Embed Size (px)
Citation preview

Research
© The Authors (2009) New Phytologist (2009) 182: 495–506 495Journal compilation © New Phytologist (2009) www.newphytologist.org 495
Blackwell Publishing Ltd
Diversispora celata sp. nov: molecular ecology and phylotaxonomy of an inconspicuous arbuscular mycorrhizal fungus
Hannes A. Gamper1, Christopher Walker2 and Arthur Schüßler3
1Netherlands Institute of Ecology (NIOO-KNAW) – Centre for Terrestrial Ecology, Department of Terrestrial Microbial Ecology, Boterhoeksestraat 48,
PO Box 40, 6666 ZG Heteren, The Netherlands; 2Royal Botanic Garden Edinburgh, 20A Inverleith Row, Edinburgh EH3 5LR, UK; 3Genetics,
Department of Biology I, Ludwig-Maximilian-University Munich, Großhadernerstrasse 4, 82152 Planegg-Martinsried, Munich, Germany
Summary
• The increasing numbers of taxonomically unassigned phylotypes reported inmolecular ecological studies contrast with the few formally described arbuscularmycorrhizal fungi (AMF; Glomeromycota). Here, a species new to science withGlomus-like spores is phylogenetically, morphologically and ecologically characterized.• From single spore isolates of a previously recognized member of the Diversisporaceaefrom Swiss agricultural grassland, 17 new nuclear internal transcribed spacer (ITS),large subunit (LSU) and small subunit (SSU) ribosomal RNA (rRNA) gene sequenceswere determined and compared with 14 newly generated sequences of two closerelatives and public database sequences, including environmental sequences, ofknown geographic origin.• SSU ribosomal DNA (rDNA) sequence signatures and phylogenies based on ITS,LSU and SSU rDNA sequences show that the fungus belongs to the genus Diversispora.It is described as Diversispora celata sp. nov. Comparison with environmentalsequences in the public domain confirmed its molecular genetic distinctiveness andrevealed a cross-continental distribution of close relatives.• The value of combining morphology and phylogeny to characterize AMF wasreinforced by the morphological similarity to other species and the inconspicuousnature of D. celata spores and mycorrhizas. Inclusion of all three nuclear rDNAregions in species descriptions will facilitate species determination from environmentalphylotypes.
Author for correspondence:Hannes GamperTel: +31 26 4791205Fax: +31 26 4723227Email: [email protected]
Received: 30 October 2008Accepted: 30 November 2008
New Phytologist (2009) 182: 495–506doi: 10.1111/j.1469-8137.2008.02750.x
Key words: arbuscular mycorrhiza, biogeography, Diversisporales, molecular phylogeny, new species, rDNA, spore morphology, taxonomy.
Introduction
The recognition of species, whether named or unnamed, isfundamental to understanding how diversity affects ecosystemprocesses (Tilman et al., 2001; Zak et al., 2003). However,for arbuscular mycorrhizal fungi (AMF) of the phylumGlomeromycota (Schüßler et al., 2001b), the detection anddiscrimination of species are difficult and many are inadequatelydefined. Natural communities usually harbour AMF fromseveral phylogenetic clades. Progress in ascribing physiologicaland ecological traits to particular AMF taxa or lineages (Hart &Reader, 2002; Munkvold et al., 2004; Maherali & Klironomos,2007) largely depends on their molecular characterization andthe availability of pure reference cultures. Once traits havebeen ascribed to particular species, or phylotypes, it may bepossible to predict the influence of AMF on plant growth in
the field and to understand ecosystem processes (van derHeijden & Scheublin, 2007). Molecular characterization,therefore, forms a foundation for ecological enquiries, whichalso require modern species descriptions to be integrated intocurrent ecological knowledge.
Species and phylotypes from the Glomeromycota are oftenwidely distributed geographically, probably as a result of their longbiogeographical history (Pirozynski & Malloch, 1975; Redeckeret al., 2000). However, the substantial environmental phylotypicrichness (Öpik et al., 2006, 2008; Vandenkoornhuyse et al., 2007,Kottke et al., 2008) is much greater than the approximately 200formally described AMF species (www.amf-phylogeny.com).Thus, the vast majority of existing species are not formallynamed. Moreover, the majority of described species have beenneither sequenced nor established as cultured mycorrhizalsymbionts, thus precluding further characterization.

New Phytologist (2009) 182: 495–506 © The Authors (2009)www.newphytologist.org Journal compilation © New Phytologist (2009)
Research496
The suitability of molecular genetic markers for comprehensiveecological community sampling can be compromised either byinadequate taxon resolution, for example as a result of DNAsequence conservation, or by failure to detect lineages, for exampleas a result of selectivity of PCR primers (Schüßler et al., 2001a;Walker et al., 2007). To address such problems, more reliable andcomprehensive sequence databases are needed that are based onaccurately identified organisms (Schüßler et al., 2003; Vilgalys,2003; Bidartondo et al., 2008; Nilsson et al., 2008).
Characterizations and descriptions of AMF species are bytheir nature limited by the state of knowledge and theavailability of reference material when they were made, andspecies identification solely on morphological grounds may beimpossible, for example in the Diversisporales and Archaeosporales(Walker & Schüßler, 2004; Walker et al., 2007). Provision ofsequence information in species descriptions is thereforeessential for correct classification and may inform new speciesdescriptions and taxonomic revisions. For molecular ecologicalinvestigations of phylotaxonomic richness and application ofAMF in agriculture and horticulture, species recognition andidentification must be simple and reliable. Methods used infield ecology should be applicable to all life stages, includingmycelia in the soil and roots, and are often based on nuclearribosomal DNA (rDNA) markers (Redecker et al., 2003).
The Diversisporales contain four AMF families, the Acau-losporaceae, Gigasporaceae, Entrophosporacea and Diversisporaceae.The first two are reasonably well circumscribed, whereas theEntrophosporaceae were emended to contain only two species(Sieverding & Oehl, 2006) without verification of their plac-ing in the Diversisporales. The Diversisporaceae encompass spe-cies with very distinct spore morphologies, including thosewith ‘Glomus-like’ (glomoid) spores that can be identified onlyby a combination of molecular and morphological characters(Walker et al., 2004a,b; Walker & Schüßler, 2004).
Here we report the characterization of such a Glomus-likefungus, which was isolated from a fertile grassland in Switzerlandwhere it is a common root symbiont (Gamper & Leuchtmann,2007). Its molecular characterization had already shown it tobelong in the Diversisporaceae, but our detailed morphologicalstudies and new sequence data for the nuclear internal tran-scribed spacer (ITS) region and large subunit (LSU) rRNAgene show that the AMF is a species new to science. Phylogeneticcomparisons with environmental rDNA sequences from publicdatabases were made to study the geographic distribution ofits known relatives. The isolates used in this study, includingthe type culture, are available to the research community ininternational culture collections.
Materials and Methods
Isolation and pure culturing
Trap cultures were established with soil from permanent mono-culture plots of Trifolium repens cv. Milkanova in the Swiss
long-term Free-Air CO2 Enrichment (FACE) experiment inEschikon, 20 km north-east of Zurich (47°27′N, 8°41′E, 550 masl). After 5 months, Plantago lanceolata L. seedlings were inoculatedwith single spores of the different morphotypes found, resultingin four pure isolates, two from plots subjected to elevatedatmospheric pCO2 field conditions (60 Pa; isolates FACE 234and FACE 272) and two from plots subjected to ambientatmospheric pCO2 field conditions (FACE 83 and FACE 410).FACE 83 was trapped with Allium porrum L. cv. Dubouchetand Glycine max (L.) Merrill as host plants, and the others withLeontodon autumnalis L. and Dipsacus fullonum L. as host plants.Single spore isolate FACE 234, specimens of which were chosenas the TYPUS of D. celata sp. nov., was registered and releasedto the scientific research community under the accession numberBEG231 by depositing it in the International Bank for theGlomeromycota (BEG; http://www.kent.ac.uk/bio/beg/). Theother isolates were also registered and are available under thefollowing numbers: BEG230 (FACE 83), BEG232 (FACE272) and BEG233 (FACE 410). All four isolates were alsosent to the International Culture Collection of ArbuscularMycorrhizal Fungi (INVAM; http://invam.caf.wvu.edu/) inthe USA. See Supporting Information Methods S1 and S2 forfurther details about the site of origin, culturing conditions,culture histories, and other cultures analysed.
DNA extraction, PCR amplification, cloning and sequencing
Genomic DNA templates for the isolates listed above and forGlomus aurantium J. Blaszk., V. Blanke, C. Renker & F. Buscotand Glomus eburneum L.J. Kenn., J.C. Stutz & J.B. Morton(INVAM AZ420A) were prepared from single and multiplespores following the protocols of Schwarzott & Schüßler(2001), Jansa et al. (2003) and Gamper & Leuchtmann (2007).Fragments of more variable domains of the nuclear LSUrRNA and SSU rRNA genes were amplified after Gamper &Leuchtmann (2007) and Helgason et al. (1998), respectively.Near full-length SSU rRNA gene fragments were generatedfollowing Schwarzott & Schüßler (2001), and the nuclear ITSregion was amplified as in Walker et al. (2007). All PCRamplicons were cloned for bidirectional plasmid sequencingon Prism®310, 3100, 3730xl ABI automated capillarysequencers (Applied Biosystems, Forsters City, CA, USA).Details are provided in Supporting Information Methods S3.
Phylogenetic sequence analyses
All three nuclear rDNA sequence partitions were phylogeneticallyanalysed using the maximum likelihood (ML) algorithms oftree-puzzle (version 5.2; Schmidt et al., 2002) and phyml(version 2.4.4; Guindon & Gascuel, 2003), Bayesian inference(BI) in MrBayes (version 3.1.2; Ronquist & Huelsenbeck,2003), and maximum parsimony (MP) and neighbour joining(NJ) tree searches in paup (version 4b10; Swofford, 2003).

