Embed Size (px)
Citation preview

The change in muscle fiber size and fiber percentage was studied in three heads of the dog quadriceps following 10 weeks of immobilization using an external skeletal fixator. Muscle biopsy morphometric analysis revealed that muscle fiber atrophy was greatest for the vastus medialis and least for the rectus femoris. The atrophic response for type 1 fibers was, in order from most to least atrophied: VM > VL > RF, whereas for type 2 fibers the corresponding order was VM = VL > RF. An increase in connective tissue was also observed for all muscles. These results are discussed in terms of skeletal muscle architecture, initial fiber-type distribution, and level of use. As a result, predictions are made as to the muscles that are most vulnera- ble to disuse atrophy, namely the postural muscles that contain a relatively large proportion of slow muscle fibers and cross a single joint. Conversely, those that are least susceptable to atrophy are those that are not used as postural muscles, that cross multiple joints, and that are predominantly composed of fast muscle fibers.
MUSCLE & NERVE 11:193-201 1988
DIFFERENTIAL RESPONSE OF THE DOG QUADRICEPS MUSCLE TO EXTERNAL SKELETAL FIXATION OF THE KNEE
RICHARD L. LIEBER, PhD, JAN 0. FRIDEN, MD, PhD, ALAN R. HARGENS, PhD, LARRY A. DANZIG, MD, and DAVID H. GERSHUNI, MD, FRCS
Skeletal muscles atrophy when the level of muscle use is decreased as a result of myopathy,31 suspension h o k i n e ~ i a , ~ ~ ~ ' ~ ? ' ~ spinal cord t ran~ect ion",~~, y'30 and limb immobiliza-
In order to determine the nature of the atrophic response, a variety of muscle groups have been studied. Investigators have demonstrated that muscles composed mainly of type 1 fibers (i.e., "slow" muscles) atrophy to a greater extent than muscles composed mainly of type 2 fibers (i.e., "fast" muscle^).^^^*'"^'^^'^."" Ad-
tion.',4,~,17,18,23,~5,32,3(i,37,42
From the Division of Orthopaedics and Rehabilitation, Deparlment of Surgery, Veterans Administration Medical Center and University of Cali- fornia, San Diego, CA.
Address reprint requests to Dr. Lieber at the Division of Orthopaedics and Rehabilitation, Department of Surgery, V-151, Veterans Adminislra- tion Medical Center, San Diego. CA 92161
Dr. Friden's address is Departmpt of Hand and Reconstructive Sur- gery Umea University. S-901 87 Umea, Sweden.
Acknowledgments: This work was supported by the Veterans Adminis- tration (DHG, ARH. RLL) and USPHS/NIH grants AM25501 and AM26344 (ARH) and AR35192 (RLL) The authors thank Tim Kim and Diana LeBow for their excellent technical assistance and Dr Bjorn Ryde- vik for his critical reading of the manuscript.
Received for publication September 26, 1986 Revised manuscript ac- cepted for publication February 12, 1987.
0 1988 John Wiley & Sons, Inc 01 48-639Xil103/0193 $04.0019
ditionally, antigravity muscles atroph to a reater
Experimentally, slow muscles (e.g., soleus) are typically compared with fast muscles (e.g., gastroc- nemius, tibialis anterior, plantaris, and extensor digitorum longus) following a period of limb im- mobilization. However, several difficulties arise in making comparisons between these two muscle types. First, the two muscles are immobilized at different lengths, which dramatically affects the atrophic response. muscle t pes normally have different activity levels. 1191'21671g Thus, the change in level of use following immobilization differs. Third, the mus- cle fiber lengths, fiber length/muscle length ratios, arid number of joints crossed differ betwen these diffi-ent muscles, 14,29240 precluding definitive de- termination of the cause of atrophy. Therefore, it is not clear whether the differences observed be- tween fast and slow muscles are due to inherent muscular differences or to other factors such as level of use, immobilization length, or architec- ture.
