Embed Size (px)
Citation preview

Molecular and Biochemical Parasitology, 45 (1990) 241-248 241
Elsevier
MOLBIO 01492
Distinct patterns of tyrosine phosphorylation during the life cycle of Trypanosoma brucei
Mari lyn Parsons j .2, Mary Valentine 3, Julie Deans 3, Gary L. Schieven 4 and Jeffrey A. Ledbet ter 3"4
~ Seattle Biomedical Research Institute, 2Departments of Pathobiology and 3Department of Microbiology, University of Washington, and 40ncogen/Bristol-Myers Squibb, Seattle, WA, U.S.A.
(Received 10 September 1990; accepted 19 October 1990)
Regulation of tyrosine phosphorylation is a critical element in controlling growth and differention in higher eukaryotes. We have determined that the protozoan Trypanosoma brucei, which diverged early in the eukaryotic lineage, possesses multiple proteins
~2 h h 1 which react with a specific anti-phosphotyrosine antiserum. Anti-phosphotyrosine immunoprecipitates of [- P]orthop osp ate- ab- eled cells were shown to contain phosphotyrosine by two-dimensional electrophoresis. Western analysis of cells from different stages of the life cycle demonstrates the appearance of tyrosine-phosphorylated proteins at 40-42 kDa during the transition from slender to stumpy bloodforms. Growth of procyclic form cells in orthovanadate resulted in increased levels of specific tyrosine- phosphorylated proteins. The demonstration of phosphotyrosine-containing proteins in T. brucei and their differential regulation during the life cycle suggests that tyrosine kinases and phosphatases may play an important role in the biology of primitive protozoa.
Key words: Difluoromethylornithine; Protein tyrosine kinase; Tyrosine phosphatase; Trypanosome; Stage regulation
Introduction
Tyrosine phosphorylation is a key regulator of cell proliferation and development in higher euka- ryotes [1]. Modulation of the level of tyrosine phosphorylation plays a major role in oncogenic transformation [2-5]. The importance of tyrosine kinases in growth and development in non-mam- malian multicellular organisms such as Drosophila has recently become evident [6]. Considering yet more divergent organisms, Dictyostelium disco- ideum [7], Saccharomyces cerevisiae [8], and Schi- zosaccharomycespombe [9] have all been shown to possess tyrosine kinase genes or activities. The ev- olutionary distance between Trypanosoma brucei and humans is more than twice the distance be-
Correspondence address: Marilyn Parsons, Seattle Biomedi- cal Research Institute, 4 Nickerson St., Seattle, WA 98109, U.S.A.
Abbreviations: DFMO, difluoromethylornithine; p-tyr, phos- photyrosine; SDS, sodium dodecyl sulfate; PAGE, polyacryl- amide gel electrophoresis.
tween humans and yeast, as calculated from analy- sis of the 16S ribosomal RNA subunits [ 10]. What is the role of tyrosine phosphorylation in protozoa such as T. brucei which diverged early in the euka- ryotic lineage? This study was designed to lay the groundwork for exploring this question.
T. brucei is an extracellular parasite with a highly regulated life cycle [11] and cell cycle [12]. Trypanosomes move into Go at specific points, and then re-enter the cell cycle and move to the next stage of the life cycle. Non-cycling stages include stumpy bloodforms, and proventricular mesocyclic and metacyclic insect forms, while cycling forms include slender bloodforms, and procyclic and epi- mastigote insect forms [11 ]. Many highly passaged laboratory strains of T. brucei are continually cy- cling as slender forms. These monomorphic strains no longer differentiate to stumpy forms. Morpho- logically stumpy forms can be induced in such strains by treatment of the host with the drug di- fluoromethylornithine (DFMO) [13]. The signals which regulate the transition from one stage of the life cycle to the next have not been defined in
0166-6851/91/$03.50 © 1991 Elsevier Science Publishers B.V. (Biomedical Division)

242
trypanosomes. By analogy with higher eukaryotes we would expect that protein phosphorylation and dephosphorylation events may play key regulatory roles in these cyclical transitions.
The data presented in this paper demonstrate that T. brucei possesses multiple proteins that react with an anti-phosphotyrosine antiserum and contain phosphotyrosine (p-tyr). Many of the tyrosine- phosphorylated proteins are detected in all avail- able stages of the life cycle. However, a doublet at 40-42 kDa which is found in both stumpy blood- forms and procyclic forms is absent in slender bloodforms. Because slender bloodforms and pro- cyclic forms are cycling, while stumpy forms are in Go, this tyrosine phosphorylation appears to be stage-regulated, rather than cell cycle-regulated. Treatment of procyclic forms with the tyrosine phosphatase inhibitor orthovanadate results in in- creased levels of tyrosine-phosphorylated sub- strates, particularly the 40/42-kDa species.
