Embed Size (px)
Citation preview

Differential Effects of Selexipag and Prostacyclin Analogsin Rat Pulmonary Artery
Keith Morrison, Rolf Studer, Roland Ernst, Franck Haag, Katalin Kauser,and Martine ClozelDrug Discovery Department, Actelion Pharmaceuticals Ltd, Allschwil, Switzerland
Received June 4, 2012; accepted August 22, 2012
ABSTRACT{4-[(5,6-Diphenylpyrazin-2-yl)(isopropyl)amino]butoxy}aceticacid (ACT-333679) is the main metabolite of the selective pros-tacyclin (PGI2) receptor (IP receptor) agonist selexipag. Thegoal of this study was to determine the influence of IP receptorselectivity on the vasorelaxant efficacy of ACT-333679 and thePGI2 analog treprostinil in pulmonary artery under conditionsassociated with pulmonary arterial hypertension (PAH). Sel-exipag and ACT-333679 evoked full relaxation of pulmonaryartery from control and monocrotaline (MCT)-PAH rats, andACT-333679 relaxed normal pulmonary artery contracted witheither endothelin-1 (ET-1) or phenylephrine. In contrast, trepro-stinil evoked weaker relaxation than ACT-333679 of controlpulmonary artery and failed to induce relaxation of pulmonaryartery from MCT-PAH rats. Treprostinil did not evoke relaxationof normal pulmonary artery contracted with either ET-1 orphenylephrine. Expression of prostaglandin E3 (EP3) receptormRNA was increased in pulmonary artery from MCT-PAH rats.
In contraction experiments, the selective EP3 receptor agonistsulprostone evoked significantly greater contraction of pulmo-nary artery from MCT-PAH rats compared with control rats. Thepresence of a threshold concentration of ET-1 significantlyaugmented the contractile response to sulprostone in normalpulmonary artery. ACT-333679 did not evoke direct contractionof rat pulmonary artery, whereas treprostinil evoked concentra-tion-dependent contraction that was inhibited by the EP3 re-ceptor antagonist (2E)-3-(3�,4�-dichlorobiphenyl-2-yl)-N-(2-thienylsulfonyl)acrylamide. Antagonism of EP3 receptors alsorevealed a relaxant response to treprostinil in normal pulmonaryartery contracted with ET-1. These data demonstrate that therelaxant efficacy of the selective IP receptor agonist selexipagand its metabolite ACT-333679 is not modified under condi-tions associated with PAH, whereas relaxation to treprostinilmay be limited in the presence of mediators of disease.
IntroductionSelexipag [2-{4-[(5,6-diphenylpyrazin-2-yl)(isopropyl)ami-
no]butoxy}-N-(methylsulfonyl)acetamide] is an orally avail-able IP receptor agonist in clinical development for the treat-ment of PAH (Simonneau et al., 2012). Both selexipag and itsactive metabolite {4-[(5,6-diphenylpyrazin-2-yl)(isopropyl)amino]butoxy}acetic acid (ACT-333679) (previously known as MRE-269)possess high selectivity for the IP receptor over other prostanoidreceptors and as such can be distinguished from PGI2 analogscurrently used in the management of PAH (Kuwano et al., 2007).Progression of PAH is associated with reduced PGI2 production(Christman et al., 1992; Tuder et al., 1999), and restoration of IP
receptor signaling using analogs of PGI2 is an effective strategy inthe treatment of the disease (Gomberg-Maitland and Olschewski,2008). Clinical efficacy and tolerability of PGI2 analogs, however,may be compromised by concomitant activation of other prosta-noid receptors. The selectivity of PGI2 replacement therapies forthe IP receptor, therefore, is emerging as an important consider-ation in the management of PAH. For example, vasorelaxation toiloprost and beraprost is attenuated in pulmonary artery frommonocrotaline (MCT)-PAH rats (Kuwano et al., 2008) via a mech-anism that involves coactivation of the contractile EP3 receptor. Incontrast, ACT-333679 has at least 130-fold selectivity for the hu-man IP receptor over other prostanoid receptors (Kuwano et al.,2007), and vasorelaxation of rat pulmonary artery evoked by ACT-333679 is not modulated by antagonism of EP3 receptors (Kuwanoet al., 2008). Moreover, gastric side effects including cramping,nausea, and vomiting may develop via EP3 receptor-dependent
Article, publication date, and citation information can be found athttp://jpet.aspetjournals.org.
http://dx.doi.org/10.1124/jpet.112.197152.
ABBREVIATIONS: PGI2, prostacyclin; IP receptor, PGI2 receptor; EP1 receptor, prostaglandin E receptor 1; EP3 receptor, prostaglandin E receptor3; DP1 receptor, prostaglandin D2 receptor 1; TP receptor, thromboxane A2 receptor; EPA, extralobar pulmonary artery; ET-1, endothelin-1; IPA,intralobar pulmonary artery; MCT, monocrotaline; PAH, pulmonary arterial hypertension; PGF2�, prostaglandin F2�; ACT-333679, {4-[(5,6-diphe-nylpyrazin-2-yl)(isopropyl)amino]butoxy}acetic acid; DBTSA, (2E)-3-(3�,4�-dichlorobiphenyl-2-yl)-N-(2-thienylsulfonyl)acrylamide; SC51322, 8-chloro-2-[3-[(2-furanylmethyl)thio]-1-oxopropyl]hydrazide, dibenz[b,f][1,4]oxazepine-10(11H)-carboxylic acid; GR32191B, (4Z)-7-[(1R,2R,3S,5S)-5-([1,1�-biphenyl]-4-ylme-thoxy)-3-hydroxy-2-(1-piperidinyl)cyclopentyl]-4-heptenoic acid; U46619, 9,11-dideoxy-9�,11�-methanoepoxy prostaglandin F2�.
1521-0103/12/3433-547–555$25.00THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 343, No. 3Copyright © 2012 by The American Society for Pharmacology and Experimental Therapeutics 197152/3803249JPET 343:547–555, 2012
547
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from

mechanisms. Analogs of PGI2 contract gastric smooth muscle viathe stimulation of EP3 receptors (Morrison et al., 2010). Activationof the EP3 receptor subtype mediates disruption of gastric contrac-tility (Pal et al., 2007; Forrest et al., 2009) and underlies thedevelopment of emesis (Kan et al., 2002). Selexipag does not evokegastric contraction or disrupt gastric function in the rat stomach(Morrison et al., 2010). A role for EP3 receptors has also beeninvoked in the development of peripheral pain reported in patientsreceiving treatment with PGI2 analogs (Minami et al., 2001;Southall and Vasko, 2001). Thus, the development of a selective IPreceptor agonist that is devoid of off-target effects may provideimproved efficacy and tolerability in patients with PAH.
Infusion of treprostinil via the subcutaneous or intrave-nous routes is effective in the clinical management of PAH(Simonneau et al., 2002; Tapson et al., 2006). In addition,treprostinil reduces pulmonary arterial pressure, as mea-sured by right ventricular systolic pressure and total pulmo-nary vascular resistance in the rat MCT-PAH model (Yanget al., 2010), but the contribution of dilatation of the pulmo-nary artery to this effect is not clear. Little information onthe direct vasorelaxant efficacy of treprostinil in pulmonaryartery ex vivo is available.
