Embed Size (px)
Citation preview

Cell, Vol. 7, 59-65, January 1976, Copyright @ 1976 by MIT
Different Lifetimes of Reticulocyte Messenger RNA
Harvey F. Lodish and Barbara Small Department of Biology Massachusetts Institute of Technology Cambridge, Massachusetts 02139
Summary
About 85-90% of cytoplasmic protein synthesized by young retlculocytes is globln, and about 10% Is a polypeptide (I) of molecular weight 64,000 dal- tons. Maturation of retlculocytes Is accompanied by selective reduction in the synthesis of polypep- tide I, relative to globin; mature reticulocytes syn- thesize globin at a high level but make no detect- able polypeptide I. Studies In which RNA from young and old reticulocytes was translated in a wheat germ ceil-free extract showed that reduction in synthesis of polypeptlde I was correlated with a re- duction in the amount of translatable mRNA for this protein. Differential destruction of mRNA thus is an important factor in determining the types of proteins made during the final stages of erythropoiesis.
Introduction
The concentration of an mRNA species in a cell is a function both of its rate of synthesis and of its rate of destruction. Much is known about the mech- anism of biosynthesis of mRNAs in eucaryotic cells (Darnell, Jelinek, and Molloy, 1973; Brawerman, 1974) and, in particular, about the rates of synthesis of specific RNAs such as globin mRNA and ovalbu- min mRNA during differentiation (Hunt, 1974a; Pal- miter, 1973, 1975). Our understanding of mRNA de- struction is much less complete. Most studies on mRNA stability use cultured human or mouse cells. The decay of poly(A)-containing mRNA in HeLa cells is complex, but can be approximated by as- suming a (stochastic) first-order decay and by assuming that 33% of the total cellular mRNA has a half-life of 1 hr, while the remainder has a half-life of 24 hr (Singer and Penman, 1973). In L cells grow- ing at different temperatures with different doubling rates, the average mRNA half-life was found to be similar to the cell doubling time (Perry and Kelley, 1973). More recent evidence indicates that much of the rapidly labeled polyadenylated mRNA in HeLa cells has a half-life of about l-2 hr (Puckett, Chambers, and Darnell, 1975). The half-life of his- tone mRNA is about 6 hr-less than that of most poly(A)-containing mRNA molecules (Perry and Kelley, 1973).
Half-lives of 1 day or greater have been reported for globin mRNA in rabbit or mouse reticulocytes (Schulman, 1968; Hunt, 1974b) and for ovalbumin mRNA in the hen oviduct (Palmiter, 1973, 1975).
Many studies on mRNA half-life use actinomycin D or other antibiotics to block synthesis of mRNA, and synthesis of a specific protein is followed. For the most part, these are impossible to evaluate, since one is not certain whether the drug inhibits synthe- sis of the mRNA in question, nor whether the drug inhibits protein synthesis by means other than blocking mRNA biogenesis. To cite one example of such side effects, in hepatoma cell cultures, actino- mycin D appears to stabilize either the enzyme tyro- sine amino-transferase (TAT) or its mRNA; this makes it difficult to study the effects of glucocorti- coid hormones on the stability of TAT mRNA (Tom- kins et al., 1972; Kenney et al., 1973; Steinberg, Levinson, and Tomkins, 1975). Studies in cultured HeLa or L cells also indicate that actinomycin D cannot be used to measure mRNA stability by moni- toring the rate of protein synthesis (Singer and Pen- man, 1972; Greenberg, 1972).
Rabbit reticulocytes are an ideal system for study- ing mRNA turnover, since these cells contain no nucleus and synthesize no RNA. About 90% of the protein synthesized in these cells is (Y and /3 globin, but a number of discrete non-globin proteins, in- cluding two membrane proteins, are also made (Lo- dish and Desalu, 1973; Lodish and Small, 1975). In this report we show that the mRNA for one non- globin protein has a much shorter lifetime than does globin mRNA. Differential destruction of mRNA thus is an important factor in determining the types of proteins made during the terminal stage of erythropoiesis, the maturation of reticulocytes to erythrocytes.
