Embed Size (px)
Citation preview

Austral Ecology
(2005)
30
, 1–13
Does biogeographical history matter? Diversity and distribution of lotic midges (Diptera: Chironomidae) in the Australian Wet Tropics
BRENDAN G. M
C
KIE,
1,2,3
* RICHARD G. PEARSON
1,2
AND PETER S. CRANSTON
2,3
1
School of Tropical Biology, James Cook University, Townsville, Queensland, Australia,
2
Rainforest Cooperative Research Centre, and
3
Entomology Department, University of California, Davis, California, USA
Abstract
We examined broad scale patterns of diversity and distribution of lotic Chironomidae (Diptera) withinthe Wet Tropics bioregion of northern Queensland, Australia. Field surveys across broad latitudinal and altitudinalgradients within the Wet Tropics revealed a fauna of 87 species-level taxa in 49 genera comprising three mainelements: a small genuinely tropical fraction, and larger cosmopolitan and Gondwanan components. The latter grouporiginated when Australia, as part of the ancient Gondwana supercontinent, was situated over Antarctic latitudeswith a cooler, wetter climate than today. In the Wet Tropics, cool Gondwanan taxa occurred predominantly in uplandand shaded lowland sites, but no species appeared narrowly temperature restricted, and there was no faunal zonationwith altitude. Most chironomid species occurred at all latitudes within the Wet Tropics, with no evidence for anenduring effect of the historical rainforest contractions on current-day distribution patterns. These findings contrastwith those for aquatic faunas elsewhere in the world and for the terrestrial Wet Tropics fauna. We relate this to thegenerally broad environmental tolerances of Australian chironomids, and comment on why the latitudinal diversitygradient does not apply to the Australian chironomid fauna.
Key words:
altitude, diversity gradients, faunal zonation, Gondwana, tropical streams.
INTRODUCTION
Knowledge of patterns of diversity and distribution oflotic organisms is deficient, especially outside thetemperate northern hemisphere and at very largespatial scales (Vinson & Hawkins 1998). Consequently,the applicability of a latitudinal diversity gradient tofreshwater ecosystems (Coffman 1989; Flowers 1991)and the generality of the humped relationship betweenaltitude and diversity exhibited by some northernhemisphere systems (Minshall 1985; Casas & Vilchez-Quero 1993) are both unclear. Previous study of thebroad-scale distributional patterns of lotic easternAustralian chironomids, assessed by sampling the castskins of emerging adults (pupal exuviae) at multiplelocations and times within a latitudinal gradient,showed no evidence for a larger regional species poolin the tropics of northern Queensland than in thetemperate south-east (Cranston 2000a). Furthermore,species believed restricted to cool south-eastern
streams, with biogeographical affinities (sister-taxonrelationships) with other Gondwanan landmasses(particularly southern South America and NewZealand) and assumedly from cool-adapted lineages,were found to have anomalous presence in Australia’stropical north (Cranston 2000a). Here we extend theseobservations by focusing on patterns of chironomiddiversity and distribution within northern Queens-land’s Wet Tropics bioregion. In particular, we assesshow restricted the cool Gondwanan component is in itsdistribution within the Wet Tropics.
The Wet Tropics bioregion, between the cities ofCooktown and Townsville along the east coast ofnorthern Queensland, Australia (Fig. 1), has highbut seasonal rainfall, with summer wet season falls(
ca
600 mm/month in the vicinity of Tully Gorge)substantially greater than those of the winter dry season(
ca
150 mm/month, Pearson 1994). These conditionsfavour tropical rainforests, which straddle a spine ofmountains that frequently rise to over 1000 m a.s.l.(Nix 1991; Goosem
et al
. 1999). Beyond the bioregion,the climate is hotter and drier, and rainforest gives wayto xeric vegetation types (Switzer 1991). Despitecovering just 0.01% of the surface of Australia(1 849 725 ha), the Wet Tropics supports a majorproportion of the continent’s terrestrial plant andanimal species, including 65% of its fern species, 36%
*Corresponding author. Present address: Department ofEcology and Environmental Science, Umeå University, SE-90187 Umeå, Sweden. E-mail: [email protected],Fax: + 46 90 7866705
.
Accepted for publication February 2004.
DIVERSITY AND DISTRIBUTION OF WET TROPICS CHIRONOMIDAE 2
of its mammals, 60% of its butterflies and 41% of itsfreshwater fishes (Pusey & Kennard 1996; Goosem
et al
. 1999). Regional endemism among the terrestrialfauna is also high, with 25% of the region’s plant andvertebrate species occurring nowhere else (Williams &Pearson 1997; Goosem
et al
. 1999). Similar data areunavailable for freshwater invertebrates, thoughdiversity of some freshwater insects (notablyTrichoptera) appears higher than in comparable areasof south-eastern Australia (Benson & Pearson 1988;
Lake
et al
. 1994), in contrast to Cranston’s (2000a)observations for chironomids.
The range of rainforests within the Wet Tropicshas oscillated with climatic fluctuations during theQuaternary, due to their specific temperature andrainfall requirements (Quilty 1994). Glaciation athigher latitudes was associated with a global reductionin rainfall, and during the last glacial maximum in thePleistocene (13 000–8000 years ago), most Australiantropical rainforest was displaced by drier sclerophyll
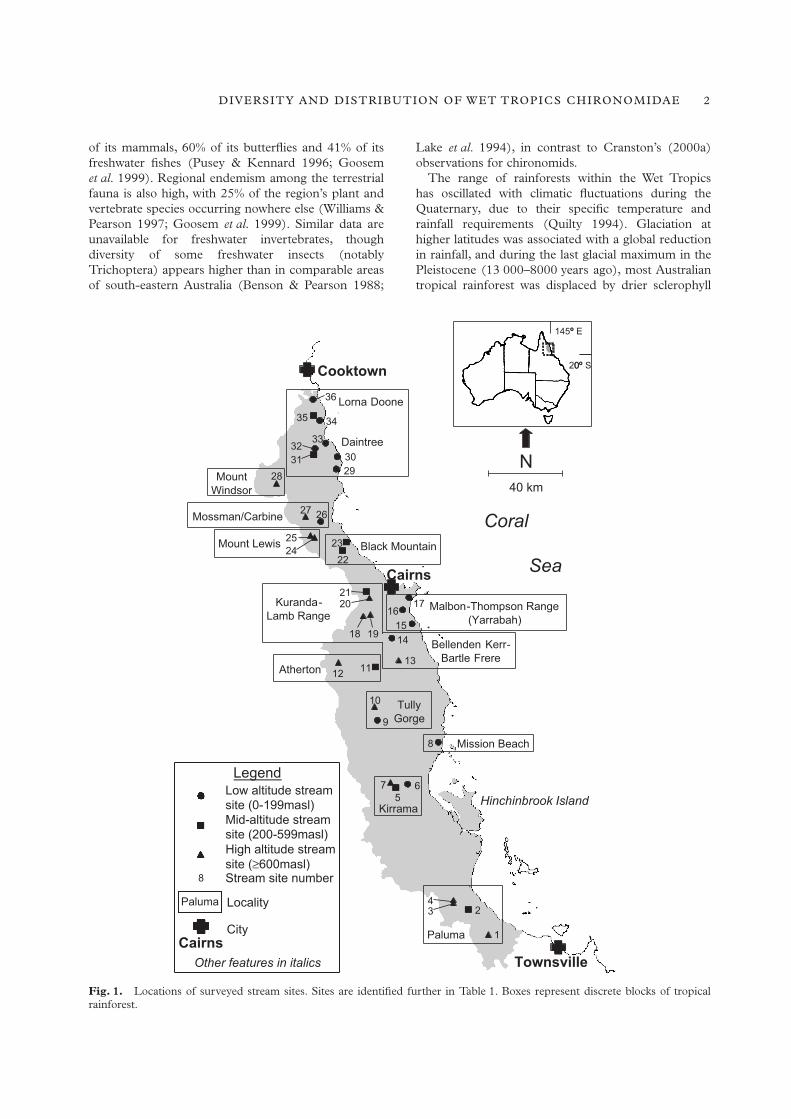
Fig. 1.