The present study sought to determine the influence of IPreceptor selectivity of ACT-333679 and treprostinil on vas-orelaxation of the rat pulmonary artery under pathologicalconditions associated with PAH. Experiments were per-formed with pulmonary artery isolated from the rat MCT-PAH model. Although development of neointimal and plexi-form lesions, which are characteristic of the human disease(Tuder et al., 2007), is not a feature of this rat model, pul-monary hypertension in MCT-PAH rats is associated withsustained pulmonary arterial vasoconstriction (Oka et al.,2008). The pulmonary artery isolated from this model pro-vides a suitable functional system in which to evaluate vas-orelaxant properties of test compounds. Dysfunction of boththe ET-1 and adrenergic systems underlies the developmentof PAH (Salvi, 1999; Rubens et al., 2001), and circulatinglevels of ET-1 and catecholamines are significantly increasedin the MCT rat model of PAH (Clozel et al., 2006). Thus, thecurrent study measured the ability of ACT-333679 andtreprostinil to relax pulmonary artery from MCT-PAH ratsand control pulmonary artery precontracted with ET-1 or the�-adrenoceptor agonist phenylephrine. The expression levelsof mRNA encoding prostanoid receptors in the MCT ratmodel of PAH were also analyzed to provide a molecularmechanistic explanation for potential differences in arterialreactivity to the test compounds.
The data presented here highlight the importance of IPreceptor selectivity for full vasorelaxant efficacy of IP recep-tor agonists under pathological conditions associated withPAH. Selexipag and its active metabolite ACT-333679 exertfull relaxant efficacy, whereas the ability of treprostinil toevoke pulmonary arterial vasorelaxation is significantly di-minished in disease. The limited relaxant efficacy of trepro-stinil observed in this study may result from coactivation ofcontractile EP3 receptors, which are up-regulated in the MCTrat model of PAH. Potentiation of EP3 receptor-mediatedcontraction by subthreshold concentrations of pathologicalmediators (ET-1 and catecholamines) in control pulmonaryarteries substantiates the impact of this contractile mecha-nism in disease development.
Materials and MethodsAnimals
Male Wistar rats (12 weeks) were obtained from Harlan Labora-tories B.V. (Venray, The Netherlands). All rats were housed in cli-mate-controlled conditions with a 12-h light/dark cycle and had freeaccess to normal pelleted rat chow and drinking water in accordancewith local guidelines (Basel-Landschaft cantonal veterinary office).In certain experiments, PAH was induced in rats by a single injectionof MCT (60 mg/kg i.p.). Vehicle control rats were treated in parallel.Disease was allowed to progress for 30 days before experimentation(Iglarz et al., 2008). Development of disease was confirmed by an in-crease in the weight of right ventricle [right ventricle/(left ventricle �septum) ratio]: control versus MCT-PAH, 0.33 � 0.03 versus 0.81 �0.1 g; P � 0.05) and endothelial dysfunction as measured by atten-uated relaxation to acetylcholine (pEC50 values: control versus MCT-PAH, 7.23 � 0.04 versus 6.06 � 0.13, P � 0.0001; Emax values:control versus MCT-PAH, 82.33 � 1.73 versus 39.79 � 5.26%, P �0.0001).
Rat Isolated Pulmonary Artery
After euthanasia by CO2 asphyxiation, rings of extralobar pulmo-nary artery (EPA) and intralobar pulmonary artery (IPA) were pre-pared from rats using standard techniques. Two or four arterialrings, EPA (1.5 mm) and IPA (1.0 mm), respectively, were preparedfrom each animal. Vessels were suspended between stainless wiresin either a standard tissue bath set-up (EPA) or a Mulvany-Halpernmyograph system (IPA) containing modified Krebs-Henseleit buffer(10 ml) of the following composition: 115 mM NaCl, 4.7 mM KCl, 1.2mM MgSO4, 1.5 mM KH2PO4, 2.5 mM CaCl2, 25 mM NaHCO3, and10 mM glucose. Care was taken to avoid damage to the endotheliumduring the preparation of the arterial rings. In certain experiments,the vascular endothelium was removed by abrasion with the tip of awatchmaker’s forceps. Bathing solution was maintained at 37°C andaerated with 95% O2/5% CO2, pH 7.4. An initial resting force of 4.9mN (EPA) or 3.9 mN (IPA) was applied to vessels, and changes inforce generation were measured by using an isometric force recorder(Multi Wire Myograph System, model 610 M, version 2.2; DMT A/S,Aarhus, Denmark) coupled to a EMKA data acquisition system(EMKA Technologies Inc., Paris, France). Viability of the pulmonaryartery was tested by measuring contraction to KCl (60 mM), and thepresence of a functional endothelium was confirmed by measuringthe ability of acetylcholine 10�5 M to relax arterial rings contractedwith phenylephrine (10�6 M; EPA) or 9,11-dideoxy-9�,11�-metha-noepoxy prostaglandin F2� (U46619) (10�6 M; IPA).
Experimental Protocols
Relaxation of Pulmonary Artery from Control and MCT-PAH Rats. Rings of pulmonary artery were contracted with prosta-glandin F2� (10�5 M). When the developed force had stabilized,cumulative concentration-relaxation curves to selexipag, ACT-333679, beraprost, or treprostinil were obtained. The interval be-tween additions of higher concentrations of compounds to the bathswas determined by the time required for the response to reachplateau.
Relaxation of Pulmonary Artery Contracted with ET-1 orPhenylephrine. Rings of EPA and IPA, in the presence or absenceof endothelium, were contracted with either ET-1 (3 � 10�10 M) orphenylephrine (1–3 � 10�6 M) before cumulative concentration-relaxation curves to ACT-333679 and treprostinil were obtained.
Contraction of Pulmonary Artery. Cumulative concentration-contraction curves to the EP3/EP1 receptor agonist sulprostone wereobtained in the rings of EPA from control and MCT-PAH rats.Separate experiments measured the contractile responses of nor-mal EPA and IPA to ACT-333679 and treprostinil. In experimentsthat sought to characterize the identity of the receptor mediatingcontraction to test compounds, rings of normal IPA were exposed
548 Morrison et al.
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from

to either drug vehicle or receptor antagonists for 20 min beforecumulative concentration-response curves to agonists were ob-tained. The choices and concentrations of receptor antagonistswere based on published data. The following receptor antagonistswere used: (2E)-3-(3�,4�-dichlorobiphenyl-2-yl)-N-(2-thienylsulfo-nyl)acrylamide (DBTSA) (EP3 receptor; Gallant et al., 2002;Kuwano et al., 2008); 8-chloro-2-[3-[(2-furanylmethyl)thio]-1-oxopropyl]hydrazide, dibenz[b,f][1,4]oxazepine-10(11H)-carboxy-lic acid (SC51322) (EP1 receptor; McCormick et al., 2010), and (4Z)-7-[(1R,2R,3S,5S)-5-([1,1’-biphenyl]-4-ylmethoxy)-3-hydroxy-2-(1-piperidinyl)cyclopentyl]-4-heptenoic acid (GR32191B) (TP receptor;Lumley et al., 1989).
