Embed Size (px)
Citation preview

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Determinación sexual en primatesDeterminación sexual en primatesneotropicales: el caso de los monosneotropicales: el caso de los monos
aulladoresaulladores
Steinberg, Eliana Ruth
2011
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
Steinberg, Eliana Ruth. (2011). Determinación sexual en primates neotropicales: el caso de losmonos aulladores. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires.
Cita tipo Chicago:
Steinberg, Eliana Ruth. "Determinación sexual en primates neotropicales: el caso de los monosaulladores". Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. 2011.

UNI VERSIDAD DE BUENOS AI RES FACULTAD DE CI ENCI AS EXACTAS Y NATURALES
DEPARTAMENTO DE ECOLOGÍ A, GENÉTI CA Y EVOLUCI ÓN
“Determ inación sexual en Pr im ates Neot ropica les: e l caso de los m onos aulladores”
Tesis presentada para optar al t ítulo de Doctor de la Universidad de Buenos Aires en el área Ciencias Biológicas
Lic. Eliana Ruth Ste inberg Directora de Tesis: Prof. Dra. Marta Dolores Mudry Consejera de Estudios: Prof. Dra. Marta Dolores Mudry Lugar de t rabajo: Grupo de I nvest igación en Biología Evolut iva (GI BE) , Departamento de Ecología, Genét ica y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires
Buenos Aires, Argent ina
Marzo, 2011

Determ inación sexual en Pr im ates Neot ropica les:
e l caso de los m onos aulladores
La Cariosistem át ica perm ite com parar taxa relacionados, en part icular
especies. Una var iable de análisis con valor diagnóst ico es el t ipo de sistema
de determ inación sexual. En este Trabajo de Tesis se analizaron m onos
aulladores del género Alouat ta con sistem as de determ inación sexual
m últ iple resultado de t ranslocaciones Y-autosom a. Se los comparó con ot ros
3 Ceboidea (Cebus, Saim ir i y Aotus) y con dos Hom inoidea (Pan t roglodytes
y Hom o sapiens) ante la diversidad de pat rones de determ inación sexual,
en part icular en m achos. Se realizó la prim era caracterización del cariot ipo
de Alouat ta pigra que m ostró 2N= 58, X1X1X2X2 / X1X2Y1Y2. Se estudió
conservación genóm ica por FI SH con las sintenias 3/ 15 y 3/ 21 evidenciando
que estas asociaciones no estarían conservadas en las especies
m esoam ericanas A. pigra y A. palliata. Este estudio junto al de hom eologías
crom osóm icas, m ost ró que los autosom as involucrados en las
t ranslocaciones que darían origen a los m ult ivalentes sexuales en las
especies sudam ericanas y m esoam ericanas serían dist intos. En el m arco
conceptual de “Evidencia Total” , el análisis combinado de variables
m oleculares y crom osóm icas resolvió las relaciones de parentesco ent re las
especies de aulladores de am bos orígenes am ericanos, dem ost rando su
ut ilidad en el esclarecim iento de cont roversias que relacionan la Taxonomía
y la Evolución de los pr imates ceboideos.
Palabras claves: Alouat ta, evolución crom osómica, Meiosis, sistem as de
determ inación sexual, Evidencia Total, SRY, sintenias cromosóm icas,
filogenias por análisis combinado.

Sex determ inat ion in neot ropica l pr im ates:
the exam ple of the How ler Monkeys
Karyosystem at ics allows for com parison of related taxa, part icular ly species.
The sex determ inat ion system const itutes a var iable with diagnost ic value.
I n this dissertat ion howler m onkeys, genus Alouat ta, with mult iple sex
determ inat ion systems product of Y-autosom e t ranslocat ions, were
analized. I n light of the diversity of sex determ inat ion pat terns observed,
part icularly in males, they were com pared with other 3 Ceboidea (Cebus,
Saim ir i and Aotus) and with 2 Hom inoidea (Pan t roglodytes and Homo
sapiens) . The first cytogenet ic character izat ion of Alouat ta pigra karyotype
was perform ed, exhibit ing a 2N= 58, X1X1X2X2/ X1X2Y1Y2. Genom ic
conservat ion was studied by FI SH analizing the syntenies 3/ 15 and 3/ 21,
showing that these asociat ions are not conserved in the m esoam erican
howlers, A. pigra y A. palliata. This analysis, together with the chrom osomal
homologies, dem onst rated that the autosom al pairs involved in the
t ranslocat ions that originate the sexual mult ivalents are different in the
southam erican and m esoam erican species. Under the fram ework of the
“Total Evidence” concept , the combined análisis of molecular and
chrom osom al variables resolved the phylogenet ic relat ionships between the
howler species of both am erican origins, demonst rat ing its ut ility to clarify
the cont roversies related to the Taxonom y and Evolut ion of Ceboidea
primates.
Key words: Alouat ta, chromosom e evolut ion, Meiosis, sex determ inat ion
system s, Total Evidence, SRY, chrom osom al syntenies, phylogenies by
combined analysis.

Agradecim ientos
A la Universidad de Buenos Aires (UBACyT X107 y X145 M.D.M.) , en
part icular a la Facultad de Ciencias Exactas y Naturales y al departam ento
de Ecología, Genét ica y Evolución que posibilitaron y facilitaron m i
form ación doctoral así cómo el desarrollo experim ental objeto de este
t rabajo de Tesis.
Al CONI CET, inst itución nacional que m ediante los subsidios PI P 5012 y
112-200801-00744 concedidos a m i Directora de Tesis y las becas internas
de postgrado t ipo I y I I que m e fueron otorgadas avaló la concreción de
este t rabajo de Tesis Doctoral.
A los directores de las I nst ituciones, tales com o Jardines Zoológicos,
Bioter ios y cent ros de cría, que perm it ieron la realización estos estudios así
como a los profesionales, veterinarios y cuidadores, que colaboraron en la
obtención de las muest ras y seguim iento de los anim ales de caut iverio
objeto de estudio.
A la Dra. Marta D. Mudry, m i directora de Tesis, m i m aest ra en la ciencia y
en la vida. Gracias por enseñarm e a am ar la Biología y a siem pre t ratar de
superarnos.
A la Dra. Mariela Nieves por su invaluable colaboración en el desarrollo e
interpretación de las técnicas citogenét icas.
Al Lic. Alejandro Laudicina, por su paciencia para responder a m is infinitas
preguntas sobre FI SH y ayudarm e a sacar adelante la técnica en los
ceboideos.
A la Dra. M. García Caldés por su cont r ibución a la discusión de las
hom eologías en aulladores.
Al equipo de invest igadores de la Unitat de Biologia Cellular i Genèt ica
Mèdica, Facultat de Medicina, de la Universidad Autónom a de Barcelona por

su colaboración en el análisis por inmunofluorescencia de células
germ inales.
Al equipo m exicano de invest igadores y a todo el equipo de cam po por su
colaboración en la obtención de las m uest ras de aulladores de dist r ibución
natural m arginal norte.
A Juan Pablo Luaces y Luis Rossi, por hacer de m i estadía en FMed una
grata experiencia y por el intercam bio de saberes en la aplicación e
interpretación de las técnicas citogenét icas.
A los jurados que evaluarán m i t rabajo de Tesis Doctoral, que con sus
aportes y sugerencias ayudarán a m ejorar lo.
A Gaby, Mae, Lucía, Nancy, Carola, Gise, Ana, Crist ina y Leandro por
com part ir conm igo el cam ino de la ciencia con m ates, aguante y cont inuas
charlas.
A m is am igos, por com prenderm e, soportarm e y acom pañarm e durante los
años que duró m i doctorado.
A Carlos, por estar a m i lado, por comprenderm e, por su paciencia y por ser
m i cable a t ierra durante todos estos años.
A m is padres y a m i herm ana, por el apoyo incondicional y por hacerm e la
persona que soy hoy.

Í ndice
1 . I nt roducción ……………………………………………………………………………………………………… 1
1.1 Rearreglos cromosóm icos y especiación………………………………………………………… 1
1.2 Cariosistemát ica………………………………………………………………………………………………. 2
1.2.1 Cromosom as sexuales, especiación y cariosistemát ica………………………… … 3
1.2.2 Mecanism os de determ inación sexual……………………………………………………... 4
1.2.2.1 Plantas……………………………………………………………………………………………………. 5
1.2.2.2 I nvertebrados……………………………………………………………………………………….. 7
1.2.2.3 Vertebrados………………………………………………………………………………………….. 10
1.2.2.3.1 Prim ates……………………………………………………………………………………………. 14
1.2.3 Sintenias crom osóm icas…………………………………………………………………………….. 17
1.2.3 1 Primates……………………………………………………………………………………………….. 19
1.2.4 El gen SRY…………………………………………………………………………………………………. 22
1.2.4.1 Primates…………………………………………………………………………………………………. 23
1.3 El m odelo de estudio………………………………………………………………………………………. 24
1.3.1 El género Alouat ta…………………………………………………………………………………….. 26
1.3.1.1 Diagnosis citogenét ica………………………………………………………………………… 29
1.3.1.2 Asociaciones crom osóm icas y sistem as de determ inación sexual……… 31
1.3.1.3 Estudios filogenét icos…………………………………………………………………………. 32
2 . Objet ivos e Hipótesis ………………………………………………………………………………………. 34
2.1 Objet ivos………………………………………………………………………………………………….... .. ... 34
2.1.1 Objet ivo General………………………………………………………………………………………… 34
2.1.2 Objet ivos específicos…………………………………………………………………………………. 34
2.2 Hipótesis de t rabajo………………………………………………………………………………………… 3
3 . Mater ia les y Métodos ………………………………………………………………………………………… 35
3.1 Material biológico…………………………………………………………………………………………….. 35
3.2 Diagnosis por caracterización citogenét ica clásica…………………………………………. 38
3.2.1 Estudios en células somát icas…………………………………………………………………… 38
3.2.1.1 Ext racción de las m uest ras………………………………………………………………….. 38
3.2.1.2 Cult ivo de sangre perifér ica…………………………………………………………………. 38
3.2.1.3 Análisis de metafases…………………………………………………………………………… 40
3.2.1.3.1 Determ inación del índice m itót ico ( I M% )………………………………………. 40
3.2.1.3.2 Determ inación del núm ero m odal (2N) y fundamental (NF)………… 40
3.2.1.4 Variables cromosóm icas………………………………………………………………………. 41
3.2.1.4.1 Mediciones cromosóm icas………………………………………………………………. 41
3.2.1.4.2 Bandas G con t inción de Wright (García Haro, 2000, m od.)…………….. 43
3.2.1.4.3 Bandas G con t r ipsina (Seabright 1971, m od.)…………………………………. 44

Í ndice
3.2.1.4.4 Bandas C (Sum ner, 1972)……………………………………………………………..... 44
3.2.1.4.5 Bandas NOR (Howel & Black, 1980)………………………...... .. . . . . . . . . . . . . .. . 45
3.2.1.5 Armado y confección de los cariot ipos………………………………………………… 45
3.2.1.6 Homeologías cromosóm icas………………………………………………………………… 47
3.2.2 Estudios en células germ inales…………………………………………………………………. 48
3.2.2.1 Ext racción de las m uest ras…………………………………………………………………. 48
3.2.2.2 Análisis con m icroscopía ópt ica…………………………………………………………… 48
3.3 Diagnosis por caracterización citom olecular………………………………………………….. 50
3.3.1 Estudios en células somát icas…………………………………………………………………… 50
3.3.1.1 Hibridación in situ Fluorescente (FI SH)………………………………………………. 50
3.3.1.1.1 Sondas…………………………………………………………………………………………….. 50
3.3.1.1.2 Extendidos………………………………………………………………………………………. 50
3.3.1.1.2.1 Preparación……………………………………………………………………………….. 50
3.3.1.1.2.2 Envejecim iento………………………………………………………………………….. 51
3.3.1.1.3 Hibridación………………………………………………………………………………………. 52
3.3.1.1.3.1 Est rategia 1: desnaturalización con form am ida……………………… . 53
3.3.1.1.3.1.1 Desnaturalización por separado………………………………………….. 53
3.3.1.1.3.1.2 Desnaturalización conjunta………………………………………………….. 54
3.3.1.1.3.2 Est rategia 2: desnaturalización con Hidróxido de sodio………….. 55
3.3.1.1.4 Revelado……………………………………………………………………………………….. 56
3.3.2 Estudios en células germ inales…………………………………………………………………. 56
3.3.2.1 Análisis con m icroscopía ópt ica de fluorescencia…………………………………. 56
3.3.2.1.1 Extensión de esperm atocitos……………………………………………………………. 57
3.3.2.1.2 I nmunofluorescencia…………………………………………………………………………. 58
3.3.2.1.2.1 Ant icuerpos prim arios…………………………………………………………………. 58
3.3.2.1.2.2 Ant icuerpos secundarios…………………………………………………………….. 58
3.3.2.1.2.3 Protocolo……………………………………………………………………………………… 59
3.3.2.1.3 Hibridación in situ…………………………………………………………………………….. 59
3.3.2.1.3.1 Sondas…………………………………………………………………………………………. 59
3.3.2.1.3.2 FI SH……………………………………………………………………………………………… 60
3.4 Estudios filogenét icos………………………………………………………………………………………. 60
3.4.1 Elección de los caracteres………………………………………………………………………….. 60
3.4.2 Confección de las mat rices…………………………………………………………………………. 61
3.4.3 Análisis filogenét ico……………………………………………………………………………………. 63
3.5 Fotografía…………………………………………………………………………………………………………. 63
4 . Resultados ………………………………………………………………………………………………………… 64
4.1 Diagnosis por fenot ipo externo………………………………………………………………………. 64

Í ndice
4.1.1 Pat rón de coloración de pelaje………………………………………………………………… 64
4.1.2 Otros…………………………………………………………………………………………………………. 69
4.2 Diagnosis por caracterización citogenét ica clásica…………………………………………. 70
4.2.1 Estudios en células somát icas……………………………………………………………………. 70
4.2.1.1 Determ inación del Í ndice Mitót ico ( I M% )……………………………………………. 70
4.2.1.2 Análisis m itót icos………………………………………………………………………………….. 72
4.2.1.2.1 Género Alouat ta (Atelidae: Ceboidea)……………………………………………. 72
4.2.1.2.1.1 Alouat ta caraya……………………………………………………………………….. 72
4.2.1.2.1.1.1 Determ inación del 2N y NF……………………………………………….. 72
4.2.1.2.1.1.2 Pat rones de bandas G y C…………………………………………………. 73
4.2.1.2.1.2 Alouat ta guariba clam itans……………………………………………………… 74
4.2.1.2.1.2.1 Determ inación del 2N y NF……………………………………………….. 74
4.2.1.2.1.2.2 Pat rones de bandas G y C…………………………………………………. 75
4.2.1.2.1.3 Alouat ta pigra………………………………………………………………………….. 76
4.2.1.2.1.3.1 Determ inación del 2N y NF……………………………………………….. 76
4.2.1.2.1.3.2 Medidas crom osóm icas……………………………………………………… 77
4.2.1.2.1.3.3 Caracter ización de los pat rones de bandas G y C……………. 79
4.2.1.2.1.3.4 Crom osomas m arcadores………………………………………………….. 81
4.2.1.2.1.4 Alouat ta palliata………………………………………………………………………. 82
4.2.1.2.1.4.1 Determ inación del 2N y NF……………………………………………….. 82
4.2.1.2.1.4.2 Pat rones de bandas G y C…………………………………………………. 83
4.2.1.2.2 Ot ros géneros de Ceboidea (Cebidae: Platyrr ini)………………………….. 84
4.2.1.2.2.1 Saim ir i boliv iensis boliviensis……………………………………………………. 84
4.2.1.2.2.1.1 Determ inación del 2N y NF…………………………………………………. 84
4.2.1.2.2.1.2 Pat rones de bandas G y C…………………………………………………… 85
4.2.1.2.2.2 Cebus libidinosus………………………………………………………………………. 86
4.2.1.2.2.2.1 Determ inación del 2N y NF…………………………………………………. 86
4.2.1.2.2.2.2 Pat rones de bandas G y C…………………………………………………… 87
4.2.1.2.2.3 Cebus nigr itus……………………………………………………………………………. 88
4.2.1.2.2.3.1 Determ inación del 2N y NF………………………………………………….. 88
4.2.1.2.2.3.2 Pat rones de bandas G y C…………………………………………………… 89
4.2.1.2.2.4 Aotus azarae……………………………………………………………………………… 90
4.2.1.2.2.4.1 Determ inación del 2N y NF…………………………………………………. 90
4.2.1.2.2.4.2 Pat rones de bandas G y C………………………………………………….. 91
4.2.1.2.3 Hom inoidea…………………………………………………………………………………….. 93
4.2.1.2.3.1 Pan t roglodytes……………………………………………………………………….. 93
4.2.1.2.3.1.1 Determ inación del 2N y NF…………………………………………………. 93
4.2.1.2.3.1.2 Pat rones de bandas G y C………………………………………………….. 93

Í ndice
4.2.1.2.3.2 Homo sapiens………………………………………………………………………… 95
4.2.1.2.3.2.1 Determ inación del 2N y NF………………………………………………. 95
4.2.1.2.3.2.2 Pat rones de bandas G y C……………………………………………….. 95
4.2.1.3 Hom eologías y reorganizaciones crom osóm icas en aulladores…….. 97
4.2.1.3.1 Hom o sapiens y Alouat ta caraya…………………………………………………. 97
4.2.1.3.2 Alouat ta caraya y Alouat ta pigra. . . .. .. . . . .. .. . .. . .. .. .. . . . . . . . . . . .. . . . . . . . 109
4.2.1.3.3 Alouat ta caraya y Alouat ta palliata. . .. . . . .. .. . .. . .. .. .. . . . . . . . . . . .. . . . . . . 118
4.2.1.3.4 Com paración con ot ros aulladores. . . .. . . . . . . . . . . .. . . . . . . .. .. . .. . . . .. .. . . . 126
4.2.2 Estudios en células germ inales……………………………………………………………….. 128
4.2.2.1 Sistem as de determ inación sexual múlt iple……………………………………… 128
4.2.2.1.1 Alouat ta caraya……………………………………………………………………………. 128
4.2.2.1.1.1 Caracter ización de los estadíos de la m eiosis………………………. 128
4.2.2.1.1.2 El cuadrivalente sexual en Diacinesis/ Metafase I …………………. 129
4.2.2.1.2 Alouat ta pigra……………………………………………………………………………….. 130
4.2.2.1.2.1 Caracterización de los estadíos de la meiosis……………………….. 130
4.2.2.1.2.2 El cuadrivalente sexual en Diacinesis/ Metafase I ………………….. 132
4.2.2.1.3 Alouat ta palliata……………………………………………………………………………… 133
4.2.2.1.3.1 Caracterización de los estadíos de la meiosis………………………… 133
4.2.2.1.3.2 El t r ivalente sexual en Diacinesis/ Metafase I …………………………. 134
4.2.2.2 Sistem as de determ inación sexual XY “Hum an- like”………………………… 135
4.2.2.2.1 Cebus libidinosus…………………………………………………………………………… 135
4.2.2.2.1.1 Caracter ización de los estadíos de la m eiosis………………………… 135
4.2.2.2.1.2 El bivalente sexual en Diacinesis/ Metafase I …………………………. 136
4.2.2.2.2 Saim iri boliviensis boliviensis………………………………………………………… 136
4.2.2.2.2.1 Caracter ización de los estadíos de la m eiosis………………………… 136
4.2.2.2.2.2 El bivalente sexual en Diacinesis/ Metafase I …………………………. 137
4.3 Diagnosis por caracterización citom olecular………………………………………………… 138
4.3.1 Estudios en células somát icas………………………………………………………………… 138
4.3.1.1 Análisis de la dist r ibución de secuencias telom éricas en Ceboidea.. . 138
4.3.1.2 Análisis de conservación de sintenias……………………………………………….. 140
4.3.1.3 Análisis de conservación del SRY……………………………………………………… 148
4.3.1.3.1 Géneros de Hominoidea………………………………………………………………. 148
4.3.1.3.2 Género Alouat ta…………………………………………………………………………… 149
4.3.1.3.3 Ot ros géneros de Ceboidea…………………………………………………………. 151
4.3.2 Estudios en células germ inales………………………………………………………………. 153
4.3.2.1 Caracter ización del pat rón de recombinación…………………………………… 153
4.3.2.1.1 Cebus libidinosus………………………………………………………………………….. 153
4.3.2.1.1.1 Apaream iento crom osóm ico y sinapsis………………………………….. 153

Í ndice
4.3.2.1.1.2 Análisis de la frecuencia de recombinación…………………………… 155
4.3.2.1.1.3 Comportam iento del par sexual XY………………………………………… 156
4.3.2.1.2 Alouat ta caraya…………………………………………………………………………….. 158
4.3.2.1.2.1 Análisis de la frecuencia de recombinación……………………………. 158
4.3.2.1.2.2 Análisis del cuadrivalente sexual……………………………………………. 160
4.4 Estudios filogenét icos…………………………………………………………………………………... 161
4.4.1 Mat riz Básica de datos…………………………………………………………………………….. 161
4.4.2 Análisis por PAUP…………………………………………………………………………………….. 163
4.4.2.1 Caracteres crom osóm icos………………………………………………………………… 163
4.2.2.2 Caracteres m oleculares…………………………………………………………………… 165
4.2.2.3 Análisis combinado………………………………………………………………………….. 167
5 . Discusión …………………………………………………………………………………………………………. 170
5.1 Citogenét ica y diagnosis taxonóm ica.………………………………………………………….. 170
5.2 Com portam iento m eiót ico de los cromosomas sexuales….…………….………….. 172
5.2.1 Diagnosis por citogenét ica clásica..………………………………………….…………….. 172
5.2.2 Diagnosis por caracterización citom olecular…….……………………………………. 173
5.2.2.1 Cebus libidinosus……………………………………………………………………………… 173
5.2.2.2 Alouat ta caraya..……………………………………………………………………………… 175
5.3 El gen SRY en Ceboidea……………………………………………………………………………..… 175
5.4 Aportes a la sistem át ica del género Alouat ta………………………………………….….. 177
5.4.1 Sistem as de determ inación sexual…………………………………………………………. 177
5.4.2 Reconst rucción filogenét ica del género………………………………………………….. 179
5.4.3 Biogeografía y crom osom as sexuales en aulladores……………………………… 180
5.4.4 Perspect ivas…………………………………………………………………………………………….. 185
6 . Conclusiones ……………………………………………………………………………………………………. 187
7 . Bibliografía ………………………………………………………………………………………………………. 189
8 . Anexos
Anexo I : I dent ificación de los ejem plares de Ceboidea (Prim ates: Platyrrhini) analizados en el presente t rabajo de Tesis (N = 43)…………………………………………….. i
Anexo I I : Soluciones de t rabajo para citogenét ica clásica.……………………………………. ii
Anexo I I I : Soluciones de t rabajo e infraest ructura para estudios citom oleculares. vi
Anexo I V: Matr ices básicas de datos………………………………………………………………………. xi
Anexo V: Fuente de las fotografías ut ilizadas en las carátulas…………………………… . xix

