Embed Size (px)
Citation preview

DINÁMICA DE LA DENSIDAD DE ÁRBOLES Y LA BIOMASA AÉREA EN
BOSQUES ANDINOS DEL DEPARTAMENTO DE ANTIOQUIA, COLOMBIA
REMEDICIÓN DE PARCELAS PERMANENTES
PROYECTO EXPEDICIÓN ANTIOQUIA
Álvaro Javier Duque Montoya
Asesor Académico
Miguel Ángel Peña
Iván López Montiel
Adriana Yépes
Felipe Cardona
Junio 10 de 2015
Medellín, Colombia
En el marco del Proyecto
Mecanismo para la Mitigación Voluntaria de Emisiones de GEI en Colombia – MVC producto 2.4
Generación de información de crecimiento y captura de carbono de especies nativas generada,
incorporada a bases de datos nacionales, y utilizada por el portafolio de proyectos de carbono
forestal

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 2
TABLA DE CONTENIDO
INTRODUCCIÓN .............................................................................................................................. 4
OBJETIVO GENERAL ...................................................................................................................... 5
OBJETIVOS ESPECÍFICOS .............................................................................................................. 5
ÁREA DE ESTUDIO ......................................................................................................................... 6
PREPARACIÓN DE LOS DATOS .................................................................................................... 9
BIOMASA AÉREA .......................................................................................................................... 10
TASAS DEMOGRÁFICAS .............................................................................................................. 10
DINÁMICA BIOMASA AÉREA ..................................................................................................... 10
RESULTADOS ................................................................................................................................. 11
Tasas de cambio por categoría diamétrica .................................................................................... 11
Tasas de cambio por categoría de densidad de la madera: ............................................................ 14
DISCUSIÓN ..................................................................................................................................... 16
BIBLIOGRAFIA ............................................................................................................................... 18

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 3
LISTA DE TABLAS
Tabla 1. Características y ubicación de las 8 parcelas permanentes establecidas en el departamento
de Antioquia, Colombia. ..................................................................................................................... 6
LISTA DE FIGURAS
Fig 1. Localización de las parcelas permanentes en el departamento de Antioquia, Colombia. Los
puntos oscuros representan las parcelas con altitudes por debajo de los 1.500 m (Tb = tierras bajas);
y los puntos grises representan las parcelas con altitudes por encima de los 1.500 m (Ta = tierras
altas). ................................................................................................................................................... 6
Fig 2. Remedición parcelas permanentes. A) medición diamétro individuos del dosel y B) del
sotobosque. .......................................................................................................................................... 8
Fig 3. Remedición parcelas permanentes. A) remedición diámetros y B) medición alturas con vara
extensible............................................................................................................................................. 9
Fig 4. (A) Número total de individuos (ha-1
); y (B) biomasa aérea total (t/ha) (D≥1cm). Las barras
contiguas indican cada censo. WD0.25: especies con densidad de madera ≤ 0.52gr cm-3
. WD0.5:
especies con densidad de madera 0.52<WD≤0.69gr cm-3
. WD0.75: especies con densidad de madera
≤ 0.69gr cm-3
. .................................................................................................................................... 12
Fig 5. Dinámica y cambio neto (% año-1
) del número de individuos (A, B y C) y de la biomasa
aérea (D, E y F), para las categorías de tamaño definidas: todo (D≥1cm; A y D), dosel (D≥10cm; B
y E) y sotobosque (1cm≤D<10cm; C y F). ....................................................................................... 13
Fig 6. Dinámica y cambio neto (% año-1
) del número de individuos (A, B y C) y de la biomasa
aérea (D, E y F), para los grupos de especies definidos a partir de la densidad de la madera: WD0.25
(A y D), WD0.5 (B y E) y WD0.75 (C y F).......................................................................................... 15

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 4
INTRODUCCIÓN
Los bosques tropicales, considerados uno de los ecosistemas más extensos y productivos
del planeta (Phillips & Gentry, 1994; Phillips et al., 2002), se estima que contienen
aproximadamente el 40-50% del carbono almacenado en toda la vegetación terrestre (Pan et
al., 2011; Saatchi et al., 2011). Por este motivo, el papel que juegan los bosques tropicales
en la regulación del carbono y mitigación del cambio climático es preponderante (Clark,
2007). Sin embargo, uno de los principales desafíos que enfrentan los proyectos de
conservación y manejo sostenible de los bosques, en especial aquellos enfocados en los
mecanismos de Reducción de emisiones por Deforestación y Degradación de bosques (e.g.
REDD+), es obtener estimados confiables de la variación espacial y temporal de los
contenidos de carbono que almacenan los bosques. En buena medida, la carencia de
información está asociada con la alta variabilidad de la biomasa aérea (BA) contenida en
los bosques, la cual representa el compartimento de carbono con mayor dinámica natural
dentro del sistema. En consecuencia, el monitoreo simultáneo de las tasas de crecimiento,
reclutamiento y mortalidad, es crucial para entender la variación temporal de la BA e inferir
acerca de los factores que determinan la pérdida y/o acumulación de carbono en los
bosques tropicales (Baker et al., 2004; Valencia, Condit, Muller-Landau, Hernández &
Navarrete, 2009).
Durante las últimas décadas, diferentes estudios han reportado un incremento en la BA
acumulada en los bosques tropicales, así como en las tasas de mortalidad y crecimiento
(Baker et al., 2004; Lewis et al., 2004; Chave et al., 2008; Phillips, Lewis, Baker, Chao &
Higuchi, 2008; Lewis et al., 2009; Brienen et al., 2015). Estos cambios funcionales y
estructurales en los bosques han sido principalmente atribuidos al continuo aumento en las
concentraciones de CO2 en la atmósfera (Lewis et al., 2004; Phillips et al., 2004; 2008;
Lewis et al., 2009). Aun así, la alta tasa de fertilización por los incrementos de la
concentración de CO2 parece estar a su vez menguando la longevidad de las especies y
aumentando la mortalidad, dando como resultado una disminución en la tasa de
acumulación de carbono de los bosques en la última década (Brienen et al., 2015). Sin
embargo, otros autores proponen mecanismo alternos asociados con procesos de respuesta a
la perturbación o disturbio, como responsables del incremento en la productividad
observada en los bosques (Chave et al., 2008). Si el aumento en las concentraciones
atmosféricas de CO2 es la principal causa del incremento en la BA acumulada por los
bosques, se espera un aumento simultáneo en el crecimiento, reclutamiento y mortalidad de
árboles, pero siendo mayor el reclutamiento que la mortalidad (Lewis et al., 2004; Phillips
et al., 2004). Si por el contrario, los bosques están en un proceso de recuperación como
respuesta a un disturbio pasado, se podría encontrar un incremento en el aumento en
abundancia y BA de especies de crecimiento lento en relación con especies de rápido
crecimiento (Chave et al., 2008). Cuando los bosques están sometidos a aclareo natural
(“self-thinning”) como recuperación de un disturbio, lo que se debe esperar es un
incremento en la mortalidad de individuos, pero con un aumento en la BA (Oliver &
Larson, 1990). Como una premisa básica, la mayoría de estudios asocian especies de rápido
crecimiento con baja densidad de madera (WD) y especies de lento crecimiento con alta
WD (Wright et al., 2010).

