Embed Size (px)
Citation preview

IntroductionOur knowledge of species ecology is the scientific background for the conservation and management of threatened species (e.g. Brito 2003, Bjorgan 2005, Richardson et al. 2006). However, individuals of marginal and isolated populations of reptiles may have different ecological features than the conspecific individuals from other “mainland” populations (e.g.: Strijbosch et al. 1980, Amat et al. 2003, Tabachishin et al. 2006), therefore, it is sometimes difficult to extend our knowledge to all populations of a certain species. On the other hand, individuals may have different ecological preferences and have dissimilar activity depending on the age (e.g. Sun et al. 2001, Amat et al. 2003, Brito 2003). This is especially true for the selection of preferable habitats (e.g. Lind 1990, Shine et al. 2002, Bjorgan 2005, Nemes et al. 2006). Ontogenetic shifts in habitat selection may reflect various age groups’ features such as predation risk (Keren-Rotem et al. 2006, Werner et al. 1983), thermal reaction habits (Cherlin 1989, Winne and Keck 2005, Blouin-Demers et al. 2007), diet (Lind 1990, Luiselli et al. 1995, Weatherhead et al. 2003), or lokomotion (Lind 1990, Shine et al. 2002, Blouin-Demers et al. 2007).
In southwestern Siberia, the Halys pit viper (Gloydius halys halys (Pallas 1776) inhabits mountains and foothills of the Altai-Sayan mountain system (Orlov and Barabanov 2000, Ananieva et al. 2004, Kuzmin and Semenov 2006, Simonov 2008). So far, the most isolated population of G. h. halys is known from the west Siberia, in the Novosibirsk region (Pestov 2003, Simonov 2007, 2008). This locality represents the northwesternmost record of G. h. halys distribution. The nearest neighbouring populations are at 190-250 km (Simonov 2008). Due to the natural habitat fragmentation, G. h. halys shows a patchy distribution, with a series of local micropopulations along the Berd’ river, where rocky habitats are available.Since 2008, this population of Halys pit viper is included in the Red data book of Novosibirsk region (in press), and the development of a conservation program based on the species ecology is a major goal to improve its protection. Habitat requirements of neonate snakes are poorly known, even for well-studied species (e.g. Blouin-Demers et al. 2007) because of their secretive style of life. Due to their high abundance (up to 267 individuals per hectare), Simonov (2007, 2008) considered G. h. halys to be a good model organism to study snake ecology. The aim of this paper is to evaluate and analyse the differences between adult and neonate G. h. halys in the habitat use preferences, in the daily activity patterns and in the preferred ambient temperatures.
Herpetology Notes, volume 2: 1-7 (2009)
Differences in habitat use, daily activity patterns and preferred ambient temperatures of adult and neonate Gloydius halys halys
from an isolated population in southwest Siberia: preliminary dataEvgeniy Simonov
Department of Zoology, Institute of Natural and Social-Economical Sciences, Novosibirsk State Pedagogical University, Viluyskaya street 28, 630126 Novosibirsk, Russia; e-mail: [email protected]
Abstract. Differences in habitat selection, daily activity and preferred ambient temperatures between adult and neonate Halys pit-vipers, Gloydius halys halys (Pallas, 1776) were studied in an isolated population in Novosibirsk region (Russia, southwest Siberia). Investigations were conducted at the end of August, due to the high abundance of neonates. Simple correspondence analysis showed that adults and neonates have different habitats and daily activity, and prefer different sun expositions. Multiple correspondence analysis showed that adult pit-vipers select rocky habitats, southwest exposition and are most active at midday. Neonates prefer stony steppes, southeast sun exposition and have activity peaks in the morning and evening. Adult snakes select significant higher ambient temperatures than neonates. Different thermal reactions and different diets may be likely the factors driving to the differences in habitat use between neonate and adult of G. h. halys.
Key words. Ecology, Gloydius halys halys, Habitat selection, Ontogenetic shifts, Southwest Siberia.