I NTRODUCCI ÓN

1. I nt roducción
1
1 .1 Rearreglos crom osóm icos y especiación
Desde que Sut ton (1902) y Boveri (1904) propusieron que los
crom osom as son los portadores del m aterial hereditar io num erosas
evidencias han dem ost rado que el núm ero, la est ructura y la arquitectura
crom osóm ica t ienen un significado funcional en el m antenim iento de la
integridad genóm ica y el aislam iento reproduct ivo en las especies (White
1977; King 1993) .
Los rearreglos o reordenam ientos cromosóm icos ( inversiones,
t ranslocaciones, fusiones y fisiones, ent re ot ros) cont r ibuyen a la
reorganización del genom a de m am íferos, generando nuevas formas
crom osóm icas, fuente de var iabilidad sobre la que puede actuar la selección
natural (King 1993; Terence J. Robinson y Aurora Ruiz-Herrera 2010). Estos cam bios
afectan tanto a los autosom as com o a los cromosom as sexuales (o
gonosom as) , pudiendo tener o no efecto visible sobre el fenot ipo y pudiendo
o no cont r ibuir a la especiogénesis de un grupo.
Bajo el Concepto Biológico de Especie (Mayr 1942) , en los taxa que
difieren de sus especies relacionadas en el núm ero cromosómico o en ot ros
rasgos car iológicos, las diferencias crom osómicas ent re especies cercanas
podrían actuar com o m ecanism os de aislam iento reproduct ivo vinculables a
una dism inución de la fert ilidad de los híbridos, en ausencia de m ecanism os
precigót icos de aislam iento que im pidan las cruzas específicas (Reig 1984) .
Del espect ro de cambios cromosóm icos que pueden ocurr ir , sólo
aquellos que causan efectos heterót icos negat ivos (es decir, que dism inuyen
el “ fitness” de los heterocigotas para estos reordenam ientos) pueden tener
un papel en el proceso cladogénico. Estos son las fusiones en tándem, las
t ranslocaciones recíprocas, las fusiones y fisiones cént r icas, las fusiones
cént r icas múlt iples, las fusiones cént r icas m últ iples que com parten
homologías braquiales, las t ranslocaciones X-autosom a y las inversiones
pericént r icas y paracént r icas (King 1987) . Sin em bargo, si ocurre una
segregación balanceada de los productos m eiót icos en los heterocigotos
para estos cam bios, estos no const ituirán un m ecanism o de aislam iento

1. I nt roducción
2
reproduct ivo postcigót ico, sino que const ituirán polimorfismos
crom osóm icos.
Sin em bargo, si un reordenam iento part icular está o no involucrado
en el proceso especiogénico es definido en últ ima instancia por el sistema
m eiót ico del individuo. Un com plejo rearreglo est ructural puede segregar de
form a balanceada en un organismo y no en ot ro, y es este único evento el
que decide si un rearreglo puede tener un rol en la especiación (King 1987) .
De aquí se desprende la im portancia de los análisis citogenét icos ( tanto
m itót icos com o m eiót icos) en los estudios taxonóm icos y evolut ivos.
1 .2 Car iosistem át ica
La Cariosistem át ica, entendida com o el estudio de las relaciones
naturales de los organismos a part ir de la inform ación brindada por los
crom osom as, perm ite la comparación detallada de los cariot ipos de especies
relacionadas y la ident ificación precisa de los cromosom as enteros o de los
segm entos crom osóm icos que part icipan en los reordenam ientos que
pueden tener un posible com prom iso con la diferenciación de las especies.
Los datos crom osómicos pueden ser usados com o m arcadores
filogenét icos ya que se heredan com o caracteres m endelianos y están
conservados dent ro de las especies, y las m utaciones que se fij an en el
car iot ipo de una especie están consideradas com o eventos “ raros” (Sankoff
2003; Dobigny et al. 2004; Stanyon et al. 2008) . Siguiendo el cr iter io de Máxima
Parsimonia, las comparaciones citogenét icas ent re taxa relacionados
perm iten ident ificar form as cromosómicas com part idas por ancest ralidad
com ún (sinaporm orfías) (Olsen et al. 1996; Dobigny et al. 2004) .
Es así com o los pat rones crom osóm icos de coloración diferencial
conocidos com o bandas G, C, ent re ot ros, ident if ican regiones part iculares
del genom a revelando aspectos de la organización est ructural, incluyendo la
caracterización de rearreglos crom osómicos con posible com prom iso
especiogénico.

1. I nt roducción
3
El pat rón de bandas G revela una sucesión de bandas longitudinales
claras (G negat ivas) y oscuras (G posit ivas) a lo largo del crom osom a que
pueden variar en ancho e intensidad y son característ icas de cada par de
crom osom as hom ólogos. La com paración de los pat rones de bandas G ent re
los cariot ipos de diferentes especies se puede ut ilizar para definir
homologías primarias a nivel cromosóm ico, ident ificar sinapomorfías y
relaciones de grupos herm anos ent re los taxa (Dobigny et al. 2004) . De esta
form a los cromosom as, analizables y com parables por tam año, m orfología y
comportam iento m itót ico o m eiót ico, colaboran en resolver dudas
taxonóm icas y ancest ralidades com o ot ro t ipo de datos o caracteres en un
análisis filogenét ico.
1 .2 .1 Crom osom as sexuales, especiación y car iosi stem át ica
Dos reglas em píricas sugieren un papel part icular de los crom osomas
sexuales en el surgim iento de barreras de aislam iento reproduct ivo
postcigót ico: la regla de Haldane y el “gran efecto del X” ( “ large X-effect ” ) .
La regla de Haldane (Haldane 1922) postula que en un cruce
interespecífico, si hay un sexo inviable o estér il en la descendencia híbrida,
éste será el sexo heterogam ét ico. Este fenóm eno se observa en todas las
especies que poseen cromosom as sexuales, incluyendo invertebrados, aves
y m am íferos (Haldane 1922; Laurie 1997; Orr 1997; Coyne y Orr 2004) . Las
explicaciones para este fenóm eno son diversas y todas involucran la
acum ulación diferencial de factores recesivos de esterilidad e inviabilidad de
los híbr idos (Graves y O'Neill 1997; Orr 1997; Presgraves 2008) .
La regla del “gran efecto del X” ( también llamada la “ regla de
Coyne” ) se refiere al efecto desproporcionado del cromosom a X versus los
autosom as en la ester ilidad e inviabilidad de los híbridos observadas en los
experim entos genét icos de ret rocruzas en Drosophila (Coyne y Orr 1989) . Este
efecto se debería a una alta densidad de factores de esterilidad en el
crom osom a X (o en el cromosom a Z) o a una m ayor expresión de factores
recesivos ligados al X (o al Z) . Este fenóm eno es observado en dípteros,
lepidópteros, aves y en el ratón (Forej t 1996; Laurie 1997; Presgraves 2008) .

1. I nt roducción
4
De lo anter iorm ente referido se desprende la im portancia de la
caracterización de los m ecanism os de determ inación cromosóm ica del sexo
en los estudios cariosistem át icos.
1 .2 .2 Mecanism os de determ inación sexual
Dentro de la determ inación sexual prim aria se observan en la
naturaleza diversos m ecanism os (Bull 1983; Charlesworth y Mank 2010) .
Se denom ina Determ inación Genét ica del Sexo (GSD, sigla de “Genet ic
Sex Determ inat ion” ) al m ecanismo m ediante el cual alelos de un locus o t ipos
alternat ivos de un grupo de ligam iento (por ejem plo cromosom as Y y W)
influencian la probabilidad del em brión de desarrollarse como m acho o
hembra. El sexo del em brión se puede predecir por el genot ipo del cigoto
(Charlesworth y Mank 2010) . Dent ro de la GSD encont ram os:
-Heterogam ia: sistem as de crom osom as sexuales o gonosom as en
donde m achos (heterogam ia m asculina) o hem bras (heterogam ia fem enina)
t ienen crom osom as heterom órficos, es decir en los que se observan
diferencias m orfológicas en un par cromosóm ico en el sexo heterogam ét ico.
Los crom osom as sexuales heterom órficos son la base hereditar ia de la
determ inación sexual, la cual es determ inada en la concepción (Charlesworth y
Mank 2010) . Dentro de los casos de heterogam ia m asculina encont ramos el
sistem a de Balance X-A (X-autosom a) en la que la razón ent re el núm ero de
crom osom as X y crom osom as autosóm icos determ ina el sexo m ediante un
sistem a de conteo de los crom osom as X. En este sistem a, el crom osom a Y
es dispensable (Bull 1983) . Ot ro caso incluido dent ro de heterogam ia
m asculina es el sistem a “Y dom inante” . En este sistem a las hem bas son XO
y los m achos XXY. El sexo es determ inado por la presencia del crom osoma
Y, independientem ente de la presencia de uno o dos crom osom as X (Bull
1983) .
-Determ inación del sexo Polifactor ial (Poligénica) : el sexo se
determ ina en el cigoto por la acción de m uchos factores con efectos

1. I nt roducción
5
individuales pequeños, pudiendo influir tam bién el am biente. El efecto
acum ulado de estos factores cont rola la determ inación sexual (Bull 1983) .
-Arrenotoquia (Haplo-diploidía) : sistem a genét ico en el cual los
m achos surgen de huevos no fert ilizados y las hembras de huevos
fert ilizados. La determ inación sexual puede ser ambiental (basada en la
fert ilización) o puede ser genot ípica (basada en el nivel de ploidía u ot ro
factor) (Bull 1983) .
Por ot ro lado, se define como Determ inación del sexo Am biental (ESD,
sigla de “Enviromental Sex Determ inat ion” ) cuando el sexo se determ ina en la
em briogénesis en respuesta al am biente local, con ciertas condiciones
produce hem bras y con ot ras condiciones produce m achos. En este caso el
sexo del individuo no se puede predecir por el genot ipo del cigoto (Bull 1983) .
A cont inuación se describirán brevem ente algunos aspectos de la
determ inación del sexo en organismos superiores.
1 .2 .2 .1 Plantas
Los sistem as de determ inación sexual en las plantas son diversos,
incluyendo m ecanismos que involucran crom osom as sexuales,
determ inantes genét icos, horm onas e influencia ambiental, así com o una
com binación de varios de estos factores. Sin em bargo son los factores
genét icos los que t ienen el papel principal en la determ inación sexual. En
las especies estudiadas hasta la fecha de la redacción de este t rabajo de
Tesis se han descr ito sistem as de determ inación del sexo de t ipo
heterogam ia m asculina, heterogam ia fem enina y balance X-A (Charlesworth
2002; Vyskot y Hobza 2004; Ming et al. 2007; Charlesworth y Mank 2010) .
En algunas plantas el sexo es determ inado por la segregación de un
único locus mendeliano com o en el pepino Ecballium elater ium , o la
segregación de varios loci com o en la ort iga anual Mercurialis annua, sin
observar diferenciación m orfológica de un par crom osóm ico en ninguno de
los dos sexos. En este caso se los denom ina “ crom osom as sexuales

1. I nt roducción
6
hom om órficos” , siendo el caso observable en la gran m ayoría de las plantas
dioicas (Vyskot y Hobza 2004; Ming et al. 2007)
Las especies en las que se ha descr ito la presencia de crom osomas
sexuales heterom órficos son pocas: las hepát icas Marchant ia polymorpha y
Sphaerocarpus donnelli, la papaya o m amón (Carica papaya) , las Collejas
Blanca y Roja (Silene lat ifolia y S. dioica respect ivam ente) , el cáñam o
Cannabis sat iva, el espárrago (Asparagus off icianalis) , el lúpulo (Humulus
lupulus y H. japonicus) , las vinagreras (Rum ex acetosa y R. acetosella) y en
el alga Sacchoriza polyschides (Charlesworth 2002; Navajas-Pérez et al. 2005; Yu et
al. 2008; Howell y Filatov 2009; Charlesworth y Mank 2010) .
La papaya es una angiosperm a polígam a que posee form as
m asculinas, fem eninas y hermafroditas. Esta especie t iene un núm ero
diploide 2n= 18 y cromosom as sexuales homomórficos. Sin em bargo, un
locus único M con 3 alelos (M1, M2 y m ) t iene un papel importante en la
determ inación sexual. Los m achos y hermafroditas son heterocigotos M1m y
M2m respect ivam ente, m ient ras que la combinación recesiva m m da origen
a hem bras. La com binación M1M2 es letal em brionaria (Yu et al. 2008) . Este
locus fue mapeado en una corta región no- recom binante de los cromosom as
sexuales. Dado que los m achos son heterocigotos y las hem bras
homocigotas se considera que la determ inación sexual en la papaya sería de
t ipo proto-XY (Y dom inante) con una incipiente diferenciación ent re los
crom osom as sexuales, uno de los pr im eros pasos para la form ación de
crom osom as sexuales heterom órficos.
En la vinagrera (Rum ex acetosa) y en el lúpulo (Hum ulus j aponicus)
se produjeron m odificaciones del sistem a XY, denom inadas “ sistem as
crom osóm icos de determ inación sexual m últ iple” , que surgen cuando uno
de los gonosom as sufre una t ranslocación hacia un autosom a o cuando uno
de los gonosom as sufre una fisión dando lugar a m ás de un crom osom a. En
estos taxa se observa un sistem a crom osómico de determ inación sexual
m últ iple XY1Y2 o de t ipo t r ivalente (se lo llama así dado que en Metafase I
form a una cadena form ada por t res elem entos) . Este sistem a podría haber
surgido por t ranslocaciones X-autosom a o por fisión del crom osom a Y. Este
1. I nt roducción
7
sistem a de determ inación sexual a su vez se encuent ra asociado con un
sistem a de determ inación del sexo por balance X/ A (Navajas-Pérez et al. 2005;
Charlesworth y Mank 2010) . En H. lupulus var. cordiflorus (Navajas-Pérez et al. 2005)
y en la colleja Silene diclinis (Howell et al. 2009) se han descrito sistem as
sexuales crom osóm icos X1X2Y1Y2 o sistem a sexual m últ iple de t ipo
cuadrivalente (pues form an una cadena de 4 elem entos en Metafase I ) , que
habrían surgido de una t ranslocación recíproca ent re el crom osoma Y y un
par autosóm ico a part ir de un sistem a XY1Y2.
1 .2 .2 .2 I nver tebrados
En invertebrados encont ram os una alta diversidad de m ecanism os de
determ inación del sexo, incluyendo tanto determ inación genét ica del sexo
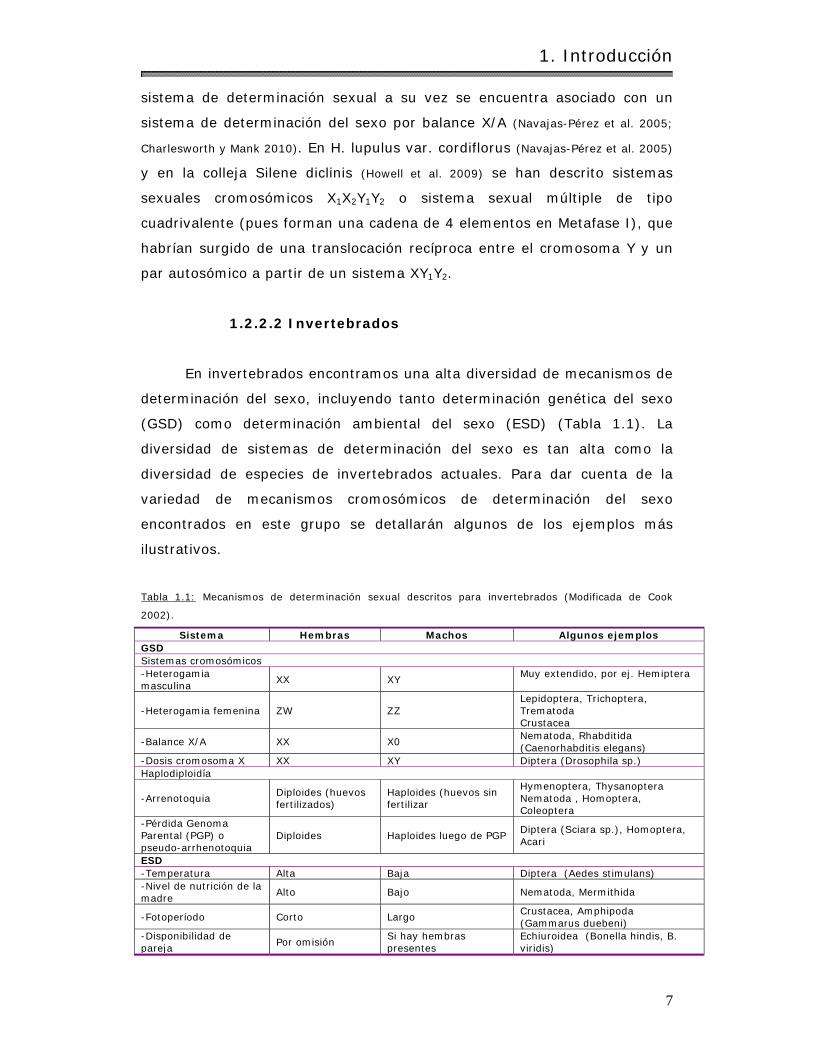
(GSD) com o determ inación am biental del sexo (ESD) (Tabla 1.1) . La
diversidad de sistem as de determ inación del sexo es tan alta com o la
diversidad de especies de invertebrados actuales. Para dar cuenta de la
var iedad de m ecanismos cromosóm icos de determ inación del sexo
encont rados en este grupo se detallarán algunos de los ejemplos más
ilust rat ivos.
Tabla 1.1: Mecanism os de determ inación sexual descritos para invertebrados (Modificada de Cook
2002) .
Sistem a Hem bras Machos Algunos e jem plos GSD Sistem as crom osóm icos -Heterogam ia m asculina
XX XY Muy extendido, por ej . Hem iptera
-Heterogam ia fem enina ZW ZZ Lepidoptera, Tr ichoptera, Trem atoda Crustacea
-Balance X/ A XX X0 Nem atoda, Rhabdit ida (Caenorhabdit is elegans)
-Dosis crom osom a X XX XY Diptera (Drosophila sp.) Haplodiploidía
-Arrenotoquia Diploides (huevos fert ilizados)
Haploides (huevos sin fert ilizar
Hym enoptera, Thysanoptera Nem atoda , Hom optera, Coleoptera
-Pérdida Genom a Parental (PGP) o pseudo-arrhenotoquia
Diploides Haploides luego de PGP Diptera (Sciara sp.) , Hom optera, Acar i
ESD -Tem peratura Alta Baja Diptera (Aedes st im ulans) -Nivel de nut r ición de la m adre
Alto Bajo Nematoda, Merm ithida
-Fotoperíodo Corto Largo Crustacea, Am phipoda (Gam m arus duebeni)
-Disponibilidad de pareja
Por om isión Si hay hem bras presentes
Echiuroidea (Bonella hindis, B. v ir idis)

1. I nt roducción
8
Un género muy estudiado en insectos es Drosophila. Hasta hace
algunos años se consideraba que en Drosophila la determ inación del sexo
ocurría por un balance X-autosoma. Una razón 2X/ 2A daba lugar a las
hembras y una razón X/ 2A daba lugar a los m achos. Recientem ente se ha
confirmado que la señal X/ A predice el dest ino sexual, si bien no la
especifica (Erickson y Quintero 2007) . El número de cromosom as X m ás que el
balance X/ A es lo que da la señal para la determ inación sexual. Hoy se sabe
que la señal prim aria es de t ipo cuant itat iva: es la cant idad de factores de
t ranscripción codificados por el crom osom a X los que determ inan si el gen
Sex- letal (Sxl) se t ranscribe tem pranam ente act ivando el prom otor
tem prano para este gen durante el estadio de blastoderm o, dando lugar a
una cascada regulator ia que lleva al desarrollo de un individuo fem enino.
Una sola copia del cromosom a X no posee la cant idad necesar ia de factores
de t ranscripción para act ivar este prom otor tem prano del gen Sxl, por lo
que la cascada regulator ia consiguiente da lugar al desarrollo de un m acho.
Para m ás inform ación sobre este m ecanism o de determ inación del sexo
consultar Sanchez (2008) y Verhulst et al. (2010) .
El crom osom a Y en Drosophila es pequeño y com pletam ente
heterocrom át ico, consist iendo m ayoritar iam ente de ADN repet it ivo y
elem entos t ransponibles. Cont iene alrededor de 20 genes, que en los
m achos codifican para funciones relacionadas con la fert ilidad, tales como la
m ot ilidad esperm át ica ( los machos X0 son estériles) (Carvalho et al. 2009) . En
este género, estudios recientes indicaron que no existe hom ología de
secuencia ent re el cromosoma X y el cromosom a Y. Actualm ente se
postulan varias hipótesis para explicar esta falta de hom ología: 1) que los
genes or iginales del Y ancest ral se perdieron y los genes que se encuent ran
hoy en el Y son adiciones subsecuentes dada la ventaja select iva para los
m achos; 2) que este cromosom a Y actual habría sido derivado de un
crom osom a B (Koerich et al. 2008; Carvalho et al. 2009; Kaiser y Bacht rog 2010).
Por ot ro lado, algunas especies de este género han adquir ido
crom osom as sexuales secundarios ( los neo-cromosom as sexuales) , los

1. I nt roducción
9
cuales se form arían por una fusión ent re un autosom a y un crom osoma
sexual (Yi y Charlesworth 2000; Kaiser y Bacht rog 2010) .
Ot ro grupo interesante lo const ituyen los lepidópteros. Los taxa m ás
basales de Lepidoptera y las especies del orden herm ano Tr ichoptera
poseen un sistem a de determ inación sexual Z0/ ZZ, lo que sugerir ía que
éste sería el estado ancest ral para esos grupos. En Lepidoptera una fusión
ent re el crom osom a Z y un autosom a habría dado lugar al crom osom a W
(que sería en realidad un neo-W) , que se encuent ra en 98% de las
especies. Tam bién se ha postulado que un crom osoma B podría haber
tom ado el rol de cromosom a W ( revisado en Traut et al. 2007). En algunos
linajes el W se perdió secundariam ente (creando sistem as Z0/ ZZ) o se
fusionó con un autosom a (creando sistem as W1W2Z/ ZZ) . En la polilla
saturnida Sam ia cynthia fusiones del Z con autosomas dieron lugar a
dist intos sistem as, encont rándose sistem as sexuales Z0/ ZZ, WZ/ ZZ y
WZ1Z2/ Z1Z1Z2Z2 en diferentes subespecies del género (Yoshido et al. 2006) .
Todas las especies del orden Hemiptera t ienen crom osom as
holocinét icos (crom osom as sin const r icción prim aria y, por ende, sin
cent róm ero localizado) y los cromosom as sexuales son aquiasm át icos en los
m achos (es decir , no recom binan) (Mola y Papeschi 2006) . En este orden se han
descrito sistem as XX/ XY, XX/ X0, sistem as múlt iples XnXn/ XnY, XnXn/ Xn0 y
XX/ XYn (or iginados por la fragm entación del X ancest ral) y sistem as con
neo-crom osom as sexuales originados por la fusión del X e Y ancest rales con
un par autosóm ico ( revisado en Papeschi y Bressa 2006).
Esta diversidad de sistem as sexuales cromosómicos en invertebrados
perm ite el estudio comparat ivo de crom osom as sexuales en dist intos
estadíos de su t ransición evolut iva desde autosom as hasta crom osom as
sexuales heterom órficos.