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 5
No obstante, nuestro conocimiento sobre la distribución espacial y temporal de la
abundancia de individuos (AI) y BA, y por ende de las reservas de carbono en los bosques
tropicales en general, continúa siendo limitado. En el caso de los bosques tropicales de Sur
América, gran parte de los estudios sobre los cambios temporales de la BA se han centrado
en bosques de tierras bajas (Phillips et al., 2004; 2008; Valencia et al., 2009; Brienen et al.,
2015), limitando nuestro conocimiento de su variación espacial y temporal en los
ecosistemas andinos. Generalmente se asumen los ecosistemas tropicales de tierras bajas,
en especial los de la Amazonía, como los bosques más productivos del planeta (Phillips et
al., 2004). Sin embargo, en bosques andinos de alturas intermedias, se pueden encontrar
valores de BA similares o incluso superiores a los reportados para la Amazonia (Sierra et
al., 2007; Yepes et al., 2015). En este estudio, que tiene como objetivo contribuir con el
conocimiento de la dinámica de la AI y BA de los bosques andinos, se analizó la
información proveniente de dos censos realizados en ocho parcelas permanentes de 1 ha
cada una, las cuales están localizadas en el noroeste de Colombia, en su mayoría en el
departamento de Antioquia. Con base en esta información, se busca dar respuesta a las
siguientes preguntas: 1) ¿Cuál es la tendencia actual en relación con el esperado equilibrio
dinámico de la AI y BA en los bosques estudiados?; 2) ¿Según la tendencia en la dinámica
de la AI y BA, que podemos deducir acerca del efecto de la fertilización por CO2 o la
respuesta al disturbio como posibles mecanismos que determinan la dinámica observada en
los bosques estudiados?
OBJETIVO GENERAL
El objetivo de este convenio es el de generar información sobre crecimiento y captura de
carbono para especies nativas, que sea incorporada en sistemas nacionales de información y
usada por los proyectos de carbono forestal del país.
OBJETIVOS ESPECÍFICOS
a) Realizar un trabajo de campo que permita la remedición de las variables
estructurales básicas (el. Diámetro, altura, taxonomía) en 8 parcelas de las 16 establecidas
en marco del proyecto Expedición Antioquia.
b) Estimar la biomasa aérea y los contenidos de carbono asociados para un segundo
periodo de medición (2013/2014), y compararlos con los obtenidos para el periodo uno
(2007/2009).
c) Identificar si existen pérdidas y ganancias en términos de carbono durante el
periodo de análisis, y determinar si en el caso de las pérdidas se observa degradación.
d) Identificar especies con crecimientos importantes en biomasa que puedan ser
potenciales para efectos de restauración ecológica y/o proyectos de carbono forestal.
e) Aportar el conocimiento de la dinámica forestal de los bosques naturales del
departamento de Antioquia.

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 6
ÁREA DE ESTUDIO
Fig. 1. Localización de las parcelas permanentes en el departamento de Antioquia,
Colombia. Los puntos oscuros representan las parcelas con altitudes por debajo de los
1.500 m (Tb = tierras bajas); y los puntos grises representan las parcelas con altitudes por
encima de los 1.500 m (Ta = tierras altas).
El área de estudio se localiza en el departamento de Antioquia (Colombia), en los
municipios de Anorí, Belmira, Caicedo, El Bagre, Nechí, Necoclí, Puerto Triunfo y
Támesis (Fig. 1). En cada uno de estos municipios se estableció una parcela permanente de
1 ha, cubriendo un gradiente altitudinal que varía entre 50 a 2.950 msnm. La precipitación
promedio anual varía entre 1.750 a 4.250 mm año-1
(Tabla 1). La topografía y la geología
de la zona es muy variable como producto de los sedimentos volcánicos y aluviales que dan
origen a la diversidad de suelos y las propiedades geomorfológicas de la región (Malagón,
2003).
Tabla 1. Características y ubicación de las ocho (8) parcelas permanentes establecidas en el
departamento de Antioquia, Colombia.

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 7
Código Municipio Coordenadas A P T
Sotobosque Dosel
n1(n2) BA1(BA2) n1(n2) BA1(BA2)
A Támesis 05°47’42” N
75°40’11” W 550 1.750 26
3.475
(2.825)
10,76
(9,98)
428
(434)
135,94
(143,91)
B Caicedo 06°23’30” N
76°01’31” W 2.670 2.200 18
5.425
(3.481)
25,45
(21,52)
1.141
(1.087)
223,75
(232,78)
F Nechí 08°03’21” N
74°46’05” W 129 3.250 28
2.206
(2.081)
7,54
(6,96)
501
(463)
199,13
(216,43)
G El Bagre 06°00’22” N
74°36’38” W 50 4.250 37
5.319
(4.906)
14,64
(16,04)
497
(469)
183,52
(185,26)
H Pto. Triunfo 07°39’30” N
74°49’07” W 192 2.250 31
5.400
(4.719)
20,26
(20,73)
365
(376)
195,23
(222,89)
J Necoclí 08°30’26” N
76°39’27” W 68 1.750 24
5.063
(3.900)
17,28
(10,79)
527
(516)
184,39
(191,94)
L Anorí 06°58’00” N
75°09’00” W 1.750 2.400 21
7.488
(7.250)
21,54
(22,70)
918
(949)
188,86
(210,34)
M Belmira 06°09’05” N
75°42”00” W 2.950 2.100 11
5.088
(5.300)
17,46
(19,60)
485
(491)
201,58
(225,76)
A=altitud (msnm); P=precipitación promedio anual (mm año-1
); T=Temperatura promedio (°C); n=número de
individuos por hectárea (D≥1cm) en el primer (n1) y segundo censo (n2); BA=biomasa aérea (t ha-1
) en el
primer (BA1) y segundo censo (BA2).
Las parcelas fueron establecidas entre los años 2006 y 2009 y la remedición de estas se
realizó entre los años 2013 y 2014. Durante el primer censo en cada parcela se marcaron,
mapearon, midieron y colectaron todos los árboles y palmas con D (diámetro a 1.3 m de
altura) mayor o igual a diez cm (dosel; Fig. 2A), los individuos con deformación en los
tallos, bambas y raíces tabulares o columnares se midieron donde el tallo era regular
(Condit, 1998). Adicionalmente, en cada parcela se estableció una subparcela de 0.16 ha
(40×40m), en la cual se muestrearon todos los árboles y palmas con D mayor o igual a 1cm
y menor a 10cm (sotobosque; Fig. 2B).

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 8
Fig. 2. Remedición parcelas permanentes. A) medición diamétro individuos del dosel y B)
del sotobosque.
En todos los casos se marcó con pintura el punto de medición del D, con el objetivo de
realizar las futuras mediciones en el mismo lugar (Fig. 3A). La medición de alturas se
realizó al 40% de los individuos; los individuos con alturas menores de 15 m se midieron
con vara extensibles (Fig. 3B) y aquellos con altura superior a 15 m con hipsómetro laser
(Hipsómetro Forestry 550). Durante el segundo censo se registró el crecimiento en el D, el
reclutamiento y la mortalidad. La determinación taxonómica y homologación del material
vegetal se llevó a cabo en el Herbario de la Universidad de Antioquia (HUA).