Measurement of Receptor mRNA Expression
Prostanoid receptor mRNA expression was measured by quanti-tative polymerase chain reaction in arteries from normal and MCT-PAH Wistar rats. Analyses were performed by using the main, left,and right branches of the pulmonary artery. Total RNA was isolatedfrom arteries (30-mg samples) by using the RNeasy micro fibroustissue kit according to the manufacturer’s protocol (QIAGEN GmbH,Hilden, Germany). Remaining genomic DNA was digested by usingthe DNase free kit (Ambion, Austin, TX). The quantity of RNA wasanalyzed by using a Nanodrop spectrophotometer (Thermo FisherScientific, Waltham, MA), and RNA quality was assessed by using aBioanalyzer 2100 (Agilent Technologies, Santa Clara, CA). TotalRNA was reverse-transcribed with a high-capacity cDNA archive kit(Applied Biosystems, Foster City, CA). Quantitative polymerasechain reaction was performed on an ABI 7500 machine using Taq-Man probes (Applied Biosystems). The TaqMan assays from AppliedBiosystems used for mRNA detection were IP-Rn01764022_m1,DP1-Rn04223963_g1, EP1-Rn01432713_s1, EP3-Rn0121735_m1;TP-Rn00690601_m1; 18s-4319413E, B2M-Rn00560865_m1 andGUS-Rn00566655_m1 were used for normalization. Results werecalculated by using the ��Ct method, which allows comparisonbetween gene expression values for different genes based on anidentical linear scale. A value of 1 was defined as no expression.Final results are presented as the PAH/control rat ratio.
Materials
Selexipag, ACT-333679, and DBTSA were synthesized by NipponShinyaku Co. Ltd (Kyoto, Japan). Beraprost, treprostinil, SC51322, andsulprostone were obtained from Cayman Chemical (Ann Arbor, MI).Acetylcholine, GR32191B, phenylephrine, prostaglandin F2�, andU46619 were purchased from Sigma (St Louis, MO).
Statistical Evaluation of Results
Responses of rat pulmonary artery to test compounds are ex-pressed either as a percentage of the precontraction or the referencecontraction to KCl (60 mM). Contraction to phenylephrine and ET-1was measured in millinewtons. Results are presented as mean �S.E.M. In some experiments, the S.E.M. values are smaller than thedata symbol. n values refer to the number of animals. pEC50 valuesare defined as the negative logarithm of the concentration of agonistthat evoked half-maximal response. The effects of receptor antago-nists on responses of pulmonary artery to analogs of PGI2 were
quantified by comparing calculated areas under the agonist concen-tration-response curves in the absence and presence of antagonists.Statistical comparisons between control and treated groups wereperformed by using either Student’s paired t test or Student’s un-paired t test (two-tailed). Significance was accepted at P � 0.05.
ResultsSelexipag/ACT-333679, but Not Prostacyclin Ana-
logs, Relax Pulmonary Artery from MCT-PAH Rats.The abilities of selexipag, metabolite ACT-333679, and theanalogs of prostacyclin, beraprost, and treprostinil to relaxrat pulmonary artery were measured. Arterial rings fromcontrol and MCT-PAH rats were contracted to equal degreeswith PGF2� (10�5 M) (control versus MCT-PAH; P 0.05).Selexipag and ACT-333679 evoked concentration-dependentrelaxation of EPA rings contracted with PGF2� (10�5 M).Relaxation was similar in EPA from control and MCT-PAHrats (Table 1; Fig. 1) (areas under curves, P 0.05). Bera-prost and treprostinil evoked concentration-dependent relax-ation of control rat EPA, but Emax values were significantlyless than that measured in response to selexipag and ACT-333679 (Table 1; Fig. 1). Likewise, relaxation of EPA fromMCT-PAH rats to beraprost and treprostinil was signifi-cantly reduced compared with control EPA (Table 1; Fig. 1)(areas under curves: beraprost, P � 0.05; treprostinil, P �0.0001). In particular, relaxation to treprostinil was abol-ished in EPA from MCT-PAH rats.
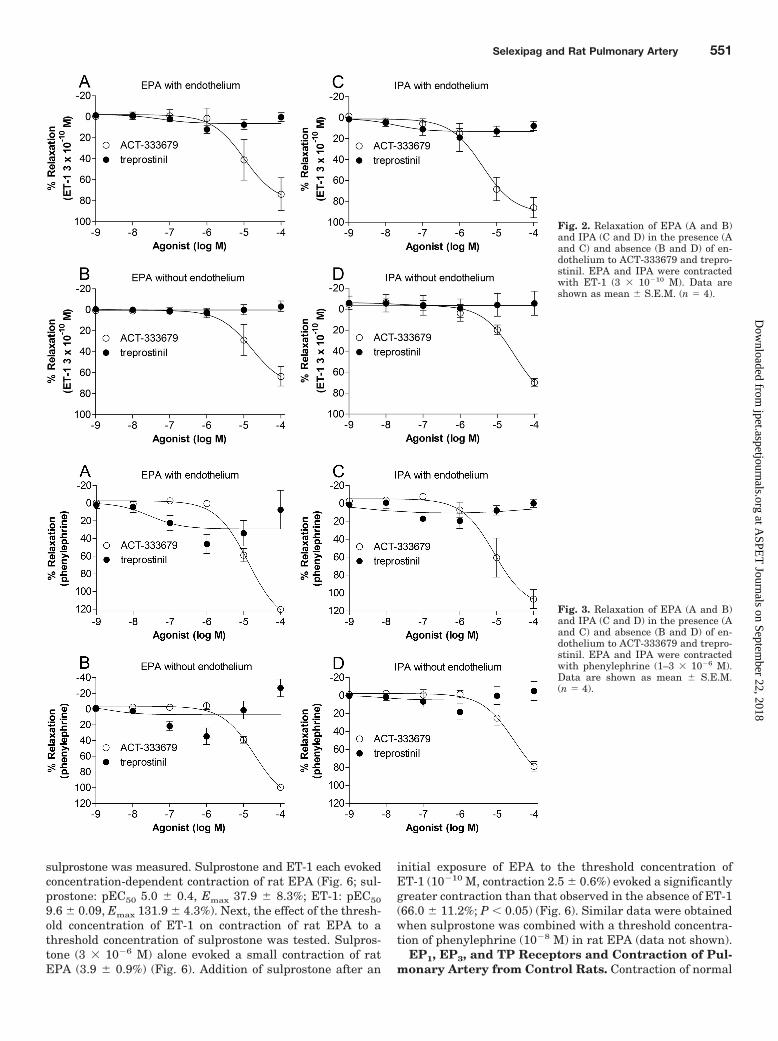
ACT-333679, but Not Treprostinil, Relaxes Pulmo-nary Artery Contracted with ET-1. ET-1 is a key media-tor in the development of PAH (Rubens et al., 2001; Clozel etal., 2006). Thus, the ability of ACT-333679 and treprostinil toevoke vasorelaxation of pulmonary artery contracted withET-1 was measured in both the presence and absence offunctional endothelium. Rings of EPA and IPA from normalrats were contracted with ET-1 (3 � 10�10 M), and when tonehad stabilized, cumulative concentration-response curves toACT-333679 and treprostinil were obtained. Contraction toET-1 was similar in the presence or absence of endotheliumwithin each pulmonary artery group (Table 2). ACT-333679evoked concentration-dependent relaxation of normal EPAand IPA rings (Table 3; Fig. 2). In contrast, treprostinil didnot evoke relaxation of either EPA or IPA rings, even at thehighest concentration tested (Table 3; Fig. 2). Removal of theendothelium did not influence the responsiveness of EPA andIPA rings to treprostinil. Removal of endothelial cells in IPA,however, caused a small, but statistically significant, de-crease in the pEC50 value for ACT-333679 (Table 3).