1. I nt roducción
10
1 .2 .2 .3 Vertebrados
Los Vertebrados, al igual que los grupos anteriorm ente refer idos,
exhiben una amplia var iedad de m ecanismos de determ inación sexual. Al
relacionar estos m ecanism os en un árbol filogenét ico (Figura 1.1) se hace
evidente la “ labilidad evolut iva” de estos m ecanism os, en la que num erosas
t ransiciones debieron haber ocurr ido para alcanzar la diversidad actual
(Janzen y Phillips 2006; Barske y Capel 2008) .
Figura 1.1: Determ inación del sexo en vertebrados actuales (Modificada de Barske y Capel
2008) . Los peces, anfibios, tortugas y lagartos exhiben más de un sistema de determ inación sexual,
incluyendo tanto GSD com o ESD. XX/ XY: Heterogam ia masculina; ZZ/ ZW: heterogam ia fem enina; Hom om órficos
refiere a crom osomas sexuales no diferenciados, Poligénica y dependiente de la densidad son los característ icos de
peces; TSD es un subt ipo dent ro de ESD, en la que el sexo se determ ina por la temperatura de incubación de los
huevos. A excepción de los mam íferos euter ios, la m ayoría de los em briones de vertebrados son suscept ibles a
reversiones del sexo inducidas por hormonas exógenas. Otros: el asterisco ( * ) indica la co-ocurrencia de TSD y GSD en
algunas especies de lagartos; el círculo ( ◦) indica el sistem a sexual complejo que presentan los monotremas (ver más
adelante) que form a una cadena en la meiosis masculina. El signo de pregunta (?) indica los casos en los que no se
t iene inform ación para ese taxón.
En peces y en anfibios, GSD es el m ecanismo de determ inación
sexual m ás extendido. En estos taxa la presencia de cromosom as sexuales

1. I nt roducción
11
heterom órficos ha sido descrita sólo en un reducido núm ero de especies
(Devlin y Nagaham a 2002; Nakam ura 2009) .
Especies cercanas o incluso poblaciones dent ro de una m ism a especie
pueden tener diferentes m ecanism os de determ inación del sexo o sus
crom osom as sexuales no presentar hom ologías. En peces, las fam ilias
Cichlidae (Perciform es) , Gasterosteidae (Gasterosteiform es) y en los peces
Medaka del género Oryzias (Cyprinodont idae) se ha descrito la presencia
tanto de sistem as crom osóm icos de determ inación sexual XX/ XY como
ZW/ ZZ (Takehana et al. 2007; Ross y Peichel 2008) . En anfibios, en la especie Rana
rugosa, las poblaciones de Japón difieren en su sistem a de determ inación
sexual según la región geográfica en la que habitan. Las poblaciones en el
noroeste presentan heterogam ia fem enina (ZW/ ZZ) y las que habitan en el
oeste poseen determ inación del sexo XX/ XY (Ogata et al. 2003) , siendo que el
par ZW de la población heterogam ét ica fem enina hom ólogo al par XY de la
población heterogam ét ica m asculina (Uno et al. 2008) .
En rept iles, la determ inación del sexo dependiente de la tem peratura
es la m ás extendida y se ha sugerido que const ituye el estado ancest ral a
part ir del cual la determ inación genét ica del sexo ZW/ ZZ y XX/ XY ha
surgido m ás de una vez de form a independiente (Janzen y Phillips 2006) .
Recientem ente se ha dem ostrado que en algunas especies de rept iles que
presentan GSD, la tem peratura de incubación de los huevos puede revert ir
la determ inación genét ica del sexo del em brión (Radder et al. 2008) , indicando
que la separación ent re TSD y GSD no sería tan m arcada com o se sostenía
t radicionalm ente, sino que representarían puntos de un cont inuo a lo largo
del cual las poblaciones pueden cam biar bajo presión select iva (Barske y Capel
2008) .
Las aves presentan heterogam ia fem enina ZW/ ZZ. El crom osom a Z
está conservado en todas las especies de aves. El cromosom a W presenta
un pat rón evolut ivo diferencial desde altam ente conservado en las aves
rat ites no voladoras, hasta altam ente diferenciado entre las aves carinadas
(Pigozzi y Solar i 1999, 2000, 2005; Ezaz et al. 2006).

1. I nt roducción
12
Las serpientes (Rept ilia) tam bién poseen heterogam ia fem enina.
Como en las aves, el cromosom a Z está altam ente conservado, m ient ras
que el cromosom a W muest ra dist intos grados de degeneración (Ezaz et al. ,
2006) . Morfológicam ente el par ZW de las aves es sim ilar al de las serpientes
y m ediante estudios citomoleculares se determ inó que estos crom osom as
no poseen homología genét ica (Matsubara et al. 2006; Kawai et al. 2007) .
Todos los m am íferos presentan heterogam ia m asculina XX/ XY
( revisado en Solar i 1993) .
Los m onot rem as (Prototheria) presentan un sistem a de
determ inación sexual m últ iple que involucra más de 4 elem entos,
característ ica única ent re los m am íferos (Wallis et al. 2008) . Los m achos de
ornitorr inco poseen un sistem a de determ inación del sexo
X1X2X3X4X5Y1Y2Y3Y4Y5, que forman en Metafase I una cadena de 10
elem entos (decavalente) . Estos cromosomas segregan en la m eiosis
m asculina de m anera alternada produciendo gam etas balanceadas,
generando esperm atozoides portadores de los 5 cromosom as Y y ot ros
portadores de los 5 crom osomas X. En las t res especies de equidnas el
sistem a de determ inación sexual crom osóm ico m últ iple en m achos está
form ado por 5 crom osom as X y 4 cromosom as Y (X1X2X3X4X5Y1Y2Y3Y4) ,
const ituyendo una cadena de 9 elem entos en Metafase I m asculina
(nonavalente) . Los sistem as de determ inación sexual del ornitorr inco y los
equidnas habrían surgido por t ranslocaciones recíprocas sucesivas ent re los
crom osom as sexuales ancest rales y autosom as (Rens et al. 2007) . Las cadenas
del ornitorr inco y los equidnas poseen elem entos hom ólogos y han
incorporado un autosom a diferente en cada taxón de m anera
independiente. Los cromosom as Y3, X4 e Y4q del ornitorr inco corresponden
al crom osoma 27 de los equidnas y los crom osom as Y4q y X5q del equidna
corresponden al crom osom a 11 del ornitorr inco (Rens et al. 2007) .
En los marsupiales (Metatheria) el sistem a de determ inación sexual
m ás extendido es el XX/ XY. Sin em bargo, el crom osom a Y es generalm ente
m uy pequeño y su influencia en la determ inación del sexo no es total.
Mient ras que los hum anos XXY son fenot ípicam ente m achos (estériles) , los

1. I nt roducción
13
canguros XXY m uest ran característ icas tanto de m acho com o de hem bra.
Los crom osom as X e Y no se aparean en m eiosis (no existe una región PAR
o pseudoautosóm ica) . El X tam bién es m ás pequeño en com paración con el
de los euter ios, com prendiendo el 3-4% del complem ento haploide (Graves y
Westerm an 2002) . Algunas especies poseen cromosom as X de mayor tam año
debido a la adquisición de cant idades variables de heterocrom at ina (Solar i et
al. 1993) . En m arsupiales se han observado sistem as de determ inación
sexual m últ iple en t res especies, el potoro de nariz larga (Potorous
t r idactylus) , el walabí de pantano (Wallabia bicolor) y el bilbi mayor
(Macrot is lagotes) , las cuales poseen un sistem a de t ipo XX/ XY1Y2 or iginado
a part ir de una t ranslocación recíproca ent re un autosom a y el crom osoma
X or iginal. El autosom a homólogo (ahora denom inado Y2) pasa a ser parte
del t r ivalente que se form a en la m eiosis masculina (Sharp 1982) . En el walabí
de anteojos (Lagorchestes conspicillatus) se ha descrito un sistem a de
intercambio com plejo denom inado t (XA1) , t (A2YA1) , A2. Este sistem a se
habría originado a part ir de t res intercambios ent re los X e Y originales y
dos pares autosóm icos (A1 y A2) ( revisado en Solar i et al. 1993).
Al igual que en m arsupiales, en m am íferos placentar ios (Eutheria) , el
sistem a de determ inación sexual cromosómico más extendido es el XX/ XY.
El crom osom a X está altam ente conservado en m amíferos euterios (Ohno
1977; Graves y Westerm an 2002) , m ient ras que el cromosom a Y m uest ra un
am plio espect ro de divergencia m orfológica y génica (Solar i 1993; Graves 2006) .
Tanto el crom osom a Y com o el crom osom a X están involucrados en
m uchos casos en la form ación de sistem as de determ inación sexual
m últ iple, aunque las t ranslocaciones X-autosom a son m enos frecuentes que
las Y-autosom a debido a que si durante el proceso de com pensación de
dosis la inact ivación del crom osoma X se expande hasta cubrir la porción
autosóm ica t ranslocada puede resultar en consecuencias deletéreas (Dobigny
et al. 2004) . Aún así, t ranslocaciones X-autosom a se han caracterizado en
algunos roedores, bóvidos y quirópteros en los que la amplificación de
secuencias heterocrom át icas ent re las secuencias del X y el autosom a,
previene la expansión de la inact ivación hacia las zonas autosóm icas (Ohno
1977; Pathak y Lin 1981; Viegas-Péquignot et al. 1982; Kasahara y Dut r illaux 1983; Solar i y

1. I nt roducción
14
Pigozzi 1994; Kingswood et al. 1994; Dobigny et al. 2004; Rodrigues Norohna et al. 2004;
Veyrunes et al. 2007) .
Ot ras m odificaciones al sistem a XX/ XY que involucran al crom osom a
X son las deleciones que se observan en los lemm ings Myopus schist icolor y
Dicrostonyx torquatus torquatus y en el ratón de cam po Akodon azarae.
Esto genera en esas especies dos t ipos de hem bras, las XX y las Xx, estas
últ im as portadoras del crom osoma X delecionado. Los m achos son siem pre
XY, ya que los m achos xY son inviables (Bianchi y Cont reras 1967; revisado en
Solar i 1993) .
Ot ra m odificación interesante al sistem a XX/ XY es la que se observa
en el cr icét ido Ellobius lutescens y en el múrido Tokudaia osim ensis. Estas
especies presentan un sistem a XX/ X0, dado que carecen de crom osom a Y
(Just et al. 2002; Sutou et al. 2001; Graves 2008) . Se postula que el cromosom a Y
podría haberse t ranslocado a un autosom a o al cromosom a X, o que ot ros
genes autosóm icos habrían tomado las funciones de los genes con función
específica en m achos que contenía el Y (Just et al. 2002; Sutou et al. 2001; Graves
2008) .
El cromosom a Y de los euter ios tam bién se encuent ra involucrado en
t ranslocaciones Y-autosom a generando sistem as de determ inación sexual
m últ iple, tales cóm o los observados en quirópteros, roedores (Revisado en
Fredga 1970; Solari 1993; Deuve et al. 2007) y en algunos primates.
1 .2 .2 .3 .1 Pr im ates
En el orden Prim ates, uno de cuyos géneros es objeto de estudio de
este t rabajo de Tesis, el sistem a de determ inación sexual m ás extendido es
el XX/ XY. Com o m odificaciones a este sistem a ancest ral encont ram os
t ranslocaciones Y-autosom a generando sistem as de determ inación sexual
m últ iple. Estos sistem as sexuales m últ iples se han ver ificado en diversos
géneros de Primates Neot ropicales (Platyrrhini) (Ma et al. 1975; Armada et al.
1987; Seuánez et al. 1989; Lim a y Seuánez 1991; Rahn et al. 1996; Mudry et al. 1998,
2001; Moura-Pensin et al. 2001; Solar i y Rahn 2005) , m ient ras que en Prim ates del

1. I nt roducción
15
Viejo Mundo (Catarrhini) , de las revisiones bibliográficas surge que, hasta el
presente, se ha descrito la presencia de sistem as de determ inación sexual
m últ iple en una única especie, Presbyt is cr istata (Bigoni et al. 1997) .
Dada esta diversidad, el pat rón de determ inación sexual const ituir ía
un carácter con valor sistem át ico en los Prim ates y en part icular en la
superfam ilia Ceboidea (Platyrrhini) . La caracterización de estos sistem as se
realiza m ediante el análisis cariot ípico de las dist intas especies de prim ates.
Tom ando en consideración que el m antenim iento de la diversidad genét ica
dent ro de las especies es dependiente de la reproducción, la caracterización
cariot ípica perm ite una correcta t ipificación, ident ificando las diferencias
cariot ípicas ent re las dist intas especies y los reordenam ientos cromosóm icos
con posible protagonism o en la especiogénesis. Con este fin, los parám et ros
m itót icos han sido am pliam ente ut ilizados en décadas pasadas. Sin
embargo, los datos m eiót icos son los que perm iten confirm ar la cariología e
ident ificar inequívocam ente el sistem a de determ inación sexual.
La caracterización m eiót ica en Primates Neotropicales no sólo es de
gran ut ilidad a la hora de proponer planes de conservación o de
reproducción en caut iver io, sino que aporta inform ación út il para
interpretaciones evolut ivas, cont r ibuyendo a su vez a resolver dudas
taxonóm icas. Sin em bargo, este t ipo de estudios son notablem ente escasos
y en algunas especies, nulos (Egozcue 1969; Ma et al. 1976; Seuánez et al. 1983;
Arm ada et al. 1987; Lim a y Seuánez 1991; Rahn et al. 1996; Mudry et al. 1998, 2001;
Nieves et al. 2005; Steinberg et al. 2007) . El car iot ipo m eiót ico se ha descrito hasta
la fecha de este t rabajo de Tesis en sólo un reducido núm ero de especies de
Platyrrhini, confirm ando la determ inación sexual XX/ XY en Cebus
paraguayanus (Seuánez et al. 1983; Mudry et al. 2001) , Ateles geoffroyi y A.
paniscus (Mudry et al. 2001; Nieves et al. 2005) y Saim ir i boliviensis boliviensis
(Egozcue 1969; Steinberg et al. 2007) . Diferentes sistem as de determ inación
sexual m últ iple se han descrito en ot ras especies, tales com o Alouat ta
caraya (Rahn et al. 1996; Mudry et al. 1998, 2001) , A. seniculus (Lim a y Seuánez
1991) , A. belzebul (Arm ada et al. 1987) , A. palliat ta (Solar i y Rahn 2005) , Aotus
azarae (Ma et al. 1976) , Callim ico sp. (Seuánez et al. 1989) y Cacajao sp.
(Moura-Pensin et al. 2001) .

1. I nt roducción
16
Los pr im ates no humanos representan el m odelo de m am ífero m ás
cercano a Hom o sapiens, const ituyendo uno de los m odelos exper im entales
m ás ut ilizados, después de los roedores. En este contexto el estudio del
comportam iento m eiót ico en pr im ates no hum anos es de ut ilidad en el
estudio de las característ icas conservadas ent re las especies de prim ates,
como los procesos de sinapsis y recombinación.
Durante la profase I de la m eiosis, los crom osom as homólogos
aparean, sinapsan y recom binan. El proceso de sinapsis comprende la
form ación del com plejo sinaptoném ico (CS) , una est ructura proteica
t r ipart ita form ada por dos elem entos laterales (EL) de 60 nm de ancho y un
espacio cent ral de alrededor de 100 nm . La región cent ral está at ravesada
por los filam entos t ransversales, sobre los cuales se deposita el elem ento
cent ral (Solar i 2004) . Es sobre esta est ructura que se depositan las proteínas
involucradas en el proceso de recombinación. Para la especie hum ana se
han caracterizado una batería de proteínas involucradas en la form ación del
CS y en el proceso de recom binación (Moens et al. 2002; Baker et al. 1996; Moses
1968; Turner et al. 2005; Garcia-Cruz et al. 2009) :
-MLH1, una proteína involucrada en el sistem a de reparación de
apaream ientos erróneos cuyos foci (sit ios dónde se detecta la señal
de hibr idación en los crom osom as) m arcan los nódulos de
recombinación tardíos, que son equivalentes a detectar y contabilizar
quiasm as en Diacinesis o Metafases I .
-SYCP3, com ponente de los elem entos laterales.
-SYCP1, com ponente de los filam entos t ransversales.
-REC8, com ponente del elem ento lateral.
-SMC1 y SMC3, cohesinas presentes tanto en m eiosis com o en
m itosis.
-BRCA1, com ponente de la m aquinar ia de “ checkpoint ” celular.
Ant icuerpos cont ra estas proteínas perm iten la visualización del
com plejo sinaptoném ico em pleando m icroscopía ópt ica de fluorescencia.
Estos estudios perm iten analizar a su vez la dinám ica crom osóm ica durante

1. I nt roducción
17
los estadíos tem pranos de la profase, perm it iendo estudiar el proceso de
sinapsis y recom binación de un individuo y dist intos m iem bros de una dada
especie.
La m ayoría de los estudios realizados en vertebrados hasta la fecha
se desarrollaron en humano, ratón o aves (Lynn et al. 2002; Hassold et al. 2004;
Codina-Pascual et al. 2005, 2006; Calderón y Pigozzi 2006) . Sin em bargo, existe una
vacancia de estos estudios en m am íferos, más aún de los relacionados con
el estudio de la dinám ica de los cromosom as sexuales en m eiosis. Lo dicho,
realza la im portancia de estos estudios para aportar nuevas evidencias
respecto a las diferencias tanto en est ructura com o en el com portam iento
de estos cromosom as, cont r ibuyendo a esclarecer el pat rón de evolución de
los sistem as de determ inación sexual en vertebrados.
1 .2 .3 Sintenias crom osóm icas
En los últ imos 20 años la aplicación de la técnica de Hibr idación in
situ Fluorescente (FI SH, “Fluorescent I n Situ Hybridizat ion” ) con sondas
específicas para crom osom as enteros (pintado crom osómico o “ chrom osom e
paint ing” ) ha probado ser un m étodo rápido y confiable para establecer
hom eologías (entendidas com o reconocim iento de las parejas de
crom osom as homólogos ent re organismos diferentes, válidas para el
análisis filogenét ico) ent re diferentes taxa y en part icular, dent ro del Orden
Primates (Wienberg et al. 1990; Consigliere et al. 1996, 1998; Morescalchi et al. 1997; de
Oliveira et al. 2002; Stanyon et al. 2004, 2008; Dum as et al. 2007; Am aral et al. 2008) .
Gracias a su precisión, esta últ im a técnica se ut ilizó sat isfactor iam ente en
sistem át ica para verificar las homologías y hom eologías hipotet izadas
previam ente en base a la morfología crom osóm ica y técnicas de t inción
diferencial por bandas cromosóm icas (Steinem ann et al. 1984) . Las hom eologías
realizadas m ediante pat rones de bandas G, si bien son de ut ilidad para
definir homologías primarias dent ro y ent re especies, ven su eficacia
reducida en el caso de los cariot ipos m uy reordenados, es decir con
num erosos rearreglos crom osóm icos (Robinson 2001; Schneider et al. 2001) . La
técnica de FI SH br inda la confirm ación inequívoca de las hom eologías
previam ente descr itas por bandas G, perm it iendo una m ayor definición a

1. I nt roducción
18
nivel m olecular para su interpretación (Wienberg et al. 1990; Wienberg 2004, 2005;
Muller 2006; Stanyon et al. 2008) .
El pintado crom osóm ico comparado en m amíferos perm it ió
determ inar un alto grado de conservación genóm ica en este taxón. Sólo
unos pocos cam bios fueron fijados en algunos taxa, tales com o fisiones,
fusiones y t ranslocaciones de brazos crom osómicos, los cuales son
considerados com o “ rearreglos m arcadores” ( “ landm ark rearrangem ents” )
út iles com o caracteres para tener en cuenta en los análisis filogenét icos
(Wienberg et al. 1990; O'Brien et al. 1999; Wienberg 2004, 2005; Müller 2006; Stanyon et
al. 2004, 2008) .
Si tenem os en cuenta que cada crom osom a reúne un cúm ulo de
inform ación part icular, que la relación ent re la citogenét ica y genét ica
m olecular han perm it ido ident if icar parcial o totalm ente (Wienberg y Stanyon
1995; Lander et al. 2001; Venter et al. 2001; Mikkelsen et al. 2005; Wienberg 2005), se
puede considerar que hay ciertos conceptos que am bas disciplinas
com parten ínt im am ente. Así, en genét ica, se define com o loci sinténicos a
dos loci que están ligados en un m ismo crom osom a (Renwick 1971; Rieger et al.
1991) . Esta definición puede ser ampliada y aplicada a nivel cromosómico,
surgiendo así el concepto de segm entos crom osóm icos sinténicos. Estas
regiones se pueden localizar com o una est ructura en un m ism o crom osoma,
que se m ant ienen en el genom a de diferentes especies, perm it iendo
ident ificar y analizar las hom eologías interespecíficas. Para ello, en el
análisis de conservación genóm ica, se aplica la hibridación cruzada
ut ilizando sondas de crom osom as hum anos y/ o de las especies de interés
part icular (Wienberg y Stanyon 1995, 1997; Müller et al. 2000; Wienberg 2005) .
Una sintenia crom osóm ica “ rota” se refiere a una región crom osómica
que en un taxón se encuent ra en un m ismo cromosom a y que en ot ro taxón
se localiza en cromosom as diferentes, es decir, ese segm ento es sinténico
sólo en uno de los dos taxa. Las sintenias pueden rom perse tanto por
fisiones o t ranslocaciones crom osóm icas y el análisis de la conservación de
estas sintenias a lo largo de la filogenia de un grupo perm ite analizar los
reordenam ientos cromosómicos con posible com prom iso especiogénico en

1. I nt roducción
19
esos taxa. Actualm ente, debido a que para los crom osom as hum anos se
han desarrollado sondas tanto a nivel de cromosom a entero como de brazos
y segm entos parciales, se ut ilizan estas sondas para analizar,
indirectam ente, la evolución de un grupo de interés part icular en relación al
cariot ipo humano (Ehrlich et al. 1997; Wienberg y Stanyon 1997; Müller 2006; Stanyon
et al. 2008) .
Al com parar los cariot ipos de dist intos órdenes de m am íferos, se
observa un alto grado de conservación sinténica (Müller 2006; Ferguson-Sm ith y
Tr ifonov 2007; CHROMOME: Nagarajan et al. 2008) . Los datos obtenidos m ediante
FI SH con sondas de cromosom as hum anos perm it ieron la realización de
inferencias sobre un posible car iot ipo ancest ral para mamíferos placentarios
con un 2n= 46 (Müller 2006; Ferguson-Sm ith y Tr ifonov 2007; CHROMOME: Nagarajan et
al. 2008; Robinson y Ruiz-Herrera 2008, 2010) . Los cromosom as hom eólogos de los
autosom as hum anos 1, 5, 6, 9, 11, 13, 17, 18 y 20 se encont rarían
conservados en el cariot ipo ancest ral como ent idades separadas (es decir ,
como cromosom as enteros) , por lo que se asum e que representarían las
form as cromosóm icas ancest rales. Otros autosom as se encuent ran divididos
en dos segm entos sinténicos, tales com o los hom eólogos de los
crom osom as 2, 7, 8, 10, 16 y 19. Los hom eólogos de los autosom as
hum anos 3, 4, 14, 15 y 21 se conservan enteros involucrados en
t ranslocaciones form ando las sintenias 3/ 21, 4/ 8p y 14/ 15, por lo que se
considera que estas sintenias serían ancest rales para los mamíferos
placentarios (Richard y Dut r illaux 1998; O´ Brien et al. 1999; Wienberg et al. 1999; Müller
et al. 2000; Wienberg 2005; Müller 2006) .
1 .2 .3 1 Pr im ates
En el caso del Orden Prim ates (grupo objeto de estudio de este
t rabajo de Tesis) , el car iot ipo ancest ral inferido m ediante FI SH con sondas
hum anas muest ra pocas diferencias con el cariot ipo ancest ral de m amíferos
placentarios (Figura 1.2) . Se postula que habrían ocurr ido fisiones
involucrando a los hom eólogos de los siguientes brazos cromosóm icos
hum anos: una fisión ent re 10p y 12pter-q23.3 con una consiguiente fusión
ent re 10p y 10q dando lugar al hom eólogo del crom osoma 10 hum ano (HSA