© The Authors (2009) New Phytologist (2009) 182: 495–506Journal compilation © New Phytologist (2009) www.newphytologist.org
Research 497
Separate best-fit sequence evolutionary models for thedifferent data sets were determined in ModelTest (version3.7; Posada & Crandall, 1998) by hierarchical likelihood ratiotesting and branch support evaluated using bootstrap analyses.
To set the results in a broader ecological and biogeographicalcontext, an additional multiple sequence alignment for theSSU rRNA gene, mainly with environmental sequences, andan alignment with ITS region sequences of members of theDiversisporaceae were created. The environmental sequencesrepresent close matches to the new cultures in BLASTn searchesin the public databases. The SSU rDNA data set covers referencesequences of the Diversisporales and all near full-lengthsequences of the Diversisporaceae. The LSU rDNA and ITSregion data sets incorporate all Diversisporaceae sequences. Allphylogenetic trees were rooted with Gigasporaceae sequences,except for the SSU rRNA gene tree of the Diversisporales forwhich members of Glomus groups A and B (Schwarzott et al.,2001) were chosen as an outgroup. The new ITS, LSU and SSUsequences were deposited in the public databases under theaccession numbers listed in Supporting Information Table S1.The alignments (SSU rDNA, LSU rDNA and ITS regionrDNA) are available at www.amf-phylogeny.com. Furtherdetails are given in Supporting Information Methods S4.
Morphological analyses
Specimens were examined morphologically using the techniquespublished in Walker et al. (1993), and the formal descriptionfollowed the rules of the current Botanical Code (http://ibot.sav.sk/icbn/main.htm). Full details of the analyses areprovided in Supporting Information Notes S1.
Results
Four single spore isolates (FACE 83, 234, 272 and 410) of anAMF producing spores morphologically indistinguishable fromeach other were established. This AMF corresponds to a commonroot symbiont at the site of origin that had previously beendetected with a taxon-specific LSU rDNA PCR amplificationassay (Gamper & Leuchtmann, 2007). Molecular analysis ofthe SSU rRNA gene and ITS rDNA region showed the isolatesto be indistinguishable from each other and different from anyexisting species characterized. Similarly, the morphologicalexamination showed that no other described species shared itscharacteristics. The conclusion was drawn that the isolatesrepresented a new species in the Diversisporaceae and a formalspecies description is presented to comply with the rules laidout in the Botanical Code. More comprehensive details areprovided in Supporting Information Notes S1–S3.
Taxonomy of Diversispora celata sp. nov.
Diversispora celata C. Walker, Gamper and Schuessler sp. nov.(Fig. 1).
Sporae juventute hyalinae, tum eburneae vel carneo-eburneae,maturitate ochraceae, globosae, subglobosae, ovoideae, obovoideaevel interdum irregulares, 5–160 × 48–192 µm. Hypha subtentensrecta, recurvata vel infundibuliformis, septo occlusa. Parietissporarum partibus tribus in turmis duabus. Pars 1 evanescens,hyalina, arcte adhaerens a partem 2 laminarem, luteum. Pars 3flexibilibus, fortasse post obturamentum sporae efficerens.
Spores formed singly and in loose clusters in the soil;colourless when young, becoming ivory (2) to pinkish cream(4), and eventually ochre (9) with age; globose to subgloboseto ellipsoid, ovoid or obovoid, occasionally irregular; 53–160 × 48–192 µm; with a straight to recurved, parallel-sidedto funnel-shaped subtending hypha, occluded by a septumformed from the laminated structural wall component. Sporewall structure of three components in two groups. Compo-nent 1 evanescent, thin (< 1 µm), colourless, often with finesoil particles incorporated in a matrix of variable thickness,tightly adherent to laminated, yellow component 2 that is2–5 µm thick. Wall component 3 flexible, much less than1 µm thick, possibly developing after spore occlusion. Placed inthe genus Diversispora (Diversisporaceae) by its SSU sequencesignatures and distinguished from other members of thegenus by its nuclear ITS region and LSU rDNA sequences.
Collections examined All from pot cultures with P. lanceolataestablished with spores from soil-trap pot cultures. Switzerland,field station of the Swiss Federal Institute of Technology(ETH) Eschikon-Lindau ZH, near Zurich (47°27′N, 8°41′E).Isolate FACE 234: single spore isolate taken 20 Jul. 2005,H. Gamper (Att 1278-2, W4718 TYPUS E, Isotype MSB);loc. cit., 27 Jul. 2005, H. Gamper (Att 1278-2, W4719) (E);loc. cit. from a subculture of Att 1278-2, H. Gamper 2 & 3Jun. 2008) (Att 1278-3, W5461, W5462); loc. cit. from asubculture of Att 1278-2, H. Gamper 2 & 15 Feb. 2008)(Att 1278-5, W5401).
Etymology From the Latin, celator, a concealer, referring tothe small size and pale colour of the spores, which make themdifficult to see (concealed) in substrate extractions.
Distribution and habitat Isolated from an agricultural grasslandin Switzerland and cultivated in pot culture with P. lanceolata,Glycine max, Allium porrum, Leontodon autumnalis and Dipsacusfullonum. Forming arbuscular mycorrhizas; vesicles not observed.
See Supporting Information Notes S1 and S2 for details ofthe morphology of D. celata sp. nov. and comparisons withother members of the Glomeromycota.
Molecular phylogeny and distinctiveness of D. celata
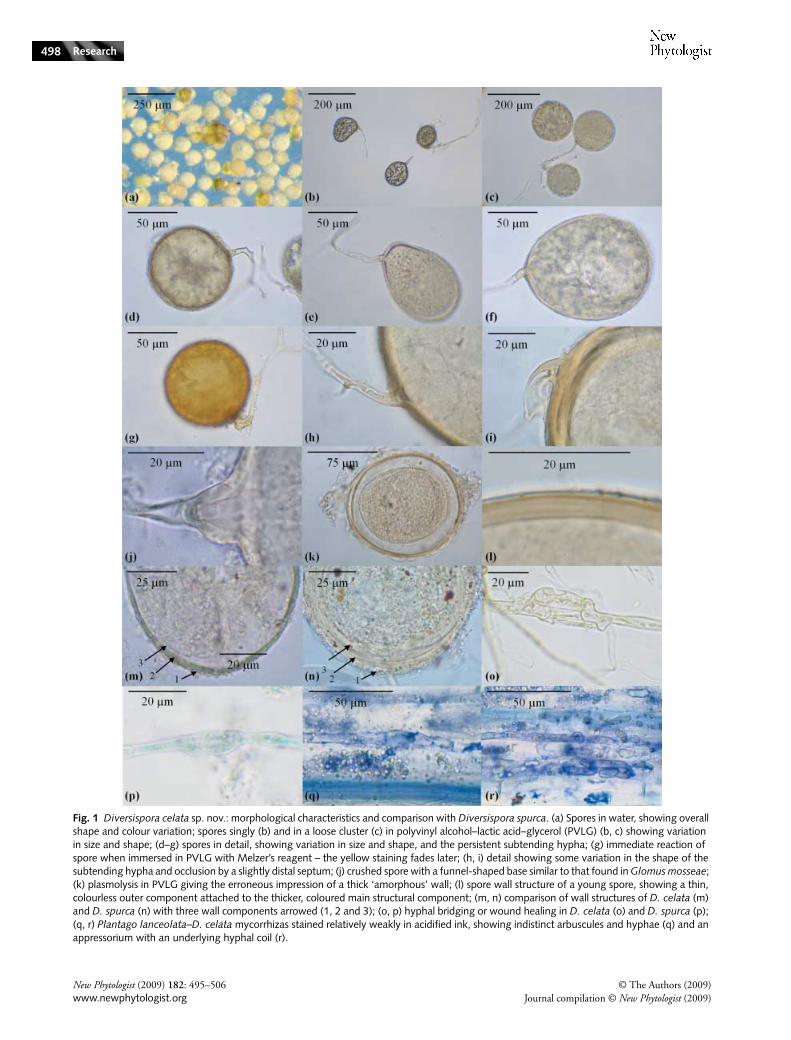
Analyses of near full-length nuclear SSU rRNA and partial LSUrRNA gene sequences revealed that D. celata is phylogeneticallyaffiliated with other members of the genus Diversispora(Figs 2, 3). In the SSU rRNA gene tree, relatively high
New Phytologist (2009) 182: 495–506 © The Authors (2009)www.newphytologist.org Journal compilation © New Phytologist (2009)
Research498
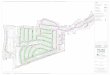
Fig. 1 Diversispora celata sp. nov.: morphological characteristics and comparison with Diversispora spurca. (a) Spores in water, showing overall shape and colour variation; spores singly (b) and in a loose cluster (c) in polyvinyl alcohol–lactic acid–glycerol (PVLG) (b, c) showing variation in size and shape; (d–g) spores in detail, showing variation in size and shape, and the persistent subtending hypha; (g) immediate reaction of spore when immersed in PVLG with Melzer’s reagent – the yellow staining fades later; (h, i) detail showing some variation in the shape of the subtending hypha and occlusion by a slightly distal septum; (j) crushed spore with a funnel-shaped base similar to that found in Glomus mosseae; (k) plasmolysis in PVLG giving the erroneous impression of a thick ‘amorphous’ wall; (l) spore wall structure of a young spore, showing a thin, colourless outer component attached to the thicker, coloured main structural component; (m, n) comparison of wall structures of D. celata (m) and D. spurca (n) with three wall components arrowed (1, 2 and 3); (o, p) hyphal bridging or wound healing in D. celata (o) and D. spurca (p); (q, r) Plantago lanceolata–D. celata mycorrhizas stained relatively weakly in acidified ink, showing indistinct arbuscules and hyphae (q) and an appressorium with an underlying hyphal coil (r).
© The Authors (2009) New Phytologist (2009) 182: 495–506Journal compilation © New Phytologist (2009) www.newphytologist.org
Research 499
Fig. 2 Phylogram of cultured and/or voucher-based members of the Diversisporales derived from nuclear small subunit (SSU) rRNA gene sequences, indicating the family affiliation of Diversispora celata (grey box). The tree topology and molecular distances (nucleotide substitutions per site) were derived from maximum likelihood (ML) inferences based on 1692 aligned and 275 parsimony-informative sites, using TREEPUZZLE. Representatives of the Glomeraceae were chosen to root the tree. Support values from quartet puzzling, Bayesian, ML, maximum parsimony (MP) and neighbour-joining distance analyses are shown. Consistent branch support of > 80% among all five analysis methods is indicated by thick branches. Branch support values are only given if at least three analysis methods yielded ≥ 50%; MP tree length 1054, consistency index (CI) = 0.503. If available, culture, attempt, voucher numbers, and geographic origin are shown. Genus abbreviations: D, Diversispora; Gi, Gigaspora; G, Glomus; P, Pacispora; S, Scutellospora. Geographic abbreviations: AZ, Arizona; CA, California; ENG, England; FL, Florida; HE, Hessen; OR, Oregon; QC, Quebec; RJ, Rio de Janeiro; SCT, Scotland; SU, Suomi; SZ, Szczecin; TA, Tres Arrovos; WV, West Virginia. Public sequence database accession numbers are shown in parentheses; consensus A, X86687 + Y17651 + AJ132666 + AJ276088; consensus B, AJ301860-63 + AJ276076 + Y17644; consensus C, AJ276077 + AJ276078 + Y17649+Y17650; consensus D, AF038590 + U31997; consensus E, AJ871274 + AJ871275; consensus F, AB041344 + AB041345; consensus G, AJ871270–AJ871272; consensus H, AJ306443 + AJ306445 + AJ306446; consensus I, AJ276092 + AJ276093; consensus J, AJ619940–AJ619943; consensus K, AJ619944–AJ619947; consensus L, AJ619948–AJ619951; consensus M, AJ619952–AJ619955.
New Phytologist (2009) 182: 495–506 © The Authors (2009)www.newphytologist.org Journal compilation © New Phytologist (2009)
Research500
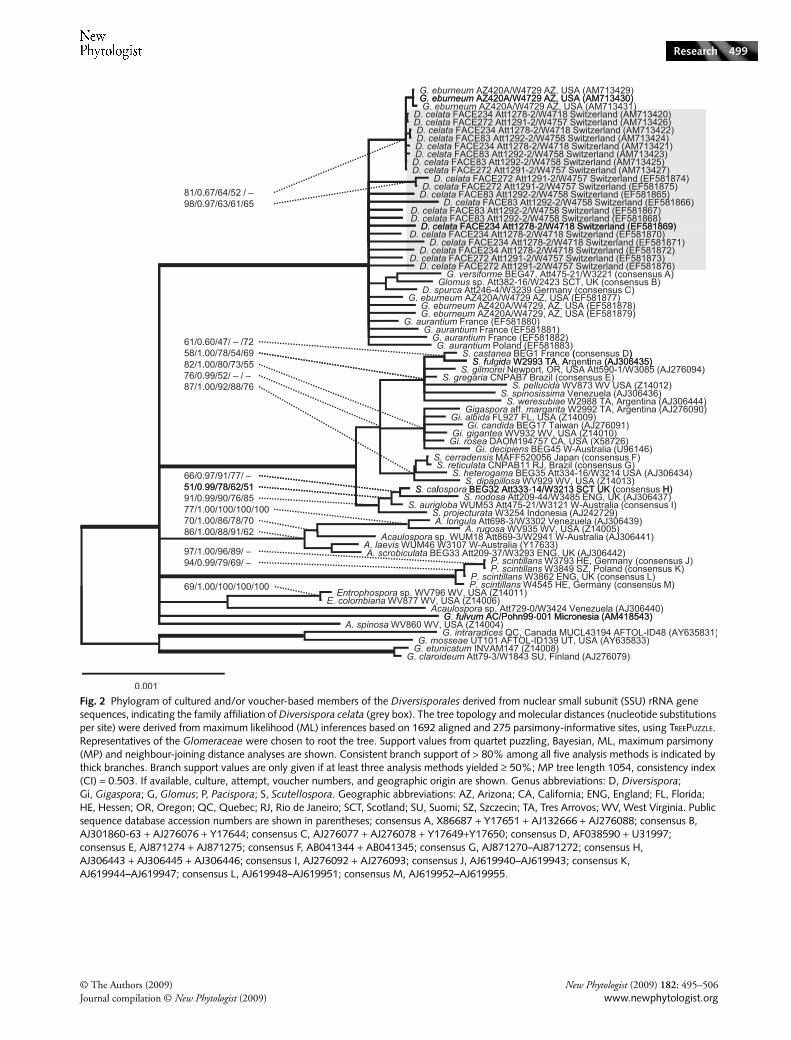
Fig. 3 Phylogram derived from nuclear large subunit (LSU) rRNA gene sequences, indicating the family affiliation of Diversispora celata (grey box) with respect to cultured and/or voucher-based members of the Diversisporales. The tree topology and molecular distances were derived from maximum likelihood (ML) inferences based on 652 aligned and 274 parsimony-informative sites, using TREEPUZZLE. Representatives of the Glomeraceae were chosen to root the tree. Maximum parsimony (MP) tree length 967, consistency index (CI) = 0.504. Annotations of support values, genus abbreviations and sequence accession numbers are as in Fig. 2. Geographic abbreviations: AZ, Arizona; IA, Iowa; ENG, England; FL, Florida; NH, New Hampshire; QC, Quebec; SCT, Scotland; SUM, Sumatra; UT, Utah; WB, West Bohemia.