Locations of surveyed stream sites. Sites are identified further in Table 1. Boxes represent discrete blocks of tropicalrainforest.

3 B. G. MCKIE
ET AL.
forests. Isolated moist refugia did remain, the largestcentred on the Thornton and Atherton uplands(Fig. 1, Nix 1991). As the climate ameliorated, rainfallincreased and rainforest expanded to its current extent(Hopkins
et al
. 1996), but modern diversity anddistribution patterns of several groups of terrestrialorganisms bear the signature of past rainforest con-tractions. For example, diversity and endemicity ofvertebrates are greatest in the large refugial areas,which maintained the largest vertebrate populationsduring the glacial period (Williams & Pearson 1997;Williams 1997; Winter 1997). Patterns for flightlessterrestrial bugs and beetles are more complex,evidently because smaller rainforest refugia main-tained significant populations of rainforest arthropods
(Yeates
et al
. 2002; C. A. M. Reid pers. comm., 2000).In contrast, current-day diversity patterns of the WetTropics freshwater fish and Trichoptera faunas seemlittle affected by the Pleistocene contractions, with mostspecies homogeneously distributed throughout theregion’s latitudinal range (Pusey & Kennard 1996;Pearson in press).
Here we characterize the biogeographical affinities ofthe regional chironomid fauna, and consider relation-ships between patterns of diversity and distribution andboth historical (the Pleistocene rainforest contractions)and ecological (especially altitude) factors. We examineevidence derived from two species-level surveys oflotic Chironomidae. The first sampled the riffle-inhabiting fauna of 32 streams, covering the full
Table 1.
Streams sampled in the riffle and leaf pack surveys
Map LocalityMapref.
StreamName
Altitude(m a.s.l.)
Stream Order & Dominant Lithology Survey sampled
Paluma 1 Keelbottom 600 3 (Granite) R942 Little Crystal 400 3 (Granite-P) LP3 Birthday 800 3 (Granite-P) R93 & LP4 Camp 850 2 (Granite) R93 & LP
Kirrama 5 Goddard 500 3 (Granite) R936 Bridge 100 3 (Granite) R937 Yuccabine 600 3 (Granite) R93
Mission Beach 8 Limbo 20 3 (Alluvium/Met.)
†
R93Tully Gorge 9 Pixies 60 3 (Volcanics) R93 & LP
10 Charappa 620 3 (Granite) R93Atherton 11 North Johnson 500 3 (Granite) R93
12 Nigger 900 3 (Granite) R94Mt. Bellenden Kerr – Mt. Bartle Frere 13 Bartle Frere 1400 1 (?) R95
14 Kearney 80 3 (Basalt/Granite)
†
R94Malbon Thompson Range (Yarrabah) 15 Malbon Thompson 100 2 (Granite) R93
16 Simmonds 40 2 (Granite) R9317 Oombunghi 40 3 (Granite) R94
Kuranda – Lamb Range 18 Kauri 900 1 (Granite) R9319 Breach 700 3 (Granite) R9320 Clohesy 700 2 (Granite) LP21 Shoteel 450 2 (Metam.) R93 & LP
Black Mountain 22 Rifle 440 3 (Metam.) R9323 Allan 350 3 (Metam.) R93
Mount Lewis 24 Windmill 900 2 (Granite) R93 & LP25 Mary 900 2 (Granite) LP
Mossman/Mt. Carbine 26 Gorge 80 2 (Hodgk./Granite)† R9427 Carbine 1175 3 (Granite) R94
Mt. Windsor 28 Mt Windsor 960 3 (Granite) R94Daintree (Mount Thornton) 29 Oliver 20 3 (Hodgk.) LP
30 Emmagen 20 4 (Hodgk.) R93 & LP31 Baird 200 4 (Hodgk.) R9332 Bloomfield 20 4 (Hodgk.) R9333 Woobadda 100 4 (Hodgk.) R93
Lorna Doone (Far Northern) 34 Gap 150 2 (Granite) R9335 Wallaby 300 2 (Alluvium) R9336 Scrubby 140 3 (Granite) R94
†
Requires on-site verification. Map locality and ref. refer to Fig. 1. Lithology: Granite P, porphyrytic granite; Hodgk., Hodg-kinson granites and metamorphics, including siliceous cherts; Met., metamorphics. Survey sampled: R93, riffle survey 1993(November – December); R94, riffle survey 1994 (May – July); R95, riffle survey 1995 (November); LP, leaf pack survey(August – September 1997).

DIVERSITY AND DISTRIBUTION OF WET TROPICS CHIRONOMIDAE 4
latitudinal and altitudinal range of the Wet Tropics; thesecond sampled the fauna of pool leaf packs from 10streams.
METHODS
Riffle survey: sampling and identification
The 36 Wet Tropics streams sampled in the riffle andleaf pack surveys are listed in Table 1, with theirlocation plotted in Figure 1. Riffle-inhabiting chirono-mids were sampled from 32 of these sites over threeyears as part of a broader study of the region’s fresh-water communities (Pearson in press). All streams flowthrough tropical rainforest and are characterized byvery soft waters (hardness 1.8–21 mg/L) of low con-ductivity (22–111
�
S/cm) and variable but usuallymildly acidic pH (5.5–6.8) (Butler & Pearson 1998).On the mountain peaks, annual mean air temperaturesgenerally fall between 17 and 21
�
C, but occasionallydrop below zero in the winter. The coastal lowlands arewarmer, especially in the north of the bioregion wherethe average annual mean temperature is often above25
�
C (Nix 1991). More southerly lowlands are a fewdegrees cooler, and regimes on the inland tablelandsare similar to those of the mountain tops. Most sites(23) were sampled in the late dry season (Novemberand December) of 1993 (R93, Table 1). The remainingsites, expanding altitudinal, latitudinal and seasonalcoverage, were sampled in May and June 1994,following the end of the wet season, except for BartleFrere (the highest mountain in the region at 1622 ma.s.l.), which was sampled in November 1995 (R94-R95, Table 1).
