Embed Size (px)
Citation preview

40
http://journals.tubitak.gov.tr/biology/
Turkish Journal of Biology Turk J Biol(2014) 38: 40-47© TÜBİTAKdoi:10.3906/biy-1304-4
Development of an improved RNA interference vector system forAgrobacterium-mediated plant transformation
Umut TOPRAK1,4,*, Cathy COUTU1, Doug BALDWIN1, Martin ERLANDSON1,2, Dwayne HEGEDUS1,3
1Agriculture and Agri-Food Canada, Saskatoon, SK, Canada2Department of Biology, University of Saskatchewan, Saskatoon, SK, Canada
3Department of Food and Bioproduct Sciences, University of Saskatchewan, Saskatoon, SK, Canada4Department of Plant Protection, Faculty of Agriculture, University of Ankara, Turkey
* Correspondence: [email protected]
1. IntroductionThe application of RNA interference (RNAi), first discovered by Fire et al. (1998), has led to a better understanding of the functions of genes from mammals to plants. The technique is based on the use of dsRNA to target a specific gene, which eventually reduces the levels of the corresponding mRNA in the cytoplasm. In entomology, various studies have shown that RNAi has potential beyond functional gene analysis, that is, the control of insects (Huvenne and Smagghe, 2010). The concept is based on the silencing of the genes essential for normal physiological processes, which leads to retardation of insect development or death. The success of gene silencing as an insect control strategy is closely related to the functional importance of the target gene and the dsRNA delivery method. Development of transgenic plants encoding dsRNA (plant-mediated RNAi) targeting midgut genes has great potential as an insect control strategy (Baum et al., 2007; Mao et al., 2007; Pitino et al., 2011; Zha et al., 2011). Furthermore, the target of dsRNA, midgut
epithelial cells, is accessible through feeding (Turner et al., 2006). Various insect midgut genes, including those encoding structural peritrophic matrix proteins (Toprak et al. 2013), digestive enzymes (Rajagopal et al., 2002), chitin deacetylase (Toprak et al., 2013), chitinases (Zhu et al., 2008), and chitin synthase-B (Arakane et al., 2005) have been successfully silenced by RNAi.
The generation of Agrobacterium-mediated transgenic plants encoding dsRNA involves i) subcloning of the sense and antisense regions in vectors such as pDONR
207 (Pitino et al., 2011) or pHANNIBAL (Huang et al., 2006; Fairbairn et al., 2007; Zha et al., 2011; Zhang et al., 2011), ii) transfer of the silencing cassette into a plant binary vector, iii) transformation of Agrobacterium with the desired plasmids, and iv) plant transformation by Agrobacterium. The procedure is relatively complex and time consuming, and problems with plasmid instability related to antibiotic selection have been reported. For example, many Agrobacterium strains, such as C58 (Huang et al., 2006), GV3101 (Pitino et al., 2011; Zhang et al.,
Abstract: Plant-mediated RNA interference (RNAi) has shown a great potential in pest control and requires i) subcloning of sense/antisense regions in compatible vectors, ii) transfer of the silencing cassette into a binary vector, iii) transformation of Agrobacterium tumefaciens with desired binary plasmids, and iv) transformation of plants with Agrobacterium. The procedure is long and should ensure plasmid backbone stability; however, plasmid recombination due to antibiotic selection is a common problem. pGSA1252 is an RNAi silencing binary vector allowing direct cloning of hairpin structure; however, it possesses a chloramphenicol selection marker leading to plasmid recombination in various Agrobacterium strains. To solve this selection marker/Agrobacterium compatibility problem and to shorten the cloning process, we developed a new RNAi vector system containing the RNAi cassette of pGSA1252 in a plant expression vector, pMDC32, which has kanamycin as the selection marker. A T7 RNA polymerase promoter was also incorporated adjacent to the multiple cloning site, allowing for in vitro dsRNA synthesis. This vector was tested by transforming Arabidopsis thaliana with 4 different dsRNA constructs specific to insect midgut genes: insect intestinal mucin 1/4, peritrophic matrix protein 1, chitin deacetylase 1, and chitin synthase-B. This improved system shows no recombination and shortens the entire cloning procedure.
