protective effects in an ALImodel. We demonstrated that D2DR plays a critical role inpreventing VEGFR2 activation and the subsequent fluid influxassociated with lung injury. Our findings highlight the promiseof dopamine or D2DR agonists as potential novel therapies forthe treatment of pulmonary edema.
GRANTS
This work was partly supported by NIH grants CA78383, HL072178, andHL70567 and Bruce and Martha Atwater Foundation grant to D. Mukhopad-hyay.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
REFERENCES
1. Baltalarli A, Goksin I, Sirin H, Ortaç R, Onem G, Baltalarli B,Rendex O, Saçar M. Does anti-inflammatory therapy attenuate the lunginjury caused by ischemia/reperfusion of the lower extremities in therabbit? Int J Thoracic Cardiovasc Surg 3: 22, 2000.
2. Basu S, Nagy JA, Pal S, Vasile E, Eckelhoefer IA, Bliss VS, ManseauEJ, Dasgupta PS, Dvorak HF, Mukhopadhyay D. The neurotransmitterdopamine inhibits angiogenesis induced by vascular permeability factor/vascular endothelial growth factor. Nat Med 7: 569–574, 2001.
3. Becker PM, Alcasabas A, Yu AY, Semenza GL, Bunton TE. Oxygen-independent upregulation of vascular endothelial growth factor and vas-cular barrier dysfunction during ventilated pulmonary ischemia in isolatedferret lungs. Am J Respir Cell Mol Biol 22: 272–279, 2000.
4. Berthiaume Y, Matthay MA. Alveolar edema fluid clearance and acutelung injury. Respir Physiol Neurobiol 159: 350–359, 2007.
5. Bertorello AM, Sznajder JI. The dopamine paradox in lung and kidneyepithelia: sharing the same target but operating different signaling net-works. Am J Respir Cell Mol Biol 33: 432–437, 2005.
6. Bhandari V, Choo-Wing R, Lee CG, Yusuf K, Nedrelow JH, Amba-lavanan N, Malkus H, Homer RJ, Elias JA. Developmental regulationof NO-mediated VEGF-induced effects in the lung. Am J Respir Cell MolBiol 39: 420–430, 2008.
7. Bhattacharya R, Sinha S, Yang SP, Patra C, Dutta S, Wang E,Mukhopadhyay D. The neurotransmitter dopamine modulates vascularpermeability in the endothelium. J Mol Signal 3: 14, 2008.
8. Bucci M, Roviezzo F, Posadas I, Yu J, Parente L, Sessa WC, IgnarroLJ, Cirino G. Endothelial nitric oxide synthase activation is critical forvascular leakage during acute inflammation in vivo. Proc Natl Acad SciUSA 102: 904–908, 2005.
9. Chakroborty D, Sarkar C, Mitra RB, Banerjee S, Dasgupta PS, BasuS. Depleted dopamine in gastric cancer tissues: dopamine treatmentretards growth of gastric cancer by inhibiting angiogenesis. Clin CancerRes 10: 4349–4356, 2004.
10. Edirisinghe I, Arunachalam G, Wong C, Yao H, Rahman A, PhippsRP, Jin ZG, Rahman I. Cigarette-smoke-induced oxidative/nitrosativestress impairs VEGF- and fluid-shear-stress-mediated signaling in endo-thelial cells. Antioxid Redox Signal 12: 1355–1369, 2010.
11. Frey EA, Finlay BB. Lipopolysaccharide induces apoptosis in a bovineendothelial cell line via a soluble CD14 dependent pathway. MicrobPathog 24: 101–109, 1998.
12. Fujita M, Kuwano K, Kunitake R, Hagimoto N, Miyazaki H, KanekoY, Kawasaki M, Maeyama T, Hara N. Endothelial cell apoptosis inlipopolysaccharide-induced lung injury in mice. Int Arch Allergy Immunol117: 202–208, 1998.