© The Authors (2009) New Phytologist (2009) 182: 495–506Journal compilation © New Phytologist (2009) www.newphytologist.org
Research 501
sequence conservation precludes resolution at the specieslevel. The partial LSU rDNA sequences appear to resolve thespecies level (Fig. 3), although this may only be because of thepresently poorer taxonomic coverage for this rRNA genepartition. Together, the SSU and LSU rRNA gene treesshowed D. celata to be closely related to Diversispora spurca,Glomus versiforme, G. aurantium and G. eburneum. The phylo-
genetic signal from the ITS rDNA region (ITS1, 5.8S rRNAgene, and ITS2) resolves D. celata as most closely related to aphylotype known from environmental studies in Africa(Namibia) and North America (Arizona, USA) (Fig. 4).Diversispora celata, G. eburneum and the environmentalphylotypes from Namibia and Arizona fall within a sisterclade to that formed by G. versiforme, G. aurantium, and an
Fig. 4 Phylogram derived from sequences of the nuclear ribosomal internal transcribed spacer (ITS) region, indicating the genus affiliation of Diversispora celata (grey box) and the distribution of its close relatives in the Diversisporaceae over the globe. Cultured and/or voucher-based members of the Diversisporaceae are shown in bold. The tree topology and molecular distances (nucleotide substitutions per site) were derived from maximum likelihood (ML) inferences based on 417 aligned and 206 parsimony-informative sites, using TREEPUZZLE. Representatives of the Gigasporaceae were chosen to root the tree (distance shortened by 50% for better display). Maximum parsimony (MP) tree length 581, consistency index (CI) = 0.410. Annotations of support values, genus abbreviations and sequence accession numbers are as in Fig. 2. Geographic abbreviations: AZ, Arizona; FL, Florida; TH, Thuringia. Consensus A: AF185690 + AF185691 + AF185695. Note: for environmental sequences and unidentified cultures the most likely genus affiliation is given and not the taxonomic affiliations mentioned in public databases.

New Phytologist (2009) 182: 495–506 © The Authors (2009)www.newphytologist.org Journal compilation © New Phytologist (2009)
Research502
‘uncultured G. versiforme’ from Europe (Turingia, Germany);the latter possibly representing another undescribed species.The recently reported ‘Glomus fulvum clade’ (Redecker et al.,2007), comprising Glomus fulvum, G. pulvinatum and Glomusmegalocarpum, forms a more distantly related lineage within theDiversisporaceae (Fig. 4), probably at the level of genus assuggested by Redecker et al. (2007). New sequences of allthree rDNA regions for G. eburneum showed it to be themost closely related named species to D. celata. We cannotmake such detailed comparison with the generic type speciesD. spurca, as its phylogenetic placement can presently only beinferred from SSU rDNA sequences.
Although not as reliable as phylogenetic analyses, pairwisedistances are an alternative approach to the evaluation of thedistinctiveness of species. The ranges of sequence identity, ascalculated for intra- and interspecific comparisons of theNS31-AM1 SSU rDNA fragment, ITS, and LR1-FLR2 LSUrDNA region (Supporting Information Table S1), andevidence for shared rDNA sequence variants among closelyrelated species within the genus Diversispora are reported asSupporting Information Notes S3.
Geographical distribution of Diversisporaceae phylotypes
Similar environmental ITS and SSU rDNA sequences frompublic databases with different geographical origins indicate aworldwide distribution of species in the Diversisporaceae.Figure 5 shows the phylogenetic relationship of D. celata toenvironmental SSU rDNA sequences from Australia, Botswana,the USA, Estonia, Ethiopia, Germany, Italy, Micronesia, theNetherlands, Panama, Portugal, South Korea, Sweden, andthe UK. The sequences of D. celata cluster together withenvironmental sequences from Africa, America, Asia and Europe,and with those of the generic type species D. spurca, G. eburneum(AZ420A), Otospora bareai, and G. versiforme (BEG47) (Fig. 5).Members of the Diversisporaceae occur in both natural anddisturbed habitats such as arable land and polluted sites. The‘G. fulvum clade’ (Redecker et al., 2007) comprises environmentalsequences from Portugal and South Korea, which demonstratesthat its members are not restricted to the tropics.
In the ITS tree (Fig. 4), AMF from two INVAM cultures,AZ237 from the USA and NB101 from Namibia, are mostclosely related to the Swiss D. celata. The most closely relateddescribed species is from the USA (G. eburneum AZ420A).Glomus aurantium from maritime dunes in Israel, G. versiforme(BEG47) from potted plants in an Oregon conservatory, andmany environmental sequences from Thuringia in Germanyare closely related but split into several additional clusters.
In contrast to the SSU and ITS region rDNA sequences,environmental LSU rDNA sequences are underrepresented inthe public databases and do not allow interpretation of thewider geographical distribution of Diversisporaceae. There isonly one group of 11 short sequences (AF396808–AF396810
and AF396819–AF396826, apparently wrongly ascribed toGigaspora) in the public databases. These were obtained bynested PCR from field-collected root material sampled within17 km of the type locality for D. celata with purportedlyGigaspora-specific primers (Jansa et al., 2003). Two of thosesequences (AF396819 and AF396820) group with G. versiformeBEG47, but the remaining sequences cannot reliably beaffiliated to any taxon, perhaps because of their shortness anda lack of sufficiently closely related phylotypes. One furtherenvironmental short LSU rDNA sequence (AB206237, fromMount Fuji in Japan; Wu et al., 2007) possibly comes fromanother member of the Diversisporaceae (not shown).
Discussion
This discovery, isolation, and comprehensive morphologicaland molecular characterization of D. celata, as a species newto science, originated from a molecular ecological initiative todetect AMF lineages directly in root samples from the field(Gamper & Leuchtmann, 2007) and to establish AMFsingle spore cultures. Four pure cultures producing sporesmorphologically indistinguishable from each other wereestablished (see Supporting Information Notes S1 and S2 fordetailed morphological comparisons). Close correspondenceamong partial LSU rRNA gene sequences from the distinctisolates (Gamper & Leuchtmann, 2007) was linked withmorphology to preliminarily characterize the organism.
For a more detailed analysis, additional LSU rDNA sequences,along with SSU and ITS rDNA region sequences, were generated,including some from closely related fungi (Supporting Informa-tion Table S1). The phylogenetic analyses (Figs 2–5) resolvedD. celata in a monophyletic clade that also included the generictype species D. spurca (Figs 2, 3). This ‘Diversispora clade’ is clearlydistinct from a sister clade comprising G. fulvum, G. pulvinatumand G. megalocarpum (Fig. 4) with divergent SSU sequencesignatures (Walker & Schüßler, 2004; Redecker et al., 2007).
Recently, our placement of D. celata within the genusDiversispora had to be reinterpreted when O. bareai wasdescribed (Palenzuela et al., 2008). Two relatively shortnuclear SSU rDNA sequences ascribed to Otospora clusterwithin the ‘Diversispora clade’ (Fig. 5). This clade was interpretedhere and in previous molecular studies as the genus level andtherefore Otospora could render Diversispora paraphyletic.However, the sequences of Otospora completely match thosesignatures formally described in the protologue of Diversispora(Walker & Schüßler, 2004) as being specific for the genus,which challenges the erection of the new genus Otospora.Moreover, the spore morphology of O. bareai is acaulosporoid,unlike the glomoid spores of all other AMF within theDiversisporaceae. Such morphological diversity is not impossible,as both acaulosporoid and glomoid spores were demonstratedto form in the genus Ambispora (Walker et al., 2007). However,as a parsimonious explanation it is also possible that the shortsequences reported as those of O. bareai could have been