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 9
Fig. 3. Remedición parcelas permanentes. A) Remedición diámetros y B) medición alturas
con vara extensible.
PREPARACIÓN DE LOS DATOS
Durante la remedición de parcelas permanentes es común encontrar algunos individuos con
crecimiento negativo o positivos muy altos (Condit et al., 2004); en estos casos se
identificaron los individuos con crecimientos extremos negativos (menores de -0.1cm año-
1) y positivos (mayores de 7.5cm año
-1). Posteriormente, el D del segundo censo de los
individuos identificados con crecimiento extremo, se modificó para que su crecimiento
diamétrico no fuera inferior o superior, según el caso, a los valores indicados anteriormente
como extremos.
Para estimar la altura de todos los individuos, se generó un modelo de regresión diámetro–
altura, para lo cual se utilizó el modelo asintótico de Weibull 𝐻 = 𝑎 × (1 − 𝑒(−𝑏 × 𝐷𝑐));
donde H es la altura, a, b y c son los parámetros a estimar y D el diámetro asociado con
cada árbol en cada medición (Feldpausch et al., 2012). Para obtener los parámetros
requeridos para el modelo se usó la información de todos los árboles con registro de altura
en las ocho parcelas. El modelo obtenido se utilizó para la estimación de la altura de todos
los árboles en los dos periodos de medición.
A cada individuo se le asignó el valor de WD empleando la información reportada en la
literatura (Chave, Muller-Landau, Baker, Easdale, Ter Steege, & Webb, 2006; Zane et al.,
2009). En caso de no encontrar la densidad a nivel de especie, se le asignó el promedio por
género, familia o el promedio de la densidad de las especies registradas en cada parcela,
respectivamente. La WD se seleccionó como un rasgo funcional clave (Wright et al., 2010),
que permite analizar la influencia de grupos de especies con diferentes estrategias de
supervivencia, crecimiento y mortalidad sobre las tasas de acumulación de BA y el
recambio de individuos. Con base en lo anterior, se empleó el valor de la WD por especies
para la definición de grupos funcionales en el presente estudio. Es decir, todos los valores
de WD, de los individuos monitoreados en el primer censo, fueron agrupados y

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 10
posteriormente divididos en dos grupos funcionales extremos, los cuales fueron definidos
por el primer (WD0.25) y cuarto cuartil (WD0.75), con un tercer grupo intermedio (WD0.5)
(Peña & Duque, 2013).
BIOMASA AÉREA
Para estimar la BA se utilizó la ecuación alométrica propuesta por Chave et al. (2014)
𝐵𝐴 = 0.0673 × (𝑊𝐷 × 𝐷2 × 𝐻)0.976; donde BA es la biomasa aérea estimada en
kilogramos, D es el diámetro del árbol, H la altura estimada y WD es el valor de la
densidad de la madera. Esta ecuación fue construida con información de 4.004 árboles con
D≥5 cm (Chave et al., 2014). Dado que en la información del presente estudio se incluyen
árboles con D≥1 cm, el uso de esta ecuación puede inducir menos errores en los estimados
que los que se obtendrían con el uso de las ecuaciones propuestas por Álvarez et al. (2012)
para los bosques naturales de Colombia, en las cuales solo se incluyeron árboles con D≥10
cm.
TASAS DEMOGRÁFICAS
Las tasas de reclutamiento y mortalidad de individuos, son usualmente estimadas a partir de
modelos logarítmicos, los cuales asumen probabilidades constantes en cada período de
tiempo (Phillips, Hall, Gentry, Sawyer & Vásquez, 1994). La tasa de mortalidad de
individuos se calculó como 𝑚𝐴𝐼 = (ln(𝑁0 𝑁𝑠⁄ ) 𝑡⁄ ) × 100, y la tasa de reclutamiento de
individuos como 𝑟𝐴𝐼 = (ln(𝑁𝑓 𝑁𝑠⁄ ) 𝑡⁄ ) × 100. El cambio neto del número de individuos se
calculó como 𝑁𝐴𝐼 = (ln(𝑁𝑓 𝑁0⁄ ) 𝑡⁄ ) × 100 (Condit et al., 2004). Donde N0 es el número
inicial de árboles, Ns el número original de árboles que sobreviven en el último censo
[𝑁𝑠 = 𝑁0 − 𝑁𝑑], Nf el número final de árboles [𝑁𝑓 = 𝑁0 − 𝑁𝑑 + 𝑁𝑟]; siendo Nd el número
de árboles muertos entre censos y t es el tiempo entre censos.
DINÁMICA BIOMASA AÉREA
La dinámica de la BA de cada parcela fue representada por las tasas anuales de mortalidad,
reclutamiento, crecimiento e incremento neto. La tasa anual de mortalidad entre los censos
se estimó como 𝑚𝐵𝐴 = (ln (𝐵𝐴1 (𝐵𝐴1 − 𝑀)⁄ ) 𝑡⁄ ) × 100. La tasa anual de reclutamiento se
estimó como 𝑟𝐵𝐴 = (ln((𝐵𝐴1 − 𝑀 + 𝑅) (𝐵𝐴1 − 𝑀)⁄ ) 𝑡⁄ ) × 100. La tasa de crecimiento
relativo de la BA entre los censos se estimó como
𝑔𝐵𝐴 = (ln((𝐵𝐴2 − 𝑅) (𝐵𝐴1 − 𝑀)⁄ ) 𝑡⁄ ) × 100. El cambio neto de la BA se estimó como
𝑁𝐵𝐴 = (ln(𝐵𝐴2 𝐵𝐴1⁄ ) 𝑡⁄ ) × 100. Donde BA1 y BA2 es la BA en el primer y segundo censo
respectivamente, M es la BA de los árboles muertos y R es la BA de los individuos
reclutados durante el segundo censo (Valencia et al., 2009; Peña & Duque, 2013).

