Embed Size (px)
Citation preview

JOURNAL OF APPLIED BIOMECHANICS, 1996,12,338-358 O 1996 by Human Kinetics Publishers, Inc.
Difierences Between One-Foot and Two-Foot
Vertical Jump Performances
Peter F. Vint and Richard N . Hinrichs
The purpose of this investigation was to quantify the differences between one- and two-foot vertical jumping performances. Fourteen subjects performed both jump styles with a four-step, self-paced approach. While overall jump and reach heights were similar between one-foot and two-foot jumps, the strategies employed to achieve these results were notably different. One-foot jumps benefited from an increased takeoff height that was largely attributable to the elevation of the free swinging leg. Further, it was suggested that the actions of this limb may have helped slow the rate of extension of the support leg during the propulsion phase. Greater flight heights were achieved during two-foot jumps, as expected, but the magnitude of this difference was only about 9 cm. It was suggested that factors associated with the development of muscular tension, vertical velocity at touchdown, and horizontal approach speed may have all contributed to the unexpectedly small differences in flight height between one-foot and two-foot jumping performances.
Jumping is a fundamental aspect of several sports including basketball, volleyball, and track and field. In addition, vertical jumping has often been used as a crude method for evaluating anaerobic muscle power (Fox, Bowers, & Foss, 1988; Sargent, 1921). Thus, jumping has received a great deal of attention from researchers in biomechanics and physi- ology. A vast body of literature has documented numerous investigations of jumping me- chanics and the determinant factors of jumping performance. While several researchers have examined biomechanical aspects of either one-foot vertical jumping (Dapena, 1980a, 1980b; Dapena & Chung, 1988; Dapena, McDonald, & Cappaert, 1990) or two-foot ver- tical jumping (Ae & Shibukawa, 1980; Bobbert, Mace, Schinkelshoek, Huijing, & Ingen Schenau, 1986; Coutts, 1982; Harman, Rosenstein, Frykman, & Rosenstein, 1990; Komi & Bosco, 1978), few have specifically investigated the differences between the two styles. Therefore, the purpose of this investigation was to quantify the differences between one- and two-foot maximum effort vertical jumping performances. It was expected that one- foot jumps would benefit from the actions of the free swinging leg during the ground support phase and from the elevation of this limb at the instant of takeoff. Two-footjumps, with essentially twice the available leg musculature to produce vertical impulses against the ground, were expected to produce greater vertical velocities at takeoff and, therefore, greater elevations of the whole body center of mass (CM) during the flight phase.
The authors are with the Exercise and Sport Research Institute, Arizona State University, Tempe, AZ 85287-0404.

Vertical Jump Performances 339
Review of Literature
Soest, Roebroek, Bobbert, Huijing, and Ingen Schenau (1985) compared one- and two- foot vertical jumping performances among 10 competitive male volleyball athletes. All jumps were performed from a standing start with a countermovement but without the use of an arm swing. In one-foot jumps, swinging of the contralateral leg was prohibited. One would expect that one-foot jump heights would be half the value of two-foot jump heights since only half of the total leg musculature is allowed to produce vertical impulses against the ground. However, Soest et al. concluded that one-foot jump height was 58.5% of two- foot jump height-a value significantly greater than 50%. The authors suggested that the difference between one- and two-foot jumps was attributable to a neural mechanism that inhibited the expression of maximal strength during the two-footjumps (Soest et al., 1985). This inhibiting mechanism, sometimes called the "bilateral deficit," has also been demon- strated in more simple tasks including unilateral versus simultaneous bilateral isometric leg extension activities (Howard & Enoka, 1991; Koh, Grabiner, & Clough, 1993; Schantz, Moritani, Karlson, Johansson, & Lundh, 1989; Vandervoort, Sale, & Moroz, 1984).
Saunders (1980) examined the effects of approach speed on one- and two-foot ver- tical jump performances. In the study, 3 volleyball players and 3 basketball players were asked to complete two-foot and one-foot jumps, respectively. During each of 10 jumps, athletes were asked to initiate approach runs of various speeds. These approach runs were expressed as a percentage of each athlete's maximum sprinting speed. Saunders (1980) concluded that vertical velocities of two-foot jumps increased with approach speeds up to 50-60% of maximum sprint speed. Similarly, vertical velocities of one-foot jumps in- creased with approach speeds up to 60-70% of maximum sprint speed. In both cases, vertical jump performances decreased after the "optimal" approach speeds were exceeded. Dapena et al. (1990) reported similar decrements in high-jump performances when opti- mal approach speeds were exceeded.
In general, however, studies have shown that vertical jump performance increases when an approach run precedes the jump. Healy (1977) reported that one-half-step and two-step approaches yielded significantly greater vertical jump heights than did jumps performed without an approach. Overall, it was found that the two-step approach yielded the greatest jump height compared to the other approach conditions. Kayambashi (1977) reported that three-step and four-step approaches resulted in greater jump heights than did two-step approaches. Similarly, Enoka (1971) reported that jumps following approach runs of one, three, and five steps resulted in higher jump and reach scores than jumps initiated from a standing position.
With regard to more conventional standing two-foot vertical jump performances, several researchers have demonstrated that a properly performed countermovement will contribute to significant increases in jump height (Asmussen & Bonde-Petersen, 1974; Cavagna, Dusman, & Margaria, 1968; Harman et al., 1990; Komi & Bosco, 1978). Dur- ing the descent of the countermovement, the hips, knees, and ankles are flexed into posi- tions which stretch the muscles that will later act to extend those same joints. It has been theorized that besides improving the force-producing capacities of the muscle itself, the countermovement utilizes some of the elastic properties of muscles and tendons (Bobbert et al., 1986; Komi & Bosco, 1978); increases the distance over which force can be ex- erted, thereby prolonging the upward propulsion phase (Anderson & Pandy, 1993; Pandy & Zajac, 1991; Zajac, 1993); and takes up some of the muscular slack that is associated with the initial stages of the development of muscular tension (Ingen Schenau, 1984; Zajac, 1993).