In order to determine the relative importance of these factors, we have studied the atrophic re- sponse of three heads of the dog quadriceps, the rectus femoris (RF), vastus lateralis (VL), and
extent than their antagonists. 4,9,23,26,&,42
5,14,33,36,37 Second, the two
MUSCLE & NERVE March 1988 193

vastus medialis (VM), when the knee is irnmobi- lized by an external skeletal fixator. 'These three muscles contain nearly identical architectures and fiber but differ in fiber type percent- age and number of joints crossed.3 The RF acts both as a knee extensor and hip flexor and is coni- posed of about 50% type 1 fibers. The VM and VL both function as knee extensors only, but the VM contains about 50% typc 1 fibers, whereas the VL contains only about 20% type 1 fibers."%34 This model thus allows comparison between the VM and VL, which can be immobilized at precisely the same length but contain different percentages of type 1 and type 2 fibers. Similarly, comparisons between the RF and VM can be made, which have similar fiber type percentages but cross different joints. Because dog muscles contain no type 2B fiber^,"'^ unequivocal identification of fiber types can be made from a single histochemical stain for myofibrillar ATPase activity. Finally, the external fixation procedure in this study permits more re- producible setting of joint angle between animals than does cast immobilization and produces rigid fixation.
The purpose of this study, therefore, was to compare the morphometric properties of three heads of the dog quadriceps after immobilization by external skeletal fixation in order to determine the relative importance of function and fiber-type distribution in the atrophic response. Portions of this material have been presented previously. l Y
MATERIALS AND METHODS
The right knees from seven mature dogs (mass range 20-25 kg) were immobilized at 90" of flex- ion for 10 weeks (Fig. 1). The external fixator used was composed of two transfixing threaded Steinman pins placed in the distal femur and two placed in the proximal tibia. The pins were con- nected to side bars such that the knee was neither compressed nor distracted. The left leg served as a nonimmobilized control. All procedures were performed in accordance with the National Re- search Council's guide for the care and use of lab- oratory animals. At 10 weeks following fixation, open biopsies (approximate size 1.5 cm") were taken with the overlying fascia from a well-de- fined superficial portion of the vastus medialis (VM), vastus lateralis (VL), and rector femoris (RF), 10 cm proximal to the knee and distant from the Steinman pin tracts. On both legs, ar- throtomies were performed to mimic the common clinical occurrence prior to postoperative immobi-
-- ---___ '. , .
FIGURE 1. Method of immobilization of right knee at 90" (A) with external skeletal fixator (6) in position C.
lization. Several factors provided support for us- ing the arthrotomized leg as a control. First, it was found that 110 significant difference in any mea- sured parameter (see below) occurred between nonoperated control legs and legs that had only received arthrotomy. Second, in a separate study of remobilization following immobilization, con- trol legs in remobilized animals were not signifi- cantly different from either the arthrotornized legs or the legs from the immobilization study. Fi- nally, fiber-type distributions from all control legs were similar to those found by Armstrong et al." and Snow et a1.34
The muscle biopsy was stretched to approxi- mate rest length (fascia used as reference), frozen in isopentane cooled by liquid nitrogen (- 159"C), and stored at -80°C for subsequent quantitative histological and enzyme histochemical analyses. Muscle cross sections (8p.m) were stained for rou- tine histological examination with Hernatoxylin & Eosin for oxidative enzyme activity with succinic
194 Differential Atrophy of Dog Quadriceps MUSCLE & NERVE March 1988

dehydrogena~e’~ and for niyofibrillar A‘T’Pase ac- tivity to distinguish between type 1 and type 2 fibers.6 Fibers were classified as either type 1 or type 2 depending on their optical density follow- ing myofibrillar ATPase staining. Intermediate staining fibers (approximately 2% in control mus- cles and 4% in experimental muscles) were classi- fied as type 2. However, subsequent data analysis with these fibers classified either as type 1 or omit- ted did not alter any statistical conclusions.
Muscle morphometric parameters were deter- mined according to the point countin stereologi- cal methods developed by Weibel.” Six fields were randomly scored for a given muscle section using an eyepiece grid inserted into the micro- scope ocular. An average of 195 fibers was in- cluded for analysis of control muscles, whereas an average of 351 fibers was included in the analysis of experimental muscles. The following parame- ters were quantified:
1. Type 1 fiber area (Fm2) 2. Type 2 fiber area &m2) 3. Percentage of each fiber type 4. Area fraction of endomysial and perimysial
connective tissue (%)
In an independent control series, it was dem- onstrated that interobserver variation in morpho- metric parameters (three observers) was about 296, and intersample variation within a muscle within an animal (4 samples in 4 animals) was about 10% if care was taken to obtain biopsies from the same superficial area of the muscle (Fri- dkn and Lieber, unpublished data).