Key words: RNA interference, plant transformation, Agrobacterium tumefaciens, chloramphenicol, kanamycin, Arabidopsis thaliana
Received: 01.04.2013 Accepted: 02.08.2013 Published Online: 02.01.2014 Printed: 24.01.2014
Research Article

TOPRAK et al. / Turk J Biol
41
2011), and A136 and BG53 (Rogers et al., 2002) that are routinely used for Arabidopsis thaliana transformation possess an inducible resistance mechanism against chloramphenicol (Tennigkeit and Matzura, 1991), suggesting that effective selection with this antibiotic is not possible with many Agrobacterium strains (Rogers et al., 2002; Bent, 2006; Szakasits et al., 2007). This response also leads to promiscuous recombination and plasmid instability (Ballester et al., 1986, 1989).
The pGSA1252 is an RNAi-silencing binary plant transformation vector that allows direct cloning without the need to subclone the silencing cassette into an intermediate vector, thus shortening the entire cloning procedure. Synthesis of the dsRNA is driven by the powerful Cauliflower mosaic virus 35S promoter in the plant; however, pGSA1252 possesses a chloramphenicol selection marker. To solve the selection marker/Agrobacterium compatibility problem and to further shorten the cloning procedure, we developed an improved vector by transferring the RNAi cassette of pGSA1252 into the plant binary vector, pMDC32, which uses kanamycin (an appropriate antibiotic for selection of Agrobacterium) as the selection marker (Bent, 2006; Szakasits et al., 2007). A T7 RNA polymerase promoter was also incorporated into the constructs allowing dsRNA synthesis in vitro. The vector was tested by transforming Arabidopsis plants with 4 different dsRNA constructs specific to genes encoding insect intestinal mucin 1/4 (McIIM1/4) (Shi et al., 2004; Toprak et al., 2010), peritrophic matrix protein 1 (McPM1) (Shi et al., 2004), chitin deacetylase 1 (McCDA1) (Toprak et al., 2008), and chitin synthase-B (McCHS-B) from Mamestra configurata, one of the major pests of Brassica crops in North America.
2. Materials and methods2.1. Cloning of sense and antisense regions in pGSA1252 in Escherichia coli A region of 500, 552, 496, and 500 bp was selected and amplified for McCDA1 (GenBank accession no.: EU660852), McIIM1/4 [a region common to McIIM1 (AY057052) and McIIM4 (FJ670569)], McPM1 (AY277403), and McCHS-B, respectively, using total RNA from M. configurata larval midgut by reverse transcription-PCR (RT-PCR), as previously described (Toprak et al., 2013). The sense primers (Fpsen and Rpsen) and antisense primers (Fpant and Rpant) used for each amplicon are shown in the Table. In the design of primers, a T7 RNA polymerase promoter was incorporated into the Fpsen and Rpant primers allowing in vitro synthesis of dsRNA from the vectors (Table). The sense and antisense RT-PCR products were sequentially ligated into the pGSA1252 vector. Sense regions (except in McPM1) were first ligated between the NcoI and AscI restriction sites; then antisense regions were ligated between the BamHI and SpeI
restriction sites. The McPM1 sense region was ligated into pGSA1252 between the AscI and SwaI sites. All pGSA1252 plasmid constructs were transformed into Escherichia coli DH10B electrocompetent cells by electroporation. Transformants were grown on Luria–Bertani (LB) agar plates containing chloramphenicol (25 µg/mL), and the cultures were grown in 2 mL of LB overnight from a single colony. Plasmid DNAs were isolated using a QIAprep spin miniprep kit (Qiagen), and sequences of the sense and antisense regions were confirmed by sequencing using the primers Fvpsen-Rvpsen and Fvpant-Rvpant for each amplicon, respectively (Table). 