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 11
Todos los análisis de la dinámica de individuos y BA indicados anteriormente se realizaron
por separado para los individuos del dosel (D≥10cm) sotobosque (1cm≤D<10cm) y todos
los individuos (D≥1cm), el cual se tomó como la suma de lo reportado para el dosel y
sotobosque. Todos los valores provenientes de las subparcelas de 0.16 ha fueron llevados a
hectárea, con el objetivo de poder hacer comparables los datos. De igual forma, dichos
análisis se realizaron de forma independiente para los tres grupos de especies definidos
anteriormente con base en la WD (WD0.25, WD0.5 y WD0.75).
RESULTADOS
Con respecto al total de todos los individuos (D≥1cm), se observó una disminución en la AI
entre censos, el cual disminuyó de un valor promedio de 5.541 ± 1.694 individuos ha-1
en el
primer censo a 4.897 ± 1.714 individuos ha-1
en el segundo (promedio ± desviación
estándar; Fig. 4A). Se observó una disminución anual promedio entre censos en la AI de -
157.34 ± 175.83 individuos ha-1
año-1
. En total, este valor representa una tasa de cambio
neto en la AI de -2.83 ± 2.77% año-1
(Fig. 5A). La cantidad promedio de BA por el
contrario, incrementó de 205.92 ± 28.64t ha-1
en el primer censo a 219.70 ± 32.90t ha-1
en
el segundo (Fig. 4B). El valor promedio de BA, tuvo un incremento anual promedio de 2.23
± 1.77t ha-1
año-1
, lo cual equivale a una tasa de cambio neto de la BA de 1.08 ± 0.81% año-
1 (Fig. 5D). La pérdida total de individuos por mortalidad (290.91 ± 152.64 individuos ha
-1
año-1
) fue mayor que la ganancia por reclutamiento (133.57 ± 107.18 individuos ha-1
año-1
).
En el caso de la BA para el total de individuos, la mayor parte de la ganancia se dio por
efecto del crecimiento (gBA=4.65 ± 1.26t ha-1
año-1
), pero siendo en este caso mayor la
mortalidad (mBA=3.06 ± 1.57t ha-1
año-1
) que el reclutamiento (rBA=0.72 ± 0.67t ha-1
año-1
).
Tasas de cambio por categoría diamétrica: En relación a los individuos del dosel
(D≥10cm), se observó una menor disminución en la AI entre censos, la cual paso de un
valor promedio de 608 ± 272 individuos ha-1
en el primer censo a 598 ± 265 individuos ha-1
en el segundo. No obstante, las parcelas A, H, L y M presentaron un incremento neto de los
individuos entre censos (Tabla 1). La disminución anual promedio entre censos en la AI fue
de -2.49 ± 5.27 individuos ha-1
año-1
. En total, este valor representa una tasa de cambio neto
en la AI de -0.36 ± 0.82% año-1
(Fig. 5B).
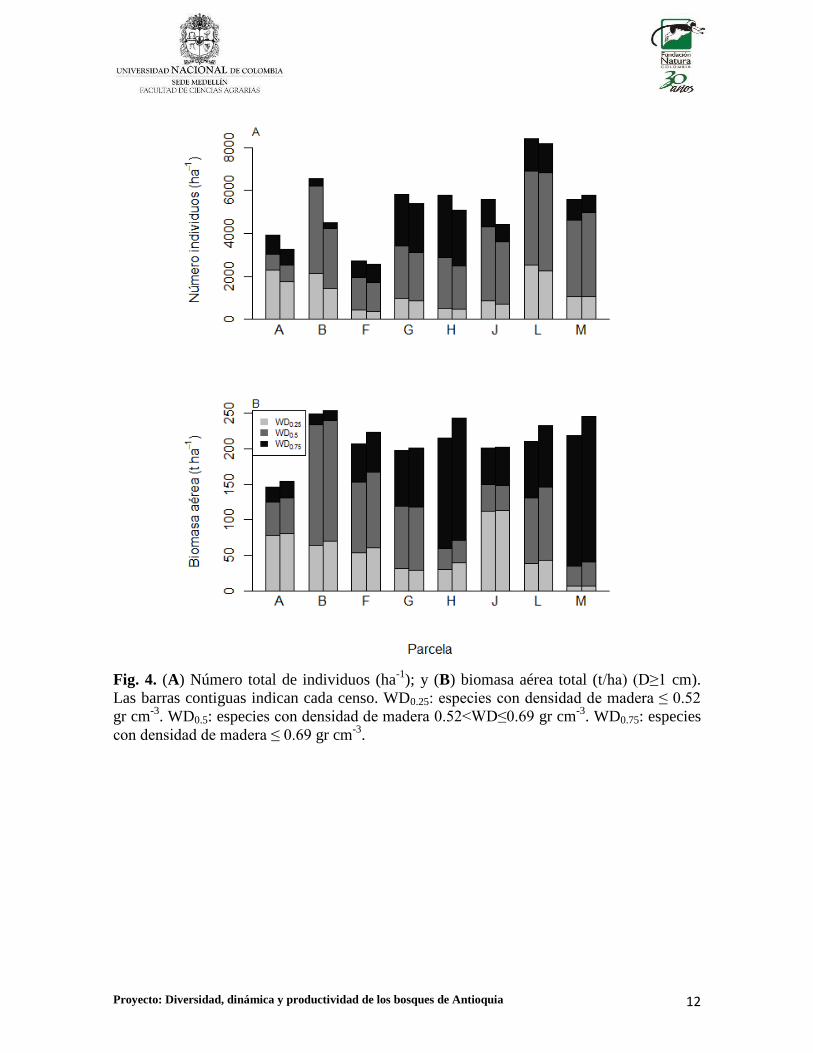
Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 12
Fig. 4. (A) Número total de individuos (ha-1
); y (B) biomasa aérea total (t/ha) (D≥1 cm).
Las barras contiguas indican cada censo. WD0.25: especies con densidad de madera ≤ 0.52
gr cm-3
. WD0.5: especies con densidad de madera 0.52<WD≤0.69 gr cm-3
. WD0.75: especies
con densidad de madera ≤ 0.69 gr cm-3
.

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 13
Fig. 5. Dinámica y cambio neto (% año-1
) del número de individuos (A, B y C) y de la
biomasa aérea (D, E y F), para las categorías de tamaño definidas: todo (D≥1cm; A y D),
dosel (D≥10cm; B y E) y sotobosque (1cm≤D<10cm; C y F).
La cantidad promedio de BA por el contrario, incrementó de 189.05 ± 25.00t ha-1
en el
primer censo a 203.66 ± 29.18t ha-1
en el segundo. El valor promedio de BA, tuvo un
incremento anual promedio de 2.44 ± 1.53t ha-1
año-1
, lo cual equivale a una tasa de cambio
neto de la BA de 1.28 ± 0.76% año-1
(Fig. 5E). La pérdida promedio de individuos por