At each stream site, two riffles were sampled bythoroughly disturbing the substrate within sixrandomly placed 25 cm
�
25 cm quadrats (three perriffle) upstream of a dip net (63
�
m mesh), withcollected material preserved in 70% ethanol. Because ofthe large number of larval chironomids found (approx.7400), individuals were identified to species level fromonly two of the six samples collected per site, one perriffle. Abundance was highly clumped, with one or twosamples from each site typically yielding many moreindividuals than the remainder, and so in each casethe samples selected were those yielding the greatestnumber of individuals. These samples represented57
�
9% (mean
�
standard deviation) of individualscollected per site. Diversity was also well represented inthese samples. For example, average richness per sitewas 71% of that found by Cranston (2000a), despitehis samples integrating all habitats, and richness of thetwo samples for one stream (Birthday Creek, 17species) was comparable to that assessed from sixriffles intensively sampled as part of another study in
1999 (19 species, BGM unpubl. data). Individualswere slide mounted in Hoyer’s aqueous solution andidentified using the keys of Cranston (1997a,b,2000b). Novel morphospecies with distinctivemouthpart morphologies were given a mnemonic(e.g. a number preceded by the letters ‘FNQ’ (FarNorth Queensland) for an undescribed putativegenus).
During the 1993 sampling trip, physico-chemicaldata were recorded from each site, as detailed inButler and Pearson (1998). Methodology followed theQueensland ‘State of the Rivers’ pro-forma (Andersen1993), and included measures of temperature, pH andconductivity, replicate measures (10) of width, depthand water flow over the stream reach sampled, as wellas estimates of substrate composition and percentcanopy cover (shading). By necessity, the exact time atwhich temperature measurements were taken variedamong sites, but all were taken in the shade duringdaylight hours, and temperatures of Wet Tropicsstreams typically vary by less than two degrees over a24-h period (B. G. McKie, P. Wulf and R. G. Pearson,unpubl. data, 2002).
Leaf-pack survey for chironomids of the Wet Tropics
As leaf packs support a different chironomid assem-blage from riffles, a survey of this habitat from 10streams was conducted in the 1997 dry season(August–September, Table 1), to broaden the under-standing of the Wet Tropics fauna. Samples consistedof fist-sized handfuls (volume approximately 300 cm
3
)of leaves, collected from dense accumulations inslower-flowing pools, which were placed directly into aplastic bag containing 70% alcohol. Five separate leafpacks were sampled over a reach of approximately100 m at each stream site. Chironomid larvae wereidentified as described above.
Biogeographical categorization
Where possible, species were allocated to biogeo-graphical groupings comprising Gondwanan taxa,with recent postulated phylogenetic (sister group)linkages to other Gondwanan landmasses, and cosmo-politan taxa. Within the Gondwanan group, twosubgroups can be recognized: cool, with currentsouthern temperate distributions; and warm, withmodern distributions in the warmer parts of thesouthern continents, often including tropical Africa.In contrast to these patterns, a cosmopolitan compo-nent comprises mostly broadly ranging taxa, bothwithin Australia and elsewhere, with various relation-ships, some to proximate landmasses to the north,

5 B. G. MCKIE
ET AL.
but without close phylogenetic association withGondwanan landmasses.
Analysis
Distributional data were ordinated using Non-metricMultidimensional Scaling (NMS, Clarke 1993).Similarities in chironomid faunal composition amongsites were assessed additionally using
UPGMA
(unweighted pair group arithmetic average) clusteranalysis (Sneath & Sokal 1973). These analyses wereconducted using the PC-ORD for Windows multi-variate analysis package (Version 4.0 1999, MjMSoftware, Gleneden Beach, Oregon, USA)
.
Addition-ally, an Analysis of Similarities (ANOSIM) wascarried out using Primer for Windows (Version 5.2.9,2002, Primer-E Ltd, Plymouth, UK) to assessdifferences between sites sampled in 1993 and 1994.Since the main interest was assessing similaritiesbetween sites, rather than groupings of sampleswithin sites, data for the two riffle or five leaf packsamples (in the riffle and leaf pack surveys, respec-tively) from each site were pooled prior to analysis.All analyses used Bray-Curtis dissimilarities as thedistance measure.
Following ordination, simultaneous plots of streamsand species in the ordination space were producedusing
PC-ORD.
Scores for the species were calculatedvia weighted averaging, whereby stream unit scores areused to weight the average position of each speciesalong the ordination axes. This allows for a visualassessment of relationships between species and sites,similar to a traditional principal components biplot, but
one which better accounts for non-linearity in theserelationships (McCune & Mefford 1999).
Riffle survey samples had been collected up to7 years prior to identification, and some were poorlypreserved. For three genera (the orthoclads
Coryno-neura
and
Thienemanniella
and the Chironominae
Tanytarsus
), antennal characteristics crucial forspecies recognition were damaged. Therefore, datawere pooled for two
Corynoneura
and two or three
Thienemanniella
species in the analyses. Two
Tanytarsus
species could be recognized from all sites, becausecharacters important for species recognition wereadequately preserved. Remaining
Tanytarsus
wereexcluded from analyses. These measures were unneces-sary for the leaf-pack survey.
Riffle survey data were additionally analysed usingunivariate regression, in order to assess relationshipsbetween species diversity and the altitude and latitudeof the study sites. Standard measures of speciesdiversity were used (evenness, species richness andthe Shannon-Wiener, Simpson and Berger-Parkerdiversity indices), calculated using the SpeciesDiversity & Richness program (Version 1.2, 1997,Pisces Conservation, Lymington, UK).
In the riffle survey, assemblage compositionappeared to be influenced by altitude (see Resultsbelow), and so univariate regression was used toexplore relationships between altitude and streamphysico-chemical characteristics. Where replicatemeasures were taken at each site (e.g. for depth,width and water flow), the response variable consistedof the average of the 10 readings. All regressions werecarried out using SPSS for Windows (version 10.0.5,1989–1999, SPSS Inc., Chicago, USA).
Fig. 2.
Riffle Survey: results from NMS ordination (Bray-Curtis dissimilarities) of ln(x + 1) transformed chironomidabundance data. Ordination in three dimensions, axes 1 & 2 plotted; stress = 0.16. (a) Stream site coordinates, with altitudecategories superimposed: (
�
) <200 m a.s.l., ( ) 200–599 m a.s.l., (
�
)
�
600 m a.s.l. (refer to Table 1 for site numbers); (b)species coordinates, calculated using weighted averaging, with biogeographical categories superimposed: (
�
) cosmopolitan ortropical, (
�
) warm Gondwanan, (
�
) cool Gondwanan, (
�
) other cool-temperate species, (+) uncertain .

DIVERSITY AND DISTRIBUTION OF WET TROPICS CHIRONOMIDAE 6
RESULTS
Composition of the Wet Tropics fauna
Raw data from the riffle and leaf pack Wet Tropicssurveys are available online at http://entomology.ucdavis.edu/chiropage/Wtsurvey. The 87 chironomidtaxa distinguished from the surveys are mostly distinctspecies, but for 4–6 genera, distinguishing larvalspecies on the basis of morphology is problematical,so the number of species found may be an underesti-mate. Fifteen of the 49 genera distinguished werenovel, and within previously recognized genera, somespecies were novel. Over half the genera found occurworldwide, though at least four are probably recentcolonisers from Asia. Approximately 25% of the fauna(at least 21 species in 14 genera) have Gondwananaffinities, either with warmer, tropical parts of theformer Gondwanan supercontinent (7 taxa, including
Nilotanypus
and
Harrisius
species)
,
or more usuallycool-temperate regions (at least 14 species, including
Echinocladius martini
,
Botryocladius grapeth
,
Australo-pelopia prionoptera
and
Riethia
species). Five of the coolGondwanan species were novel, but their morpho-logical affinity with described Gondwanan species iseasily recognized (McKie 2002). Several other speciesalso may have been Gondwanan, as otherwise world-wide genera such as
Rheotanytarsus
,
Polypedilum
and
Cricotopus
include species in the Wet Tropics withpresent-day distributional patterns centred onAustralia’s temperate south-east, similar to the coolGondwanan taxa mentioned above, suggestingcommon phyletic origins. Hereafter, these are referredto as ‘temperate-zone species’.