Results
Protein Synthesis in Maturing Reticulocytes Polyacrylamide gel electrophoresis in the presence of 6 M urea and 0.1% sodium dodecyl sulfate re- solves the cytoplasmic proteins synthesized by young rabbit reticulocytes into two predominant species (Figures 1A and 1 D; see also Lodish and Desalu, 1973). About 90% of 3XS-methionine radio- activity incorporated into cytoplasmic proteins co- migrates with globin; analysis of tryptic digests of this species confirmed that it contained equal amounts of labeled (Y and ,!3 chains (data not shown). About 12% of the incorporated radioactivity is in a single species-peptide l-of apparent molec- ular weight 64,000 daltons (Table 1). Essentially the same pattern was obtained from reticulocyte-rich blood taken from rabbits made anemic by phenylhy- drazine injection (Figure 1A) or by daily bleeding (Figure 1 D). In some cases, other minor non-globin cytoplasmic peptides are also made. These reticulo- cytes also make two membrane proteins (Lodish, 1973; Lodish and Small, 1975), but only the cyto-

Cell 60
A. Phenylhydrazine - Anemic Cells 96% Reticulacytes
Glabin
I
Day I T
El. Recovery - Doy 4
C. Recovery - Doy 7
n IJ
IO 8 6 4 2 C
D. Phlebotomized - Anemic Cells 84 % Reticulocytes
Glo,bin Day I
E. Recovery - Doy 4
AF. Recovez
I I IO 8 6 4 2 (top
‘rn
Figure 1. Synthesis of Globin and Non-Globin Proteins by Reticulocytes of Various Ages
Two rabbits were used, one made anemic by injection with phenylhydrazine (A-C) and one made anemic by bleeding (D-F) (see Experimental Procedures). For both anemic rabbits, day 1 is the day of maximum reticulocytosis (see Experimental Procedures). The animals were allowed to recover, with no further injections or massive bleedings. At 4 days (6 and E) and 7 days (C and F), about 5 ml of blood were withdrawn. Cells were labeled in vitro with Wj-methionine as described in Experimental Procedures; the rates of protein synthesis by the different cell populations are given in Table 1. An amount of cell supernatant containing 0.5 mg hemoglobin was analyzed by electrophoresis through cylindrical polyacrylamide gels; shown are scans of the radioautograms of the dried gels. Exposure of the X-ray film was for 6 days.
Table 1. Synthesis of Globin and Non-Globin Proteins by Reticulocytes of Different Ages
Total Protein Synthesis cpm W+Methionine/hr/mg Hemoglobin
Phenylhydrazine- Day 1 99,000 Anemic Rabbit
Ratio Polypeptide I/Globin
0.122
Phlebotomized Anemic Rabbit
Day 4 61,000 0.060
Day 7 23,000 0.00
Day 1 76.000 0.116
Day 4 62,000 0.035
Day 7 41,000 0.001
The cell preparations used are described in the legend to Figure 1. Shown is the total amount of XSS-methionine incorporated per hour by these cell populations and the ratio of synthesis of polypeptide I to that of globin. The latter data were calculated from the areas under the respective peaks in Figure 1.

Different Lifetimes of Reticulocyte Messenger RNA 61
plasmic fraction of reticulocytes was used in these studies.
Figure 1 also shows the results of experiments in which anemic rabbits were allowed to recover. By day 4, about half the peripheral blood stained with new methylene blue as reticulocytes, and the amount of radioactive methionine incorporated per cell was 60-80% that of reticulocyte-rich blood on day 1 (Table 1). Table 1 and Figures 1 B and 1E show the important result that the synthesis of polypeptide I, relative to that of globin, was marked- ly reduced. By day 7, the remaining reticulocytes still synthesize a significant amount of globin (Table l), but make no detectable amount of polypeptide I.
The ratio of synthesis of polypeptide I to that of globin was independent of the time of incorporation of 35S-methionine; in particular, cells at day 6 syn- thesized no detectable polypeptide I, even in label- ing periods as short as 5 min (data not shown). These results tend to rule out selective turnover of polypeptide I as a cause of the different profiles ob- tained using reticulocytes of different ages.