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 14
mortalidad (10.78 ± 4.97 individuos ha-1
año-1
) fue mayor que la ganancia por reclutamiento
(8.29 ± 3.63 individuos ha-1
año-1
; Fig. 5B). Con relación a la BA de los individuos del
dosel, la mayor parte de la ganancia se dio por efecto del crecimiento (gBA=4.27 ± 1.26t ha-
1 año
-1), pero con una mayor mortalidad (mBA=2.41 ± 1.33 t ha
-1 año
-1) que reclutamiento
(rBA=0.65 ± 0.66t ha-1
año-1
; Fig. 5E).
Con respecto al sotobosque (1cm≤D<10cm), se observó una disminución en el promedio de
la AI entre censos, el cual disminuyó de un valor promedio de 4 933 ± 1 546 individuos ha-
1 en el primer censo a 4 298 ± 1 614 individuos ha
-1 en el segundo. Solo la parcela M
presentó un incremento en el número de individuos del sotobosque. La disminución anual
promedio entre censos en la AI fue de -158.14 ± 178.87 individuos ha-1
año-1
. En total, este
valor representa una tasa de cambio neto en la AI de -3.23 ± 3.33% año-1
(Fig. 5C). De
igual forma, la cantidad promedio de BA, disminuyó de 16.87 ± 5.82t ha-1
en el primer
censo a 16.04 ± 6.05t ha-1
en el segundo. Esta disminución en la BA obedeció
principalmente a la alta disminución en la BA de la parcela J, mientras que las parcelas G,
H, L y M presentaron tasas de cambio neto positivas (Fig. 5F). La disminución promedio
anual de la BA fue de -0.28 ± 0.75t ha-1
año-1
, lo cual equivale a una tasa de cambio neto de
la BA en el sotobosque de 1.61 ± 4.19% año-1
(Fig. 5F). La pérdida total de individuos por
mortalidad (284.51 ± 155.50 individuos ha-1
año-1
) superó la ganancia por reclutamiento
(126.37 ± 107.40 individuos ha-1
año-1
; Fig. 5C). La pérdida promedio de BA por
mortalidad (mBA=0.70 ± 0.54t ha-1
año-1
) en el sotobosque fue superior al crecimiento
(gBA=0.36 ± 0.25t ha-1
año-1
), con una tasa de crecimiento promedio de la BA muy baja
(rBA=0.08 ± 0.09t ha-1
año-1
; Fig. 5F).
Tasas de cambio por categoría de densidad de la madera: Los límites establecidos para
las categorías funcionales según la WD fueron 0.52 gr cm-3
, para el primer cuartil (WD0.25),
0.69 gr cm-3
, para el tercer cuartil (WD0.75), y una media de 0.60 gr cm-3
para el grupo
promedio. La AI por categoría de WD disminuyó entre censos, con un cambio neto
promedio de NAI WD0.25=-3.75 ± 2.69% año-1
, NAI WD0.5=-2.05 ± 2.98% año-1
y NAI
WD0.75=-2.93 ± 3.59% año-1
. En todos los casos, la tasa de mortalidad de individuos
promedio fue superior a la tasa de reclutamiento (Fig. 6A, B y C). En promedio, la BA en
las tres categorías de WD incrementó entre censos, con una tasa de cambio neto de BAn
WD0.25=1.34 ± 2.38% año-1
, BAn WD0.5=0.76 ± 1.28% año-1
y BAn WD0.75=0.94 ± 0.86%
año-1
. La tasa de crecimiento de BA presentó en todos los casos un valor promedio superior
a las tasas de mortalidad y reclutamiento. En promedio, la pérdida promedio anual de BA
por mortalidad (2.29 ± 1.27% año-1
), así como la tasa de crecimiento de BA (2.51 ± 0.47%
año-1
), en la categoría de WD0.5, fue mayor que en las categorías de WD0.25 (mBA= 1.92 ±
1.78% año-1
; gBA= 2.38 ± 1.14% año-1
) y WD0.75 (mBA= 1.11 ± 0.63% año-1
; gBA= 1.92 ±
0.40% año-1
), respectivamente. Por el contrario, la tasa de reclutamiento fue mayor en las
especies de la categoría WD0.25 (rBA= 0.95 ± 1.82% año-1
) que en las categorías de WD0.5
(rBA= 0.62 ± 0.43% año-1
) y WD0.75 (rBA= 0.15 ± 0.13% año-1
; Fig. 6D, E y F). Sin
embargo, no se encontraron diferencias significativas en el promedio de las tasas de

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 15
mortalidad, reclutamiento, crecimiento e incremento neto de la BA observado en cada
parcela, entre categorías de WD (mBA: F-Value=1.673; P-Value=0.212; rAI: F-Value=1.098;
P-Value=0.352; gAI: F-Value=1.403; P-Value=0.268; NAI: F-Value=0.261; P-
Value=0.773).
Fig 6. Dinámica y cambio neto (% año-1
) del número de individuos (A, B y C) y de la
biomasa aérea (D, E y F), para los grupos de especies definidos a partir de la densidad de la
madera: WD0.25 (A y D), WD0.5 (B y E) y WD0.75 (C y F).

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 16
DISCUSIÓN
En promedio, el incremento neto en la BA de los bosques del área de estudio (D≥1
cm=2.23 ± 1.77 t ha-1
año-1
; y dosel D≥10 cm=2.44 ± 1.53t ha-1
año-1
) fue superior a lo
reportado para bosques tropicales de tierras bajas de la Amazonía (Baker et al., 2004;
Chave et al., 2008; Phillips et al. 2008; Brienen et al., 2015). Si asumimos que el 50% de la
biomasa seca es carbón, los bosques monitoreados acumulan en promedio 1.11 tC ha-1
año-
1. El incremento en la BA fue direccionado principalmente por el crecimiento de los árboles
de dosel, en donde el promedio de la tasa de crecimiento de la BA (2.23 ± 0.50% año-1
) fue
superior a la tasa de mortalidad (1.25 ± 0.65% año-1
). Los individuos del sotobosque por su
parte, disminuyeron BA a una tasa de cambio neto promedio de -1.61 ± 4.19% año-1
, lo
cual obedeció principalmente a la alta pérdida de BA por mortalidad (4.21 ± 2.84% año-1
).
Este resultado, resalta la importancia de la inclusión de árboles con D<10 cm para la
cuantificación de la BA y su dinámica en bosques tropicales (Valencia et al., 2009). Según
los resultados obtenidos, la no inclusión del sotobosque, puede conducir a una sobre
estimación del 11% en el cambio neto de la BA en el área de estudio.
El recambio de individuos, contrario a lo reportado para los bosques de tierras bajas de la
Amazonia (Phillips et al., 2004), presentó una tendencia negativa (-2.83 ± 2.77% año-1
). Si
bien dicha tendencia fue consistente en las dos categorías diamétricas definidas, se observó
una mayor pérdida de individuos en el sotobosque, donde la tasa promedio de mortalidad
dobló la tasa de reclutamiento. Las diferencias en las tasas de mortalidad entre el dosel y el
sotobosque, son similares a lo reportado recientemente para los bosques de la Amazonía
(Brienen et al., 2015), donde se encontró un incremento en la pérdida de BA por
mortalidad, la cual es determinada principalmente por la mortalidad de árboles pequeños.
No obstante, en el presente estudio las altas tasas de mortalidad aún no parecen afectar las
tasas de acumulación de BA. En general, se asume que altas tasas de mortalidad pueden
emerger como consecuencia de la presencia de condiciones desfavorables para el
crecimiento de los árboles (Chave et al., 2008). Por ejemplo, es probable que factores
externos, como la sequía provocada por el fenómeno del niño del año 2010, sea uno de los
mecanismos que permita explicar la alta mortalidad observada en estos bosques. Esta
explicación es consistente con los resultados reportados para los bosques de la Amazonía
(Phillips et al., 2009), los cuales indican que los árboles tropicales son vulnerables al estrés
por sequías prolongadas. Sin embargo, dado que en el área de estudio se presentó una
mayor mortalidad en los individuos del sotobosque y en las especies de la categoría de
WD0.25, las altas tasas de mortalidad parecen estar en parte asociadas a una reducción en la
disponibilidad de luz en el sotobosque, como consecuencia de un mayor crecimiento en la
dominancia de árboles del dosel (Lewis et al., 2004).
Las tendencias encontradas en la dinámica de la BA y la AI en las categorías de tamaño y
de WD definidas, permiten asumir que en estos bosques se está presentando una
disminución en los individuos con valores bajos de WD, así como un incremento en la
abundancia y dominancia de árboles grandes. Con respecto a la dinámica de la BA, en los