Materials and Methods
Trypanosomes. Monomorphic strain EATRO 164, clone IHRI1 [14], and pleiomorphic strain TREU667 [15] were used for all studies. Blood- forms were grown in either irradiated or non-ir- radiated rats, harvested, and separated from the host blood cells by DEAE chromatography [16]. Intermediate and stumpy forms of EATRO164 were induced in monomorphic strains by treatment of the host with 2% DFMO in buffered drinking water for 2 and 4 days, respectively. The popula- tions were characterized as to their composition by morphological analysis following Giemsa staining. Procyclic culture forms were grown in medium SDM-79 containing 10% SeruMax (Sigma) and harvested in mid-log phase.
Immunoblot analyses. Antisera were raised and immunoblots performed according to the pro- cedure of Kamps and Sefton [17]. Briefly, rabbits were immunized with random copolymers of p-tyr, alanine, and glycine conjugated to keyhole limpet hemocyanin. Antibodies were adsorbed onto solid phase phosphotyramine. Following washing with buffers containing phosphoserine and phosphothreonine, the antibodies were eluted with phenyl phosphate, and extensively dialyzed.
This antiserum does not react with phosphoserine or phosphothreonine.
In preparation for immunoblotting, cells were rinsed free of serum, lysed in hot sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS- PAGE) sample buffer containing 50 ~tM ortho- vanadate, and immediately boiled for 5 min. Ali-
• - 6 7
quots containing 3 x 10 to 10 cells (15-50~tg pro- tein) were loaded onto 10% SDS-PAGE gels. Fol- lowing electrophoresis the proteins were electro- phoretically transferred to polyvinylidene difluor- ide (PVDF) membranes. The blots were incubated with anti-p-tyr antibodies and bound antibodies were revealed by [JzSI]protein A.
For solubility analyses, procyclic form cells were lysed in lysis buffer (50 mM Tris-HCl/150 mM NaC1/1 mM orthovanadate/1 mM phenylme- thylsulfonyl fluoride, pH 7.5) plus 2 mM EGTA, 50
- 1 • •
~tg ml leupeptm, and various detergents. The so- lubilized proteins were separated from the pellet by centrifugation at 15 000 x g for 5 min.
Metabolic labeling immunoprecipitation and phosphoaminoacid analysis. For phosphoami- noacid analysis, 5 x 108 mid-log phase TREU667 procyclic forms were grown overnight in 5 ml phosphate-free Cunningham's medium [18] con- taining 2 mCi [32p]orthophosphate. For SDS- PAGE analysis of total cellular phosphoproteins the incubation was scaled down 10-fold. The cells were washed twice in phosphate-free medium and lysed in 25 mM Tris-HCl containing 0.5% SDS and 1 mM dithiothreitol. The lysate was immediately placed in a boiling water bath for 5 rain to inactivate phosphatases and proteases. The sample was di- luted with 4 vols. of the lysis buffer (described above) supplemented with 0.5% aprotinin, 1 mM dithiothreitol, 1% NP-40, and 1% sodium cholate, and incubated on 10 rain on ice to allow detergent exchange. The insoluble material was removed by centrifugation. 10 ~tg of purified anti-p-tyr was added and incubated at 4°C overnight, and then the immune complexes were collected on protein A- Sepharose. The immunoprecipitates were washed four times with washing buffer (50 mM Tris-HC1/ 0.45 M NaC1/0.5% NP-40, pH 8.3), and then bound proteins were released with SDS-PAGE sample buffer. Control experiments determined that cells incubated overnight in phosphate-free medium still

possessed significant levels of immunoreactive proteins. The same procedure was used to affinity purify immunoreactive proteins from unlabeled cells.
Phosphoamino acid analysis followed the me- thod of Cooper et al. [19]. Briefly, radiolabeled samples were spotted onto polyvinylidene difluor- ide paper, and hydrolysed for 1 h at 110°C in con- stant boiling HCI. After drying, the samples were resuspended in first dimension buffer (pH 1.9) con- taining unlabeled phosphoserine, phosphothreon- ine, and p-tyr as internal standards. The samples were electrophoresed at 1000 V for 45 min on 10 x 10 cm plates. The plates were dried, wet in second dimension buffer (pH 3.5), and run at 1000 V for 18 min. The standards were revealed with 0.2% ninhy- drin, and the radiolabeled phosphoamino acids by autoradiography.
Results
Western immunoblot analysis provides a sensi- tive, specific method for detecting proteins phos- phorylated on tyrosine. We analyzed lysates of trypanosome procyclic forms grown in culture for the presence of proteins reactive with an affinity- purified anti-p-tyr antiserum. As shown in Fig. 1 A, many proteins ranging from 35 to 200 kDa were re- vealed. As shown in Fig. 1 B, the reactivity is in- hibited by 40 mM phenyl phosphate, demonstrat- ing its specificity. Many of the bands observed, particularly those in the 40-55 kDa region, react strongly with monoclonal anti-p-tyr antibody 6G9, [20] (not shown). Another monoclonal antibody (5F 11; ref. 20) reacted with other subsets of the pro- teins detected by the polyclonal antibodies.