L191DOPAMINE INHIBITS PULMONARY EDEMA IN LUNG INJURY
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00274.2010 • www.ajplung.org
by 10.220.33.4 on April 19, 2017
http://ajplung.physiology.org/D
ownloaded from

13. Fukumura D, Gohongi T, Kadambi A, Izumi Y, Ang J, Yun CO,Buerk DG, Huang PL, Jain RK. Predominant role of endothelial nitricoxide synthase in vascular endothelial growth factor-induced angiogenesisand vascular permeability. Proc Natl Acad Sci USA 98: 2604–2609, 2001.
14. Fulton D, Gratton JP, McCabe TJ, Fontana J, Fujio Y, Walsh K,Franke TF, Papapetropoulos A, Sessa WC. Regulation of endothelium-derived nitric oxide production by the protein kinase Akt. Nature 399:597–601, 1999.
15. Godzich M, Hodnett M, Frank JA, Su G, Pespeni M, Angel A,Howard MB, Matthay MA, Pittet JF. Activation of the stress proteinresponse prevents the development of pulmonary edema by inhibitingVEGF cell signaling in a model of lung ischemia-reperfusion injury inrats. FASEB J 20: 1519–1521, 2006.
16. Gropper MA, Wiener-Kronish J. The epithelium in acute lung injury/acute respiratory distress syndrome. Curr Opin Crit Care 14: 11–15, 2008.
17. Guerrero C, Lecuona E, Pesce L, Ridge KM, Sznajder JI. Dopamineregulates Na-K-ATPase in alveolar epithelial cells via MAPK-ERK-dependent mechanisms. Am J Physiol Lung Cell Mol Physiol 281: L79–L85, 2001.
18. Gurkan OU, O’Donnell C, Brower R, Ruckdeschel E, Becker PM.Differential effects of mechanical ventilatory strategy on lung injury andsystemic organ inflammation in mice. Am J Physiol Lung Cell Mol Physiol285: L710–L718, 2003.
19. Hatakeyama T, Pappas PJ, Hobson RW 2nd, Boric MP, Sessa WC,Duran WN. Endothelial nitric oxide synthase regulates microvascularhyperpermeability in vivo. J Physiol 574: 275–281, 2006.
20. Hess W, Bruckner JB, von Faber du Faur J, Schmidt D, Tarnow J.[The haemodynamic effects of dobutamine and dopamine in patients withcoronary artery disease A study performed under general anaesthesia(author’s transl)]. Anaesthesist 28: 316–321, 1979.
21. Hollenberg SM, Guglielmi M, Parrillo JE. Discordance between micro-vascular permeability and leukocyte dynamics in septic inducible nitricoxide synthase deficient mice. Crit Care 11: R125, 2007.
22. Hotchkiss CE, Hall PD, Cline JM, Willingham MC, Kreitman RJ,Gardin J, Latimer A, Ramage J, Feely T, DeLatte S, Tagge EP,Frankel AE. Toxicology and pharmacokinetics of DTGM, a fusion toxinconsisting of a truncated diphtheria toxin (DT388) linked to humangranulocyte-macrophage colony-stimulating factor, in cynomolgus mon-keys. Toxicol Appl Pharmacol 158: 152–160, 1999.
23. Jacobson JR, Garcia JG. Novel therapies for microvascular permeabilityin sepsis. Curr Drug Targets 8: 509–514, 2007.
24. Jonkam CC, Bansal K, Traber DL, Hamahata A, Maybauer MO,Maybauer DM, Cox RA, Lange M, Connelly RL, Traber LD, DjukomCD, Salsbury JR, Herndon DN, Enkhbaatar P. Pulmonary vascularpermeability changes in an ovine model of methicillin-resistant Staphylo-coccus aureus sepsis. Crit Care 13: R19, 2009.
25. Kaner RJ, Crystal RG. Pathogenesis of high altitude pulmonary edema:does alveolar epithelial lining fluid vascular endothelial growth factorexacerbate capillary leak? High Alt Med Biol 5: 399–409, 2004.