It is assumed in these studies that the more active reticulocytes found in the circulation on day 1, whether induced by phenylhydrazine or by bleed- ing, mature into the less active reticulocytes charac- teristic of day 4, although direct proof of this would be difficult to obtain. Since the same proteins are made irrespective of the method of induction of anemia, it seems improbable that the cells on day 1 represent an abnormal type of reticulocyte.
As shown in Figure 2, cell-free extracts from blood cells of an anemic rabbit (day 1) synthesize appreciable amounts of polypeptide I as well as of globin. Extracts from older cells (day 6) synthesize 80% as much globin as do young cells (day l), but make little detectable polypeptide I. These results support the notion that synthesis of polypeptide I declines much faster than that of globin during mat- uration of reticulocytes to erythrocytes.
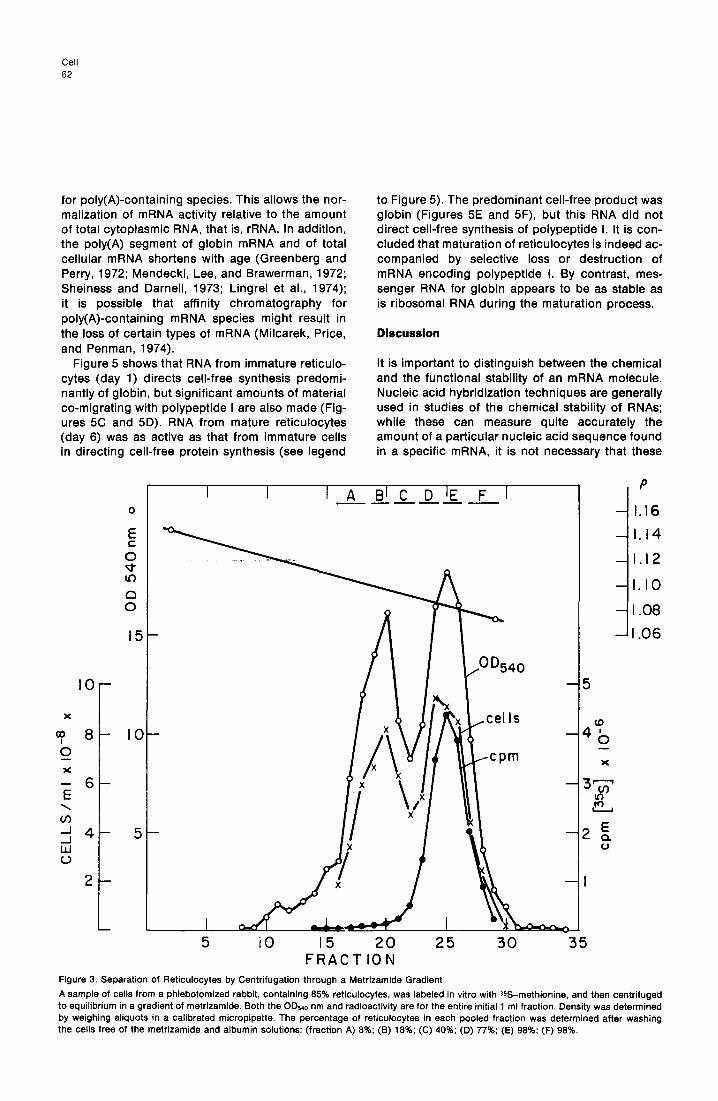
Fractlonatlon of Reticulocytes of Metrizamlde Gradient Centrifugation Further correlation between reticulocyte maturation and the selective reduction in synthesis of polypep- tide I requires fractionation of a population of retic- ulocytes according to age. To this end, we centri- fuged a sample of blood cells from a phlebotomized rabbit (85% reticulocytes) through a linear gradient of metrizamide (Lodish, Small, and Chang, 1975). The cells had been previously labeled in vitro with 35S-methionine, since younger reticulocytes are more active in protein synthesis than older ones. Figure 3 shows that younger cells (high ratio of cpm per cell) band at a lighter density than do more ma- ture reticulocytes, as is the case as well in gradients of bovine serum albumin (Glowacki and Millett,
- Globin
ABCD Figure 2. Analysis of Protein Synthesis by Reticulocytes and Their Cell-Free Extracts by Slab Gel Electrophoresis
Exposure of the dried gel to film was for 7 days. (A) Protein synthesis by phenylhydrazine-anemic cells (day 1); 8000 cpm (0.1 mg hemoglobin) of cell supernatant were used. (B) Protein synthesis by cells from a phenylhydrazine-anemic rabbit which had been allowed to recover for 6 days: 3600 cpm (0.1 mg hemoglobin) of cell supernatant were used. (C) Protein synthesis by cell-free extracts of cells used in (A); pro- teins synthesized by 2.4 ~1 of cell-free reaction (13,100 cpm of WS-methionine) were analyzed. (D) Protein synthesis by cell-free extracts of the cell population used in (B); a 2.4 ~1 aliquot containing 11,600 cpm XS-methionine- labeled proteins was used.