Simonov2
Materials and Methods
Study areaThe area of investigation is located in the southeastern part of Novosibirsk region, on the territory of Maslyanino district (Rus-sia). The study site is a 100m wide strip along the right bank of the river Berd (54º30´N, 84º03´E), on an elevation between 160 to 219 m a.s.l. At this place, the bedrock foundation are made of numerous rocks, blocks and stony slide-rocks (Fig. 1c). Berd is a permanent lowland river (with the exception of the upper end) 25-30m wide (at the study place), showing spring flood. Average air temperature ranges from -19,1ºC in January to 17,7ºC in July, annual average air temperature is -0,3ºC, (Kravcov and Donuka-lova 1996). The average rainfall is 471 mm/year, and the average monthly rainfall ranges from 14 mm in February to 75 mm in July, while the precipitation-evaporation ratio is 1,3 (Kravcov and Donukalova 1996). Due to the agriculture, the valley of the river is very disturbed, and the native vegetation is preserved only along the river. The study area is a wood-meadow area, where birches (Betula sp.) and aspens (Populus sp.) dominate; small number of Pinus sylvestris and Larix sp. also occur. On the cliffs are often steppe and petrophyte plants associations.
Field methodsObservations were conducted from 23rd to 30th August 2008. This time was chosen due to the following reasons: (1) previously
observations (Simonov 2007, 2008) showed that neonate (and juveniles) snakes occur extremely rare during the May–August period, (2) in this population, females of G. h. halys give birth at the end of August (3) September weather conditions are often non optimal for snake observations, and therefore unusable for data collection. During this short period 25 adult (SVL > 420 mm) and 15 neonate snakes (SVL < 170 mm) were observed.Surveys of G. h. halys were conducted repeatedly, eight times during the day (between 08.00-21.00) on transects with non-limi-ted width in the following types of the habitat: (1) “rocky area”, represented by stony slide-rocks, bulks of large stones and other rocky areas with similar microclimatic and vegetable conditions (projective cover of plants less than 25%; basic plant species are Artemisia frigida Wild., Rubus caesius L., Galium ruthenicum Willd., Chelidonium majus L., Allium nutans L.) (Fig. 1b); (2) “stony steppe”, areas where arid soil contains numerous stones, average projective cover 50-60%, basic plant species are Stipa sp., Orostachys spinosa (L.) C. A. Meyer, Spiraea hypericifolia L., Caragana frutex (L.) C. Koch, Artemisia frigida Wild., Artemisia sp. (Fig. 1a); (3) “bottomland”, flood-plain of Berd’ river, where projective cover of miscellaneous herbs is close to 100%. Single trees as birches (Betula pendula Roth), aspens (Populus tremula L.) and willows (Salix rosmarinifolia L.) occour. In contrast to previous habitats, it is a flat area.Estimation of snakes’ activity via visual counts was used, although this method has obvious lacks, being influenced by weather con-
Figure 1. Main habitats of G. h. halys in Novosibirsk region: A – portion of stony steppe; B – portion of stony slide-rocks (the one variant of “rocky area”); C – view on study site from far bank (stony steppes mainly on a right side of hill, rocky areas mainly on a left side).

Habitat differences in adult and neonate of G. h. halys 3
ditions, by the snake size, and by the plant coverage (Sun et al. 2001). The following variables were recorded for each individu-al: time of the day (morning - 09 to 11 a.m., midday – 11 a.m. to 04 p.m., afternoon – 04 p.m. to 08 p.m.), habitat type (rocky area, stony steppe, bottomland; see above for explanation), expositi-on (southwest (SW), southeast (SE) or flat area), air temperature (Ta), air temperature at 5 cm from ground (Ta2), temperature of substrate (Ts). Temperatures of air and substrate were measured using a digital thermometer (accuracy 0.1ºC).
Statistical proceduresSimple and multiple correspondence analysis (Greenacre 1984) was used to analyse associations between groups (adults and neonates) and selected variables (habitat, exposition and daily activity). All temperature variables (Ta, Ta2, Ts) did not deviate from normality in both groups (Kolmogorov-Smirnov test for normality: p > 0.2), but homogeneity of variances between diffe-rent groups was significant only for Ta and Ts (Levene’s test: p > 0.1), Ta2 was not significant (Levene’s test: p < 0.01). Thus, to in-
vestigate the differences between groups, for Ta and Ts the T-test was applied, while the nonparametric Mann-Whitney U Test was applied for Ta2. The statistical processing of data was conducted with Statistica 6.0.
Figure 2. Associations of adult and neonate of G. h. halys in the space as described by the first two correspondent axes of multiple correspondence analysis.