340 Vint and Hinrichs
Dapena and Chung (1988) noted that during one-foot high-jump performances, ver- tical velocity of the CM was positive throughout all but the first 0.01 s of the takeoff phase of the jump. The fact that the CM moved upward despite continued flexion of the support leg seemed to indicate that the mechanism by which one-foot jumps load the support leg musculature is not at all similar to the traditional countermovement action that is often performed during two-foot jumps. Rather, Dapena and Chung (1988) suggested that one- foot jumps may have derived a beneficial eccentric loading effect from the radial motion of the CM moving over the base of support. The radial motions of the segmental and whole-body centers of mass, with respect to the support foot, may have helped develop faster eccentric or slower concentric conditions of the support leg musculature. This would have helped maintain the active musculature in a more favorable region of the force- velocity relationship (Fenn & Marsh, 1935; Komi, 1973; Perrine & Edgerton, 1978).
Deterministic Factors of Vertical Jumping
In a maximal-effort vertical jump performance, the goal of the task is simply to jump and reach as high as possible. In the absence of air resistance and other external forces, the upward projection of the whole-body CM is completely determined by the vertical veloc- ity at the instant of takeoff and the acceleration due to gravity. However, this quantity does not completely describe the overall jump and reach height that is observed.
A simple deterministic model (Hay & Reid, 1988) can be used to show that the maximum height to which an individual can jump and reach may be described by the sum of four lesser heights: takeoff height, flight height reach height, and loss height (see Fig- ure I). Takeoff height may be defined as the height of the CM at the instant the individual leaves the ground. Flight height refers to the actual height to which the CM is elevated during the in-flight phase of the jump. Reach height describes the vertical distance from the CM to the fingertips at the instant the maximum jump and reach height is evaluated. Loss height refers to the difference between the peak height of the CM and the height of the CM at the instant the maximum jump and reach height is evaluated. This last factor can usually be attributable to a mistiming of the final reach.
In standing two-foot vertical jumps (with arm swing and countermovement), take- off height, flight height, and reach height account for 41%, 17%, and 42% of the overall jump and reach height, respectively. Loss height is negligible, accounting for about 0.2% of the overall jump and reach height (Hinrichs & Vint, 1994). It may be surprising to note that the contribution of flight height is so much smaller than that of takeoff height and reach height. Regardless, it is clear that an overwhelming percentage of the overall jump and reach height is determined not by the vigorous muscular effort required to propel the body upward but simply by the position and orientation of the body about the CM at the instant of takeoff and again when the maximum height is evaluated.
Procedures
Fourteen male physical education students (mean age 22.5 years; height 181.6 cm and mass 80.1 kg with shoes on) volunteered to participate as subjects in the experiment. In- formed consent was obtained from each subject prior to testing. Following a brief warm-up and stretching period, each subject was asked to perform a block of five one-foot and five two-foot maximum-effort vertical jumps. For both jump styles, subjects were allowed to use a four-step, self-paced approach run-up. Jump and reach heights were measured for each trial using a simple vertical jump height measuring device (Vertec, Sports Imports, Columbus, OH). Subjects were asked to jump and reach as high as possible and contact the Vertec with

Vertical Jump Performances
Jump and Reach
Takeoff Height
Jump and Reach I / Height
v/ W Reach Height
Figure 1 -Deterministic model and illustration of simple factors that determine overall jump and reach height (following guidelines of Hay & Reid, 1988). Note that relative i f the four subheights are drawn approximately to scale. Refer to Table 1 for exact proportions. From "Vertical Jumping Performance: One-Foot Vs. Two-Foot Takeoff Techniques," by P. Vint, 1996, Performance and Conditioning for Volleyball, 3, pp. 4-7. Copyright 1996 by Conditioning Press, P.O. Box 6819, Lincoln, NE 68506.
their dominant hand. For each jump style and for each subject, the trial that yielded the highest measured jump and reach height was considered for further analysis.
Two-dimensional video data were collected at 60 Hz in the sagittal plane. APanasonic model D5 100 video camera was placed approximately 30 m away from the subjects in an effort to reduce perspective error. Selected jumps were digitized with the Peak Perfor- mance motion measurement system (Englewood, CO). Digitizing began approximately 10 video fields before the last heel strike of the approach and ended approximately 10 video fields after the vertical jump and reach measurement device was contacted. If, dur- ing two-foot jumps, subjects planted one foot and then the other, digitizing was initiated approximately 10 video fields before the final heel strike of the first foot to contact the ground. This asynchronous foot planting technique (termed the "step-close" technique by Coutts, 1982) was observed to be the norm rather than the exception.
Mathematical Model and Body Segment Parameters
Twenty-one points were used to define a 14-segment model of the body. In addition, the fingertip of the dominant hand was digitized at the instant the Vertec was contacted. Seg-

342 Vint and Hinrichs
mental masses and center of mass locations obtained from the mean data of Clauser, McConville, and Young (1969) as adjusted by Hinrichs (1990) were used to compute the whole-body CM location throughout the entire trial. The centers of mass for various sys- tems of segments were computed from the general equation:
P = (Crn,Pi)J(Crni> (1)
where m, is the mass of the ith segment, and P, and P are the vectors locating the centers of mass of the ith segment and the system of segments, respectively, relative to the fixed 0:XY reference frame. The various systems of segments that were defined in this study included the head and trunk, right arm, left arm, both arms, right leg, left leg, both legs, and the whole body.
Smoothing and Differentiation
Raw two-dimensional data were rotated into a true vertical-horizontal lab reference frame. To reduce the endpoint effects of the Butterworth digital filter, all coordinate data were padded with 20 points on either side of the original array using a linear extrapolation scheme (Smith, 1989). Horizontal (X) and vertical (Y) coordinates of each data point were smoothed with a fourth order, zero-lag Butterworth digital filter (Winter, Sidwell, & Hohson, 1974). Cutoff frequencies ranged from 3.25 to 9.75 Hz and were selected auto- matically by an algorithm based on the method of Wells and Winter (1980). All velocity data were derived from finite difference differentiation of the smoothed position data (Miller & Nelson, 1973). Velocity data were not subjected to further smoothing.
Kinematic Factors
Takeoff height, flight height, reach height, and loss height were calculated using the ver- tical position of the CM at the appropriate events. Average horizontal approach speed was calculated over approximately the last 0.1 s prior to the final heel strike of the approach. Similarly, average horizontal flight speed was calculated over approximately the last 0.1 s prior to the instant the Vertec was contacted.
Segmental Contributions to Lift
The relative momentum approach of Ae and Shibukawa (1980) was used to determine the contributions of the arms, head and trunk, and legs to the change in vertical momentum of the body during the propulsion phase of the jump. The change in vertical momentum will be hereafter referred to as "lift" and will be used to represent the net impulse from the vertical forces over the propulsion phase (Hinrichs, Cavanagh, & Williams, 1987). For both jump styles, the propulsion phase was defined by the interval between the instant of zero vertical velocity of the CM during the ground support phase and the instant of takeoff.
The relative momentum for each arm, each leg, and the head and trunk were com- puted as follows:

Vertical jump Performances 343
where m represents mass, v represents velocity, and the subscripts A and T denote the arms, and the trunk and head, respectively. Equation 2 was used to compute the relative momentum of the right and left arms separately. For each leg during two-foot jumps, and for the support leg during one-foot jumps, relative momentum was calculated from Equa- tion 4, where the subscript SL denotes the support leg. For the free swinging leg during one-foot jumps, relative momentum was calculated from Equation 5, where the subscript FL denotes the free swinging leg. The subscripts SSN and H, preceded by a slash, were used to indicate the velocities relative to the supersternale and hips (midpoint between the right and left hip), respectively (e.g., v,,, describes the relative velocity of the center of mass of the entire arm segment with respect to the supersternale).
In Equations 2-5, the terms with a solid underline were considered to be the mo- mentum generated by the support leg or legs, while those with a dotted underline defined the momentum generated by the trunk. The arms were left only with the momentum asso- ciated with the motion of the arms relative to the supersternale (mAvNSs,). During one-foot jumps, the momentum of the free swinging leg was determined solely by the relative motion of the free swinging leg with respect to hips (m,v,). Rather than using the midpoint of the shoulders (Ae & Shibukawa, 1980) or the chin-neck intersect (Hinrichs et al., 1987), we designated the supersternale as the reference point for defining the relative motion of the arms. This assignment was more consistent with the chosen set of body segment parameters (Hinrichs, 1990). Further details about the methods involved in deter- mining the relative segmental contributions to vertical impulse may be found in Ae and Shibukawa (1980) and Hinrichs et al. (1987).
Radial and Vertical Motions
In order to investigate the mechanisms responsible for loading the support leg muscula- ture, we analyzed the radial and vertical motions of the body throughout the ground sup- port phase. The radial motion methodology was adapted from Dapena and Chung (1988) to accommodate an analysis of one-foot and two-foot jumps.
Analysis of the radial motions of the segmental and whole-body centers of mass depends on the definition of a single hinge point over which the body rotates. For one-foot jumps, the hinge point was defined by the average of the X coordinates of the heel and toe of the support leg when the foot was firmly planted on the ground (Figure 2a). For two- foot jumps, the hinge point was defined by the average of the X coordinates of the heel and toe points of both feet when they were both firmly planted on the ground (Figure 2b). For both jump styles, the Y coordinate of the hinge point was defined as zero throughout the duration of the support phase.
Throughout the duration of the ground support phase of the jump, the radial direc- tion vector was defined by a segment that originated at the hinge and terminated at the whole-body center of mass. Position vectors were defined from the hinge to the centers of mass of each of the various systems of segments. In addition, for one-foot jumps, a posi- tion vector was defined from the hinge to the hip of the support leg. For two-foot jumps, a similar vector was defined from the hinge to the midpoint of the right and left hip. The magnitude of each position vector was projected onto the radial direction vector using the dot product relationship. This projected magnitude was defined as the radial distance for each of the segments or points of interest. Radial velocities were obtained from finite difference differentiation of the radial distance arrays. The radial velocity of the whole- body center of mass may be described by the sum of the radial velocities of the individual systems of segments. Therefore, segmental contributions to the radial velocity of the whole- body center of mass (V,) were determined by the relationship shown in Equation 6:

344 Vint and Hinrichs
Figure 2 - Stick figure representations of typical (a) one-foot and (b) two-foot jump performances (representative subject left-handed). Horizontal spacing between adjacent stick figures bas been exaggerated for clarity. Trajectory of whole-body center of mass has been overlaid on stick figure profiles. Insets illustrate location of hinge point during ground support phase for typical (a) one-foot jump and (b) two-foot jump performances. From "Vertical Jumping Performance: One-Foot Vs. Two-Foot Takeoff Techniques," by P. Vint, 1996, Performance and Conditioning for Volleyball, 3, pp. 4-7. Copyright 1996 by Conditioning Press, P.O. Box 6819, Lincoln, NE 68506.