ACT-333679, but Not Treprostinil, Relaxes Pulmo-nary Artery Contracted with Phenylephrine. Hyperac-tivation of the sympathetic nervous system is reported in
TABLE 1Summary of relaxation data for selexipag, ACT-333679, beraprost, and treprostinil in pulmonary artery from control and MCT-PAH rats
Selexipag ACT-333679 Beraprost Treprostinil
Control ratspEC50 5.2 � 0.1 5.9 � 0.1 6.18 � 0.2 6.88 � 0.1Emax 98.4 � 2.1 102.3 � 0.7 81.1 � 6.9* 72.8 � 3.7**
MCT-PAH ratspEC50 4.9 � 0.1 5.5 � 0.1 4.7 � 0.2 N.D.Emax 104.6 � 2.4 104.5 � 1.0 57.5 � 7.8** 4.3 � 2.3***
Emax, percentage of reference contraction to KCl (60 mM); N.D., not determined.*, P � 0.05; **, P � 0.001; ***, P � 0.0001 comparison with Emax value for ACT-333679.
Selexipag and Rat Pulmonary Artery 549
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from

PAH (Haneda et al., 1983; Salvi, 1999; Nagaya et al., 2000;Velez-Roa et al., 2004), and circulating levels of cat-echolamines are increased in the MCT model of PAH (Clozelet al., 2006). Vasorelaxation of EPA and IPA rings to ACT-333679 and treprostinil, in the presence and absence of en-dothelium, was therefore measured after contraction of ringswith the �1-adrenoceptor agonist phenylephrine. ACT-333679evoked concentration-dependent relaxation of normal EPAand IPA rings contracted with phenylephrine, whereastreprostinil did not evoke relaxation (Table 3; Fig. 3). Re-moval of the endothelium did not influence the responsive-ness of EPA and IPA rings to treprostinil. A small, but
statistically significant, decrease in the pEC50 value for ACT-333679 was measured upon removal of the endothelium inIPA (Table 3), but no significant differences in responses ofIPA, with or without endothelium, were observed over thefull range of concentrations tested (areas under curve: withand without endothelium, P 0.05).
Sulprostone Contracts Pulmonary Artery from Con-trol and MCT-PAH Rats. The potential role of EP3 recep-tors in modulating vasoreactivity of the pulmonary arteryfrom MCT-induced PAH rats was investigated. First, theability of the selective EP3 receptor agonist sulprostone tocontract rat EPA from control and MCT-PAH rats was mea-sured. Sulprostone evoked concentration-dependent contrac-tion of EPA, which was significantly more pronounced inrings from MCT-PAH compared with control rats (Fig. 4)(Emax values control versus MCT-PAH: 62.7 � 7.8 versus168.5 � 20.6%, P � 0.001).
Differential Expression of Prostanoid Receptorsfrom Control and MCT-PAH Rats. The expression profileof prostanoid receptors in pulmonary artery from control andMCT-PAH rats was measured (Fig. 5). TP and EP1 showedthe highest basal expression, IP and EP3 showed moderateexpression, and DP1 showed low expression (data notshown). mRNA that encoded the IP receptor was signifi-cantly increased in all segments of pulmonary artery ana-lyzed from MCT-PAH rats (P � 0.01). Likewise, EP3 wassignificantly up-regulated in the main branch of MCT-PAHpulmonary artery (P � 0.05). In contrast, DP1 (P � 0.05),EP1 (P � 0.001), and TP (P � 0.01) were significantly down-regulated in arteries from MCT-PAH rats.
Effect of Threshold Concentration of ET-1 on Con-traction of Pulmonary Artery to Sulprostone. Becausethe function of the EP3 receptor subtype seems to be in-creased in pulmonary artery from MCT-PAH rats, the influ-ence of ET-1 on contraction of control pulmonary artery to
Fig. 1. Relaxation of pulmonary artery(EPA) from control and MCT-PAH ratsto selexipag (A), ACT-333679 (B), bera-prost (C), and treprostinil (D). Rings ofEPA were contracted with PGF2� (10�5
M). Data are shown as mean � S.E.M.(n 4).
TABLE 2Contraction of rat pulmonary artery to ET-1 and phenylephrine
Contractile Agent/Relaxant
EPA IPA
WithEndothelium
WithoutEndothelium
WithEndothelium
WithoutEndothelium
mN
ET-1/ACT-333679 8.9 � 1.3 10.3 � 1.1 3.5 � 0.4 3.2 � 0.3ET-1/treprostinil 8.1 � 0.9 10.5 � 1.0 2.7 � 0.3 2.8 � 0.3Phenylephrine/
ACT-3336795.5 � 0.8 7.6 � 1.2 3.1 � 0.2 2.8 � 0.3
Phenylephrine/treprostinil
6.5 � 0.9 6.7 � 0.5 3.1 � 0.1 3.4 � 0.2
TABLE 3Relaxant potency of ACT-333679 in rat pulmonary artery
ContractileAgent
EPA IPA
WithEndothelium
WithoutEndothelium
WithEndothelium
WithoutEndothelium
pEC50
ET-1 5.0 � 0.3 4.8 � 0.3 5.4 � 0.2 4.5 � 0.2*Phenylephrine 4.9 � 0.1 4.7 � 0.1 5.1 � 0.2 4.5 � 0.1*
*, P � 0.05, comparison between IPA with and without endothelium.
550 Morrison et al.
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from
sulprostone was measured. Sulprostone and ET-1 each evokedconcentration-dependent contraction of rat EPA (Fig. 6; sul-prostone: pEC50 5.0 � 0.4, Emax 37.9 � 8.3%; ET-1: pEC50
9.6 � 0.09, Emax 131.9 � 4.3%). Next, the effect of the thresh-old concentration of ET-1 on contraction of rat EPA to athreshold concentration of sulprostone was tested. Sulpros-tone (3 � 10�6 M) alone evoked a small contraction of ratEPA (3.9 � 0.9%) (Fig. 6). Addition of sulprostone after an
initial exposure of EPA to the threshold concentration ofET-1 (10�10 M, contraction 2.5 � 0.6%) evoked a significantlygreater contraction than that observed in the absence of ET-1(66.0 � 11.2%; P � 0.05) (Fig. 6). Similar data were obtainedwhen sulprostone was combined with a threshold concentra-tion of phenylephrine (10�8 M) in rat EPA (data not shown).