2 df P
Age * Habitat 28.25 2 < 0.001
Age * Exposition 18.06 2 < 0.001
Age * Part of the day 7.53 2 < 0.05
Table 1. Summary of simple correspondence analysis.
Adults Neonates Habitat type
Stony steppe Rocky area Bottomland
Exposition South-eastSouth-westFlat area
Part of the day Morning Midday Afternoon
1213
6163
01312
1320
1410
456
Table 2. Observed frequencies of character states analysed in multiple correspondence analyses.

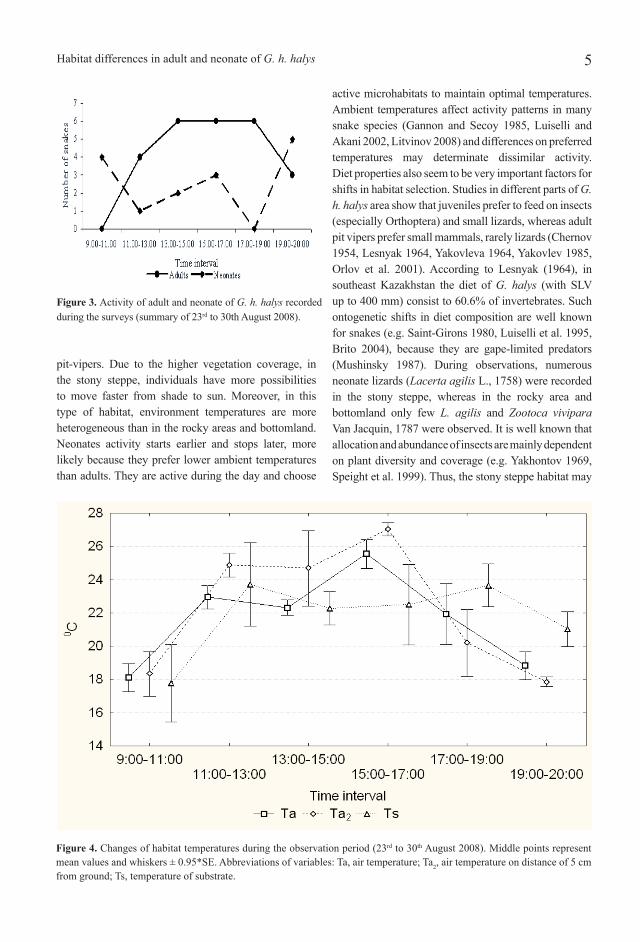
ResultsResults of simple correspondence analyses (Table 1) show significant associations between age and habitat type, exposition, and daily activity. Multiple correspondence analysis shows very close associations of neonates and adult snakes with habitat and exposition, while associations for the time of the day are not so strong, but also relevant (Fig. 2, Table 2). Thus, in this season, neonates preferred stony steppes, slopes of southeast sun exposition and were active during the whole day. In contrast, adult snakes preferred rocky areas, southwest sun exposition and were active mostly at midday and during the afternoon. In the bottomland, only adult pit-vipers were recorded. Figure 3 displays differences in daily activity among selected age groups in two-hour intervals. Neonates of G. h. halys had two peaks of activity at 09.00-11.00 and 19.00-21.00. Adult snakes exhibit in front of their hidings after 11.00. Their activity reaches a plateau between 13.00-19.00. Around 21.00 adult of G. h. halys return to their night shelters.Changes of environmental temperatures during observations are presented in Fig. 4. During snake activity Ta ranges from 16.5 to 28.5ºC, Ta2 from 16.6 to 27.8 ºC and Ts from 14.0 to 25.0 ºC. During observation period, adult snakes significantly prefered higher Ta, Ta2 and Ts than neonates (see Tab. 3).