Vertical Jump Performances 345
where V,, represents the radial velocity of the center of mass of the ith system of seg- ments, m, represents the mass of the i" system of segments, and M represents the whole body mass (Dapena & Chung, 1988). The m,/M term in Equation 6 is included to ensure that the segmental contributions to the whole-body radial velocity are weighted appro- priately.
Statistical Analysis
Matched-pairs t tests were performed on selected means to detect significant differences between one-foot and two-foot jumping performances (SPSS, Chicago, IL). Twenty-five statistical tests were conducted in all. In an effort to compromise between the effects of Type I and Type I1 errors, the overall, experiment-wise confidence level was chosen at 0.10. The Bonferroni procedure (Stevens, 1990) was used to adjust the confidence level for each test from 0.1 to 0.0042 (the latter derived from 1 - [l - 0.1]'R5).
Results
Table 1 summarizes some of the basic performance characteristics of one-foot and two- foot jumps. Both the absolute and relative magnitudes of these deterministic factors were similar to those observed for standing two-foot jumps (Hinrichs & Vint, 1994). Takeoff height was significantly higher in one-foot jumps, while flight height was significantly higher for two-foot jumps. Overall jump and reach height, reach height, and loss height were not different between jump styles (p = .7, .9, and .07, respectively).
Mean horizontal approach velocities were 3.77 + 0.41 m . s-I and 3.41 f 0.55 m . s-I for one-foot and two-foot jumps, respectively. Although one-foot jumps tended to be some- what faster, the difference between jump styles was not statistically significant (p = .06). Compared to two-foot jumps, one-foot jumps carried significantly more hori- zontal velocity into the flight phase (p < .0042). Horizontal flight velocity was 1.88 f 0.1 m . s-' for one-foot jumps and 1.01 f 0.22 m . s-' for two-foot jumps.
Table 1 Mean and STD Values (in meters) for Basic Deterministic Factors of Vertical Jumping During One- and Two-Foot Jumps, Plus Percentages of Overall Jump and Reach Height
One-foot M STD %
TWO- foot M STD %
Overall height Takeoff height Flight height Reach height Loss height
Note. Statistical tests were only performed on the absolute height values. *Two-foot significantly less than one-foot ( p < .0042). ?Two-foot significantly greater than one-foot (p < .0042).
346 Vinf and Hinrichs
Table 2 Mean and STD Values for Segmental Contributions to Lift During Propulsion Phase of Jump (Values Expressed in Absolute Units and as Percentages of Total Whole- Body Lift)
Absolute lift (N . s) Percentage of total lift (%)
One-foot Two-foot One-foot Two-foot M STD M STD M STD M STD
Whole body Head and trunk Reach arm Lowered arm Sum of arms Support leg(s) Free swinging leg Sum of legs
Note. Statistical tests were only performed on the absolute lift values. *Two-foot signifi- cantly less than one-foot (p < .0042). $Two-foot significantly greater than one-foot (p < .0042). $One-foot significantly less than zero (p < .0042).
Segmental Contributions to Li f i
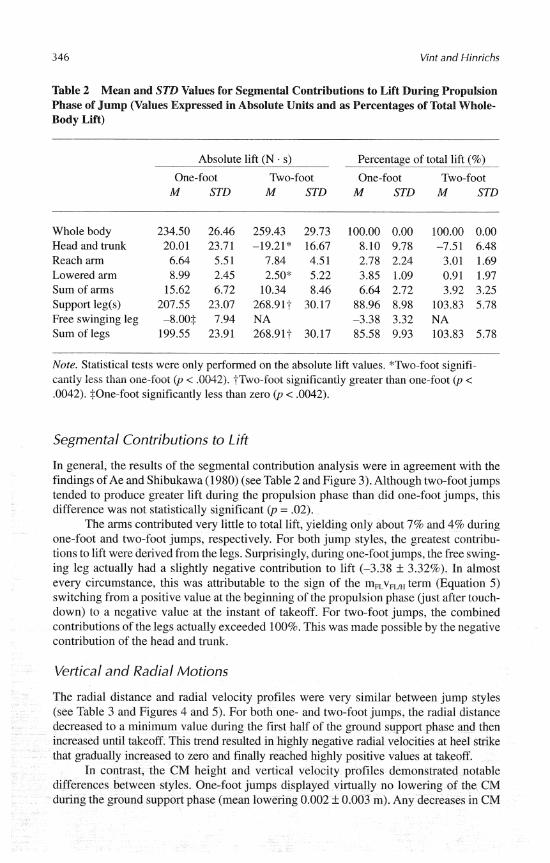
In general, the results of the segmental contribution analysis were in agreement with the findings of Ae and Shibukawa (1980) (see Table 2 and Figure 3). Although two-foot jumps tended to produce greater lift during the propulsion phase than did one-foot jumps, this difference was not statistically significant (p = .02).
The arms contributed very little to total lift, yielding only about 7% and 4% during one-foot and two-foot jumps, respectively. For both jump styles, the greatest contribu- tions to lift were derived from the legs. Surprisingly, during one-foot jumps, the free swing- ing leg actually had a slightly negative contribution to lift (-3.38 If: 3.32%). In almost every circumstance, this was attributable to the sign of the m,v, term (Equation 5) switching from a positive value at the beginning of the propulsion phase (just after touch- down) to a negative value at the instant of takeoff. For two-foot jumps, the combined contributions of the legs actually exceeded 100%. This was made possible by the negative contribution of the head and trunk.
Vertical and Radial Motions
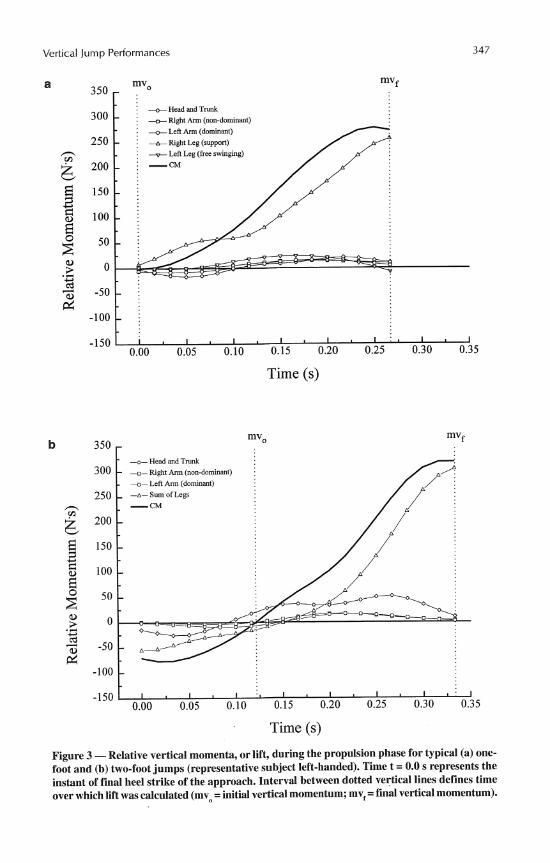
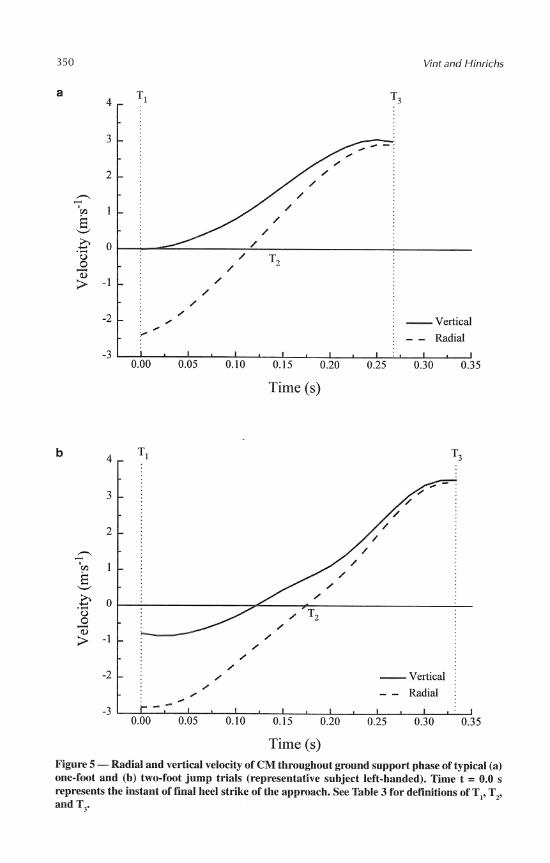
The radial distance and radial velocity profiles were very similar between jump styles (see Table 3 and Figures 4 and 5). For both one- and two-foot jumps, the radial distance decreased to a minimum value during the first half of the ground support phase and then increased until takeoff. This trend resulted in highly negative radial velocities at heel strike that gradually increased to zero and finally reached highly positive values at takeoff.
In contrast, the CM height and vertical velocity profiles demonstrated notable differences between styles. One-foot jumps displayed virtually no lowering of the CM during the ground support phase (mean lowering 0.002 f 0.003 m). Any decreases in CM
Vertical Jump Performances
: -LeftLeg(free swinging)
5
Time (s)
mvo rnVf
-0- Head and Trunk
-0- Left Arm (dominant)
5
Time (s)
Figure 3 -Relative vertical momenta, or lift, during the propulsion phase for typical (a) one- foot and (b) two-foot jumps (representative subject left-handed). T i e t = 0.0 s represents the instant of final heel strike of the approach. Interval between dotted vertical lines defines time over which lift was calculated (mvo = initial vertical momentum; mv, = final vertical momentum).


