Embed Size (px)
Citation preview

411INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSESRevista Chilena de Historia Natural77: 411-437, 2004
Disruption of ecosystem processes in western North Americaby invasive species
Alteración de procesos en ecosistemas en el oeste de Norteamérica producidospor especies invasoras
JEFFREY S. DUKES1 & HAROLD A. MOONEY
Department of Biological Sciences, Stanford University, Stanford, California 94305-5020 USACurrent address1: Biology Department, University of Massachusetts, Boston, Massachusetts 02125 USA;
e-mail: [email protected]
ABSTRACT
Many ecosystems of western North America have been dramatically changed by non-native species. Here, wereview ecological impacts of 56 plant, animal, fungus, and protist species that were brought to this region byhumans. We discuss characteristics of invasive species that can lead to major ecosystem impacts, and explorehow invasive species alter many different attributes of ecosystems. Specifically, we include examples ofinvasive species that affect geomorphology, fire regimes, hydrology, microclimate, atmospheric composition,nutrient cycling, and productivity. Finally, we review the direct consequences of biological invasions forsome native species. We summarize examples from this paper in Appendix 1. Our examples illustrate how, asinvasive species have become dominant across large areas of western North America’s grassland, shrubland,dune, riparian, and estuarine ecosystems, the properties and functioning of these systems have changed. Todate, some systems in this region, such as its forests, remain relatively unaffected by invasive species.However, recent attacks of forest pathogens highlight the potential vulnerability of these ecosystems.
Key words: biological invasions, ecosystem functioning, community structure, exotic species, impact.
RESUMEN
Muchos ecosistemas de Norteamérica occidental han cambiado dramáticamente a causa del efecto producidopor especies no autóctonas. Aquí se muestra una revisión del impacto ecológico producido por 56 especiesdiferentes de plantas, animales y hongos, y especies de protistas que fueron traídos a esta región por humanos.Discutimos las características de las especies invasoras que pueden producir un gran impacto en elecosistema, y exploramos cómo las especies invasoras pueden alterar de forma muy diferente los atributos deun ecosistema. Específicamente, incluimos ejemplos de especies invasoras que afectan a la geomorfología, alos regímenes del fuego, a la hidrología, al microclima, a la composición atmosférica, al ciclo de nutrientes, ya la productividad. Finalmente, revisamos las consecuencias directas de invasiones biológicas de algunasespecies autóctonas. Resumimos los ejemplos de este artículo en el Anexo 1. Nuestros ejemplos ilustrancómo, a medida que la especie invasora llega a ser dominante a lo largo de áreas extensas de ecosistemascomo los prados del oeste de Norteamérica occidental, en zonas arbustivas, dunas, cauces de ríos y estuarios,las propiedades y el funcionamiento de estos ecosistemas han cambiado. Hasta ahora, algunos ecosistemas enesta región, como los bosques, permanecen relativamente intactos por efecto de la especies invasoras. Sinembargo, ataques recientes de patógenos a los bosques ponen de manifiesto la vulnerabilidad potencial deestos ecosistemas.
Palabras clave: invasiones biológicas, funcionamiento ecosistémico, estructura de comunidades, especiesexóticas, impacto.
INTRODUCTION
As global t ransport becomes faster andcheaper, the distant corners of our planetbecome increasingly connected. People andtheir products, traveling from continent tocontinent, provide opportunities for thousands
of plant and animal species to be transported,or even to hitchhike along. Most of thehitchhiking species do not survive in their newenvironment. However, some thrive, and someof those that thrive cause great ecological oreconomic harm. Many alien species attack oroutcompete nat ive species, and a small

412 DUKES & MOONEY
percentage cause major changes in theappearance and operation of ecosystems(Vitousek et al. 1997, Sala et al. 1999).Invasive species (those aliens that thrive andincrease their ranges) have already done greateconomic harm in countries around the world,either by depressing growth or populations ofmore valuable species, or by directly impedinghuman activity (e.g., Pimentel et al. 2000).The acceleration of international trade islikely to increase the number of propagulesthat are transported out of their home rangeseach day. Thus, unless measures are taken toprevent propagules from hitching rides, theongoing expansion of global commerce islikely to exacerbate the problem of biologicalinvasions.
Here, we examine some of the ecologicalimpacts of a variety of alien species, includingseveral invasives. Firs t , we examinecharacteristics of species that can lead to largeecosystem impacts after their introduction.Then, we explore how alien species areal ter ing many different at t r ibutes ofecosystems, such as geomorphology, fireregime, hydrology, microclimate, atmosphericcomposi t ion, nutr ient cycl ing, andproductivity. Finally, we review the directconsequences of biological invasions for somenative species . Where possible we useexamples of invasive species in western NorthAmerican ecosystems. These examples aresummarized in Appendix 1.
Invasive species that affect ecosystemprocesses may indirectly impact populations ofnative species. A simplified conceptual modelof direct and indirect interactions among nativeand alien species is shown in Fig. 1.
We must emphasize that the ecologicalimpacts of many of western North America’sinvasive species have not been studied, and sothis review should not be viewed ascomprehensive. We have simply attempted tocompile a survey of some invaders’ impacts(and potential impacts) in this region.
WHICH BIOLOGICAL INVADERS ARE MOST LIKELY
TO ALTER ECOSYSTEMS?
Much of western North America’s currentbiota is non-native. For instance, the latestsurveys show that 1,109 of California’s 8,274catalogued species (13.4 %) were introducedfrom elsewhere (Hobbs & Mooney 1998).Which of the invaders have the potential todisrupt ecosystems? Invasive species that
differ from natives in some trait, behavior, orfunct ion increasingly al ter ecosystemproperties and processes as their populationsexpand. Such species can be grouped into twocategories: discrete t rai t invaders andcontinuous trai t invaders. Discrete trai tinvaders add a new function to the invadedecosystem, such as ni t rogen f ixat ion,hydraulic lift, or predation on a particulartrophic level. Continuous trait invaders differfrom nat ives only in t rai ts that arecontinuously distributed among species suchas litter quality or relative growth rate. Chapinet al. (1996) argue that discrete trait invadersare more likely than continuous trait invadersto have large ecosystem effects, and a recentmeta-analysis of invaders’ effects ondisturbance regimes supports this argument(D’Antonio et al. 1999). However, continuoustrai t invaders can also al ter ecosystemstructure and functioning, especially if theyconst i tute a large proport ion of theecosystem’s biomass at one trophic level.
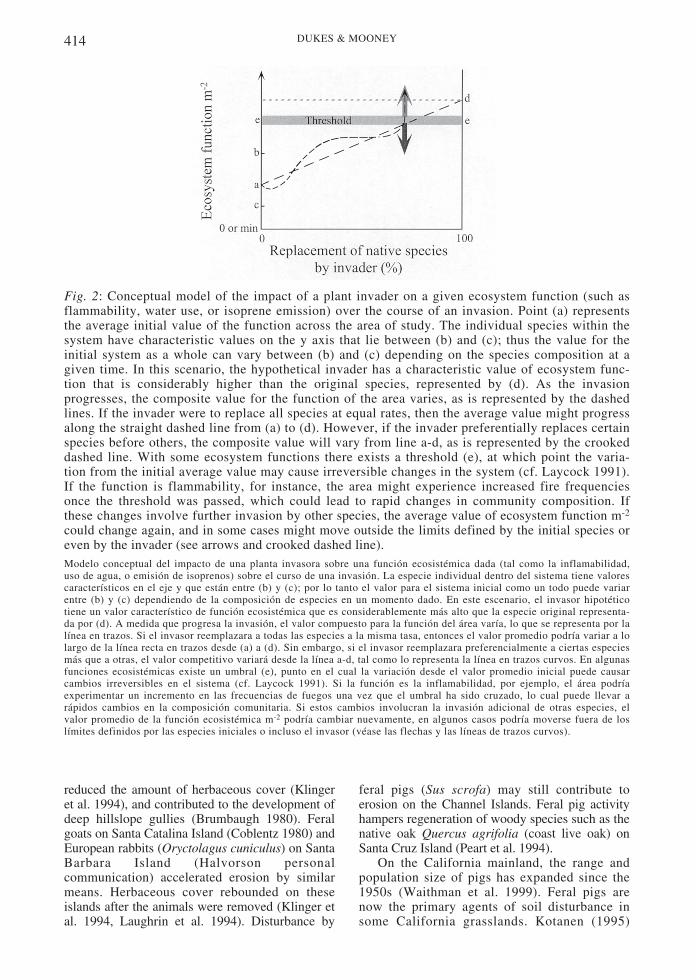
The conceptual model in Fig. 2 illustrateshow an invader that replaces other species inits trophic level can alter properties of anecosystem such as water use, flammability, orisoprene emission. In this example, theinvading species has a higher inherent valuefor the hypothetical ecosystem function thanthe native species (although this value couldjust as easily be lower than that of the native).In an uninvaded ecosystem, the value of theecosystem function may vary over time due toshifts in species dominance. As an invasionprogresses, the invader makes up anincreasing proportion of biomass at its trophiclevel. This forces the value of the functiontoward the inherent value of the invader. If thefunction crosses some threshold (increasedwater use draws down water tables below acertain level , increased f lammabil i tyaccelerates the fire cycle, increased predationon an herbivore reduces vegetat iondisturbance, among others) , the speciescomposition of the region may change, eitheras a result of the local elimination of a nativespecies that required the pre- invasionconditions for survival or due to an increase inthe susceptibility of the system to invasion byother species. Species composition changecould further displace the ecosystem functionor trait from its initial value. However,competition with native species may preventthe invader from achieving a great enoughdominance to force the value of the ecosystemfunction across a threshold.

413INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
Fig. 1: Conceptual model of interactions among biological invaders, native species, ecosystemprocesses, global commerce, and global change. Arrows show directions of influence, and symbolsnext to arrows (in descending order: +, -/+, -) indicate whether a given influence is generallythought to be positive or negative. Because “ecosystem processes” encompasses several indepen-dent elements that are not easily generalized, arrows from this compartment are left blank. Globaltransport has brought invasive species to new regions, where some of these species suppress nativepopulations. Together, native and alien species modulate ecosystem processes. Elements of globalchange such as nitrogen deposition, habitat fragmentation, and global change affect both ecosystemprocesses and the balance between native and alien species (Dukes & Mooney 1999).Modelo conceptual de interacciones entre invasores biológicos, especies nativas, procesos ecosistémicos, comercio globaly cambio global. Las flechas indican dirección de las interacciones, y los símbolos al lado (en orden ascendente odescendente +,-/+, -) indican si una influencia dada es en general interpretada como positiva o negativa. Debido a que elconcepto de “procesos ecosistémicos” abarca varios elementos independientes que no son fácilmente generalizables, lasflechas desde este compartimiento se dejan en blanco. El transporte global ha llevado especies invasoras a nuevas regiones,donde algunas de estas especies suprimen a las poblaciones de especies nativas. En conjunto, las especies nativas yforáneas modulan los procesos ecosistémicos. Elementos del cambio global tales como la deposición de nitrógeno, frag-mentación de hábitat, y cambio global afectan tanto los procesos y el balance entre las especies nativas y foráneas (Dukes& Mooney 1999).
IMPACTS OF NON-NATIVE SPECIES
Geomorphology and soil disturbance regimes
Introduced animals and plants have alteredgeomorphic processes in many of westernNorth America’s ecosystems. Beaver (Castorcanadensis) are native to some parts of thewest, but have been introduced in areas beyondtheir original range (Johnson & Harris 1990).By building dams, beaver directly modify themorphology and hydrology of streams. Damscreate ponds, slow the stream current, increasesediment retention, alter seasonal streamdischarge regimes, and expand the influence ofthe water table (Naiman et al . 1988).Decomposition, nutrient cycling, and waterquality are also altered by beaver ponds(Naiman et al. 1986). These effects caninfluence the composition of downstream plantand animal communities (Pollock et al. 1995).
Although the introduction of beaver cansubstantially alter stream geomorphology, theextent of these changes is limited by thenumber of suitable sites for beaver ponds(Johnston & Naiman 1990).
Just as beaver dams trap substrate and increasesedimentation in an area, other introduced animalscause soil to be lost. Many species contribute toerosion by disturbing the soil or overgrazingvegetation. Studies of these phenomena onCalifornia’s Channel Islands led to theimplementation of new land managementpractices. For instance, feral goats (Capra hircus)and sheep (Ovis aries) were removed orexterminated on some islands once their impactson geomorphology and vegetation were understood(Schuyler 1987, Keegan et al. 1994, Laughrin et al.1994). Before their eradication in 1987, sheepcompacted the soils of Santa Cruz Island andovergrazed the plants. These activities suppressedwoody species regeneration (Wehtje 1994),
414 DUKES & MOONEY
reduced the amount of herbaceous cover (Klingeret al. 1994), and contributed to the development ofdeep hillslope gullies (Brumbaugh 1980). Feralgoats on Santa Catalina Island (Coblentz 1980) andEuropean rabbits (Oryctolagus cuniculus) on SantaBarbara Island (Halvorson personalcommunication) accelerated erosion by similarmeans. Herbaceous cover rebounded on theseislands after the animals were removed (Klinger etal. 1994, Laughrin et al. 1994). Disturbance by
Fig. 2: Conceptual model of the impact of a plant invader on a given ecosystem function (such asflammability, water use, or isoprene emission) over the course of an invasion. Point (a) representsthe average initial value of the function across the area of study. The individual species within thesystem have characteristic values on the y axis that lie between (b) and (c); thus the value for theinitial system as a whole can vary between (b) and (c) depending on the species composition at agiven time. In this scenario, the hypothetical invader has a characteristic value of ecosystem func-tion that is considerably higher than the original species, represented by (d). As the invasionprogresses, the composite value for the function of the area varies, as is represented by the dashedlines. If the invader were to replace all species at equal rates, then the average value might progressalong the straight dashed line from (a) to (d). However, if the invader preferentially replaces certainspecies before others, the composite value will vary from line a-d, as is represented by the crookeddashed line. With some ecosystem functions there exists a threshold (e), at which point the varia-tion from the initial average value may cause irreversible changes in the system (cf. Laycock 1991).If the function is flammability, for instance, the area might experience increased fire frequenciesonce the threshold was passed, which could lead to rapid changes in community composition. Ifthese changes involve further invasion by other species, the average value of ecosystem function m-2
could change again, and in some cases might move outside the limits defined by the initial species oreven by the invader (see arrows and crooked dashed line).Modelo conceptual del impacto de una planta invasora sobre una función ecosistémica dada (tal como la inflamabilidad,uso de agua, o emisión de isoprenos) sobre el curso de una invasión. La especie individual dentro del sistema tiene valorescaracterísticos en el eje y que están entre (b) y (c); por lo tanto el valor para el sistema inicial como un todo puede variarentre (b) y (c) dependiendo de la composición de especies en un momento dado. En este escenario, el invasor hipotéticotiene un valor característico de función ecosistémica que es considerablemente más alto que la especie original representa-da por (d). A medida que progresa la invasión, el valor compuesto para la función del área varía, lo que se representa por lalínea en trazos. Si el invasor reemplazara a todas las especies a la misma tasa, entonces el valor promedio podría variar a lolargo de la línea recta en trazos desde (a) a (d). Sin embargo, si el invasor reemplazara preferencialmente a ciertas especiesmás que a otras, el valor competitivo variará desde la línea a-d, tal como lo representa la línea en trazos curvos. En algunasfunciones ecosistémicas existe un umbral (e), punto en el cual la variación desde el valor promedio inicial puede causarcambios irreversibles en el sistema (cf. Laycock 1991). Si la función es la inflamabilidad, por ejemplo, el área podríaexperimentar un incremento en las frecuencias de fuegos una vez que el umbral ha sido cruzado, lo cual puede llevar arápidos cambios en la composición comunitaria. Si estos cambios involucran la invasión adicional de otras especies, elvalor promedio de la función ecosistémica m-2 podría cambiar nuevamente, en algunos casos podría moverse fuera de loslímites definidos por las especies iniciales o incluso el invasor (véase las flechas y las líneas de trazos curvos).
feral pigs (Sus scrofa) may still contribute toerosion on the Channel Islands. Feral pig activityhampers regeneration of woody species such as thenative oak Quercus agrifolia (coast live oak) onSanta Cruz Island (Peart et al. 1994).
On the California mainland, the range andpopulation size of pigs has expanded since the1950s (Waithman et al. 1999). Feral pigs arenow the primary agents of soil disturbance insome California grasslands. Kotanen (1995)

415INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
found that pigs overturned 7.4 % of the surfaceof five Californian coastal meadows annually,changing the species composition and richnessof the grubbed areas. Pigs may increasesiltation of streams by turning up soil alongstreambanks and wallowing in the channel (Ray1988, Peart et al. 1994).
Mountain goats (Oreamnos americanus),which are native to some parts of Washington,were transported outside of their range to thatstate’s Olympic Mountains in the 1920s. Thegoats caused serious damage to nativevegetation and increased erosion in alpine areasof Olympic National Park, leading to thedevelopment of a goat population managementplan (Carlquist 1990).
We found no research on the effects of alieninvertebrates on the geomorphology of westernecosystems. It seems likely that some burrowinginvertebrate invaders have affected the movementof the substrate in which they live. For instance,since at least 1893, San Francisco Bay’s terracesand margins have been under attack from theisopod Sphaeroma quoyanum. This native ofAustralia and New Zealand riddles sediments andstructures with half-centimeter-wide burrows.Such burrows may increase erosion at the Bay’sedge (Cohen & Carlton 1995).
Plant invaders can affect geomorphology byaltering the stability of the substrate in which theylive. For instance, two species of introducedbeachgrass have slowed dune movement on thewest coast of United States. The most widespreadof these, Ammophila arenaria (Europeanbeachgrass), was imported from northern Europearound 1869 to stabilize dunes in San Francisco’sGolden Gate Park (Lamb 1898). Three factorsmay have contributed to A. arenaria’s subsequentcolonization of the majority of dunes on theUnited States’ Pacific coast. First, widespreadplanting of A. arenaria continued for a period of100 years (Wiedemann & Pickart 1996). Second,lateral growth of rhizomes allowed the grass tospread rapidly. Finally, living rhizome fragmentsmay have washed down the shore to colonize newsites (Wallén 1980).
Ammophila arenaria collects sand moreeffectively than the previously dominant nativegrass Leymus mollis (native dune grass; Barbouret al. 1985, Barbour & Johnson 1988), and itsinvasion has caused the rapid development ofsteep, continuous foredunes along the coast. Insome locations, foredunes have grown to aheight of 10 m (Wiedemann & Pickart 1996).These large foredunes, which border the upperbeach above the mean high tide line, may starveactive inland dune systems of sand, causingthem to become more static and allowing
vegetation to become established (Wiedemann &Pickart 1996). This phenomenon, in combinationwith the rapid spread of A. arenaria on thedunes themselves, has reduced the area of opendunes on the coast (Wiedemann 1984). Inaddition, the changes in foredune characteristicshave led to a new alignment of some inlanddunes. The dunes and swales inland of Leymusforedunes were oriented roughly perpendicularto the shore, but those inland of A. arenariaforedunes tend to run parallel to the coast(Barbour & Johnson 1988).
Wiedemann & Pickart (1996) argue that thePacific northwest coast has experienced periodsof slow foredune stabilization followed bystrong erosion events for thousands of years, andsuggest that although A. arenaria hasaccelerated sand stabilization, native plantswould also eventually cause the formation of anunbroken foredune. However, a native-dominated foredune might not attain the heightor strength of A. arenaria foredunes, and mightbe more rapidly eroded by waves during storms.
Another introduced species of beachgrassnow dominates the foredunes of southernWashington, and alternates with A. arenaria asthe dominant foredune species in the northernpart of the state (Seabloom & Wiedemann1994). This species is Ammophila brevigulata(American beachgrass), a native of the east coastand Great Lakes regions of North America. BothAmmophila species cause the formation of long,unbroken foredunes, but those formed by A.brevigulata are lower than those formed by A.arenaria (Seabloom & Wiedemann 1994).
As introduced beachgrasses reshape the westcoast’s dunes, introduced cordgrass species(Spartina spp.) stabilize sediments in itsestuaries. The most widespread alien cordgrassis Spartina alterniflora (smooth cordgrass). Thisnative of North America’s east coast now growsin San Francisco Bay, Suislaw Estuary inOregon, and in two locations in Washington(Daehler & Strong 1996). Spartina alternifloraoutcompetes the native cordgrass Spartinafoliosa in parts of San Francisco Bay where thetwo species co-occur (Callaway & Josselyn1992). The upper boundaries of these twoperennials are roughly the same, but S.alterniflora can colonize areas 9 to 20 cm belowthe lower limit of the native (Callaway &Josselyn 1992). This encroachment into lowertidal areas extends the marshlands and reducesmudflat area. The denser growth and thickerstems of S. alterniflora slow the tidal flow moreeffectively, causing suspended sediment toprecipitate and become trapped in the alien’sthick network of roots and rhizomes (Daehler &

416 DUKES & MOONEY
Strong 1996). In a study of a New Zealandestuary, Bascand (1970) found that areascolonized by S. alterniflora accumulated up to 5cm of sediment per year, while open mud flatstrapped little or no sediment. Sayce (1991)1,who has studied S. alterniflora in Washington,asserts that the invader can trap as much as 15cm of material annually. As sedimentaccumulates in formerly open areas, these areasmay rise above the intertidal zone. In someestuaries, sediment accretion and growth of S.alterniflora has restricted tidal channels andwaterways (Asher 1991). For example, theinvader colonized and threatened the flowcapacity of a major flood control channel in SanFrancisco Bay, leading to the initiation of aneradication program (Daehler 1996).
Three other introduced cordgrass species, S.anglica, S. densiflora, and S. patens, now growin estuarine ecosystems of the Pacific coast(Daehler & Strong 1996), and may cause similarchanges to those driven by S. alterniflora.Spartina anglica, which has invaded PugetSound in Washington, is particularly well knownfor its ability to rapidly colonize mudflats andaccrete sediment in European marshlands(Thompson 1991, Daehler & Strong 1996).
Exotic invaders also affect ripariangeomorphology. Shrubs of the genus Tamarixhave caused the most widespread changes.Recent estimates suggest that Tamarix spp.(tamarisk, salt cedar) has invadedapproximately 4,700 km2 of western UnitedStates floodplain (Zavaleta 2000). Blackburn etal. (1982) studied the impact of Tamarix spp.invasion on sedimentation processes in theBrazos River in Texas. As the phreatophyticshrub encroached onto formerly unoccupiedsandbanks along the river, i t stabilizedsediments and slowed water velocity. As waterslowed, sediment deposition increased and theriver channel narrowed. When channel sizes arereduced, flooding frequencies and flood levelsincrease. Tamarix has caused similar changesto the geomorphology of the Green River inCanyonlands National Park, Utah (Graf 1978).
Other exotic plant species may also increasesediment deposition, although the evidence isless solid. Arundo donax (arundo, giant reed)has invaded many waterways of southern andcentral coastal California (Hickman 1993,Dudley & Collins 1995). This tall perennialreed purportedly traps and stabilizes more
sediment than native vegetation, thusdecreasing channel sizes (Frandsen & Jackson1993). Arundo is also said to grow denselyenough to substantially reduce the carryingcapacity of small waterways (Robbins et al.1951). However, no data have been publishedto support these observations. In larger streamsand rivers, rafts of the reed can lodge againstnatural obstructions, bridges, or culverts,forming debris dams (Frandsen & Jackson1993). Arundo clogging is suspected to haveplayed a role in the bursting of a levee on theSanta Margarita river, which caused $12.5million dollars in damage to the CampPendleton military base (La Rue 1996). Mats ofSenecio mikanoides (German ivy) may alsoform debris dams in some Californiawaterways, redirecting water out of its channel(Chippin personal communication).
Another exotic plant species stabilizessediment in some Arizona streams. Duringfloods, mats of Cynodon dactylon(bermudagrass) protect streambanks fromerosion and shelter the basal fragments of nativeaquatic macrophytes. Recovery of the aquaticmacrophyte communities proceeds more rapidlyin these stabilized sites than in areas without C.dactylon (Dudley & Grimm 1994).
Since its introduction for landscapingpurposes, Cortaderia jubata (pampas grass) hascolonized disturbed areas such as erodingbanks, dry washes, cliffs, and logged redwoodforests throughout coastal California (Kerbavaz1985). Dense stands of the perennial must altererosion rates from invaded areas, but no studieshave quantified these changes.
Invading plants can also increase rates oferosion. The biennial forb Centaurea maculosa(spotted knapweed) is replacing nativebunchgrasses throughout many rangelands ofwestern North America (Roché & Roché 1988,Tyser & Key 1988, Lindquist et al. 1996).Lacey et al. (1989) found greater losses ofsediment and greater runoff from areasdominated by C. maculosa than frombunchgrass communities. The C. maculosacommunity’s larger fraction of bare groundmay explain these differences.
From the above examples it is clear that inthe case of both plants and animals, someinvaders accelerate the process of erosion, andothers stabilize substrates or trap sediments. Ofthe invasive plants that affect geomorphology,most slow erosion. Of the nonindigenousanimals, most accelerate erosion, especially onislands. In addition to the types of speciesmentioned above, some biological invadersinfluence geomorphic processes through their
1 SAYCE K (1991) Species displaced by Spartina in thePacific Northwest. In: Mumford TF Jr, P Peyton, JR Sayce& S Harbell (eds) Spartina workshop record: 26-27. Was-hington Sea Grant Program, Seattle, Washington, USA.

417INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
effects on disturbance regimes. For instance,alien plants that alter the fire frequency orintensity in an area also affect erosion becausefire reduces plant and litter cover (Swanson1981). We discuss invaders that alter fireregimes in the next section.
Fire regimes
Exotic grasses have replaced or currentlythreaten vast regions of western deserts andshrublands through their influence on fireregimes (D’Antonio & Vitousek 1992). Theinvading grasses occupy gaps between nativeplants in these sparsely vegetated systems,creating a continuous path of fine fuel thatpromotes the spread of fire. The grasspopulations tend to rebound quickly after fires,but many of the native perennials recoverslowly. Short fire-return intervals decimatepopulations of many shrubs and desertperennials. Thus, by accelerating firefrequencies, grass invaders can reduce thedensity of widely spaced perennials, turningshrublands to annual grasslands.
The most dramatic conversion of this kind isoccurring in the mixed-shrub steppe of Nevada,western Utah, southern Idaho, eastern Oregon,and eastern Washington. Many areas formerlydominated by Artemisia tridentata (bigsagebrush) and other shrubs are today coveredwith exotic annual grasses, primarily Bromustectorum (cheatgrass; Young & Evans 1978,Whisenant 1990, Billings 1994, Pellant & Hall1994) and Taeniatherum caput-medusae(medusahead, Young 1992). Mack (1981)chronicled the invasion of B. tectorum into theregion, and Whisenant (1990) documented theaccompanying change in fire-return intervals.Pinyon-juniper woodland ecosystems of theGreat Basin region have undergone a similarmetamorphosis due to anthropogenicdisturbance of native vegetation and the spreadof B. tectorum (Billings 1994).
Introduced grasses and forbs also threaten toaccelerate fire cycles in portions of the Mojaveand Sonora deserts. The most prevalent exoticannuals in these deserts are Bromus madritensisssp. rubens (red brome, syn. Bromus rubens) andB. tectorum, but B. trinii (Chilean chess),Schismus barbatus (Mediterranean grass), andErodium cicutarium (redstem filaree) are alsocommon in some areas (Brown & Minnich 1986,Hunter 1991, Rundel & Gibson 1996, Brooks1999). Hunter (1991) studied populations ofbrome grasses in the transition zone between theMojave and Great Basin deserts, and found thata series of wet years allowed B. madritensis to
become quite dense. At its peak in 1988, B.madritensis produced 34 g m-2, which was 97 %of that year’s total biomass production.Although exotic annuals are the most prolificinvaders of North America’s deserts, perennialspecies also pose a threat to the Sonoran desert.Cenchrus ciliaris (Buffel grass, syn. Pennisetumciliare) has been widely planted in northernMexico as a forage species for cattle, and wasplanted in the southwestern United States by theSoil Conservation Service and the ArizonaDepartment of Transportation (Pater personalcommunication). This southern Africanperennial can survive in a wide variety ofSonora desert microenvironments, and hasspread into many undisturbed areas (Burgess etal. 1991, Búrquez & Quintana 1994). Lehmannlovegrass (Eragrostis lehmanniana), anotherAfrican perennial that was once recommendedby the Soil Conservation Service, has alsospread from planted areas and become dominantin some areas of Arizona’s Sonora desert(Anable et al. 1992). Grass litter decomposesslowly in dry desert climates, maintaining acontinuous fuel load through years of lowbiomass production. Fires carried by grass litterthreaten populations of native annuals (Hunter1991) or long-lived perennials (Brown &Minnich 1986, Búrquez & Quintana 1994) in atleast three types of North American deserts.
Deliberate post-fire seeding and accidentalinvasion of non-native annuals into chaparraland coastal sage scrub may facilitate an increasein fire frequencies in some areas of southernCalifornia. Planted annuals such as the grassesLolium multiflorum (Italian ryegrass) and Vulpiamyuros (rattail fescue) and the mustardHirschfeldia incana, and invaders such asBromus madritensis and B. diandrus (ripgutbrome) have recently increased in dominance inthese ecosystems (Keeley 1995, Minnich &Dezzani 1998). These exotics persist throughfrequent fires more successfully than nativeshrubs such as Adenostoma fasciculatum(chamise), Ceanothus oliganthus, and Salviamellifera (black sage; Zedler et al. 1983, Keeley1995). A combination of increased firefrequencies, competition from introducedannuals, and other anthropogenic factors maydrive the replacement of chaparral and coastalsage scrub ecosystems by grassland in manyareas of southern California (Keeley 1995,Minnich & Dezzani 1998).
Fire-promoting exotic grasses also threatenriparian ecosystems. Arundo donax (arundo,giant reed) has invaded many waterways ofsouthern and central coastal California. The tallperennial reed quickly colonizes areas left bare

418 DUKES & MOONEY
from flooding, achieving dominance alongriverbanks and even in some estuaries (Dudley& Collins 1995). Arundo increases the fuel loadin riparian zones and provides an unbroken fuelcorridor along which fire can spread (Jackson1993, Scott 1993). Increased fire frequenciesmay prevent recovery of native plant speciesand purportedly changes the successional cycleof the cottonwood-willow riparian system,converting the vegetation to an Arundomonoculture (Bell 1993).
In western North America, most of theinvasive species that affect fire regimes aregrasses, and most of these species decreasefire-return intervals. Many native species,especially longer-lived plants in arid regions,cannot tolerate these frequent fires. Of all theways that invasive species modify ecosystemsin western North America, this impact on fireregime may have the most widespreadrepercussions for native species.
Hydrology
The invasion of exotic plants into NorthAmerican ecosystems has altered the hydrologyof vast areas of western North America.Invasions of Tamarix spp. have lowered watertables in many riparian zones of thesouthwestern United States (e.g., Neill 1983,Weeks et al. 1987). On a per-unit-leaf-areabasis, water loss of Tamarix is comparable tothat of native phreatophytes (Sala et al. 1996,Cleverly et al. 1997), so what characteristics ofTamarix lead to such great water loss? Twomechanisms have been proposed. First ,monospecific stands of the invasive shrub maydevelop a higher leaf area index (LAI) thanwould be found in native stands. Second,Tamarix stands tend to occupy a wider cross-section of the riparian zone than native stands(Sala et al. 1996).
Spanish colonists began altering thecomposition and hydrology of Californiagrasslands around 1769 when they introducedplants from Mediterranean Europe (Frenkel1970). Until that time, perennial grasses suchas Nassella pulchra (purple needlegrass)probably dominated most California grasslands(Heady 1988; but see Mooney et al. 1986).Under conditions of heavy grazing and possiblydrought, introduced annuals outcompeted mostof the original species (Hendry 1931, Mack1989, Rejmánek et al. 1991). The annualgrasses such as Avena spp. and Bromus spp.that now dominate extensive areas use less ofthe available deep water over the course of agrowing season than do native perennial
grasses, probably because they senesce earlierand have shallower roots (Holmes & Rice1996, Gerlach 2000). The excess water left byannual grasses may have created favorableconditions for Centaurea solstitialis (yellowstarthistle, Dyer & Rice 1999), a more recentinvader of these grasslands (Maddox &Mayfield 1985). Centaurea is a late-seasonannual that now draws deep soil moisture downto levels found under native perennial grasses(Gerlach 2000). Borman et al. (1992) observedsimilar soil moisture dynamics amongintroduced annual and native perennial grassesand C. solstitialis in southwestern Oregon.Hydrologic changes caused by grasslandinvaders may affect the establishment of nativewoody perennials (Da Silva & Bartolome 1984,Gordon & Rice 1993).
The replacement of native and naturalizedsystems by Eucalyptus spp. forests (Boyd 1985,Bulman 1988, Westman 1990) may havealtered the hydrology of large tracts ofCalifornia, although no studies havequantitatively documented these changes.Eucalyptus (primarily Eucalyptus globulus,blue gum) forests have replaced many differentecosystem types in California. These forestsprobably altered hydrology most drasticallywhere they replaced grassland. Eucalyptusroots grow much deeper than those of grasslandspecies (Canadell et al. 1996), and extractwater from lower in the soil profile. Eucalyptusglobulus is the only widespread woody alienknown to transport deep soil moisture toshallower layers through hydraulic lift (Dawsonpersonal communication). It is not knownwhether this process eases drought stress fornearby shallow-rooted plants, as occurs aroundother hydraulic l ifters (Dawson 1993).Evergreen eucalypts transpire year-round, butCalifornia’s grasslands are mostly dormant inthe summer. Although Eucalyptus forestsprobably transpire more water than grasslandson an annual, per-area basis, the forests havegreater surface roughness and deeper litterlayers than grasslands (Poore & Fries 1985,Robles & Chapin 1995), and may lose lesswater to surface evaporation. On balance,eucalypt forests probably extract more waterfrom the ground than California grasslands.Such a difference has been observed in SouthAfrica, where Van Lill et al . (1980)documented a dramatic reduction in runofffrom a grassland after its conversion to an E.grandis plantation. In areas where Eucalyptusforests have replaced native forests orwoodlands, alterations to local hydrology wereprobably less drastic.

419INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
Replacement of native perennial vegetationby Bromus tectorum in western shrublands (seeabove) reduced rooting depths and shortenedthe period when plants in these systems arephotosynthetically active. As a consequence,annual evapotranspiration has declined in someof these systems (Cline et al. 1977, Kremer &Running 1996).
Although relatively few studies havecompared the hydrology of invaded and pristineplant communities in western North America,these few studies have examined changescaused by some of the most widespread speciesthat are likely to have effects. Two aliengenera, Tamarix and Eucalyptus, probablyincrease water use rates beyond what theinvaded ecosystems experienced previously.The invasion of California’s alien-dominatedannual grasslands by Centaurea solstitialis maymove the hydrological cycle closer to a pre-European settlement dynamic. In some parts ofwestern North America, the invasion of annualgrasses has reduced plant water use, primarilyby reducing the abundance of deeply rootedspecies.
Climate and microclimate
Biological invasions have altered moisturetransport and land surface characteristics of alarge portion of western North America (seeabove). However, the extent and implicationsof these changes are poorly understood. Recentstudies have indicated that changes invegetation types can alter local or regionalclimatic patterns (Lean & Warrilow 1989,Shukla et al. 1990, Chase et al. 1999, Hoffman& Jackson 2000). While it seems possible thatsome invaders of western North America,particularly the annual grasses, have altered theland surface characteristics and hydrology ofsufficient area to affect the regional climate,this hypothesis has not yet been tested.
Plant invaders can also alter the microclimateof invaded areas. For instance, dense stands ofAmmophila arenaria sharply reducetemperatures and available light at theunderlying surface of the Pacific coast’s dunesrelative to stands of the native grass Leymusmollis (Barbour et al. 1985). Spartinaalterniflora may similarly reduce light levelsunder the plant canopy of marshes, which coulddepress estuarine algal production (Callaway &Josselyn 1992). Soil temperature, soil moisture,and light conditions under the plant canopyaffect the germination and establishment successof plants (Evans & Young 1970, Evans & Young1972), and the suitability of habitat for animals.
Invasive species have undoubtedly alteredmicroclimates in many other ecosystems, butwe did not find studies that documented thesechanges.
Composition of the atmosphere
Biological invaders can alter the flux of gasesbetween the land surface and the atmosphere.In the United States, annual emissions ofvolatile organic compounds (VOCs) fromvegetation equal or exceed anthropogenicemissions (Guenther 1997), although thebiogenic output is more evenly distributedacross the landscape. Because vegetationcontributes such a great proportion ofatmospheric VOCs, and because plant speciesvary widely in their rate of VOC emission(Evans et al. 1982, Winer et al. 1992, Arey etal. 1995), regional atmospheric VOC poolsdepend largely on the species composition oflocal plants. Invasive species that overrun largeareas can alter regional VOC emissions andatmospheric composition (Monson et al. 1995),with important consequences for atmosphericchemistry and air quality (Mooney et al. 1987).Air quality of the west coast may have beenadversely affected by the introduction ofEucalyptus globulus and Arundo donax, whichemit high levels of isoprene relative to manynative species (Evans et al. 1982, Hewitt et al.1990, Arey et al. 1995).
The invasion of plants of one growth forminto a region dominated by another may alterthe local rate of CO2 uptake and storage. Forinstance, the replacement of shrublands andpinyon-juniper woodlands by annual grasslandsprobably reduces long-term carbon storage inbiotic pools. Conversely, replacement ofgrassland with Eucalyptus globulus or otherwoody species may increase both biotic carbonstorage and net primary productivity (NPP,Robles & Chapin 1995).
Invaders may also alter the emission ofNOx, N2O, NH3, and CH4 from the landscape.Chatigny et al. (1996) found evidence that thespecies composition of a plant communityaffects local rates of nitrification anddenitrification, which in turn moderate theemission of nitrogenous gases by the microbialcommunity (Schlesinger 1991). By creatingponds, beaver (Castor canadensis) cansubstantially increase methane emissions froman area (Yavitt et al. 1992). Naiman et al.(1991) estimated that North America’sexpanding beaver population has contributed 1% of the recent rise in atmospheric methane.Methane release rates from wetlands also

420 DUKES & MOONEY
depend on the biomass and structural propertiesof the inhabitant vascular plants (Sebacher etal. 1985, Schimel 1995, Verville et al. 1998).Invaders such as Lepidium latifolium (perennialpepperweed) and Lythrum salicaria (purpleloosestrife) change the amount and compositionof wetland vegetation in western NorthAmerica, and may alter regional methaneemission, although this remains unstudied.
Plant invaders can also affect atmosphericcomposition by altering fire frequencies.During fires, carbon, nitrogen and otherelements enter the atmosphere throughgasification, volatilization, and convection(Christensen 1994). However, the contributionof exotic grass-fueled fires to changes inatmospheric composition is estimated to besmall (D’Antonio & Vitousek 1992).
The effects of biological invaders on thecomposition of the atmosphere remain largelyunstudied. Although these effects are probablysmall at the global and regional scales, theymay in some cases (such as near large eucalyptforests, in the case of VOCs) be locallyimportant.
Nutrient cycling and soil chemistry
Non-native plants and animals can alterecosystem nutrient cycling and soil chemistrythrough a number of mechanisms. Nitrogen-fixing invaders increase the rate of N input to asystem when they replace non- or less-efficiently fixing plants, and when theycolonize open areas. Vitousek & Walker (1989)and Vitousek et al. (1987) found that theinvasion of an N-fixing plant into a youngecosystem in Hawaii increased the rate ofecosystem N accumulation more than fourfold.
Horticulturists have introduced manyspecies of leguminous European shrubs to thewestern United States, including gorse Ulexeuropaea, and the brooms Cytisus scoparius,Genista monspessulana and Spartium junceum.The nitrogen-fixing capacity of brooms hasstimulated research into their potential as yieldenhancing understory shrubs in commercialPseudotsuga menziesii (Douglas-fir)plantations. Studies have focused on C.scoparius, which fixes nitrogen year-roundunder mild conditions, albeit at relatively lowlevels (Wheeler et al. 1979, Wheeler et al.1987). Helgerson et al. (1979) integrated ayear’s worth of nitrogenase activitymeasurements on a young stand of broom inOregon, and estimated an annual fixation rateof 35 kg N ha-1 year-1. This value itselfrepresents a substantial input, but because this
method of estimation is imprecise, the actualfixation rate could be twice as high (Wheeler etal. 1987). In addition to the brooms and gorse,several leguminous annual and perennial herbssuch as Medicago polymorpha (burr medic),Melilotus alba (white sweetclover), andTrifolium hirtum (rose clover) have invadedWestern ecosystems (Hickman 1993). It is notknown whether all of the invasive legumesactively fix N.
Nitrogen inputs to a system from N-fixingalien plants may be constrained by thecompatibili ty of the plants with localsymbionts. Absence of a compatible Rhizobiumstrain could explain the low nodulation on C.scoparius roots observed by Wheeler et al.(1987) in Oregon and Scotland, although acidicsoil conditions or other factors could also havelimited nodulation.
Some plant invaders may decrease nitrogeninputs in their vicinity by leaching chemicalsthat reduce the ability of other species to fix N(Rice 1992). In glasshouse and pasture studiesin New Zealand, Wardle et al. (1994) foundevidence that decomposing leaves of theinvasive thistle Carduus nutans inhibit nitrogenfixation by Trifolium repens. Carduus nutanshas invaded many areas of western NorthAmerica, but no published studies haveexamined whether this thistle adversely affectsgrowth and nitrogen fixation of native legumes.
Both N-fixing and non-fixing plants directlyaffect the nutrient retention of ecosystems bymoderating erosion of nutrient-rich topsoil, andby sequestering available soil nutrients, thusreducing leaching losses. Gholz et al. (1985)found that the invasive annual Seneciosylvaticus took up a large fraction of thenutrients released from unburned clear-cutstands of old-growth Douglas-fir in Oregon.Invaders can also indirectly modulate N lossesby influencing soil moisture and nitrate levels,which can constrain denitrification rates. Wefound no studies of invasive plants thatexamined indirect effects on nutrient retention.
Although fires increase short-term N-availability in a system, frequent fires generallycause long-term loss of N (Ojima et al. 1994),depending on grazing practices (Hobbs et al.1991). Alien species that accelerate fire cycles(such as those mentioned above) couldeventually increase N losses from ecosystems.
Plant species strongly influence the rate atwhich nutrients cycle within an ecosystemthrough litter-quality feedbacks (Wedin &Tilman 1996, Evans et al. 2001; for review seeHobbie 1992). The invasion of a species withrapidly decomposing litter into an ecosystem

421INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
dominated by plants with slow-decomposinglitter will accelerate net nutrient mineralizationin the system (Van Vuuren et al. 1992, VanVuuren & Berendse 1993, Van Vuuren et al.1993). For instance, leaves of the notoriouswetland invader Lythrum salicaria (purpleloosestrife) have higher phosphorus (P)concentrations and decompose more quicklythan shoots of native Typha spp. (Emery &Perry 1996). These characteristics will forcechanges in the nutrient dynamics of invadedwetlands that may accelerate eutrophication ofdownstream water bodies.
Introduced detrit ivores also alterecosystems’ internal nitrogen cycling. InKansas’ tallgrass prairie, James (1991)observed that the invasion of Europeanearthworms has decreased nutrientmineralization and soil turnover rates. At least45 species of exotic earthworms have beenintroduced to North America north of Mexico(Reynolds 1995). These species have beenparticularly successful in disturbed habitats,and also dominate some wildland habitatsincluding southern California chaparral andriparian zones (Kalisz & Wood 1995). It isunclear how detritivore invasions have affectedthese ecosystems.
A few of western North America’s mostinvasive plants release compounds that alter thesoil’s nutrient availability or suitability forother species of plants. For instance, theubiquitous tumbleweed Salsola tragus (syn. S.iberica) releases oxalate in leachate from itscanopy and litter (Cannon et al. 1995). Theleached oxalic acid increases phosphorus (P)availability in the soil by solubilizing it fromthe pool of inorganic-bound soil P. Otherwestern invaders such as Halogeton glomeratusand some plants in the Oxalidaceae probablyaffect P availability similarly, as they alsoproduce high concentrations of oxalic acid(Kingsbury 1964, Whitson et al. 1996).
The iceplant Mesembryanthemumcrystallinum exploits its high salt tolerance tooutcompete native species in coastal areas ofCalifornia. This South African annualstockpiles salts in living tissue. Once the tissuehas senesced, rainfall and fog drip leach thesalts out and deposit them on the soil surface.The high concentrations of salt that accumulatearound populations of this grassland invaderexclude competitors through osmoticinterference (Vivrette & Muller 1977).
Another iceplant that plagues California’scoastal plant communities, Carpobrotus edulis,modifies soil in a different way. This rapidlyspreading succulent acidifies the soil around its
roots (D’Antonio 1990), through an as yetunstudied mechanism. Investigations inEngland suggest that the common west coastinvader Ulex europaea may affect soilssimilarly (Grubb et al. 1969, Grubb & Suter1971). Changes in soil pH can influence thedominance of different plant species in oldfields (Tilman & Olff 1991) and montaneforests (Goldberg 1985).
Other introduced species release compoundsthat can inhibit their own growth, as well asthat of competitors. Phytotoxic chemicals thatleach from the leaves and litter of eucalyptsduring rainfall and fog drip events can directlyinhibit germination and retard seedling growthof grasses, as well as of the eucalyptsthemselves (del Moral & Muller 1969, delMoral & Muller 1970).
Nutrient cycles and soil properties aresubject to change by many of western NorthAmerica’s invasive species. The mostwidespread change may be an increase in Ninputs from nonindigenous legumes. However,few researchers have studied the amount ofatmospheric N fixed by these species. Exoticearthworms may also have caused importantand widespread changes in nutrient cycles, butthese changes remain unstudied.
Productivity and decomposition
An ecosystem’s live biomass (LB) and NPPmay respond to the addition of a species withnovel traits. For instance, invaders that accessor use existing resources more completely orefficiently than native plant species, or thatproduce more readily mineralizable litter thannative species, may cause increases in LB andNPP. Examples of exotic species that accesspreviously untapped water and nutrient storescan be found in previous sections (see sectionson hydrology and nutrient cycling). Invadersthat represent a new life form or that eliminatea prominent life form may also alter an area’sLB and NPP. Unfortunately, relatively fewstudies of invaders in western North Americahave included data on these basic ecosystemproperties.
Along the edges of San Francisco Bay,invading Spartina alterniflora produces six toseven times as much aboveground biomass perunit area as the native cordgrass Spartinafoliosa , and 1.6 to 3.2 times as muchbelowground biomass (Callaway & Josselyn1992). The great aboveground production of S.densiflora, deposited as wrack on the uppermarsh in Humbolt Bay, California, smothersnative marsh species, opening space for further