To confirm that the material detected by the anti- body was truly p-tyr, we performed phosphoamino acid analyses on the immunoreactive proteins. On two-dimensional electrophoresis of total 32P-lab- eled trypanosome lysates, the p-tyr spot was unde- tectable, indicating that it represents a very small proportion of the total phosphoamino acids (not shown). Therefore, we used the anti-p-tyr antibody plus protein A-sepharose to enrich the material of interest. As shown by Western analysis (Fig. 2A), our protocol removes the immunoreactive material from the cell lysate and the reactive proteins can be eluted from the beads by SDS-PAGE sample
243
2 1 9 -
l O 0
68 iil
4 3
A B 1 2 1 2
27
Fig. 1. Specificity of reactivity with anti-p-tyr antibodies. Ly- 7 sate from 10 EATRO164 procychc forms was loaded per lane
on a 10% SDS-PAGE gel. Proteins were transferred to poly- vinylidene difluoride membranes, incubated with anti-p-tyr antibodies, followed by [~25I]protein A. (A) 0.25 lag ml ~ anti- p-tyr antibodies. (B) 1 I-tg ml ~ anti-p-tyr antibodies, plus 40 mM phenyl phosphate. Molecular weight markers are indi- cated. Lanes 1 and 2 differ only in the concentration of ortho-
vanadate used during cell processing.
buffer. Procyclic form cells were labeled with [32p]orthophosphate and the proteins reactive with anti-p-tyr antibody were isolated by immunopreci- pitation.Thepro- teins were eluted, hydrolyzed, and two-dimen- sional electrophoresis performed to identify the ra- diolabelled trypanosome phosphoamino acids. P- tyr was clearly present in these immunoprecipit- ates, as was phosphoserine and phosphothreonine (Fig. 2B). In control immunoprecipitates using nor- mal rabbit serum only a faint phosphoserine spot, and no p-tyr, was observed (not shown). These data confirm the presence ofp-tyr in T. brucei procyclic forms and indicate that it is indeed a parasite prod- uct.
The availability of parasites from different stages allows examination of tyrosine phosphoryl- ation patterns as they relate to the life cycle. We compared procyclic forms with bloodforms from the monomorphic strain EATROI64 and from the pleiomorphic (naturally differentiating) strain TREU667. Lysates prepared from buffy coats col- lected at high parasitemias and the bloodform
244
A 1 2 3
2 1 9 -
1 0 0 -
4 3 - ~ ; ~
ii ii!!i !i ~ ili!~!i~ ~i~i!!~iiii~i~i! i
B
S
Y
Fig. 2. Immunoprecipitation and phosphoamino acid analy- sis. (A) Western analysis of anti-p-tyr immunoprecipitates. Immunoreactive proteins were enriched by immunoprecipita- tion as described in Methods. Each lane was loaded with ma- terial from the same amount of cell lysate. Lanes: (1) total cell lysate; (2) lysate cleared of proteins reactive with anti-p-tyr; (3) eluate from anti-p-tyr immunoprecipitation. (B) Two-di- mensional phosphoamino acid analysis. Procyclic forms were metabolically labeled with [32p]orthophosphate, and proteins reactive with the anti-p-tyr antiserum were enriched by immu- noprecipitation as above. The proteins were hydrolyzed and phosphoamino acids separated by two-dimensional electro- phoresis. The radioactive spots co-migrated exactly with phos- phoamino acid standards revealed with ninhydrin: S, serine; T,
threonine; Y, tyrosine.
trypanosomes subsequently purified from them showed no consistent differences in p-tyr protein patterns (not shown), indicating that the purifi- cation of the parasite did not alter the phosphoryla- tion of the major substrates. The most significant contaminant of the bloodform trypanosomes is red blood cells (< 1%). When red blood cells equivalent to the typical contamination were analyzed, p-tyr containing proteins were not seen (not shown), in- dicating that the tyrosine-phosphorylated proteins are probably not derived from the host. This is sup- ported by the observation that most of the tyrosine- phosphorylated proteins seen in bloodforms are the same as those seen in procyclic forms grown in the absence of host cells.