2.2. Cloning of dsRNA cassette (sense-spacer-antisense regions in pGSA1252) in pMDC32 in E. coli To clone the sense-spacer-antisense cassettes of pGSA1252 constructs into pMDC32, the pGSA1252/McCDA1, pGSA1252/McIIM1/4, and pGSA1252/McCHS-B were first digested with XhoI, followed by removal of the XhoI site by the large fragment of DNA polymerase I (Klenow). The constructs were then digested with SpeI. In parallel, pMDC32 plasmid was first digested with PspOMI, treated with Klenow, and then digested with SpeI. The sense-spacer-antisense cassettes were then ligated into pMDC32. For the construction of the pMDC32/McPM1 construct, pGSA1252/McPM1 was digested with AscI and SpeI. In parallel, pMDC32 plasmid was digested with AscI and SpeI. The McPM1 sense-spacer-antisense cassette was then ligated into the AscI and SpeI restriction sites of pMDC32. E. coli DH10B electrocompetent cells were transformed with the pMDC32 plasmid constructs using electroporation. Transformants were grown on LB agar plates containing kanamycin (50 µg/mL), and the cultures were grown in 2 mL of LB overnight from a single colony. Plasmid DNAs were isolated using a QIAprep spin miniprep kit, and sequences were confirmed using the T7 primers.2.3. Transformation of Agrobacterium with pMDC32-containing sense-spacer-antisense and verification of backbone stability of plasmid isolated from transformed AgrobacteriumThe pMDC32 plasmids containing sense-spacer-antisense cassette were introduced into A. tumefaciens strain GV3103 pMP90 by electroporation. Transformants were grown on LB agar plates containing gentamycin (30 µg/mL) and kanamycin (50 µg/mL) for 2 days at 28 °C. A single colony was selected and grown (28 °C at 225 rpm) overnight in 10 mL of LB medium containing the corresponding antibiotics.
Plasmid DNAs were isolated using a QIAprep spin miniprep kit (the volumes of buffers P1, P2, and N3 were doubled to compensate for the high culture volume) and digested with HincII for the verification of plasmid backbone stability.

TOPRAK et al. / Turk J Biol
42
Table. RT-PCR primers1 used in the study.
Primer Sequence
McCDA1
Fpsen2 CATGCCATGGTAATACGACTCACTATAGGACTATGGCCTTACACCCT
Rpsen TTGGCGCGCCTGCATCCAGTAGTCCATTCC
Fpant CGCGGATCCTGCATCCAGTAGTCCATTCC
Rpant2 CTAGACTAGTTAATACGACTCACTATAGGACTATGGCCTTACACCCT
McIIM1/4
Fpsen2 CATGCCATGGTAATACGACTCACTATACCTGTCGACTTCACCATCCA
Rpsen TTGGCGCGCCGATGCTGGAGTCAACTGGGC
Fpant CGCGGATCCGATGCTGGAGTCAACTGGGC
Rpant2 CTAGACTAGTTAATACGACTCACTATACCTGTCGACTTCACCATCCA
McPM1
Fpsen2 TTGGCGCGCCTAATACGACTCACTATAGTGCGCCGAAGGTAGACCAG
Rpsen GGCATTTAAATGTTGGTCACAGCTGGGTTGG
Fpant CGCGGATCCGTTGGTCACAGCTGGGTTGG
Rpant2 CTAGACTAGTTAATACGACTCACTATAGTGCGCCGAAGGTAGACCAG
McCHS-B
Fpsen2 CATGCCATGGTAATACGACTCACTATAGGCATAGTCCTCCAGATCGT
Rpsen TTGGCGCGCCTCTGCTTCTTCGACGGAGGC
Fpant CGCGGATCCTCTGCTTCTTCGACGGAGGC
Rpant2 CTAGACTAGTTAATACGACTCACTATAGGCATAGTCCTCCAGATCGT
Other primers
Fvpsen ACGCTCGAGTATAAGAGCTC
Rvpsen ACCACGATGCCATGTTCATC
Fvpant CCATCAGCGATCTCTTTG
Rvpant TCTCATTAAAGCAGGACTC
Fpgus AGATGAACATGGCATCGTGG
Rpgus CTTGGGTGGTTTTTGTCACG
1Fpsen and Rpsen primers were used for the amplification of sense regions, while Fpant and Rpant primers were used for the amplification of antisense regions for McCDA1, McIIM1/4, McPM1, and McCHS-B. Fvpsen-Rvpsen and Fvpant-Rvpant primers were used for verifying the sequence of the sense and antisense regions, respectively. Fpgus and Rpgus primers were used to verify that the transgene was present in transformed Arabidopsis plants.2Underlined sequences in the Fpsen and Rpant primers indicate the site of T7 RNA polymerase promoter that allows dsRNA synthesis from the vectors in vitro.