EP1, EP3, and TP Receptors and Contraction of Pul-monary Artery from Control Rats. Contraction of normal
Fig. 2. Relaxation of EPA (A and B)and IPA (C and D) in the presence (Aand C) and absence (B and D) of en-dothelium to ACT-333679 and trepro-stinil. EPA and IPA were contractedwith ET-1 (3 � 10�10 M). Data areshown as mean � S.E.M. (n 4).
Fig. 3. Relaxation of EPA (A and B)and IPA (C and D) in the presence (Aand C) and absence (B and D) of en-dothelium to ACT-333679 and trepro-stinil. EPA and IPA were contractedwith phenylephrine (1–3 � 10�6 M).Data are shown as mean � S.E.M.(n 4).
Selexipag and Rat Pulmonary Artery 551
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from

rat EPA and IPA to ACT-333679 and treprostinil was mea-sured. ACT-333679 did not contract rings of pulmonary ar-tery, whereas treprostinil evoked concentration-dependentcontraction of both EPA and IPA. Contraction to treprostinilwas more pronounced in IPA than in EPA rings (EPA: Emax
22.5 � 5.8%; IPA: Emax 62.8 � 11.9%) (Fig. 7). The EP3
receptor antagonist DBTSA (3 � 10�6 M) abolished contrac-tion of rat IPA to treprostinil (Fig. 7). The EP1 receptorantagonist SC51322 (10�6 M) had no effect on contraction ofIPA to treprostinil (control versus treated: pEC50, 4.8 � 0.15versus 4.8 � 0.15, P 0.05; Emax, 52.5 � 5.5 versus 50.8 �5.6%, P 0.05). Likewise, antagonism of TP receptors withGR32191B (10�6 M) did not significantly inhibit contractionto treprostinil (control versus treated: pEC50, 4.5 � 0.4 ver-sus 4.5 � 0.3, P 0.05; Emax, 34.8 � 11.3 versus 22.0 � 5.2%,P 0.05).
EP3 Receptors and Relaxation of IPA from NormalRats. The influence of DBTSA on the relaxant response of ratIPA to treprostinil was measured next. Rings of IPA, in theabsence or presence of DBTSA, were contracted to the samelevel with ET-1 (control versus DBTSA groups, 3.08 � 0.5versus 2.8 � 0.6 mN; P 0.05). In the absence of DBTSA,treprostinil evoked minimal relaxation of IPA (Fig. 8). An-tagonism of EP3 receptors with DBTSA, however, revealed asignificant concentration-dependent relaxation of rat IPA totreprostinil (Fig. 8) (control versus treated: areas undercurves, P � 0.05).
DiscussionSelexipag and the active metabolite ACT-333679 can be
distinguished from analogs of PGI2 based on vasorelaxantefficacy in the rat pulmonary artery. Specifically, both sel-exipag and ACT-333679, but not the PGI2 analog treprosti-
nil, evoke concentration-dependent relaxation of pulmonaryartery under pathological conditions associated with PAH.Increased expression of mRNA that encodes EP3 receptorsand enhanced pharmacological activity of this receptor sub-type are measured in pulmonary artery from the MCT ratmodel of PAH, but do not influence the relaxant efficacy ofthe selective IP receptor agonist ACT-333679. The limitedvasorelaxant efficacy of treprostinil was partially restoredafter EP3 receptor blockade, indicating that coactivation ofcontractile EP3 receptors attenuates the relaxant action oftreprostinil.
ACT-333679 is highly selective for the human IP receptor(Kuwano et al., 2007). The Ki value for ACT-333679 at the IPreceptor was 130-fold lower than that at other prostanoidreceptors. The affinity value for ACT-333679 at the rat IPreceptor was 2.2 � 10�7 M and would be expected to activateIP receptors at the concentrations used in the current study.Treprostinil, however, is a nonselective agonist at prostanoidreceptors (Whittle et al., 2012). In particular, treprostinilbinds and activates DP1 and EP2 receptors in addition to theIP receptor. Treprostinil regulates macrophage function andpromotes survival of bacteria via an EP2 receptor-dependentmechanism (Aronoff et al., 2007). In addition, treprostinilactivates contractile EP3 receptors in gastric (Morrison et al.,2010) and vascular smooth muscle (Orie and Clapp, 2011).
The relaxant response to ACT-333679 was similar in pul-monary artery from control and MCT-PAH rats. These dataconfirm previous findings (Kuwano et al., 2008). In contrast,treprostinil evoked weaker relaxation than ACT-333679 inpulmonary artery from control rats and failed to relax pul-monary artery from MCT-PAH rats. Diminished relaxationof pulmonary artery from MCT-PAH rats to iloprost andberaprost has been reported (Kuwano et al., 2008), buttreprostinil seems to be particularly sensitive to the patho-logical conditions associated with PAH. This possibility wasfurther examined in experiments that measured the relaxantefficacy of ACT-333679 and treprostinil in pulmonary arterycontracted with ET-1 or the �1-adrenoceptor agonist phenyl-ephrine. Dysfunction of both the ET-1 and adrenergic sys-tems underlie the progression of PAH (Salvi, 1999; Rubenset al., 2001), and circulating levels of ET-1 and cat-echolamines are significantly increased in the rat MCT-PAHmodel (Clozel et al., 2006). The relaxant profile of ACT-333679 and treprostinil was remarkably similar to that ob-served in MCT-PAH rats: ACT-333679 evoked concentration-dependent relaxation of all arterial rings tested, whereastreprostinil was unable to relax pulmonary artery contractedwith ET-1 or phenylephrine.
Treprostinil can activate contractile EP3 receptors thatcounter the vasorelaxant properties of the drug (Orie andClapp, 2011). Contraction evoked by the selective EP3 recep-tor agonist sulprostone (e.g., Dong et al., 1986) is signifi-cantly greater in pulmonary artery from MCT-PAH rats thanin control rats and is consistent with previous findings (Ku-wano et al., 2008). Hyper-responsiveness of the MCT-PAHpulmonary artery to sulprostone may reflect an increase inthe number of EP3 receptors. Receptor density is a key de-terminant of both potency and efficacy of an agonist (Neubiget al., 2003). Molecular analyses of pulmonary artery fromcontrol and MCT-PAH rats revealed a modest, but statisti-cally significant, up-regulation of the expression of mRNAencoding EP3 receptors in the disease condition. Thus, both
Fig. 4. Contraction of pulmonary artery (EPA) from control and MCT-PAH rats to sulprostone. Data are shown as mean � S.E.M. (n 6).
Fig. 5. Expression of prostanoid receptors in pulmonary artery fromcontrol and MCT-PAH rats. Data are expressed as the PAH/control ratratio (n 6). S.E.M. values are omitted for clarity. �, P � 0.05; ��, P �0.01; ���, P � 0.001.