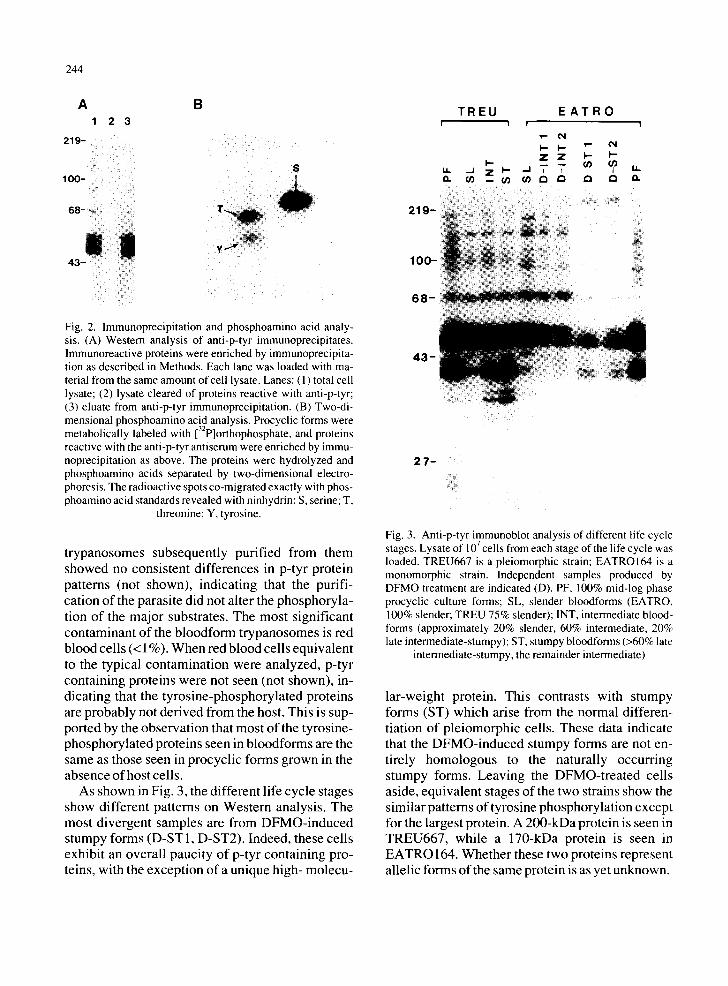
As shown in Fig. 3, the different life cycle stages show different patterns on Western analysis. The most divergent samples are from DFMO-induced stumpy forms (D-ST1, D-ST2). Indeed, these cells exhibit an overall paucity of p-tyr containing pro- teins, with the exception of a unique high- molecu-
219-
100-
6 8 -
T R E U i i
I-- u,. ,_1 z I - a . ( / ) m ¢ / )
E A T R O ! !
I - - I - " ' = " ¢ q Z Z I - - I - - _ - - ( / )
. . I 14. I /
4 3 -
i ¸¸
2 7 ~
Fig. 3. Anti-p-tyr immunoblot analysis of different life cycle 7 stages. Lysate of 10 cells from each stage of the hfe cycle was
loaded. TREU667 is a pleiomorphic strain; EATROI64 is a monomorphic strain. Independent samples produced by DFMO treatment are indicated (D). PF, 100% mid-log phase procyclic culture forms; SL, slender bloodforms (EATRO, 100% slender; TREU 75% slender); INT, intermediate blood- forms (approximately 20% slender, 60% intermediate, 20% late intermediate-stumpy); ST, stumpy bloodforms (>60% late
intermediate-stumpy, the remainder intermediate)
lar-weight protein. This contrasts with stumpy forms (ST) which arise from the normal differen- tiation of pleiomorphic cells. These data indicate that the DFMO-induced stumpy forms are not en- tirely homologous to the naturally occurring stumpy forms. Leaving the DFMO-treated cells aside, equivalent stages of the two strains show the similar patterns of tyrosine phosphorylation except for the largest protein. A 200-kDa protein is seen in TREU667, while a 170-kDa protein is seen in EATRO 164. Whether these two proteins represent allelic forms of the same protein is as yet unknown.
219 -
1 0 0 -
1 2 3 4 P ~ P .q T P
68
4 3 -
27
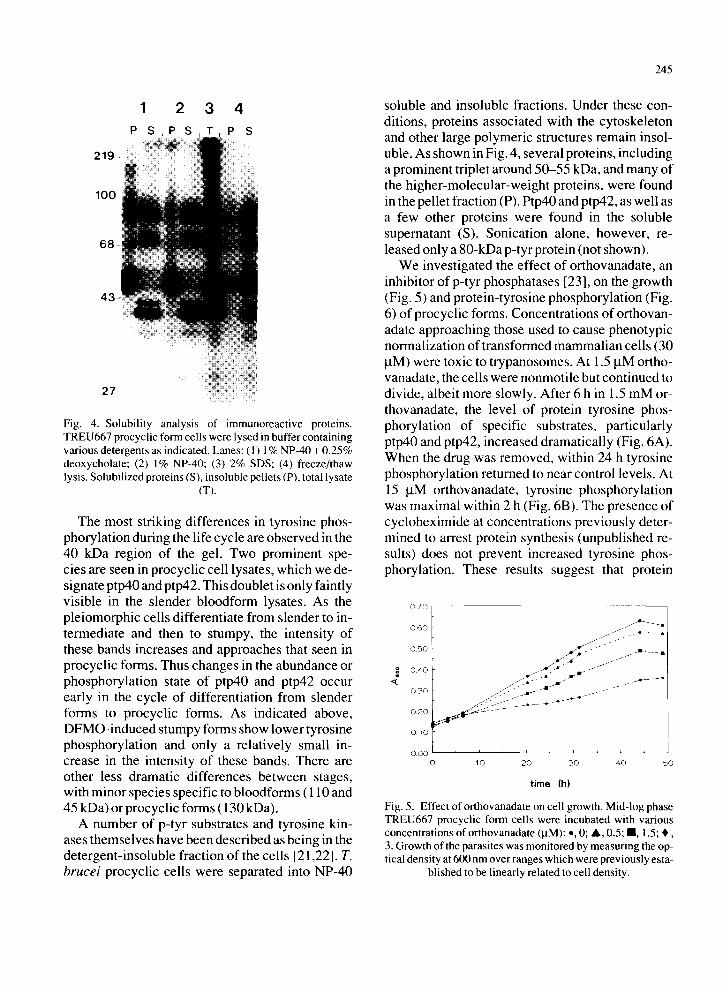
Fig. 4. Solubility analysis of immunoreactive proteins. TREU667 procyclic form cells were lysed in buffer containing various detergents as indicated. Lanes: ( 1 ) 1% NP-40 + 0.25% deoxycholate; (2) 1% NP-40; (3) 2% SDS; (4) freeze/thaw lysis. Solubilized proteins (S), insoluble pellets (P), total lysate
(T).