DiscussionResults of this study clearly show ontogenetic differences in all examined variables. Habitat preferences and daily activity of G. h. halys have been studied in different populations and several subspecies (Chernov 1954, Lesnyak 1964, Yakovleva 1964, Ataev 1985, Yakovlev 1985, Borkin et al. 1990, Prokopov et al. 2000, Orlov et al. 2001, Kolobaev 2006), but these investigations
were concentrated mainly on adults and did not take in consideration neonate ecological features. The same differences in the habitat use between adult and neonate of Halys pit vipers were recorded from the Zeya sanctuary (the northeasternmost point of G. halys distributional range) (N.N. Kolobaev, unpublished data), where neonates and juveniles prefer more shaded and humid habitats. In previous studies (Simonov 2007, 2008) it has been shown that adult individuals of the G. h. halys population in Novosibirsk region display ecological differences from populations of the “mainland” area. Main differencies are the following: they are stenotopic and have an higher density, which determines the specific population structure; they are more active during the day, whereas G. halys from the central part of its range are more active in the dusk and at night (in summer). Differences of preferable ambient temperatures may determine the habitat split among neonate and adult pit-vipers. As ectotherms, reptiles manipulate habitat choice and time of activity in an attempt to achieve body temperatures in preferred ranges (Heath 1965, Huey, Pianka and Hoffman 1977, Meek and Jayes 1982, Christian et al. 1983, Litvinov 2008), whereas body size (and accordingly age) influences thermoregulatory requirements of ectothermic animals because smaller individuals warm and cool more quickly than larger ones (Schmidt-Nielsen 1979, Stevenson 1985). In fact, differences of preferable habitat and body temperatures among age groups are common in reptiles, and young individuals usually choose lesser temperatures than adults (e.g. Carretero, Marcos and de Prado 2006, Hitchcock and McBrayer 2006, Xu and Ji 2006). In this context, detected significant differences of preferable environmental temperatures can explain shifts in habitat selection and activity time of adult and neonate
Variable Adults Neonates T-test Mann-Whitney U p-value
Ta 22.9 ± 0.7 16.5-28.5
24
20.5 ± 0.8 16.5-26.5
152.17 - < 0.05
Ta2
23.1 ± 1.1 17.0-27.8
21
18.3 ± 0.9 16.6-22.6
15- 2.33 < 0.05
Ts22.7 ± 0.8 18.9-30.0
21
19.5 ± 1.4 14.0-25.0
112.11 - < 0.05
Table 3. Descriptive statistics of ambient temperatures for adult and neonate of G. halys and their comparison. Descriptive statis-tics are given as mean ± SE (top), range (middle) and sample size n (bottom). Abbreviations of variables: Ta, air temperature; Ta2, air temperature on distance of 5 cm from ground; Ts, temperature of substrate.
Simonov4
pit-vipers. Due to the higher vegetation coverage, in the stony steppe, individuals have more possibilities to move faster from shade to sun. Moreover, in this type of habitat, environment temperatures are more heterogeneous than in the rocky areas and bottomland. Neonates activity starts earlier and stops later, more likely because they prefer lower ambient temperatures than adults. They are active during the day and choose
active microhabitats to maintain optimal temperatures. Ambient temperatures affect activity patterns in many snake species (Gannon and Secoy 1985, Luiselli and Akani 2002, Litvinov 2008) and differences on preferred temperatures may determinate dissimilar activity.Diet properties also seem to be very important factors for shifts in habitat selection. Studies in different parts of G. h. halys area show that juveniles prefer to feed on insects (especially Orthoptera) and small lizards, whereas adult pit vipers prefer small mammals, rarely lizards (Chernov 1954, Lesnyak 1964, Yakovleva 1964, Yakovlev 1985, Orlov et al. 2001). According to Lesnyak (1964), in southeast Kazakhstan the diet of G. halys (with SLV up to 400 mm) consist to 60.6% of invertebrates. Such ontogenetic shifts in diet composition are well known for snakes (e.g. Saint-Girons 1980, Luiselli et al. 1995, Brito 2004), because they are gape-limited predators (Mushinsky 1987). During observations, numerous neonate lizards (Lacerta agilis L., 1758) were recorded in the stony steppe, whereas in the rocky area and bottomland only few L. agilis and Zootoca vivipara Van Jacquin, 1787 were observed. It is well known that allocation and abundance of insects are mainly dependent on plant diversity and coverage (e.g. Yakhontov 1969, Speight et al. 1999). Thus, the stony steppe habitat may
Figure 3. Activity of adult and neonate of G. h. halys recorded during the surveys (summary of 23rd to 30th August 2008).
Habitat differences in adult and neonate of G. h. halys 5
Figure 4. Changes of habitat temperatures during the observation period (23rd to 30th August 2008). Middle points represent mean values and whiskers ± 0.95*SE. Abbreviations of variables: Ta, air temperature; Ta2, air temperature on distance of 5 cm from ground; Ts, temperature of substrate.