Riffle survey
Ordination and cluster analyses
Two separate analyses of riffle survey data were carriedout. The first focused only on the sites sampled in 1993whereas the second included data for all sites (seeTable 1). An ANOSIM testing for differences betweensamples collected in 1993 and 1994 was not significant(Global R = 0.094,
P
= 0.174), and since both the1993-only and combined-years analyses yielded similarresults, only the second is presented here.
Combined-years data were ordinated in threedimensions. In confirmation of the ANOSIM analysis,sampling season appeared not to affect assemblagecomposition, as sites sampled in 1994 were inter-spersed with those sampled in 1993. Rather, assem-blage composition was influenced by altitude, as mosthigh altitude sites separated from the remainder alongaxis 1 (Fig. 2a). Prominent in chironomid assemblages
Fig. 3.
Riffle Survey: Classification of all sites by
UPGMA
cluster analysis, superimposed with latitude ((
�
) >19
�
S, (
�
) 18–19
�
S,(
�
) 17–17.99
�
S, (
�
)
�
16
�
S) and altitude ((
�
) <200 m a.s.l., ( ) 200–599 m a.s.l., (
�
)
�
600 m a.s.l.) categories. The arrowedcluster contains all Daintree sites, other geographic groupings (Fig. 1) are widely dispersed through the dendrogram.

7 B. G. MCKIE
ET AL.
from these sites were several cool (southern) Gond-wanan species from the Orthocladiinae (for example,species j, o, p and q in Fig. 2b:
Echinocladius martini
,
Botryocladius grapeth
, novel species FNQ-2 and
Sticto-cladius
sp.) and species from cosmopolitan genera withmodern-day cool temperate, south-east Australiandistributions (temperate-zone species), includingspecies of
Rheotanytarsus
(species d and m in Fig. 2b)
,Polypedilum
(species k) and
Cricotopus
(species b andc). Mid and low altitude sites that were positioned withthe high altitude sites in the ordination (towards theright of axis one), such as Goddard, Bridge, Rifle andGap Creeks (streams 5, 6, 22 and 34 in Fig. 2) alsosupported several species from these biogeographicalgroupings (including the cool Gondwanan aphrotenine
Aphroteniella filicornis
, point r). Overall, speciesrichness of Gondwanan and other temperate-zonespecies was significantly related to altitude (regressionanalysis: richness = 3.6 + 0.002
�
altitude,
R
2
= 0.25,
P
= 0.003). The mean and standard deviation of rich-ness values for these groups in low-, mid- and highaltitude bands (as defined in Fig. 2a) were 3.7
�
1.7,4.6
�
1.7 and 5.7
�
1.6 species, respectively.The only cool-Gondwanan taxa found in multiple
sites positioned towards the opposite (left) side of the
ordination space were two
Riethia
species (the twospecies marked i in Fig. 2b) and the tanypod
Australo-pelopia prionoptera
(species l). No other temperate-zonespecies were common in these streams. More typical oflowland sites were species from tropical and cosmo-politan genera. For example, a cosmopolitan species of
Rheocricotopus
was rare at high altitudes but often veryabundant at lower altitude sites (point f), while thetropical
Thienemannimyia occurred exclusively atsites of lower altitude (point e). Tropical and cosmo-politan species of Rheotanytarsus, Dicroctendipes andPolypedilum (the three species plotted around point gin Fig. 2b) occurred at all altitudes, as did the warmGondwanan Nilotanypus and Harrisius (points hand n).
Some mid-and low altitude streams were positionedto the right of the ordination space but somewhat apartfrom the main cluster of high altitude sites. Mostdistant was Simmonds Creek (stream 16), which hadan unusual fauna with species that were rare orabsent elsewhere, including a novel cool-Gondwananorthoclad species (point s). Of the two sites occurringin the upper right-hand corner of the ordinationspace, Oombunghi Creek (stream 17) supported somecool Gondwanan and southern elements, such asEukiefferiella (species a) whereas North Johnson(stream 11) was a very low diversity site that groupedwith Oombunghi because of the mutual occurrence ofa few cosmopolitan and tropical species.
No groupings due to latitude were apparent andoverall, geographical proximity explained little of theclustering pattern in the UPGMA analysis (Fig. 3). Forexample, geographically adjacent Paluma sites,Birthday and Camp Creeks (numbers 3 and 4),and Lamb Range sites, Kauri and Breach Creeks(numbers 18 and 19), occur on widely divergentterminal branches. Similarly, although all fourDaintree sites (numbers 30–33) occur in a singlelarge cluster (arrowed in Fig. 3), within that clusterthese sites were mostly well separated. However,certain altitudinally similar sites clustered despitebeing widely separated geographically, includinghigh elevation sites Birthday Creek and Bartle Frere(13), and low elevation sites Kearney (14) and GorgeCreeks (26).
Table 2. Effect of stream altitude on three physical variables: mean � standard deviation (sd) for three altitudinal bands
Altitudinal band(m a.s.l.) n
Water Temperature (�C)(mean � SD)
Stream Width (m)(mean � SD)
Current Velocity (m s-1) (mean � SD)
< 200 9 23.77 � 2.07 16.02 � 6.06 0.21 � 0.12200–599 7 21.33 � 1.32 11.59 � 5.50 0.20 � 0.13� 600 7 19.57 � 2.11 9.56 � 2.24 0.10 � 0.03
Regression analyses: temperature = 24.2–0.006 � altitude, R2 = 0.64, P < 0.001; Width = 16.1–0.009 � altitude, R2 = 0.28,P = 0.009*; Velocity = –0.2–0.0004 � altitude, R2 = 0.23, P = 0.019*.
*Response variable was average of 10 readings, log transformed.
Fig. 4. Relationship between altitude and stream temper-ature of the 23 Riffle Survey sites sampled in the first trip(Table 1), with canopy shade categories ((�) <25%, (�) 25–49%, (�) 50–74%, () >75%) superimposed and selectedsites labelled with their stream number. Multiple regressionanalysis: Altitude t = –7.8, P < 0.001; Shade t = –4.7,P < 0.001; R2 = 0.832.

DIVERSITY AND DISTRIBUTION OF WET TROPICS CHIRONOMIDAE 8
The distribution of diversity and abundance
There were no relationships between either latitude oraltitude and any measure of species diversity, regardlessof whether regression analyses focused only on sitessampled in one season or on the combined dataset (e.g. regression analysis of species richness fromthe 1993 survey: richness = 12.8 + 0.002 � altitude,R2 = 0.049, P = 0.23; relationships with other indicesof even lower significance).
Altitude and stream physico-chemical characteristics
Three physico-chemical variables measured from the23 streams sampled during the (1993) survey tripdecreased significantly with increasing altitude: watertemperature, stream width and water current (Table 2).Multiple regression analysis indicated that both altitudeand shade affect stream temperatures in the WetTropics (altitude, t = –7.8, P < 0.001; shade t = –4.7,P < 0.001; R2 = 0.832). Heavily shaded low- and mid-altitude streams consistently fell below the regressionline plotted in Fig. 4, which predicts temperature forstreams of a given altitude.