Vertical Jump Performances
i - Vertical
Time (s)
Time (s)
2.0
1.5
n E w a, 1.0-
8 C, rn .- n
0.5
0.0
Figure 4 -Radial distance and height of CM throughout ground support phase of typical (a) one-foot and (b) two-foot jump trials (representative subject left-handed). Time t = 0.0 s represents the instant of final heel strike of the approach. See Table 3 for definitions of T,, T , and T,.
- , - Vertical i - - Radial i
- i
- . . . - - i - - - -
- i
l . l , l , l I 1 . 1 . 1 1 ' 1
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
Vint and Hinrichs
- .
- .
7
/ Tz /
- . / /
/ #
- : 0 . /
; V e r t i c a l :/ i - - Radial I I . I . I . I , I : , I . ~
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
Time (s)
/ /
/ / V e r t i c a l i
# 0 - - Radial i
Time (s) Figure 5 -Radial and vertical velocity of CM throughout ground support phase of typical (a) one-foot and (b) two-foot jump trials (representative subject left-handed). Time t = 0.0 s represents the instant of final heel strike of the approach. See Table 3 for definitions of T,, T,, and T,.

Vertical Jump Performances 351
height that did occur during one-foot jumps did so within the first 0.014 t 0.018 s after heel strike. Conversely, two-foot jumps elicited a mean lowering of 0.064 k 0.023 m through the first 0.11 1 f 0.023 s. Peak negative vertical velocities for one-foot and two- foot jumps during the ground support phase were -0.05 + 0.17 m . s-' and -0.93 + 0.24 m - s-I, respectively (p < .0042).
At the instant of final heel strike during the approach for two-foot jumps, both the radial and vertical velocities of the CM were negative. One-foot jumps, by contrast, dis- played negative radial velocity but negligible vertical velocity at heel strike. For both jump styles, the CM was already moving upward with appreciable vertical velocity by the time the minimum radial distance was achieved. At this time, the vertical velocity for one- foot jumps was about twice that of two-foot jumps. By takeoff, however, the vertical velocity for two-foot jumps had surpassed that for one-foot jumps.
The relative radial velocity of the CM with respect to the hip of the support leg was expected to indicate the state of activity of the support leg musculature. According to Dapena and Chung (1988),
For a given radial velocity of the CM, the faster that body segments move radially away from the foot, the faster the hip joint will move radially toward the foot, or the slower it will move radially away from the foot. Therefore, positive radial motions of the body segments will put the leg muscles in faster eccentric or slower concen- tric conditions, thus enabling them to exert larger tensions. (p. 293)
Segmental contributions to the radial velocity of the CM were derived from Equation 6 and were expressed relative to the radial velocity of the hip (see Figure 6).
For one-foot and two-foot jumps, the radial velocity of the CM with respect to the hip (or hips) (V,,) was negative throughout approximately the first quarter of the ground support phase and then remained positive until takeoff. Early in the ground support phase for one-foot jumps, the sum of the negative contributions of the arms and free swinging leg was greater than the sum of the positive contributions of the head and trunk and the support leg. Later, when V,, was positive, the positive contributions of the head and trunk, the arms, and the free swinging leg outweighed the modest negative contributions of the support leg.
The pattern of segmental contributions to V,,was very similar for two-foot jumps. The main differences between jump styles were found in the contributions of the legs. Figure 6b shows that the contribution of the left leg was initially positive and then became negative toward the end. This was likely due to the asynchronous step-close foot plant technique (Coutts, 1982) used by most subjects during two-foot jumps. For example, dur- ing most of the two-foot jump trials, left-handed subjects were found to plant the left foot and then the right foot. In a similar manner, right-handed subjects typically employed a right-left foot planting technique.
Discussion
The purpose of this study was to quantify the differences between one-foot and two-foot vertical jump performances. Fourteen male subjects were asked to complete maximal effort one-foot and two-foot vertical jumps using a four-step, self-paced running approach. Two-dimensional video data were used to study jumping performances from the last step of the approach through the instant that the maximum jump and reach height was reached.
The simple deterministic model presented in Figure 1 indicates that the overall jump and reach height depends upon the height and vertical velocity of the CM at takeoff and

Vint and Hinrichs
+-Head and Trunk
-A- Right Leg (support)
-o- Left Leg (free swinging)
Time (s)
+Head and Tmnk 4- Right Ann (non-dominant)
1.25 4- Left Arm (dominant)
l o I -A- Right Leg
Time (s)
Figure 6 - Segmental contributions to the relative radial velocity of the CM with respect to the (a) hip of the support leg throughout the ground support phase of a typical one-foot jump trial and (b) mid-hip throughout the ground support phase of a typical two-foot jump trial (representative subject left-handed). Time t = 0.0 s represents the instant of final heel strike of the approach. Positive values indicate segment is contributing to slower concentric o r faster eccentric conditions of support leg musculature. Negative values indicate segment is contributing to a cushioning effect.