1. I nt roducción
20
10 por cromosom a 10 de Hom o sapiens) ; una fisión ent re 16q y 19q; una
fisión ent re los hom eólogos a HSA 4 and HSA 8p. Estos reordenam ientos
darían com o resultado un cariot ipo ancest ral con 2n= 50 cromosomas,
conservando las sintenias ancest rales para m am íferos 3/ 21, 14/ 15, 12/ 22
(dos bloques) y 7/ 16 y los bloques sinténicos 1, 2 (part ida en dos bloques) ,
4, 5, 6, 7, 8, 9, 10, 11, 13, 17, 18, 19 (part ida en dos bloques) , 20, X e Y
(Müller 2006; Ferguson-Sm ith y Tr ifonov 2007; CHROMOME: Nagarajan et al. 2008; Stanyon
et al. 2008) .
Figura 1.2: Car iot ipo ancest ral infer ido para mamíferos placentar ios y para primates. A la
izquierda de cada crom osom a se indica la asignación de hom eología m ediante sondas de crom osom as
hum anos. Se indican los rearreglos infer idos ent re uno y ot ro cariot ipo (m odificada de Müller 2006) .
Este m ism o análisis puede ser aplicado dentro del Orden Prim ates
para postular los rearreglos crom osómicos m arcadores que darían lugar al
cariot ipo hum ano actual (Figura 1.3) . Luego de la divergencia de los
St repsirrhini y Ant ropoidea se postularon los siguientes cam bios
crom osóm icos: fisión ent re los dos pares con hom eología 12/ 22 y posterior
fusión dando origen a los hom eólogos de los actuales cromosom as HSA 12 y
HSA 22; fusión de los dos pares hom eólogos a HSA 19 dando lugar al
hom eólogo del crom osom a actual; fisión de la sintenia 7/ 16. En la
divergencia ent re Monos del Nuevo Mundo y del Viejo Mundo encont ram os
como reordenam ientos m arcadores a los siguientes rearreglos: fisión de la

1. I nt roducción
21
sintenia ancest ral 3/ 21 para dar a los hom eólogos actuales HSA 3 y HSA
21; fusión de los dos hom eólogos a HSA 7 para dar lugar al hom eólogo del
crom osom a actual; fusión de los dos hom eólogos a HSA 16 para dar lugar al
hom eólogo del crom osoma actual. Siguiendo la filogenia, en la separación
ent re los Monos del Viejo Mundo y los Sim ios (grupo que comprende a los
gibones, orangutanes, gorilas, chimpancés y al hum ano) encont ram os com o
m arcador la rotura de la sintenia ancest ral 14/ 15 para dar lugar a los
hom eólogos de los cromosom as HSA 14 y HSA 15 actuales. Por últ imo, el
crom osom a 2 hum ano se habría form ado de la fusión ent re los dos
hom eólogos ancest rales.
Figura 1.3: Reordenam ientos crom osóm icos m arcadores en la evolución de los prim ates
(m odificada de Wienberg 2005) .
Al analizar la conservación de las sintenias crom osóm icas, aparece
una característ ica interesante en los Primates del Nuevo Mundo que es la
ausencia de hom eología del crom osom a Y de Platyrrhini con el cromosom a Y
hum ano. En estudios previos de FISH con sondas de crom osom a Y hum ano
sobre m etafases de Prim ates Neot ropicales, la sonda no m ost ró señal de
hibridación en ningún crom osoma de las m etafases m itót icas analizadas en

1. I nt roducción
22
dist intos ceboideos, en cont raposición con la m arcada conservación del
crom osom a X descrita en los m am íferos (Consigliere et al. 1996, 1998; Mudry et al.
2001; de Oliveira et al. 2002) . Por el cont rar io, en m onos del Viejo Mundo sí se
ha descr ito la conservación del cromosoma Y (Archidiacono et al. 1998) . Esta
falta de hom eología y el com prom iso del crom osom a Y en Platyrrhini en
sistem as de determ inación sexual m últ iple sugieren una histor ia evolut iva
diferencial para este cromosom a en el linaje de Prim ates del Viejo Mundo
con respecto al linaje del Nuevo Mundo.
De lo anteriorm ente expuesto cobra im portancia el estudio de la
conservación de las sintenias cromosóm icas para la m ejor com prensión de
la sistem át ica y la filogenia de un grupo, en especial en el de los prim ates
donde se ha postulado la especiación crom osóm ica com o posible
m ecanism o evolut ivo (Clem ente et al. 1990) .
1 .2 .4 El gen SRY
A nivel génico, la determ inación génica del sexo m asculino en
m am íferos depende de la presencia del gen SRY ( “Sex-determ ining Region
of the Y” ) , ubicado en hum anos en Yp11.3. Codifica para un pépt ido de 204
am inoácidos, con un dom inio de unión al ADN, “HMG box” ( “H igh-Mobility
Group box” ) (Sinclair et al. 1990) . Este dom inio se encuent ra en factores de
t ranscripción y proteínas crom at ínicas no histónicas e induce una torsión en
la región de ADN a la que se une. En los m achos, la expresión del SRY inicia
la cascada de eventos que lleva al desarrollo de la gónada indiferenciada a
test ículo.
El gen que codifica para el SRY se encuent ra casi en el borde (a 5 kb)
de la región pseudoautosóm ica (PseudoAutosom al Region o PAR en inglés) ,
por lo que no es raro que, a veces, ocurra un ent recruzam iento por error
ent re el X y el Y en esa zona y sea este gen t ransfer ido al cromosom a X.
Cuando esto ocurre se pueden observar individuos herm afroditas 46 XX
m osaicos. El SRY se inact iva al azar por el fenóm eno de com pensación de
dosis y se observa m asculinización incompleta. Esto estaría indicando que

1. I nt roducción
23
se necesita un nivel um bral de SRY para asegurar una norm al
determ inación test icular (Veit ia et al. 2001) .
1 .2 .4 .1 Pr im ates
Cóm o ya se refir ió previam ente, el crom osom a Y de Prim ates se
caracter iza citogenét icam ente por una alta var iabilidad est ructural y por su
comprom iso en diversos pat rones de determ inación sexual ( revisado en Solari
1993) . En Primates del Viejo Mundo se ha descr ito la conservación tanto del
crom osom a Y como del gen SRY hum ano (Archidiacono et al. 1998) . En Primates
Neot ropicales, por el cont rario, se ha descrito la ausencia de hom eología del
crom osom a Y de Platyrrhini con el crom osom a Y humano m ediante técnicas
de FI SH.
Respecto al gen SRY, Moreira (2002) , estudió su evolución m ediante
técnicas de biología molecular (PCR, “Polym erase chain react ion” ) en 7
géneros de Ceboidea, observando 7 sust ituciones en la región HMG ent re
hum anos y ceboideos. Considerando estos antecedentes, se abordó este
tem a en el GI BE y m ediante amplificación por PCR, se detectó la presencia
del SRY en 5 ceboideos (Alouat ta caraya, Saim ir i boliviensis boliviensis,
Cebus libidinosus (ex paraguayanus) , Ateles cham ek y A. paniscus) con
tem peraturas de “annealing” ent re 47˚ C y 50˚ C y bandas diferentes
especie-específicas para la secuencia de la HMG box del SRY humano (299
pb) , observándose dos bandas en la calle del gel correspondiente a Alouat ta
caraya (Nieves et al. 2003) .
Estudios previos por hibridación in situ con una sonda del SRY
hum ano m ostraron que en Prim ates del Viejo Mundo el SRY se encuent ra
próximo a la región pseudoautosóm ica (Gläser et al. 1998) . En ese m ismo
t rabajo se estudiaron especies de Primates del Nuevo Mundo de los géneros
Callithr ix y Ateles en las cuales el SRY se observó a su vez próxim o a la
PAR.
Dada la presencia de sistem as de determ inación sexual múlt iple en
Alouat ta, cuyos rearreglos crom osómicos involucran al crom osom a Y, se

1. I nt roducción
24
pone en evidencia la importancia del estudio de la posición del SRY en el
cariot ipo y de su part icipación en los sistem as sexuales m últ iples respecto
de los sistem as XY, inform ación que aún no ha sido objeto de publicaciones
especializadas según las revisiones actualizadas realizadas al m om ento de
redacción de la presente Tesis.
El análisis de la conservación y posición del gen SRY humano en los
sexuales m últ iples de aulladores, com parado con los sistem as XY presente
en ot ros ceboideos, brindará inform ación de ut ilidad para proponer hipótesis
sobre los rearreglos que pudieron dar lugar a los sistem as sexuales
m últ iples observables hoy en aulladores.
1 .3 El m odelo de estudio
Las form as actuales del Orden Primates com prenden dos subórdenes,
St repsirrhini, que incluye a los lem úridos, quirogaleidos, lorísidos, aye-ayes,
índr idos y gálagos, y los Haplorrhini, que agrupa a los Tarsiform es, a los
Primates del Nuevo (Platyrrhini) y del Viejo Mundo (Catarrhini) , incluyendo
en este últ im o grupo a los Hom inoidea (Groves 2001) . Los pr im ates no
hum anos habitan actualm ente en 5 de los 7 cont inentes, asociados en
general a climas t ropicales que aseguran sus recursos indispensables para
sobrevivir . Si bien son la excepción, unas pocas especies viven en climas
tem plados (Fleagle 1999; Rowe 1996; Groves 2001; Rylands y Mit term eier 2009) .
Los Prim ates del Nuevo Mundo se dist r ibuyen desde el sur de México
hasta el norte de Argent ina. Hasta el presente no existe un acuerdo en el
núm ero de fam ilias y especies que com prenden estos prim ates
neot ropicales.
A nivel sistem át ico se han realizado num erosos estudios ut ilizando
tanto caracteres morfológicos com o genét icos. Brevem ente, a nivel
supragenérico estudios realizados en base a caracteres m orfológicos
reconocen 2 fam ilias, Callit r ichidae (Cebuella, Callithr ix, Saguinus, Callim ico
y Leontopitecus) y Cebidae (Aotus, Callicebus, Pithecia, Chiropotes,
Cacajao, Alouat ta, Saim ir i, Cebus, Ateles, Lagothr ix y Brachyteles) (Sim pson

1. I nt roducción
25
1945; Hill 1962; Cabrera 1957) . Hershkovitz (1977) elevó a Callim ico a su propia
fam ilia, Callim iconidae, reconociendo así 3 fam ilias m ediante caracteres
m orfológicos. Callim ico se caracteriza por tener t res m olares como los
cébidos, caracteres bioquím icos de los callit r ícidos y, de acuerdo a Egozcue
(1971) , un cariot ipo más cercano a los callit r ícidos que a los cébidos. Ese
m ismo año, ot ro análisis por caracteres m orfológicos consideraba a este
género com o “ t it ís anóm alos” y lo ubicaba dent ro de los callit r ícidos
(Thorington 1977) .
Esta est ructura de clasificación se m antuvo hasta 1981, año en que
Rosenberger publica su estudio basado en datos postcraneales y dentar ios y
propone la clasificación en dos fam ilias: Cebidae (Saim ir i, Cebus, Cebuella,
Callithr ix, Saguinus, Callim ico y Leontopitecus) y Atelidae (Callicebus,
Aotus, Pithecia, Chiropotes, Cacajao, Alouat ta, Ateles, Lagothrix y
Brachyteles) . Esta últ ima clasificación fue avalada parcialm ente por las
nuevas evidencias genét icas, las cuales m uest ran t res clados m onofilét icos:
el pr im ero incluye al grupo form ado por Callicebus y los pitécidos (Pithecia,
Chiropotes y Cacajao) form ando la fam ilia Pitheciidae, el segundo incluye a
los Alouat ta con los atelinos (Ateles, Lagothr ix y Brachyteles) form ando la
fam ilia Atelidae y el tercero incluye a los géneros Cebus y Saim ir i j unto con
los callit r ícidos (Callithr ix, Cebuella, Leontopithecus, Saguinus y Callim ico)
form ando la fam ilia Cebidae (Schneider et al. 1993, 2000; Schneider y Rosenberger
1996; Harada et al. 1995; Barroso et al. 1997) .
Aún siguen en discusión la posición de ciertos géneros, tales com o
Aotus, que algunos autores colocan dent ro de la fam ilia Aot idae (Groves
2001; Rylands y Mit term eier 2009) , m ient ras que ot ros colocan a Aotus dent ro de
la fam ilia Pitheciidae (Rosenberger y Tejedor 2011) .
En el m arco de la “Evidencia Total” , tom ando en cuenta que la
definición de una especie debe considerar tanto datos provenientes de la
m orfología, la anatomía, la genét ica y la ecología de un taxón, en este
t rabajo de Tesis se consideró la clasificación compuesta por 4 fam ilias:
Cebidae, Callithr icidae, Pitheciidae y Atelidae basada en la clasificación
propuesta por Schneider et al. (2001) con m odificaciones (Tabla 1.2) , en la que

1. I nt roducción
26
se tom ó en consideración tanto caracteres genét icos (m oleculares y
citogenét icos) com o m orfológicos.
Tabla 1.2: Clasif icación de los Pr im ates del Nuevo Mundo (Schneider et al. 2001 m odificada
según Mudry et al. 2006) . * modificaciones
I nfraorden: Platyrrhini
Superfam ilia: Ceboidea
Fam ilia : Atelidae
Subfam ilia : Alouat t inae *
Alouat ta
Subfam ilia : Atelinae
Lagothrix
Brachyteles
Ateles
Fam ilia : Cebidae
Subfam ilia : Cebinae
Cebus
Subfam ilia : Saim ir inae
Saim ir i
Subfam ilia : Aot inae
Aotus
Fam ilia : Callit r ichidae *
Cebuella
Callithrix
Leontopithecus
Saguinus
Callim ico
Fam ilia : Pitheciidae
Subfam ilia : Pitheciinae
Pithecia
Chiropotes
Cacajao
Callicebus
1 .3 .1 El género Alouat ta
Dentro de la fam ilia Atelidae encontram os a los monos aulladores
(género Alouat ta) que poseen una de las dist r ibuciones geográficas m ás
am plias regist radas al día de hoy para los Prim ates Neot ropicales (Figura

1. I nt roducción
27
1.4) ya que se ext iende desde el sur de México hasta el norte de Argent ina
(Crocket t y Eisenberg 1987; Rylands et al. 2000) .
Figura 1.4: Dist r ibución actual del género Alouat ta. Mapa confeccionado por la autora de este
t rabajo de Tesis con datos de Rylands (2000) , Groves (2001) y Hirsch (2002) . Fotos tom adas de Rowe
(1996) .
Los aulladores habitan un rango de ambientes m uy diverso,
incluyendo la selva t ropical lluviosa, selva de inundación, bosques en
galería, parches de bosque y am bientes estacionales deciduos y
sem ideciduos (Crocket t y Eisenberg 1987; Zunino et al. 2001) .

1. I nt roducción
28
Tradicionalm ente los monos aulladores fueron agrupados en su
clasificación en 6 especies: A. palliata, A. pigra, A. seniculus, A. belzebul, A.
fusca y A. caraya (Cabrera 1958; Hill 1962) . Dos revisiones sistem át icas
poster iores elevaron al rango de especie a varias subespecies (Rylands 2000;
Groves 2001, 2005; Gregorin 2006; Rylands y Mit termeier 2009) . Rylands (2000)
reconoció 9 especies: A. seniculus, A. sara, A. nigerr im a, A. belzebul, A.
guariba (= fusca) , A. palliat ta, A. coibensis, A. caraya, A. pigra m ient ras que
unos años después Groves (2001, 2005) refir ió 10 especies de aulladores al
incorporar a A. m acconnelli com o especie plena.
Más recientem ente (Gregorin 2006) , un estudio sobre las especies
brasileñas basado en el pat rón de coloración de pelaje reconoció 10
especies, elevando num erosas subespecies a la categoría de especies
plenas: A. caraya, A. belzebul, A. discolor, A. ululata, A. fusca, A. clam itans,
A. juara, A. m acconnelli, A. nigerr im a, A. puruensis. Este estudio fue
reconocido por Rylands y Mit termeier (2009) quienes a su vez, elevaron el
núm ero de especies reconocidas a 14: A. seniculus, A. arctoidea, A.
m acconnelli, A. juara, A. puruensis, A. sara, A. nigerr im a, A. belzebul, A.
discolor, A. ululata, A. guariba, A. caraya, A. palliata y A. pigra. Sin
embargo, el pat rón de coloración de pelaje por sí sólo no es caracter
suficiente para la correcta ident if icación taxonóm ica dada su naturaleza
polimórfica, m áxim e para un género en el que se han descrito en vida
silvest re individuos con pat rón de coloración m ixto, posiblem ente de
procedencia híbrida (Aguiar et al. 2007; Agost ini et al. 2008) , y en el que se ha
confirm ado m ediante estudios genét icos la presencia de híbr idos en la
naturaleza (Cortés-Ort iz et al. 2007) . Por esta razón en este t rabajo de Tesis se
adhiere a la clasificación propuesta por Groves (2001, 2005) en la que se
reconocen 10 especies para el género.
Sin em bargo, aún existe una falta de consenso en la com unidad de
pr imatólogos en cuanto al reconocim iento del número de especies dent ro
del género así com o sus posibles relaciones de parentesco. Por ello, los
estudios genét icos (citogenét icos y m oleculares) se han convert ido en una
im portante y valiosa herram ienta que com plem enta los análisis fenot ípicos y
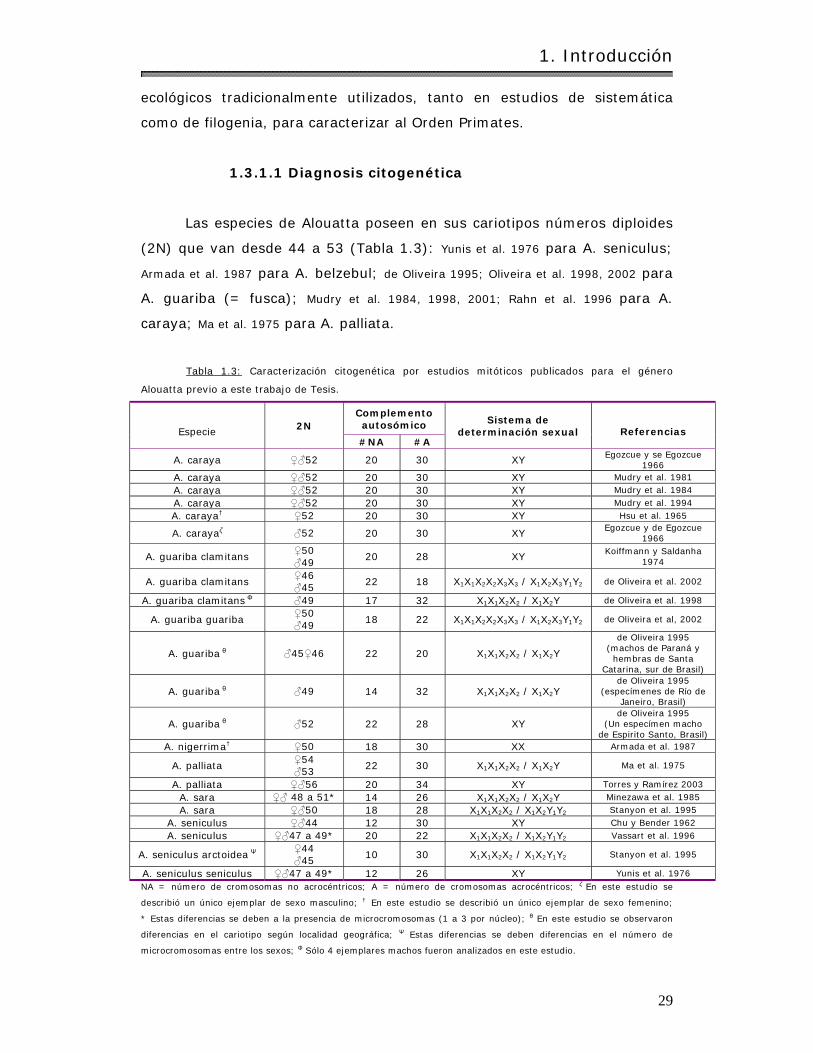
1. I nt roducción
29
ecológicos t radicionalm ente ut ilizados, tanto en estudios de sistem át ica
como de filogenia, para caracter izar al Orden Prim ates.
1 .3 .1 .1 Diagnosis citogenét ica
Las especies de Alouat ta poseen en sus cariot ipos núm eros diploides
(2N) que van desde 44 a 53 (Tabla 1.3) : Yunis et al. 1976 para A. seniculus;
Arm ada et al. 1987 para A. belzebul; de Oliveira 1995; Oliveira et al. 1998, 2002 para
A. guariba (= fusca) ; Mudry et al. 1984, 1998, 2001; Rahn et al. 1996 para A.
caraya; Ma et al. 1975 para A. palliata.
Tabla 1.3: Caracter ización citogenét ica por estudios m itót icos publicados para el género
Alouat ta previo a este t rabajo de Tesis.
Especie 2 N Com plem ento
autosóm ico Sistem a de determ inación sexual Referencias
# NA # A
A. caraya ♀♂52 20 30 XY Egozcue y se Egozcue
1966 A. caraya ♀♂52 20 30 XY Mudry et al. 1981
A. caraya ♀♂52 20 30 XY Mudry et al. 1984
A. caraya ♀♂52 20 30 XY Mudry et al. 1994
A. caraya† ♀52 20 30 XY Hsu et al. 1965
A. carayaζ ♂52 20 30 XY Egozcue y de Egozcue
1966
A. guariba clam itans ♀50 ♂49
20 28 XY Koiffmann y Saldanha
1974
A. guariba clam itans ♀46 ♂45
22 18 X1X1X2X2X3X3 / X1X2X3Y1Y2 de Oliveira et al. 2002
A. guariba clam itans Φ ♂49 17 32 X1X1X2X2 / X1X2Y de Oliveira et al. 1998
A. guariba guariba ♀50 ♂49
18 22 X1X1X2X2X3X3 / X1X2X3Y1Y2 de Oliveira et al, 2002
A. guariba θ ♂45♀46 22 20 X1X1X2X2 / X1X2Y
de Oliveira 1995 (m achos de Paraná y
hem bras de Santa Catarina, sur de Brasil)
A. guariba θ ♂49 14 32 X1X1X2X2 / X1X2Y de Oliveira 1995
(especímenes de Río de Janeiro, Brasil)
A. guariba θ ♂52 22 28 XY de Oliveira 1995
(Un especímen macho de Espir ito Santo, Brasil)
A. nigerr im a† ♀50 18 30 XX Arm ada et al. 1987
A. palliata ♀54 ♂53
22 30 X1X1X2X2 / X1X2Y Ma et al. 1975
A. palliata ♀♂56 20 34 XY Torres y Ram írez 2003
A. sara ♀♂ 48 a 51* 14 26 X1X1X2X2 / X1X2Y Minezawa et al. 1985
A. sara ♀♂50 18 28 X1X1X2X2 / X1X2Y1Y2 Stanyon et al. 1995
A. seniculus ♀♂44 12 30 XY Chu y Bender 1962
A. seniculus ♀♂47 a 49* 20 22 X1X1X2X2 / X1X2Y1Y2 Vassart et al. 1996
A. seniculus arctoidea Ψ ♀44 ♂45
10 30 X1X1X2X2 / X1X2Y1Y2 Stanyon et al. 1995
A. seniculus seniculus ♀♂47 a 49* 12 26 XY Yunis et al. 1976
NA = núm ero de crom osom as no acrocéntr icos; A = núm ero de crom osomas acrocént r icos; ζ En este estudio se
descr ibió un único ejem plar de sexo masculino; † En este estudio se descr ibió un único ejem plar de sexo femenino;
* Estas diferencias se deben a la presencia de m icrocrom osom as (1 a 3 por núcleo) ; θ En este estudio se observaron
diferencias en el cariot ipo según localidad geográfica; Ψ Estas diferencias se deben diferencias en el número de
m icrocromosom as ent re los sexos; Φ Sólo 4 ejemplares m achos fueron analizados en este estudio.