Simonov6
provide the highest prey density for neonate pit-vipers. Differences in diet preference may also lead to shifts in the habitat use of neonate and juvenile snakes of various species (e.g. Lind 1990, Weatherhead et al. 2003). Additional data on the ecology of neonate, juvenile as well as for subadult of G. h. halys are needed to confirm these observations, and data on feeding and prey availability in different types of habitat are particularly needed, because they will help understanding habitat preferences of pit-vipers and will allow to infer prognosis on habitat suitability (of interest for conservation actions). Furthermore, data on preferred body temperatures are needed to provide the correct explanation of shifts in habitat selection, and represent some of the purposes for future research.Summarizing, this study indicates differences in habitat selection, daily activity and preferred ambient temperatures between neonate and adult Halys pit vipers in the examined population. Thermal requirements, diet and other biotic factors (such as predation risk) may be the most likely causes of ontogenetic changes in habitat selection. My preliminary results indicate that the stony steppe must be considered as the critical habitat for the reproduction of G. h. halys population in Novosibirsk region.
AcknowledgementsI thank V.K. Zinchenko (Institute of Animal Systematic and Ecology, Russia) for his help with fieldwork and N.N. Kolobaev (Norskiy reserve, Russia) who provided his unpublished data about ecology of G. h. halys in the Zeya sanctuary. Also, I thank anonymous reviewers for useful comments.
ReferencesAmat, F., Llorente, G.A., Carretero, M.A. (2003): A preliminary
study on thermal ecology, activity times and microhabitat use of Lacerta agilis (Squamata: Lacertidae) in the Pyrenees. Folia Zool. 52 (4): 413-422.
Ananieva, N.B., Orlov, N.L., Halikov, R.G., Darevskiy, I.S., Ry-abov, S.A., Barabanov, A.B. (2004): Atlas of North Eurasian retiles. St. Petersburg, Zoological Institute of Russian Acad-emy of Sciences. [in Russian]
Ataev, Ch., (1985): Reptiles of Turkmenistan’s mountains. Ash-khabad, Ylym. [in Russian]
Bjorgan, L. (2005): Habitat use and movement patterns of juve-nile black ratsnakes (Elaphe obsoleta) and their conservation implications. M.Sc. Thesis, University of Ottawa.
Blouin-Demers, G., Bjorgan, L. P.G., Weatherhead, P.J. (2007): Changes in habitat use and movement patterns with body size in black ratsnakes (Elaphe obsoleta). Herpetologica 63(4): 421-429.
Borkin, L.J., Munkhbayar, Kh., Orlov, N.L., Semenov, D.V., Terbish, Kh. (1990): Distribution of Reptiles in Mongolia. In: Reptiles of mountain and arid territories: systematics and
distribution. USSR Acad. Sci., Proc. Zool. Inst., Vol. 207, p. 22-137. Borkin, L.J., Ed., Leningrad. [in Russian]
Brito, J.C. (2003): Seasonal and daily activity patterns of Vipera latastei in northern Portugal. Amphibia-Reptilia 24: 497-508.
Brito, J.C. (2004): Feeding ecology of Vipera latastei in northern Portugal: ontogenetic shifts, prey size and seasonal variations. Herpetol. J. 14: 13-19.
Carretero, M.A., Marcos, E., de Prado, P. (2006): Intraspecific variation of preferred temperatures in the NE form of Podarcis hispanica. In: Mainland and insular lacertid lizards: a Mediter-ranean perspective, p. 55-64. Corti, C., Lo Cascio, P., Biaggini, M., Eds., Florence, Firenze University Press.
Cherlin, V.A. (1989): Age differences of thermobiological char-acteristics of some lizards. Proc. Acad. Sci. Turkmenian SSR 1: 35-38. [in Russian]
Chernov, S.A. (1954): Ecological and faunistic survey of reptiles in the south of Volga-Ural interfluve. Proc. Zool. Inst. USSR Acad. Sci. 16: 137-158. [in Russian]
Christian, K., Tracy, C.R., Porter, W.P. (1983): Seasonal shifts in body temperature and use of microhabitats by Galapagos land iguanas (Conolophus pallidus). Ecology 64: 463-468.