The most striking differences in tyrosine phos- phorylation during the life cycle are observed in the 40 kDa region of the gel. Two prominent spe- cies are seen in procyclic cell lysates, which we de- signate ptp40 and ptp42. This doublet is only faintly visible in the slender bloodform lysates. As the pleiomorphic cells differentiate from slender to in- termediate and then to stumpy, the intensity of these bands increases and approaches that seen in procyclic forms. Thus changes in the abundance or phosphorylation state of ptp40 and ptp42 occur early in the cycle of differentiation from slender forms to procyclic forms. As indicated above, DFMO-induced stumpy forms show lower tyrosine phosphorylation and only a relatively small in- crease in the intensity of these bands. There are other less dramatic differences between stages, with minor species specific to bloodforms ( 110 and 45 kDa) or procyclic forms (130 kDa).
A number of p-tyr substrates and tyrosine kin- ases themselves have been described as being in the detergent-insoluble fraction of the cells [21,22]. T. brucei procyclic cells were separated into NP-40
245
soluble and insoluble fractions. Under these con- ditions, proteins associated with the cytoskeleton and other large polymeric structures remain insol- uble. As shown in Fig. 4, several proteins, including a prominent triplet around 50-55 kDa, and many of the higher-molecular-weight proteins, were found in the pellet fraction (P). Ptp40 and ptp42, as well as a few other proteins were found in the soluble supernatant (S). Sonication alone, however, re- leased only a 80-kDa p-tyr protein (not shown).
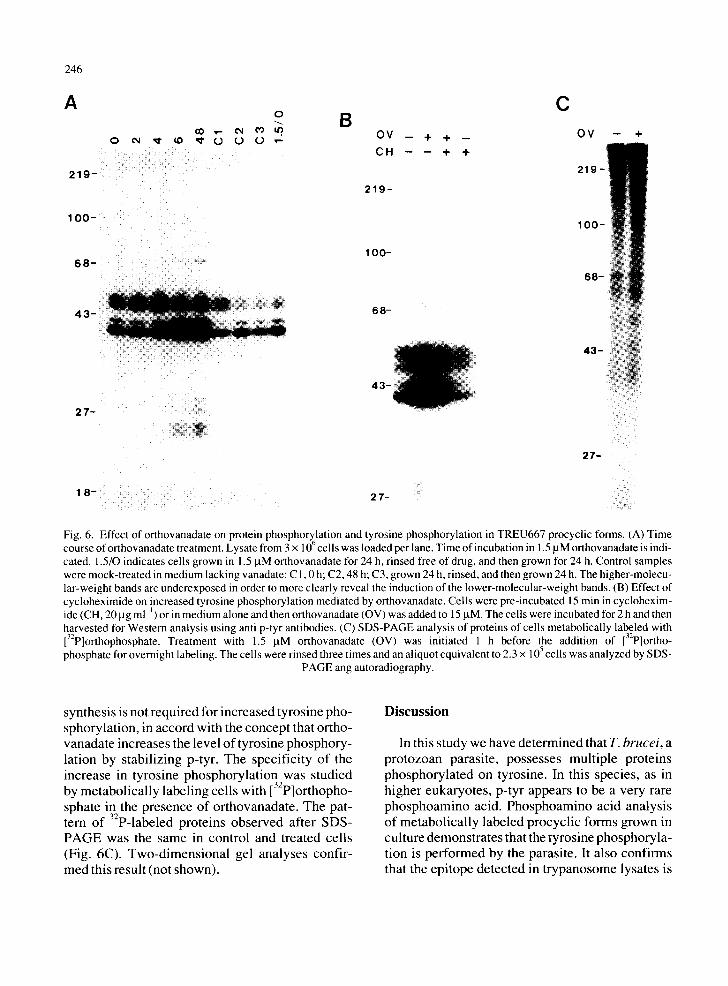
We investigated the effect of orthovanadate, an inhibitor of p-tyr phosphatases [23], on the growth (Fig. 5) and protein-tyrosine phosphorylation (Fig. 6) of procyclic forms. Concentrations of orthovan- adate approaching those used to cause phenotypic normalization of transformed mammalian cells (30 taM) were toxic to trypanosomes. At 1.5 laM ortho- vanadate, the cells were nonmotile but continued to divide, albeit more slowly. After 6 h in 1.5 mM or- thovanadate, the level of protein tyrosine phos- phorylation of specific substrates, particularly ptp40 and ptp42, increased dramatically (Fig. 6A). When the drug was removed, within 24 h tyrosine phosphorylation returned to near control levels. At 15 ~tM orthovanadate, tyrosine phosphorylation was maximal within 2 h (Fig. 6B). The presence of cycloheximide at concentrations previously deter- mined to arrest protein synthesis (unpublished re- suits) does not prevent increased tyrosine phos- phorylation. These results suggest that protein
0 7 0
g 4
0 . 6 0
0 . 5 0
0 . 4 0
0 3 0
0 . 2 0
I 0 . 1 0
0 . 0 0 0
. . A ~ i . . r
. ~11 f J
i , i , i ,
10 2 0 3 0 4 0 5 0
time (h)
Fig. 5. Effect of orthovanadate on cell growth. Mid-log phase TREU667 procyclic form cells were incubated with various concentrations of orthovanadate (laM): o, 0; A, 0.5; II, 1.5; , , 3. Growth of the parasites was monitored by measuring the op- tical density at 600 nm over ranges which were previously esta-
blished to be linearly related to cell density.