1. I nt roducción
30
En este género se ha observado la presencia de sistem as de
determ inación sexual múlt iple, que se habrían originado a part ir de
t ranslocaciónes Y-autosom a, com o se ejem plifica en la Figura 1.5.
Resultados de estudios m itót icos (Tabla 1.3) han sugerido la presencia de
sistem as de t ipo X1X2X3Y1Y2 (que form aría una cadena de 5 elem entos o
pentavalente en Metafase I ) en A. guariba (de Oliveira et al. 2002) y de t ipo
X1X2Y1Y2 en A. sara (Minezawa et al. 1985; Consigliere et al. 1996) , m ient ras que
ot ros t rabajos sugieren la presencia de sistem as sexuales en m achos de
t ipo XY (A. seniculus: Yunis et al. 1976; A. guariba clam itans: Koiffm ann y Saldanha 1974;
A. palliata: Torres y Ram írez 2003) . Estos resultados aún esperan confirm ación
por estudios m eiót icos.
Figura 1.5: Posible or igen para el sistem a de determ inación sexual m últ iple X1X2Y1Y2 en los
m achos de Alouat ta caraya (m od. de Rahn et al., 1996) . En esta especie el par autosóm ico que estaría
involucrado en la t ranslocación Y-autosoma sería el par ACA7. El X ancest ral se muest ra de color blanco,
el crom osom a Y ancest ral en punteado gr is sobre blanco y el par autosóm ico 7 (A7) se m uest ra relleno
de rayas t ransversales gr is oscuro. A part ir de un sistem a de t ipo XY, roturas sim ultáneas en A7pprox y
en la región distal de Yq, seguido de una t ranslocación recíproca ent re ellos, habría dado lugar a los
crom osom as Y1 e Y2. El crom osom a hom ólogo del par A7 que no estuvo involucrado en la t ranslocación
const ituir ía el crom osom a X2 y el X ancest ral se denom inaría ahora X1.
La presencia de sistem as de determ inación sexual m últ iples sólo se
ha confirm ado hasta la fecha por estudios m eiót icos en m achos (Tabla 1.4) :
de t ipo X1X2Y1Y2 ( formando una cadena de 4 elem entos o cuadrivalente en
Metafase I ) en Alouat ta seniculus (Lim a & Seuánez, 1991) , A. caraya (Mudry et
al. 1998, 2001) ; de t ipo X1X2Y ( form ando una cadena de 3 elem entos o
t r ivalente en Metafase I ) en A. belzebul (Arm ada et al. 1987) y A. palliat ta
(Solar i y Rahn 2005) .
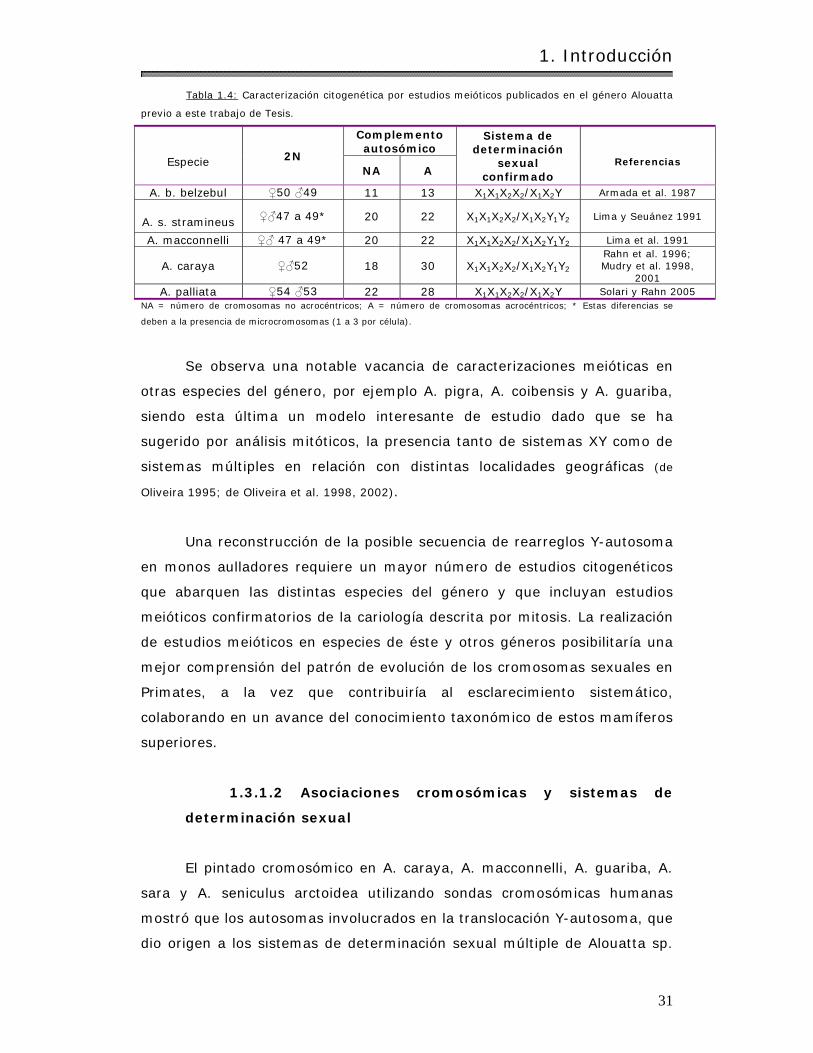
1. I nt roducción
31
Tabla 1.4: Caracter ización citogenét ica por estudios m eiót icos publicados en el género Alouat ta
previo a este t rabajo de Tesis.
Especie 2 N
Com plem ento autosóm ico
Sistem a de determ inación
sexual confirm ado
Referencias NA A
A. b. belzebul ♀50 ♂49 11 13 X1X1X2X2/ X1X2Y Arm ada et al. 1987
A. s. st ram ineus ♀♂47 a 49* 20 22 X1X1X2X2/ X1X2Y1Y2 Lim a y Seuánez 1991
A. macconnelli ♀♂ 47 a 49* 20 22 X1X1X2X2/ X1X2Y1Y2 Lim a et al. 1991
A. caraya ♀♂52 18 30 X1X1X2X2/ X1X2Y1Y2 Rahn et al. 1996; Mudry et al. 1998,
2001 A. palliata ♀54 ♂53 22 28 X1X1X2X2/ X1X2Y Solari y Rahn 2005
NA = núm ero de cromosomas no acrocént r icos; A = número de cromosomas acrocént r icos; * Estas diferencias se
deben a la presencia de m icrocrom osom as (1 a 3 por célula) .
Se observa una notable vacancia de caracterizaciones m eiót icas en
ot ras especies del género, por ejem plo A. pigra, A. coibensis y A. guariba,
siendo esta últ im a un m odelo interesante de estudio dado que se ha
sugerido por análisis m itót icos, la presencia tanto de sistem as XY com o de
sistem as m últ iples en relación con dist intas localidades geográficas (de
Oliveira 1995; de Oliveira et al. 1998, 2002) .
Una reconst rucción de la posible secuencia de rearreglos Y-autosom a
en m onos aulladores requiere un m ayor núm ero de estudios citogenét icos
que abarquen las dist intas especies del género y que incluyan estudios
m eiót icos confirm ator ios de la car iología descr ita por m itosis. La realización
de estudios m eiót icos en especies de éste y ot ros géneros posibilitaría una
m ejor com prensión del pat rón de evolución de los crom osom as sexuales en
Primates, a la vez que cont r ibuiría al esclarecim iento sistem át ico,
colaborando en un avance del conocim iento taxonóm ico de estos m am íferos
superiores.
1 .3 .1 .2 Asociaciones crom osóm icas y sistem as de
determ inación sexual
El pintado cromosóm ico en A. caraya, A. m acconnelli, A. guariba, A.
sara y A. seniculus arctoidea ut ilizando sondas cromosómicas hum anas
m ost ró que los autosom as involucrados en la t ranslocación Y-autosom a, que
dio or igen a los sistemas de determ inación sexual múlt iple de Alouat ta sp.

1. I nt roducción
32
exhiben señal de hibr idación para los cromosom as 3 y 15 hum anos (Figura
1.6) (Consigliere et al. 1996, 1998; de Oliveira et al. 2002; Mudry et al. 2001) .
Figura 1.6: Pat rón de Hibr idación in situ Fluorescente con las sondas de crom osomas humanos
X, 3 y 15 en las especies de Alouat ta sp. con determ inación sexual en machos X1X2Y1Y2. Los núm eros a
la derecha de los crom osom as indican la señal de hibr idación de cada crom osom a hum ano en su
correspondiente hom eólogo en Alouat ta (Tom ada de Consigliere et al. 1996) .
Esta sintenia 3/ 15 fue descrita en autosom as de especies de Atelidae
tales com o Ateles geoffroyi y A. belzebul hibr idus (Morescalchi et al. 1997; García
Haro 2001) si bien no fue observada en ot ros géneros de Prim ates
Neot ropicales tales com o Cebus libidinosus (= C. paraguayanus) o Saim ir i
boliviensis boliviensis (Mudry et al. 2001) .
En base a estos hallazgos se puede interpretar que los sistem as
sexuales m últ iples se habrían originado a part ir del m ismo par cromosómico
para todas las especies de aulladores descritas hasta la fecha y que estos
reordenam ientos deberían haber ocurr ido m uy tem pranam ente en la
divergencia del género. Con esta interpretación com o m arco teór ico, esta
sintenia puede ser considerada com o la condición ancest ral para la fam ilia
Atelidae, donde la asociación con los crom osom as sexuales m últ iples
const ituir ía una novedad evolut iva sinapom órfica.
1 .3 .1 .3 Estudios f ilogenét icos
En 1989 Kluge int rodujo en los estudios de Sistem át ica la expresión
“Evidencia Total” para el análisis filogenét ico e incluyendo en este concepto
todos los caracteres (entendidos com o datos) disponibles en una única

1. I nt roducción
33
m atriz, donde no sólo se consideraran caracteres morfológicos, sino
tam bién genét icos, fisiológicos, ecológicos, com portam entales, ent re ot ros,
con el objet ivo de alcanzar la m ayor congruencia taxonóm ica posible (Kluge
1989) . Y en este t rabajo de Tesis es bajo este nuevo enfoque
m ult idisciplinar io que se considera im portante adoptar el concepto al
considerar que aplicándolo se pueden lograr importantes avances en el
conocim iento y aportar cont r ibuciones novedosas a la sistem át ica de una
especie y/ o género.
Las filogenias propuestas hasta ahora para Alouat ta ut ilizaron en
algunos casos, m arcadores m oleculares (Meireles et al. 1999; Cortés-Ort iz et al.
2003) y en ot ros, caracteres citogenét icos (de Oliveira et al. 2002) , y pocos son
los t rabajos que han com parado y tom ado en cuenta am bos t ipos de
caracteres (Bonvicino et al. 2001; Villalobos et al. 2004) . Estos últ im os t rabajos
incluyeron caracteres crom osómicos sólo para las especies sudam ericanas,
dado que las especies cent roam ericanas se encontraban escasam ente
caracterizadas citogenét icam ente para el m om ento de la publicación, y no
se realizó un análisis combinado de los m ism os, sino que se superpuso la
inform ación cromosóm ica sobre la filogenia obtenida a part ir de los
caracteres m oleculares.
En estas filogenias se ha propuesto la ancest ralidad del sistem a
sexual X1X1X2X2/ X1X2Y1Y2. Sin em bargo, com o discuten Solari y Rahn (2005) , el
sistem a sexual t r ivalente es m ás simple que el cuadrivalente y está
presente en ot ros géneros de Prim ates Neot ropicales, com o Aotus, Callim ico
y Cacajao (Ma et al. 1976; Seuánez et al. 1989; Mourá-Pensin et al. 2001) , con lo que
una hipótesis de ancest ralidad de este sistem a de determ inación sexual
X1X1X2X2/ X1X2Y parecería ser m ás parsimoniosa.
Por lo anteriorm ente expresado, la realización en este t rabajo de
Tesis de nuevos análisis filogenét icos incluyendo caracteres citogenét icos y
m oleculares brindará inform ación de im portancia para el esclarecim iento de
la sistem át ica de este género y perm it irá confirm ar la evolución
crom osóm ica com o est rategia especiogénica, en el marco de un análisis de
“Evidencia Total” .

OBJETI VOS E HI PÓTESI S

2. Objet ivos e Hipótesis
34
2 .1 Objet ivos
2 .1 .1 Objet ivo Genera l
Caracterizar el sistem a de determ inación sexual en aulladores
(género Alouat ta, Prim ates: Platyrrhini) y proponer una interpretación del
posible pat rón evolut ivo así com o de su relación con los sistem as de
determ inación sexual en ot ros Prim ates.
2 .1 .2 Objet ivos Específ icos
1. Caracterizar los sistem as de determ inación sexual en aulladores
(género Alouat ta) a part ir de estudios citogenét icos en células som át icas y
germ inales.
2. Proponer una filogenia para los aulladores (género Alouat ta) y su
relación con ot ros Ceboidea teniendo en cuenta parám et ros citogenét icos
(propios de este t rabajo y de la literatura) y de secuencias de ADN
(obtenidas de la literatura) .
3. Analizar la conservación y posición crom osóm ica del gen SRY
hum ano en Ceboidea a fin de proponer un posible m odelo evolut ivo.
2 .2 Hipótesis de t raba jo
1. En el género Alouat ta, el sistem a de determ inación sexual
ancest ral en m achos es de t ipo X1X2Y.
2. En aulladores (género Alouat ta) la sintenia 3/ 15 se encuent ra
conservada y asociada a los cromosom as sexuales.
3. En Ceboidea el gen SRY hum ano se halla conservado, incluyendo
las especies con sistem as sexuales m últ iples, y se ubica próximo a la PAR
del crom osom a Y.

MATERI ALES Y MÉTODOS

3. Materiales y m étodos
35
3 .1 Mater ia l Biológico
Para el presente t rabajo de Tesis se realizaron análisis genét icos a
part ir de m uest ras de sangre perifér ica y biopsias de tej ido test icular. Se
analizó un total de 43 ejem plares del Orden Primates.
Se estudiaron ejem plares de prim ates ceboideos y hom inoideos
procedentes tanto de vida silvest re como de caut iver io ( jardines
zoológicos y estaciones de cría) en Argent ina y en México.
Se t rabajó con m uest ras de sangre perifér ica hum ana (Homo
sapiens) correspondientes a donantes voluntar ios sanos, adultos de
am bos sexos.
La Tabla 3.1 resum e la información detallada de las especies
estudiadas, núm ero de ejem plares, sexo, procedencia y t ipo de muest ra.
Los prim ates no-humanos (Ceboidea y Hom inoidea) alojados en
zoológicos y cent ros de cría, fueron inm ovilizados por sus respect ivos
cuidadores y anestesiados por profesionales veter inarios según los
procedim ientos habituales de cada inst itución.
En el caso de los ejem plares de Ceboidea m uestreados en vida
silvest re, dent ro de la dist r ibución geográfica m arginal norte del género
Alouat ta, se estudiaron 4 m achos y 4 hem bras de edad adulta de
Alouat ta pigra (especie que hasta este t rabajo de Tesis no había sido
caracterizada citogenét icam ente) . Los ejemplares fueron capturados en 2
localidades de Cam peche, México, en el rango de dist r ibución natural de
la especie: Rancho “Montebravo” (A: 18°51´ 46,7”N; 9 0°46´ 8,6”O) y
Rancho “Don Álvaro” (B: 18°34´ 13,4”N; 90°26´ 18”O) .
En el caso de las m uest ras de sangre perifér ica de Hom o sapiens,
las m ismas fueron ext raídas por la Dra. Ana María Buzaleh del
Departam ento de Quím ica Biológica de la FCEyN, UBA.

3. Materiales y m étodos
36
En todos los casos, cada individuo analizado fue regist rado con un
núm ero en el libro de ent rada y procesam iento de m uestras del GI BE. En
Anexo I (adjunto al f inal de este t rabajo de Tesis) se detalla el listado
com pleto de individuos con su núm ero de ident ificación, su procedencia y
dem ás característ icas que hacen a la ident ificación de la muest ra.
En Ceboidea se realizó la ident ificación fenot ípica de cada ejem plar
según los pat rones de coloración del pelaje especie-específicos descritos
para cada especie analizada: Alouat ta caraya (Cabrera 1958) , A. pigra (Smith
1970) , A. palliata (Sm ith 1970) , A. guariba clam itans (Cabrera 1958) , Saim ir i
boliv iensis boliviensis (Hershkovit z 1984) , Cebus libidinosus (Cabrera 1958) , C.
nigritus (Cabrera 1958) , Aotus azarae (Cabrera 1958) .
El sexo de cada ejemplar de Ceboidea fue determ inado m ediante
observación directa. En los casos en que fue posible, la edad de los
m ism os, se determ inó m ediante el pat rón de erupción dental (para una
revisión del tem a ver Henderson 2003) .
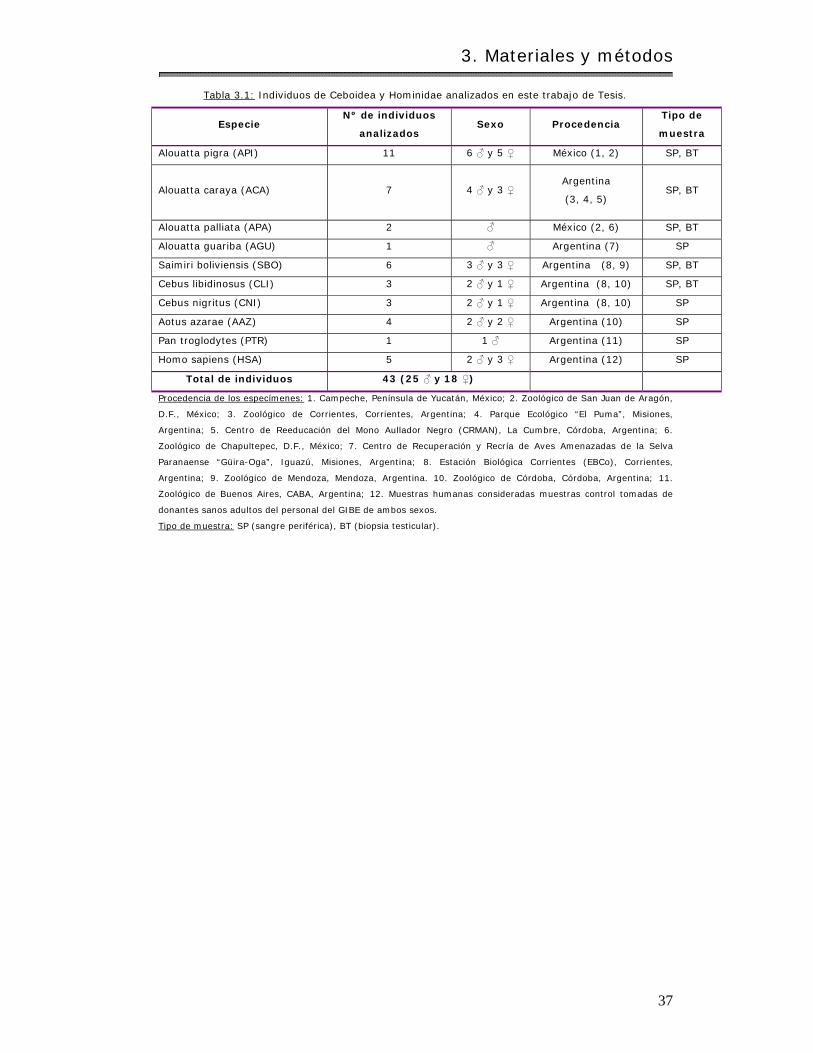
3. Materiales y m étodos
37
Tabla 3.1: I ndividuos de Ceboidea y Hom inidae analizados en este t rabajo de Tesis.
Especie Nº de individuos
analizados Sexo Procedencia
Tipo de
m uest ra
Alouat ta pigra (API ) 11 6 ♂ y 5 ♀ México (1, 2) SP, BT
Alouat ta caraya (ACA)
7
4 ♂ y 3 ♀ Argent ina
(3, 4, 5) SP, BT
Alouat ta palliata (APA) 2 ♂ México (2, 6) SP, BT
Alouat ta guar iba (AGU) 1 ♂ Argent ina (7) SP
Saim ir i boliviensis (SBO) 6 3 ♂ y 3 ♀ Argent ina (8, 9) SP, BT
Cebus libidinosus (CLI ) 3 2 ♂ y 1 ♀ Argent ina ( 8, 10) SP, BT
Cebus nigr itus (CNI ) 3 2 ♂ y 1 ♀ Argent ina ( 8, 10) SP
Aotus azarae (AAZ) 4 2 ♂ y 2 ♀ Argent ina ( 10) SP
Pan t roglodytes (PTR) 1 1 ♂ Argent ina ( 11) SP
Hom o sapiens (HSA) 5 2 ♂ y 3 ♀ Argent ina ( 12) SP
Total de individuos 4 3 ( 2 5 ♂ y 1 8 ♀)
Procedencia de los especímenes: 1. Campeche, Península de Yucatán, México; 2. Zoológico de San Juan de Aragón,
D.F., México; 3. Zoológico de Corr ientes, Corr ientes, Argent ina; 4. Parque Ecológico “El Puma” , Misiones,
Argent ina; 5. Cent ro de Reeducación del Mono Aullador Negro (CRMAN) , La Cum bre, Córdoba, Argent ina; 6.
Zoológico de Chapultepec, D.F., México; 7. Cent ro de Recuperación y Recría de Aves Am enazadas de la Selva
Paranaense “Güira-Oga” , I guazú, Misiones, Argent ina; 8. Estación Biológica Corr ientes (EBCo) , Corr ientes,
Argent ina; 9. Zoológico de Mendoza, Mendoza, Argent ina. 10. Zoológico de Córdoba, Córdoba, Argent ina; 11.
Zoológico de Buenos Aires, CABA, Argent ina; 12. Muestras hum anas consideradas m uest ras cont rol tom adas de
donantes sanos adultos del personal del GI BE de am bos sexos.
Tipo de m uest ra: SP (sangre perifér ica) , BT (biopsia test icular) .