Statistical analysis of the morphometric param- eters was accomplished using the BMDP statisical package.8 A two-way analysis of variance (ANOVA, Program P7D) was used with side (im- mobilized or control) and muscle (VM, VL, or RF) as the grouping variables. In this way, potential interactions between side and muscle could be in- vestigated. Subsequent paired comparisons were made a posteriori using the Bonferonni approximation.g5 Following the ANOVA, in or- der to determine the parameters that best dis- criminated between control and immobilized VL, VM, and RF, discriminant analysis (BMDP pro- gram P7M) was performed with F-to-enter = 3.000 and F-to-remove = 2.996. An additional discriminant analysis was performed to discrimi- nate only between the immobilized and control- sides, independent of muscle. All statistical results were considered significant for P < 0.05.
RESULTS
In general, specimens from the immobilized side displayed muscle fibers with a variable distribu- tion of size, shape, and staining intensities relative to the nonimmobilized side (Figs. 2-4). Neither necrotic nor inflammatory cells were observed. The oxidative activity of the immobiliLed muscles, as evidenced by histochemical staining intensity, decreased. However, type 2 fibers retained their normal oxidative staining pattern, with the high- est mitochondria1 staining density on the fiber pe- riphery. Qualitatively, the VM appeared to atro- phy to the greatest extent (Fig. 2), whereas the RF atrophied the least (Fig. 3 ) . The VL atrophic re- sponse was intermediate between the VM and RF (Fig. 4).
No difference in type 1 or type 2 fiber areas was observed between any of the muscles on the control side (Table 1, Figs. 5 and 6). Fiber-type percentages of the control muscles obtained agreed with those reported by Armstrong et for the superficial portion of all muscles. The 10 weeks of immobilization significantly decreased both type 1 (Fig. 5, P < 0.01) and type 2 (Fig. 6, P < 0.001) muscle fiber area. Although no signif- icant difference was observed between the type 1 areas of the three immobilized muscles (Fig. 5, P > 0.4), a significant difference was observed be- tween the type 2 areas of the immobilized muscles ( P < 0.05). Specifically, the type 2 fiber area of the immobilized VM and VL were significantly less than the type 2 area of the immobilized RF (Fig. 6, P < 0.05).
Immobilization caused proliferation of en- domysial and perimysial connective tissue relative to the control leg (P < 0.03), with a significantly greater increase in the immobilized VM and VL than the immobilized RF (Table 1). In addition, a significant increase in fast fiber percentage fol- lowing immobilization was observed in the VM (Table 1). The atrophic response for type 1 fibers was thus, in order from most to least atrophied: VM > VL > RF; whereas for type 2 fibers the corresponding order was VM = VL > RF.
Discriminant analysis revealed that the param- eter that best discriminated between the six groups was type 2 fiber area. Following type 2 fi- ber area, the next-best parameter was the percent- age of type 1 fibers. Using the discriminant func- tion to distinguish retrospectively between the various muscles under study produced mixed re- sults, depending on the muscle. The aggregate correct classification percentage was 56%. The in- dividual correct classification percentages were:
Differential Atrophy of Dog Quadriceps MUSCLE & NERVE March 1988 195

FIGURE 2. Photomicrograph of dog vastus medialis muscle stained for myofibrillar ATPase activity following preincubation at pH 10.0. Type 2 fibers stain black. (A) Control muscle, (B) immobilized muscle. Bar = 60 km.
control VM (17%), control VL (86%), control RF (SO%), immobilized VM (75%, irnmobilized VL (75%), immobilized RF (33%). Therefore, the ability to classify muscles based on type 2 fiber area and type 1 fiber percentage was fairly suc- cessful (>SO%) except for the control VM and im- mobilized RF. Discriminant analysis was much more successful in distinguishing between the im- mobilized and control sides, independent of nius- cle. The aggregate correct classification percent- age was 81%. The most difficult muscle to fit into a classification scheme was the RF. All of the in- correctly classified immobilized muscles (31 14) were from the RF. As a result of the relatively small degree of type 2 fiber atrophy, these were classified as control muscles.