552 Morrison et al.
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from

expression and pharmacological activity of contractile EP3
receptors are enhanced in pulmonary artery from MCT-PAHrats. A differential regulation of IP and TP receptor mRNAexpression was also noted: an up-regulation of IP receptormRNA whereas down-regulation of mRNA encoding TP re-ceptors was recorded. These data are suggestive of a phar-macological compensatory mechanism to combat disease pro-gression through improved PGI2 signaling (IP receptor) anddiminished deleterious effects of thromboxane A2 (TP recep-tor) (Christman et al., 1992; Tuder et al., 1999). Publishedinformation on the regulation of prostanoid receptors in PAH
is, however, contradictory. Down-regulation in expression ofIP receptors is reported in pulmonary arterial smooth musclecells cultured from MCT-PAH rats (Lai et al., 2008) andpulmonary arterial smooth muscle cells and nondefined lungtissue from human patients with PAH (Lai et al., 2008;Falcetti et al., 2010). In contrast, a genomewide RNA expres-sion study did not observe significant regulation of IP or EP3
receptor mRNA in lung tissue samples from patients withPAH (Rajkumar et al., 2010). Differences in the observationsbetween studies may reflect the experimental approachesadopted. For example, in the present study receptor mRNA
Fig. 6. Contraction of rat pulmonaryartery (EPA) to sulprostone and ET-1.A and B, concentration-dependentcontraction of EPA to sulprostone (A)and ET-1 (B). C and D, representativetraces of contractile responses of EPAto sulprostone (3 � 10�6 M) alone (C)or in combination with ET-1 (10�10 M)(D). E, quantification of contractiledata shown in C and D. Data areshown as mean � S.E.M. (n 4). �,P � 0.05.
Fig. 7. Contraction of rat pulmonary artery to ACT-333679 and treprostinil. A and B, concentration-dependent contraction of EPA and IPA toACT-333679 (A) and treprostinil (B). C, DBTSA (3 � 10�6 M) inhibits contraction of IPA to treprostinil. Data are expressed as mean � S.E.M. (n 4). �, P � 0.05.
Selexipag and Rat Pulmonary Artery 553
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from

was measured in arterial blood vessels that were directlyisolated from MCT-PAH rats. An additional cell culture stagewas included in previous work and may have incurredchanges in receptor expression (e.g., Boess et al., 2003). Inaddition, nonvascular components in snap-frozen lung tissuemay have altered the receptor expression profile. Despite thevariability of results, any potential changes in the expressionof IP receptors should affect the interaction with selexipagand treprostinil equally, because both drugs bind the humanIP receptor with similar affinity (Ki values, 2 � 10�8 and3.2 � 10�8 M, respectively) (Kuwano et al., 2007; Whittle etal., 2012). Vascular responsiveness to a nonselective prosta-noid receptor agonist, such as treprostinil, however, may bemore susceptible to changes in the overall expression patternand pharmacological activity of prostanoid receptors.
Increased pharmacological activity of the EP3 receptor wasalso evident in combination experiments that measured con-traction to sulprostone and ET-1 or phenylephrine in pulmo-nary arteries isolated from control rats. Contraction of pul-monary artery to sulprostone was significantly augmented inthe presence of a threshold concentration of ET-1 or phenyl-ephrine and raises the possibility of a synergistic interactionbetween the EP3 and endothelin and adrenergic receptorsystems. Contractile synergism between ET-1 and serotoninis reported in the human mammary artery (Yang et al.,1990), and significant synergy exists between the �-adreno-ceptor and EP3 receptors in rat femoral artery (Hung et al.,2006). Coactivation of EP3 receptors under pathological con-ditions of PAH may limit the therapeutic efficacy of trepros-tinil. The current study and other work (Orie and Clapp,2011) demonstrate a clear role for the EP3 receptor subtypein vasoconstriction to treprostinil. The beneficial effects of IPreceptor signaling on vascular wall remodeling in PAH canpotentially be tempered by excessive vasoconstriction. In-deed, the EP3 receptor mediates contraction of human pul-monary artery (Maddox et al., 1985; Haye-Legrand et al.,1987; Norel et al., 1991; Qian et al., 1994; Walch et al., 1999)and may modulate relaxant IP receptor function (Jones et al.,1997; Chan and Jones, 2004). In addition, EP3 receptor sig-naling is profibrotic (White et al., 2008; Bozyk and Moore,2011; Harding and LaPointe, 2011) and may limit the bene-ficial effects of treprostinil in PAH.
Failure of treprostinil to evoke relaxation of the pulmonaryartery from MCT-PAH rats may not be fully explained by thesmall down-regulation of mRNA encoding relaxant DP1 re-ceptors in this model. A role for EP2 receptors can similarlybe discounted because no change in the expression of mRNA
encoding this receptor subtype is measured in MCT-PAHrats (Lai et al., 2008). Indeed, agonism at DP1 and EP2
receptors does not evoke relaxation of human pulmonaryartery (Walch et al., 1999).
The vasorelaxant efficacy of ACT-333679 measured here inrat pulmonary artery ex vivo reflects the high selectivity ofthis compound for the relaxant IP receptor (Kuwano et al.,2007). Furthermore, vasorelaxation evoked by ACT-333679under pathological conditions associated with PAH (absenceof endothelium, ET-1, and �-adrenoceptor stimulation) sup-ports the therapeutic profile of selexipag for the treatment ofPAH. The combined vasodilator and antiproliferative prop-erties of ACT-333679 (Kuwano et al., 2007, 2008) probablylead to the prolonged survival of MCT-PAH rats (Kuwanoet al., 2008) and contribute to the significant decrease inpulmonary vascular resistance in patients with PAH (Simo-nneau et al., 2012). In contrast, the present data suggest thatthe therapeutic benefits of treprostinil in the management ofPAH may not be attributed solely to direct vasodilatation.Indeed, only a small, but statistically significant, reductionin right ventricular systolic pressure is reported in the MCT-PAH rat (Yang et al., 2010). Infusion of treprostinil in atherapeutic regimen effectively prevents smooth muscle pro-liferation and progression of PAH in the rat MCT-PAH model(Yang et al., 2010), although the impact on survival is notreported. In addition, treprostinil exerts antiproliferative ef-fects via an IP receptor-dependent mechanism in a recombi-nant cell system (Falcetti et al., 2007).
In conclusion, these data demonstrate that selexipag/ACT-333679 and treprostinil differ in their vasorelaxant effects inpulmonary artery. Selexipag/ACT-333679 relax pulmonaryartery under conditions associated with PAH, whereastreprostinil fails to evoke relaxation. The relaxant efficacy oftreprostinil depends on the nature of the agonist used tocontract the pulmonary artery and is also attenuated byoff-target agonism at EP3 receptors. Thus, a high degree ofselectivity for the target IP receptor may help maintain ther-apeutic efficacy and minimize unwanted side effects of IPreceptor agonists.
Acknowledgments
We thank Marie Schnoebelen for technical assistance.
Authorship Contributions
Participated in research design: Morrison, Studer, Kauser, andClozel.
Conducted experiments: Studer, Ernst, and Haag.Performed data analysis: Morrison, Studer, Ernst, and Haag.Wrote or contributed to the writing of the manuscript: Morrison,
Studer, Kauser, and Clozel.