TOPRAK et al. / Turk J Biol
43
2.4. Development of transgenic Arabidopsis plants and verification of dsRNA synthesis in transgenic plants by RT-PCRTransgenic A. thaliana plants were generated by Agrobacterium-mediated transformation using the floral dip method (Bechtold et al., 1993). A 50-mL culture of Agrobacterium containing the pMDC32/sense-spacer-antisense plasmids was prepared in LB with the corresponding antibiotics. Cultures were resuspended in 5% sucrose, 0.05% Silwet L77 to an OD600 of 0.8. Plants with young buds were dipped into the Agrobacterium solution, vacuum-infiltrated at 25 mmHg for 30 s and then slowly returned to normal atmospheric pressure. They were then grown in a growth chamber at 22 ± 1 °C until mature. Several thousand T0 seeds were collected and sown on selection medium [For 1 L: 2.15 g of MS salts, 10 g of sucrose, 7 g of agar, 500 µL of 100 mg/mL hygromycin (25 µg/mL), and 1.5 mL of 200 mg/mL Timentin (300 µg/mL); pH 5.7]. Seedlings were allowed to grow at 22 ± 1 °C for approximately 14 days with a 16 h day/8 h night cycle, and then 28 plants were gently removed from the plates and transferred to soil in the greenhouse. When plants had matured, several leaves were collected and stored at –80 °C. Total RNA was extracted using TRIzol reagent (Invitrogen) from 2–3 small leaves from 10 T1 plants. The presence of the transgene was confirmed by RT-PCR for pMDC32/McCDA1, pMDC32/McCHS, pMDC32/McIIM4, and pMDC32/McPM1 plants using total leaf RNA and the GUS-specific primers (Fpgus and Rpgus) representing the common spacer element (Table), as previously described (Toprak et al., 2013).
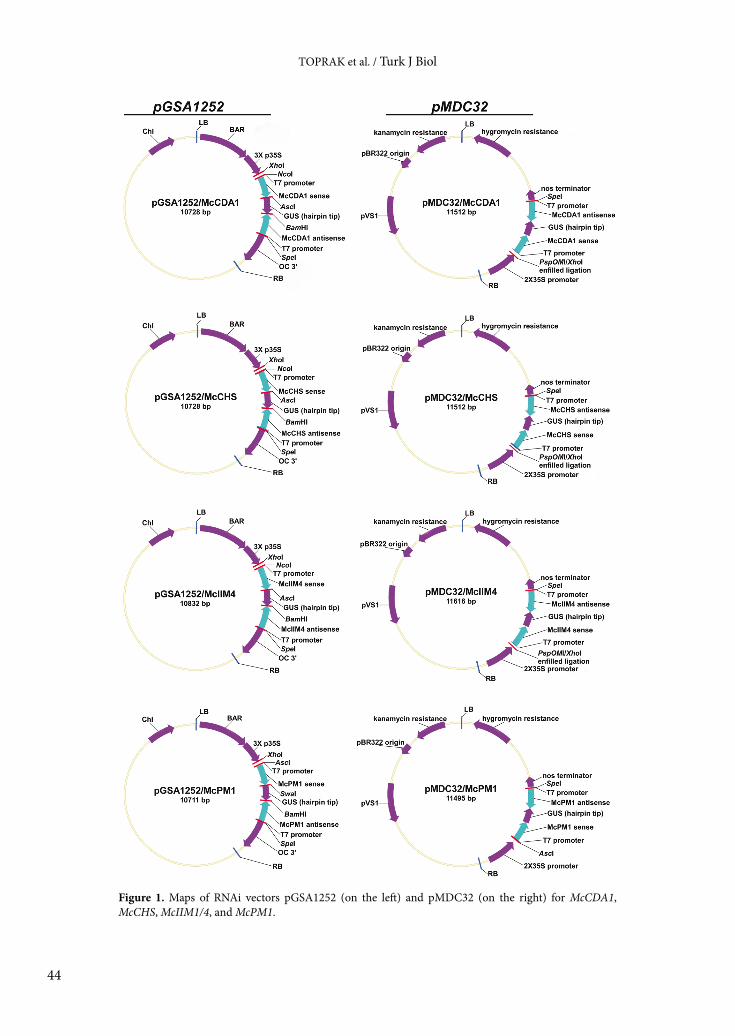