DISCUSSION
Skeletal muscle that is immobilized in a shortened position atrophies, its force generating capacity is decreased, and it demonstrates reduced oxidative activity. The response of different muscle groups to decreased use is a function of their normal use arid their fiber-type distribution.
Increased epimysial and perimysial connective tissue has been observed by others',37241 and re- sults in increased muscle stiffness.37341 Clinically, this results in decreased joint range of motion.14 Appel et al.' reported that the initial response to iminobilization is increased perimysial tissue, and only under prolonged immobilization conditions (such as those reported here) is the proliferation in epirnysial connective tissue observed.
196 Differential Atrophy of Dog Quadriceps MUSCLE & NERVE March 1988

FIGURE 3. Photomicrograph of dog rectus femoris muscle stained for myofibrillar ATPase activity following preincubation at pH 10.0. Type 2 fibers stain black. (A) Control muscle, (B) immobilized muscle. Bar = 60 Fm.
'l'he differences observed between the VM and \'I. cannot be explained by clif'fkrences in immobi- lization length. Si~nilarly, differences observed be- tween the VM antl RF cannot be explaiiiecl h y dif- ferences in fiber tyrlc distribution. In order to explain the differential atrophy of the V M , \'I,, and KF, several factors must be coiisicleretl. First. all three muscles are innervated h y the fenioi-a1 nerve, so that trophic influences probably do i i o t
explain the observed dif'fkreiices. 'l'he ar-chitec- ture of all three muscles is very siniilar. ' l l i e ratio of fiber length/niuscle length has been studied in h~1mans"" and guinea pip" and approxiniates 0.2 for all three muscles; tlius, archi(ecture dif'ter- ences cannot account for the diffei-ences ob- served.
T h e VM demonstrated a significantly greater amount of atrophy than the RF (Figs. 2, 3, 5, antl
6 arid Table I ) . This is interesting in light of the f'act that the two muscles initially contained nearly ideritical fibei--type disti-ibutions (Table 1). ' Ihere- fore, initial fiber-type distribution alone does not dictate the inagnitude o f the atrophic response. Hobvever. the KF crosses both the hip and knee. t'unctioniiig as a knee extensor arid hip flexor, \vhei-eas the VM arid VI, cross only the knee, f.unctioniiig ;is knee extensors. 'I'he KF is there- fine less rigidly immol)ilizecl than either of the \,asti, pi-ol)al)ly explaining the differential re- sponse.
~ l ' h e type 1 fibers of the VM atrophied to a greater extent thaii those of ' the VL (Fig. 5). Both the Vh.1 a n t l VI, were immobilized at the same length, since they both arise from the proxinial fe- I I ILII . and insert together with the rectus tendon onto the patella. I t seems unlikely. then, that the
Differential Atrophy of Dog Quadriceps MUSCLE 8, NERVE March 1988 197

FIGURE 4. Photomicrograph of dog vastus lateralis muscle stained for myofibrillar ATPase activity following preincubation at pH 10.0. Type 2 fibers stain black. (A) Control muscle, (6) immobilized muscle. Bar = 60 pm.
small difference in anatomical location could ac- count for the markedly different response. The VM initially contained a much larger proportion of type 1 fibers than the VL. This indicates that the VM is largely used as an antigravity muscle, since muscle fiber type distributions rovide in-
fore, following immobilization, the changp in the amount of VM activation was probably greater than the change for the VL, even though the ab- solute levels following immobilization may have been similar. The large change in VM activation level could account for the slow-to-fast transfor- mation. For example, Haggmark and coworkers15 observed a significant increase in VL, type 2 fiber percentage following knee surgery. The magni- tude of the transformation was correlated with the change in use, since elite athletes demon-
sights into muscle activation history. 1<),8.32 There-
strated the largest degrees of transformation. Two difficulties arise in interpreting the data pre- sented in this study. First, because of the type 1 to type 2 trailsformation observed in the VM, many of the VM type 2 fibers may actually be type 1 f i - bers that havc atrophied and then transformed. Thus, it may not be justifiable to speak of “type 2 atrophy” in the VM, since the fibers may have at- rophied while they were type 1. Second, the fiber sizes on the nonimmobilized leg may be larger than normal if there was additional weight-bear- ing while carrying the immobilized leg. This pos- sible effect was considered small and was not in- vestigated further.