1965). The ratio (cpm per cell) increases monotoni- cally across the gradient.
Figure 4 shows that the younger reticulocytes (fraction E) synthesize twice as much polypeptide I relative to globin as do the more mature cells in fraction B. We conclude that maturation of reticulo- cytes is indeed correlated with selective loss in the capacity to synthesize polypeptide I.
Messenger RNA for Globin and for Polypeptlde I To quantitate the amount of mRNAs for these polypeptides, we studied the translation of RNA from reticulocytes of various ages in cell-free ex- tracts of wheat germ. We used total cytoplasmic RNA, not fractionated by affinity chromatography
Cell 62
for poly(A)-containing species. This allows the nor- malization of mRNA activity relative to the amount of total cytoplasmic RNA, that is, rRNA. In addition, the poly(A) segment of globin mRNA and of total cellular mRNA shortens with age (Greenberg and Perry, 1972; Mendecki, Lee, and Brawerman, 1972; Sheiness and Darnell, 1973; Lingrel et al., 1974); it is possible that affinity chromatography for poly(A)-containing mRNA species might result in the loss of certain types of mRNA (Milcarek, Price, and Penman, 1974).
Figure 5 shows that RNA from immature reticulo- cytes (day 1) directs cell-free synthesis predomi- nantly of globin, but significant amounts of material co-migrating with polypeptide I are also made (Fig- ures 5C and 5D). RNA from mature reticulocytes (day 6) was as active as that from immature cells in directing cell-free protein synthesis (see legend
to Figure 5). The predominant cell-free product was globin (Figures 5E and 5F), but this RNA did not direct cell-free synthesis of polypeptide I. It is con- cluded that maturation of reticulocytes is indeed ac- companied by selective loss or destruction of mRNA encoding polypeptide I. By contrast, mes- senger RNA for globin appears to be as stable as is ribosomal RNA during the maturation process.
Discussion
It is important to distinguish between the chemical and the functional stability of an mRNA molecule. Nucleic acid hybridization techniques are generally used in studies of the chemical stability of RNAs; while these can measure quite accurately the amount of a particular nucleic acid sequence found in a specific mRNA, it is not necessary that these
I 1 A Ed C D ‘E F 1 y--m--
5 i0 15 20 25 30 FRACT ION
P I. 16
I. I4
I.12
I. IO
I .08
I .06
Figure 3. Separation of Reticulocytes by Centrifugation through a Metrizamide Gradient
A sample of cells from a phlebotomized rabbit, containing 85% reticulocytes, was labeled in vitro with SLS-methionine, and then centrifuged to equilibrium in a gradient of metrizamide. Both the OD 540 nm and radioactivity are for the entire initial 1 ml fraction. Density was determined by weighing aliquots in a calibrated micropipette. The percentage of reticulocytes in each pooled fraction was determined after washing the cells free of the metrizamide and albumin solutions: (fraction A) 8%: (B) 18%; (C) 40%; (D) 77%: (E) 98%: (F) 98%.