3. Results and discussionA fragment of 500, 552, 496, and 500 bp specific to 4 different midgut genes encoding McCDA1, McIIM1/4, McPM1, and McCHS-B, respectively, was amplified by RT-PCR using total midgut RNA. RT-PCR products were cloned into the RNAi vector, pGSA1252 (9706 bp), between NcoI and AscI restriction sites to create the sense strand (except for McPM1) and between the BamHI and SpeI restriction sites to create the antisense strand. The McPM1 sense region was cloned between the AscI and SwaI restriction sites. Sequencing and HincII restriction patterns of isolated plasmids from transformed E. coli indicated that the plasmid backbone was intact, and no recombination had occurred (data not shown). The pGSA1252 constructs for McCDA1, McIIM1/4, McPM1, and McCHS-B are 10728, 10832, 10711, and 10728 bp in length, respectively (Figure 1, left panel).
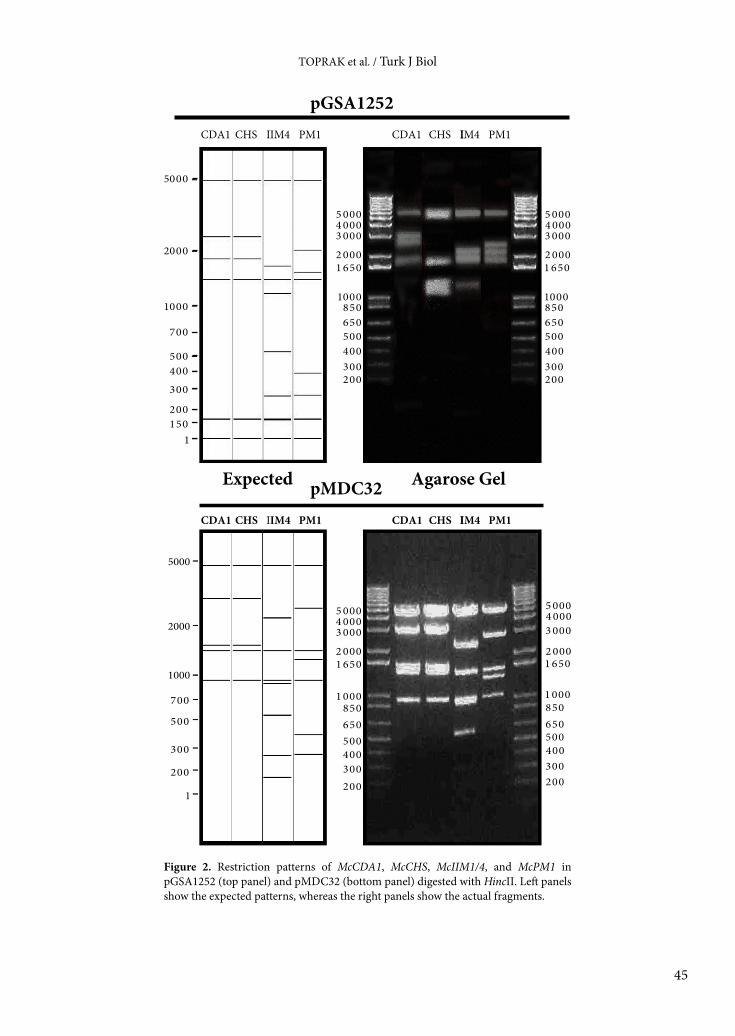
When the pGSA1252 constructs were introduced into Agrobacterium strains GV3103 or LBA4404, the isolated pGSA1252 plasmids revealed various recombination events, as detected by sequencing and HincII restriction
patterns (Figure 2, top panel). Similar recombinations were found to occur in pGSA1252 constructs containing shorter sense and antisense sequences (150 bp long; data not shown). pGSA1252 has a chloramphenicol-resistant selection marker, and many Agrobacterium strains such as C58 (Huang et al., 2006), GV3101 (Pitino et al., 2011; Zhang et al., 2011), and A136 and BG53 (Rogers et al., 2002) that are routinely used for A. thaliana transformation have been reported to possess an inducible resistance mechanism against this antibiotic (Tennigkeit and Matzura, 1991), suggesting that chloramphenicol selection causes recombination with many Agrobacterium strains (Rogers et al., 2002; Bent, 2006; Szakasits et al., 2007). Therefore, we decided to use a vector with a kanamycin-selectable marker, pMDC32, since kanamycin is an appropriate antibiotic for selection of Agrobacterium (Bent, 2006; Szakasits et al., 2007).