26. Kaner RJ, Ladetto JV, Singh R, Fukuda N, Matthay MA, Crystal RG.Lung overexpression of the vascular endothelial growth factor geneinduces pulmonary edema. Am J Respir Cell Mol Biol 22: 657–664, 2000.
27. Kosmidou I, Karmpaliotis D, Kirtane AJ, Barron HV, Gibson CM.Vascular endothelial growth factors in pulmonary edema: an update. JThromb Thrombolysis 25: 259–264, 2008.
28. Kroll J, Waltenberger J. VEGF-A induces expression of eNOS andiNOS in endothelial cells via VEGF receptor-2 (KDR). Biochem BiophysRes Commun 252: 743–746, 1998.
29. Leaver SK, Evans TW. Acute respiratory distress syndrome. BMJ 335:389–394, 2007.
30. Marwick JA, Edirisinghe I, Arunachalam G, Stevenson CS, MacneeW, Kirkham PA, Rahman I. Cigarette smoke regulates VEGFR2-mediated survival signaling in rat lungs. J Inflamm 7: 11, 2010.
31. Matthay MA. Alveolar fluid clearance in patients with ARDS: does itmake a difference? Chest 122: 340S–343S, 2002.
32. Matthay MA, Brower RG, Carson S, Douglas IS, Eisner M, Hite D,Holets S, Kallet RH, Liu KD, Macintyre N, Moss M, Schoenfeld D,Steingrub J, Thompson BT. Randomized, placebo-controlled clinicaltrial of an aerosolized beta-2 agonist for treatment of acute lung injury. AmJ Respir Crit Care Med 184: 561–568, 2011.
33. Matthay MA, Calfee CS. Therapeutic value of a lung protective venti-lation strategy in acute lung injury. Chest 128: 3089–3091, 2005.
34. Matthay MA, Clerici C, Saumon G. Invited review: Active fluid clearancefrom the distal air spaces of the lung. J Appl Physiol 93: 1533–1541, 2002.
35. Matthay MA, Robriquet L, Fang X. Alveolar epithelium: role in lungfluid balance and acute lung injury. Proc Am Thorac Soc 2: 206–213,2005.
36. Matthay MA, Uchida T, Fang X. Clinical acute lung injury and acuterespiratory distress syndrome. Curr Treat Options Cardiovasc Med 4:139–149, 2002.
37. Matthay MA, Zimmerman GA. Acute lung injury and the acute respi-ratory distress syndrome: four decades of inquiry into pathogenesis andrational management. Am J Respir Cell Mol Biol 33: 319–327, 2005.
38. Medford AR, Douglas SK, Godinho SI, Uppington KM, Armstrong L,Gillespie KM, van Zyl B, Tetley TD, Ibrahim NB, Millar AB. Vascularendothelial growth factor (VEGF) isoform expression and activity inhuman and murine lung injury. Respir Res 10: 27, 2009.
39. Mullane KM, Kraemer R, Smith B. Myeloperoxidase activity as aquantitative assessment of neutrophil infiltration into ischemic myocar-dium. J Pharmacol Methods 14: 157–167, 1985.
40. Murohara T, Asahara T, Silver M, Bauters C, Masuda H, Kalka C,Kearney M, Chen D, Symes JF, Fishman MC, Huang PL, Isner JM.Nitric oxide synthase modulates angiogenesis in response to tissue isch-emia. J Clin Invest 101: 2567–2578, 1998.
41. Patel GP, Grahe JS, Sperry M, Singla S, Elpern E, Lateef O, Balk RA.Efficacy and safety of dopamine versus norepinephrine in the managementof septic shock. Shock 33: 375–380, 2010.
42. Perkins GD, McAuley DF, Thickett DR, Gao F. The beta-agonist lunginjury trial (BALTI): a randomized placebo-controlled clinical trial. Am JRespir Crit Care Med 173: 281–287, 2006.