246
A
2 1 9 -
1 0 0 -
6 8 -
4 3
o B
0 ~ '~ tO ~' rO ¢0 ~ ',,=" OV
CH
2 1 9 -
1 0 0 -
6 8 -
- ÷ ÷
+ ÷
C OV
2 1 9 -
1 0 0 -
6 8 -
- - .4-
2 7 -
4 3 -
1 8 - 2 7 -
4 3 -
i i i i
2 7 -
~ ~;~;~i~i~i~ili~) ~ ~
Fig. 6. Effect of orthovanadate on protein phosphorY61ation and tyrosine phosphorylation in TREU667 procyclic forms. (A) Time course of orthovanadate treatment. Lysate from 3 × 10 cells was loaded per lane. Time of incubation in 1.5 gM orthovanadate is indi- cated. 1.5/0 indicates cells grown in 1.5 JaM orthovanadate for 24 h, rinsed free of drug, and then grown for 24 h. Control samples were mock-treated in medium lacking vanadate: C 1,0 h; C2, 48 h; C3, grown 24 h, rinsed, and then grown 24 h. The higher-molecu- lar-weight bands are underexposed in order to more clearly reveal the induction of the lower-molecular-weight bands. (B) Effect of cycloheximide on increased tyrosine phosphorylation mediated by orthovanadate. Cells were pre-incubated 15 min in cyclohexim- ide (CH, 20 lag ml ~) or in medium alone and then orthovanadate (OV) was added to 15 laM. The cells were incubated for 2 h and then harvested for Western analysis using anti p-tyr antibodies. (C) SDS-PAGE analysis of proteins of cells metabolically labeled with 132P]orthophosphate. Treatment with 1.5 laM orthovanadate (OV) was initiated 1 h before the addition of 132p]ortho - phosphate for overnight labeling. The ceils were rinsed three times and an aliquot equivalent to 2.3 x 105 cells was analyzed by SDS-
PAGE ang autoradiography.
synthesis is not required for increased tyrosine pho- sphorylation, in accord with the concept that ortho- vanadate increases the level of tyrosine phosphory- lation by stabilizing p-tyr. The specificity of the increase in tyrosine phosphorylation was studied by metabolically labeling cells with [32p]orthopho- sphate in the presence of orthovanadate. The pat- tern of 32p-labeled proteins observed after SDS- PAGE was the same in control and treated cells (Fig. 6C). Two-dimensional gel analyses confir- med this result (not shown).
Discussion
In this study we have determined that T. brucei, a protozoan parasite, possesses multiple proteins phosphorylated on tyrosine. In this species, as in higher eukaryotes, p-tyr appears to be a very rare phosphoamino acid. Phosphoamino acid analysis of metabolically labeled procyclic forms grown in culture demonstrates that the tyrosine phosphoryla- tion is performed by the parasite. It also confirms that the epitope detected in trypanosome lysates is

p-tyr, and not some unusual phosphoserine or phos- phothreonine derivative as has been observed in yeast using a different anti-p-tyr antiserum [24]. The T. brucei proteins reacting with the anti-p-tyr antiserum, or proteins associated with them, also contain phosphoserine and phosphothreonine (as do most other tyrosine-phosphorylated proteins in other species; ref. 25). This raises the possibility of intricate interactions between various kinases and their substrates, suggesting the potential for com- plex regulation.
Our studies indicate that a subset of p-tyr con- taining proteins, including ptp40 and ptp42, are dif- ferentially regulated during the life cycle. Ptp40 and ptp42 are absent in immunoblots of slender bloodforms, but intermediate and stumpy blood- forms show levels approaching those observed in procyclic forms. Since stumpy forms are non-di- viding, while procyclic forms are dividing, the abundance of ptp40 and ptp42 appears to be corre- lated with the stage of the life cycle, rather than with quiescent or dividing states. It is intriguing that in- creases in ptp40 and ptp42 occur early in the differ- entiation of bloodform parasites. It is possible that phosphorylation of these substrates is important for controlling the differentiation process. The exact relationship of DFMO-induced stumpy forms to naturally occurring stumpy forms is not clear; how- ever, they share a number of metabolic character- istics [26,27]. Our data indicate that these similar- ities do not extend to the level of tyrosine phosphorylation, suggesting that the DFMO- treated stumpy form is distinct from the naturally occurring stumpy form.