Different Lifetimes of Reticulocyte Messenger RNA 63
be in functional mRNA molecules. A case in point is a recent study of E. coli trp mRNA synthesized either under the control of the normal 0-p operon promotor or under the control of a phage lambda promotor (Yamamoto and Imamoto, 1975). When formation of new mRNA is inhibited, the capacity to serve as template for trp enzyme synthesis de- clines at a comparable rate for the two types of trp mRNAs, and the two mRNAs are translated with equal efficiency. In contrast to this similar functional stability, trp mRNA synthesized under control of the lambda promotor shows more than a 10 fold greater chemical stability than does the normal trp mRNA.
Most studies on stability of mRNA in cultured mammalian cells use the amount of cytoplasmic polyadenylated RNA as the measure of mRNA. It is not clear, however, whether all this material is in
G’“lbin 9
IO 8 6 4 2 (top cm
Figure 4. Polyacrylamide Gel Electrophoresis of Cytoplasmic Extracts of Reticulocytes Fractionated on a Metrizamide Gradient
The letters B-E refer to 4 samples taken from the gradient shown in Figure 3. Shown are scans of radioautograms of the dried gels, each of which was used to analyze an amount of cell supernatant containing about 10,000 cpm of %S-methionine-labeled protein and from 0.05 to 0.25 mg hemoglobin. Exposure of the X-ray film was 6 days. Since equal amounts of radioactivity were added to the gels, the areas under the peaks corresponding to globin and polypeptide I do not represent the absolute rate of protein synthesis by each gradient fraction (compare Figure 3) but can be used to calculate the relative synthesis of the two proteins.
fact functional mRNA, particularly when older poly(A)-containing RNAs are under study. Only in cases where one can study synthesis of a specific protein, such as globin (Schulman, 1968; Hunt, 1974b), ovalbumin (Palmiter, 1973, 1975) or silk fi- broin (Suzuki and Suzuki, 1974) is it possible to obtain reliable information concerning the dynam- ics of mRNA synthesis and destruction. Ideally, both a functional assay of mRNA-ability to direct cell- free synthesis of the specific protein-and a chemi- cal-or hybridization-assay should be used.
In the present study, mRNAs for three reticulocyte proteins-the (Y and /3 chains of hemoglobin and a non-globin protein (polypeptide I) of molecular weight 64,000 daltons-were assayed by means of
- 6 an d I
- G II0 bin
ABCDEFGH Figure 5. Synthesis of Reticulocyte Proteins in Cell-Free Extracts of Wheat Germ
Reactions of 50 gl are described in Experimental Procedures. 20 pl of each reaction were analyzed by electrophoresis through a slab gel of polyacrylamide. Given below are the amounts of reticulo- cyte RNA used and, in brackets, the incorporation into protein of 3%-methionine by a 3 ~1 aliquot. Exposure of the gel to film was for 6 days, The same animal was used for RNA isolation (C-F) and for whole cell labeling (G and H). (A) No added RNA (7200 cpm). (6) No added RNA (9200 cpm). (C) 0.16 A260 units RNA from phenylhydrazine-anemic cells, day 1 (72,000 cpm). (D) 0.32 Asao units of RNA used in (C). (E) 0.15 AsbO units of RNA from phenylhydrazine-anemic cells which had been allowed to recover for 6 days (52,000 cpm). (F) 0.30 AZ60 units of RNA used in (E) (68,000 cpm). (G) Supernatant from phenylhydrazine-anemic cells (day 1) which had synthesized a%-methionine-labeled proteins in vitro. (H) As in (G), except that cells from the animal which had been allowed to recover for 6 days were used.

Cell 64
cell-free protein synthesis. It was shown that during maturation of reticulocytes, the amount of translat- able mRNA for polypeptide I declines at a much faster rate than does mRNA for globin (Figure 5) and that this is reflected in decreased synthesis of polypeptide I relative to that of globin during matu- ration of reticulocytes (Figures 1, 2, and 4).