I t was interesting that the technique of discri- minant analysis had the most difficulty in classify- ing the immobilized RE’ (33% correct) and control VM (17% correct). This was expected for the RF,
198 Differential Atrophy of Dog Quadriceps MUSCLE & NERVE March 1988
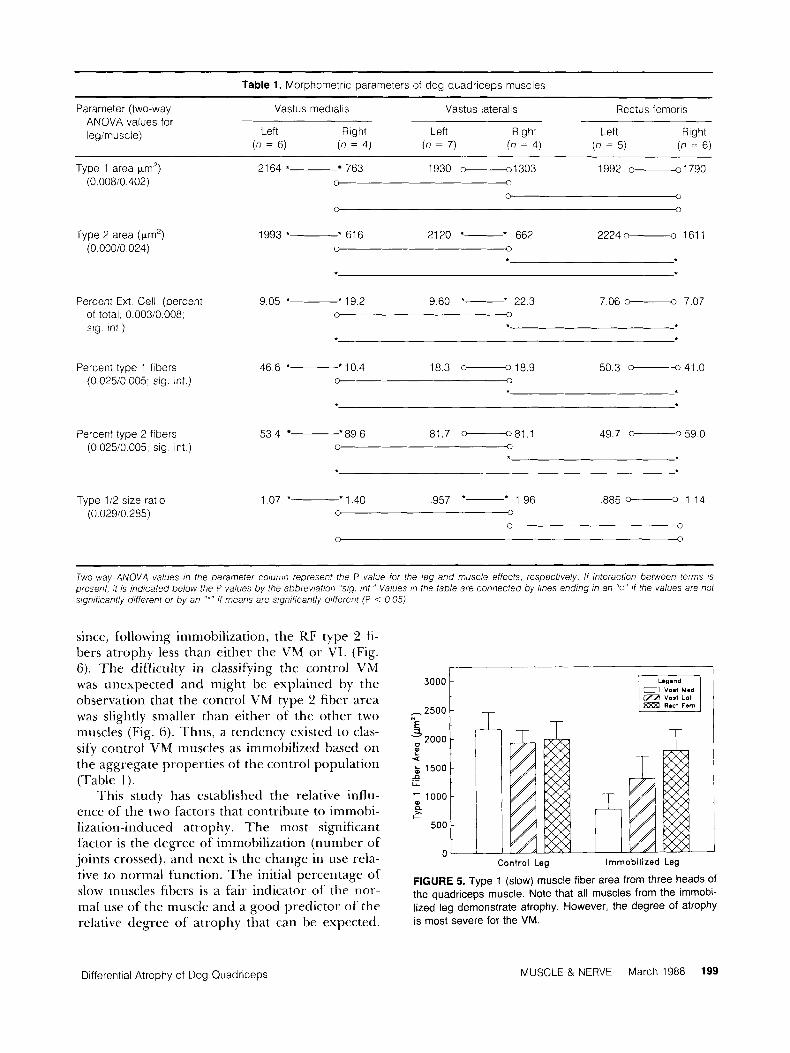
Table 1. Morphornetric parameters of dog quadriceps muscles
Parameter (two-way Vastus rnedialis Vastus lateralis Rectus fernoris ANOVA values for legirnuscle) Left Right Left Right Left Right
(n = 6) (n = 4) (n = 7) (n = 4) (n = 5) (n = 6 )
Type 1 area pm') 2164 * 763 1930 -1303 1992 0--01791790 (0 00810 402) 0 0
0 0
0 3
Type 2 area (prn') 1993 *- * 616 2120 *- * 662 2224 o------o 161 1 (0 00010 024) 0
Percent Ext Cell (percent 9 0 5 *- ' 1 9 2 9 6 0 *- * 2 2 3 7 06 -0 7 07 of total 0 00310 008, 0 0
sig int )
Percent type 1 fibers 4 6 6 + * l o 4 183 -189 5 0 3 -41 0 (0 02510 005 sig. int ) 0 0
Percent type 2 fibers 5 3 4 *89 6 81 7 -81 1 49.7 - 59 0 (0 02510 005, sig int ) 0 -0
Type 112 size ratio (0.02910.285)
1.07 *- * 1.40 ,957 *- * 1.96 ,885 -0 1.14 0
0 0
0 0
~
Two-way ANOVA values ,n the parameter coluiiin represent the P value for the leg and muscle effects, respectively. If interaction between terms is present, i t is indicated below the P values by the abbreviation "sig. mt." Values in the table are connected by lines ending in an "0" i f the values are not significant/y different or by an "*" if means are significantly different (P < 0 05)
since, following immobilization, the RF type 2 fi- bers atrophy less than either the VM or V I A (Fig. 6). The difficulty in classifying the control VM was unexpected and might be explained by the observation that the control V M type 2 fiber area was slightly smaller than either of the other two muscles (Fig. 6). Thus, a tendency existed to clas- sify control VM muscles as immobilized based on the aggregate properties of the control population (Table 1).