The RNAi-silencing cassettes (sense-spacer-antisense regions) for McCDA1, McIIM1/4, and McCHS-B were cut from pGSA1252 using XhoI and SpeI, followed by removal of the XhoI site by Klenow filling and ligation into SpeI and Klenow-treated PspOMI restriction sites in the plant binary vector pMDC32. The silencing cassette for pGSA1252/McPM1 was digested with AscI and SpeI and cloned into the same restriction sites in pMDC32. After transformation into Agrobacterium and selection by kanamycin and gentamycin, isolated pMDC32 plasmids containing the RNAi-silencing cassette were checked for correct insertion and backbone integrity via digestion with HincII; no recombination was found to have occurred in the new constructs (Figure 2, bottom panel). The inserts were also confirmed by DNA sequencing. Therefore, the new vector, pMDC32 containing the silencing cassette and kanamycin/hygromycin selection markers, is compatible with Agrobacterium transformation. The pMDC32 constructs for McCDA1, McIIM1/4, McPM1, and McCHS-B are 11,512, 11,616, 11,495, and 11,512 bp in length, respectively (Figure 1, right panel).
Arabidopsis thaliana plants transformed by Agrobacterium containing the pMDC32 constructs were grown on selection media containing hygromycin. The leaves of 10 F1 plants (containing pMDC32/McCDA1, pMDC32/McCHS, pMDC32/McIIM4, and pMDC32/McPM1) were tested for the presence of the T-DNA insert cassette by RT-PCR using total plant RNA. In this analysis, primers were designed to be specific for the common spacer (GUS) due to the folding of the gene-specific sense-antisense regions, which makes PCR amplification difficult. This analysis produced a band of 189 bp, verifying dsRNA synthesis for each transgenic plant (Figure 3).
The new vector developed in the current study has several improvements other than selection marker/Agrobacterium compatibility. First, it allows one-step
TOPRAK et al. / Turk J Biol
44
Figure 1. Maps of RNAi vectors pGSA1252 (on the left) and pMDC32 (on the right) for McCDA1, McCHS, McIIM1/4, and McPM1.
TOPRAK et al. / Turk J Biol
45
CDA1 CHS IIM4 PM1 CDA1 CHS IIM4 PM1
CDA1 CHS IIM4 PM1 CDA1 CHS IIM4 PM1
500040003000
2000
1000850650500
1650
400300200
pGSA1252
pMDC32Expected Agarose Gel
500040003000
2000
1000850650500
1650
400300200
500040003000
2000
1000850650500
1650
400300200
500040003000
2000
1000850650500
1650
400300200
5000
2000
1000
700
500400
300
200150
1
7
5000
2000
1000
00
500
300
200
1
Figure 2. Restriction patterns of McCDA1, McCHS, McIIM1/4, and McPM1 in pGSA1252 (top panel) and pMDC32 (bottom panel) digested with HincII. Left panels show the expected patterns, whereas the right panels show the actual fragments.