Vertical Jump Performances 353
upon the position and orientation of the body about the CM at the instant the maximum height is evaluated. The results of this study indicate that while overall jump and reach heights were similar between one-foot and two-foot jumps, strategies employed to achieve these results were notably different.
During one-foot jumps, subjects benefited from an increased takeoff height that was largely attributable to the elevation of the free swinging leg. This conclusion is sup- ported by the fact that at takeoff, the height of the CM of the free swinging leg was 0.35 m higher than the CM of the support leg. In addition, it appeared that there was a slightly greater degree of extension in the support leg during one-foot jumps. At takeoff, the aver- age height of the CM of the support leg (or legs) was 0.74 f 0.04 m and 0.71 f 0.03 m for one-foot and two-foot jumps, respectively. During two-foot jumps, subjects were able to achieve greater flight heights, perhaps due to the greater availability of musculature that could be recruited to generate vertical impulse against the ground. This issue will be dis- cussed in greater detail later. Reach heights were virtually identical between jump styles. Loss heights were negligible for both jump styles and were similar in magnitude to those found in standing two-foot jump performances (Hinrichs & Vint, 1994). In addition, al- though one-foot jumps carried significantly faster horizontal velocity throughout the in- flight phase of the jump, subjects did not differ between jump styles in their ability to coordinate the final reach with the apex of the CM.
The relative momentum approach of Ae and Shibukawa (1980) was used to quantify segmental contributions to the change in vertical momentum, or lift, during the propulsion phase of jumping. Although whole-body lift was not statistically different between jump styles, there was a strong tendency for more lift to be produced during two- foot jumps ( p = .02). While this tendency was consistent with the greater flight heights observed for two-foot jumps, the lack of significance may be explained as a Type I1 error arising from the stringent experiment-wise confidence level (a = .0042).
In general, the legs were the greatest contributors to lift for both jump styles. The summation of the lift derived from both legs was about 86% and 104% for one-foot and two-foot jumps, respectively. For one-foot jumps, the free-swinging leg was found to have a negative contribution to lift. While this finding may seem somewhat surprising, it should be recalled that the lift contributed by the free swinging leg is solely determined by the m,v, term in Equation 5. At the final heel strike of the approach, the relative verti- cal velocity of the free swinging leg with respect to the hip of the same leg was approxi- mately zero. Throughout the majority of the ground support phase, this leg was swinging upward with a velocity greater than that of the hip. However, immediately before takeoff, the free swinging leg decelerated to a speed that was slower than the hip of the same leg (see Figure 3a).
The fact that the free swinging leg had a negative contribution to lift during one- foot jumps does not necessarily mean that it had a deleterious effect on overall perfor- mance. During the time when the free swinging leg was accelerating upward, it would have exerted a downward force on the torso. This downward force would have slowed the rate of extension of the support leg, thereby maintaining these muscles in a more favor- able portion of the force-velocity curve and facilitating the development of greater mus- cular tension (Fenn & Marsh, 1935; Perrine & Edgerton, 1978). Near takeoff, the free swinging leg decelerated and exerted an upward force on the torso. Although this upward force would have tended to reduce the force-developing capacities of the support leg mus- culature, it would have done so at a time when the support leg was near full extension and was in a poor position to develop further propulsive forces. (It is also possible that this upward force was responsible for the slightly greater support leg extension and, hence, the

354 Vint and Hinrichs
increased takeoff height observed during one-foot jumps.) This mechanism has been used to describe the contribution of the arm swing during standing two-foot jumps (Harman et al., 1990) and may also be used to account for the relatively small contribution of the arms to overall lift in the present study. We therefore suggest that care be taken when interpret- ing results obtained from the Ae and Shibukawa (1980) relative momentum methodology. While this approach is both intuitively and computationally appealing, investigators must also consider the dynamic effects of the limbs throughout the activity of interest.
Previously, the radial motion methodology had been used exclusively to study the mechanics of the high-jump approach (Dapena & Chung, 1988). In the present investiga- tion, this methodology was adapted to study the function of the approach during one- and two-foot jumps. Although we provided a somewhat different definition for the hinge point, the patterns of the radial motions of segmental and whole-body centers of mass were very similar to those presented by Dapena and Chung (1988). While one-foot jumps displayed virtually no lowering of the CM during the ground support phase, two-foot jumps demon- strated a pattern of CM lowering that was consistent with traditional countermovement actions. However, both jump styles displayed highly negative radial velocities at heel strike that gradually increased to zero and finally reached highly positive values at takeoff. It has been suggested that radial velocity profiles are consistent with the well-known stretch- shortening cycle for previously stretched muscle (Dapena & Chung, 1988). If this is true, it appears that both one-foot and two-foot jumps may benefit from this mechanism.
While the radial motions of the CM can provide some information about the gen- eral conditions of the support leg musculature, the radial velocity of the hip of the support leg can provide a better source of this information (Dapena & Chung, 1988). The differ- ence between radial velocity of the CM and radial velocity of the hip is attributable to the relative motion of the body segments about the hip. The relative radial velocity of the CM with respect to the hip (V,,,) can then be used to understand how various body segments contribute to development of tension in the support leg musculature. Computationally, this is accomplished by expressing both sides of Equation 6 with respect to the radial velocity of the hip. For both one-foot and two-foot jumps, the initial negative values for V,,, indicated that the radial motions of the body segments had a net cushioning effect early in the ground support phase (see Figure 6). For one-foot jumps, this was caused by negative contributions of the arms and free swinging leg that essentially overrode the positive contributions of the head and trunk and the support leg. For two-foot jumps, this cushioning effect was caused by the radial motions of the arms and the first foot to plant during the ground support phase. It is conceivable that this was one of the reasons why most of our subjects adopted an asynchronous foot-planting technique in preference to a simultaneous landing technique. Coutts (1982) suggested that while the asynchronous (or "step-close") landing technique does not offer any distinct advantage over the simulta- neous (or "hop") landing technique in the development of vertical velocity at takeoff, it may help to reduce peak impact forces and attenuate the impact impulse by increasing ground contact time. This, in turn, may help to reduce the predisposition for injury. Future studies should investigate the advantages and disadvantages of these landing strategies as they relate to vertical jump performances.
For both jump styles, when V,,, became and then remained positive, radial mo- tions of the body segments may have helped place the muscles of the support leg (or legs) in either faster eccentric or slower concentric conditions (Dapena & Chung, 1988). This again would have allowed the leg extensor muscles to develop greater forces and therefore increase the vertical impulses against the ground. This finding is important in that it helps to explain the results of previous research demonstrating that vertical jump