© The Authors (2009) New Phytologist (2009) 182: 495–506Journal compilation © New Phytologist (2009) www.newphytologist.org
Research 503
derived from xenobiotic DNA (Schüßler et al., 2003) of aDiversispora sp. present in the mixed trap pot culture. Furtherverification of the systematic position of Otospora is needed,as it cannot, in our opinion, yet be reliably placed in aphylogenetic framework.
rDNA-based species identification of D. celata
The SSU rDNA analyses placed the new AMF species in thegenus Diversispora, defined by the clade containing D. spurca.From our analyses, the LSU and ITS region rDNA sequences
Fig. 5 Phylogram derived from nuclear small subunit (SSU) rRNA gene sequences, indicating the phylogenetic relationship of Diversispora celata (grey box) with environmental sequences from public databases and their geographic origin. Cultured and/or voucher-based members of the Diversisporaceae are shown in bold. The tree topology and molecular distances (nucleotide substitutions per site) were derived from maximum likelihood inferences based on 1743 aligned and 134 parsimony-informative sites, using TREEPUZZLE. Representatives of the Gigasporaceae were chosen to root the tree. Maximum parsimony (MP) tree length 329, consistency index (CI) = 0.763. Annotations of support values, genus abbreviations and sequence accession numbers are as in Fig. 2. Geographic abbreviations: AZ, Arizona; CA, California; SCT, Scotland; TA, Tres Arrovos. Consensus A, AJ301860-63 + AJ276076 + Y17644; consensus B, X86687 + Y17651 + AJ132666 + AJ276088; consensus C, AJ276077 + AJ276078 + Y17649 + Y17650; consensus D, AF038590 + U31997; concatenated E: AM400229 + AM905318 (reported as Otospora bareai, but see Discussion). Note: for environmental sequences and unidentified cultures the most likely genus affiliation is given, if possible, and not the taxonomic affiliations mentioned in public databases.

New Phytologist (2009) 182: 495–506 © The Authors (2009)www.newphytologist.org Journal compilation © New Phytologist (2009)
Research504
probably provide resolution down to the species level. MP,NJ, tree puzzling, Bayesian, and ML analyses yielded similarSSU and LSU tree topologies and branch support values, exceptfor some terminal branches and the usual higher posteriorprobability values from Bayesian analyses, which, for thehighly variable ITS region, showed some deviations in treebranching. However, problems with Bayesian analyses ofrather short, rapidly evolving sequences are known and makethese topologies less reliable than those from ML inferences.
Presently, molecular characterization of taxa in Glomeromycotais best achieved using the rDNA region. However, as forAmbispora (Walker et al., 2007), the SSU rDNA does notresolve species in Diversispora. It is important to note thatmost previous molecular ecological studies were concernedwith a taxonomic rank higher than that of species, becauseSSU rDNA was analysed. For species-level identification,rapidly evolving rDNA regions such as the ITS region willinform future molecular ecological studies, once the phylo-genetic ‘backbone’ is sufficiently robust. Therefore, althoughpronounced intraspecific variation may impose some limita-tions, for species characterization it is desirable also toreport the more variable ITS region and partial LSU rDNAsequences in future species descriptions. Such data wouldbe essential for comparison of the findings of molecularecological studies. In future, short, distinct nucleotide sequences,representing taxon-specific DNA signatures or ‘microcodes’(Summerbell et al., 2005; Landis & Gargas, 2007), mayhelp in the endeavour to ascribe species to environmentalsequence types.
Living cultures for species descriptions and future studies
Diversispora celata was characterized by examining isolatesobtained from independent soil-trap cultures, allowing analysis,to some degree, of intraspecific variation. Some previousAMF species descriptions have examined variation withinsingle spore isolates (Walker & Vestberg, 1998; Declercket al., 2000) and among more than one single spore isolate(Walker et al., 2007). Similar characterization of new specieswould be facilitated by the existence of living pure cultureswhich also would permit future investigations onmorphological, developmental and physiological variation,and also allow predictions of responses to environmentalinfluences (van der Heijden & Scheublin, 2007). The presentspecies description is comprehensive and combines morphologyand molecular phylogeny, but it is just the first step towardsunderstanding the ecological traits of D. celata. It is desirablethat the ‘whole fungus’ (Kendrick, 1979; Walker, 1985) beconsidered when describing AMF to achieve biologicallyand ecologically meaningful species characterizations. Suchholistic species descriptions will also consider other aspects ofthe biology of AMF, such as anastomosis compatibility andsymbiotic functional traits.
Molecular ecology and global distribution of Diversisporaceae
Molecular investigations are indispensable for the study of theecology of AMF, with their cryptic lifestyles. The majority ofAMF species will remain uncultured for the near future,and therefore molecular detection and identification areparticularly important. Uncovering AMF molecular diversityin geographically distant locations will provide importantinformation about global taxon distribution.
The SSU and ITS rDNA sequences of D. celata, whencompared with environmental sequences from publicdatabases, confirmed the molecular distinctiveness of the newspecies and revealed a global distribution of close relatives. Forthe Diversisporaceae, the geographical annotations of databaseentries suggest worldwide occurrence (Fig. 5) and evidencefor undescribed species (Fig. 4). The two undescribed AMFin INVAM cultures AZ237 (USA) and NB101 (Namibia)were closest to D. celata and clusters of environmentalsequences from Germany may represent further Diversisporaspecies. In contrast to the apparently better taxonomic resolu-tion of the ITS rDNA data set, poor geographical coverageimpaired inferences about the global distribution of theDiversisporaceae, because most studies employing the ITSregion as a marker were for relatively restricted areas of centralEurope. For LSU rDNA the low number of publicly availablesequences prevented analysis of the geographical distribution.The only environmental Diversisporaceae sequences originatefrom near the type locality of D. celata in Switzerland andfrom Japan.
Evidence from several molecular ecological studies may beseverely biased by the PCR primer combinations used. Thismay be exemplified by the supposedly AMF-specific LSUrDNA primer pair FLR3/FLR4 (Gollotte et al., 2004), whichshows poor binding site matching for members of theDiversisporaceae. In particular, the reverse primer FLR4 showsseveral mismatches at its 3′ end to all known Diversisporaceaesequences, which will therefore be overlooked in studies inwhich these primers are used. Results from studies usingnonphylogenetically inclusive primers must therefore beinterpreted carefully. For example, D. celata was commonlyfound in Trifolium repens root samples from Switzerlandwhen species-specific PCR primers were used (Gamper &Leuchtmann, 2007) but would not have been detected ifFLR4 had been used. Similar problems exist for other AMFlineages when using the SSU rDNA primer AM1 (Helgasonet al., 1998).
Future species detection and descriptions in Glomeromycota
Our preliminary analysis of the geographical distribution ofmembers of the Diversisporaceae underlines the importanceof increasing the number of well-annotated reference