For several reasons, our data by themselves can- not be used to calculate the absolute half-lives of the mRNAs for globin and for polypeptide I. Normal- ly, reticulocytes are confined to the marrow and ma- ture into erythrocytes within 1 day. During severe anemia, reticulocytes are prematurely released into the peripheral blood and may comprise as much as 90% of the circulating red blood cells. We do not know with precision the length of the reticulo- cyte period during erythropoiesis, nor do we know the rate of maturation of reticulocytes to erythro- cytes. Moreover, in the studies in which successive- ly older populations of reticulocytes were obtained from animals recovering from anemia (Figure 1) we do not know the rate at which the maturing reticulo- cytes in the blood were being replaced by new retic- ulocytes from the marrow. Nonetheless, it is rea- sonable to assume a half-life of globin mRNA of about 24 hr, as has been measured in several recent studies on erythropoiesis (Schulman, 1968; Hunt, 1974b). In this case, the data in Figures 1 and 4 would indicate that the half-life of the mRNA for polypeptide I would be about 5 to 8 hr.
Maturation of reticulocytes is accompanied by loss of ribosomes. More mature reticulocytes con- tain fewer ribosomes than young cells, and a smaller fraction of the ribosomes are found on poly- somes and are active in protein synthesis (Danon, Zechavi-Willner, and Berman, 1965; Glowacki and Millett, 1965; Marks, Rifkind, and Danon, 1963). Maturation is also accompanied by loss of one or more protein factors required for initiation of pro- tein synthesis (Herzberg, Revel, and Danon, 1969). Reticulocytes also contain one or two membrane proteins not found in erythrocyte membranes and which are presumably lost at some stage in matura- tion (Koch, Gardner, and Carter, 1973; Lodish and Small, 1975).
It should be noted that not only does differential turnover of reticulocyte mRNA influence the protein composition of mature erythrocytes, but that dif- ferential turnover of reticulocyte proteins is also im- portant. Subunits of reticulocyte supernatant pro- teins which are preferentially degraded during maturation are, on the average, larger than those which are stable, and are more sensitive to in vitro de- struction by proteases (Lodish et al., 1975). These correlations between turnover rate, size of polypep- tide subunits, and susceptibility to proteases have been noticed in a variety of other systems as well
(Goldberg and Dice, 1974; Schimke, 1973; Gold- berg et al., 1974). In HeLa cells, the more stable mRNA population has a slightly smaller average size than do mRNAs which turn over more rapidly (Singer and Penman, 1973). Our results on stability of reticulocyte mRNAs are consistent with this cor- relation between mRNA size and relative rate of mRNA destruction, but it is clear that a much larger number of specific mRNAs in a variety of systems must be studied to test this generalization.
Experimental Procedures
Phenylhydrazine-Anemic Rabbits New Zealand white rabbits weighing 4 to 6 lb were given subcuta- neous injections for 5 days of 2 ml 1.2% acetylphenylhydrazine. On
day 7, the animal was bled from the ear. As determined by staining with methylene blue, over 90% of the blood cells were reticulocytes.
Phlebotomlzed Anemic Rabbits On day I,2 cc acetylphenylhydrazine were injected subcutaneous- ly, and 40 ml of blood were removed from an ear. On each day following, 40-50 ml of blood were taken from one ear. On days 3 and 6, the rabbit was also given an intramuscular injection of 1 cc iron-dextran (50 mg Fe/ml, Imferon, Lakeside Laboratories, Milwaukee, Wisconsin). By day 10, generally over 80% of the blood cells were reticulocytes.
Preparation of Cell Supernatants Blood was collected in 50 ml solution A (0.14 M NaCI, 0.0015 M magnesium acetate, 0.005 M KCI) containing 0.001% heparin. The cells were collected by centrifugation at 4000 rpm for 5 min and washed by centrifugation 3 times in solution A. The huffy coat con- taining white cells was removed after the first 2 centrifugations.
In labeling experiments, the cells were incubated in vitro with X-methionine for 60 min at 37°C (Lodish and Oesalu, 1973). Ali- quots of 5 pl taken at 20 min intervals were used to determine the amount of radioactivity incorporated into protein. Hemoglobin was quantitated by absorbance at 540 nm of a dilution of the cells in Drabkin’s solution (Wintrobe, 1974).