This study has established the relative influ- ence of the two factors that contribute to immobi- lization-induced atrophy. The most significant factor is the degree of immobilization (number of' joints crossed), and next is the change in use rela- tive to normal function. The initial percentage of slow muscles fibers is a fair indicator of the nor- mal use of the muscle and a good predictor of the relative degree of atrophy that can be expected.
2500
T
t
n I-
500
Control Leg immobilized Leg
FIGURE 5. Type 1 (slow) muscle fiber area from three heads of the quadriceps muscle. Note that all muscles from the immobi- lized leg demonstrate atrophy. However, the degree of atrophy is most severe for the VM.
Differential Atrophy of Dog Quadriceps MUSCLE & NERVE March 1988 199

N 30001 2500 i 1 ,
e
n 1
500
n Control Leg Immobilized Leg
FIGURE 6. Type 2 (fast) muscle fiber area from three heads of the quadriceps muscle. Note that all muscles from the immobi- lized leg demonstrate significant atrophy. However, the degree of atrophy is most severe for the VM and VL.
These data also indicate that a blanket concept o f "slow fiber atrophy" does not apply to all muscles. Rather, it is a combination of factors that deter- mines the muscular response to decreased use.
Given the structure and fiber type distribu- tions of the various muscles in the human, it is possible to predict those that are most vulnerable to immobilization-induced atrophy, i.e., those that function as antigravity muscles, cross a single joint, and contain a relatively large proportion of
REFERENCES
1. Appell HJ: Morphology of immobilized skeletal muscle and the effects of a pre- and p"stimmobi1izatioii training program. IritJ Sports Med 7:6-12, 1986.
2. Appell HJ: Skeletal muscle atrophy during immobiliza- tion. f n t J Sports Mrd 7:l-5, 1986.
3. Armstrong RB, Saubert CW IV, Seeherman HJ, Taylor CR: Distribution of fiber types in locomotory musclrs of
4. Booth FW: Effect of limb immobilization on skeletal mus- cle.JApplPhysio1 52:1113-1118, 1982.
5. Boyes G, Johnston I: Muscle fibre composition of rat vastus intermedius following immobilisation at different rnusclc lengths. Pfluegm Arch 381: 195-200, 1979.
6. Brooke MH, Kaiser KK: Muscle fiber types: how many and what kind? Arch Neurol 23:369-379, 1970.
7. Corky K, Kowalchuk N , McComas AJ: Contrasting effects of suspension on hind limb rnuscles in the hamster. E x p Neurvl 85:30-40, 1984.
8. Dixon WJ: BMDP Statistical Software. I m Angeles, Uni- versity of California Press, 1983.
9. Edgerton VR, Barriard RJ, Peter JB, Maier A, Sinipson DR: Properties of immobilized hind-limb muscles of the Galugo senegaltnsis. Exp Neurol 46:115-131, 1975.
10. Edstrom L: Selective atrophy of red muscle fibres in the quadriceps in long-standing kneejoint dysfunction inju- ries to the anterior cruciate ligament. J Neurol Sri 11:551-558, 1970.