© The Authors (2009) New Phytologist (2009) 182: 495–506Journal compilation © New Phytologist (2009) www.newphytologist.org
Research 505
sequences from identified AMF for future molecular ecologicaland biogeographical examination. Where doubt exists concerningspecies determination, database entries should indicate this.A change of policy by the public sequence databases(Bidartondo et al., 2008) would allow improvements in thetaxonomic annotation of older sequence accessions.
To enable natural classification (Woese et al., 1990), taxonomyshould ideally be informed by phylogenetic analyses, whichwould also help to avoid misclassifications in the wronggenera. The Botanical Code, however, only requires thedesignation of a dead type specimen along with a validlypublished description, although living cultures are encouragedin recommendation 8B.1, which reads ‘Whenever practicablea living culture should be prepared from the holotype materialof the name of a newly described taxon of fungi or algae anddeposited in at least two institutional culture or geneticresource collections’. This recommendation, although importantfor follow-up studies, is not always followed. Consequentlymany named species exist without cultures available to allowfurther study of their biology, genetics and ecology. If livingcultures cannot be provided (many AMF appear to berecalcitrant), inclusion of comprehensive molecular evidenceas part of species descriptions is essential for future revisionswithin a natural classification system, or for comparison withmolecular ecological data.
This work continues the trend to integrate morphologicaland molecular evidence for AMF characterization and speciesdescriptions, and proposes that, as far as possible, a ‘wholefungus’ approach should be encouraged by use of type isolatesand their descendant cultures made generally available to thescientific community for future investigations. Particular careshould be taken to ensure that public database entries areaccurately named, where possible, and clearly labelled as‘unidentified’ if not well characterized.
Acknowledgements
We are grateful to J. Blaszkowski, who generously providedspores of G. aurantium, and J. M. Trappe for the Latin diagnosis.HG received funding through research grants from the ETH,the BBSRC and the NIG during the time this study wascarried out. AS received funding from the DFG, and workcarried out in Germany was also funded by the DFG.
References
Bidartondo MI, Bruns TB, Blackwell M, Edwards I, Taylor AFS, Horton T, Zhang N, Kõljalg U, May G, Kuyper TW. 2008. Preserving accuracy in GenBank. Science 319: 1616.
Declerck S, Cranenbrouck S, Dalpé Y, Seguin S, Grandmougin Ferjani A, Fontaine J, Sancholle M. 2000. Glomus proliferum sp. nov.: a description based on morphological, biochemical, molecular and monoxenic cultivation data. Mycologia 92: 1178–1187.
Gamper H, Leuchtmann A. 2007. Taxon-specific PCR primers to detect two inconspicuous arbuscular mycorrhizal fungi from temperate agricultural grassland. Mycorrhiza 17: 145–152.
Gollotte A, van Tuinen D, Atkinson D. 2004. Diversity of arbuscular mycorrhizal fungi colonising roots of the grass species Agrostis capillaris and Lolium perenne in a field experiment. Mycorrhiza 14: 111–117.
Guindon S, Gascuel O. 2003. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Systematic Biology 52: 696–704.
Hart MM, Reader RJ. 2002. Taxonomic basis for variation in the colonization strategy of arbuscular mycorrhizal fungi. New Phytologist 153: 335–344.
van der Heijden MGA, Scheublin TR. 2007. Functional traits in mycorrhizal ecology: their use for predicting the impact of arbuscular mycorrhizal fungal communities on plant growth and ecosystem functioning. New Phytologist 174: 244–250.
Helgason T, Daniell TJ, Husband R, Fitter AH, Young JPW. 1998. Ploughing up the wood-wide web? Nature 394: 431–443.
Jansa J, Mozafar A, Kuhn G, Anken T, Ruh R, Sanders IR, Frossard E. 2003. Soil tillage affects the community structure of mycorrhizal fungi in maize roots. Ecological Applications 13: 1164–1176.
Kendrick B. 1979. Introduction. In: Kendrick B, ed. The whole fungus, Vol. 1. Ottawa, Canada: National Museum of Natural Sciences, Canada & The Kananaskis Foundation, 11–15.
Kottke I, Haug I, Setaro S, Suárez JP, Weiß M, Preußing M, Nebel M, Oberwinkler F. 2008. Guilds of mycorrhizal fungi and their relation to trees, ericads, orchids and liverworts in a neotropical mountain rain forest. Basic and Applied Ecology 9: 13–23.
Landis FC, Gargas A. 2007. Using ITS2 secondary structure to create species-specific oligonucleotide probes for fungi. Mycologia 99: 681–692.
Maherali H, Klironomos JN. 2007. Influence of phylogeny on fungal community assembly and ecosystem functioning. Science 316: 1746–1748.
Munkvold L, Kjoller R, Vestberg M, Rosendahl S, Jakobsen I. 2004. High functional diversity within species of arbuscular mycorrhizal fungi. New Phytologist 164: 357–364.
Nilsson RH, Kristiansson E, Ryberg M, Hallenberg N, Larsson KH. 2008. Intraspecific ITS variability in the kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evolutionary Bioinformatics 4: 193–201.
Öpik M, Moora M, Liira J, Zobel M. 2006. Composition of root-colonizing arbuscular mycorrhizal fungal communities in different ecosystems around the globe. Journal of Ecology 94: 778–790.
Öpik M, Moora M, Zobel M, Saks I, Wheatley R, Wright F, Daniell T. 2008. High diversity of arbuscular mycorrhizal fungi in a boreal herb-rich coniferous forest. New Phytologist 179: 867–876.
Palenzuela J, Ferrol N, Boller T, Azcon-Aguilar C, Oehl F. 2008. Otospora bareai, a new fungal species in the Glomeromycetes from a dolomitic shrub land in Sierra de Baza National Park (Granada, Spain). Mycologia 100: 296–305.
Pirozynski KA, Malloch DW. 1975. Origin of land plants – a matter of mycotropism. Biosystems 6: 153–164.
Posada D, Crandall KA. 1998. Modeltest: testing the model of DNA substitution. Bioinformatics 14: 817–818.
Redecker D, Hijri I, Wiemken A. 2003. Molecular identification of arbuscular mycorrhizal fungi in roots: perspectives and problems. Folia Geobotanica 38: 113–124.
Redecker D, Kodner R, Graham LE. 2000. Glomalean fungi from the Ordovician. Science 289: 1920–1921.
Redecker D, Raab P, Oehl F, Camacho FJ, Courtecuisse R. 2007. A novel clade of sporocarp-forming species of glomeromycotan fungi in the Diversisporales lineage. Mycological Progress 6: 35–44.
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572–1574.
Schmidt HA, Strimmer K, Vingron M, von Haeseler A. 2002. TREE-PUZZLE: maximum likelihood phylogenetic analysis using quartets and parallel computing. Bioinformatics 18: 502–504.

New Phytologist (2009) 182: 495–506 © The Authors (2009)www.newphytologist.org Journal compilation © New Phytologist (2009)
Research506
Schüßler A, Gehrig H, Schwarzott D, Walker C. 2001a. Analysis of partial Glomales SSU rRNA gene sequences: implications for primer design and phylogeny. Mycological Research 105: 5–15.
Schüßler A, Schwarzott D, Walker C. 2001b. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycological Research 105: 1413–1421.
Schüßler A, Schwarzott D, Walker C. 2003. Glomeromycota rRNA genes – the diversity of myths? Mycorrhiza 13: 233–236.
Schwarzott D, Schüßler A. 2001. A simple and reliable method for SSU rRNA gene DNA extraction, amplification, and cloning from single AM fungal spores. Mycorrhiza 10: 203–207.
Schwarzott D, Walker C, Schüßler A. 2001. Glomus, the largest genus of the arbuscular mycorrhizal fungi (Glomales), is nonmonophyletic. Molecular Phylogenetics and Evolution 21: 190–197.
Sieverding E, Oehl F. 2006. Revision of Entrophospora and description of Kuklospora and Intraspora, two new genera in the arbuscular mycorrhizal Glomeromycetes. Journal of Applied Botany and Food Quality-Angewandte Botanik 80: 69–81.
Summerbell RC, Levesque CA, Seifert KA, Bovers M, Fell JW, Diaz MR, Boekhout T, de Hoog GS, Stapers J, Crous PW. 2005. Microcoding: the second step in DNA barcoding. Philosophical Transactions of the Royal Society B 360: 1897–1903.
Swofford DL. 2003. PAUP: phylogenetic analysis using parsimony and other methods, version 4.0.0b10 beta. Sunderland, MA, USA: Sinauer Associates.
Tilman D, Reich PB, Knops J, Wedin D, Mielke T, Lehman C. 2001. Diversity and productivity in a long-term grassland experiment. Science 294: 843–845.
Vandenkoornhuyse P, Mahé S, Ineson P, Staddon P, Ostle N, Cliquet JB, Francez AJ, Fitter AH, Young JPW. 2007. Active root-inhabiting microbes identified by rapid incorporation of plant-derived carbon into RNA. Proceedings of the National Academy of Sciences, USA 104: 16970–16975.
Vilgalys R. 2003. Taxonomic misidentifications in public DNA databases. New Phytologist 160: 4–5.
Walker C. 1985. Taxonomy of the Endogonaceae. In: Trappe JM, Molina R, Berch S, eds. Proceedings of the Sixth North American Conference on Mycorrhizae. Corvallis, OR, USA: Forestry Sciences Laboratory, Oregon State University, 193–198.
Walker C, Blaszkowski J, Schwarzott D, Schüßler A. 2004a. Gerdemannia gen. nov., a genus separated from Glomus, and Gerdemanniaceae fam. nov., a new family in the Glomeromycota. Mycological Research 108: 707–718.
Walker C, Gianinazzi-Pearson V, Marion-Espinasse H. 1993. Scutellospora castanea, a newly described arbuscular mycorrhizal fungus. Cryptogamie, Mycologie 14: 279–286.
Walker C, Schüßler A. 2004. Nomenclatural clarifications and new taxa in the Glomeromycota. Mycological Research 108: 979–982
Walker C, Vestberg M, Demircik F, Stockinger H, Saito M, Sawaki H, Nishmuar I, Schüßler A. 2007. Molecular phylogeny and new taxa in the Archaeosporales (Glomeromycota): Ambispora fennica gen. sp. nov., Ambisporaceae fam. nov., and emendation of Archaeospora and Archaeosporaceae. Mycological Research 111: 137–153.
Walker C, Vestberg M, Schüßler A. 2004b. Nomenclatural clarifications in Glomeromycota. Mycological Research 111: 253–255.
Walker C, Vestberg V. 1998. Synonymy amongst the arbuscular mycorrhizal fungi: Glomus claroideum, G. maculosum, G. multisubstensum, and G. fistulosum. Annals of Botany 82: 601–624.
Woese C, Kandler O, Wheelis M. 1990. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proceedings of the National Academy of Sciences, USA 87: 4576–4579.
Wu B, Hogetsu T, Isobe K, Ishii R. 2007. Community structure of arbuscular mycorrhizal fungi in a primary successional volcanic desert on the southeast slope of Mount Fuji. Mycorrhiza 17: 495–506.
Zak DR, Holmes WE, White DC, Peacock AD, Tilman D. 2003. Plant diversity, soil microbial communities, and ecosystem function: are there any links? Ecology 84: 2042–2050.
Supporting Information
Additional supporting information may be found in theonline version of this article.
Table S1 Database accession numbers of sequences of Diver-sispora celata and its close relatives Glomus aurantium andGlomus eburneum
Methods S1 Soil and climatic conditions at the site of originand establishment of trap and single spore cultures.
Methods S2 Additionally analysed living reference cultures ofDiversispora celata, Glomus aurantium and Glomus eburneum.
Methods S3 Details of DNA extraction and PCR amplification.
Methods S4 Details of sequence alignments and phylogeneticanalyses.
Notes S1 Spore and hyphal morphology of Diversisporacelata, as recorded for multiple isolates.
Notes S2 Morphological determination of Diversispora celatasp. nov. and comparison with arbuscular mycorrhizal fungi ofsimilar spore morphology.
Notes S3 Pairwise nucleotide sequence identities and sharedrDNA sequence variants within and among members ofDiversispora.
Notes S4 References to Methods S1–S4, Notes S1–S3 andTable S1.
Please note: Wiley-Blackwell are not responsible for the contentor functionality of any supporting information supplied bythe authors. Any queries (other than missing material) shouldbe directed to the New Phytologist Central Office.