3. Materiales y m étodos
38
3 .2 Diagnosis por caracter ización citogenét ica clás ica
3 .2 .1 Estudios en célu las som át icas
Los protocolos para la preparación de las soluciones empleadas en
los estudios en células som át icas de este t rabajo de Tesis se encuent ran
descritos en el Anexo I I .
3 .2 .1 .1 Ext racción de m uest ras
Los m uest reos se realizaron evitando el sufr im iento innecesario de
los anim ales. La autora de este t rabajo de Tesis y su directora adhieren a
la “Guía para el Cuidado y Uso de Anim ales de Laboratorio” del Consejo
Canadiense para el Cuidado Animal ( “Canadian Council of Anim al Care” ) y a los
Principios de la Sociedad Am ericana de Pr imatología ( “Am erican Society of
Primatologists, ASP” ) para el Tratam iento Ét ico de los Pr imates No Hum anos.
Profesionales veter inarios ext rajeron de ejemplares ceboideos de
am bos sexos, bajo efecto de anestesia (Zelazol, Fort Dodge, o Ketam ina,
Parke Davis, según preferencia del veterinario a cargo, en dosis 10
m g/ kg) ent re 1 y 2 m l de sangre perifér ica con jer inga descartable estér il
(Terum o) heparinizada (Heparina FADA PHARMA 500 UI / m l) . En el caso
de Homo sapiens, personal especializado ext rajo ent re 2 y 5 m l de sangre
per ifér ica con jer inga descartable estéril (Terumo) heparinizada
(Heparina FADA PHARMA 500 UI / m l) .
3 .2 .1 .2 Cult ivo de sangre per ifér ica
Los cult ivos de linfocitos de sangre perifér ica de 72 horas se
realizaron por duplicado a part ir de las m uest ras de sangre perifér ica
entera según la técnica estándar (Mudry et al. 1990) .
Protocolo
1) En ester ilidad, se adicionó ent re 0,8 y 1 m l de sangre perifér ica
entera en frascos de vidr io de 50 m l conteniendo el m edio de

3. Materiales y m étodos
39
cult ivo (ver Anexo I I ) ajustado a un volum en final de 10 m l. Cada
cult ivo se realizó por duplicado.
2) Los cult ivos así preparados fueron incubados en estufa (San Jor)
a 37º C durante 72 horas.
3) Una hora antes de cumplido ese t iem po se adicionó 0,1 m l de
colchicina (Sigm a, 100µg/ m l) en esterilidad.
4) Se regresaron los frascos a la estufa, donde se m antuvieron a
37º C durante una hora y m edia.
5) El m ater ial de cult ivo se t ransfir ió a tubos cónicos de cent rífuga
de 14 m l, ident ificados con el núm ero de m uest ra
correspondiente y se cent r ifugó 10 m inutos a 1800 rpm en una
cent rífuga de m esa (Rolco, Modelo 3070) .
6) Luego de descartar el sobrenadante, a cada tubo cónico se le
adicionaron 8 m l de solución hipotónica (KCl 0,075 M) , dejándose
en esta hipotonía por 30 m inutos en estufa a 37º C. Se detuvo la
hipotonía con 3 gotas de fij ador fr ío (Carnoy 3: 1) .
7) Se repit ió la cent r ifugación y luego de descartar el sobrenadante,
el m aterial se resuspendió en 8 m l de fij ador fr ío (Carnoy 3: 1) .
Estas fij aciones se llevaron a heladera (4º C) por 30 m inutos.
8) Luego de ese lapso se realizaron lavados consistentes en
cent r ifugar 10 m inutos a 1800 rpm, descartar el sobrenadante y
agregar 8 m l de fij ador. Esta operación se repit ió hasta obtener
un botón celular blancuzco y un sobrenadante t raslúcido t ratando
de perder la m enor proporción posible de m aterial para estudio
citogenét ico.
9) Luego del últ imo lavado, el botón celular se resuspendió en una
cant idad apropiada de fijador, según el tam año del m ismo,
considerándose com o adecuado un volum en de fij ador dos veces
superior al tam año del botón celular obtenido.
10) Las preparaciones cromosóm icas se realizaron sobre portaobjetos
previam ente desengrasados y conservados en etanol a 4º C. Se
gotearon 12 µl (m edidos con m icropipeta Gilson de 20 µl) de
suspensión celular sobre portaobjetos em pañados con vapor,
dejándose secar al aire el extendido así obtenido.

3. Materiales y m étodos
40
11) Estos extendidos se colorearon durante 5 m inutos en Giem sa
10% en agua corr iente.
12) Los extendidos se lavaron con agua corr iente, se secaron al aire
y se observaron al m icroscopio ópt ico a 400x (Leica DMLB) para
corroborar el estado del cult ivo y proceder al análisis
citogenét ico.
3 .2 .1 .3 Análisis de m etafases
3 .2 .1 .3 .1 Determ inación del índice m itó t ico ( I M% )
Para caracter izar el estado y rendim iento celular del cult ivo, en los
extendidos realizados a part ir de cada tubo de cult ivo se determ inó el
índice m itót ico ( I M% = porcentaje de metafases observadas / núcleos
est im ulados) . Se contó un total de 1000 núcleos est im ulados en cada uno
de los individuos cuya sangre perifér ica entera fuera cult ivada por
duplicado para lo cual se preparararon extendidos de los dos tubos de
cult ivo y se analizaron a ciegas para evitar sesgos de interpretación en la
act iv idad de reconocer y contar las m etafases. El operador siem pre fue el
m ism o, es decir la t itular y responsable de los hallazgos reunidos en este
t rabajo de Tesis.
3 .2 .1 .3 .2 Determ inación del núm ero m odal ( 2 N) y
fundam enta l ( NF)
Para establecer el núm ero modal o diploide (2N) y el núm ero de
brazos o fundam ental (NF) correspondiente a los individuos de am bos
sexos de cada especie en estudio se analizaron con m icroscopía ópt ica
(Leica DMLB) al m enos 50 m etafases de cada ejem plar y se regist raron
los datos en una tabla según m odelo descrito en la Tabla 3.2.

3. Materiales y m étodos
41
Tabla 3.2: Modelo de tabla utilizado para la determinación del número diploide (2N), ejemplificado para el caso de Alouatta caraya (2N= 52)
2N ≤ 49 50 51 52 53 54 ≥ 55 Total
Nº DE CÉLULAS
1 2 6 33 4 2 2
50
PORCENTAJE (%) 2 4 12 66 8 4 4 100
Hipoploidías: 9 (18%) Hiperploidías: 8 (16%)
3 .2 .1 .4 Var iables crom osóm icas
3 .2 .1 .4 .1 Mediciones crom osóm ica s
El conjunto de crom osom as de una célula const ituye el
com plem ento cromosómico. El estudio de sus característ icas m orfológicas
externas ( form a, tam año y núm ero) se lleva a cabo ordenando los pares
de cromosom as hom ólogos de acuerdo a su tam año y form a, lo que
const ituye el cariot ipo de la especie en estudio (Lacadena 1996) . El cariot ipo
se determ ina para cada especie y se ut ilizan para ello parám et ros
m itót icos com o el núm ero m odal o diploide (2N) , el núm ero fundam ental
(NF) , las m edidas cromosóm icas (Porcentaje de la Longitud Total del
Com plem ento o “% LTC” , relación de brazos o “ r” y el Í ndice Cent rom érico
o “ I C” ) en t inción uniform e o clásica y luego se aplican técnicas de
bandas (G, C, DAPI , NOR, Q, R) a fin de confirm ar lo observado y
regist rado con t inción clásica. Los crom osom as m etafásicos están
form ados por dos brazos y su morfología está dada por la posición del
cent róm ero y la relación de brazos en función de la const r icción prim aria
(Lacadena, 1996) . Para la com paración de los cariot ipos de dist intas
especies o taxa en estudio, se realizaron los análisis cromosóm icos
teniendo en cuenta los parám et ros siguientes (Levan et al. 1964 m od.) :
• I ndice cent rom érico ( I C)
I C =
Donde
C = longitud total del crom osom a
p = longitud del brazo corto ( “p” ) .
p x 100
C

3. Materiales y m étodos
42
• Relación de brazos ( r)
Este índice determ ina la m orfología crom osóm ica.
r =
Donde
p = longitud del brazo corto
q = longitud del brazo largo
En base a los valores del I C y el r se determ inó la morfología
cromosóm ica ut ilizando com o referencia los valores de la Tabla 3.3.
Tabla 3.3: Clasif icación morfológica de los cromosom as (Levan et al. 1964 m odif.)
Mor fología I C r
Metacént r ico 0,50 1,00
Subm etacént r ico 0,25< I C< 0,37 1,71< r< 3,00
Acrocént r ico 0,00< I C< 0,12 - - -
I C: índice cent rom érico; r : relación de brazos
• Porcentaje de la Longitud total del Com plem ento (% LTC)
% LTC =
Donde
LT= largo total del com plem ento
C = longitud del cromosoma
Este parám et ro representa la proporción que ocupa cada
cromosom a en el com plem ento cromosóm ico teniendo en cuenta, según
acuerdo internacional, el cromosom a X. En este t rabajo de Tesis se
incluyeron las m edidas correspondientes al crom osom a X de cada
ejemplar estudiado.
Actualm ente existen programas para la m edición de
cromosom as, tales com o el Microm easure v3.3
(ht tp: / / www.colostate.edu/ Depts/ Biology/ MicroMeasure) , que realiza m ediciones a part ir
de fotografías de las m etafases seleccionadas para su m edición, tom adas
q
p
C x 100
LT

3. Materiales y m étodos
43
todas en la m ism a escala y con la m ism a m icroscopía (Leica DMLB;
X1000) .
En este t rabajo de Tesis para las m ediciones cromosóm icas se
emplearon dos m etodologías: la clásica (m edición con papel m ilimetrado)
y la inform át ica (Micromeasure) . Se compararon las m ediciones
obtenidas con am bas m etodologías y se corroboró que ambas brindaron
resultados que no m ostraron diferencias estadíst icas significat ivas luego
de aplicar un análisis de Chi cuadrado (Excel, Microsoft corp.) .
Para la m edición clásica, se realizaron las m edidas crom osómicas a
part ir de m etafases de los ejem plares estudiados. Dado que el grado de
cont racción cromosómica es var iable en cada m etafase, se m idieron los
dos cromosom as hom ólogos para cada par y para cada m etafase en 3
m etafases con t inción convencional para cada espécimen, ut ilizando
papel m ilim et rado. Luego se calcularon los prom edios de esas m ediciones
y los respect ivos errores y se hizo la tabla de valores correspondientes a
cada par cromosóm ico para los ejemplares estudiados.
3 .2 .1 .4 .2 Bandas G co n t inción de W right ( García Haro 2 0 0 1 m od.)
Procedim iento
1) Los extendidos ut ilizados tenían al m enos 2 días de envejecim iento
mantenidos a temperatura ambiente.
2) Se int rodujo el extendido en un vaso de Coplin con 2xSSC a 65º C,
com enzando con un t iem po m ínim o de 10 segundos (este t iem po
es variable según el envejecim iento del extendido y la ant igüedad
de la fij ación con la que fuera realizado el preparado) .
3) Se lavó el extendido con abundante agua dest ilada y se dejó secar
al aire.
4) Se preparó la solución de colorante de Wright en el m om ento de
realizar la t inción.
5) Se cubrió uniform em ente el extendido con la solución colorante y
se dejó actuar 2 m inutos y m edio, luego de lo cual se ret iró el
colorante con agua corr iente.

3. Materiales y m étodos
44
6) Se dejó secar al aire y se observó con m icroscopía ópt ica (Leica
DMLB a X1000) .
3 .2 .1 .4 .3 Bandas G con t r ipsina ( Seabr ight 1 9 7 1 m od.)
Procedim iento
1) Los extendidos ut ilizados tenían al m enos 1 día de envejecim iento
a tem peratura ambiente (se considera un estándar de 25º C) . El
día previo a la realización de la técnica se colocaron los extendidos
en estufa a 60º C toda la noche para completar el proceso de
envejecim iento y asegurar que el extendido estuviera
completam ente seco al m om ento de realizar el protocolo.
2) Se int rodujo el extendido en un vaso de Coplin con PBS sin Ca2+ ni
Mg2+ a tem peratura ambiente por 10 m inutos.
3) Se t ransfir ió inm ediatam ente el extendido a una solución de
t r ipsina 0,05% en agua dest ilada a tem peratura am biente. El
t iem po m ínim o ut ilizado fue de 10 segundos (variable según el
envejecim iento del extendido y la ant igüedad de la fij ación
ut ilizada) .
4) Se ret iró lentam ente el extendido de la solución de t r ipsina. Se lo
observó con m icroscopio ópt ico bajo cont raste de fases para ver el
estado de los crom osom as y el grado de digest ión alcanzado.
5) Se coloreó el extendido con Giem sa 3% en agua corr iente durante
3 m inutos y se observó con m icroscopía ópt ica (Leica DMLB a
X1000) .
3 .2 .1 .4 .4 Bandas C ( Sum ner 1 9 7 2 )
Procedim iento
1) Se ut ilizaron extendidos con por lo m enos 1 día de envejecim iento
y no m ás de 2 m eses de ant igüedad.
2) Se colocaron los extendidos en HCl 0,2 N en un vaso de Coplin
durante 20 m inutos a tem peratura am biente.
3) Pasado ese t iem po se lavaron con abundante agua dest ilada.

3. Materiales y m étodos
45
4) Se colocaron los extendidos en Ba(OH) 2 al 5% en “baño María” a
50º C. El t iem po de digest ión dependió del t iem po de
envejecim iento de los preparados, el t ipo celular y la especie en
estudio, pudiendo variar de 10 segundos a 3 m inutos.
5) Se lavaron con abundante agua dest ilada en un vaso de Coplin.
6) Se sum ergieron en solución 2xSSC a 60º C por 1 hora.
7) Pasado este lapso, se lavaron con abundante agua corr iente.
8) Se colorearon con Giem sa 3% en agua corr iente durante 5
m inutos.
9) Se lavó el colorante con abundante agua corr iente y los preparados
se dejaron secar al aire. Se observó cada preparado con
m icroscopía ópt ica (Leica DMLB a X1000) .
3 .2 .1 .4 .5 Bandas NOR ( How ell y Black 1 9 8 0 )
Procedim iento
1) Se colocaron sobre el extendido 30 μl de solución gelat ina (Sigm a)
y 20 μl de solución de nit rato de plata (Sigm a) , am bas preparadas
según protocolo descrito en el Anexo I I .
2) Se colocó sobre el extendido un cubreobjeto previam ente
sum ergido en alcohol y secado con una gasa lim pia.
3) Se calentó el extendido en cám ara húm eda dent ro de un baño
térm ico a 60º C.
4) Se cont roló cada 5 m inutos la t inción con m icroscopio ópt ico (Leica
DMLB a X1000) . Cuando se alcanzó la intensidad de coloración
deseada se removió el cubreobjeto haciéndolo flotar en un
recipiente con agua dest ilada.
5) Se lavó cuidadosam ente el extendido con agua dest ilada.
6) Se dejó secar al aire y se observó con m icroscopía ópt ica (Leica
DMLB a X1000) .
3 .2 .1 .5 Arm ado y confección de los car i ot ipos
Se fotografiaron (ver apartado 3.5) al m enos 10 m etafases con
t inción uniform e, 10 m etafases con bandas G y 10 m etafases con

3. Materiales y m étodos
46
bandas C por individuo de cada especie y subespecie analizada. Estas
fotografías fueron almacenadas en el archivo digital del GI BE y se
hicieron copias de las m ism as en papel fotográfico.
Se ordenaron las m etafases con t inción estándar considerando los
cromosom as por t ipo de morfología (según posición del cent róm ero:
m orfologías m etacéntr ica, subm etacént r ica y acrocént r ica) y en orden de
tam año decreciente para cada t ipo de m orfología. En el caso de las
m etafases con t inción de bandas G, para su arm ado se consideró el
pat rón de bandas longitudinales, oscuras y claras, puesto en evidencia
por esta técnica y característ ico de cada crom osom a.
Estas m etafases así arm adas fueron archivadas en una carpeta
para cada ejemplar. En cada carpeta figura una ficha con los parám et ros
citogenét icos analizados, según el siguiente modelo de la Figura 3.1.
Figura 3.1: Modelo de ficha ut ilizar para regist rar los parámet ros citogenét icos analizados en
este t rabajo de Tesis (confeccionada por la Dra. M. Nieves, 2000) .
Individuo: Origen:
Sexo:
Cultivo Fecha de entrada ........./ ......../ .........
Fecha de salida ......../ ........./ ......... Resultado
Indice mitótico
Número modal
2N
Nº DE CÉLULAS
PORCENTAJE
Técnicas de bandas
CONVENCIONAL BG BC G/C SEC. BG= bandas G; BC= bandas C; G/C= bandas G/C

3. Materiales y m étodos
47
3 .2 .1 .6 Hom eologías crom osóm icas
Se realizó el análisis de las hom eologías crom osóm icas basado en
la comparación del pat rón de bandas G de las siguientes especies y
subespecies:
-Alouat ta caraya (ACA)
-Alouat ta pigra (API )
-Alouat ta palliata (APA)
-Alouat ta guar iba clam itans (AGUc)
-Alouat ta guar iba guar iba (AGUg)
-Alouat ta m acconelli (AMA)
-Alouat ta seniculus seniculus (ASEs)
-Alouat ta seniculus arctoidea (ASEa)
-Alouat ta sara (ASA)
-Alouat ta belzebul (ABE)
-Homo sapiens (HSA)
Una vez ordenadas las m etafases de los ejemplares procesados en
este t rabajo de Tesis (ACA, API , APA y AGUc) siguiendo el protocolo
descripto en el apartado 3.2.4.5 se realizó la comparación de los pat rones
de bandas G. Se ut ilizaron los cariot ipos ya descritos para ACA, AGUg y
AGUc (de Oliveira et al. 2002; Stanyon et al. 2011), AMA (de Oliveira et al. 2002) , ASEa
(Consigliere et al. 1996) , ASEs (Torres y Leibovici 2001) , ABE (Consigliere et al. 1998), y ASA
(Consigliere et al. 1996) .

3. Materiales y m étodos
48
3 .2 .2 Estudios en célu las germ inales
Los protocolos para la preparación de las soluciones empleadas en
los estudios en células som át icas de este t rabajo de Tesis se encuent ran
descritos en el Anexo I I .
3 .2 .2 .1 Ext racción de las m uest ras
Con el objet ivo de realizar la caracterización meiót ica y confirm ar
el sistem a de determ inación sexual observado en los estudios m itót icos, a
cada ejem plar m acho adulto de las especies Alouat ta caraya, A. pigra, A.
palliata, Saim ir i boliviensis boliviensis y Cebus libidinosus se le realizó
una biopsia test icular de 6 m m 3 bajo efecto de anestesia (Zelazol, Fort
Dodge, en dosis 10m g/ kg) . Las 8 biopsias fueron realizadas por m édicos
veterinarios pertenecientes al grupo estable de profesionales de cada una
de las inst ituciones en las que se alojaban los ejemplares, respetando las
condiciones de esterilidad requeridas para el estudio citogenét ica de los
m ism os.
En el caso de los ejem plares de Alouat ta caraya y Cebus libidinosus
la biopsia ext raída se dividió en dos partes: una porción se dest inó al
análisis bajo m icroscopía ópt ica y la ot ra porción se dest inó al análisis
bajo m icroscopía de fluorescencia.
3 .2 .2 .2 Análisis con m icroscopía ópt ica
Com o protocolo para la extensión de esperm atocitos para su
análisis con m icroscopía ópt ica se ut ilizó Evans et al. (1964) m odificado
para m axim izar la eficiencia de los análisis considerando experiencias
previas del grupo de t rabajo en ot ros Ceboidea (Steinberg et al. 2007) . Estos
estudios previos nos perm it ieron corroborar que el procedim iento, cuando
es realizado en ester ilidad, no ocasiona una reducción en la fert ilidad del
ejemplar ni ot ro t ipo de inconveniente o incom pat ibilidad con el norm al
desarrollo del m ismo.

3. Materiales y m étodos
49
El m aterial de biopsia fue procesado en un lapso no m ayor a 3
horas después de su ext racción.
Procedim iento
1) Una vez obtenida la biopsia se la liberó de grasa y de ot ros tej idos,
removiendo la túnica albugínea, dejando expuestos los túbulos
sem iníferos.
2) Se colocó el tej ido en cit rato t r isódico 1% en caja de Petr i,
resuspendiendo suavem ente con pipeta Pasteur.
3) Se dejó en esta solución hipotónica por 20 m inutos. Durante este
periodo se m aceraron y vaciaron los túbulos con dos agujas de
disección.
4) Se t ransfir ió la suspensión a un tubo de cent rífuga de 14 m l y se
cent r ifugó 5 m inutos a 500 rpm en una cent rífuga de m esa (Rolco,
Modelo 3070) .
5) Una vez descartado el sobrenadante, se agregó lentam ente en
cada tubo 1 m l de fij ador a 4º C (Carnoy 3: 1) , resuspendiendo, y
se dejó fijando el m aterial a 4º C aproxim adam ente 30-40 m inutos.
6) Se cent r ifugó nuevam ente durante 5 m inutos a 500 rpm ,
descartando el sobrenadante. Este lavado se repit ió 4 veces, luego
de lo cual se resuspendió el botón celular en una cant idad
apropiada de fij ador, dependiendo del tam año del m ismo,
considerándose com o adecuado un volum en de fij ador dos veces
superior a su tam año.
7) Se gotearon t res o cuat ro gotas de la suspensión celular en un
portaobjeto limpio y fr ío y se secaron los preparados en corr iente
de aire caliente.
8) El 70% de los preparados se colorearon con Giemsa al 5% en agua
dest ilada. El 30% restante se dejó envejecer para som eter los a la
técnica de bandas C (Sumner 1972) .

3. Materiales y m étodos
50
3 .3 Diagnosis por caract er ización citom olecular
3 .3 .1 Estudios en célu las som át icas
3 .3 .1 .1 H ibr idación in situ Fluorescente ( FI SH)
Para la realización de estos estudios se contó con la colaboración
de la Dra. Mariela Nieves (GI BE-FCEyN-UBA) y el asesoram iento del
Lic. Alejandro Laudicina (Laborator io de Citogenét ica Molecular, I NTI ) .
3 .3 .1 .1 .1 Sondas
-Sonda de Locus SRY hum ano (LSI -SRY Spect rum Orange,
Vysis, USA) .
-Sondas de crom osom a entero de los crom osom as 3, 15, 21, X
e Y hum anos (PCT3 Cy3, PCT15 FI TC, PCT21 FI TC, PCTX FI TC, PCTY Cy3,
Lexel S.R.L., Argent ina) .
-Sonda pantelom érica de t ipo PNA ( telóm eros sintét icos en
los que los grupos fosfato han sido reem plazados por una cadena de
pépt ido/ poliam ida neut ra) com ercial (Telom ere PNA FI SH kit / Cy3, DAKO,
Glost rup, Dinam arca) .
3 .3 .1 .1 .2 Extendidos
3 .3 .1 .1 .2 .1 Preparación
Las preparaciones cromosóm icas se realizaron sobre portaobjetos
previam ente desengrasados y conservados en etanol a 4º C. Para
asegurar el éxito de la técnica fue necesario disponer de preparaciones
cromosóm icas limpias y libres de restos celulares, dado que estos
interfieren en la hibr idación, evitando que la sonda pueda hibridar con el
ADN blanco.