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 17
individuos del dosel se observó una alta tasa de crecimiento de BA y poca pérdida por
mortalidad. Mientras que en lo relacionado con la AI, el cambio neto de individuos
presentó mayor disminución en el sotobosque (-3.23 ± 3.33% año-1
) y en las especies de la
categoría WD0.25 (-3.75 ± 2.69% año-1
). En consecuencia, los resultados encontrados
indican que los bosques del área de estudio se encuentran en un proceso de recuperación de
un disturbio pasado. No obstante, la dinámica de la BA y la AI presentó variaciones
importantes entre algunas de las parcelas monitoreadas (Fig. 5 y 6), lo cual puede ser una
consecuencia de la alta variación en el régimen de perturbación natural de los bosques
(Baker et al., 2004; Marvin et al., 2014). Por ejemplo, en tres de las ocho parcelas se
encontró un cambio neto de la BA inferior a 1.00t ha-1
año-1
(parcelas B, G y J; Fig. 5D).
En el caso de las parcelas B y G, este patrón fue direccionado principalmente por la
mortalidad de varios individuos de gran porte. Mientras que en la parcela J, esta tendencia
es direccionada principalmente por la alta pérdida de BA en el sotobosque, como
consecuencia del alto número de árboles pequeños muertos por la caída de dos individuos
del dosel.
Otra fuente importante de variabilidad en el área de estudio es la relacionada con las
diferencias en la composición florística de las parcelas, lo cual es notorio en el aporte que
realizan las tres categorías de especies definidas a partir de la WD a la AI y la BA (Fig. 4).
Por ejemplo, en las parcelas M y H, las especies pertenecientes a la categoría WD0.75
aportan más del 70% de la BA total de las parcelas, mientras que las especies con WD0.25
aportaron 3% y 14% de la BA total, respectivamente (Fig. 4B). Entre las especies más
representativas en las parcelas, se destaca la gran abundancia de individuos y el alto aporte
en BA que realizó la especie Quercus humboldtii (Bonpl.), la cual es reconocida en los
bosques de alta montaña por su gran porte, madera valiosa y adaptación natural a las bajas
temperaturas. Esta especie contribuyó en el segundo censo con el 13% del total de la BA
acumulada en todas las parcelas y el 87% de la BA en la parcela M. En la región en la cual
se encuentra la parcela M y en menor proporción la parcela L, municipios de Belmira y
Anorí respectivamente, se encuentran fragmentos de bosque con importantes asociaciones
de Q humboldtii, los cuales según los resultados del presente estudio tienen un alto
potencial de acumulación de carbono en la BA. En conclusión, las diferencias observadas
en la dinámica de la BA y la AI entre las parcelas, así como diferencias en la composición
florística como lo indicado anteriormente, resaltan la alta variabilidad espacial de los
bosques andinos, lo cual limita la comprensión de su estructura y la variación de la BA a
una escala regional (Marvin et al., 2014). En este sentido, los resultados del presente
estudio resaltan la importancia de contar con muestreos lo suficientemente robustos, ya sea
en área y/o en el número de parcelas, que permitan capturar la variación espacial y temporal
de los bosques tropicales.
Por último, se puede asumir que los bosques del área de estudio actualmente actúan como
un sumidero importante de carbono. No obstante, es pertinente continuar con su monitoreo,
con el fin de corroborar si las altas tasas de acumulación de BA se mantienen en el tiempo,

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 18
o si por el contrario se presenta una disminución en estas, similar a lo reportado para otros
bosques tropicales (Brienen et al., 2015). Los resultados del presente estudio resaltan la
importancia de los bosques andinos, no solo en lo relacionado con la acumulación de
carbono, sino también por su importancia como refugio de la flora (Duque et al., 2014). En
este sentido, es importante emprender acciones encaminadas a garantizar su conservación y
el entendimiento de su dinámica, más aún si tenemos en cuenta la alta intervención
antrópica a la cual han estado sometidos históricamente estos bosques (Cabrera et al.,
2011). Adicionalmente, las tendencias relacionadas con la dinámica de la BA, corroboran el
gran potencial que estos ecosistemas tienen para la implementación de programas
relacionados con el pago por servicios ambientales, tales como la captura y regulación del
ciclo de carbono.
BIBLIOGRAFIA
Álvarez, E., Duque, A., Saldarriaga, J., Cabrera, K., De Las Salas, G., Del Valle, I., Lema,
A., Moreno, F., Orrego, S. & Rodríguez, L. (2012). Tree above-ground biomass allometries
for carbon stocks estimation in the natural forests of Colombia. Forest Ecology and
Management, 267, 297-308.
Baker, T. R., Phillips, O. L., Malhi, Y., Almeida, S., Arroyo, L., Di Fiore, A., Erwin, T.,
Higuchi, N., Killeen, T. J., Laurance, S. G., Laurance, W. F., Lewis, S. L., Monteagudo, A.,
Neill, D. A., Núñez Vargas, P., Pitman, N. C. A., Silva, J. N. M. & Vásquez Martínez, R.
(2004). Increasing biomass in Amazonian forest plots. Philosophical Transactions of the
Royal Society of London. Series B: Biological Sciences, 359, 353-365.
Brienen, R. J. W., Phillips, O. L., Feldpausch, T. R., Gloor, E., Baker, T. R., Lloyd, J.,
Lopez-Gonzalez, G., Monteagudo-Mendoza, A., Malhi, Y., Lewis, S. L., Vásquez
Martinez, R., Alexiades, M., Álvarez Dávila, E., Alvarez-Loayza, P., Andrade, A., C.
Aragao, L. E. O., Araujo-Murakami, A., M. Arets, E. J. M., Arroyo, L., Aymard G. A.,
Bánki, O. S., Baraloto C., Barroso, J., Bonal, D., Boot, R. G. A., Camargo, J. L. C.,
Castilho, C. V., Chama, V., Chao, K. J., Chave, J., Comiskey, J. A., Cornejo Valverde, F.,
da Costa, L., de Oliveira, E. A., Di Fiore, A., Erwin, T. L., Fauset, S., Forsthofer, M.,
Galbraith, D. R., Grahame, E. S., Groot, N., Hérault, B., Higuchi, N., Honorio Coronado, E.
N., Keeling, H., Killeen, T. J., Laurance, W. F., Laurance, S., Licona, J., Magnussen, W. E.,
Marimon, B. S., Marimon-Junior, B. H., Mendoza, C., Neill, D. A., Nogueira, E. M.,
Núñez, P., Pallqui Camacho, N. C., Parada, A., Pardo-Molina, G., Peacock, J., Peña-Claros,
M., Pickavance, G. C., Pitman, N. C. A., Poorter, L., Prieto, A., Quesada, C. A., Ramírez,
F. Ramírez-Angulo, H., Restrepo, Z., Roopsind, A., Rudas, A., Salomao, R. P., Schwarz,
M. Silva, N., Silva-Espejo, J. E., Silveira, M., Stropp, J., Talbot, J., ter Steege, H., Teran-
Aguilar, J., Terborgh, J., Thomas-Caesar, R., Toledo, M., Torello-Raventos, M., Umetsu,
R. K., van der Heijden, G. M. F., van der Hout, P., Guimaraes Vieira, I. C., Vieira, S. A.,