Supporting Information 1
2
3
4
5
6
7
8
Table S1 Database accession numbers of sequences of Diversispora celata and its close
relatives Glomus aurantium and G. eburneum. Species affiliation of the different nuclear
ribosomal DNA regions (SSU: small subunit, ITS: internal transcribed spacer, LSU: large
subunit) are indicated. New sequences are typed in bold and the approximate relative
coverage of the rDNA regions is indicated with black bars.
Culture [attempt, voucher]
Accession number SSU rDNA ITS rDNA region LSU rDNA
Diversispora celata AM713423-5# FACE 83 EF581865-8¶ [Att 1292-2, W4758] AY639225-32§ FACE 234 AM713420-2# [Att 1278-2, W4718, EF581869-72¶ W4719] AM713402-4* AY639233-5§
AY639306§
AM713417-9* FACE 272 AM713426-7# [Att 1291-2, W4757] EF581873-6¶ AY639236-41§ FACE 410 [Att 1500-2, W5306, W5307] DQ350448-53§
Glomus eburneum AM713429-31#
AZ420A [W4729] EF581877-9¶ EF0678886-8§ AM713405-
12*
AM713413* AM713414-6* Glomus aurantium EF581880-3¶ [W4728] EF581860-4§
9 10
11
12
13
* PCR primer SSU-Glom1 (Renker et al., 2003) and LSU-Glom1b (Walker et al., 2007) used
¶ PCR primer NS31 (Simon et al., 1992) and AM1 (Helgason et al., 1998) used
§ PCR primer LR1 (van Tuinen et al., 1998) and FLR2 (Trouvelot et al., 1999) used
# PCR primer pair GeoA1/ART4 and nested pair GeoA2/Geo11b (Schwarzott & Schüßler, 2001) used
1

Supporting Information 1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
Methods S1 Soil and climatic conditions at the site of origin and
establishment of trap and single spore cultures
The climatic conditions at the field site in Eschikon (Switzerland) are characterised by
wet winters (mean annual precipitation 1108 mm yr-1) and a mean annual temperature
of 8.6°C. The soil is a fertile, eutric cambisol (clay loam according to the US
classification, 28% clay, 33% silt; 36% sand, 2.9-5.1% organic material) with pH
(water) values between 6.5 and 7.6 and available phosphorus and potassium (after
Dirks & Scheffer, 1930) ranging from 1.2 to 6.0 and from 18 to 49 mg kg-1 soil,
respectively. The monoculture plots had been fertilised at non-limiting levels to
achieve maximum forage production and kept weed free manually. Fusarium and
Sclerotinia trifoliorum disease was controlled by a springtime aqueous soil drench (5
g m-2 of 1 g l-1) of Benlate (DuPont; Wilmington, DE, USA).
For trap culturing approx. 20 ml of top soil were mixed with a
potting substrate of pasteurised field soil, expanded attapulgite clay (Oil Dry Chem-
Sorb WR 24/18, Brenntag Mediterranée Export, Vitrolles Cedex, France), and silica
sand (0.7–1.2 mm) at 1:2:2 (v/v/v) in 750 ml pots. The trap cultures were kept in a
greenhouse under a 16/8 h day/night photoperiod, a minimum illumination of 400 mol
photons m–2 s–1, an approx. 25/20°C day/night temperature regime, and 40–50%
relative humidity. Tensiometer-controlled drip-irrigation (Tropf-Blumat®, Blumat,
Austria) maintained substrate humidity at 50-60% of water holding capacity, and
plants were fertilised monthly with 20 ml of Hoagland nutrient solution containing
1% of the original P concentration (Sylvia & Hubbell, 1986).
1

2
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
After five months, spores were extracted from the trap culture
substrate by wet sieving and decanting (Gerdemann, 1963) and spores with healthy
appearance were selected under a dissecting microscope. Single spore culture attempts
were established by placing one spore on a root of a Plantago lanceolata seedling,
which was planted into a 750 ml pot with the same substrate as described above and
grown, unfertilised, for approximately four months. Three such attempts were
successful. One single spore isolate, designated FACE 83 (Att 1292-1), was from a
trap culture with two hosts, Glycine max and Allium porrum cv Dubouchet. Two
isolates, FACE 234 (Att 1278-1) and FACE 272 (Att 1291-1), were derived from trap
cultures with both Leontodon autumnalis and Dipsacus fullonum as host plants, as
was a fourth culture (FACE 410, Att 1500-1) resulting from a failed attempt to
establish a single spore culture of a Scutellospora sp. Most likely this spore was
occupied by the glomoid species (Koske, 1984). This isolate (FACE 410) can,
therefore, not be termed an isolate, since the Scutellospora spore may have contained
several spores.
The four AMF parent cultures were subcultured from air-dried pot-
substrate for further study, first on P. lanceolata and Zea mays cv Corso (substrate as
above), and subsequently on P. lanceolata in a 1:1 (v/v) autoclaved expanded
attapulgite clay (Terragreen, TDS043 GRAN, Oil Dri, UK, Ltd) and quartz sand
(WBB Minerals Brookside Hall, Sandbach, UK), amended with bone meal, and
fertilised weekly with P-free Rorison’s nutrient solution (Hewitt, 1966).

Supporting Information 1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
Methods S2 Additionally analysed living reference cultures of
D. celata, G. aurantium and G. eburneum
Besides the TYPUS E (Isotype MSB), represented by the single spore isolate
FACE 234, three further isolates of D. celata FACE 272, FACE 83, and FACE 410
were studied morphologically: Isolate FACE 272: loc. cit. from a single spore pot
culture with P. lanceolata established from a soil trap pot culture, 30 Oct. 2005, H.
Gamper (Att 1291-2, W4757) (E); loc. cit. 7 Jan. 2006, H Gamper (Att 1291-2,
W4773) (E); loc. cit. from a subculture of Att 1291-2 with P. lanceolata, 15 Feb.
2008, H. Gamper (Att 1291-5, W5399) (E). Isolate FACE 83: loc. cit. from a single
spore pot culture with P. lanceolata established from a soil trap pot culture, 30 Oct.
2005, H. Gamper (Att 1292-2, W4758) (E); loc. cit. 7 Jan. 2006, H. Gamper
(Att 1292-2, W4774) (E); loc. cit. from a subculture of Att 1292-2 with P. lanceolata,
15 Feb. 2008, H. Gamper (Att 1292-4, W5397) (E). Culture FACE 410: loc. cit. from
a pot culture with P. lanceolata established from a single presumably dead spore of
Scutellospora sp., 30 Oct. 2005 H. Gamper (Att 1500-1, W5306); loc. cit. 15 Jul.
2006, H. Gamper (Att 1500-2, W5307) (E); loc. cit. from a subculture of Att 1500-2
with P. lanceolata, 15 Feb. 2008, H. Gamper (Att 1500-4, W5400) (E).
Fresh pot material from an ex-type culture descendent pot culture of
G. eburneum (AZ420A) was directly obtained from the International Culture
Collection of Arbuscular Mycorrhizal fungi (INVAM, http://invam.caf.wvu.edu/), Att
1290-5 voucher W4729. A sample of fresh spores of G. aurantium from a trap culture
was obtained from Dr. Janusz Blaszkowski, Att 1296-0, voucher W4728.
1

Supporting Information 1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
Methods S3 Details of DNA extraction and PCR amplification
Genomic DNA of the four Swiss pure AMF cultures was prepared by crushing individual
spores in 5 μl of PCR-grade water, with a flame-sealed Pasteur pipette, adding 5 µl of 20%
(w/v) Chelex-100 resin (Bio-Rad, Laboratories, Hercules, CA, USA) incubating the
suspension at 95°C for 3 min, and cooling it on ice. To prepare more concentrated DNA
extracts multiple spores were collected with forceps in 7 μl PCR grade water in the lid of a
0.2 ml PCR tube, crushed, vortexed, and boiled at 95°C for 5 min. Multi-spore DNA extracts
were prepared for Glomus aurantium from 65 spores and for G. eburneum from 120 spores in
30 μl 20% (w/v) Chelex-100 resin. About 250 spores were used for multi-spore DNA
extracts from all four isolates of the Swiss fungus, which were, however, prepared by using
the DNeasy Plant Mini Kit (Qiagen GmbH, Hilden, Germany), instead of Chelex-100 resin.
Nested PCR amplification from single spore DNA templates with the primer
pairs ITS3/NDL22 (White et al., 1990; van Tuinen et al., 1998b) and LR1/ FLR2 (van
Tuinen et al., 1998a; Trouvelot et al., 1999) was used to obtain a fragment from the 3' end of
the nuclear LSU rRNA genes (Gamper & Leuchtmann, 2007). Partial sequences of the
nuclear SSU rRNA genes were amplified from 0.5 μl of multispore DNA extracts in 25 μl
PCR reactions with the primers NS31 and AM1 (Helgason et al., 1998). The near full-length
nuclear SSU rRNA gene sequences were obtained from multi-spore or single spore DNA
extracts as described in Schwarzott & Schüßler (2001), except that the Phusion proof reading
polymerase (Finnzymes, Espoo, Finland) was used. The ITS region was amplified using the
primers SSU-Glom1 (Renker et al., 2003) and LSU-Glom1b (Walker et al., 2007), using the
thermal cycling conditions described in Walker et al. (2007).
1