3. Materiales y m étodos
51
Con este objet ivo, para la concreción de la técnica de FI SH se
ut ilizaron dos protocolos:
Extendidos crom osóm icos fij ados en Carnoy: Se gotearon 12 µl de
suspensión celular preparada en solución de Carnoy 3: 1 sobre vidr ios
empañados con vapor. Se dejó secar al aire los preparados. Con esta
técnica se obtuvieron resultados sat isfactor ios tanto para la técnica de
desnaturalización con Form am ida com o para la desnaturalización con
Hidróxido de Sodio.
Extendidos crom osómicos resuspendidos en acét ico 60% : Se
gotearon 12 µl de suspensión celular resuspendida en solución de ácido
acét ico al 60% en agua dest ilada sobre vidrios em pañados con vapor.
I nm ediatam ente después se gotearon sobre el extendido 60 µl de fij ador
Carnoy recién preparado. Se dejó secar al aire. Esta técnica es m ás
eficiente para liberar los extendidos de restos de citoplasm a y se
obtuvieron resultados sat isfactorios para la técnica de desnaturalización
con Form am ida, no así para la técnica con la desnaturalización con
Hidróxido de Sodio.
3 .3 .1 .1 .2 .2 Envejecim iento
El t ratam iento de desnaturalización del ADN blanco puede llevar a
la pérdida de morfología crom osómica, generando poster iorm ente una
reducción en la señal de hibr idación. El envejecim iento de los extendidos
previo a la desnaturalización evita la pérdida de morfología cromosómica.
En este t rabajo de Tesis se ut ilizaron diferentes est rategias que se
aplicaron según el éxito obtenido con cada una de ellas en cada individuo
y/ o especie (ver Anexo I I I ) .
Para la hibr idación con la sonda pantelomérica los preparados
fueron realizados en Carnoy 48 horas antes de realizar la técnica,
m antenidos hasta ese m om ento a tem peratura ambiente.

3. Materiales y m étodos
52
3 .3 .1 .1 .3 Hibr idación
Para la Hibr idación in situ sobre ADN cromosómico, el ADN blanco
y la sonda deben estar desnaturalizados (es decir , en estado de simple
cadena) para así poder hibr idar ent re sí. El alcanzar una adecuada
hibridación depende de la capacidad del ADN desnaturalizado disponible
para renaturalizar con una cadena com plem entaria a una tem peratura
debajo de su temperatura de desnaturalización (Tm o “m elt ing point ” , la
temperatura a la cual el 50% del ADN está en form a de simple cadena) .
Las temperaturas de desnaturalización del ADN son altas (superiores a
85º C en la m ayoría de los casos) y a estas tem peraturas se pierde
m orfología crom osómica. La presencia del solvente orgánico Form amida
en la m ezcla de hibr idación y en la solución de desnaturalización reduce
la tem peratura de desnaturalización del ADN, perm it iendo así conservar
una adecuada morfología crom osóm ica (Boehringer Mannheim 1995;
Schwarzacher 2000) .
Dos m etodologías se ut ilizan para desnaturalizar el ADN blanco: la
desnaturalización sim ultánea con la sonda (en cuyo caso se ut iliza una
concent ración m enor de Form am ida y una tem peratura m ayor) y la
desnaturalización por separado. En este últ im o caso varía tanto la
temperatura com o el t iempo de desnaturalización, si bien se m ant iene
constante la concent ración de Form am ida (Boehringer Mannheim 1995;
Schwarzacher 2000) . Recientem ente se ha desarrollado ot ra m etodología
para la desnaturalización por separado del ADN blanco ut ilizando
Hidróxido de Sodio (NaOH) como solvente orgánico (Laudicina,
com unicación personal) . La ventaja de esta últ im a est rategia es que se
realiza a tem peratura ambiente y que luego de alcanzada una correcta
desnaturalización, los crom osomas exhiben un pat rón de bandas sim ilar
al obtenido m ediante las técnicas de bandas G o C (el t ipo de pat rón
obtenido depende de las condiciones de hum edad y tem peratura
am biental) .
Se llevaron a cabo t res est rategias de hibr idación, que fueron
aplicadas según el éxito obtenido con cada est rategia en cada individuo

3. Materiales y m étodos
53
y/ o especie. Com o pr imera aproximación se ut ilizó la desnaturalización
con Form am ida (est rategia 1) com enzando con el protocolo de
desnaturalización por separado. Si con esta técnica no se obtenía una
buena desnaturalización de los extendidos se intentaba el m étodo de
desnaturalización conjunta con Form amida. De lo cont rar io se ut ilizaba la
est rategia de desnaturalización con Hidróxido de Sodio (est rategia 2) .
Para el caso de la sonda pantelom érica para la hibridación se siguió
el protocolo sugerido por el fabricante (DAKO, Glost rup, Dinamarca) . Para
esta m etodología se contó con la colaboración del Dr. Alejandro Bolzán,
del I MBI CE-CONI CET, La Plata.
3 .3 .1 .1 .3 .1 Est ra tegia 1 : Desnatura lización con form am ida 3 .3 .1 .1 .3 .1 .1 Desnatura lización por separado
Procedim iento
1) Las regiones a hibridar se m arcaron sobre los extendidos con lápiz
con punta de diam ante para vidr io en la cara opuesta del
portaobjeto que soporta dicha preparación.
2) Los extendidos se sum ergieron en Formamida 70% en 2xSSC a
PH= 7 a una tem peratura de ent re 56° y 70º C, durante 15
segundos a 5 m inutos (según el envejecim iento del extendido y la
ant igüedad de la fij ación) .
3) El extendido se enjuagó en etanol 70% agitando durante 2
segundos, para quitar el exceso de Form amida.
4) El extendido se deshidrató en alcoholes de concent ración creciente
(etanol 70% , etanol 90% y etanol 100% , en ese orden) 2 m inutos
en cada uno.
5) Se dejó secar al aire.
6) En un tubo de 0,5 m l se m ezcló 0,5 µl de sonda + 0,5 µl de agua
MiliQ + 2,5 µl de Mezcla de Hibridación (LiVe) , en el caso de
hibr idar con una sola sonda. Si se hibr idó con dos sondas se
preparó la solución con 0,5 µl de la sonda 1 + 0,5 µl de la sonda 2
+ 2,5 µl de Mezcla de Hibridación (LiVe) .

3. Materiales y m étodos
54
7) Se calentó esta solución durante 7 m inutos a 94º C. Se dejó
enfr iar.
8) Se agregó esta solución de sonda sobre el extendido en la región a
hibr idar previam ente m arcada.
9) Se cubrió con un cubreobjeto de tam año adecuado para abarcar
totalm ente la región a hibridar y se fijó todo el contorno del
cubreobjeto con pegam ento sintét ico (Hey-Cola, Heyco) de m anera
de evitar el drenaje de sonda.
10) El extendido sellado se incubó dent ro de una cám ara húm eda en
estufa (San Jor) a 45º C durante un m ínimo de 16 y un m áxim o de
72 horas.
3 .3 .1 .1 .3 .1 .2 Desnatura lización conjunta
Procedim iento
1) Se m arcaron sobre el extendido las regiones a hibridar con lápiz
con punta de diam ante para vidrio sobre la cara opuesta del
portaobjeto que soporta dicha preparación.
2) En un tubo de 0,5 m l se m ezclaron 0,5 µl de sonda + 3 µl de
Mezcla de Hibridación (LiVe) , en el caso de hibridar con una sola
sonda. Si se hibr idó con dos sondas se preparó la solución con 0,5
µl de la sonda 1 + 0,5 µl de la sonda 2 + 3 µl de Mezcla de
Hibridación (LiVe) .
3) Esta solución se calentó durante 7 m inutos a 94º C. Se dejó enfr iar.
4) La solución de sonda se agregó sobre la región a hibr idar.
5) Se cubrió con un cubreobjeto de tam año adecuado para que
abarcara totalmente la región a hibridar y se fij ó todo el contorno
del cubreobjeto con pegam ento sintét ico (Hey-Cola, Heyco) de
manera de evitar el drenaje de sonda.
6) La desnaturalización conjunta se realizó en un termociclador
BOECO con adaptador para hibridación in situ. El t iempo de
desnaturalización varió ent re 5 y 8 m inutos a 70°-7 8°C,
dependiendo de la especie de prim ate en estudio.

3. Materiales y m étodos
55
7) El extendido sellado se incubó dentro de una cám ara húm eda en
estufa (San Jor) a 45º C durante un m ínimo de 16 y un m áxim o de
72 horas.
3 .3 .1 .1 .4 .2 Est ra tegia 2 : Desnatura l ización con
Hidróx ido de sodio ( NaOH)
Procedim iento
1) Las regiones a hibridar se m arcaron sobre el extendido con lápiz
con punta de diam ante para vidrio sobre la cara opuesta del
portaobjeto que soporta dicha preparación.
2) El extendido se colocó en NaOH 100m M durante 3 segundos a 10
m inutos (según el envejecim iento del extendido y de la ant igüedad
de la fij ación) .
3) El extendido se lavó con abundante agua de la canilla.
4) Este preparado se deshidrató con alcoholes de concent raciones
crecientes (Etanol 70% , 90% , 100% ) por 2 m inutos en cada
alcohol
5) Se dejó secar al aire.
6) En un tubo de 0,5 m l se m ezclaron 0,5 µl de sonda + 0,5 µl de
agua MiliQ + 2,5 µl de Mezcla de Hibridación (LiVe), en el caso de
hibr idar con una sola sonda. Si se hibr idó con dos sondas se
preparó la solución con 0,5 µl de la sonda 1 + 0,5 µl de la sonda 2
+ 2,5 µl de Mezcla de Hibridación (LiVe) .
7) Esta solución se calentó durante 7 m inutos a 94º C. Se dejó enfr iar.
8) La m ezcla de hibridación se colocó sobre la región a hibridar y se
cubrió el extendido con un cubreobjeto. Todo el contorno del
cubreobjeto se fijó con pegam ento sintét ico (Hey-Cola, Heyco) de
manera de evitar el drenaje de sonda.
9) El preparado sellado se incubó dent ro de una cám ara húm eda en
estufa (San Jor) a 45º C durante un m ínimo de 16 y un m áxim o de
72 horas.

3. Materiales y m étodos
56
3 .3 .1 .1 .5 Revelado
Procedim iento 1) Se ret iró cuidadosam ente el pegam ento sintét ico de los bordes del
cubreobjeto.
2) Se removió el cubreobjeto sum ergiendo el preparado en 2XSSC a
tem peratura am biente durante algunos m inutos.
3) Se realizó un enjuague en un vaso de Coplin con 0,4XSSC + 0,3%
Tween 20 en baño térm ico (Vicking) a 70º C por 2 m inutos.
4) Se t ransifir ió el extendido a un vaso de Coplin con 2xSSC + 0,1%
Tween 20 a tem peratura am biente y se incubó por 2 m inutos.
5) Se escurr ió el extendido.
6) Se agregaron 15 µl de solución de t rabajo de DAPI .
7) Se cubrió con un cubreobjeto de tam año adecuado para abarcar
totalm ente la región hibr idada y se presionó suavem ente con papel
secante para elim inar el excedente de DAPI .
8) El análisis se realizó en un m icroscopio Leica DLMB equipado con
filt ros para fluorescencia (ver anexo I V) y con una cám ara DFC 340
FX (x1000) . Se fotografiaron 10 m etafases con cada filt ro, las que
luego fueron analizadas ut ilizando los program as Photoshop CS
(Adobe) e I m age Pro-Plus 4.5 (Media Cybernet ics I nc.) .
9) Luego de la observación en el m icroscopio de fluorescencia se
guardaron los preparados envueltos en papel secante a –20º C.
3 .3 .2 Estudios en células germ ina les
3 .3 .2 .1 Análisis con m icroscopía de f luorescen cia
Para este análisis se contó con la colaboración del equipo de
invest igadores de la Unitat de Biologia Cellular i Genèt ica Mèdica, Facultat
de Medicina, de la Universidad Autónom a de Barcelona: Dra. Montserrat
García Caldés, Dra. Aurora Ruiz-Herrera Moreno, Dra. Raquel García-
Cruz, Dr. Roig I gnasi, Dr. Pedro Robles, Dra. Nuria Cam ats, Dr. Miguel
Angel Brieño y Magíster Sarai Pacheco Piñol en el m arco de un convenio

3. Materiales y m étodos
57
interinst itucional de intercam bio de experiencias citomoleculares con
m aterial humano y de ot ros pr imates.
Las secciones de cada biopsia dest inadas a este análisis fueron
congeladas en 1,5 m l de I sopentano (Merck) a -80º C inm ediatam ente
después de su ext racción.
Com o protocolo para la extensión de esperm atocitos para su
análisis mediante inm unofluorescencia se ut ilizó Roig et al. (2004) . Los
extendidos fueron realizados y analizados en la Unitat de Biologia Cellular
i Genèt ica Mèdica, Facultat de Medicina, de la Universidad Autónom a de
Barcelona, España.
3 .3 .2 .1 .1 Extensión de esperm atocitos
Protocolo
1) Cortar un t rozo de biopsia test icular congelado a -80º C sobre un
portaobjeto.
2) Disgregar el tej ido por todo el portaobjeto y añadir 20 μl de PBS
para que no quede seco.
3) Transferir 5 μl de la anterior suspensión celular sobre ot ros
portaobjetos.
4) Añadir sobre cada extendido 80 μl de solución hipotónica (Lipsol
1% en agua desionizada) . Dejar actuar durante 15 m inutos.
5) Añadir 90 ul de solución fij adora (ver Anexo I I ) .
6) Mantener los extendidos con esta solución fij adora durante 2 horas
en una cám ara húm eda parcialmente abierta a tem peratura
am biente.
7) Destapar la cám ara húm eda y dejar que los extendidos se sequen
a tem peratura am biente.
8) Lavar 4 veces en Agepon (AGFA) al 1% en agua bidest ilada
durante un m inuto.
9) Realizar la técnica de I nm unofluorescencia en el m om ento
(protocolo 3.2.7.) o guardar los portabojetos a -20º C.

3. Materiales y m étodos
58
3 .3 .2 .1 .2 I nm unofluorescencia
3 .3 .2 .1 .2 .1 Ant icuerpos pr im ar ios
-Suero policlonal de ratón cont ra la cohesina REC8 (gent ilm ente
cedidos por JL.Barbero) .
-Suero policlonal de conejo cont ra la proteína SYCP1 del elem ento
cent ral del complejo sinaptoném ico (gent ilm ente cedido por C.Heyt ing) .
-Suero policlonal de conejo cont ra la proteína SYCP3 de los
elem entos laterales del com plejo sinaptoném ico (gent ilm ente cedido por
C.Heyt ing) .
-Ant icuerpo policlonal de conejo cont ra BRCA1 (Abcam ) , proteína de
la m aquinaria de “ checkpoint ” celular.
-Ant icuerpo m onoclonal de ratón cont ra MLH1 (Pharm ingen) ,
m arcadora de sit ios de recombinación.
-Ant icuerpo m onoclonal de ratón cont ra RPA (Calbiochem ) ,
com ponente de los nódulos m eiót icos t ransicionales.
-Suero policlonal cont ra la proteína CenP proveniente de suero
autoinm une hum ano de un paciente con síndrom e de CREST (cedido
gent ilm ente por M. Frit zler) , m arcadora de la posición de los
cent róm eros.
3 .3 .2 .1 .2 .2 Ant icuerpos secundarios ut ili zados
-Ant icuerpo m onoclonal de cabra cont ra conejo conjugado con el
fluorocrom o Cy3 (Jackson I m m unoResearch Laboratories) .

3. Materiales y m étodos
59
-Ant icuerpo m onoclonal de cabra cont ra hum ano conjugado con el
fluorocrom o Cy5 (Jackson I m m unoResearch Laboratories) .
-Ant icuerpo m onoclonal de cabra cont ra ratón conjugado con el
fluorocrom o FI TC (Jackson I m m unoResearch Laborator ies) .
3 .3 .2 .1 .2 .3 Protocolo
1) Los ant icuerpos y sueros pr imarios fueron diluidos en “buffer”
PBTG en proporciones 1/ 200 (REC8, SYCP3, CenP, BRCA1) y 1/ 100
(MLH1, RPA, ARN Pol I I , SYCP1 y ant icuerpos secundarios) .
2) Los ant icuerpos pr im arios se incubaron toda la noche a 4º C en
cám ara húm eda. Los ant icuerpos secundarios se incubaron 1 hora
a 37º C en cám ara húm eda.
3) Se realizaron 2 lavados de 5 m inutos cada uno en “buffer” PBTG
con agitación para elim inar el exceso de ant icuerpo secundario.
4) Se fij ó la señal del ant icuerpo secundario aplicando una solución de
Form aldehído al 1% en PBS e incubando durante 10 m inutos a
tem peratura ambiente.
5) Se lavó el portaobjetos en glicina al 0,5% en PBS en agitación
durante 5 m inutos.
6) Se lavó el portaobjetos con PBTG a 37º C en agitación durante 10
m inutos.
7) Se aplicaron 30 μl de solución com puesta por 0,1 μg/ m l de DAPI
en Vectashield.
3 .3 .2 .1 .3 H ibr idación in situ
3 .3 .2 .1 .3 .1 Sondas
-Sonda gen específica 21q22 (CAMBI O) .
-Sonda de crom osom a entero hum ano X (CAMBI O) .

3. Materiales y m étodos
60
3 .3 .2 .1 .3 .2 FI SH
1) Se lavó el extendido durante 2 m inutos en agua dest ilada.
2) Se dejó secar al aire.
3) Se desnaturalizó el extendido durante 5 m inutos en form am ida
70% en 2xSSC.
4) Se lavó el m aterial durante un m inuto en agua dest ilada.
5) Se aplicó sobre el extendido 100μl de NaSCN 1M y se incubó
durante 3 horas en cám ara húm eda a 65º C.
6) Se dejó secar al aire.
7) Se desnaturalizó nuevam ente el extendido durante 5 m inutos en
Form am ida 70% en 2xSSC.
8) Se deshidrató el extendido en soluciones de Etanol 70% , 80% y
100% de a -20º C durante 1 m inuto en cada una de ellas.
9) Se aplicaron las sondas (21q22 y X) sobre el extendido ya seco,
previam ente desnaturalizadas durante 10 m inutos a 75º C según
especificaciones del fabricante.
10) Se incubó durante 72 horas a 37º C en cám ara húm eda.
11) Se lavó el extendido con 0,05xSSC a 45º C en agitación durante 5
m inutos.
12) Se aplicaron 30μl de una solución compuesta por 0,1μg/ m l de
DAPI en Vectashield.
13) Se procedió a su análisis en el m icroscopio de fluorescencia
(Olym pus BX70, x1000) y procesadas con el sofware “Sm art
Capture” (Vysis) para luego equiparar la intensidad de
fluorescencia con Adobe PhotoShop CS (Adobe) .
3 .4 Estudios f ilogenét icos
3 .4 .1 Elección de los caracteres
Para este análisis se ut ilizaron:
-Datos obtenidos por Hibridación in situ Fluorescente con sondas
de crom osom as enteros hum anos propios de en este t rabajo de Tesis y

3. Materiales y m étodos
61
ot ros previam ente publicados (Consigliere et al. 1996, 1998; Morescalchi et al.
1997; García et al. 2000, 2002; Stanyon et al. 2001, 2003, 2004, 2011; Mudry et al.
2001; Neusser et al. 2001; de Oliveira et al. 2002, 2005; Ruiz-Herrera et al. 2005;
Am aral et al. 2008) .
-Datos de secuencias m oleculares obtenidos para el gen
m itocondrial para el Citocromo b (Cytb) a part ir de la base de datos de
Genbank (Cortés-Ort iz et al. 2003; Lorenz et al. 2005; Nascim iento et al. 2005; Harr is
et al. 2005; Casado et al. 2010) , núm eros de ent rada (Accession num bers) :
-Cebus apella: FJ529102.1,
-Lagothr ix lagothicha: AY671799.1,
-Alouat ta belzebul: AY374348.2,
-A. caraya: AY374359.2,
-A. seniculus seniculus: AY065886.1,
-A. sara: AY065887.1,
-A. m acconnelli: AY065888.1,
-A. guariba guariba: AY065899.1,
-A. guariba clam itans: DQ679782.1,
-A. pigra: AY065884.1,
-A. palliata: AY065879.1
Para obtener el pat rón de Hibridación in situ Fluorescente para las
especies de dist r ibución marginal norte (A. pigra y A. palliata) se
ut ilizaron las hom eologías crom osómicas, siguiendo el pat rón de
hibridación para A. caraya descrito por ot ros autores (de Oliveira et al. 2002;
Stanyon et al. 2011) .
3 .4 .2 Confección de las m at r ices
Se confeccionó una m atr iz binaria de datos, considerando la
presencia o ausencia de asociaciones sinténicas ent re segm entos
hom eólogos, según la nom enclatura descrita en Neusser et al., 2001 y en de
Oliveira et al. (2002) , en concordancia con las bandas cromosóm icas del
idiogram a humano (Tabla 3.4) .

3. Materiales y m étodos
62
Las secuencias m oleculares se alinearon m ediante el program a
CLUSTALW (Thom pson et al. 1994) .
Tabla 3.4: Caracteres ut ilizados para confeccionar la mat r iz binar ia de homeologías
crom osóm icas (m odificada de Neusser et al. 2001; de Oliveira et al. 2002) .
1. 1p21-pter/ 1p12-21
2. (5q31.3-qter/
7p22; q11 q21) 1
3. 5pter-q31.2/ 5q31.3-qter
4. 2pter q12/ 16q
5. 4q31.3-qter/ 4q23-q31.2
6. 4q23-q31.2/ 4pter-q22
7. (10q/ 16p) 3
8. 6
9. 8p/ 18
10. 15q21.3-q24/ 15q13-q21.2
11. 15q11 q13; q25 qter
12. 7p21 p11; q11 q21; q22
qter
13. 8q
14. 12
15. 11
16. 13
17. 9
18. 3pter p24; p21 p12; q12
q13; q27 qter
19. 3p24 q21; q13 q26
20. 1q32 qter
21. 1q21 q31
22. 3p12/ 21
23. 10p
24. 22
25. X
26. Y
27. 5pter-q31.2
28. (5q31.3-qter/
7p22; q11 q21) 2
29. 1p21-pter
30. 1p12-21
31. 4q31.3-qter
32. 4q23-q31.2/ 15q13-q212
33. 4pter-q22
34. 14/ 15q21.3-q24
35. (10q/ 16p) 2/ ( 10q/ 16p) 1
36. 15q11 q13; q25 qter/ Y
37. 2pter q12
38. 16q
39. 3p24 q21; q13 q26/
15q11 q13; q25 qter
40. (11/ 5pter-q31.2) 1
41. 5pter-q31./ 7p22; q11; q21
42. 12/ 9
43. 1p21-pter/ 2pter q12
44. 16q/ 4pter-q22
45. 22/ 14
46. 2q13 qter/ 20
47. 2q13 qter/ 4q23-q31.2
48. 8q/ 2q13 qter
49. 7p22; q11 q21/ 8q
50. 17/ 2pter q12
51. 17/ 2pter q12/ 12
52. 2pter q12/ 12
53. 1q32 qter/ 11
54. 1q32 qter/ (11/ 5pter-
q31.2) 2
55. (11/ 5pter-q31.2) 2
56. 18/ 14
57. 3pter p24; p21 p12; q12
q13; q27 qter/ 15q21.3-q24
58. 3pter p24; p21 p12; q12
q13; q27 qter/ 15q21.3-
q24/ 16q
59. 15q21.3-q24/ 16q
60. 17/ 10p
61. 17/ 10p/ 19
62. 10p/ 19
63. 22/ 20
64. 22/ 20/ 1q21 q31
65. 20/ 1q21 q31
66. (11/ 5pter-q31.2) 3
67. 1p12-21/ 8p
68. 7p21 p11; q11 q21; q22
qter / 14/ 15q21.3-q24
69. (10q/ 16p) 2
70. 11/ (10q/ 16p) 2
71. (10q/ 16p) 1
72. (10q/ 16p) 1/ 4pter-q22
73. 10p/ (10q/ 16p) 1/ 4pter-q22
74. 10p/ (10q/ 16p) 1
75. 19/ 13
76. 19/ 22
77. 22/ 1p21-pter
78. 19/ 22/ 1p21-pter
79. 2q13 qter/ 4q23-q31.2
80. 6/ 1p12-21
81. Y/ 15q11 q13; q25 qter/
3p24 q21; q13 q26
82. 1p12-21/ 5pter-q31.2/ 7p22;
q11 q21/ 5q31.3-qter/
7p22; q11 q21
83. 9/ 22
84. 17/ 11
85. 3pter p24; p21 p12; q12
q13; q27 qter/ 8p
86. 15q21.3-q24/ 1q32 qter
87. 16q/ 15q21.3-q24/ 1q32
qter
88. 4pter-q22/ 1p12-21
89. 14
90. 2pter q12/ 4pter-q22
91. 15q13-q21.2/ 7p22; q11
q21/ 5q31.3-qter/
7p22; q11 q21
92. 6/ 15q21.3-q24
93. 14/ 1p12-21
94. 6/ 15q21.3-q24/ 14/ 1p12-21
95. 17/ 8p/ 18
96. 22/ (10q/ 16p) 1
97. 2q13 qter/ 11
98. (10q/ 16p) 2/ 1q21 q31/ 20

3. Materiales y m étodos
63
3 .4 .3 Análisis f ilogenét ico
Se aplicó un análisis bajo el cr iter io de m áxim a parsim onia
ut ilizando el programa PAUP (Phylogenet ic Analysis Using Parsimony, Swofford
2002) . Todos los caracteres fueron considerados con igual peso, basados
en la prem isa de que los rearreglos cromosómicos ocurren con igual
probabilidad (de Oliveira et al. 2002) .
3 .5 Fotografía
Para la confección de las figuras de este t rabajo de Tesis, se
emplearon las fotografías tom adas con una cám ara analógica MPS 30
(ut ilizada durante el período comprendido ent re los años 2006 a 2008
inclusive) , adosada al m icroscopio Leica DMLB situado en el GI BE. En el
t ranscurso del t rabajo de Tesis Doctoral esta cám ara fue reem plazada por
una cám ara DFC 340 FX (ut ilizada durante el período com prendido ent re
los años 2009 y 2010) , dado que m ediante un subsidio PME 2006 Nº
1821 se agregaron al m icroscopio Leica DMLB filt ros de fluorescencia
(Leica A, I 3 y N2.1; ver Anexo I V para ver el listado de fluorocromos
visibles con estos filt ros) y la cám ara digital anteriorm ente m encionada.
Para el caso de la cám ara analógica, se ut ilizaron películas I lforpan
50 blanco y negro y el t rabajo fotográfico ( revelado y posit ivado) fue
realizado por el Laboratorio de Fotografía de la Facultad de Ciencias
Exactas y Naturales (UBA) .
Para el caso de la cámara digital, las fotos fueron procesadas por la
autora de este t rabajo de Tesis ut ilizando los program as PhotoShop CS
(Adobe) e I m age Pro-Plus 4.5 (Media Cybernet ics I nc.) en los laboratorios
del GI BE, FCEyN, UBA.