Gannon, V.P.J., Secoy, D.M. (1985): Seasonal and daily activity patterns in a Canadian population of the prairie rattlesnake, Crotalus viridis viridis. Can. J. Zool. 63: 86-91.
Greenacre, M.J. (1984): Theory and applications of correspon-dence analysis. New York, Academic Press.
Heath, J.E. (1965): Temperature regulation and diurnal activity in horned lizards. Univ. Calif. Publ. Zool. 64: 97-136.
Hitchcock, M., McBrayer, L.D. (2006): Thermoregulation in noc-turnal ecthotherms: seasonal and intraspecific variation in the Mediterranean gecko (Hemidactylus turcicus). J. Herpetol. 40: 185-195.
Huey, R.B., Pianka, E.R., Hoffman, J.A. (1977): Seasonal varia-tion in thermoregulatory behavior and body temperature of diurnal Kalahari lizards. Ecology 58: 1066-1075.
Keren-Rotem, T., Bouskila, A., Geffen, E. (2006): Ontogenetic habitat shift and risk of cannibalism in the common chame-leon (Chamaeleo chamaeleon). Behav. Ecol. Sociobiol.Behav. Ecol. Sociobiol. 59: 723–731.
Kolobaev, N.N. (2006): Influence of microclimate on morpholog-ical features of Halys Pit Viper. In: Climate change impact on ecosystems of Amur River basin, p. 120-128. Darman, U.A., Kokorin, A.O., Minin, A.A., Eds., Moscow, WWF Russia. [in Russian]
Kravcov, V.M., Donukalova, R.P. (1996): Geography of Novo-sibirsk region. Novosibirsk, Design studio “INFOLIO”. [in Russian]
Kuzmin S.L., Semenov, D.V. (2006): Synopsis of amphibian and reptilian fauna of Russia. Moscow, Association of science edi-tions KMK. [in Russian]
Lesnyak A.P. (1964): Ecology and maintenance in captivity of Halys pit viper Ancistrodon halys (Pallas 1776). Abstract of Ph.D. Dissertation. Tashkent. [in Russian]
Lind, A.J. (1990): Ontogenetic changes in the foraging behavior, habitat use and food habits of the western aquatic garter snake, Thamnophis couchii, at Hurdygurdy Creek, Del Norte County, California. M.Sc. Thesis, Humboldt State University.

Habitat differences in adult and neonate of G. h. halys 7
Litvinov N.A. (2008): The body temperature and microclimatic conditions of habitat for reptiles in the Volga river basin. Zool. Zh. 87(1): 1-13. [in Russian]
Luiselli, L., Akani, G.C. (2002): Is thermoregulation really unim-portant for tropical reptiles? Comparative study of four sympa-tric snake species from Africa. Acta Oecol. 23: 59-68.
Luiselli, L., Anibaldi, C., Capula, M. (1995): The diet of juvenile adders, Vipera berus, in an alpine habitat. Amphibia-Reptilia 16: 404-407.
Meek, R., Jayes, A.S. (1982): Body temperatures and activity pat-terns of Testudo graeca in North West Africa. Br. J. Herpet. 6: 194-197.
Mushinsky, H.R. (1987): Foraging ecology. In: Snakes - Ecology and Evolutionary Biology, p. 302-334. Seigel, R.A., Collins, J.T., Novak, S.S., Eds., New York, McGraw Hill.
Nemes, S., Vorgin, M., Hartel, T., Ollerer, K. (2006): Habitat se-lection at the sand lizard (Lacerta agilis): ontogenetic shifts. North-West. J. Zool. 2 (1):17-26.
Orlov, N. L., Barabanov, A. V. (2000) Classification of the Ag-kistrodon halys-intermedius complex: a critical review. Russ. J. Herpet. 6 (3): 167-192.