Leaf pack survey
Leaf pack survey data were ordinated in two dimen-sions (Fig. 5), with higher altitude sites separating outfrom the remainder along axis 1. As in the riffle survey,species richness of cool Gondwanan and other tem-perate zone chironomids was significantly related toaltitude (regression analysis: richness = 2.6 + 0.005 �altitude, R2 = 0.47, P = 0.029). Cool-Gondwanan
species associated with upland sites included B. grapethand E. martini (points b and n in Fig. 5b), novelorthoclad species FNQ-3, FNQ-2 and FNQ-1 (pointsa, j and s), and to a lesser extent, the aphrotenineA. filicornis (point h). Also associated with higher alti-tude sites were temperate-zone species from the other-wise cosmopolitan genera Rheotanytarsus, Corynoneuraand Polypedilum (points p, q and r). Other species fromthese genera, known only from the tropics, were foundpredominately at low altitudes or at all elevations(points f, g and i), as were representatives of thecosmopolitan Rheocricotopus and Dicrotendipes (pointsk and o). As in the riffle survey, the cool Gondwanantanypod A. prionoptera (point d) and the warmGondwanan tanypod Nilotanypus and ChironominiHarrisius (points c and l) were found equally at allaltitudes, but in contrast with the earlier survey, the twocool Gondwanan Riethia species (points e and m) werefound predominately at high altitudes.
UPGMA analysis of the leaf pack data defined twodistinct clusters, one consisting entirely of sites fallingwithin the 17–16�S latitudinal band, and another withall the more southern sites plus the more northernShoteel and Windmill Creeks (numbers 21 and 24,Fig. 6). The two Paluma sites (numbers 3 and 4)clustered together, but other geographically adjacentsites occurred on widely separated branches (e.g.numbers 20 and 21, 24 and 25, 29 and 30).
DISCUSSION
Consistent trends in lotic chironomid distributionswithin the Wet Tropics bioregion related to altitude,
Fig. 5. Leaf Pack Survey: results from NMS ordination (Bray-Curtis dissimilarities) of ln(x + 1) transformed chironomidabundance data. Ordination in two dimensions, stress = 0.13. (a) Stream site coordinates, with altitude categories superimposed:(�) <200 m a.s.l., ( ) 200–599 m a.s.l., (�) �600 m a.s.l.; (b) species coordinates, calculated using weighted averaging, withbiogeographical categories superimposed (refer to Table 1 for site numbers); (b) species coordinates, calculated using weightedaveraging, with biogeographical categories superimposed: (�) cosmopolitan or tropical, (�) warm Gondwanan, (�) coolGondwanan, (+) other cool-temperate species, (+) uncertain.

9 B. G. MCKIE ET AL.
with high elevation assemblages more likely to includeorthoclad taxa with cool (southern) Gondwananaffinities (e.g. E. martini, B. grapeth), together withspecies from cosmopolitan genera with modern-day predominately cool temperate, south-easternAustralian distributions (e.g. Rheotanytarsus flabellatus,‘Cricotopus conicornis’). Some, if not all, cool-Gond-wanan genera appear to have originated in the Creta-ceous or early Tertiary (Brundin 1966; Cranston &Edward 1992; Cranston & Edward 1999) when proto-Australia, as part of the fragmenting Gondwanansupercontinent, was cooler than now (annual polar airtemperature range estimate: –5 to +8�C), with sea-iceprobably forming in the winter (Cranston & Naumann1991; Quilty 1994). Preponderance of cool Gond-wanan and other temperate zone taxa in cooler uplandlocations in an otherwise warm region seems related totemperature, one of the few stream physico-chemicalcharacteristics to vary systematically with altitude in theWet Tropics.
Cool Gondwanan orthoclads do occur in lower-elevation streams, although less frequently. Loweraltitudes in the Wet Tropics also support cooler-adapted terrestrial vertebrate faunas, but ‘only inthose areas characterized by high annual mean rainfalland frequent cloud cover, where mountains and ridgesdrop steeply to the sea’ (Nix 1991). This ‘mountainmass’ (‘massenerhebung’) effect (Nix 1991) mightsimilarly affect the fauna of streams that drop rapidlydown steep coastal escarpments (e.g. those east of therange in the Daintree and Malbon-Thompson low-lands; Fig. 1), limiting the scope for warming. Streamshading further suppresses lowland water temperatures(Fig. 4), with shaded lowland sites, such as Goddard(5), Bridge (6) and Gap (34) Creeks supporting coolGondwanan Orthocladiinae and other temperate zonespecies. In contrast, the less-shaded upland creeksYuccabine (7) and Charappa (10) were between twoand three degrees warmer than predicted for theiraltitude, and yielded no cool Gondwanan orthoclads.Thus upland and cooler lowland sites appear to permitpersistence in northern Queensland of cool temperate
southern elements, including several cool Gondwananspecies.
In contrast, a suite of apparently warm-tolerantspecies dominated the warmer lowland streams. Thissuite comprised representatives of a general Queens-land ‘background fauna’, which characterize the warm-hot lowland, often ephemeral, streams flowing throughopen shrubland, savannah and grassland ecosystemsoutside the Wet Tropics (e.g. species of Rheocricotopusand Dicrotendipes), circum-tropical taxa (e.g. Thiene-mannimyia and a species of Rheotanytarsus which isidentifiable with the Northern Territory and south-eastAsian Rheotanytarsus oss, Kyerematen et al. 2000) andwarm Gondwanan species of Nilotanypus andHarrisius. Cool Gondwanan species occurring fre-quently in warmer lowland systems may either haveevolved away from characteristic ‘cool-Gondwanan’physiologies (as has occurred for species from cool-Gondwanan radiations inhabiting warm systems incentral Australia, e.g. Cranston & Edward 1999)or else may be capable of behaviourally maintainingpreferred body temperatures (possible for the tanypodA. prionoptera, which being free-roaming may be betterable to exploit microclimate temperature differencesthan the more sedentary tubicolous Gondwananorthoclads).
However, whether any Australian lotic chironomidspecies is strictly ‘stenothermic’, or able to operate withmaximal efficiency over a narrow temperature range, isdebatable (Brundin 1966; McKie et al. in press). NoWet Tropics chironomid seemed narrowly constrainedby temperature in this study. Even cool Gondwananchironomids occur over a relatively broad range oftemperatures, particularly at a continental scale (watersbelow 8�C in Australia’s south-east, and above 20�C inthe tropics), compared with strictly cool stenothermousspecies from other regions. For example, Rossaro(1991a, 1991b) and Casas and Vilchez-Quero (1993)clearly distinguished alpine and/or winter-active cold-stenothermous chironomids in Italy and Spain, respec-tively. In tropical Africa and the Americas, high altitudemacroinvertebrate faunas (presumed cold-adapted) in
Fig. 6. Leaf pack survey: Classification of all sites by UPGMA cluster analysis, superimposed with ((�) >19�S, (�) 18–19�S,(�) 17–17.99�S, (�) �16�S) and altitude ((�) <200 m a.s.l., ( ) 200–599 m a.s.l., (�) �600 m a.s.l.) categories.