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 19
Vilanova, E., Vos, V. A. & R. J. Zagt (2015). Long-term decline of the Amazon carbon
sink. Nature, 519(7543), 344-348.
Cabrera, E., Vargas, D. M., Galindo, G., García, M. C., Ordoñez, M. F., Vergara, L. K., &
Giraldo, P. (2011). Memoria técnica de la cuantificación de la deforestación histórica
nacional - escalas gruesa y fina [PDF version]. Retrieve from
https://www.siac.gov.co/documentos/DOC_Portal/DOC_Bosques/131112_MT_Cuantif_Ta
sa_Deforestacion.pdf.
Chave, J., Muller-Landau, H. C., Baker, T. R., Easdale, T. A., Ter Steege, H. & Webb, C.
O. (2006). Regional and phylogenetic variation of wood density across 2456 Neotropical
tree species. Ecological Applications, 16, 2356-2367.
Chave, J., Condit, R., Muller-Landau, H. C., Thomas, S. C., Ashton, P. S.,
Bunyavejchewin, S., Co, L. L., Dattaraja, H. S., Davies, S. J., Esufali, S., Ewango, C. E. N.,
Feeley, K. J., Foster, R. B., Gunatilleke, N., Gunatilleke, S., Hall, P., Hart, T. B.,
Hernández, C., Hubbell, S. P., Itoh, A., Kiratiprayoon, S., LaFrankie, J. V., de Lao, S. L.,
Makana, J. R., Supardi Noor, Md. N., Rahman-Kassim, A., Samper, C., Sukumar, R.,
Suresh, H. S., Tan, S., Thompson, J., Tongco, M. D. C., Valencia, R., Vallejo, M., Villa,
G., Yamakura, T., Zimmerman, J. K. & Losos, E. C. (2008). Assessing evidence for a
pervasive alteration in tropical tree communities. PLoS Biology, 6, 455-462.
Chave, J., Réjou‐Méchain, M., Búrquez, A., Chidumayo, E., Colgan, M. S., Delitti, W. B.,
Duque, A., Eid, T., Fearnside, P. M., Goodman, R. C., Henry, M., Martínez-Yrízar, A.,
Mugasha, W. A., Muller-Landau, H. C., Mencuccini, M., Nelson, B. W., Ngomanda, A.,
Nogueira, E. M., Ortiz-Malavassi, E., Pélissier, R., Ploton, P., Ryan, C. M., Saldarriaga, J.
G. & Vieilledent, G. (2014). Improved allometric models to estimate the aboveground
biomass of tropical tres. Global Change Biology, 20, 3177-3190.
Clark, D. A. (2007). Detecting tropical forests responses to global climatic and atmospheric
change: current challenges and a way forward. Biotropica, 39, 4-19.
Condit, R. (1998). Tropical forest census plot. Springer-verlag, Berlin, and R.G. Landes
Company, Georgetown, Texas.
Condit, R., Aguilar, S., Hernandez, A., Perez, R., Lao, S., Angehr, G., Hubbell, S. P. &
Foster, R. B. (2004). Tropical forest dynamics across a rainfall gradient and the impact of
an El Nino dry season. Journal of Tropical Ecology, 20, 51-72.
Duque, A., Feeley, K. J., Cabrera, E., Callejas, R., & Idarraga, A. (2014). The dangers of
carbon-centric conservation for biodiversity: a case study in the Andes. Tropical
Conservation Science, 7, 178-191.
Feeley, K. J., Wright, S. J., Nur Supardi, M. N., Kassim, A. R. & Davies, S. J. (2007).
Decelerating growth in tropical forest trees. Ecology Letters, 10, 461-469.

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 20
Feldpausch, T., Lloyd, T., Lewis, S. L., Brienen, R. J. W., Gloor, E., Monteagudo
Mendoza, A., Lopez-Gonzalez, G., Banin, L., Abu Salim, K., Affum-Baffoe, K., Alexiades,
M., Almeida, S., Amaral, I., Andrade, A., Aragâo, L. E. O. C., Araujo Murakami, A., Arets,
E. J. M. M., Arroyo, L., Aymard, C., G. A., Baker, T. R., Bánki, O. S., Berry, N. J.,
Cardozo, N., Chave, J., Comiskey, J. A., D´avila, E. A., de Oliveira, A., DiFiore, A.,
Djagbletey, G., Domingues, T. F., Erwin, T. L., Fearnside, P. M., Franca, M. B., Freitas, M.
A., Higuchi, N., Honorio, C. E., Iida, Y., Jiménez, E., Kassim, A. R., Killeen, T. J.,
Laurance, W. F., Lovett, J. C., Malhi, Y., Marimon, B. S., Marimon-Junior, B. H., Lenza,
E., Marshall, A. R., Mendoza, C., Metcalfe, D. J., Mitchard, E. T. A., Nelson, B. W., Nilus,
R., Nogueira, E. M., Parada, A., Peh, K. S. H., Pena Cruz, A., Peñuela, M. C., Pitman, N.
C. A., Prieto, A., Quesada, C. A., Ramírez, F., Ramírez-Angulo, H., Reitsma, J. M., Rudas,
A., Saiz, G., Salomâo, R. P., Schwarz, M., Silva, N., Silva-Espejo, J. E., Silveira, M.,
Sonké, B., Stropp, J., Taedoumg, H. E., Tan, S., ter Steege, H., Terborgh, J., Torello-
Raventos, M., van der Heijden, G. M. F., Vásquez, R., Vilanova, E., Vos, V., White, L.,
Wilcock, S., Woell, H. & Phillips, O. L. (2012). Tree height integrated into pantropical
forest biomass estimates. Biogeosciences, 9, 3381-3403.
Lewis, S. L., Phillips, O. L., Baker, T. R., Lloyd, J., Malhi, Y., Almeida Higuchi, N.,
Laurance, W. F., Neill, D. A., Silva, J. N. M., Terborgh, J., Torres Lezama, A., Vásquez
Martínez, R., Brown, S., Chave, J., Kuebler, C., Núñez Vargas, P. & Vinceti, B. (2004).
Concerted changes in tropical forest structure and dynamics: evidence from 50 South
American long-term plots. Philosophical Transactions of the Royal Society of London.
Series B: Biological Sciences, 359, 421-436.
Lewis, S. L., Lopez-Gonzalez, G., Sonke, B., Affum-Baffoe, K., Baker, T. R., Ojo, L. O.
Phillips, O. L., Reitsma, J. M., White, L., Comiskey, J. A., Djuikouo, M., Ewango, C. E.
N., Feldpausch, T. R., Hamilton, A. C., Gloor, M., Hart, T., Hladik, A., Lloyd, J., Lovett, J.
C., Makana, J. R., Malhi, Y., Mbago, F. M., Ndangalasi, H. J., Peacock, J., Peh, K. S. H.,
Sheil, D., Sunderland, T., Swaine, M. D., Taplin, J., Taylor, D., Thomas S. C., Votere, R. &
Wöll, H. (2009) Increasing carbon storage in intact African tropical forests. Nature, 457,
1003-1006.
Malagón, D. (2003). Ensayo sobre tipología de suelos colombianos, énfasis en génesis y
aspectos ambientales. Revista de la academia Colombiana de ciencias exactas, físicas y
naturales, 27, 319-341.
Marvin, D. C., Asner, G. P., Knapp, D. E., Anderson, C. B., Martin, R. E., Sinca, F. &
Tupayachi, R. (2014). Amazonian landscapes and the bias in field studies of forest structure
and biomass. Proceedings of the National Academy of Sciences, 111, E5224-E5232.
Oliver, C. D. & Larson, B. C. (1990). Forest Stand Dynamics. McGraw-Hill, New York.
Pan, Y., Birdsey, R. A., Fang, J., Houghton, R., Kauppi, P. E., Kurz, W. A., Phillips, O. L.,
Shvidenko, A., Lewis, S. L., Canadell, J. G., Ciais, P., Jackson, R. B., Pacala, S. W.,