TOPRAK et al. / Turk J Biol
46
direct cloning into a plant transformation vector without subcloning of the sense and antisense regions in an intermediate vector followed by cloning into a binary plant vector. Thus, sense and antisense regions of any target gene could be easily cloned into the XhoI/AscI and BamHI/SpeI, respectively, in the binary vector (except
pMDC32/McPM1). Several RNAi-silencing vectors such as pHANNIBAL (Huang et al., 2006; Fairbairn et al., 2007; Zha et al., 2011; Zhang et al., 2011) have been used for plant-mediated RNAi. However, expression of dsRNA constructs in these vectors requires an additional cloning step into a binary vector (Fairbairn et al., 2007; Zha et al., 2011; Zhang et al., 2011). As a second improvement, the constructs allow in vitro synthesis of dsRNA through the addition of the T7 RNA polymerase promoter so that the intervening DNA templates can be used for in vitro synthesis of dsRNAs.
Plant-mediated RNAi possesses great potential as an insect control strategy in the field (Gordon and Waterhouse, 2007; Huvenne and Smagghe, 2010). The RNAi-silencing cassettes developed in this study were designed to target the genes McCDA1, McIIM1/4, McPM1, and McCHS-B, encoding proteins important for midgut physiology. Several RNAi-silencing vectors have been designed to target other midgut genes encoding proteins such as vacuolar ATPase subunit A (Baum et al., 2007), cytochrome P450 monooxygenase, and glutathione-S-transferase (Mao et al., 2007); receptor of activated kinase C (Pitino et al., 2011); hexose transporter; carboxypeptidase; and trypsin-like serine protease (Zha et al., 2011). Targeting midgut genes through plant-mediated RNAi could have great potential as an insect control strategy because of its essential role in insect digestive physiology and the indirect contact with the external environment, which allows for a per os application.
In conclusion, the new RNAi silencing system has several benefits, such as a selection marker compatible with Agrobacterium transformation, the T7 promoter allowing in vitro dsRNA synthesis, and one-step cloning without the subcloning of hairpin structure in an intermediate vector.
AcknowledgementsThe authors would like to thank Mrs Diana Bekkaoui (Agriculture and Agri-Food Canada) for technical assistance in the study.
M cC DA1
M 1 2 3 4 5 6 7 8 9 1 0
M cP M1
M cC HS
M cI IM 4
Figure 3. RT-PCR analysis to detect dsRNA synthesis in A. thaliana plants transformed by Agrobacterium containing pMDC32/McCDA1, pMDC32/McCHS, pMDC32/McIIM4, and pMDC32/McPM1 plasmids.
References
Arakane Y, Muthukrishnan S, Kramer KJ, Specht CA, Tomoyasu Y, Lorenzen MD, Kanost MR, Beeman RW (2005). The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix, respectively. Insect Mol Biol 14: 453–463.
Ballester S, Lopez P, Alonso JC, Espinosa M, Lacks SA (1986). Selective advantage of deletions enhancing chloramphenicol acetyltransferase gene expression in Streptococcus pneumoniae plasmids. Gene 41: 153–163.
Ballester S, Lopez P, Espinosa M, Alonso JC, Lacks SA (1989). Plasmid structural instability associated with pC194 replication functions. J Bacteriol 171: 2271–2277.
Baum JA, Bogaert T, Clinton W, Heck GR, Feldmann P, Ilagan O, Johnson S, Plaetinck G, Munyikwa T, Pleau M et al. (2007). Control of coleopteran insect pests through RNA interference. Nat Biotechnol 25: 1322–1326.
Bechtold N, Ellis J, Pelletier G (1993). In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. CR Acad Sci Paris, Life Sciences 316: 1194–1199.

TOPRAK et al. / Turk J Biol
47
Bent AF (2006). Arabidopsis thaliana floral dip transformation method. In: Wang K, editor. Agrobacterium Protocols. Totowa, NJ, USA: Humana Press, pp. 87–103.
Fairbairn DJ, Cavallaro AS, Bernard M, Mahalinga-Iyer J, Graham MW, Botella JR (2007). Host-delivered RNAi: an effective strategy to silence genes in plant parasitic nematodes. Planta 226: 1525–1533.
Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC (1998). Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391: 806–811.