Vertical Jump Performances 355
performance increases when the jump is preceded by a running approach (e.g., Enoka, 1971).
With essentially twice the available leg musculature, two-foot jumps were expected to demonstrate a clear superiority in the development of vertical velocity at takeoff. How- ever, while the difference between one-foot and two-foot flight height was statistically significant, the magnitude of this difference was only 9 cm or about 17%. Why was this difference so small? After all, Soest et al. (1985) demonstrated a difference of nearly 40% in jump height between one-foot and two-foot standing jumps. Although it is impossible to completely ascertain the source of the unexpectedly small differences in flight height, a number of explanations for this finding may be plausible.
One of the most obvious explanations for this difference is that the support leg musculature was probably more active during one-foot jumps. Since the weight of the body was unchanged between jump conditions, one-foot jumps would have required one leg to support the entire body weight. Two-foot jumps, conversely, would have allowed the weight to be distributed across two legs. This would have resulted in a greater muscu- lar tension, per support leg, during one-foot jumps compared to two-foot jumps. It is likely that this increase in tension would have been due to relatively greater neural activa- tion of the support leg musculature during one-foot jumps.
Soest et al. (1985) suggested that the differences between one-foot and two-foot vertical jumps were consistent with the literature related to the bilateral deficit phenom- enon. A complete discussion of the manifestations of the bilateral deficit is beyond the scope of the present investigation (for excellent reviews, see Ohtsuki, 1994, and Archontides & Fazey, 1993); however, we will note that a number of investigators have reported that the strength developed during simultaneous bilateral exercise is significantly lower than the sum of that developed during unilateral efforts (Howard & Enoka, 1991; Schantz et al., 1989; Vandervoort et al., 1984). In addition, there is evidence suggesting that the bilateral deficit increases with increasing movement speed (Vandervoort et al., 1984; Vandervoort, Sale, & Moroz, 1987). It has been suggested that the bilateral deficit may be attributable to a decreased neural drive that results in an inability to fully recruit large, high-threshold motor units during maximal voluntary contractions (Koh et al., 1993; Vandervoort et al., 1984,1987). Acloser inspection of the Soest et al. (1985) data revealed that the angular velocities of the hip and knee were notably greater during two-foot jump- ing performances. Therefore, in addition to eliciting the force reduction that would be expected from the well-known force-velocity properties of muscle, faster leg extensions during two-foot jumps may have also suffered from an increased inhibition in the expres- sion of maximum force due to the nature of the bilateral deficit phenomenon. These speed- dependent issues may have played a significant role in the findings of Soest et al. (1985) and should be considered when comparing independent unilateral and simultaneous bilat- eral performances.
In the present study, it was suggested that radial motion parameters would better indicate the state of the support leg musculature. At takeoff, the radial velocity of the CM was significantly faster for two-foot jumps (see Table 3). Therefore, it is conceivable that two-foot jumps were subject to decreases in musculature tension that would have been brought about by the force-velocity relationship and the movement speed dependence of the bilateral deficit. However, during the time when the CM was moving in the positive radial direction (between times T, and T,), the average radial velocities of the CM were 1.86 m . s-' and 1.85 m - s-' for one-foot and two-foot jumps, respectively. Since the aver- age radial velocities of the CM were virtually identical, it seems unlikely that these speed- related factors played a significant role in limiting two-foot jump performances.

356 Vint and Hinrichs
Another possible explanation for the relatively small difference in flight height is that one-foot jumps were simply more effective in developing propulsive impulses against the ground. One-foot jumps displayed virtually no lowering of the CM during the ground support phase. In fact, the magnitude of the peak negative velocity was only -0.05 f 0.17 m . s-' and this was overcome in the first 0.014 + 0.01 8 s after heel strike. Therefore, since virtually no effort was required to stop the downward motion of the CM, more energy could be expended to propel the body upward. Any beneficial effects that may have been derived from the radial motions of the body would have helped to provide higher impulses against the ground while the body was already moving upward. Two-foot jumps, while probably deriving eccentric loading effects early in the ground support phase from the radial and vertical motions of the body, still had to overcome the downward momentum of the body before efforts could be made to directly increase the vertical velocity in the desired direction. This fact alone may have put the two-foot jumps in a less than ideal situation for propelling the body upward.
Horizontal approach speeds were 3.77 m . s ' and 3.41 m . s-'for one-foot and two- foot jumps, respectively. While these values were not statistically different @ = .06), there was clearly a trend for one-foot jumps to employ faster approaches. This would imply that the kinetic energy carried into the ground support phase would have also tended to be greater for one-foot jumps. If a portion of this increased energy could be stored and subse- quently reutilized during the takeoff phase, this would afford one-foot jumps an advan- tage over two-foot jumps. In this study, subjects were allowed to use a four-step, self- paced approach. It is conceivable that had subjects been permitted to use an unconstrained run-up, there would have been a greater difference in the approach speed and hence in the initial kinetic energy between jump styles.
Although the overall jump and reach heights were virtually identical between jump styles, the manners in which these results were achieved were notably different. One- foot jumps benefited from an increased takeoff height that was largely attributable to the elevation of the free swinging leg. Further, it was suggested that the actions of this limb may have helped slow the rate of extension of the support leg during the propul- sion phase. This would have resulted in the development of higher muscular tensions and therefore greater flight heights than would have otherwise been expected. Two-foot jumps were expected to produce greater vertical velocities at takeoff and, therefore, greater elevations of the whole-body center of mass (CM) during the flight phase. Al- though greater flight heights were achieved during two-foot jumps, the magnitude of this difference was only about 9 cm. Factors associated with the development of muscular tension, vertical velocity at touchdown, and horizontal approach speed may have all con- tributed to the unexpectedly small differences in flight height between one-foot and two- foot jumping performances.
References
Ae, M., & Shibukawa, K. (1980). A biomechanical method for the analysis of the body segments in human movement. Japanese Journal of Physical Education, 25,233-243.
Anderson, F.C., & Pandy, M.G. (1993). Storage and utilization of elastic strain energy during jump- ing. Journal ofBiomechanics, 26, 1413-1428.
Archontides, C., & Fazey, J. (1993). Inter-limb interactions and constraints in the expression of maximum force: A review, some implications and suggested underlying mechanisms. Jour- ltQl of Sports Sciences, 11, 145-158.
Asmussen, E., & Bonde-Petersen, F. (1974). Apparent efficiency and storage of elastic energy in human muscles during exercise. Acta Physiologica Scandinavica, 91, 385-392.