The existence of tyrosine phosphorylation in T. brucei implies the existence of tyrosine kinases. It is now important to identify and characterize these kinases. Unicellular organisms are generally thought not to possess receptor type tyrosine kin- ases, because they do not exist in a 'cellular' en- vironment where intercellular communication is vital [2]. However, parasites such as T. brucei exist in a cellular environment, and may have evolved mechanisms to sense the host status, allowing them to regulate their proliferation or differentiation. Evidence for extracellular factors affecting the growth and development of T. brucei has been ob- tained. The most clear-cut evidence is the demon- stration of an epidermal growth factor receptor on
247
T. brucei [28]. Although the molecular nature of the T. brucei receptor has not been described, in mam- malian cells the epidermal growth factor receptor is a protein tyrosine kinase [2]. Other data indicate that serum factors and possibly parasite-derived factors affect the differentiative capacity of trypanosomes [29,30]. By analogy to other eukary- otes we hypothesize that signal transduction in T. brucei may include either receptor or cytoplasmic tyrosine kinases.
In conjunction with tyrosine kinases, tyrosine phosphatases regulate tyrosine phosphorylation of physiological substrates [31 ]. The ability of ortho- vanadate, a phosphotyrosine phosphatase inhibi- tor, to increase the level of p-tyr containing pro- teins, suggests that such phosphatases also function in these ancient organisms. As in other systems [3,4], orthovanadate specifically increases the level of tyrosine phosphorylation, but does not cause major alterations in overall protein phosphoryla- tion (which is primarily phosphoserine and phos- phothreonine). However, the effects of vanadate are complex as it is an inhibitor of other enzymes not normally thought to function in tyrosine phos- phorylation pathways (see refs. 3 and 24). The rela- tive contribution of increased tyrosine phosphoryl- ation to the inhibition of growth remains to be elucidated. Whatever the mechanism, vanadate treatment provides a simple method for increasing the tyrosine phosphorylation of specific proteins, in particular ptp40 and ptp42 kDa, for future bio- chemical analysis and immunoaffinity purifi- cation.
The presence of regulated, phosphotyrosine- containing proteins in this ancient species suggests that phosphotyrosine regulatory circuits may be important in the growth and development of T. bru- cei and other primitive eukaryotes. Thus, the characterization of T. brucei protein tyrosine kin- ases, tyrosine phosphatases, and their inhibitors be- comes important as a potential route for the devel- opment of anti-parasite agents. Further investigation of these regulatory circuits will aid our understanding of the evolution of growth con- trol and host-parasite interactions.
Acknowledgements
The authors would like to thank Victoria Carter

248
and Karen Licciardi for their exce l len t technical as- sistance. Michel le Maddox provided secretarial support. This work was supported in part by NIH
AI22635, GM42508 , and the Murdock Char i table Trust.
References
1 Hunter, T. and Cooper, J.A. (1985) Protein-tyrosine kin- ases. Annu. Rev. Biochem. 54,897-930.
2 Yarden, Y. and Ullrich, A. (1989) Growth factor receptor tyrosine kinases. Annu Rev. Biochem. 57,443-478.
3 Karlund, J. (1985) Transformation of cells by an inhibitor of phosphatases acting on phosphotyrosine proteins. Cell 41,707-717.
4 Feldman, R.A., Lowy, D.R. and Vass, W.C. (1990) Selec- tive potentiation of c-fps/fes transforming activity by a phosphatase inhibitor. Oncogene Res. 5, 187-197.
5 Uehara, Y., Hori, M., Takeuchi, T. and Umezawa, H. (1985) Screening of agents which convert 'transformed morphology' of Rous sarcoma virus-infected rat kidney cells to 'normal morphology': identification of an active agent as herbimycin and its inhibition of intracellular src kinase. Jpn. J. Cancer Res. 76,672-675.
6 Hoffman, F.M. (1989) Roles of Drosophila proto-onco- gene and growth factor homologs during development of the fly. Curr. Top. Microbiol. Immunol. 147, 1-29.
7 Tan, J.L. and Spudich, J.A. (1990) Developmentally regu- lated protein-tyrosine kinase genes in Dictyostelium disco- ideum. Mol. Cell. Biol. 10, 3578-3583.
8 Schieven, G., Thorner, J. and Martin, G.S. (1986) Protein- tyrosine kinase activity in Saccharomyces cerevisiae. Science 231,390-393.
9 Gould, K.L. and Nurse, P. (1989) Tyrosine phosphoryla- tion of the fission yeast cdc2 protein kinase regulates entry into mitosis. Nature 342, 39-45.