DIVERSITY AND DISTRIBUTION OF WET TROPICS CHIRONOMIDAE 10
mountain ranges over 2000 m a.s.l. can be identifiedthat are quite distinct from those inhabiting thesurrounding tropical lowlands (Harrison & Hynes1988; Flowers 1991; collated data in Jacobsen et al.1997). In the Wet Tropics, where the highest peak isunder 1700 m a.s.l., there is no discrete uplandchironomid fauna, only species that exhibit greaterabundance and frequency in upland locations. Even therestricted areas of south-eastern Australia subject toannual snowfall do not support chironomid faunasdistinct from nonalpine montane streams. Limitedsampling of streams in the alpine Kosciosko NationalPark (c. 36�S, > 2000 m a.s.l.) produced severalGondwanan species that also occur in the QueenslandWet Tropics, as well as species occurring at much loweraltitudes in south-eastern Australia (Boothroyd &Cranston 1995; Cranston 1997a; Cranston & Edward1999). The widespread distribution across broadlatitudinal and altitudinal bands of Australian runningwater chironomids show their relatively unconstrainedecology (Brundin 1966), as is substantiated byecophysiological studies demonstrating broad temper-ature tolerances, even for cool Gondwanan species(McKie et al. in press). Such broad tolerances mayreflect the paucity of alpine and strongly seasonalsystems, and the drought-related thermal fluctuationsthat Australian lotic systems are regularly but unpre-dictably subjected to (Finlayson & McMahon 1988;Lake 1995; Closs & Lake 1996). This reasoning doesnot apply to lentic chironomids, which seem moretemperature-restricted than the lotic fauna (Wright &Cranston 2000; Dimitriadis & Cranston 2001).
Surveys of lotic chironomid pupal exuviae con-ducted during 1998 (Cranston unpubl. data) alsorevealed a substantial cool-Gondwanan componentthat was distributed across altitudinal bands in the WetTropics. The exuvial surveys, which sampled midgesemerging from all in-stream habitats, including themost cryptic, also found several species unrecognizedin the larval survey, notably additional species ofAphroteniinae, Podonominae and Riethia, which addto the pool of Gondwanan taxa occurring in the WetTropics. Most of these Wet Tropics Gondwananchironomids are not endemic to the bioregion, sincethey are known from Australia’s south-east (andexceptionally the south-west). This contrasts with theterrestrial Wet Tropics biota, in which many of theGondwanan plant and vertebrate species are uniqueendemics (Winter 1997; Goosem et al. 1999). Thus,whereas the terrestrial rainforest community representsthe remaining fragments of a formerly widespreadecosystem that has now disappeared from most ofAustralia, the Wet Tropics populations of most Gond-wanan chironomids appear to represent ‘northernoutliers’ of distributions that remain variably extensivein cooler regions of the continent. This reflects thevagility of chironomids, and also the broad physio-
logical tolerances that characterize the Australian fauna(McKie et al. in press). Nevertheless, some putativeGondwanan taxa found in the Wet Tropics have notbeen recorded previously in Australia, either represent-ing formerly widespread Gondwanan taxa that havebecome extinct elsewhere in Australia, or species thatwere always more restricted in the former supercontinent (possibly evolving during Australia’snorthwards drift) and remain so today.
There is no evidence for an enduring effect ofhistorical rainforest contractions on current-daydistribution patterns of Wet Tropics Chironomidae.Our surveys showed no ‘hotspots’ of species richnessassociated with rainforest persistence during dry glacialperiods, as observed for terrestrial fauna (Williams1997). Such regions might be recognized also by theoccurrence of distinct assemblages of sensitive speciesmissing from localities affected more profoundly byrainforest contractions, but even the relativelytemperature-sensitive Gondwanan taxa occur in allmajor rainforest blocks, including the northern- andsouthernmost. These findings are understandable ifPleistocene aridity and cooling allowed maintenance ofstream shade and flow even as drier sclerophyllousvegetation replaced wet tropical rainforest. In thecurrent landscape, chironomid assemblages associatedwith suitably cool streams flowing through opensclerophyll forest, such as Davies Creek (520 m a.s.l. inthe Kuranda Range), harbour Gondwanan taxa andare undifferentiated from assemblages in closed rain-forest streams (P. S. Cranston unpubl. data, 1998).Furthermore, where Pleistocene aridity did inducexeric vegetation, streams often may have maintainedriparian strips of closed rainforest vegetation, as seentoday at the periphery of the southern Wet Tropics,where rainforest extends into blocks of sclerophyllousvegetation alongside wet stream channels (Pearson inpress). Where streams dried out completely, the vagilityof adult chironomids would allow recolonization asclimate ameliorated and flow returned, obscuring anyeffects of these events on distributional patterns.
Apart from the tendency for richness of cool Gond-wanan chironomids to increase with altitude, thegeneral distribution of diversity within the Wet Tropicswas homogeneous, with no linear or non-linearrelationships detected between species richness (orany other index) and either altitude or latitude (thoughthe latitudinal range covered in the surveys waslimited). Previous surveys elsewhere (Coffman 1989)have suggested higher chironomid diversity at middleto lower altitudes, supporting predictions (e.g. of theRiver Continuum Concept) relating to thermal habitatheterogeneity: high elevation streams tend to fluctuateless in temperature (being more buffered by ground-water and/or snow), restricting the range of thermaloptima for species to exploit (Vannote & Sweeney1980; Ward & Stanford 1982; Minshall 1985). It is

11 B. G. MCKIE ET AL.
unknown if the extent of diel and seasonal thermalfluctuation varies with altitude for Wet Tropics streams,but the broad thermal tolerances that seem character-istic of Australian lotic chironomids may preclude fine-scale niche differentiation according to temperature,reducing tracking of environmental differences alonggradients (McKie et al. in press). This may not applyfor other Australian freshwater groups, as diversity ofTrichoptera relates to altitude in the Wet Tropics, withhigher elevation sites supporting more species (Pearsonin press), possibly indicating greater stenothermy thanin the Chironomidae, or alternatively greater exploit-ation of specialized food sources or habitats rarer atlower altitudes (Wiggins & Mackay 1978).
Lotic ecosystems have been argued to provide ageneral exception to the trend for species diversity tobe greater in tropical regions (Coffman 1989; Flowers1991; Allan & Flecker 1993). Greater plant diversity oftropical rainforests, a postulated associate with speci-ation in terrestrial insect assemblages (Erwin 1988), isless likely to drive speciation among lotic insectsbecause aquatic leaf-shredders consume decayed andsoftened material leached of defensive chemicals, andthus are more generalist than terrestrial phytophages(Flowers 1991). The relative uniformity of habitatarchitecture characteristic of forested streams world-wide (with little variation in the basic substrates: rocks,sand, wood, leaves) also militates against divergence inspecies richness according to latitude – as suggested byCoffman and De La Rosa (1998), ‘the action offlowing water on geological materials is not latitudedependent’. Nonetheless, some evidence suggestsmacroinvertebrate species richness may be greater intropical streams (Fittkau 1971; Stout & Vandermeer1975; Minshall 1985; Pearson et al. 1986; Outridge1987; Lake et al. 1994; Yule 1995), although most ofthese comparisons suffer from inconsistent samplingmethods and/or taxonomy (as discussed by Lake et al.1994; Jackson & Sweeney 1995a). The Cranston(2000a) systematic survey does not suffer from theseproblems and revealed no latitudinal diversity gradientfor eastern Australian lotic chironomids, even account-ing for seasonal fluctuations in species composition andrichness. Maintenance of high diversity is favouredwhere species responses to environmental variation arewell-differentiated (Chesson 2000), but temperaturetolerances of Australian chironomids seem generallybroad (McKie et al. in press), and Wet Tropics loticmacroinvertebrates in general, and chironomids inparticular, tolerate remarkably wide oxygen, nutrientand sedimentation ranges (Connolly & Pearson 1998;Pearson & Connolly 2000; Connolly et al. 2004), allfactors which can vary in concert with flow in theAustralian environment (Lake 1995; Boulton & Brock1999). Such broad ecological tolerances make it lesslikely that environmental differences between tropicaland temperate systems would be associated with
divergence in species richness. However, Lake et al.(1994) also sampled eastern Australian streams in acomparable manner, and concluded that overall diver-sity of macroinvertebrates was higher in the tropics.This discrepancy may reflect differences betweenrichness at the sample scale, assessed by Lake et al.(1994), and the overall in-stream species pool, ordifferences in the ecological characteristics of the focaltaxa (chironomids compared with the entire macro-invertebrate assemblage). Clearly more research isrequired before generalizations can be made about theglobal patterning of lotic diversity and the processesunderlying those patterns.