422 DUKES & MOONEY
S. densiflora invasion (Daehler & Strong1996).
Along the California coast, Ammophilaarenaria-dominated beach communities haveup to three times as much standing biomass asnative-dominated communities (Barbour &Robichaux 1976, Pavlik 1983a). The differencein aboveground biomass of stands of A.arenaria and the native perennial grass Leymusmollis stems from A. arenaria’s higher nitrogenuse efficiency, greater allocation of nitrogenand photosynthetic assimilate to leaf blades,different architecture, and slower leafsenescence (Pavlik 1983a, 1983b, 1983c).
Robles & Chapin (1995) compared adjacentexotic-dominated annual grassland andeucalypt-covered sites in the San Francisco Bayarea. Annual aboveground production ofEucalyptus globulus forests was more thantwice that of grassland, and the layer of slow-decomposing E. globulus litter had grown ninetimes thicker than the litter layer of thegrassland.
Growth and decomposition rates of primaryproducers can be affected by organisms onother trophic levels. Plant pathogens that attackone of the dominant species in an ecosystemcan, at least temporarily, lower the system’sproductivity and live biomass. At least threeexotic fungi are causing widespread damage towestern trees, and must have temporarilylowered the productivity of some forests. Thefungal pathogen Fusarium subglutinans f. sp.pini, endemic to the southeastern United States,causes pitch canker disease in a number ofconiferous tree species (Storer et al. 1994). Thedisease appeared in California in 1986 (McCainet al. 1987), and spread rapidly, killing offPinus radiata stands along much of the coast.The disease has now reached all three ofCalifornia’s relictual P. radiata stands (Gordonet al. 1997), and may eventually infect as manyas 85 % of the trees in these stands (Woodpersonal communication). Pitch canker has alsobeen found in a native Pinus attenuata standnear Mendocino (Storer et al. 1994).
The fungus Cronartium ribicola, whichcauses white pine blister rust, has infectedpines in the Cascades, Rocky Mountains, andthe Sierra Nevada (Kinloch & Dulitz 1990).Growth of the pathogen can rapidly girdle andkill shoots of pines in the subgenus Strobus(white pines), or lead to their attack byDioryctria spp. larvae. White pine blister rustepidemics generally lead to the loss of allinfected seedlings and saplings, and the deathof many adult trees (Kinloch & Dulitz 1990,Tomback et al. 1995).
A third imported fungus has attacked theroots of Port-Orford-cedar (Chamaecyparislawsoniana) throughout its native range insouthwestern Oregon and northwesternCalifornia. This fungus, Phytophthora lateralis,probably arrived from Asia on ornamentalplants sometime in the 1920s. A field survey ofthree infested drainages found 46 % mortalityof Port-Orford-cedar and 10 % mortality ofanother native, Pacific yew (Taxus brevifolia,Murray & Hansen 1997).
The recent invasion of Asian clams(Potamocorbula amurensis) has probablyincreased consumption rates of bacterioplanktonand phytoplankton in San Francisco Bay. Theseclams, which can reach densities as high as10,000 individuals m-2 (Carlton et al. 1990),filter the water column more than once per dayin deep waters, and as much as 13 times per dayin shallow waters (Werner & Hollibaugh 1993).This rapid filtration equals or exceeds planktonicgrowth rates, and may affect the standing crop ofplankton and intensity of the annual spring algalblooms in San Francisco Bay.
Although relatively few studies havecompared the productivity of invaded anduninvaded communities in western NorthAmerica, it seems likely that most invasiveplants have increased resource use of invadedcommunities, and also increased ecosystem-level productivity. Invasive species that alterdisturbance regimes or otherwise eliminateother life forms from the community may bethe most common exceptions to this trend. Theeffects of invasive animals on primaryproductivity are rarely examined, and we canonly speculate that these species have hadrelatively little effect on the productivity ofmost ecosystems, excepting the case of Asianclams in San Francisco Bay.
Native species
Although biological invaders add to the speciesrichness of an area upon their arrival, some caneventually cause the decline or even extinctionof native species through predation,competition, disease, or replacement ofresource species. In fact, the spread ofbiological invaders is generally regarded to bethe second greatest agent of speciesendangerment and extinction after habitatdestruction (Wilcove et al. 1998). As a generalrule, native populations are more likely to beimpacted by invaders if they are in isolatedsystems such as on islands, in lakes or instreams than if they are on the mainland(D’Antonio & Dudley 1995).

423INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
Introduced fish and amphibians havesuppressed native fish and amphibianpopulations in the majority of lakes and riversin western North America. Non-nativebullfrogs (Rana catesbeiana) prey on andcompete with yellow-legged frogs (Ranaboylii) (Moyle 1973, Kupferberg 1997), andintroduced predatory fish appear to be a factorin the decline of mountain yellow-legged frogs(Rana pipiens) in Yosemite National Park(Drost & Fellers 1996). Bullfrogs and exoticfish both may have contributed to the decline inred-legged frog (Rana aurora) populations(Kiesecker & Blaustein 1998, Adams 2000).
Nonindigenous fish species are dominantthrough most of the San Joaquin river drainage,and the entire Colorado river drainage, wheremost of the native fish species are listed asthreatened or endangered (Moyle 1986). Theexotic protozoan Myxobolus cerebralis, thecausative agent of whirling disease, is likelyresponsible for recent declines in populationsof the rainbow trout (Oncorhynchus mykiss)(Nehring & Walker 1996, Bergersen &Anderson 1997). However, in many othercases, the decline of native fish can be directlyattributed to competition with or predation byfish species that were introduced forsportfishing. Such is the case with theendangered razorback sucker (Xyrauchentexanus) in the Colorado river basin (Minckleyet al . 1991). There, heavy predation byintroduced fish on larval razorback suckersprevents the regrowth of native populations.Some suspect that the thicktail chub (Gilacrassicauda), a native fish species that oncepopulated the Sacramento-San Joaquin delta,was extirpated by the predation of introducedlargemouth bass (Micropterus salmoides) andstriped bass (Morone saxatilis , Cohen &Carlton 1995). Although non-native fishspecies have depressed populations of nativefauna in many western rivers, globalextinctions such as that of the thicktail chub arerare (Moyle & Light 1996).
Just downstream from the thicktail chub’sformer habitat lies one of the world’s mostbiologically polluted ecosystems, the SanFrancisco Bay estuary (Cohen & Carlton 1998).Invaders have relegated native species toobscurity in much of this isolated system. Themost abundant invader is probably the Asianclam (Potamocorbula amurensis). This smallbivalve blankets sediments in many regions ofthe bay (see above), precluding theestablishment of other benthic organisms(Carlton et al. 1990, Cohen & Carlton 1995).Another invader, the mudsnail Ilyanassa
obseleta, has usurped much of the formerhabitat of Cerithidea californica, relegating thenative mudsnail to the estuary’s highly salinemargins (Race 1982). Advancing across themudflats, Spartina alterniflora has reducedfeeding habitat for many species of shorebirds(Callaway & Josselyn 1992). In creeks thatfeed into San Francisco Bay, two species ofintroduced crayfish (Orconectes virilis andPacifastacus leniusculus) may have contributedto the extinction of the native sooty crayfish(Pacifastacus nigrescens). These invaders maynow be factors in the decline of the Shastacrayfish (Pacifasctacus fortis) in other regionsof California (Light et al. 1995).
At one time, 17 of 19 threatened orendangered plant species on California’sChannel Islands were imperiled by exoticspecies, primarily feral animals (D’Antonio &Dudley 1995). Feral animal disturbance alsolowered soil mite diversity (Bennett 1987).Removal of livestock and feral fauna may haveallowed regrowth of threatened populations onsome of these islands, but Foeniculum vulgare(fennel) and other invasive plants have alsobenefited from the reduction in herbivory(Beatty & Licari 1992, Brenton & Klinger1994). Persistent alien plants may now pose thegreatest risk to some of the Channel Islands’beleaguered native plant populations.
Although native populations in isolatedsystems are at the greatest risk from biologicalinvaders, native species in mainland terrestrialecosystems can also be affected. Somedramatic instances have been mentioned in theabove sections (e.g., displacement of westernshrublands and California’s native grasslandcommunity by introduced annual grasses,attack of California’s relictual Pinus radiatastands by the pitch canker fungus Fusariumsubglutinans, among others); we add andexpand on a few examples here.
Argentine ants (Linepithema humile) havebeen introduced to every continent exceptAntarctica (Hölldobler & Wilson 1990), andhave spread to much of western North America,including California, Arizona, southernNevada, and Mexico (Wheeler & Wheeler1986). The invaders displace native antcolonies in California (e.g., Ward 1987, Human& Gordon 1996, Human & Gordon 1997), andpossibly throughout the zone of invasion. Thedisplacement of native seed-burying ants byArgentine ants has reduced the establishment ofsome native shrubs in South Africa (Bond &Slingsby 1984, Slingsby & Bond 1985), andmay affect plant species distributions inCalifornia grasslands as well (Human 1996).

424 DUKES & MOONEY
Argentine ants are known to tend aphids andscale insects; the ecological consequences ofthis behavior have yet to be investigated(Human 1996).
Non-native eastern gray (Sciuruscarolinensis) and fox (S. niger) squirrels havedeveloped large populations in and aroundCalifornia’s suburban environments, particularlyin the San Francisco Bay area. Native westerngray squirrels (S. griseus) have been displacedfrom some areas where they overlapped with theintroduced species, but they maintain dominancein xeric sites (Byrne 1979).
Physical disturbance by feral burros (Equusasinus) once caused contamination of watersources and local elimination of native plantspecies in Death Valley, California. Theseburros may have once contributed to thedecline of native bighorn sheep (OvisCanadensis , McMichael 1964 as cited byWoodward 1976), but the population of burroshas now been effectively controlled (Loope etal. 1988).
According to Vuilleumier (1991), most ofMediterranean California’s non-native birdspecies have only small populations or arerestricted to urban environments, and areunlikely to adversely impact native species.However, some of the state’s most abundantbird species were introduced from elsewhere,including the European starling (Sturnusvulgaris), house sparrow (Passer domesticus),and rock dove (Columbia livia). Starlings arethought to have the most negative impact onnative bird species, as they occupy the nestsites of other cavity-nesting birds. Thisbehavior may be contributing to the decline ofthe purple martin (Progne subis) in California(Small 1994).
Exotic plant invasions can also reduce theamount of habitat available for native birds. Aninvasion of Tamarix spp. at Eagle Borax Springin Death Valley, California, caused a largemarsh to dry up, reducing habitat for migratorybirds (Neill 1983). Subsequent removal of theinvader led to the return of surface water andwildlife. Along western North American rivers,stands of Tamarix spp. (Cohan et al. 1978,Hunter et al. 1988) and Elaeagnus angustifolia(Russian-olive, Knopf & Olson 1984) support amore depauperate avian fauna than do nativestands. Government-subsidized replacement ofcottonwood-dominated riparian vegetation withinvasive Russian-olive may have reducedhabitat for cavity-nesting birds (Olson & Knopf1986). Invasions of Lythrum salicaria (purpleloosestrife) are thought to have degradedwetland habitat for waterfowl and other
wildlife throughout much of North America(Thompson 1987) although many claims fordamage wrought by this species are as yetunsubstantiated (Anderson 1995, Hager &McCoy 1998). In the Sonora desert, areasdominated by the introduced perennial grassEragrostis lehmanniana provide undesirablehabitat for scaled quail (Callipepla squamata,Medina 1988).
In addition to providing poor foraging forquail, Eragrostis-dominated desert supports amore depauperate faunal community thannative-dominated areas (Bock et al. 1986).Similarly, Pacific coast dunes dominated by theintroduced beachgrass Ammophila arenariasupport fewer species of arthropods(Slobodchikoff & Doyen 1977) and plants(Barbour et al. 1976, Boyd 1992) than native-dominated dunes. The Eurasian perennial herbEuphorbia esula (leafy spurge), which as of1997 infested more than 110 km2 in the UnitedStates and Canada (including parts of Idaho,Montana, Wyoming, and Colorado, Lajeunesseet al. 1999), decreases habitat quality for bison(Bos bison) and deer (Odocoileus spp.) inTheodore Roosevelt National Park, NorthDakota (Trammell & Butler 1995).
Invasive exotic plants can threatenpopulations of rare native plant species, but fewcases have been documented. Some findings:on Montana rangeland, introduced Centaureamaculosa can reduce recruitment andpopulation growth of the rare native Arabisfecunda (Lesica & Shelly 1996). Phalarisarundinacea (reed canarygrass), which hasgenotypes native to both northern NorthAmerica and northern Europe, appears to havedisplaced the endangered aquatic annualHowellia aquatilis (Lesica 1997) from parts oftwo Montana marshes. In central New Mexico,habitat for the endemic thistle Cirsiumvinaceum, a federally listed threatened species,is being taken over by the Eurasian biennialDipsacus fullonum (teasel). Studies byHuenneke & Thompson (1995) suggest that thenative thistle could decline if the Dipsacusinvasion continues unchecked.
Plant invaders can also depress fungalcommunities. Allen et al. (1995) report lowerfungal diversity and colony numbers underintroduced annual grasses and forbs comparedwith neighboring coastal sage habitat. In theintermountain west, most native plant speciesare mycorrhizal, whereas some invaders are not.When nonmycorrhizal species such as Salsolatragus invade rangeland in this region,populations of vesicular-arbuscular mycorrhizaedecline (Goodwin 1992).

425INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
Although we found many reports of declinesof native biodiversity following invasions, wealso found some cases where native speciespreferentially made use of habitat created byintroduced species. In these cases theintroduced species were generally replacinghabitat that had formerly been provided bynative species. For instance, monarchbutterflies (Danaus plexippus) that overwinteron the west coast of United States mostcommonly roost in groves of Eucalyptusglobulus. For the monarchs, this introducedspecies probably replaces habitat that was lostwhen groves of native trees were logged in thelate 1800s (Lane 1993). However, the eucalyptsmay not provide suitable replacement habitatfor all the species that had used the nativegroves.
It is clear from the above examples thatinvasive species pose significant threats tonative populations. A handful of the mostsuccessful invasive species has contributed todeclines in populations of several native speciesthrough competition, predation, and habitatalteration. Native populations on islands or inisolated systems such as creeks and estuariesseem much more likely to be impacted byinvasive species than mainland populations.Considering the number of alien species thathave become established in mainland systems, itseems that a minority of these invaders haswidespread and adverse impacts on nativespecies. However, effects of the vast majority ofmainland invaders remain unstudied.
CONCLUSIONS
A small number of biological invaders havedrastically changed the structure andfunctioning of ecosystems in western NorthAmerica, and thousands of other invaders havewrought more subtle changes. The ecologicaldisruption caused by invasive species, incombination with other factors (including landdevelopment and elements of global changesuch as N deposition, climate change, amongothers), threatens to drive species extinctionsand to reduce the dominance of natives in manyecosystems, altering the character of much ofwestern North America (Fig. 1).
Many of western North America’sgrassland, shrubland, dune, riparian, andestuarine ecosystems are already dominated,probably irreversibly, by non-native species.Some of California’s remaining coastal scrub isundergoing annual grass invasion, and some ofthe state’s chaparral faces threats from
management practices that encourage annualgrass dominance. The forests of the west coasthave escaped threats from biological invadersmore successfully than other systems.However, the current declines of Port-Orford-cedar, relictual stands of Pinus radiata, andfive-needled pines in the Sierra Nevada, all dueto introduced fungi, highlight the potentialvulnerability of these ecosystems.
We know very little about the impacts ofmost biological invaders on native species andon ecosystem functioning (Levine et al. 2003).Even some potentially dramatic impacts of themost widespread invasions have yet to bestudied. For example, the large-scalereplacement of mixed-shrub steppe with annualgrasslands must have affected the energy andwater balance of the intermountain west, andthus may have affected regional weatherpatterns. Transformations such as this wouldhave important ecological and economicconsequences, and should be examined. We doknow that some invaders are causing importantecological changes. If we observe and quantifythese changes, and we identify viableecosystem restoration strategies, then oursociety (and its land managers) will be betterable to make informed decisions about whichof these invaders are noxious enough to meritlarge-scale eradication campaigns.
ACKNOWLEDGMENTS
We thank J. Canadell, C. D’Antonio, G. Joel, J.Polsenberg, J. Randerson, C. Still, J. Vervilleand an anonymous reviewer for helpfulcriticism of drafts of this manuscript. We aregrateful to Marta Berrocal-Lobo for help withSpanish translations. J.S.D. received supportfrom a NASA Earth System ScienceFellowship.
LITERATURE CITED
ADAMS MJ (2000) Pond permanence and the effects ofexotic vertebrates on anurans. EcologicalApplications 10: 559-568.
ALLEN MF, SJ MORRIS, F EDWARDS & EB ALLEN(1995) Microbe-plant interactions inMediterranean-type habitats: Shifts in fungalsymbiotic and saprophytic functioning in responseto global change. In: Moreno JM & WC Oechel(eds) Global change and Mediterranean-typeecosystems: 287-305. Springer-Verlag, New York,New York, USA.
ANABLE ME, MP MCCLARAN & GB RUYLE (1992)Spread of introduced Lehmann lovegrass Eragrostislehmanniana Nees. in southern Arizona. BiologicalConservation 61: 181-188.