ReferencesAronoff DM, Peres CM, Serezani CH, Ballinger MN, Carstens JK, Coleman N, Moore
BB, Peebles RS, Faccioli LH, and Peters-Golden M (2007) Synthetic prostacyclinanalogs differentially regulate macrophage function via distinct analog-receptorbinding specificities. J Immunol 178:1628–1634.
Boess F, Kamber M, Romer S, Gasser R, Muller D, Albertini S, and Suter L (2003)Gene expression in two hepatic cell lines, cultured primary hepatocytes, and liverslices compared to the in vivo liver gene expression in rats: possible implicationsfor toxicogenomics use of in vitro systems. Toxicol Sci 73:386–402.
Bozyk PD and Moore BB (2011) Prostaglandin E2 and the pathogenesis of pulmonaryfibrosis. Am J Respir Cell Mol Biol 45:445–452.
Chan KM and Jones RL (2004) Partial agonism of taprostene at prostanoid IPreceptors in vascular preparations from guinea-pig, rat, and mouse. J CardiovascPharmacol 43:795–807.
Christman BW, McPherson CD, Newman JH, King GA, Bernard GR, Groves BM,
Fig. 8. Effect of DBTSA (3 � 10�6 M) on relaxation of rat pulmonaryartery (IPA, with endothelium) to treprostinil. IPA was contracted withET-1 (3 � 10�10 M). Data are expressed as mean � S.E.M. (n 4).
554 Morrison et al.
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from

and Loyd JE (1992) An imbalance between the excretion of thromboxane andprostacyclin metabolites in pulmonary hypertension. N Engl J Med 327:70–75.
Clozel M, Hess P, Rey M, Iglarz M, Binkert C, and Qiu C (2006) Bosentan, sildenafil,and their combination in the monocrotaline model of pulmonary hypertension inrats. Exp Biol Med (Maywood) 231:967–973.
Dong YJ, Jones RL, and Wilson NH (1986) Prostaglandin E receptor subtypes insmooth muscle: agonist activities of stable prostacyclin analogues. Br J Pharmacol87:97–107.
Falcetti E, Flavell DM, Staels B, Tinker A, Haworth SG, and Clapp LH (2007) IPreceptor-dependent activation of PPAR� by stable prostacyclin analogues.Biochem Biophys Res Commun 360:821–827.
Falcetti E, Hall SM, Phillips PG, Patel J, Morrell NW, Haworth SG, and Clapp LH(2010) Smooth muscle proliferation and role of the prostacyclin (IP) receptor inidiopathic pulmonary arterial hypertension. Am J Respir Crit Care Med 182:1161–1170.
Forrest AS, Hennig GW, Jokela-Willis S, Park CD, and Sanders KM (2009) Prosta-glandin regulation of gastric slow waves and peristalsis. Am J Physiol GastrointestLiver Physiol 296:G1180–G1190.
Gallant M, Carriere MC, Chateauneuf A, Denis D, Gareau Y, Godbout C, Greig G,Juteau H, Lachance N, Lacombe P, et al. (2002) Structure-activity relationship ofbiaryl acylsulfonamide analogues on the human EP3 prostanoid receptor. BioorgMed Chem Lett 12:2583–2586.
Gomberg-Maitland M and Olschewski H (2008) Prostacyclin therapies for the treat-ment of pulmonary arterial hypertension. Eur Respir J 31:891–901.
Haneda T, Nakajima T, Shirato K, Onodera S, and Takishima T (1983) Effects ofoxygen breathing on pulmonary vascular input impedance in patients with pul-monary hypertension. Chest 83:520–527.
Harding P and LaPointe MC (2011) Prostaglandin E2 increases cardiac fibroblastproliferation and increases cyclin D expression via EP1 receptor. ProstaglandinsLeukot Essent Fatty Acids 84:147–152.
Haye-Legrand I, Bourdillat B, Labat C, Cerrina J, Norel X, Benveniste J, and BrinkC (1987) Relaxation of isolated human pulmonary muscle preparations with pros-tacyclin (PGI2) and its analogs. Prostaglandins 33:845–854.
Hung GH, Jones RL, Lam FF, Chan KM, Hidaka H, Suzuki M, and Sasaki Y (2006)Investigation of the pronounced synergism between prostaglandin E2 and otherconstrictor agents on rat femoral artery. Prostaglandins Leukot Essent Fatty Acids74:401–415.
Iglarz M, Binkert C, Morrison K, Fischli W, Gatfield J, Treiber A, Weller T, Bolli MH,Boss C, Buchmann S, et al. (2008) Pharmacology of macitentan, an orally activetissue-targeting dual endothelin receptor antagonist. J Pharmacol Exp Ther 327:736–745.
Jones RL, Qian YM, Wise H, Wong HN, Lam WL, Chan HW, Yim AP, and Ho JK(1997) Relaxant actions of nonprostanoid prostacyclin mimetics on human pulmo-nary artery. J Cardiovasc Pharmacol 29:525–535.
Kan KK, Jones RL, Ngan MP, and Rudd JA (2002) Actions of prostanoids to induceemesis and defecation in the ferret. Eur J Pharmacol 453:299–308.
Kuwano K, Hashino A, Asaki T, Hamamoto T, Yamada T, Okubo K, and KuwabaraK (2007) 2-{4-[(5,6-Diphenylpyrazin-2-yl)(isopropyl)amino]butoxy}-N-(methylsul-fonyl)acetamide (NS-304), an orally available and long-acting prostacyclin recep-tor agonist prodrug. J Pharmacol Exp Ther 322:1181–1188.
Kuwano K, Hashino A, Noda K, Kosugi K, and Kuwabara K (2008) A long-acting andhighly selective prostacyclin receptor agonist prodrug, 2-{4-[(5,6-diphenylpyrazin-2-yl)(isopropyl)amino]butoxy}-N-(methylsulfonyl)acetamide (NS-304), amelioratesrat pulmonary hypertension with unique relaxant responses of its active form,{4-[(5,6-diphenylpyrazin-2-yl)(isopropyl)amino]butoxy} acetic acid (MRE-269), onrat pulmonary artery. J Pharmacol Exp Ther 326:691–699.
Lai YJ, Pullamsetti SS, Dony E, Weissmann N, Butrous G, Banat GA, Ghofrani HA,Seeger W, Grimminger F, and Schermuly RT (2008) Role of the prostanoid EP4receptor in iloprost-mediated vasodilatation in pulmonary hypertension. Am JRespir Crit Care Med 178:188–196.
Lumley P, White BP, and Humphrey PP (1989) GR32191, a highly potent andspecific thromboxane A2 receptor blocking drug on platelets and vascular andairways smooth muscle in vitro. Br J Pharmacol 97:783–794.
Maddox Y, Cunard CM, Shapiro R, Kawaguchi A, Goldman M, Lower RR, andRamwell PW (1985) A comparison of the contractile responses of rodent andhuman pulmonary vascular segments to Eicosanoids, in Prostaglandins, Leuko-trienes and Lipoxins (Martyn Bailey J ed) pp 267–272, Plenum Press, New York.