11. Fischbach GD, Robbins N: Changes in contractile proper- ties of disused soleus muscles. ,] Pliyszol (Lond) 201:305-320, 1969.
12. FouInier M , Roy RR, Perhani H, Simard CP, Edgerton
dogs. A m J Anal 153~87-98, 1982.
slow libers. This description fits the soleus, vastus medialis, and vastus intermedius. The next class of muscles susceptible to immobilization-iri~luced atrophy would be antigravity muscles, predomi- nantly slow that cross multiple joints, namely the longissimus and transversospinalis (erector spi- nae), gastrocnemius, and rectus femoris. These muscle groups should be immobilized conserva- tively to avoid severe strength loss. Conversely, phasically activated, predominantly fast rnuscles (e.g., tibialis anterior, extensor digitoruni longus, and biceps) can be inimobilized with less loss ot' strength. This heirarchy of susceptibility to immo- bilization is supported by the data of Musacchia et al., who studied the atrophic response second- ary to hindlimb suspension. They noted that mus- cles atrophied in the order (most to least atrophy) soleus > gastrocnemius = plantaris > extensor di- gitorum longus. Similarly Edgerton et aL9 mea- sured morphological, biochemical, and physiolog- ical properties of immobilized hindlimb muscles from Galego senegalensis (bushbaby) and found that muscles atrophied in the order (most to least atrophy) soleus > plantaris > vastus iritermedius = vastus lateralis > gastrocnemius > tibialis ante- rior = rectus femoris, agreeing well with the priri- ciples stated above.
26
VR: Is limb iinrnobilization a model of muscle disuse? E x p Neurol 80:147-156, 1983.
19. Fridcn 10, Lieber RL, Gershuiii DII, Kim T K , Hargcns AR: Differeniial response of 1 he dog quadriceps muscle to rigid knee immobilimtion. Trans Orthop Re.! Soc 11: 1x4, 1986.
14. Gossman MR, Rose SJ, Sahrmann SA, Katholl CR: Length and circumference measurements in one-joint and niulti- joint muscles in rabbits after immobilization. P h y Ther 66:516-520, 1986.
15. Haggmark T, Eriksson E, Jansson E: Muscle fiber type changes in human skeletal muscle after injuries and im- mobilization. Orthopedics '3: 181-185, 1986.
16. Hnik P, Vejsada R, Goldspink DF, Kasicki S, Krekule I: Quantitative evaluation of electromyogram activity in rat extensor and flexor muscles immobilized at different lengths. Exp Nettrol 88:515-528, 1985.
17. Jaffe DM, Terry RD, Sprio AJ: Disuse atrophy of skeletal muscle: a morphometric study using image analysis.] Neu- rol Sci 35:189-200, 1978.
18. Joki P, Konstadt S: The effect of limb irnmobilization on muscle function arid protein composition. Cliri O r h p 174:222-228, 1983.
19. Jolesz F, Sreter FA: Development, innervation, and activi- ty-pattern induced changes in skeletal muscle. Aiinzr Re71 I'hysiol 43:531-552, 1981.
20. LeBlanc A , March C, Evans H , Johnson P, Schneider V, Jhingran S: Bone and niuscle atrophy with susprnsion of the rat.] AppZ PIiyiol 68:1669-3675, 1985.
21. I.ieber RL, Fridkri .YO, Hargens AR, E'eringa ER: Long- term effects of spinal cord transection on Cast and slow rat
200 Differential Atrophy of Dog Quadriceps MUSCLE & NERVE March 1988

skeletal muscle. 11. Morphonietric properties. Exp Neurul
22. Lieber KL., Johaiisson C H . Vahlsing HI., Hargens AR, Fe- ringa ER: Long-term effects of spinal cord transection on fast and slow rat skeletal muscle I . Contractile properties. Exp Neurul 9 1 :423-434, 1986a.
23. Maier A, Cocket JL, Sinipson DR, Saubert CW I V , Edger- ton VR: Properties of immobilized guinea pig hindlimb muscles. Am J Physiul 231:1520-1526, 1976.