426 DUKES & MOONEY
ANDERSON MG (1995) Interactions between Lythrumsalicaria and native organisms: a critical review.Environmental Management 19: 225-231.
AREY J, DE CROWLEY, M CROWLEY, M RESKETO &J LESTER (1995) Hydrocarbon emissions from na-tural vegetation in California’s south coast airbasin. Atmospheric Environment 29: 2977-2988.
ASHER R (1991) Spartina introduction in New Zealand.In: Mumford Jr. TF, P Peyton, JR Sayce & SHarbell (eds) Spartina workshop record: 23-24.Washington Sea Grant Program, Seattle, Washing-ton, USA.
BARBOUR MG, TM DE JONG & AF JOHNSON (1976)Synecology of beach vegetation along the PacificCoast of the United States of America: a firstapproximation. Journal of Biogeography 3: 55-69.
BARBOUR MG, TM DEJONG & BM PAVLIK (1985)Marine beach and dune plant communities. In:Chabot BF & HA Mooney (eds) Physiologicalecology of North American plant communities:296-322. Chapman and Hall, New York, New York,USA.
BARBOUR MG & AF JOHNSON (1988) Beach and dune.In: Barbour MG & J Major (eds) Terrestrialvegetation of California: 223-261. California NativePlant Society, Sacramento, California, USA.
BARBOUR MG & RH ROBICHAUX (1976) Beachphytomass along the California coast. Bulletin ofthe Torrey Botanical Club 103: 16-20.
BASCAND LD (1970) The roles of Spartina species inNew Zealand. Proceedings of the New ZealandEcological Society 17: 33-40.
BEATTY SW & DL LICARI (1992) Invasion of fennel(Foeniculum vulgare) into shrub communities onSanta Cruz Island, California. Madroño (UnitedStates) 39: 54-66.
BELL GP (1993) Biology and growth habits of giant reed(Arundo donax). In: Jackson NE, P Frandsen & SDuthoit (eds) Proceedings of the Arundo donaxworkshop, Ontario, California: 1-6. California ExoticPest Plant Council, Berkeley, California, USA.
BENNETT SG (1987) The effects of feral animals on soilmites recovered from Catalina ironwood groves(Lyonothamnus floribundus) on Santa CatalinaIsland, California. In: Hochberg FG (ed) ThirdCalifornia islands symposium: recent advances inresearch in the California islands: 155-170. SantaBarbara Museum of Natural History, Santa Barbara,California, USA.
BERGERSEN EP & DE ANDERSON (1997) Thedistribution and spread of Myxobolus cerebralis inthe United States. Fisheries 22: 6-7.
BILLINGS WD (1994) Ecological impacts of cheatgrassand resultant fire on ecosystems in the westernGreat Basin. In: Monsen SB & SG Kitchen (eds)Proceedings in ecology and management of annualrangelands: 22-30. United States Department ofAgriculture, Forest Service Intermountain ResearchStation, Ogden, Utah, USA.
BLACKBURN WH, RW KNIGHT & JL SCHUSTER(1982) Saltcedar influence on sedimentation in theBrazos River. Journal of Soil and WaterConservation 37: 298-301.
BOCK CE, JH BOCK, KL JEPSON & JC ORTEGA(1986) Ecological effects of planting Africanlovegrasses in Arizona. National GeographicResearch 2: 456-463.
BOND W & P SLINGSBY (1984) Collapse of an ant-plantmutualism: the Argentine ant (Iridomyrmex humilis)and myrmecochorous Proteaceae. Ecology 65:1031-1037.
BORMAN MM, DE JOHNSON & WC KRUEGER (1992)Soil moisture extraction by vegetation in aMediterranean/marit ime climatic regime.Agronomy Journal 84: 897-904.
BOYD D (1985) Status report on invasive weeds:Eucalyptus. Fremontia (United States) 12: 19-20.
BOYD RS (1992) Influence of Ammophila arenaria onforedune plant microdistributions at Point ReyesNational Seashore, California. Madroño (UnitedStates) 39: 67-76.
BRENTON B & R KLINGER (1994) Modeling theexpansion and control of fennel (Foeniculumvulgare) on the Channel Islands. In: Halvorson WL& GJ Maender (eds) The fourth California islandssymposium: update on the status of resources: 497-504. Santa Barbara Museum of Natural History,Santa Barbara, California, USA.
BROOKS ML (1999) Alien annual grasses and fire in theMojave desert. Madroño 46: 13-19.
BROWN DE & RA MINNICH (1986) Fire and changes increosote bush scrub of the western Sonora Desert,California. American Midland Naturalist 116: 411-422.
BRUMBAUGH RW (1980) Recent geomorphic and vege-tal dynamics on Santa Cruz Island, California. In:Power DM (ed) The California islands: proceedingsof a multidisciplinary symposium: 139-158. SantaBarbara Museum of Natural History, Santa Barbara,California, USA.
BULMAN TL (1988) The eucalyptus in California.Fremontia (United States) 16: 9-12.
BURGESS TL, JE BOWERS & RM TURNER (1991)Exotic plants at the desert laboratory, Tucson,Arizona. Madroño (United States) 38: 96-114.
BÚRQUEZ A & M QUINTANA (1994) Islands ofdiversity: Ironwood ecology and the richness ofperennials in a Sonora desert biological reserve. In:Nabhan GP & JL Carr (eds) Ironwood: anecological and cultural keystone of the Sonoradesert: 9-27. Conservation International, Washing-ton, District of Columbia, USA.
BYERS JE (2000) Competition between two estuarinesnails: implications for invasions of exotic Species.Ecology 81: 1225-1239.
BYRNE S (1979) The distribution and ecology of the non-native tree squirrels Sciurus carolinensis andSciurus niger in northern California. Ph.D. Thesis,University of California, Berkeley, California. 190pp.
CALLAWAY JC & MN JOSSELYN (1992) Theintroduction and spread of smooth cordgrass(Spartina alterniflora) in south San Francisco Bay.Estuaries 15: 218-226.
CANADELL J, RB JACKSON, JR EHLERINGER, HAMOONEY, OE SALA & ED SCHULZE (1996)Maximum rooting depth of vegetation types at theglobal scale. Oecologia 108: 583-594.
CANNON JP, EB ALLEN, MF ALLEN, LM DUDLEY &JJ JURINAK (1995) The effects of oxalatesproduced by Salsola tragus on the phosphorusnutrition of Stipa pulchra. Oecologia 102: 265-272.
CARLQUIST B (1990) An effective management plan forthe exotic mountain goats in Olympic National Park,Washington USA. Natural Areas Journal 10: 12-18.
CARLTON JT, JK THOMPSON, LE SCHEMEL & FHNICHOLS (1990) Remarkable invasion of San Fran-cisco Bay (California, USA) by the Asian clamPotamocorbula amurensis : introduction anddispersal. Marine Ecology Progress Series 66: 81-94.
CHAPIN FS III, HL REYNOLDS, CM D’ANTONIO &VM ECKHART (1996) The functional role of

427INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
species in terrestrial ecosystems. In: Walker B & WSteffen (eds) Global change and terrestr ialecosystems: 403-428. Cambridge University Press,Cambridge, United Kingdom.
CHASE TN, RA PIELKE, TGF KITTEL, JS BARON & TJSTOHLGREN (1999) Potential impacts on ColoradoRocky Mountain weather due to land use changes onthe adjacent Great Plains. Journal of GeophysicalResearch Atmospheres 104: 16673-16690.
CHATIGNY MH, D PREVOST, DA ANGERS, LPVEZINA & FP CHALIFOUR (1996) Microbialbiomass and N transformations in two soils croppedwith annual and perennial species. Biology andFertility of Soils 21: 239-244.
CHEATER M (1992) Alien invasion. Nature Conservancy(United States) 42: 24-29.
CHRISTENSEN NL (1994) The effects of fire on physicaland chemical properties of soils in Mediterranean-climate shrublands. In: Moreno JM & WC Oechel(eds) The role of fire in Mediterranean-typeecosystems: 79-95. Springer-Verlag, New York,New York, USA.
CLEVERLY JR, SD SMITH, A SALA & DA DEVITT(1997) Invasive capacity of Tamarix ramosissima ina Mojave desert floodplain: the role of drought.Oecologia 111: 12-18.
CLINE JF, DW URESK & WH RICKARD (1977)Comparison of water used by a sagebrush-bunchgrass community and a cheatgrasscommunity. Journal of Range Management 30: 199-201.
COBLENTZ BE (1980) Effects of feral goats on the SantaCatalina Island ecosystem. In: Power DM (ed) TheCalifornia islands: proceedings of amultidisciplinary symposium: 167-170. SantaBarbara Museum of Natural History, Santa Barbara,California, USA.
COHAN DR, BW ANDERSON & RD OHMART (1978)Avian population responses to saltcedar along thelower Colorado River. Forest Service GeneralTechnical Report WO-12: 371-381, United StatesForest Service, Washington, USA.
COHEN AN & JT CARLTON (1995) Non-indigenousaquatic species in a United States estuary: a casestudy of the biological invasions of the San Francis-co Bay and Delta. United States Fish and WildlifeService, Washington, District of Columbia, USA.272 pp.
COHEN AN & JT CARLTON (1998) Acceleratinginvasion rate in a highly invaded estuary. Science279: 555-558.
CORRELL JC, TR GORDON, AH MCCAIN, JW FOX,CS KOEHLER, DL WOOD & ME SCHULTZ(1991) Pitch canker disease in California:pathogenicity, distribution, and canker developmenton Monterey pine (Pinus radiata). Plant Disease 75:676-682.
D’ANTONIO CM (1990) Invasion of coastal plantcommunities by the introduced succulent ,Carpobrotus edulis (Aizoaceae). Ph.D. Thesis,University of California, Santa Barbara, California,USA. 212 pp.
D’ANTONIO CM & TL DUDLEY (1995) Biologicalinvasions as agents of change on islands versusmainlands. In: Vitousek PM, LL Loope & HAdsersen (eds) Islands: biological diversity andecosystem function: 103-121. Springer-Verlag, NewYork, New York, USA.
D’ANTONIO CM, TL DUDLEY & M MACK (1999)Disturbance and biological invasions: direct effectsand feedbacks. In: Walker LR (ed) Ecosystems of
disturbed ground: 413-452. Elsevier, Amsterdam,The Netherlands.
D’ANTONIO CM & PM VITOUSEK (1992) Biologicalinvasions by exotic grasses, the grass/fire cycle, andglobal change. Annual Review of Ecology andSystematics 23: 63-87.
DA SILVA PG & JW BARTOLOME (1984) Interactionbetween a shrub, Baccharis pilularis consanguinea(Asteraceae), and an annual grass, Bromis mollis(Poaceae), in coastal California. Madroño (UnitedStates) 31: 93-101.
DAEHLER CC (1996) Spartina invasions in Pacificestuaries: biology, impact, and management. In:Sytsma MD (ed) Proceedings of the symposium onnon-indigenous species in western aquaticecosystems. Portland State University Lakes andReservoirs Program, Publication (USA) 96-98: 1-6.
DAEHLER CC & DR STRONG (1996) Status, predictionand prevention of introduced cordgrass Spartinaspp. invasions in Pacific estuaries, USA. BiologicalConservation 78: 51-58.
DAWSON TE (1993) Hydraulic lift and water use byplants: implications for water balance, performanceand plant-plant interactions. Oecologia 95: 565-574.
DEL MORAL R & CH MULLER (1969) Fog drip: amechanism of toxin transport from Eucalyptusglobulus. Bulletin of the Torrey Botanical Club 96:467-475.
DEL MORAL R & CH MULLER (1970) The allelopathiceffects of Eucalyptus camaldulensis. The AmericanMidland Naturalist 83: 254-282.
DROST CA & GM FELLERS (1996) Collapse of a regio-nal frog fauna in the Yosemite area of theCalifornia Sierra Nevada, USA. ConservationBiology 10: 414-425.
DUDLEY T & B COLLINS (1995) Biological invasions inCalifornia Wetlands. Pacific Institute for Studies inDevelopment, Environment, and Security, Oakland,California, USA. 62 pp.
DUDLEY TL & NB GRIMM (1994) Modification ofmacrophyte resistance to disturbance by an exoticgrass, and implications for desert streamsuccession. Verhandlungen der InternationalenVereinigung für Theoretische und AngewandteLimnologie 25: 1456-1460.
DUKES JS & HA MOONEY (1999) Does global changeincrease the success of biological invaders? Trendsin Ecology and Evolution 14: 135-139.
DYER AR & KJ RICE (1999) Effects of competition onresource availability and growth of a Californiabunchgrass. Ecology 80: 2697-2710.
EGUNJOBI JK (1969) Dry matter and nitrogenaccumulation in secondary successions involvinggorse (Ulex europaeus L.) and associated shrubs andtrees. New Zealand Journal of Science 12: 175-193.
EGUNJOBI JK (1971) Ecosystem processes in a stand ofUlex europaeus L. I. Dry matter production, litterfall and efficiency of solar utilization. Journal ofEcology 59: 31-38.
EL-GHAREEB RM (1991) Suppression of annuals byTribulus terrestris in an abandoned field in thesandy desert of Kuwait. Journal of VegetationScience 2: 147-154.
EMERY SL & JA PERRY (1996) Decomposition rates andphosphorus concentrations of purple loosestrife(Lythrum salicaria) and cattail (Typha spp.) infourteen Minnesota wetlands. Hydrobiologia 323:129-138.
EVANS RA & JA YOUNG (1970) Plant litter and esta-blishment of alien annual weed species in rangelandcommunities. Weed Science 18: 697-703.

428 DUKES & MOONEY
EVANS RA & JA YOUNG (1972) Microsite requirementsfor establishment of annual weeds. Weed Science20: 350-356.
EVANS RC, DT TINGEY, ML GUMPERTZ & WFBURNS (1982) Estimates of isoprene andmonoterpene emission rates in plants. BotanicalGazette 143: 304-310.
EVANS RD, R RIMER, L SPERRY & J BELNAP (2001)Exotic plant invasion alters nitrogen dynamics in anarid grassland. Ecological Applications 11: 1301-1310.
FRANDSEN P & N JACKSON (1993) Impact of Arundodonax on flood control and endangered species. In:Jackson NE, P Frandsen & S Duthoit (eds)Proceedings of the Arundo donax workshop,Ontario, California: 13-16. California Exotic PestPlant Council, Berkeley, California, USA.
FRENKEL RE (1970) Ruderal vegetation along someCalifornia roadsides. University of California Press,Berkeley, California, USA. 163 pp.
GERLACH JD, Jr (2000) A model experimental system forpredicting the invasion success and ecosystemimpacts of non-indigenous summer-floweringannual plants in California’s central valleygrasslands and oak woodlands. Ph.D. Thesis,University of California, Davis, California, USA. v+ 102 pp.
GHOLZ HL, GM HAWK, A CAMPBELL & KCROMACK, Jr (1985) Early vegetation recoveryand element cycles on a clear-cut watershed inwestern Oregon. Canadian Journal of ForestResearch 15: 400-409.
GOLDBERG DE (1985) Effects of soil pH, competition,and seed predation on the distributions of two treespecies. Ecology 66: 503-511.
GOODWIN J (1992) The role of mycorrhizal fungi incompetetive interactions among nativebunchgrasses and alien weeds: a review andsynthesis. Northwest Science 66: 251-260.
GORDON DR & KJ RICE (1993) Competitive effects ofgrassland annuals on soil water and blue oak(Quercus douglasii) seedlings. Ecology 74: 68-82.
GORDON TR, KR WIKLER, AJ STORER & DL WOOD(1997) Pitch canker and its potential impacts onMonterey pine forests in California. Fremontia(United States) 25: 5-9.
GRAF WL (1978) Fluvial adjustments to the spread oftamarisk in the Colorado plateau region. GeologicalSociety of America Bulletin 89: 1491-1501.
GROZHOLZ ED, GM RUIZ, CA DEAN, KA SHIRLEY,JL MARON & PG CONNORS (2000) The impactsof a nonindigenous marine predator in a Californiabay. Ecology 81: 1206-1224.
GRUBB PJ, HE GREEN & RCJ MERRIFIELD (1969) Theecology of chalk heath: its relevance to thecalcicole-calcifuge and soil acidification problems.Journal of Ecology 57: 175-212.
GRUBB PJ & MB SUTER (1971) The mechanism ofacidification of soil by Calluna and Ulex and thesignificance for conservation. In: Duffey E & AS Watt(eds) The scientific management of animal and plantcommunities for conservation: 115-133. BlackwellScientific Publications, Oxford, United Kingdom.
GUENTHER A (1997) Seasonal and spatial variations innatural volatile organic compound emissions.Ecological Applications 7: 34-45.
HAGER HA & KD MCCOY (1998) The implications ofaccepting untested hypotheses: a review of theeffects of purple loosestrife (Lythrum salicaria) inNorth America. Biodiversity and Conservation 7:1069-1079.
HEADY HF (1988) Valley grassland. In: Barbour MG & JMajor (eds) Terrestrial vegetation of California:491-514. California Native Plant Society, Sacra-mento, California, USA.
HELGERSON OT, CT WHEELER, DA PERRY & JCGORDON (1979) Annual nitrogen fixation inScotchbroom. In: Gordon JC, CT Wheeler & DAPerry (eds) Symbiotic nitrogen fixation in themanagement of temperate forests. Oregon StateUniversity, Corvallis, Oregon, USA.
HENDRY GW (1931) The adobe brick as a historicalsource. Agricultural History 5: 110-127.
HEWITT CN, RK MONSON & R FALL (1990) Isopreneemissions from the grass Arundo donax L. are notlinked to photorespiration. Plant Science 66: 139-144.
HICKMAN JC (ed) (1993) The Jepson manual: higherplants of California. University of California Press,Berkeley, California, USA. xvii + 1,400 pp.
HOBBIE SE (1992) Effects of plant species on nutrientcycling. Trends in Ecology and Evolution 7: 336-339.
HOBBS NT, DS SCHIMEL, CE OWENSBY & DS OJIMA(1991) Fire and grazing in the tallgrass prairie:contingent effects on nitrogen budgets. Ecology 72:1374-1382.
HOBBS RJ & HA MOONEY (1998) Broadening theextinction debate: Population deletions andadditions in California and western Australia.Conservation Biology 12: 1-14.
HOFFMAN WA & RB JACKSON (2000) Vegetation-climate feedbacks in the conversion of tropicalsavanna to grassland. Journal of Climate 13: 1593-1602.
HÖLLDOBLER B & EO WILSON (1990) The ants.Harvard Universi ty Press, Cambridge,Massachusetts, USA. 732 pp.
HOLMES TH & KJ RICE (1996) Patterns of growth andsoil-water utilization in some exotic annuals andnative perennial bunchgrasses of California. Annalsof Botany 78: 233-243.
HOLWAY DA (1999) Competi t ive mechanismsunderlying the displacement of native ants by theinvasive Argentine ant. Ecology 80: 238-251.
HUENNEKE LF & JK THOMSON (1995) Potentialinterference between a threatened endemic thistleand an invasive non-native plant. ConservationBiology 9: 416-425.
HUMAN KG (1996) Interactions between the invasiveArgentine ant, Linepithema humile, and native antspecies. Ph.D. Thesis, Stanford Universi ty,Stanford, California, USA. 121 pp.
HUMAN KG & DM GORDON (1996) Exploitation andinterference competition between the invasiveArgentine ant, Linepithema humile, and native antspecies. Oecologia 105: 405-412.
HUMAN KG & DM GORDON (1997) Effects ofArgentine ants on invertebrate biodiversity innorthern California. Conservation Biology 11:1242-1248.
HUNTER R (1991) Bromus invasions on the Nevada testsite: present status of B. rubens and B. tectorumwith notes on their relationship to disturbance andaltitude. Great Basin Naturalist 51: 176-182.
HUNTER WC, RD OHMART & BW ANDERSON (1988)Use of exotic saltcedar (Tamarix chinensis) by birdsin arid riparian systems. Condor 90: 113-123.
JACKSON N (1993) The story of team arundo. CaliforniaEPPC News (United States) 1: 6-7.
JAMES SW (1991) Soil, nitrogen, phosphorus, and organicmatter processing by earthworms in tallgrassprairie. Ecology 72: 2101-2109.

429INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
JOHNSON V & J HARRIS (1990) Beaver. In: Zeiner DC,WF Laudenslayer, Jr, KE Mayer & M White (eds)California’s wildlife: mammals: 60-61. Departmentof Fish and Game, Sacramento, California, USA.
JOHNSTON CA & RJ NAIMAN (1990) The use of ageographic information system to analyze long-termlandscape alteration by beaver. Landscape Ecology4: 5-20.
KALISZ PJ & HB WOOD (1995) Native and exoticearthworms in wildland ecosystems. In: Hendrix PF(ed) Earthworm ecology and biogeography in NorthAmerica: 117-126. Lewis Publishers, Boca Raton,Florida, USA.
KEEGAN DR, BE COBLENTZ & CS WINCHELL (1994)Ecology of feral goats eradicated on San Clementeisland, California. In: Halvorson WL & GJ Maender(eds) The fourth annual California islandssymposium: update on the status of resources: 323-330. Santa Barbara Museum of Natural History,Santa Barbara, California, USA.
KEELEY JE (1995) Future of California floristics andsystematics: wildfire threats to the California flora.Madroño (United States) 42: 175-179.
KERBAVAZ JH (1985) Status reports on invasive weeds:pampas grass. Fremontia (United States) 12: 18-19.
KIESECKER JM & AR BLAUSTEIN (1998) Effects ofintroduced bullfrogs and smallmouth bass onmicrohabitat use, growth, and survival of nativered-legged frogs (Rana aurora). ConservationBiology 12: 776-787.
KINGSBURY JM (1964) Poisonous plants of the UnitedStates and Canada. Prentice-Hall, EnglewoodCliffs, New Jersey, USA. 626 pp.
KINLOCH BB, JR & D DULITZ (1990) White pine blisterrust at Mountain Home Demonstration State Forest:a case study of the epidemic and prospects forgenetic control. Research Paper PSW-204. UnitedStates Department of Agriculture, Forest Service,Pacific Southwest Research Station, Berkeley,California, USA.
KLINGER RC, PT SCHUYLER & JD STERNER (1994)Vegetation response to the removal of feral sheepfrom Santa Cruz island. In: Halvorson WL & GJMaender (eds) The fourth annual California islandssymposium: update on the status of resources: 341-350. Santa Barbara Museum of Natural History,Santa Barbara, California, USA.
KNOPF FL & TE OLSON (1984) Naturalization ofRussian-olive: implications to Rocky Mountainwildlife. Wildlife Society Bulletin 12: 289-298.
KOTANEN PM (1995) Responses of vegetation to achanging regime of disturbance: effects of feralpigs in a Californian coastal prairie. Ecography 18:190-198.
KREMER RG & SW RUNNING (1996) Simulating seasonalsoil water balance in contrasting semi-arid vegetationcommunities. Ecological Modelling 84: 151-162.
KUPFERBERG SJ (1997) Bullfrog (Rana catesbeiana)invasion of a California river: the role of larvalcompetition. Ecology 78: 1736-1751.
LA RUE S (1996). Seeds of destruction: exotic invaderscrowd out native plant species. The San DiegoUnion-Tribune, San Diego, California, USA,October 16, pp E1, E4.
LACEY JR, CB MARLOW & JR LANE (1989) Influenceof spotted knapweed (Centaurea maculosa) onsurface runoff and sediment yield. WeedTechnology 3: 627-631.
LAJEUNESSE S, R SHELEY, C DUNCAN & R LYM(1999) Leafy Spurge. In: Sheley RL & JK Petroff(eds) Biology and management of noxious
rangeland weeds: 249-260. Oregon State UniversityPress, Corvallis, Oregon, USA.
LAMB FH (1898) Sand dune reclamation on the Pacificcoast. The Forester 4: 141-142.
LANE J (1993) Overwintering monarch butterflies inCalifornia: past and present. In: Malcolm SB & MPZalucki (eds) Biology and conservation of themonarch butterfly: 335-344. Natural HistoryMuseum of Los Angeles County, Los Angeles,California, USA.
LAUGHRIN L, M CARROLL, A BROMFIELD & JCARROLL (1994) Trends in vegetation changeswith removal of feral animal grazing pressures onSanta Catalina island. In: Halvorson WL & GJMaender (eds) The fourth annual California islandssymposium: update on the status of resources: 523-530. Santa Barbara Museum of Natural History,Santa Barbara, California, USA.
LAYCOCK WA (1991) Stable states and thresholds of rangecondition on North American rangelands: a viewpoint.Journal of Range Management 44: 427-433.
LEAN J & DA WARRILOW (1989) Simulation of the re-gional climatic impact of Amazon deforestation.Nature 342: 411-413.
LESICA P (1997) Spread of Phalaris arundinaceaadversely impacts the endangered plant Howelliaaquatilis. Great Basin Naturalist 57: 366-368.
LESICA P & JS SHELLY (1996) Competitive effects ofCentaurea maculosa on the population dynamics ofArabis fecunda. Bulletin of the Torrey BotanicalClub 123: 111-121.
LEVINE JM, M VILÁ, CM D’ANTONIO, JS DUKES, KGRIGULIS & S LAVOREL (2003) Mechanismsunderlying the impacts of exotic plant invasions.Proceedings of the Royal Society of London B 270:775-781.
LIGHT T, DC ERMAN, C MYRICK & J CLARKE (1995)Decline of the Shasta crayfish (Pacifastacus fortisFaxon) of northeastern California. ConservationBiology 9: 1567-1577.
LINDQUIST JL, BD MAXWELL & T WEAVER (1996)Potential for controlling the spread of Centaureamaculosa with grass competition. Great BasinNaturalist 56: 267-271.
LOOPE LL, PG SÁNCHEZ, PW TARR, WL LOOPE &RL ANDERSON (1988) Biological invasions ofarid land nature reserves. Biological Conservation44: 95-118.
MACK RN (1981) Invasion of Bromus tectorum L. intowestern North America: an ecological chronicle.Agro-Ecosystems 7: 145-165.
MACK RN (1989) Temperate grasslands vulnerable toplant invasions: characteristics and consequences.In: Drake JA, HA Mooney, F di Castri, RH Groves,FJ Kruger, M Rejmánek & M Williamson (eds)Biological invasions: a global perspective: 155-179.John Wiley & Sons, Chichester, New York, USA.
MADDOX DM & A MAYFIELD (1985) Yellow starthistleinfestat ions are on the increase. CaliforniaAgriculture 39: 10-12.
MCCAIN AH, CS KOEHLER & SA TJOSVOLD (1987)Pitch canker threatens California pines. CaliforniaAgriculture 41: 22-23.
MEDINA AL (1988) Diets of scaled quail in southernArizona. Journal of Wildlife Management 52: 753-757.
MINCKLEY WL, PC MARSH, JE BROOKS, JEJOHNSON & BL JENSEN (1991) Managementtoward recovery of the razorback sucker. In:Minckley WL & JE Deacon (eds) Battle againstextinction: native fish management in the American

430 DUKES & MOONEY
west: 303-357. The University of Arizona Press,Tucson, Arizona, USA.
MINNICH RA & RJ DEZZANI (1998) Historical declineof coastal sage scrub in the Riverside-Perris Plain,California. Western Birds (United States) 29: 366-391.
MONSON RK, MT LERDAU, TD SHARKEY, DSSCHIMEL & R FALL (1995) Biological aspects ofconstructing volatile organic compound emissioninventories. Atmospheric Environment 29: 2989-3002.
MOONEY HA, SP HAMBURG & JA DRAKE (1986) Theinvasions of plants and animals into California. In:Mooney HA & JA Drake (eds) Ecology ofbiological invasions of North America and Hawaii:250-272. Springer-Verlag, New York, New York,USA.
MOONEY HA, PM VITOUSEK & PA MATSON (1987)Exchange of materials between terrestr ialecosystems and the atmosphere. Science 238: 926-932.
MOYLE PB (1973) Effects of introduced bullfrogs, Ranacatesbeiana, on the native frogs of the San Joaquinvalley, California. Copeia 1973: 18-22.
MOYLE PB (1986) Fish introductions into North America:Patterns and ecological impact. In: Mooney HA &JA Drake (eds) Ecology of biological invasions ofNorth America and Hawaii: 27-43. Springer-Verlag,New York, New York, USA.
MOYLE PB & T LIGHT (1996) Fish invasions inCalifornia: do abiotic factors determine success?Ecology 77: 1666-1670.
MURRAY MS & EM HANSEN (1997) Susceptibility ofPacific Yew to Phytophthora lateralis . PlantDisease 81: 1400-1404.
NAIMAN RJ, CA JOHNSTON & JC KELLEY (1988)Alteration of North American streams by beaver.Bioscience 38: 753-762.
NAIMAN RJ, T MANNING & CA JOHNSTON (1991)Beaver population fluctuations and troposphericmethane emissions in boreal wetlands.Biogeochemistry 12: 1-16.
NAIMAN RJ, JM MELLILO & JE HOBBIE (1986)Ecosystem alteration of boreal forest streams bybeaver (Castor canadensis). Ecology 67: 1254-1269.
NEHRING RB & PG WALKER (1996) Whirling diseasein the wild: the new reality in the intermountainwest. Fisheries 21: 28-30.
NEILL WM (1983) The tamarisk invasion of desert riparianareas. Educational Bulletin 83-84, Desert ProtectiveCouncil, Spring Valley, California, USA. 4 pp.
OJIMA DS, DS SCHIMEL, WJ PARTON & CEOWENSBY (1994) Long- and short-term effects offire on nitrogen cycling in tallgrass prairie.Biogeochemistry 24: 67-84.
OLSON TE & FL KNOPF (1986) Agency subsidation of arapidly spreading exotic. Wildlife Society Bulletin14: 492-493.
PAVLIK BM (1983a) Nutrient and productivity relationsof the dune grasses Ammophila arenaria andElymus mollis. I. Blade photosynthesis and nitrogenuse efficiency in the laboratory and field. Oecologia57: 227-232.
PAVLIK BM (1983b) Nutrient and productivity relationsof the dune grasses Ammophila arenaria andElymus mollis. II. Growth and patterns of drymatter and nitrogen allocation as influenced bynitrogen supply. Oecologia 57: 233-238.
PAVLIK BM (1983c) Nutrient and productivity relationsof the dune grasses Ammophila arenaria and
Elymus mollis . III. Spatial aspects of clonalexpansion with reference to rhizome growth anddispersal of buds. Bulletin of the Torrey BotanicalClub 110: 271-279.
PEART D, DT PATTEN & SL LOHR (1994) Feral pigdisturbance and woody species seedlingregeneration and abundance beneath coast live oaks(Quercus agrifolia) on Santa Cruz island,California. In: Halvorson WL & GJ Maender (eds)The fourth annual California islands symposium:update on the status of resources: 313-322. SantaBarbara Museum of Natural History, Santa Barbara,California, USA.
PELLANT M & C HALL (1994) Distribution of twoexotic grasses on intermountain rangelands: statusin 1992. In: Monsen SB & SG Kitchen (eds)Proceedings in ecology and management of annualrangelands: 109-112. United States Department ofAgriculture, Forest Service Intermountain ResearchStation, Ogden, Utah, USA.
PIMENTEL D, L LACH, R ZÚÑIGA & D MORRISON(2000) Environmental and economic costs ofnonindigenous species in the United States.Bioscience 50: 53-65.
POLLOCK MM, RJ NAIMAN, HE ERICKSON, CAJOHNSTON, J PASTOR & G PINAY (1995)Beaver as engineers: influences on biotic andabiotic characteristics of drainage basins. In: JonesCG & JH Lawton (eds) Linking species andecosystems: 117-126. Chapman and Hall, NewYork, New York, USA.
RACE MS (1982) Competitive displacement and predationbetween introduced and native mud snails .Oecologia 54: 337-347.
RAY JC (1988) Wild pigs in California: a major threat inCalifornia. Fremontia (United States) 16: 3-8.
REJMÁNEK M, CD THOMSEN & ID PETERS (1991)Invasive vascular plants of California. In: GrovesRH & F di Castri (eds) Biogeography ofMediterranean invasions: 81-101. CambridgeUniversity Press, Cambridge, United Kingdom.
REYNOLDS JW (1995) Status of exotic earthwormsystematics and biogeography in North America. In:Hendrix PF (ed) Earthworm ecology andbiogeography in North America: 1-27. LewisPublishers, Boca Raton, Florida, USA.
RICE EL (1992) Allelopathic effects on nitrogen cycling.In: Rizvi SJH & V Rizvi (eds) Allelopathy: basicand applied aspects: 31-58. Chapman and Hall,London, United Kingdom.
ROBBINS WW, MK BELLUE & WS BALL (1951)Weeds of California. Printing Division, Sacramen-to, California, USA. 547 pp.
ROBLES M & FS CHAPIN, III (1995) Comparison of theinfluence of two exotic communities on ecosystemprocesses in the Berkeley hills. Madroño (UnitedStates) 42: 349-357.
ROCHÉ CT & BF ROCHÉ, Jr (1988) Distribution andamount of four knapweed (Centaurea L.) species ineastern Washington. Northwest Science 62: 242-253.
RUNDEL PW & AC GIBSON (1996) Ecologicalcommunities and processes in a Mojave desertecosystem: Rock Valley, Nevada. CambridgeUniversity Press, Cambridge, United Kingdom.369 pp.
SALA A, SD SMITH & DA DEVITT (1996) Water use byTamarix ramosissima and associated phreatophytesin a Mojave Desert f loodplain. EcologicalApplications 6: 888-898.
SALA OE, FS CHAPIN, III, RH GARDNER, WKLAUENROTH, HA MOONEY & PS

431INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
RAMAKRISHNAN (1999) Global change,biodiversity and ecological complexity. In: Walker B,W Steffen, J Canadell & J Ingram (eds) The terrestrialbiosphere and global change: implications for naturaland managed ecosystems: 304-328. CambridgeUniversity Press, Cambridge, United Kingdom.
SANDERS NJ, NJ GOTELLI, NE HELLER & DMGORDON (2003) Community disassembly by aninvasive species. Proceedings of the NationalAcademy of Sciences of the United States 100:2474-2477.
SCHIMEL JP (1995) Plant transport and methaneproduction as controls on methane flux from arcticwet meadow tundra. Biogeochemistry 28: 183-200.
SCHLESINGER WH (1991) Biogeochemistry: an analysisof global change. Academic Press, San Diego,California, USA. 443 pp.
SCHUYLER P (1987) Control of feral sheep (Ovis aries)on Santa Cruz island, California. In: Hochberg FG(ed) Third California islands symposium: recentadvances in research in the California islands: 443-452. Santa Barbara Museum of Natural History,Santa Barbara, California, USA.
SCOTT GD (1993) Fire threat from Arundo donax. In:Jackson NE, P Frandsen & S Duthoit (eds)Proceedings of the Arundo donax workshop,Ontario, California: 17-18. California Exotic PestPlant Council, Berkeley, California, USA.
SEABLOOM EW & AM WIEDEMANN (1994)Distribution and effects of Ammophila breviligulataFern (American beachgrass) on the foredunes of theWashington coast. Journal of Coastal Research 10:178-188.
SEBACHER DI, RC HARRISS & KB BARTLETT (1985)Methane emissions to the atmosphere throughaquatic plants. Journal of Environmental Quality14: 40-46.
SHUKLA J, C NOBRE & P SELLERS (1990) Amazondeforestation and climate change. Science 247:1322-1325.
SLINGSBY P & WJ BOND (1985) The influence of antson the dispersal distance and seedling recruitmentof Leucospermum conocarpodendron (L.) Buek(Proteaceae). South African Journal of Botany 51:30-34.
SLOBODCHIKOFF CN & JT DOYEN (1977) Effects ofAmmophila arenaria on sand dune arthropodcommunities. Ecology 58: 1171-1175.
SMALL A (1994) California birds: their status anddistribution. Ibis Publishing Company, Vista,California, USA. 342 pp.
STORER AJ, TR GORDON & PL DALLARA (1994)Pitch canker kills pines, spreads to new species andregions. California Agriculture 48: 9-13.
SUAREZ AV & TJ CASE (2002) Bottom-up effects onpersistence of a specialist predator: ant invasionsand horned lizards. Ecological Applications 12:291-298.
SWANSON FJ (1981) Fire and geomorphic processes. In:Mooney HA, NL Bonnicksen, NL Christensen, JELotan & WA Reiners (eds) Fire regimes andecosystem properties: 401-420. United StatesDepartment of Agriculture, Forest Service, Was-hington, District of Columbia, USA.
THOMPSON DQ (1987) Spread, impact, and control ofpurple loosestrife (Lythrum salicaria) in NorthAmerican wetlands. United States Department ofthe Interior, Fish and Wildlife Service, Washington,District of Columbia, USA. 55 pp.
THOMPSON JD (1991) The biology of an invasive plant.Bioscience 41: 393-401.
TILMAN D & H OLFF (1991) An experimental study ofthe effects of pH and nitrogen on grasslandvegetation. Acta Oecologia 12: 427-442.
TOMBACK DF, JK CLARY, J KOEHLER, RJ HOFF &SF ARNO (1995) The effects of blister rust on post-fire regeneration of whitebark pine: the sundanceburn of northern Idaho (USA). ConservationBiology 9: 654-664.
TRAMMELL MA & JL BUTLER (1995) Effects of exoticplants on native ungulate use of habitat. Journal ofWildlife Management 59: 808-816.
TYSER RW & CH KEY (1988) Spotted knapweed in natu-ral area fescue grasslands: an ecologicalassessment. Northwest Science 62: 151-160.
VAN LILL WS, FJ KRUGER & DB VAN WYK (1980)The effect of afforestation with Eucalyptis grandisHill ex Maiden and Pinus patula Schlecht et Chamon streamflow from experimental catchments atMokobulaan, Transvaal. Journal of Hydrology 48:107-118.
VAN VUUREN MMI, R AERTS, F BERENDSE & W DEVISSER (1992) Nitrogen mineralization inheathland ecosystems dominated by different plantspecies. Biogeochemistry 16: 151-166.
VAN VUUREN MMI & F BERENDSE (1993) Changes insoil organic matter and net nitrogen mineralizationin heathland soils, after removal, addition orreplacement of litter from Erica tetralix or Moliniacaerulea. Biology and Fertility of Soils 15: 268-274.
VAN VUUREN MMI, F BERENDSE & W DE VISSER(1993) Species and si te differences in thedecomposition of l i t ters and roots from wetheathlands. Canadian Journal of Botany 71: 167-173.
VERVILLE JH, FS CHAPIN, III, SE HOBBIE & DUHOOPER (1998) Response of tundra CH4 and CO2flux to manipulation of temperature and vegetation.Biogeochemistry 41: 215-235.
VITOUSEK PM & LR WALKER (1989) Biologicalinvasion by Myrica faya in Hawaii: Plantdemography, nitrogen fixation, ecosystem effects.Ecological Monographs 59: 247-266.
VITOUSEK PM, CM D’ANTONIO, LL LOOPE, MREJMÁNEK & R WESTBROOKS (1997)Introduced species: a significant component ofhuman-caused global change. New Zealand Journalof Ecology 21: 1-16.
VITOUSEK PM, LR WALKER, LD WHITEAKER, DMUELLER-DOMBOIS & PA MATSON (1987)Biological invasion by Myrica faya al tersecosystem development in Hawaii. Science 238:802-804.
VIVRETTE NJ & CH MULLER (1977) Mechanism ofinvasion and dominance of coastal grassland byMesembryanthemum crystall inum . EcologicalMonographs 47: 301-318.
WAITHMAN JD, RA SWEITZER, D VANVUREN, JDDREW, AJ BRINKHAUS & IA GARDNER (1999)Range expansion, population sizes, andmanagement of wild pigs in California. Journal ofWildlife Management 63: 298-308.
WALLÉN B (1980) Changes in structure and function ofAmmophila during primary succession. Oikos 34:227-238.
WARD PS (1987) Distribution of the introduced Argentineant (Iridomyrmex humilis) in natural habitats of thelower Sacramento River Valley and its effects onthe indigenous ant fauna. Hilgardia 55: 1-16.
WARDLE DA, KS NICHOLSON, M AHMED & ARAHMAN (1994) Interference effects of the

432 DUKES & MOONEY
invasive plant Carduus nutans L. against thenitrogen fixation ability of Trifolium repens L.Plant and Soil 163: 287-297.
WEDIN DA & D TILMAN (1996) Influence of nitrogenloading and species composition on the carbon ba-lance of grasslands. Science 274: 1720-1723.
WEEKS EP, HL WEAVER, GS CAMPBELL & BDTANNER (1987) Water use by saltcedar and byreplacement vegetation in the Pecos Riverfloodplain between Acme and Artesia, NewMexico. United States Geological SurveyProfessional Paper 491-G. 33 pp.
WEHTJE W (1994) Response of a bishop pine (Pinusmuricata) population to removal of feral sheep onSanta Cruz Island, California. In: Halvorson WL &GJ Maender (eds) The fourth annual Californiaislands symposium: update on the status ofresources: 331-340. Santa Barbara Museum of Na-tural History, Santa Barbara, California, USA.
WERNER I & JT HOLLIBAUGH (1993) Potamocorbulaamurensis: comparison of clearance rates andassimilation efficiencies for phytoplankton andbacterioplankton. Limnology and Oceanography 38:949-964.
WESTMAN WE (1990) Park management of exotic plantspecies problems and issues. Conservation Biology4: 251-260.
WETTERER JK, AL WETTERER & E HEBARD (2001)Impact of the Argentine ant, Linepithema humile onthe native ants of Santa Cruz island, California.Sociobiology 38: 709-721.
WHEELER CT, OT HELGERSON, DA PERRY & JCGORDON (1987) Nitrogen fixation and biomassaccumulation in plant communities dominated byCytisus scoparius L. in Oregon and Scotland.Journal of Applied Ecology 24: 231-237.
WHEELER CT, DA PERRY, O HELGERSON & JCGORDON (1979) Winter fixation of nitrogen inScotch broom (Cytisus scoparius L.) . NewPhytologist 82: 697-701.
WHEELER GC & JN WHEELER (1986) The ants of Ne-vada. Natural History Museum of Los AngelesCounty and Allen Press, Inc., Lawrence, Kansas,USA. 138 pp.
WHISENANT SG (1990) Changing fire frequencies onIdaho’s snake river plains: ecological andmanagement implications. In: McArthur ED, EMRomney, SD Smith & PT Tueller (eds) Proceedingsof the symposium on cheat grass invasion, shrub
die-off, and other aspects of shrub biology andmanagement: 4-10. Intermountain Research Station,United States, Forest Service, Ogden, Utah, USA.
WHITSON TD, LC BURRILL, SA DEWEY, DWCUDNEY, BE NELSON, RD LEE & R PARKER(1996) Weeds of the west. Pioneer of Jackson Hole,Jackson, Wyoming, USA. 630 pp.
WIEDEMANN AM (1984) The ecology of Pacificnorthwest coastal sand dunes: a community profile.FWS/OBS-84/04. United States Fish and WildlifeService, Washington, District of Columbia, USA.
WIEDEMANN AM & A PICKART (1996) TheAmmophila problem on the northwest coast ofNorth America. Landscape and Urban Planning 34:287-299.
WILCOVE DS, D ROTHSTEIN, J DUBOW, A PHILLIPS& E LOSOS (1998) Quantifying threats toimperiled species in the United States. BioScience48: 607-615.
WINER AM, J AREY, R ATKINSON, SM ASCHMANN,WD LONG, CL MORRISON & DM OLSZYK(1992) Emission rates of organics from vegetationin California’s central valley. AtmosphericEnvironment 26: 2647-2659.
WOODWARD SL (1976) Feral burros of the ChemehueviMountains, California: the biogeography of a feralexotic. Ph.D. Thesis, University of California, LosAngeles, California, USA. 178 pp.
YAVITT JB, LL ANGELL, TJ FAHEY, CP CIRMO & CTDRISCOLL (1992) Methane fluxes, concentrations,and production in two Adirondack beaverimpoundments. Limnology and Oceanography 37:1057-1066.
YOUNG JA (1992) Ecology and management ofmedusahead (Taeniatherum caput medusae ssp.asperum [Simk.] Melderis). Great Basin Naturalist52: 245-252.
YOUNG JA & RA EVANS (1978) Population dynamicsafter wildfires in sagebrush grasslands. Journal ofRange Management 31: 283-289.
ZAVALETA E (2000) Valuing ecosystem services lost toTamarix invasion in the United States. In: MooneyHA & RJ Hobbs (eds) Invasive species in achanging world: 261-300. Island Press, Washing-ton, District of Columbia, USA.
ZEDLER PH, CR GAUTIER & GS MCMASTER (1983)Vegetation change in response to extreme events: theeffect of a short interval between fires in Californiachaparral and coastal scrub. Ecology 64: 809-818.
Associate Editor: Pablo Marquet

433INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
AP
PE
ND
IX 1
Som
e w
este
rn N
orth
Am
eric
an e
cosy
stem
dis
rupt
ors
and
thei
r ef
fect
sA
lgun
os d
isru
ptor
es e
cosi
stém
icos
del
oes
te d
e N
orte
amér
ica
y su
s ef
ecto
s
Inva
sive
spe
cies
Are
as a
ffec
ted1
Dis
rupt
ion
Evi
denc
e2R
efer
ence
PL
AN
TS
3 (l
ife
form
4 )A
mm
ophi
la a
rena
ria
(PG
)P
acif
ic c
oast
dun
esA
lter
s du
ne m
orph
olog
yO
-RB
arbo
ur &
Joh
nson
(19
88)
- O
pen
dune
are
aO
-RW
iede
man
n &
Pic
kart
(19
96)
- P
lant
div
ersi
tyO
Wie
dem
ann
(198
4)-
Art
hrop
od d
iver
sity
OD
-RB
arbo
ur e
t al
. (1
976)
+ P
rodu
ctiv
ity
OD
-RS
lobo
dchi
koff
& D
oyen
(19
77)
Exc
lude
s na
tive
pla
nts
OD
-RP
avli
k (1
983a
)A
lter
s m
icro
clim
ate
O-R
Bar
bour
at
al.
(198
5)O
D-R
Bar
bour
et
al.
(198
5)
Am
mop
hila
bre
vigu
lata
(P
G)
Pac
ific
nor
thw
est
dune
sA
lter
s du
ne m
orph
olog
yO
D-R
Sea
bloo
m &
Wie
dem
ann
(199
4)A
rund
o do
nax
(PG
)C
alif
orni
a ri
pari
an z
ones
+ F
ire
freq
uenc
yO
/SB
ell
(199
3),
Jack
son
(199
3)A
lter
s fl
ood
regi
mes
O/S
Fra
ndse
n &
Jac
kson
(19
93)
- P
lant
div
ersi
tyO
/SB
ell
(199
3)+
VO
C e
mis
sion
sO
D-R
Bas
ed o
n H
ewit
t et
al.
(19
90)
Bro
mus
dia
ndru
s (A
G)
Pa c
ific
sta
tes,
Bri
tish
Alt
e rs
hydr
olog
yE
M-R
Hol
me s
& R
ice
(199
6)C
olum
bia
to M
exic
o
Bro
mus
mad
rite
nsis
ssp
. ru
bens
(A
G)
Pa c
ific
sta
tes,
Bri
tish
+ F
ire
fre q
uenc
yS
D-R
Bro
wn
& M
inni
c h (
1986
)C
olum
bia
to M
exic
oH
unte
r (1
991)
OD
& E
M-R
Bro
oks
(199
9)
Bro
mus
te c
toru
m (
AG
)W
e ste
rn N
orth
Am
e ric
a+
Fir
e fr
e que
ncy
OD
-RW
hise
nant
(19
90)
Alt
e rs
N c
ycli
ngO
D-R
Eva
ns e
t a l
. (2
001)
- E
vapo
tra n
spir
a tio
nO
D-R
Cli
ne e
t a l
. (1
977)
, K
rem
e r &
Run
ning
(19
96)
Car
duus
nut
ans
(BF
)W
ide s
pre a
d in
Uni
ted
- N
inp
uts
EM
-RW
a rdl
e e t
al.
(19
94)
Sta
tes
a nd
Can
ada
Car
pobr
otus
edu
lis
(S/V
)C
oast
a l r
e gio
ns n
orth
e rn
- S
oil
pHO
DD
’Ant
onio
(19
90)
Ca l
ifor
nia
to M
exic
o;us
uall
y on
sa n
d
Cen
taur
e a m
acul
osa
(BF
)W
e ste
rn r
a nge
land
s+
Ero
sion
EM
-RL
a ce y
et
a l.
(198
9)
Cen
taur
e a s
olst
itia
lis
(AF
)C
a lif
orni
a , O
rego
n,A
lte r
s hy
drol
ogy
OD
Bor
man
et
a l.
(199
2),
Ge r
lac h
(20
00)
Wa s
hing
ton
gra s
sla n
ds,
patc
hy i
n Id
a ho