McCormick C, Jones RL, Kennedy S, and Wadsworth RM (2010) Activation ofprostanoid EP receptors by prostacyclin analogues in rabbit iliac artery: implica-tions for anti-restenotic potential. Eur J Pharmacol 641:160–167.
Minami T, Nakano H, Kobayashi T, Sugimoto Y, Ushikubi F, Ichikawa A, NarumiyaS, and Ito S (2001) Characterization of EP receptor subtypes responsible forprostaglandin E2-induced pain responses by use of EP1 and EP3 receptor knockoutmice. Br J Pharmacol 133:438–444.
Morrison K, Ernst R, Hess P, Studer R, and Clozel M (2010) Selexipag: a selectiveprostacyclin receptor agonist that does not affect rat gastric function. J PharmacolExp Ther 335:249–255.
Nagaya N, Nishikimi T, Uematsu M, Satoh T, Kyotani S, Sakamaki F, Kakishita M,Fukushima K, Okano Y, Nakanishi N, et al. (2000) Plasma brain natriuretic
peptide as a prognostic indicator in patients with primary pulmonary hyperten-sion. Circulation 102:865–870.
Neubig RR, Spedding M, Kenakin T, Christopoulos A, and International Union ofPharmacology Committee on Receptor Nomenclature and Drug Classification(2003) International Union of Pharmacology Committee on Receptor Nomencla-ture and Drug Classification. XXXVIII. Update on terms and symbols in quanti-tative pharmacology. Pharmacol Rev 55:597–606.
Norel X, Labat C, Gardiner PJ, and Brink C (1991) Inhibitory effects of BAY u3405on prostanoid-induced contractions in human isolated bronchial and pulmonaryarterial muscle preparations. Br J Pharmacol 104:591–595.
Oka M, Fagan KA, Jones PL, and McMurtry IF (2008) Therapeutic potential ofRhoA/Rho kinase inhibitors in pulmonary hypertension. Br J Pharmacol 155:444–454.
Orie NN and Clapp LH (2011) Role of prostanoid IP and EP receptors in mediatingvasorelaxant responses to PGI2 analogues in rat tail artery: evidence for Gi/omodulation via EP3 receptors. Eur J Pharmacol 654:258–265.
Pal A, Brasseur JG, and Abrahamsson B (2007) A stomach road or “Magenstrasse”for gastric emptying. J Biomech 40:1202–1210.
Qian YM, Jones RL, Chan KM, Stock AI, and Ho JK (1994) Potent contractile actionsof prostanoid EP3-receptor agonists on human isolated pulmonary artery. Br JPharmacol 113:369–374.
Rajkumar R, Konishi K, Richards TJ, Ishizawar DC, Wiechert AC, Kaminski N, andAhmad F (2010) Genomewide RNA expression profiling in lung identifies distinctsignatures in idiopathic pulmonary arterial hypertension and secondary pulmo-nary hypertension. Am J Physiol Heart Circ Physiol 298:H1235–H1248.
Rubens C, Ewert R, Halank M, Wensel R, Orzechowski HD, Schultheiss HP, andHoeffken G (2001) Big endothelin-1 and endothelin-1 plasma levels are correlatedwith the severity of primary pulmonary hypertension. Chest 120:1562–1569.
Salvi SS (1999) �1-Adrenergic hypothesis for pulmonary hypertension. Chest 115:1708–1719.
Simonneau G, Barst RJ, Galie N, Naeije R, Rich S, Bourge RC, Keogh A, Oudiz R,Frost A, Blackburn SD, et al. (2002) Continuous subcutaneous infusion of trepro-stinil, a prostacyclin analogue, in patients with pulmonary arterial hypertension:a double-blind, randomized, placebo-controlled trial. Am J Respir Crit Care Med165:800–804.
Simonneau G, Torbicki A, Hoeper MM, Delcroix M, Karlocai K, Galie N, Degano B,Bonderman D, Kurzyna M, Efficace M, et al. (2012) Selexipag, an oral, selective IPreceptor agonist for the treatment of pulmonary arterial hypertension. Eur RespirJ http://dx.doi.org/10.1183/09031936.00137511.
Southall MD and Vasko MR (2001) Prostaglandin receptor subtypes, EP3C and EP4,mediate the prostaglandin E2-induced cAMP production and sensitization of sen-sory neurons. J Biol Chem 276:16083–16091.
Tapson VF, Gomberg-Maitland M, McLaughlin VV, Benza RL, Widlitz AC, Krich-man A, and Barst RJ (2006) Safety and efficacy of IV treprostinil for pulmonaryarterial hypertension: a prospective, multicenter, open-label, 12-week trial. Chest129:683–688.
Tuder RM, Cool CD, Geraci MW, Wang J, Abman SH, Wright L, Badesch D, andVoelkel NF (1999) Prostacyclin synthase expression is decreased in lungs frompatients with severe pulmonary hypertension. Am J Respir Crit Care Med 159:1925–1932.
Tuder RM, Marecki JC, Richter A, Fijalkowska I, and Flores S (2007) Pathology ofpulmonary hypertension. Clin Chest Med 28:23–42, vii.
Velez-Roa S, Ciarka A, Najem B, Vachiery JL, Naeije R, and van de Borne P (2004)Increased sympathetic nerve activity in pulmonary artery hypertension. Circula-tion 110:1308–1312.
Walch L, Labat C, Gascard JP, de Montpreville V, Brink C, and Norel X (1999)Prostanoid receptors involved in the relaxation of human pulmonary vessels. Br JPharmacol 126:859–866.
White KE, Ding Q, Moore BB, Peters-Golden M, Ware LB, Matthay MA, and OlmanMA (2008) Prostaglandin E2 mediates IL-1�-related fibroblast mitogenic effects inacute lung injury through differential utilization of prostanoid receptors. J Immu-nol 180:637–646.
Whittle BJ, Silverstein AM, Mottola DM, and Clapp LH (2012) Binding and activityof the prostacyclin receptor (IP) agonists, treprostinil and iloprost, at humanprostanoid receptors: treprostinil is a potent DP1 and EP2 agonist. Biochem Phar-macol 84:68–75.
Yang J, Li X, Al-Lamki RS, Southwood M, Zhao J, Lever AM, Grimminger F,Schermuly RT, and Morrell NW (2010) Smad-dependent and Smad-independentinduction of id1 by prostacyclin analogues inhibits proliferation of pulmonaryartery smooth muscle cells in vitro and in vivo. Circ Res 107:252–262.
Yang ZH, Richard V, von Segesser L, Bauer E, Stulz P, Turina M, and Luscher TF(1990) Threshold concentrations of endothelin-1 potentiate contractions to norepi-nephrine and serotonin in human arteries. A new mechanism of vasospasm?Circulation 82:188–195.
Address correspondence to: Dr. Keith Morrison, Drug Discovery Depart-ment, Actelion Pharmaceuticals Ltd., Gewerbestrasse 16, Allschwil CH4123,Switzerland. E-mail: [email protected]
Selexipag and Rat Pulmonary Artery 555
at ASPE
T Journals on Septem
ber 22, 2018jpet.aspetjournals.org
Dow
nloaded from





