43. Pham I, Uchida T, Planes C, Ware LB, Kaner R, Matthay MA, Clerici C.Hypoxia upregulates VEGF expression in alveolar epithelial cells in vitroand in vivo. Am J Physiol Lung Cell Mol Physiol 283: L1133–L1142,2002.
44. Pyne-Geithman GJ, Caudell DN, Cooper M, Clark JF, Shutter LA.Dopamine D2-receptor-mediated increase in vascular and endothelialNOS activity ameliorates cerebral vasospasm after subarachnoid hemor-rhage in vitro. Neurocrit Care 10: 225–231, 2009.
45. Qvist J, Brynjolf I, Munck O. The effects of dopamine and noradrenalineon cardiovascular function in patients with acute respiratory failure. Eur JAnaesthesiol 11: 107–110, 1994.
46. Sarkar C, Chakroborty D, Mitra RB, Banerjee S, Dasgupta PS, BasuS. Dopamine in vivo inhibits VEGF-induced phosphorylation ofVEGFR-2, MAPK, and focal adhesion kinase in endothelial cells. Am JPhysiol Heart Circ Physiol 287: H1554–H1560, 2004.
47. Sinha S, Vohra PK, Bhattacharya R, Dutta S, Mukhopadhyay D.Dopamine regulates phosphorylation of VEGF receptor 2 by engagingSrc-homology-2-domain-containing protein tyrosine phosphatase 2. J CellSci 122: 3385–3392, 2009.
48. Tassiopoulos AK, Carlin RE, Gao Y, Pedoto A, Finck CM, LandasSK, Tice DG, Marx W, Hakim TS, McGraw DJ. Role of nitric oxideand tumor necrosis factor on lung injury caused by ischemia/reperfusion ofthe lower extremities. J Vasc Surg 26: 647–656, 1997.
49. Voelkel NF, Vandivier RW, Tuder RM. Vascular endothelial growthfactor in the lung. Am J Physiol Lung Cell Mol Physiol 290: L209–L221,2006.
50. Ware LB, Kaner RJ, Crystal RG, Schane R, Trivedi NN, McAuley D,Matthay MA. VEGF levels in the alveolar compartment do not distin-guish between ARDS and hydrostatic pulmonary oedema. Eur Respir J26: 101–105, 2005.
51. Ware LB, Matthay MA. The acute respiratory distress syndrome. N EnglJ Med 342: 1334–1349, 2000.
52. Ware LB, Matthay MA. Alveolar fluid clearance is impaired in themajority of patients with acute lung injury and the acute respiratorydistress syndrome. Am J Respir Crit Care Med 163: 1376–1383, 2001.
53. Wheeler AP, Bernard GR. Acute lung injury and the acute respiratorydistress syndrome: a clinical review. Lancet 369: 1553–1564, 2007.
54. Yano K, Liaw PC, Mullington JM, Shih SC, Okada H, Bodyak N,Kang PM, Toltl L, Belikoff B, Buras J, Simms BT, Mizgerd JP,Carmeliet P, Karumanchi SA, Aird WC. Vascular endothelial growthfactor is an important determinant of sepsis morbidity and mortality. J ExpMed 203: 1447–1458, 2006.
55. Zhang S, Rahman M, Qi Z, Herwald H, Thorlacius H. Simvastatinregulates CXC chemokine formation in streptococcal M1 protein-inducedneutrophil infiltration in the lung. Am J Physiol Lung Cell Mol Physiol300: L930–L939, 2011.
56. Zheng H, Chen XL, Han ZX, Zhang Z, Wang SY, Xu QL. Ligustrazineattenuates acute lung injury after burn trauma. Burns 31: 453–458, 2005.
L192 DOPAMINE INHIBITS PULMONARY EDEMA IN LUNG INJURY
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00274.2010 • www.ajplung.org
by 10.220.33.4 on April 19, 2017
http://ajplung.physiology.org/D
ownloaded from





