The cells were recovered by centrifugation, washed once in solu- tion A, lysed in 5.6 vol of 5P8 [O.OOS M sodium phosphate (pH 8) Fairbanks, Steck, and Wallach, (1971)] and centrifuged at 15,000 rpm for 20 min. The supernatants were stored frozen until used. Extracts from unlabeled cells were prepared by lysing the packed cells in an equal volume of water. Following centrifugation at 15,000 rpm for 20 min, the supernatants were stored frozen.
Metrlzamlde Gradlents An aqueous metrizamide solution (Gallard-Schlesinger Corp., Carle Place, New York) of 34.77% (w/v) is isotonic. A 14.6 ml linear gradient was made in a tube for the Beckman SW27.1 rotor. The mixing chamber (27% metrizamide) contained 5.67 ml of the isoton- ic metrizamide solution and 1.83 ml of solution B [0.118 M NaCI, 0.005 M KCI, 0.0167 M Na2HP0,, 0.00063 M CaCb, 0.00179 M MgSO,, 4% bovine serum albumin, (pH 7.5)]; the reservoir (12% metrizamide) contained 2.52 ml isotonic metrizamide solution and 4.78 ml solution B. Washed cells from a phlebotomized anemic rabbit were labeled in vitro with ‘5S-methionine, washed, and re- suspended in 5 vol of solution 8; 3 cc of this were layered atop this gradient. Centrifugation was at 15,000 rpm at 4°C for 1 hr in a Beckman ultracentrifuge. The gradients were collected from the bottom by a peristaltic pump.
Isolation of Messenger RNA and Cell-Free Protein Synthesls Total RNA was isolated from the cytoplasm of reticulocytes using procedure 2 of Nienhuis, Fulvey, and Anderson (1974). Extracts

Different Lifetimes of Reticulocyte Messenger RNA 65
from wheat germ were prepared by the procedure of Roberts and Patterson (1973). Reactions (50 ~1) contained 15 pl wheat germ extract, 1.3 mM ATP, 0.26 mM GTP, 15 mM creatine phosphate, 520 pg creatine phosphokinase, 15 mM HEPES (pH 7.0) 5 mM dithiothreitol, 62 mM KCI, 0.65 mM spermidine, 0.13 mM each of 19 unlabeled amino acids, and 10 pCi ass-methionine (200,000 mCi/mmole; New England Nuclear Corp.). Incubation was at 25°C for 2 hr. Proteins were precipitated by addition of 1 cc acetone. The precipitate was dried and dissolved in Laemmli’s sample buffer (Laemmli, 1970). Conditions for reticulocyte cell-free protein syn- thesis were detailed previously (Lodish. 1971; Lodish and Desalu, 1973).
Polyacrylamlde Gel Elactrophoreslo Two types of electrophoresis systems were used. Cell-free reaction products (Figures 2 and 5) were resolved by electrophoresis through a 13% polyacrylamide slab gel by the procedure of Laemmli (1970).
Supernatants of labeled reticulocyte cells (Figures 1 and 4) were added to the urea-SDS-phosphate gel solution used previously (Lodish and Desalu, 1973) and placed in a boiling water bath for 2 min. They were analyzed by electrophoresis through cylindrical 7.5% polyacrylamide gels containing 6 M urea and 0.1% sodium dodecyl sulfate, as detailed previously (Lodish and Desalu, 1973). Staining of the gels with Coomassie Blue and destaining followed the procedure of Fairbanks et al. (1971).
Gels were dried and subjected to radioautography against Kodak RP/RB54 X-ray films. Films were scanned with a Joyce-Loebl mi- crodensitometer, where full pen deflection was equivalent to 1.49 OD units.
Acknowledgments
We thank Mr. Tom Alton for providing the wheat germ extracts and for advice on the conditions for cell-free protein synthesis. This work was supported by grants from the NIH. H. F. L. is recipient of Research Career Development Award from the NIH.
Received August 28, 1975; revised September 29, 1975
References
Brawerman, G. (1974). Ann. Rev. Biochem. 43, 621-642.
Danon, D., Zechavi-Willner. T.. and Berman, G. (1965). Proc. Nat. Acad. Sci. USA 54, 873-879.