ACKNOWLEDGEMENTS
Jacqui Nolan, Gordie Kovacs, Niall Connolly, LindaDavis and other staff of the Australian Centre forTropical Freshwater Research assisted with samplecollection and sorting. Funding for the riffle survey (aspart of the ‘Stream Classification Project’) was pro-vided by the Land and Water Resources Research andDevelopment Corporation (LWRRDC), and the Rain-forest Cooperative Research Centre funded the leafpack survey. The senior author was supported by anAustralian Postgraduate Award. We are grateful toLeon Barmuta, Andrew Boulton and Athol Mclachlan,who read the doctoral thesis on which this manuscriptis based and made several suggestions that greatlyimproved its quality, as did the comments of twoanonymous referees.
REFERENCES
Allan J. D. & Flecker A. S. (1993) Biodiversity conservation inrunning waters: identifying the major factors that threatendestruction of riverine species and ecosystems. Bioscience 43,32–43.
Andersen J. R. (1993) State of the Rivers Project Report 2:Implementation Manual. Department of Primary Industries,Queensland, Brisbane.
Benson L. J. & Pearson R. G. (1988) Diversity and seasonality ofadult Trichoptera captured in a light-trap at YuccabineCreek, a tropical Australian rainforest stream. Aust. J. Ecol.13, 337–44.
Boothroyd I. K. G. & Cranston P. S. (1995) Two Orthocladiinae(Chironomidae) genera common to New Zealand andAustralia: Pirara N. Gen. and Eukiefferiella Thienemann. In:Chironomids, From Genes to Ecosystems (ed. P. S. Cranston)pp. 389–408. CSIRO, Melbourne.
Boulton A. J. & Brock M. A. (1999) Australian FreshwaterEcology: Processes and Management. Gleneagles Publishing,Glen Osmond, South Australia.
Brundin L. (1966) Transantarctic Relationships and Their Signifi-cance as Evidenced by Chironomid Midges. Almqvist & Wiksell,Stockholm.
Butler B. & Pearson R. G. (1998) Overview of Physical, Chemicaland Biological Features of Wet Tropics Streams. ACTFR Report

DIVERSITY AND DISTRIBUTION OF WET TROPICS CHIRONOMIDAE 12
no. 98/26. Australian Centre for Tropical FreshwaterResearch, James Cook University, Townsville.
Casas J. J. & Vilchez-Quero A. (1993) Altitudinal distribution oflotic chironomid (Diptera) communities in the SierraNevada mountains (Southern Spain). Annls Limnol. 29,175–87.
Chesson P. (2000) Mechanisms of maintenance of speciesdiversity. Annu. Rev. Ecol. Syst. 31, 343–66.
Clarke K. R. (1993) Non-parametric multivariate analyses ofchanges in community structure. Aust. J. Ecol. 18, 117–43.
Closs G. P. & Lake P. S. (1996) Drought, differential mortalityand the coexistence of native and an introduced fish speciesin a south east Australian intermittent stream. Env. Biol.Fishes 47, 17–26.
Coffman W. P. (1989) Factors that determine the species rich-ness of lotic communities of Chironomidae. Acta Biol. Debr.Oecol. Hung. 3, 95–100.
Coffman W. P. & De La Rosa C. (1998) Taxonomic Compo-sition and Temporal Organization of Tropical and TemperateAssemblages of Lotic Chironomidae. J. Kansas Entomol. Soc.71, 388–406.
Connolly N. M. & Pearson R. G. (1998) Macroinvertebrates asMonitors of River Health in the Tropics: Appendix D – the Effectof Sedimentation on Benthic Invertebrates in Tropical Streams.LWRRDC report no. JCU 11 & ACTFR no. 98/20.Australian Centre for Tropical Freshwater Research, JamesCook University, Townsville.
Connolly N. M., Crossland M. R. & Pearson R. G. (2004) Effectof low dissolved oxygen on survival, emergence, and drift oftropical stream macroinvertebrates. J. N. Am. Benthol. Soc.23, 251–70.
Cranston P. S. (1997a) Identification Guide to the Chironomidae ofNew South Wales. Australian Water Technologies Pty Ltd,West Ryde.
Cranston P. S. (1997b) The Australian RheotanytarsusThienemann and Bause (Diptera: Chironomidae) revised,with emphasis on the immature stages. Invert. Taxon. 11,705–34.
Cranston P. S. (2000a) August Thienemann’s influence onmodern chironomidology – an Australian perspective. Verh.Internat. Verein. Limnol. 27, 278–83.
Cranston P. S. (2000b) Three new species of Chironomidae(Diptera) from the Australian Wet Tropics. Mem. QueenslandMus. 46, 107–27.
Cranston P. S. & Edward D. H. D. (1992) A sytematicreappraisal of the Australian Aphroteniinae (Diptera:Chironomidae) with dating from vicariance biogeography.Syst. Entomol. 17, 41–54.
Cranston P. S. & Edward D. H. D. (1999) Botryocladius Gen.n.:a new transantarctic genus of orthocladiine midge (Diptera:Chironomidae). Syst. Entomol. 24, 305–33.
Cranston P. S. & Naumann I. D. (1991) Biogeography. In: TheInsects of Australia, 2nd edn (ed I. D. Neumann) pp. 180–97.Melbourne University Press, Melbourne.
Dimitriadis S. & Cranston P. S. (2001) An Australian Holoceneclimate reconstruction using Chironomidae from a tropicalvolcanic maar lake. Palaeogeog. Palaeoclimatol. Palaeoecol.176, 109–31.
Erwin T. L. (1988) The tropical forest canopy: the heart of bioticdiversity. In: Biodiversity (eds E. O. Wilson & F. M. Peter)pp. 123–9. National Academy Press, Washington DC.
Finlayson B. L. & McMahon T. A. (1988) Australia vs. theworld: A comprehensive analysis of streamflow character-istics. In: Fluvial Geomorphology in Australia (ed. R. F.Warner) pp. 17–40. Academic Press, Sydney.
Fittkau E. J. (1971) Distribution and ecology of Amazonianchironomids (Diptera). Can. Entomol. 103, 407–13.
Flowers R. W. (1991) Diversity of stream-living insects innorth western Panama. J. North Am. Benthol. Soc. 10,322–34.