Supporting Information 1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
Methods S4 Details of sequence alignments and phylogenetic analyses
The alignment of the SSU rDNA sequences covered the Diversisporales and all near
full-length sequences of the Diversisporaceae available in the public databases. Using
members of Glomus group A and B (Schwarzott et al., 2001) as outgroup, 72
sequences could be analysed. Another dataset was assembled by aligning SSU rDNA
sequences of reference isolates and of environmental sequences with closest BLASTn
hits (33 sequences) to the newly described AMF. Members of the Gigasporaceae
were chosen as outgroup for the SSU as well as the LSU rDNA phylogenetic tree. The
LSU rDNA alignment covered all available 74 Diversisporaceae sequences. The
alignment with 57 sequences of the ITS region (ITS1, 5.8S rDNA, ITS2) comprised
all available Diversisporaceae sequences, including representatives of distinct
phylotypes from published molecular ecological studies. Rooting of the ITS tree with
Gigasporaceae sequences caused an ambiguous alignment of the outgroup taxa at
many sites. The position of the root was, therefore, verified by an analysis of the more
conserved SSU rDNA.
For the phylogenetic analyses the following numbers of aligned sites
could be used: 1692 for the SSU rDNA of Diversisporales, 1743 for the SSU rDNA
of Diversisporaceae, 652 for the LSU rDNA of Diversisporales, and 417 for the ITS
region rDNA of Diversisporaceae. Best-fit models of sequence evolution were
determined in ModelTest (version 3.7, Posada & Crandall, 1998) by hierarchical
likelihood ratio testing. The evaluated parameter settings were used for neighbour
joining (NJ) maximum likelihood distance analyses with random sequence addition,
1

2
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
carried out in PAUP (v.4b10, Swofford, 2003) and for Bayesian phylogenetic
inferences (BI) with MrBayes (v.3.1.2, Ronquist & Huelsenbeck, 2003). For the latter,
four Markov chains were run in parallel over 3 *106 generations (for ITS region
6 *106 generations), ensuring that the runs converged to the same tree topology and
that the standard deviations between the runs fell below 0.01. Fifty percent consensus
trees and posterior probabilities for branch support were calculated from the last 75%
of trees that were sampled every 100th generation. In maximum parsimony (MP)
heuristic tree searches in PAUP the nucleotide sites were classified as unordered
characters of equal weight and gaps treated as missing data. Starting trees were
obtained by random sequence addition and 10 trees were held in memory for 100
replicates of tree-bisection-reconnection branch swapping with the ‘MulTrees’ option
kept in effect. For maximum likelihood analyses in TREE-PUZZLE (v.5.2, Schmidt et
al., 2002) and PHYML (v.2.4.4, Guindon & Gascuel, 2003) the general time
reversible (GTR) model, with gamma distributed heterogeneous rates were used.
Branch support values were calculated for NJ and PHYML from 5000 and for MP
from 200 non-parametric bootstrap pseudoreplicates by using the same tree search
strategies. Trees were assembled and edited with the programs TREEME
(http://science.do-mix.de/software.php), TreeView (http://taxonomy.zoology. gla.ac.
uk/rod/treeview.html) and Microsoft PowerPoint.

Supporting Information 1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
Notes S1 Spore and hyphal morphology of D. celata
Spores and fragments of the extraradical mycelia were extracted from fresh potting
substrate by shaking in water and decanting through paired sieves (500 μm and 30 μm
mesh size), collected with forceps, and examined in water, or after storage in 75%
ethanol, through a dissecting microscope. Selected specimens were prepared and
examined microscopically following the procedures outlined in Walker et al. (2007),
and described in Walker et al. (1993).
The spores of D. celata form blastically on a subtending hypha as
chlamydospores, singly and in loose clusters in the soil (Fig. 1a-c). There is no
evidence of sporocarp production, or of spores produced in large, dense clusters,
either in the soil or on the substrate surface. Most spores are globose to subglobose to
ovoid, though a few may be irregular (Fig. 1a-g). The subtending hypha (Fig. 1d-j)
varies in shape and length, sometimes being straight, but often recurved, usually
funnel-shaped (tapered) to some degree, and sealed by a distal septum (Fig. 1h-j)
formed by fusion of the structural spore wall component. The subtending hypha varies
from being thin-walled and parallel-sided, merely continuous with the spore bearing
hypha with a wall thickness less than 1 µm, to being sharply tapering, straight, or
recurved, with a proximal thickness of up to 3 µm, tapering to less than 1 µm where it
joins the mycelium. The length of the subtending hypha ranges from as little as 4 µm
to more than 40 µm, with attached mycelium (from the point where the tapered
subtending hypha equals the thickness of the sporogenous hypha), often being
1

24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
retained to a length of up to 100 µm. Some spores are occluded by a plug formed at
the spore base.
Spore colour (Anonymous, 1969) examined in water (Fig. 1a) was
consistent among five separate sievings, from among four isolates. There were also no
differences detected among spores from fresh pot culture substrate, dried material re-
wetted with water, or 75% EtOH preservative. Spores are sometimes colourless (when
small and presumably immature), but most are ivory (2) to pinkish cream (4),
eventually becoming ochre (9) (in vernacular terms, mature spores are pale yellow to
yellow). After immersion in Melzer’s reagent or PVLG with Melzer’s reagent the
overall spore colour darkens to become orange (Fig. 1g), but there is no clear
dextrinoid or amyloid reaction, and the colour fades within a few days of slide
preparation.
The spores of D. celata are often covered by a layer of fine soil
particles embedded in a matrix, and have three wall components in two groups. The
main structural spore wall (Group 1) comprises two components. Wall component
(wc) 1 is evanescent. At first it is thin (<1 µm) and tightly adherent to wc 2, but it
expands somewhat and then disintegrates over time. Component 2 is coloured,
laminated, and up to 4 µm thick (Fig. 1l, m). Group 2 is a third (internal), very thin
flexible, membranous component present on mature spores (Fig. 1m), similar to that
in D. spurca (Fig. 1n), though it is possible that this is merely a developing layer of
the laminated component or an artefact caused by plasmolysis of the cytoplasm. In
spores mounted in PVLG, the contents, bounded by component 3, may shrink away
due to plasmolysis (Fig. 1k), leaving a space that gives the erroneous impression of an
amorphous wall component such as is found in some Scutellospora and Acaulospora
spp.
2

49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
For size measurements on spores five different collections from two
independent single spore isolates were assessed: Spores of isolate FACE 272, W4757
(30 Oct. 2005) and W4773 (7 Jan. 2006), both from Att 1291-2, were 57-128 × 57-
192 (mean 88 × 89) µm (n=100) S.D. length = 14.9, S.D. width=19.3, and 66-160 ×
61-130 (mean 96 × 90) µm (n=71) S.D. length = 15.1 S.D. width = 13.6 in size,
respectively; specimens of isolate FACE 83, W4758 (30 Oct. 2005) from Att 1292-2
measured 64-112 × 64-110 (mean 88 × 86) µm (n=32) S.D. length = 11.4, S.D. width
=12.7; spores of isolate FACE 234 (2 and 3 Jan. 2008) yielded measurements of 54-
160 × 54-139 (mean 82 × 82) µm, S.D. length = 18.1, S.D. width = 14.7, (W5461, n =
100), and 54-128 × 59-133 (mean 86 × 86) µm, S.D. length = 14.1, S.D. width = 14.9,
(W5462, n = 100), respectively, and two spore collections from the culture FACE 410
gave the following measurements: W5306 (30 Jun. 2002) 58-96 × 54-99 (mean 75 ×
72) µm S.D. length = 8.8 S.D. width = 9.0 (n=33), and W5307 (Jul. 2006) from
Att 1500-2 53-112 × 48-101 (mean 75 × 73) µm S.D. length = 10.1 S.D. width = 10.6
(n=100). The resultant overall measurements for the four reference isolates of
D. celata are 53-160 × 48-192 (mean 84 × 83) µm S.D. length = 15.6 S.D. width =
15.8 (n=536).
Self-anastomosis (Fig. 1o) appears to represent a frequent
phenomenon in this species, as was observed in large coils of extraradical mycelium,
which were recovered when washing growth substrate for spore and root collection.
The hyphal bridging occurs in the manner described for Gigaspora spp. as ‘wound
healing’ (Gerdemann, 1955) and is a characteristic, which is shared with D. spurca
(Fig. 1p).
3

4
72
73
74
75
76
77
78
79
80
81
82
83
Mycorrhizas with P. lanceolata became only weakly coloured in
several attempts of staining using different procedures (Vierheilig et al., 2005). All
intraradical structures, arbuscules, hyphal coils and normal hyphae, remained only
weakly stained. Figures 1q,r show hyphal coils, and very finely branched arbuscules,
after staining with acidified blue Quink™ ink (Walker, 2005). In contrast to
intraradical structures, appressoria and spores stained darkly, also with Trypan blue.
Vesicles were not observed, although this does not imply they do not occur. It is
notable, that successful fungal propagation is difficult to confirm from the poorly
stained mycorrhiza. Colonisation is sparse and patchy and intraradical hyphae are
often only recognisable because of light refracting lipid droplets. In contrast, well-
established cultures soon sporulate abundantly, spores thus being a much better
indicator of successful propagation.

Supporting Information 1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
Notes S2 Morphological determination of Diversispora celata sp. nov.
and comparison with AMF of similar spore morphology
Diversispora celata exemplifies the difficulty of determining species and genera of
glomeromycotan fungi because spores from systematically widely separated taxa may appear
similar or indistinguishable. Consequently mistakes may be made by inadvertently identifying
similar spores of different species as conspecific in ecological studies, leading to biased
conclusions. It may also result in mixed species cultures when living cultures are initiated
from multiple spores. For this reason the present species characterisation was based on
monospecific spore populations from four pure cultures, three of which originated from single
spores.
D. celata, as described in the species diagnosis, is best distinguishable from
all described AMF species by the combination of its small yellow spores and the occurrence
of wound healing or hyphal bridging. In water under the dissecting microscope, the spores
appear superficially similar to small ectocarpic spores of G. mosseae (Gerdemann & Trappe,
1974), because of their pale yellow colour and sometimes funnel-shaped bases (Fig. 1j).
However, D. celata is not known to form sporocarps and the spores are on average much
smaller (mean 84-83 µm) than those of the G. mosseae isotype (voucher W728) [mean 190-
185, N=50]. A thick septum, which occludes spores of D. celata is formed from the entire
laminated wall component, whereas spores of G. mosseae are sealed by a thin curved septum
formed from the inner layers of the laminated wall component (Nicolson & Gerdemann,
1968). Moreover, spores of G. mosseae can be distinguished from those of D. celata, because
they lack an apparent third wall component. The evanescent outermost spore wall component
of G. mosseae is more prominent than that of D. celata before disintegration, and often
disappears completely in the former, whereas fragments usually remain attached to the main
1