Darnell. J. E., Jelinek. W. R.. and Molloy, G. R. (1973). Science 181, 1215-1218.
Fairbanks, G., Steck, T. L., and Wallach, D. F. H. (1971). Biochem. 7 0, 2606-2618.
Glowacki. E., and Millett, R. (1965). J. Mol. Biol. 17, 116-127.
Goldberg, A. I., and Dice, J. F. (1974). Ann, Rev. Biochem. 43, 835-869.
Goldberg, A. I., Howell, E. M., Martel, S. B., Li, J. B., and Prouty, W. F. (1974). Fed. Proc. 33, 1112-l 120.
Greenberg, J. (1972). Nature 240, 102-I 04.
Greenberg, J., and Perry, R. (1972). Biochim. Biophys. Acta 287, 361-366.
Herzberg, M., Revel, M.. and Danon, D. (1969). Eur. J. Biochem. 77, 148-153.
Hunt, J. A. (1974a). Biochem. J. 738, 499-510.
Hunt, J. A. (1974b). Biochem. J. 738, 487-498.
Kenney, F. T.. Lee, K., Stiles, D., and Fritz, J. (1973). Nature New Biol. 246, 208-210.
Koch, P., Gardner, F., and Carter, J. (1973). Biochem. Biophys. Res. Commun. 54, 1296-1299.
Laemmli, 0. (1970). Nature 227, 680-684.
Lingrel, J. B., Morrison, M., Gorski. J., and Merkel, C. (1974). Ann. NY Acad. Sci. 247, 156-169.
Lodish, H. F. (1971). J. Biol. Chem. 246, 7131-7138.
Lodish, H. F. (1973). Proc. Nat. Acad. Sci. USA 70, 1526-1530.
Lodish, H. F., and Desalu, 0. D. (1973). J. Biol. Chem. 248, 3520- 3527.
Lodish, H. F., and Small, B. (1975). J. Cell Biol. 65, 51-64.
Lodish, H. F., Small, B., and Chang, H. (1975). Dev. Biol., in press.
Marks, P., Rifkind, R., and Danon, D. (1963). Proc. Nat. Acad. Sci. USA 50. 336-342.
Mendecki, J., Lee, S-Y., and Brawerman, G. (1972). Biochem. 77, 793-798.
Milcarek, C., Price, R., and Penman, S. (1974). Cell 3, l-10.
Nienhuis, A., Fulvey. A. K., and Anderson, W. F. (1974). Methods Enzymol. 30, 621-631.
Palmiter. R. Cl. (1973). J. Biol. Chem. 248, 8260-8270.
Palmiter, R. D. (1975). Cell 4, 189-197.
Perry, R. P., and Kelley, D. E. (1973). J. Mol. Biol. 79. 681-696.
Puckett, L., Chambers, S., and Darnell, J. E. (1975). Proc. Nat. Acad. Sci. USA 72, 389-393.
Roberts, B., and Paterson, B. (1973). Proc. Nat. Acad. Sci. USA 70, 2330-2334.
Schimke, R. T. (1973). Advan. Enzymol. 37, 135-187.
Schulman, H. M. (1968). Biochim. Biophys. Acta 155, 253-261,
Sheiness. D., and Darnell. J. E. (1973). Nature New Biol. 241, 265-268.
Singer, R., and Penman, S. (1972). Nature 240, 100-102.
Singer, R. H., and Penman, S. (1973). J. Mol. Biol. 78, 321-334.
Steinberg, R. A., Levinson, B. B., and Tomkins, G. M. (1975). Cell 5, 29-35.
Suzuki, Y., and Suzuki, E. (1974). J. Mol. Biol. 88, 393-407.
Tomkins, G., Levinson, B., Baxter, J., and Dethlefson, L. (1972). Nature New Biol. 239, 9-14.
Wintrobe. M. M. (1974). Clinical Hematology (Philadelphia: Lea and Feibiger).
Yamamoto, T., and Imamoto, F. (1975). J. Mol. Biol. 92, 289-309.





