Gordon KHJ, Waterhouse PM (2007). RNAi for insect-proof plants. Nat Biotechnol 25: 1231–1232.
Huang G, Allen R, Davis EL, Baum TJ, Hussey RS (2006). Engineering broad root-knot resistance in transgenic plants by RNAi silencing of a conserved and essential root-knot nematode parasitism gene. Proc Natl Acad Sci USA 103: 14302–14306.
Huvenne H, Smagghe G (2010). Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: a review. J Insect Physiol 56: 227–235.
Mao YB, Cai WJ, Wang JW, Hong GJ, Tao XY, Wang LJ, Huang YP, Chen XY (2007). Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat Biotechnol 25: 1307–1313.
Pitino M, Coleman AD, Maffei ME, Ridout CJ, Hogenhout SA (2011). Silencing of aphid genes by dsRNA feeding from plants. PLoS ONE 6:e25709.
Rajagopal R, Sivakumar S, Agrawal N, Malhotra P, Bhatnagar RK (2002). Silencing of midgut aminopeptidase N of Spodoptera litura by double-stranded RNA establishes its role as Bacillus thuringiensis toxin receptor. J Biol Chem 277: 46849–46851.
Rogers EJ, Rahman MS, Hill RT, Lovett PS (2002). The chloramphenicol-inducible catB gene in Agrobacterium tumefaciens is regulated by translation attenuation. J Bacteriol 184: 4296–4300.
Shi X, Chamankhah M, Visal-Shah S, Hemmingsen SM, Erlandson M, Braun L, Alting-Mees M, Khachatourians GG, O’grady M, Hegedus DD (2004). Modeling the structure of the type I peritrophic matrix: characterization of a Mamestra configurata intestinal mucin and novel peritrophin containing 19 chitin binding domains. Insect Biochem Mol Biol 34: 1101–1115.
Szakasits D, Siddique S, Bohlmann H (2007). An improved pPZP vector for Agrobacterium-mediated plant transformation. Plant Mol Biol Rep 25: 115–120.
Tennigkeit J, Matzura H (1991). Nucleotide sequence analysis of a chloramphenicol-resistance determinant from Agrobacterium tumefaciens and identification of its gene product. Gene 98: 113–116.
Toprak U, Baldwin D, Erlandson M, Gillott C, Hou X, Coutu C, Hegedus DD (2008). A chitin deacetylase and putative insect intestinal lipases are components of the Mamestra configurata (Lepidoptera: Noctuidae) peritrophic matrix. Insect Mol Biol 17: 573–585.
Toprak U, Baldwin D, Erlandson M, Gillott C, Hegedus DD (2010). Insect intestinal mucins and serine proteases associated with the peritrophic matrix from feeding, starved and molting Mamestra configurata larvae. Insect Mol Biol 19: 163–175.
Toprak U, Baldwin D, Erlandson M, Gillott C, Harris S, Hegedus DD (2013). In vitro and in vivo application of RNA interference for targeting genes involved in peritrophic matrix synthesis in a lepidopteran system. Insect Science 20: 92–100.
Turner CT, Davy MW, MacDiarmid RM, Plummer KM, Birch NP, Newcomb RD (2006). RNA interference in the light brown apple moth, Epiphyas postvittana (Walker) induced by double-stranded RNA feeding. Insect Mol Biol 15: 383–391.
Zha W, Peng X, Chen R, Du B, Zhu L, He G (2011). Knockdown of midgut genes by dsRNA-transgenic plant-mediated RNA interference in the hemipteran insect Nilaparvata lugens. PLoS ONE 6: e20504.
Zhang M, Wang Q, Xu K, Meng Y, Quan J, Shan W (2011). Production of dsRNA sequences in the host plant is not sufficient to initiate gene silencing in the colonizing oomycete pathogen Phytophthora parasitica. PLoS ONE 6: e28114.
Zhu Q, Arakane Y, Beeman RW, Kramer KJ, Muthukrishnan S (2008). Functional specialization among insect chitinase family genes revealed by RNA interference. Proc Natl Acad Sci USA 105: 6650–6655.





