2
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
structural component in the latter. The lamination and colour of the main structural spore wall
components are also more obvious in G. mosseae than in D. celata, mainly because of the
larger spore size and thicker walls. In particular, the outermost spore wall component of
G. mosseae usually shows marked red or pink reactivity to Melzer reagent, whereas spores of
D. celata show only transient darkening.
The wall structure of D. celata spores is similar to that of D. spurca
(formerly G. spurcum) (Pfeiffer et al., 1996), having a colourless evanescent outer wall
component to which soil particles may adhere, a laminated structural component, and an inner
flexible wall component, the latter often difficult to see. The outermost spore wall component
of both species disintegrates. Spores of D. celata can be distinguished from those of D. spurca
by their colour (yellow compared with colourless) and their more persistent subtending
hyphae. Paraglomus occultum has similarly small spores, but these are colourless and lack the
innermost wall component. Kennedy et al. (1999) questioned the presence of this innermost
wall component in D. spurca, though it can be seen in their illustration as internal wrinkling
(their Figs. 2-4) and is clearly illustrated in the protologue (Pfeiffer et al., 1996). Examination
of spores from descendents of the ‘type culture’ showed the apparent existence of this
innermost wall component (Fig. 1n), though it might be an artefact of specimen preparation.
As a consistent feature and thus useful distinguishing taxonomic character we use it in our
species description.
Anastomoses of extraradical hyphae in the form of hyphal bridging are
known from D. spurca (Fig. 1p), A. nicolsonii (Walker et al., 1984), and members of the
genus Scutellospora (de Souza & Declerck, 2003). This form of hyphal bridging is different
from the type of anastomoses formed by members of the Glomerales (Voets et al., 2006).
Comparative studies among phylogenetic clades within the Glomeromycota are needed to
establish if different types of anastomosis formation by extraradical hyphae may serve as a
taxonomically informative character.

Supporting Information 1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
Notes S3 Pairwise nucleotide sequence identities and shared rDNA
sequence variants within and among members of Diversispora
There is an overlap of the ranges of pairwise distances between the partial SSU rDNA
sequences (NS31-AM1 region) within and among D. celata, G. aurantium, and G. eburneum
with over 99% nucleotide sequence identity, in agreement with the poor phylogenetic
resolution of the nuclear SSU rRNA gene tree. One sequence type was even found to be
shared among G. eburneum AZ420A (AM713429) and three isolates of D. celata (FACE 83:
AM713424, FACE 234: AM713422, FACE 272: AM713427). Another sequence type was
found in two of the D. celata isolates (FACE 83: EF581867, FACE 272: EF581873) and in
G. eburneum AZ420A (EF581877). Also in agreement with the higher phylogenetic
resolution, there is, however, no partial LSU rRNA gene sequence type (LR1-FLR2
fragment), shared among these three closely related Diversispora species, nor is there any
overlap in the ranges of intra- versus interspecific sequence similarities, except for
G. aurantium (Table S3). Glomus aurantium showed an unusually high intraspecific LSU
rDNA sequence polymorphism, with the lowest pairwise sequence identity of 93.4%,
compared to >98.6% for G. eburneum and >97.5% for D. celata. The interspecific LSU rRNA
sequence identities between D. celata and its closest relative, G. eburneum, were 94.7-96.8%,
and between D. celata and G. aurantium 91.1-94.6%. The values for G. aurantium and
G. eburneum were in a comparable range of 91.1-95.0%, indicating probably a comparable
molecular taxonomic status of these species. Intraspecific variability in the ITS rDNA region
revealed >97.6% nucleotide identity among the sequences of D. celata and >97.8% identity
among those of G. eburneum, and therefore was higher than the 96.0-97.5% interspecific
sequence identity between these two species, but still confirming occurrence of considerable
intraspecific ITS sequence diversity in certain members of the Glomeromycota.
1

Supporting Information 1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
Notes S4 References to Methods S1–S4, Notes S1–S3 and Table S1
Anonymous. 1969. Flora of the British Fungi. Colour Identification Chart. Royal
Botanic Garden Edinburgh. Her Majesty’s Stationery Office, Glasgow.
de Souza FA, Declerck S. 2003. Mycelium development and architecture, and spore
production of Scutellospora reticulata in monoxenic culture with Ri T-DNA
transformed carrot roots. Mycologia 95: 1004-1012.
Dirks B, Scheffer F. 1930. Der Kohlensäure-Bikarbonatauszug und der
Wasserauszug als Grundlage zur Ermittlung der Phosphorsäurebedürftigkeit der
Böden. Landwirtschaftliche Jahrbücher 71: 73-99.
Gamper H, Leuchtmann A. 2007. Taxon-specific PCR primers to detect two
inconspicuous arbuscular mycorrhizal fungi from temperate agricultural
grassland. Mycorrhiza 17: 145-152.
Gerdemann JW. 1955. Wound healing of hyphae in a phycomycetous mycorrhizal
fungus. Mycologia 47: 916-918.
Gerdemann JW. 1963. Spores of mycorrhizal Endogone species extracted from soil
by wet sieving and decanting. Transactions of the British Mycological Society
46: 235-244.
Gerdemann JW, Trappe JM. 1974. The Endogonaceae in the Pacific Northwest.
Mycologica Memoir No. 5, The New York Botanical Garden, New York, USA.
Guindon S, Gascuel O. 2003. A simple, fast, and accurate algorithm to estimate large
phylogenies by maximum likelihood. Systematic Biology 52: 696-704.
1

Helgason T, Daniell TJ, Husband R, Fitter AH, Young JPW. 1998. Ploughing up
the wood-wide web? Nature 394: 431-43.
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
Hewitt EJ. 1966. Sand and water culture methods used in the study of plant nutrition,
2nd ed. Commonwealth Agricultural Bureaux, Technical Communication No. 22,
East Malling, Kent, UK.
Kennedy LJ, Stutz JC, Morton JB. 1999. Glomus eburneum and G. luteum, two
new species of arbuscular mycorrhizal fungi, with emendation of G. spurcum.
Mycologia 91: 1083-1093.
Koske RE. 1984. Spores of VAM fungi inside spores of VAM fungi. Mycologia 76:
853-862.
Nicolson TH, Gerdeman JW. 1968. Mycorrhizal Endogone species. Mycologia 60:
313-325.
Pfeiffer CM, Walker C. Bloss HE. 1996. Glomus spurcum: a new endomycorrhizal
fungus from Arizona. Mycotaxon 54: 373-382.
Posada D, Crandall KA. 1998. Modeltest: testing the model of DNA substitution.
Bioinformatics 14: 817-818.
Renker C, Heinrichs J, Kaldorf M, Buscot F. 2003. Combining nested PCR and
restriction digest of the internal transcribed spacer region to characterize
arbuscular mycorrhizal fungi on roots from the field. Mycorrhiza 13: 191-198.
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesian phylogenetic inference
under mixed models. Bioinformatics 19: 1572-1574.
Schmidt HA, Strimmer K, Vingron M, von Haeseler A. 2002. TREE-PUZZLE:
maximum likelihood phylogenetic analysis using quartets and parallel
computing. Bioinformatics 18: 502-504.
2

Schwarzott D, Schüßler A. 2001. A simple and reliable method for SSU rRNA gene
DNA extraction, amplification, and cloning from single AM fungal spores.
Mycorrhiza 10: 203-207.
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
Schwarzott D, Walker C, Schüßler A. 2001. Glomus, the largest genus of the
arbuscular mycorrhizal fungi (Glomales), is nonmonophyletic. Molecular
Phylogenetics and Evolution 21: 190–197.
Simon L, Lalonde M, Bruns TD. 1992. Specific amplification of 18S fungal
ribosomal genes from vesicular arbuscular endomycorrhizal fungi colonizing
roots. Applied and Environmental Microbiology 58: 291-295.
Swofford DL. 2003, PAUP: Phylogenetic analysis using parsimony and other
methods. Version 4.0.0b10 beta, Sinauer Associates, Sunderland, MA, USA.
Sylvia DM, Hubbel DH. 1986. Growth and sporulation of vesicular arbsucular
mycorrhizal fungi in aeroponic and membrane systems. Symbiosis 1: 259-267.
Trouvelot S, van Tuinen D, Hijri M, Gianinazzi-Pearson V. 1999. Visualization of
ribosomal DNA loci in spore interphasic nuclei of glomalean fungi by
fluorescence in situ hybridization. Mycorrhiza 8: 203–206.
van Tuinen D, Jacquot E, Zhao B, Gollotte A, Gianinazzi-Pearson V. 1998a.
Characterization of root colonization profiles by a microcosm community of
arbuscular mycorrhizal fungi using 25S rDNA-targeted nested PCR. Molecular
Ecology 7: 879–887.
van Tuinen D, Zhao B, Gianinazzi-Person V. 1998b. PCR in studies of AM fungi:
from primers to application. Ed Varma A. In: Mycorrhiza Manual, Springer
Lab Manual, pp. 387-399.
3

4
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
Vierheilig H, Schweiger P, Brundrett M. 2005. An overview of methods for the
detection and observation of arbuscular mycorrhizal fungi in roots. Physiologia
Plantarum 125: 393-404.
Voets L, de la Providencia E, Declerck S. 2006. Glomeraceae and Gigasporaceae
differ in their ability to form hyphal networks. New Phytologist 172: 185-188.
Walker C. 2005. A simple blue staining technique for arbuscular mycorrhizal and
other root-inhabiting fungi. Inoculum 56, 68-69.
Walker C, Gianinazzi-Pearson V, Marion-Espinasse H. 1993. Scutellospora
castanea, a newly described arbuscular mycorrhizal fungus. Cryptogamie,
Mycologie 14: 279-286.
Walker C, Reed LE, Sanders FE. 1984. Acaulospora nicolsonii, a new
endogonaceous species from Great Britain. Transactions of the British
Mycological Society 83: 360-364.
Walker C, Vestberg M, Demircik F, Stockinger H, Saito M, Sawaki H, Nishmuar
I, Schüßler A. 2007. Molecular phylogeny and new taxa in the Archaeosporales
(Glomeromycota): Ambispora fennica gen. sp. nov., Ambisporaceae fam. nov.,
and emendation of Archaeospora and Archaeosporaceae. Mycological Research
111: 137-153.
White TJ, Bruns TD, Lee S, Taylor JW. 1990. Amplification and direct sequencing
of fungal ribosomal RNA genes for phylogenetics. Eds Innes MA, Gelfand DH,
Sninsky JJ, White TJ. In: PCR Protocols: a guide to methods and applications,
Academic Press, New York pp. 315-322.