Goosem S., Morgan G. & Kemp J. E. (1999) Chapter 7. WetTropics. In: The Conservation Status of Queenslands Bio-regional Ecosystems (eds P. Sattler & R. Williams) pp. 1–55.Environmental Protection Agency, Brisbane.
Harrison A. D. & Hynes H. B. N. (1988) Benthic fauna ofEthiopian mountain streams and rivers. Arch. Hydrobiol.Supplement 81, 1–36.
Hopkins M., Head J., Ash J., Hewett R. & Graham A. (1996)Evidence of a Holocene and continuing recent expansion oflowland rain forest in humid, tropical North Queensland.J. Biogeog. 23, 737–45.
Jackson J. K. & Sweeney B. W. (1995a) Present status and futuredirections of tropical stream research. J. North Am. Benthol.Soc. 14, 5–11.
Jacobsen D., Schultz R. & Encalada A. (1997) Structure anddiversity of stream invertebrate assemblages: the influence oftemperature with altitude and latitude. Freshwater Biol. 38,247–61.
Kyerematen R., Andersen T. & Saether O. (2000) A review ofOriental Rheotanytarsus Thienemann & Bause, with descrip-tions of some new species (Insecta, Diptera, Chironomidae).Spixiana 23, 225–58.
Lake P. S. (1995) Of floods and droughts: river and streamecosystems of Australia. In: Ecosystems of the World, Vol. 22:River and Stream Ecosystems (eds C. E. Cushing, K. W.Cummins & G. W. Minshall) pp. 659–94. Elsevier,Amsterdam.
Lake P. S., Schreiber E. S. G., Milne B. J. & Pearson R. G. (1994)Species richness in streams: Patterns over time, with streamsize and with latitude. Verh. Internat. Verein. Limnol. 25,1822–6.
McCune B. & Mefford M. J. (1999) Multivariate Analysis ofEcological Data Vesion 4.0 (PC-Ord Help System). MjMSoftware, Gleneden Beach, Oregon.
McKie B. G. L. (2002) Multiscale abiotic, biotic and bio-geographic influences on the ecology and distribution of loticChironomidae (Diptera) in the Australian Wet Tropics. PhDThesis, School of Tropical Biology, James Cook University,Townsville.
McKie B. G., Cranston P. S. & Pearson R. G. (in press)Gondwanan mesotherms and cosmopolitan eurytherms:effects of temperature on the development and survival ofAustralian Chironomidae (Diptera) from tropical andtemperate populations. Mar. Freshwater Res.
Minshall G. W. (1985) Species richness in streams of differentsize from the same drainage basin. Am. Nat. 125, 16–38.
Nix H. A. (1991) Biogeography: pattern and process. In:Rainforest Animals: Atlas of Vertebrates Endemic to Australia’sWet Tropics (Kowari 1) (eds H. A. Nix & M. A. Switzer)pp. 10–39. Australian National Parks and Wildlife Service,Canberra.
Outridge P. M. (1987) Possible causes of high species diversityin tropical Australian freshwater macrobenthic communities.Hydrobiologia 150, 95–107.
Pearson R. G. (1994) Limnology in the northeastern tropics ofAustralia, the wettest part of the driest continent. Mitt.Internat. Verein. Limnol. 24, 155–63.
Pearson R. G. (in press) Biodiversity of the freshwater inver-tebrates of the Wet Tropics region of north-eastern Australia:patterns and possible determinants. In: Tropical Rainforests:

13 B. G. MCKIE ET AL.
Past, Present and Future (eds B. Birmingham, C. Dick &C. Moritz). University of Chicago Press, Chicago.
Pearson R. G., Benson L. G. & Smith R. E. W. (1986) Diversityand abundance of the fauna in Yuccabine Creek, a tropicalrainforest stream. In: Limnology in Australia (eds P. DeDecker & W. D. Williams) pp. 329–42. CSIRO, Melbourne.
Pearson R. G. & Connolly N. M. (2000) Nutrient enhancement,food quality and community dynamics in a tropical rain-forest stream. Freshwater Biol. 43, 31–42.
Pusey B. J. & Kennard M. J. (1996) Species richness andgeographical variation in assemblage structure of the fresh-water fish fauna of the wet tropics region of northernQueensland. Mar. Freshwater Res. 47, 563–73.
Quilty P. G. (1994) The background: 144 million years ofAustralian palaeoclimate and palaeogeography. In: History ofthe Australian Vegetation: Cretaceous to Recent (ed. R. S. Hill)pp. 14–43. Cambridge University Press, Cambridge.
Rossaro B. (1991a) Chironomids and water temperature. Aquat.Ins. 13, 87–98.
Rossaro B. (1991b) Factors that determine Chironomidaespecies distribution in fresh waters. Boll. Zool. 58, 281–6.
Sneath P. H. A. & Sokal R. R. (1973) Numerical Taxonomy. W.H.Freeman, San Francisco.
Stout J. & Vandermeer J. (1975) Comparison of species richnessfor stream-inhabiting insects in tropical and mid-latitudestreams. Am. Nat. 109, 263–80.
Switzer M. A. (1991) Introduction. In: Rainforest Animals: Atlasof Vertebrates Endemic to Australia’s Wet Tropics (Kowari 1) (edsH. A. Nix & M. A. Switzer) pp. 1–9. Australian NationalParks and Wildlife Service, Canberra.
Vannote R. L. & Sweeney B. W. (1980) Geographical analysis ofthermal equilibria: a conceptual model for evaluating theeffect of natural and modified thermal regimes on aquaticinsect communities. Am. Nat. 115, 667–95.
Vinson M. R. & Hawkins C. P. (1998) Biodiversity of streaminsects: variation at local, basin and regional scales. Annu.Rev. Entomol. 43, 271–93.
Ward J. V. & Stanford J. A. (1982) Thermal responses in theevolutionary ecology of aquatic insects. Annu. Rev. Entomol.27, 97–117.
Wiggins G. B. & Mackay R. J. (1978) Some relationshipsbetween systematics and trophic ecology in Nearctic aquaticinsects, with special reference to Trichoptera. Ecology 59,1211–29.
Williams S. E. (1997) Patterns of mammalian species richness inthe Australian tropical rainforests: are extinctions duringhistorical contractions of the rainforest the primary deter-minants of current regional patterns in biodiversity. Wildl.Res. 24, 513–30.
Williams S. E. & Pearson R. G. (1997) Historical rainforestcontractions, localized extinctions and patterns of vertebrateendemism in the rainforests of Australia’s wet tropics. Proc.R. Soc. Lond. B264, 709–16.
Winter J. W. (1997) Responses of non-volant mammals to thelate Quaternary climatic changes in the Wet Tropics regionof north-eastern Australia. Wildl. Res. 24, 493–511.
Wright I. & Cranston P. S. (2000) Are Australian lakes different?– Chironomid and chaoborid exuviae from Lake McKenzie,a coastal temperate dune lake. Verh. Internat. Verein. Limnol.27, 303–8.
Yeates D. K., Bouchard P. & Monteith G. B. (2002) Patternsand levels of endemism in the Australian Wet Tropicsrainforest: evidence from flightless insects. Invert. Syst. 16,605–19.
Yule C. M. (1995) Benthic invertebrate fauna of an aseasonaltropical mountain stream on Bougainville Island, Papua NewGuinea. Mar. Freshwater Res. 46, 507–18.





