24. Mayer RF, Burke RE, ~I'oop J, Walnisley 9, Hodgson J A : The effect of spinal cord transection on the motor units in cat medial gastrocnemius muscles. Muscle Nerve 7:23-31, 1981.
25. Mayer KF, Burke RE, Toper J , Hodgson JA, Kanda K, Walrnsley B: ~I 'he effect of long-term jmmobilimtion on the motor unit populaton of the cat medial gastrocnemius muscle. NeurusciPncP 6:725-739, 198 1 .
26. Musacchia XJ. Steffen JM. Deaver DR: Rat hindlimb nius- cle responses to suspension hypohinesia/hypodynaniia. Aiial Spare Envirun Med 54:1015-1020, 1983.
27. Nachlas MM,'l'sou K-C, de Souza E. Cheng C-S. Seligman AM: Cytochemical demonstration of succinic dehydroge- nase by the use of a new p-nitrophenyl substituted ditetra- zole. J Mirtuchem Cylocliem 13:420-436, 1957.
28. Pette I), Vrbova G: Neural control of phenotypic expres- sion in mammalian muscle fibers. h f w c l e Nenir 8:676-689, 1985.
29. Powell PI., Roy KR, Kanini P, Bella M, Edgerton VR: Pre- dictability of skeletal muscle tension from architectural de-
91 1435-448, 1986b.
terminat;ons in guinea pig hindlimbs. J Ap/d Physiol 57: I7 15- 172 I , 1984.
30. Rov RR, Sacks RD, Baldwin K M , Short M, Edgerton VK: Interrelationships of contraction time, I/,,,, and myosin A'I'I'ase after spinal transection. J Appl p / i y ~ / ( ~ / 56: 1594- 160 1 , 1981.
31. Sabbadini RA, Baskin RJ: Active state of normal wd dys- trophic mouse muscle. Am J Plryszol 230: 1 138- 1 147, 1976.
32. Salmons S , IIenriksson J : The adaptive response of skcle- tal muscle to increased use. Mwck Nerve 4:94- 105, 1% 1.
33. Simard CP. Spector SA, Edgerton VR: Contractile proper- ties of fat hind limb muscles immobilized at different lengths. EX)J Nezcrol 77:467-482, 1982.
34. Snow D l l , Bi lke r K, Mascarello F, Carpene E, Rowlerson A, Jenny E: N o classical type 1 I R fibres in dog skeletal muscle. flzitochetnist? 75:63-65, 1982.
35. Sokal RR, Rohlf FJ: Riotnehy, 2nd ed. San Fraricisco, W. €i. Freeman and Company, 1981.
36. Spector SA, Simard CP, Fournier M , Sternight E, Edger- ton VR: Architecwral alterations of rat hind-limb skeletal muscles immobilized at different lengths. Exp Ne7truI
37. Tabary JC, Tabdry C. 'l'ardieu C;, 'l'ardieu G , Goldspink G: Physiological and structural changes in the cat's solrus muscle due to immobilization at different lengths. J I'h~siol
38. Templeton GH, Padalino M , Manton 1, Glasber M, Silver CJ, Silver P. DeMartino G. Ixconey T, Klug C , Hagler t i , Stuko JI.: Influence of suspension hypokineasia on rat so- leus musrle. J Appl Ply io l 56:278-286, 1984.
3Y. Weibel ER: Sl~reulopcal hfellioh, Vol. I : Prur/icnl MPtliuds fur Riulupcul MoiphoniPtq. New York, Academic Press, 1980.
40. Wickiewicz TI., Roy KR, Powell PI, Edgerton VR: Muscle architecture of the human lower liinh. Chi1 Orlhop
41. Williams PE, Goldspink G : Connective tissue changes in immobilised muscle. J Anal I38:343-350, 1984.
42. Witzmann FA, Kim DH, Fitts R l l : Hindlimb irnmobiliza- tion: length-tension and contractile properties of skeletal muscle. J A w l Physiul 63:335-345. 1982.
76:94-110, 1982.
(Lolid) 2241231-244, 1972.
179~317-325, 1983.
Differential Atrophy of Dog Quadriceps MUSCLE & NERVE March 1988 201





