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 21
McGuire, D., Piao, S., Rautiainen, A., Sitch, S. & Hayes, D. (2011). A large and persistent
carbon sink in the world’s forests. Science, 333, 988-993.
Peña, M. A. & Duque, A. (2013). Patterns of stocks of aboveground tree biomass,
dynamics, and their determinants in secondary Andean forests. Forest Ecology and
Management, 302, 54-61.
Phillips, O. L. & Gentry, A. H. (1994). Increasing turnover through time in tropical forest.
Science, 263, 954-958.
Phillips, O. L., Hall, P., Gentry, A. H., Sawyer, S. A. & Vásquez, R. (1994). Dynamics and
richness of tropical rain forest. Proceedings of the National Academy of Sciences of the
United States of America, 91, 2805-2809.
Phillips, O. L., Malhi, Y., Vinceti, B., Baker, T., Lewis, S. L., Higuchi, N., Laurance, W.
F., Núñez Vargas, P., Vásquez Martinez, R., Laurance, S., Ferreira, L. V., Stern, M.,
Brown, S. & Grace, J. (2002). Changes in growth of tropical forests: evaluating potential
biases. Ecological Applications, 12, 576-587.
Phillips, O. L., Baker, T. R., Arroyo, L., Higuchi, N., Killeen, T., Laurance, W. F., Lewis,
S. L., Lloyd, J., Malhi, Y., Monteagudo, A., Neill, D., Núñez-Vargas, P., Silva, N.,
Terborgh, J., Vásquez-Martínez, R., Alexiades, M., Almeida, S., Brown, S., Chave, J.,
Comiskey, J. A., Czimczik, C. I., Di Fiore, A., Erwin, T., Kuebler, C., Laurance, S. G.,
Nascimento, H. E. M., Palacios, W., Patiño, S., Pitman, N., Olivier, J., Quesada, C. A.,
Saldias, M., Torres-Lezama, A. & Vinceti, B. (2004). Pattern and process in Amazon tree
turnover, 1976-2001. Philosophical Transactions of the Royal Society B: Biological
Sciences, 359, 381-407.
Phillips, O. L., Lewis, S. L., Baker, T. R., Chao, K. J. & Higuchi, N. (2008). The changing
Amazon forest. Philosophical Transactions of the Royal Society B: Biological Sciences,
363, 1819-1827.
Phillips, O. L., Aragão, L., Lewis, S. L., Fisher, J. B., Lloyd, J., López-González, G.,
Malhi, Y., Monteagudo, A., Peacock, J., Quesada, C. A., van der Heijden, G., Almeida, S.,
Amaral, I., Arroyo, L., Aymard, G., Baker, T. R., Bánki, O., Blanc, L., Bonal, D., Brando,
P., Chave, J., de Oliveira, A. C. A., Cardozo, N. D., Czimczik, C. I., Feldpausch, T. R.,
Freitas, M. A., Gloor, E., Higuchi, N., Jiménez, E., Lloyd, G., Meir, P., Mendoza, C.,
Morel, A., Neill, D. A., Nepstad, D., Patiño, S., Peñuela, M. C., Prieto, A., Ramírez, F.,
Schwarz, M., Silva, J., Silveira, M., Thomas, A. S., Ter Steege, H., Stropp, J., Vásquez, R.,
Zelazowski, P., Alvarez Dávila, E., Andelman, S., Andrade, A., Chao, K., Erwin, T., Di
Fiore, A., Honorio, C. E., Keeling, H., Killeen, T. J., Laurance, W. F., Peña Cruz, A.,
Pitman, N. C. A., Núñez Vargas, P., Ramírez-Angulo, H., Rudas, A., Salamão, R., Silva,
N., Terborgh, J. & Torres-Lezama, A. (2009). Drought sensitivity of the Amazon rainforest.
Science, 323, 1344-1347.

Proyecto: Diversidad, dinámica y productividad de los bosques de Antioquia 22
Saatchi, S. S., Harris, N. L., Brown, S., Lefsky, M., Mitchard, E. T. A., Salas, W., Zutta, B.
R., Buermann, W., Lewis, S. L., Hagen, S., Petrova, S., White, L., Silman, M. & Morel, A.
(2011). Benchmark map of forest carbon stocks in tropical regions across three continents.
Proceedings of the National Academy of Sciences of the United States of America, 108,
9899-9904.
Sierra, C. A., del Valle, J. I., Orrego, S. A., Moreno, F. H., Harmon, M. E., Zapata, M.,
Colorado, G. J., Herrera, M. A., Lara, W., Restrepo, D. E., Berrouet, L. M., Loaiza, L. M.
& Benjumea, J. F. (2007). Total carbon stocks in a tropical forest landscape of the Porce
region, Colombia. Forest Ecology and Management, 243, 299-309.
Valencia, R., Condit, R., Muller-Landau, H. C., Hernandez, C. & Navarrete, H. (2009).
Dissecting biomass dynamics in a large Amazonian forest plot. Journal of Tropical
Ecology, 25, 473-482.
Wright, S. J., Kitajima, K., Kraft, N. J. B., Reich, P. B., Wright, I. J., Bunker, D. E., Condit,
R., Dalling, J. W., Davies, S. J., Díaz, S., Engelbrecht, B. M. J., Harms, K. E., Hubbell, S.
P., Marks, C. O., Ruiz-Jaen, M. C., Salvador, C. M. & Zanne, A. E. (2010). Functional
traits and the growth–mortality trade-off in tropical trees. Ecology, 91, 3664-3674.
Yepes, A., Herrera, J., Phillips, J. F., Cabrera, E., Galindo, G., Granados, E., Duque, A.,
Barbosa, A., Olarte, C. & Cardona, M. (2015). Contribución de los bosques tropicales de
montaña en el almacenamiento de carbono en Colombia. International Journal of Tropical
Biology and Conservation, 63, 69-82.
Zanne, A. E., López-González, G., Coomes, D. A., Ilic, J., Jansen, S., Lewis, S. L., Miller,
R. B., Swenson, N. G., Wiemann, M. C. & Chave, J. (2009). Data from: Towards a
worldwide wood economics spectrum. Dryad Digital Repository. doi:10.5061/dryad.234.





