434 DUKES & MOONEY
Inva
sive
spe
cies
Are
as a
ffec
ted1
Dis
rupt
ion
Evi
denc
e2R
efer
ence
Cyn
odon
dac
tylo
n (P
G)
Cal
ifor
nia,
Ari
zona
,S
tabi
lize
s st
ream
bank
sO
D-R
Dud
ley
& G
rim
m (
1994
)so
uthe
rn U
nite
d S
tate
s
Cyt
isus
sco
pari
us (
S)
Pac
ific
coa
stal
ran
ge o
f+
N i
nput
s to
sys
tem
OD
-RW
heel
er e
t al
. (1
987)
Uni
ted
Sta
tes,
pat
ches
OO
-RD
ance
r et
al.
(19
77)
in S
ierr
an f
ooth
ills
Dip
sacu
s fu
llon
um (
BF
)W
ides
prea
d in
Thr
eate
ns n
ativ
e pl
ants
EM
-RH
uenn
eke
& T
hom
pson
(19
95)
Nor
th A
mer
ica
Ela
eagn
us a
ngus
tifo
lia
(S/T
)W
este
rn d
eser
t ri
pari
anA
lter
s w
ildl
ife
habi
tat
SO
lson
& K
nopf
(19
86)
zone
s
Euc
alyp
tus
spp.
(T
)C
alif
orni
a ri
pari
an+
Pro
duct
ivit
yO
D-R
Rob
les
& C
hapi
n (1
995)
zone
s, C
alif
orni
a+
Lit
ter
laye
rO
D-R
Rob
les
& C
hapi
n (1
995)
gras
slan
ds+
VO
C e
mis
sion
sO
D-R
Eva
ns e
t al
. (1
982)
, A
rey
et a
l. (
1995
)A
lter
s hy
drol
ogy
OO
-RV
an L
ill
et a
l. (
1980
)+
All
elop
athi
c ch
emic
als
EM
-RD
el M
oral
& M
ulle
r (1
969,
197
0)
Eup
horb
ia e
sula
(P
F)
Nor
ther
n U
nite
d S
tate
s,-
Hab
itat
qua
lity
OD
-RT
ram
mel
l &
But
ler
(199
5)so
uthe
rn C
anad
a
Fe s
tuc a
aru
ndin
ace a
(A
G)
Wid
e spr
e ad
in+
Ero
sion
SC
hea t
e r (
1992
)N
orth
Am
e ric
a
Ly t
hrum
sal
icar
ia (
S)
Wid
e spr
e ad
in N
orth
- W
ildl
ife
habi
tat
OT
hom
pson
et
a l.
(198
7)A
me r
ica ,
esp
. n.
Alt
e rs
P c
ycli
ngO
D-R
Em
e ry
& P
e rry
(19
96)
Uni
ted
Sta
tes
Me s
e mbr
y ant
hem
um c
ryst
alli
num
(A
F)
Coa
sta l
blu
ffs,
cli
ffs,
+ S
a lin
ity
of t
opso
ilO
D&
EM
-RV
ivre
tte
& M
ulle
r (1
977)
nort
hern
Ca l
ifor
nia
toM
exic
o, A
riz o
na
Sals
ola
trag
us (
AF
)D
istu
rbe d
inl
a nd
a re a
s+
P a
vail
a bil
ity
EM
-RC
anno
n e t
al.
(19
95)
Alt
e rs
fung
a l c
omm
unit
yO
-RG
oodw
in (
1992
)
Schi
smus
spp
. (A
G)
Sou
thw
e ste
rn+
Fir
e fr
e que
ncy
OD
& E
M-R
Bro
oks
(199
9)U
nite
d S
tate
s
Sene
c io
mik
anio
ide s
(V
)C
oast
a l n
orth
e rn
Ca l
ifor
nia
Alt
e rs
stre
amfl
owS
D.
Chi
ppin
g (p
e rso
nal
c om
mun
ica t
ion)
Sene
c io
sylv
atic
us (
AF
)N
orth
e rn
Ca l
ifor
nia
- N
utri
e nt
loss
fro
m c
lea r
c uts
OD
-RG
holz
et
a l.
(198
5)to
Bri
tish
Col
umbi
a

435INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
Inva
sive
spe
cies
Are
as a
ffec
ted1
Dis
rupt
ion
Evi
denc
e2R
efer
ence
Spar
tina
alt
erni
flor
a (P
G)
San
Fra
ncis
co B
ayC
olon
izes
bar
e m
ud f
lats
OD
-RC
alla
way
& J
osse
lyn
(199
2)C
alif
orni
a; S
uisl
aw+
Sed
imen
tati
onO
D-R
Bas
cand
(19
70)
estu
ary,
Ore
gon;
+ P
rodu
ctiv
ity
OD
-RC
alla
way
& J
osse
lyn
(199
2)W
illa
pa b
ay,
Was
hing
ton;
Pug
etS
ound
, W
ashi
ngto
n
Tae
niat
heru
m c
aput
-med
usae
(A
G)
Wid
espr
ead
in w
este
rn+
Fir
e fr
eque
ncy
O-R
You
ng (
1992
)U
nite
d S
tate
s,es
peci
ally
Pac
ific
nort
hwes
t
Tam
arix
spp
. (S
/T)
Sou
thw
este
rn U
nite
d+
Wat
er u
seO
Nei
ll (
1983
), W
eeks
et
al.
(198
7)S
tate
s &
Baj
a+
Sed
imen
tati
onO
D-R
Bla
ckbu
rn e
t al
. (1
992)
, G
raf
(197
8)C
alif
orni
a ri
pari
an-
Bir
d ha
bita
tO
D-R
Hun
ter
et a
l. (
1988
)zo
nes
OD
Coh
an e
t al
. (1
978)
Tri
bulu
s te
rres
tris
(A
F)
Pat
chy
thro
ugho
ut+
All
elop
athi
c ch
emic
als
OO
-RE
l-G
hare
eb (
1991
)N
orth
Am
eric
a
Ule
x e u
ropa
ea (
S)
Coa
sta l
are
a s f
rom
+ N
inp
ut t
o sy
stem
OO
-RE
gunj
obi
(196
9)c e
ntra
l C
a lif
orni
a to
+ P
rodu
c tio
nO
O-R
Egu
njob
i (1
971)
Ba j
a C
a lif
orni
a ,-
Soi
l pH
OO
-RG
rubb
et
a l.
(196
9),
Gru
bb &
Sut
e r (
1971
)pa
tche
s in
low
e rc a
sca d
e s a
nd s
ierr
a s
FU
NG
IC
rona
rtiu
m r
ibic
ola
Ca s
c ade
Ra n
ge,
Roc
kyK
ills
na t
ive
whi
te p
ine s
OD
Kin
loch
& D
ulit
z (1
990)
(ca u
ses
whi
te p
ine
blis
ter
rust
)M
ount
a ins
, S
ierr
aN
e va d
a fo
rest
s
Fus
ariu
m s
ubgl
utin
ans
Pin
us r
a dia
ta s
tand
s in
Kil
ls n
a tiv
e tr
e es
OD
-RC
orre
ll e
t a l
. (1
991)
, S
tore
r e t
al.
(19
94)
(ca u
ses
pitc
h c a
nke r
)c e
ntra
l a n
d so
uthe
rnC
a lif
orni
a
Phy
toph
thor
a la
tera
lis
Sou
thw
e st
Ore
gon,
Kil
ls P
ort-
Orf
ord-
c eda
rO
D-R
Mur
ray
& H
anse
n (1
997)
Nor
thw
e st
Ca l
ifor
nia
INV
ER
TE
BR
AT
ES
Apo
rre c
tode
a c a
ligi
nosa
(e a
rthw
orm
)-
Soi
l tu
rnov
e rO
D&
EM
-RJa
me s
(19
91)
- N
utri
e nt
min
e ra l
iza t
ion
OD
&E
M-R
Jam
e s (
1991
)
436 DUKES & MOONEY
Inva
sive
spe
cies
Are
as a
ffec
ted1
Dis
rupt
ion
Evi
denc
e2R
efer
ence
Bat
illa
ria
attr
amen
tari
a (m
udsn
ail)
Nor
ther
n C
alif
orni
a ba
ysD
ispl
aces
nat
ive
mud
snai
lE
M-R
Bye
rs (
2000
)
Car
cinu
s m
aena
s (g
reen
cra
b)P
acif
ic c
oast
: ce
ntra
l-
Inve
rteb
rate
abu
ndan
ceO
D &
EM
-RG
rosh
olz
et a
l. (
2000
)C
alif
orni
a to
cen
tral
Mix
es s
edim
ents
O-R
Coh
en e
t al
. (1
995)
Was
hing
ton
Ilya
nasi
a ob
seli
tis
(mud
snai
l)S
an F
ranc
isco
Bay
Dis
plac
es n
ativ
e m
udsn
ail
OD
&E
M-R
Rac
e (1
982)
Lin
epit
hem
a hu
mil
e (A
rgen
tine
ant
)C
alif
orni
a, A
rizo
na,
Rep
lace
s na
tive
ant
sO
D&
EM
-RH
uman
& G
ordo
n (1
996)
sout
hern
Nev
ada,
OD
-RW
ard
(198
7),
Wet
tere
r et
al.
(20
01)
Méx
ico
EM
-RH
olw
ay (
1999
)A
lter
s na
tive
com
mun
ity
OD
-RS
ande
rs e
t al
. (2
003)
-Hor
ned
liza
rd g
row
thO
D&
EM
-RS
uare
z &
Cas
e (2
002)
Oct
olas
ion
cyan
eum
(ea
rthw
orm
)-
Soi
l tu
rnov
erO
D&
EM
-RJa
mes
(19
91)
- N
utri
ent
min
eral
izat
ion
Orc
onec
tes
viri
lis
(vir
ile
cray
fish
)S
an F
ranc
isco
Bay
Dis
plac
es n
ativ
e cr
ayfi
shO
Coh
en &
Car
lton
(19
95)
Del
ta,
nort
hern
Cal
ifor
nia
irri
gati
ondi
tche
s
Pac
ifas
tac u
s le
nius
c ulu
s (s
igna
l c r
a yfi
sh)
Sa n
Fra
ncis
c o B
a yD
ispl
a ce s
na t
ive
c ra y
fish
OC
ohen
& C
a rlt
on (
1995
)
Pot
amoc
orbu
la a
mur
e nsi
s (A
sia n
cla
m)
San
Fra
ncis
c o B
a y+
Fil
tra t
ion
rate
of
wa t
e rE
M/O
D-R
We r
ner
& H
olli
baug
h (1
993)
Alt
e rs
sedi
men
t c h
a ra c
teri
stic
sO
-RC
a rlt
on e
t a l
. (1
990)
Spha
e rom
a qu
oyan
um (
isop
od)
San
Fra
ncis
c o B
a y+
Ero
sion
O/S
Coh
en &
Ca r
lton
(19
95)
VE
RT
EB
RA
TE
SC
apra
hir
c us
(goa
t)C
a lif
orni
a C
hann
e l I
sla n
ds+
Ero
sion
OC
oble
ntz
(198
0)
Cas
tor
c ana
dens
is (
bea v
e r)
Wid
e spr
e ad
in U
nite
dA
lte r
s re
gion
a lO
D-R
Na i
man
et
a l.
(198
6),
Na i
man
et
a l.
(198
8)S
tate
s a n
d C
anad
a ,ge
omor
phol
ogy,
e xc l
udin
g de
sert
shy
drol
ogy
(na t
ive
to m
ost
of+
Me t
hane
em
issi
onO
D-R
Na i
man
et
a l.
(199
1),
Ya v
itt
e t a
l. (
1992
) t
his
regi
on)
Equ
us a
sinu
s (b
urro
)C
a lif
orni
a (D
e ath
va l
ley)
Kil
ls n
a tiv
e ve
geta
tion
OL
oope
et
a l.
(198
8)
Lep
omis
mac
roc h
irus
(bl
uegi
ll)
Wid
e spr
e ad
inP
reys
on/
c om
pete
s w
ith
OC
ohen
& C
a rlt
on (
1995
)C
a lif
orni
a (n
a tiv
e to
nati
ve s
pec i
e se a
ste r
n a n
d c e
ntra
lN
orth
Am
e ric
a )

437INVASIVE SPECIES DISRUPT ECOSYSTEM PROCESSES
Inva
sive
spe
cies
Are
as a
ffec
ted1
Dis
rupt
ion
Evi
denc
e2R
efer
ence
Lep
omis
cya
nell
us (
gree
n su
nfis
h)W
ides
prea
d in
wes
tern
Pre
ys o
n/co
mpe
tes
wit
hO
Coh
en &
Car
lton
(19
95)
Uni
ted
Sta
tes
(nat
ive
nati
ve s
peci
esto
eas
tern
and
cen
tral
Nor
th A
mer
ica)
Mic
ropt
erus
sal
moi
des
Wid
espr
ead
inP
reys
on/
com
pete
s w
ith
OC
ohen
& C
arlt
on (
1995
)(l
arge
mou
th b
ass)
Cal
ifor
nia
(nat
ive
tona
tive
spe
cies
east
ern
and
cent
ral
Nor
th A
mer
ica)
Mor
one
saxa
tili
s (s
trip
ed b
ass)
Wid
espr
ead
in w
este
rnP
reys
on/
com
pete
s w
ith
OC
ohen
& C
arlt
on (
1995
)ri
vers
and
bay
sna
tive
spe
cies
(nat
ive
to A
tlan
tic
coas
t)
Ore
amno
s am
eric
anus
(m
ount
ain
goat
)W
ashi
ngto
n (O
lym
pic
+ E
rosi
onO
Car
lqui
st (
1990
), B
ratt
on (
1982
)m
ount
ains
)
Ovu
s ar
ies
(she
ep)
Cal
ifor
nia
Cha
nnel
+ E
rosi
onO
Bru
mba
ugh
(198
0)Is
land
s
Ran
a c a
tesb
e ian
a (b
ullf
rog)
Wid
e spr
e ad
in w
e ste
rnP
reys
on/
c om
pete
s w
ith
OD
-RM
oyle
(19
73)
Uni
ted
Sta
tes
(na t
ive
nati
ve s
pec i
e sO
D&
EM
-RK
upfe
rbe r
g (1
997)
to e
a ste
rn a
nd c
e ntr
a lN
orth
Am
e ric
a )
Rat
tus
norv
e gic
us (
Nor
way
ra t
)C
oast
a l a
nd c
e ntr
a lP
reys
on
nati
ve s
pec i
e sO
Coh
en &
Ca r
lton
(19
95)
Ca l
ifor
nia
Stur
nus
v ulg
aris
(E
urop
e an
sta r
ling
)W
ide s
pre a
d in
Nor
thD
ispl
a ce s
na t
ive
c avi
ty-
OS
ma l
l (1
994)
Am
e ric
ane
stin
g bi
rds
Sus
scro
fa (
pig)
Wid
e spr
e ad
in C
a lif
orni
aD
istu
rbs
soil
sO
D-R
Kot
a ne n
(19
95)
Vul
pes
v ulp
e s (
red
fox)
Wid
e spr
e ad
in C
a lif
orni
aP
reys
on
nati
ve s
pec i
e sO
Coh
en &
Ca r
lton
(19
95)
PR
OT
IST
My x
obol
us c
e re b
rali
sP
a cif
ic s
tate
s, n
orth
e rn
Kil
ls r
a inb
ow &
bro
wn
OD
Neh
ring
& W
a lke
r (1
996)
,U
nite
d S
tate
str
out
by c
a usi
ngB
e rge
rse n
& A
nde r
son
(199
7) (
rang
e da
ta)
whi
rlin
g di
sea s
e
1In
form
a tio
n in
“a r
e as
a ffe
c te d
” is
la r
gely
de r
ive d
fro
m H
ickm
an (
1993
) a n
d W
hits
on e
t a l
. (1
996)
, or
fro
m t
he c
ite d
re f
e re n
c es.
2K
ey t
o e v
ide n
c e c
olum
n: e
xpe r
imen
tal
man
ipul
a tio
n, E
M;
obse
rva t
ion
wit
h da
ta,
OD
; ob
serv
a tio
n w
itho
ut d
a ta ,
O;
obse
rva t
ion
outs
ide
of w
e ste
rn N
orth
Am
e ric
a , O
O;
spe c
ula t
ion
wit
h da
ta,
SD
; sp
e cul
a tio
n, S
. B
oldf
a ce
is u
sed
to d
e not
e pr
ima r
y re
fere
nce s
. T
he s
uffi
x -R
de n
ote s
evi
denc
e pu
blis
hed
in p
e er-
revi
e we d
sc i
e nti
fic
lite
ratu
re.
3P
lant
nom
enc l
a tur
e fr
om H
ickm
an (
1993
).4
Key
to
pla n
t li
fe f
orm
s: t
ree ,
T;
shru
b, S
; vi
ne,
V;
a nnu
a l g
rass
, A
G;
pere
nnia
l gr
a ss,
PG
; a n
nua l
for
b, A
F;
pere
nnia
l fo
rb,
PF





