Orlov, N.L., Ananieva, N.B., Ryabov, S.A., Halikov, R.G. (2001): Distribution and biology of pit-vipers in East and South-East Asia. In: Problems of Herpetology. Proceedings of the I con-gress of the Nikolsky’s herpetological society, p. 213-216. Ananieva, N.B., Darevskiy, I.S., Dunaev, E.A., Iordanskiy N.N., Kuzmin S.L., Orlova V.F., Eds., Pushino-Moscow. [in Russian]
Pestov, M.V. (2003): Halys pit viper – the new species of No-vosibirsk region’s fauna. In: Amphibians and reptiles in West Siberia, p. 35-38. Novosibirsk, “Revi-K” Press. [in Russian]
Prokopov, K.P., Staricov, S.V., Bratash, I.B. (2000): Vertebrates of East Kazakhstan. Ust’-Kamenogorsk, EKSU Press. [in Rus-sian]
Richardson, M.L., Weatherhead, P.J., Brawn, J.D. (2006): Habitat use and activity of Prairie Kingsnakes (Lampropeltis calliga-ster calligaster) in Illinois. J. Herpetol. 40(4): 424-428.
Saint-Girons, H. (1980): Modifications sélectives du régime des vipères (Reptilia: Viperidae) lors de la croissance. Amphibia-Reptilia 1: 127-136.
Schmidt-Nielsen, K. (1979): Desert animals: physiological pro-blems of heat and water. New York, Dover Publishers.
Shine, R., Sun, L., Kearney, M., Fitzgerald M. (2002): Why do juvenile Chinese pit-vipers (Gloydius shedaoensis) select ar-boreal ambush sites? Ethology 108: 897-910.
Simonov, E.P. (2007): Distribution and some ecological aspects of Mamushi (Gloydius halys) in the north of its habitat in the Novosibirsk region. Povolzhskiy J. Ecol. 2007(1): 71-74. [in Russian]
Simonov, E.P. (2008): Revision of north distribution’s board of Halys pit viper (Gloydius (Agkistrodon) halys) and its bioto-pical location on West Siberia territory. Bull. Mordovia Univ., Biol. Sci. 2: 65-70. [in Russian]
Speight, M.R., Hunter, M.D., Watt, A.D. (1999): Ecology of insects: concepts and applications. Oxford, Blackwell Publi-shing.
Stevenson, R.D. (1985): The relative importance of behavioral and physiological adjustments controlling body-temperature in terrestrial ectotherms. Am. Nat. 126: 362-386.
Strijbosch, H., Bonnemayer, J.J.A.M., Dietvorst, P.J.M. (1980): The Northenmost population of Podacris mularis (Lacertilia, Lacertidae). Amphibia-Reptilia 1: 161-172.
Sun, L., Shine, R., Debi, Z., Zhengren, T. (2001): Biotic and abi-otic influences on activity patterns of insular pit-vipers (Glo-ydius shedaoensis, Viperidae) from north-eastern China. Biol. Conserv. 97: 387-398.
Tabachishin, V.G., Zavialov, E.V., Tabachishina, I.E. (2006): Spa-tial distribution of Eremias arguta (Pallas, 1773) in north of its Volga habitat. Mod. Herpetol. 5/6: 117-124. [in Russian]
Weatherhead, P.J., Blouin-Demers, G., Cavey, K.M. (2003): Sea-sonal and prey-size dietary patterns of black rat snakes (Elaphe obsoleta obsoleta). Am. Midl. Nat. 150: 275–281.
Werner, E.E., Gilliam, J.F., Hall, D.J., Mittelbach, G.G. (1983): An experimental test of the effects of predation risk on habitat use in fish. Ecology 64:1540–1548.
Winne, C.T., Keck, M.B. (2005): Intraspecific differences in ther-mal tolerance of the diamondback watersnake (Nerodia rhom-bifer): effects of ontogeny, latitude, and sex. Comp. Biochem. Physiol., Part A Mol. Integr. Physiol. 140(1): 141-149.
Xu, X.F., Ji, X. (2006): Ontogenetic shifts in thermal tolerance, selected body temperature and thermal dependence of food as-similation and locomotor performance in a lacertid lizard, Ere-mias brenchleyi. Comp. Biochem. Physiol., Part A Mol. Integr. Physiol. 143(1): 118-124.
Yakhontov, V.V. (1969): Ecology of Insects. Moscow, Higher school. [in Russian]
Yakovlev, V.A. (1985): Amphibians and reptiles of the Altaic re-serve. Abstract of Ph.D. Dissertation. Leningrad. [in Russian]
Yakovleva, I.D. (1964): Reptiles of Kirghizia. Frunze. [in Rus-sian].






