10 Sogin, M.L., Gunderson, J.H., Elwood, H.J., Alonso, R.A. and Peattie, D.A. (1989) Phylogenetic meaning of the kingdom concept: an unusual ribosomal RNA from Giar- dia lamblia. Science 243, 75-77.
11 Vickerman, K. (1985) Developmental cycles and biology of pathogenic trypanosomes. Br. Med. Bull. 41,105-114.
12 Woodward, R. and Gull, K. (1990) Timing of nuclear and kinetoplast DNA replication and early morphological events in the cell cycle ofTrvpanosoma brucei. J. Cell Sci. 95, 49-57.
13 Bacchi, C.J., Garofalo, J., Mockenhaupt, D., McCann, P.P., Diekema, K.A., Pegg, A.E., Nathan, H.C., Mullaney, E.A., Chunosoff, L., Sjoerdsma, A. and Hunter, S.H. (1983) In vivo effects of alpha-DL-difluoromethylomithine on the metabolism and morphology of Trypanosoma bru- cei. Mol. Biochem. Parasitol. 7,209-225.
14 Stuart, K., Gobright, E., Jenni, L., Milhausen, M., Thoma- show, L.S. and Agabian, N. (1984) The IsTaR serodeme of Trypanosoma brucei: development of a new serodeme. Pa- rasitology 70, 747-754.
15 Michelotti, E.F. and Hajduk, S.L. (1987) Developmental
regulation of trypanosome mitochondrial gene expression. J. Biol. Chem. 262, 927-932.
16 Lanham, S.M. (1968) Separation of trypanosomes from the blood of infected rats and mice by anion-exchangers. Na- ture 218, 1273-1274.
17 Kamps, M.P. and Sefton, B.M. (1988) Identification of multiple novel polypeptide substrates of the v-src, v-yes, v- fps, v-ros, and v-erb-B oncogenic tyrosine protein kinases utilizing antisera against phosphotyrosine. Oncogene 2, 305-315.
18 Cunningham, I. (1977) New culture medium for mainten- ance of tsetse tissues and growth of trypanosomatids. J. Protozool. 24,325-329.
19 Cooper, J.A., Sefton, B.M. and Hunter, T. (1983) Detec- tion and quantification of phosphotyrosine in proteins. Me- thods Enzymol. 99,387--402.
20 Kanner, S.B., Reynolds, A.B., Vines, R.R. and Parsons, J.T. (1990) Monoclonal antibodies to individual tyrosine- phosphorylated protein substrates of oncogene-encoded tyrosine kinases. Proc. Natl. Acad. Sci. USA 87, 3328- 3332.
21 Zippel, R., Morello, L., Brambilla, R., Comoglio, P.M., AI- gerghina, L. and Sturani, E. (1989) Inhibition of phospho- tyrosine phosphatases reveals candidate substrates of the PDGF receptor kinase. Eur. J. Cell Biol. 50, 428-434.
22 Gold, M.R., Law, D.A. and DeFranco, A.L. (1990) Stimu- lation of protein tyrosine phosphorylation by the B-lym- phocyte antigen receptor. Nature 345, 810-813.
23 Swarup, G., Cohen, S. and Garbers, D.L. (1982) Inhibition of membrane phosphotyrosyl-protein phosphatase activity by vanadate. Biochem. Biophys. Res. Commun. 107, 1104-1109.
24 Kanik-Ennulat, C. and Neff, N. (1990) Vanadate-resistant mutants of Saccharomyces cerevisiae show alterations in protein phosphorylation and growth control. Mol. Cell. Biol. 10, 898-909.
25 Kanner, S.B., Reynolds, A.B. and Parsons, J.T. (1989) Im- munoaffinity purification of tyrosine-phosphorylated cellular proteins. J. Immunol. Methods 120, 115-124.
26 Vickerman, K. (1965) Polymorphism and mitochondrial activity in sleeping sickness trypanosomes. Nature 208, 762-766.
27 Feagin, J.E., Jasmer, D.P. and Stuart, K. (1986) Differen- tial mitochondrial gene expression between slender and stumpy bloodforms of Trypanosoma brucei. Mol. Bio- chem. Parasitol. 20,207-214.
28 Hide, G., Gray, A., Harrison, C.M. and Tait, A. (1989) Identification of an epidermal growth factor receptor homologue in trypanosomes. Mol. Biochem. Parasitol. 36, 51-60.
29 Otigbuo, I.N. and Woo, P.T.K. (1987) Trypanosoma b. brucei: a culture medium reverting a monomorphic to a pleiomorpbic strain. Trans. R. Soc. Trop. Med. Hyg. 81, 408-410.
30 Seed, J.R. and Sechelski, J.B. (1989) Mechanism of long slender (LS) to short stumpy (SS) transformation in Af- rican trypanosomes. J. Protozool. 36,572-577.
31 Hunter, T. (1989) Protein-tyrosine phosphatases: the other side of the coin. Cell 58, 1013-1016.





