Vertical Jump Performances 357
Bobbert, M., Mace, M., Schinkelshoek, D., Huijing, P.A., & Ingen Schenau, G.J., van. (1986). Bio- mechanical analysis of drop and countermovement jumps. European Journal ofApplied Physi- ology, 54,566-573.
Cavagna, G.A., Dusman, B., & Margaria, R. (1968). Positive work done by previously stretched muscle. Journal ofApplied Physiology, 24,21-32.
Coutts, K.D. (1982). Kinetic differences of two volleyball jumping techniques. Medicine and Sci- ence in Sports and Exercise, 14,57-59.
Clauser, C.E., McConville, J.T., & Young, J.W. (1969). Weight, volume, and center of mass of the segments of the human body (AMRL Technical Report #69-70, AD-710-622). Dayton, OH: Wright-Patterson Air Force Base.
Dapena, J. (1980a). Mechanics of translation in the Fosbury-flop. Medicine and Science in Sports and Exercise, 12,37-44.
Dapena, J. (1980b). Mechanics of rotation in the Fosbuly-flop. Medicine and Science in Sports and Exercise, 12,45-53.
Dapena, J., & Chung, C.S. (1988). Vertical and radial motions of the body during the take-off phase of high jumping. Medicine and Science in Sports and Exercise, 20,290-302.
Dapena, J., McDonald, C., & Cappaert, J. (1990). Aregression analysis of high jumping technique. International Journal of Sport Biomechanics, 6,246-261.
Enoka, R.M. (1971). The effect of different lengths of run-up on the height to which a spiker in volleyball can reach. New Zealand Journal of Health, Physical Education, and Recreation, 4, 5-15.
Fenn, W.O., & Marsh, B.S. (1935). Muscular force at different speeds of shortening. Journal of Physiology, 85,277-297.
Fox, E.L., Bowers, R.W., & Foss, M.L. (1988). The physiological basis of physical education and athletics. Dubuque, IA: Brown.
Harman, E.A., Rosenstein, M.T., Frykman, P.N., & Rosenstein, R.M. (1990). The effects of arms and countermovement on vertical jumping. Medicine and Science in Sports and Exercise, 22, 825-833.
Hay, J.G., & Reid, J.G. (1988). Anatomy, mechanics, and human motion. Englewood Cliffs, NJ: Prentice Hall.
Healy, J. (1977). Effects of various approaches on the verticaljump in volleyball. Unpublished master's thesis, Western Illinois University, Macomb.
Hinrichs, R.N. (1990). Adjustments to the segment center of mass proportions of Clauser et al. (1969). Journal of Biomechanics, 23,949-951.
Hinrichs, R.N., Cavanagh, P.R., &Williams, K.R. (1987). Upper extremity function in running: I. Center of mass and propulsion consideration. International Journal of Sport Biomechanics, 3, 222-241.
Hinrichs, R.N., & Vint, P.F. (1994). A comparison of Sargent jump height and actualflight height in vertical jumping. Paper presented at the American Society of Biomechanics Annual Meeting, Columbus, OH.
Howard, J.D., & Enoka, R.M. (1991). Maximum bilateral contractions are modified by neurally mediated interlimb effects. Journal ofApplied Physiology, 70, 306-316.
Ingen Schenau, G.J. van. (1984). An alternative view of the concept of utilisation of elastic strain energy in human movement. Human Movement Science, 3,301-336.
Kayambashi, K. (1977). Effects of approaches and takeoffs on the vertical jump in volleyball. Un- published master's thesis, Western Illinois University, Macomb.
Koh, T.J., Grabiner, M.D., & Clough, C.A. (1993). Bilateral deficit is larger for step than for ramp isometric contractions. Journal of Applied Physiology, 74, 1200-1205.
Komi, P.V. (1973). Measurement of the force-velocity relationship in human muscle under concen- tric and eccentric contractions. In S. Cerquiglini, A. Venerando, & J. Wartenweiler (Eds.), Biomechanics III (pp. 224-229). Baltimore, MD: University Park Press.
Komi, P.V., & Bosco, C. (1978). Utilization of stored elastic energy in leg extensors by men and women. Medicine and Science in Sports and Exercise, 10,261-265.
Miller, D.L, & Nelson, R.C. (1973). Biomechanics of sport. PhiIadelphia: Lea & Febiger.

358 Vint and Hinrichs
Ohtsuki, T. (1994). Changes in strength, speed, and reaction time induced by simultaneous bilateral muscular activity. In S.P. Swinnen, H. Heuer, J. Massion, & P. Casaer (Eds.), Interlimb coor- dination: Neural, dynamical, and cognitive constraints @p. 259-274). San Diego, CA: Aca- demic Press.
Pandy, M.G., & Zajac, F.E. (1991). Optimal muscular coordination strategies for jumping. Journal of Biomechanics, 24(1), 1-10.
Penine, J.J., & Edgerton, V.R. (1978). Muscle force-velocity and power-velocity relationships under isokinetic loading. Medicine and Science in Sports and Exercise, 10, 159- 166.
Sargent, D.A. (1921). The physical test of a man. American Physical Education Review, 26, 188- 194.
Saunders, H.L. (1980). A cinematographical study of the relationship between speed of movement and available force. Unpublished doctoral dissertation, Texas A&M University, College Sta- tion.
Schantz, P.G., Moritani, T., Karlson, E., Johansson, E., & Lundh, A. (1989). Maximal voluntary force of bilateral and unilateral leg extension. Acta Physiologica Scandinavica, 136, 185- 192.
Smith, G. (1989). Padding point extrapolation techniques for the Buttenvorth digital filter. Journal of Biomechanics, 22,967-97 1.
Soest,A.J. van, Roebroek, M.E., Bobbert, M.F., Huijing, P.A., & Schenau, G.J. van. (1985).Acom- parison of one-legged and two-legged countermovement jumps. Medicine and Science in Sports and Exercise, 17, 635-639.
Stevens, J. (1990). Applied multivariate statistics for the social sciences (2nd ed.). Hillsdale, NJ: Lawrence Erlhaum.
Vandervoort, A.A., Sale, D.G., & Moroz, J. (1984). Comparison of motor unit activity during unilat- eral and bilateral leg extension. Journal of Applied Physiology, 56,46-51.
Vandervoort, A.A., Sale, D.G., & Moroz, J. (1987). Strength-velocity relationship and fatigability of unilateral versus bilateral arm extension. European Journal of Applied Physiology, 56,201- 205.
Wells, R.P., & Winter, D.A. (1980). Assessment of signal and noise in the kinematics of normal, pathological, and sporting gaits. Human locomotion I (pp. 92-93). London, ON: Canadian Society for Biomechanics.
Winter, D.A. (1990). Biomechanics and motor control of human movement (2nd ed.). New York: Wiley.
Winter, D.A., Sidwell, H.G., & Hobson, D.A. (1974). Measurement and reduction of noise in kine- matics of locomotion. Journal of Biomechanics, 7, 157-159.
Zajac, F.E. (1993). Muscle coordination of movement: A perspective. Journal of Biomechanics, 26(S l), 109-124.
Acknowledgments
We would like to express our sincere thanks to John DeWitt, Tony Marsh, Scott McLean, Rachel Seidler, Chris Shenvood, Dr. Phil Martin, and Steve D'Amico for their assistance throughout various stages of this project. We also wish to thank Dr. James G. Hay from the University of Iowa and Dr. Jesus Dapena from Indiana University for their valuable comments throughout the develop- ment of this manuscript.
Peter Vint is now with the Department of Exercise and Sport Science, University of North Carolina at Greensboro, 250 HHP Building, Greensboro, NC 27412-5001. Richard Hinrichs is with the Exercise and Sport Research Institute, Arizona State University, Tempe, AZ 85287-0404.





