RESULTADOS

4. Resultados
64
4 .1 Diagnóst ico por fenot ipo externo
4 .1 .1 Pat rón de coloración de pela je
La ident ificación fenot ípica de cada ejem plar se realizó previam ente a
la tom a de muest ras de sangre perifér ica. Se tuvo en consideración la edad
y el sexo de los ejemplares y los pat rones de coloración del pelaje especie-
específicos descr itos para cada especie analizada:
Alouat ta caraya (Cabrera 1958) : los m achos adultos t ienen pelaje
homogéneo de color negro y las hem bras dorado claro a am arillo intenso.
Desde que nacen hasta que alcanzan la m adurez sexual, tanto los m achos
como las hembras t ienen coloración sim ilar a la que m uest ra el pelaje de las
hem bras adultas (Figura 4.1) .
Figura 4.1: Pat rón de coloración de pelaje de Alouat ta caraya (escala 1: 20) . a) Hem bra adulta
con cría; b) Macho adulto. Fotos gent ileza de Gabriela Bruno.
A. guariba clam itans (Cabrera 1958) : tanto los machos com o las
hembras t ienen pelaje de color negro o m arrón oscuro a roj izo,
generalm ente con las puntas del pelo m ás claras, lo que les da un efecto
“ listado” ( “br indled” ) (Figura 4.2) . En general se puede observar que los
m achos t ienden a ser m ás claros que las hem bras.

4. Resultados
65
Figura 4.2: Pat rón de coloración de pelaje de Alouat ta guariba clam itans (escala 1: 20) . Foto
gent ileza de I lar ia Agost ini.
A. pigra (Sm ith 1970) : tanto los m achos com o las hem bras t ienen
pelaje de color negro, con tonalidad am arronada en las m ej illas, hombros y
parte anterior del dorso (Figura 4.3) .
Figura 4.3: Pat rón de coloración de pelaje de un macho de Alouat ta pigra. a) Pat rón vent ral y
facial; b) Pat rón lateral (escala 1: 20) . Fotos gent ileza de Ariadna Rangel.

4. Resultados
66
A. palliata (Sm ith 1970) : tanto los m achos com o las hem bras t ienen
pelaje de color negro, con los flancos de color m arrón o dorado. Poseen
m anchas blancas en la cola y en las alm ohadillas ( “ fr ict ion skin” ) de m anos y
pies, cuya coloración y form a pueden variar de individuo a individuo,
const ituyendo junto a ot ras característ icas, una var iable m ás a considerar y
regist rar al mom ento de ident ificar al ejem plar (Figura 4.4) .
Figura 4.4: Pat rón de coloración de pelaje de un m acho de Alouat ta palliata (APA) . a) Pat rón
dorsal/ lateral. Foto gent ileza de Mariela Nieves (escala 1: 20) ; b) Alm ohadilla plantar m ost rando el
pat rón de manchas claras característ ico de ese ejemplar (APA 537) . Foto gent ileza de Mariela Nieves
(escala 1: 10) c) Alm ohadilla vent ral de la cola m ost rando las m anchas claras característ icas de ese
ejem plar (APA 537) . Foto gent ileza de Ar iadna Rancel (escala 1: 10) .
Saim ir i boliviensis boliviensis (Hershkovit z 1984) : am bos sexos t ienen la
frente de color agut í gr isáceo a enteram ente negro como la coronilla, con
bandas blancuzcas ausentes; región superciliar casi desnuda o con poco
pelo de color agut í o blancuzco; fina cola de pelaje negruzco; coronilla y
parche pre-auricular negruzco tanto en los m achos como en las hem bras;
superficie superior de la cola, a excepción de la porción negruzca term inal,
de color gr isáceo o agut í a negro (Figura 4.5) .

4. Resultados
67
Figura 4.5: Pat rón de coloración de pelaje de los ejem plares de Saim ir i boliv iensis boliviensis.
a) Pat rón facial y lateral. Foto gent ileza de Melina Alvarez (escala 1: 15) . b) Pat rón dorsal con vista
com pleta de m anto y cola (escala 1: 10) .
Cebus libidinosus (Cabrera 1958) : posee un copete de pelos cortos y
negros, que puede estar dividido en dos puntas o penachos a cada lado de
la cabeza. Estos penachos (en inglés “ tuft ” ) se pueden observar en am bos
sexos, con frecuencias variables en altura y presencia. Los hom bros son de
color m ás claro que el resto del cuerpo, variando desde m arrón claro a
m arrón oscuro en el m anto y en la zona vent ral. Las m anos y pies son
negros y la cola es oscura en el ext rem o distal (Figura 4.6a) .
C. nigritus (Cabrera 1958) : tanto los m achos com o las hem bras t ienen
pelaje de color negro a m arrón oscuro homogéneo desde la cabeza hasta la
ext rem idad de la cola, tanto dorsal com o vent ralm ente (Figura 4.6b) .

4. Resultados
68
Figura 4.6: Pat rón de coloración de pelaje de las especies del género Cebus analizadas. a)
Pat rón lateral/ dorsal de Cebus libidinosus (escala 1: 20) . b) Pat rón lateral de Cebus nigritus. Foto
gent ileza de Marta D. Mudry (escala 1: 20) .
Aotus azarae (Cabrera 1958) : el pelaje es de color agut í en el cuello,
garganta y vient re, con predom inio del anaranjado. Alrededor de los ojos
t ienen un pat rón de coloración facial con zonas blancas y negras ( “ant ifaz” ) .
El manto es agut í, gr is veteado a m arrón. El ext rem o de la cola es de color
negro (Figura 4.7) .
Figura 4.7: Pat rón de coloración de pelaje de los ejem plares de Aotus azarae. Fotos gent ileza
de Carola Milozzi. a) Pat rón facial y vent ral (escala 1: 15) . b) Pat rón lateral (escala 1: 20) .

4. Resultados
69
Pan t roglodytes (Bourne 1969) : tanto los m achos com o las hem bras
t ienen pelaje de color negro. Las crías y juveniles t ienen pelaje blanco en el
contorno de la cara y un penacho blanco en la cola que se oscurece cuando
los individuos llegan a adultos. Tanto los machos como las hembras t ienen
barbas blancas (Figura 4.8) .
Figura 4.8: Pat rón de coloración de pelaje de un m acho de Pan t roglodytes (escala 1: 20) . Fotos
gent ileza de Juliana Rosa. a) Pat rón vent ral. b) Pat rón lateral.
4 .1 .2 Ot ros parám et ros
Para la diagnosis, tom ando en consideración el fenot ipo externo en
m ás de la dist r ibución y coloración del pelaje, se consideraron los valores
m orfom ét r icos, el peso corporal y la fórm ula dentaria de todos los
ejemplares, si bien no siem pre se logra el acuerdo en la tom a del m ism o
listado de m edidas. Estas var iables, en general, las tom an los técnicos o los
m édicos veterinarios de cada inst ituto que en cada circunstancia colaboran
con el estudio de los especím enes. A m odo de ejem plo se indican los datos
m orfom ét r icos en sus valores m edios para algunas de las especies de
Primates Neotropicales analizadas (Tabla 4.1) .
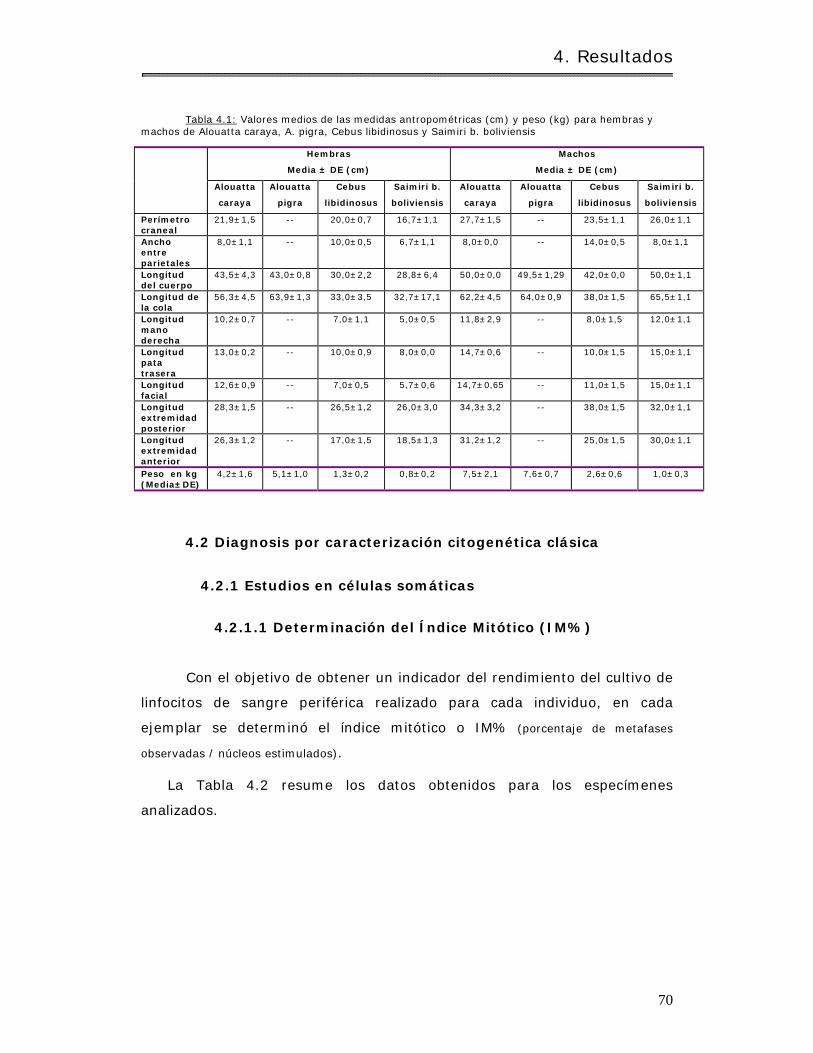
4. Resultados
70
Tabla 4.1: Valores m edios de las m edidas ant ropom ét r icas (cm ) y peso (kg) para hem bras y
m achos de Alouat ta caraya, A. pigra, Cebus libidinosus y Saim ir i b. boliv iensis
4 .2 Diagnosis por caracter ización citogenét ica clás ica
4 .2 .1 Estudios en células som át icas
4 .2 .1 .1 Determ inación del Í ndice Mitót ico ( I M % )
Con el objet ivo de obtener un indicador del rendim iento del cult ivo de
linfocitos de sangre per ifér ica realizado para cada individuo, en cada
ejemplar se determ inó el índice m itót ico o I M% (porcentaje de metafases
observadas / núcleos est im ulados) .
La Tabla 4.2 resum e los datos obtenidos para los especím enes
analizados.
Hem bras
Media ± DE ( cm )
Machos
Media ± DE ( cm )
Alouat ta
caraya
Alouat ta
pigra
Cebus
libidinosus
Saim ir i b.
boliviensis
Alouat ta
caraya
Alouat ta
pigra
Cebus
libidinosus
Sa im ir i b.
boliviensis
Perím et ro craneal
21,9± 1,5 - - 20,0± 0,7 16,7± 1,1 27,7± 1,5 - - 23,5± 1,1 26,0± 1,1
Ancho ent re par ie ta les
8,0± 1,1 - - 10,0± 0,5 6,7± 1,1 8,0± 0,0 - - 14,0± 0,5 8,0 ± 1,1
Longitud de l cuerpo
43,5± 4,3 43,0± 0,8 30,0± 2,2 28,8± 6,4 50,0± 0,0 49,5± 1 ,29 42,0± 0,0 50,0± 1,1
Longitud de la cola
56,3± 4,5 63,9± 1,3 33,0± 3,5 32,7± 17,1 62,2± 4,5 64,0± 0,9 38,0± 1,5 65,5± 1,1
Longitud m ano derecha
10,2± 0,7 - - 7,0± 1,1 5,0± 0,5 11,8± 2,9 - - 8,0± 1,5 12, 0± 1,1
Longitud pata t rasera
13,0± 0,2 - - 10,0± 0,9 8,0± 0,0 14,7± 0,6 - - 10,0± 1,5 1 5,0± 1,1
Longitud facia l
12,6± 0,9 - - 7,0± 0,5 5,7± 0,6 14,7± 0,65 - - 11,0± 1,5 1 5,0± 1,1
Longitud ex t rem idad p oster ior
28,3± 1,5 - - 26,5± 1,2 26,0± 3,0 34,3± 3,2 - - 38,0± 1,5 32,0± 1,1
Longitud ex t rem idad anter ior
26,3± 1,2 - - 17,0± 1,5 18,5± 1,3 31,2± 1,2 - - 25,0± 1,5 30,0± 1,1
Peso en kg ( Media± DE)
4,2± 1,6 5,1± 1,0 1,3± 0,2 0,8± 0,2 7,5± 2,1 7,6± 0,7 2,6 ± 0,6 1,0± 0,3
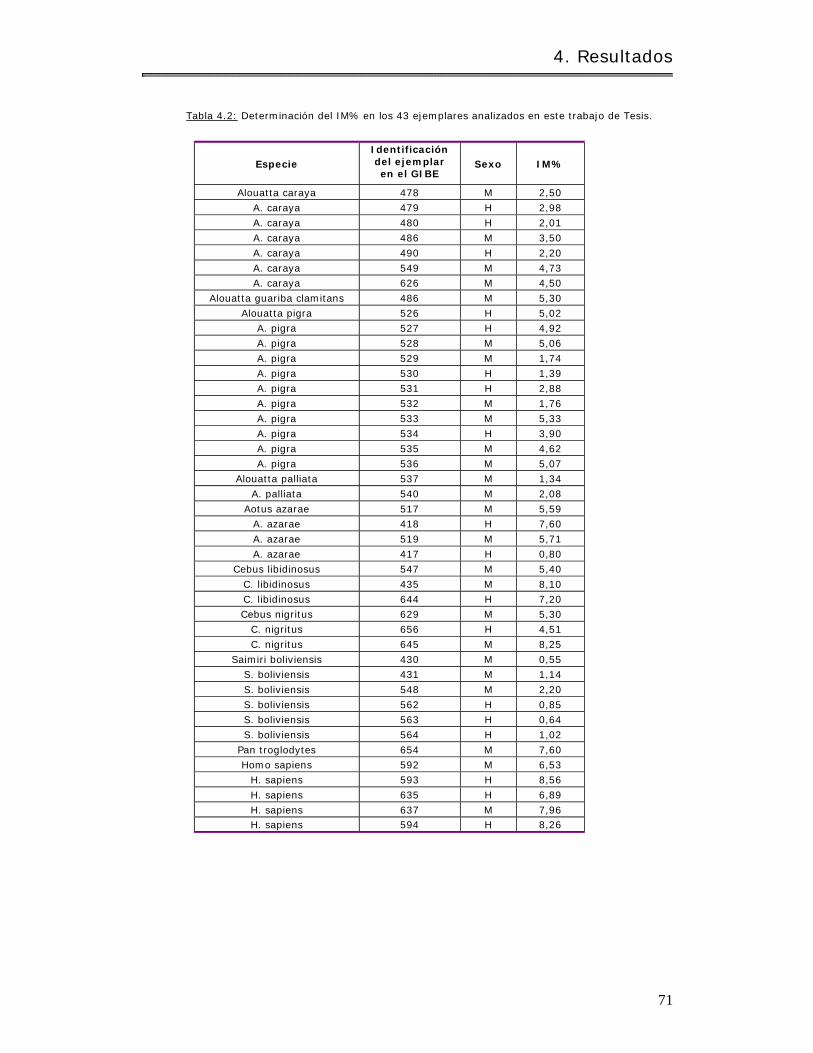
4. Resultados
71
Tabla 4.2: Determ inación del I M% en los 43 ejem plares analizados en este t rabajo de Tesis.
Especie
I dent if icación del e jem plar
en e l GI BE Sexo I M%
Alouat ta caraya 478 M 2,50
A. caraya 479 H 2,98
A. caraya 480 H 2,01
A. caraya 486 M 3,50
A. caraya 490 H 2,20
A. caraya 549 M 4,73
A. caraya 626 M 4,50
Alouat ta guariba clam itans 486 M 5,30
Alouat ta pigra 526 H 5,02
A. pigra 527 H 4,92
A. pigra 528 M 5,06
A. pigra 529 M 1,74
A. pigra 530 H 1,39
A. pigra 531 H 2,88
A. pigra 532 M 1,76
A. pigra 533 M 5,33
A. pigra 534 H 3,90
A. pigra 535 M 4,62
A. pigra 536 M 5,07
Alouat ta palliata 537 M 1,34
A. palliata 540 M 2,08
Aotus azarae 517 M 5,59
A. azarae 418 H 7,60
A. azarae 519 M 5,71
A. azarae 417 H 0,80
Cebus libidinosus 547 M 5,40
C. libidinosus 435 M 8,10
C. libidinosus 644 H 7,20
Cebus nigr itus 629 M 5,30
C. nigr itus 656 H 4,51
C. nigr itus 645 M 8,25
Saim ir i boliv iensis 430 M 0,55
S. boliviensis 431 M 1,14
S. boliviensis 548 M 2,20
S. boliviensis 562 H 0,85
S. boliviensis 563 H 0,64
S. boliviensis 564 H 1,02
Pan t roglodytes 654 M 7,60
Hom o sapiens 592 M 6,53
H. sapiens 593 H 8,56
H. sapiens 635 H 6,89
H. sapiens 637 M 7,96
H. sapiens 594 H 8,26

4. Resultados
72
4 .2 .1 .2 Análisis m itót icos 4 .2 .1 .2 .1 Género Alouat ta ( Ate lidae: Ceboidea)
4 .2 .1 .2 .1 .1 Alouat ta caraya
4 .2 .1 .2 .1 .1 .1 Determ inación del 2 N y NF
Para establecer el núm ero diploide (2N) y el núm ero fundam ental
(NF) correspondiente a los individuos de ambos sexos de cada especie
analizada, se tom aron en cuenta al m enos 50 m etafases con t inción
uniform e de cada ejem plar, bajo m icroscopía ópt ica. En todos los
ejemplares se confirmó, en m ás del 60% de las m etafases analizadas, un
2N= 52, X1X1X2X2/ X1X2Y1Y2 en coincidencia con el 2N descr ito previam ente
para la especie (Mudry et al. 1981, 1984a, 1998, 2001; Rahn et al. 1996) . La Tabla
4.3 resum e los resultados obtenidos.
Tabla 4.3: Determ inación del Núm ero Diploide (% 2N) y el Núm ero Fundam ental (NF) en los
especím enes de A. caraya analizados.
I ndividuo
Sexo
NF
% 2 N
≤49 50 51 5 2 53 ≥54
478 M 72 4,0 7,0 8,0 6 5 ,0 10,0 6,0
479 H 74 20,0 8,0 2,0 6 0 ,8 10,2 0,0
480 H 74 13,0 9,0 9,0 6 6 ,0 2,0 1,0
486 M 72 8,0 7,0 3,0 7 6 ,0 6,0 0,0
490 H 74 10,0 16,0 4,0 6 0 ,0 8,0 2,0
549 M 72 3,0 7,0 4,5 7 6 ,5 6,0 3,0
626 M 72 15,0 9,5 4,5 6 4 ,0 7,0 0,0
Se corroboró la fórmula crom osómica dado que se observó un
com plem ento autosóm ico com puesto por 8 pares crom osómicos de
m orfología subm etacént r ica, 2 pares m etacént r icos y 15 pares
acrocént r icos. En los m achos se observaron 7 pares cromosóm icos
submetacént r icos, 2 pares m etacént r icos y 15 pares de m orfología
acrocént r ica debido a la presencia de un sistem a de determ inación sexual
m últ iple.

4. Resultados
73
4 .2 .2 .1 .1 .2 Pat rones de bandas G y C
Los pat rones de bandas G de los 7 ejem plares analizados perm it ieron
corroborar la asignación de especie a Alouat ta caraya (Figura 4.9) . En los
ejem plares m achos se observa un sistem a de determ inación sexual múlt iple
de t ipo X1X2Y1Y2. El m ism o estaría com puesto por X1 y X2 de morfología
submetacént r ica y cromosom as Y1 e Y2 de morfología acrocént r ica. El par
crom osóm ico ACA7 estaría involucrado en la t ranslocación Y-autosom a en
los m achos.
Figura 4.9: Pat rón de bandas G de una hem bra adulta de A. caraya (Barra= 10 μm) . El recuadro
m uest ra los crom osom as sexuales observados en los m achos. El par crom osóm ico ACA7 estaría
involucrado en la t ranslocación Y-autosom a en los machos.

4. Resultados
74
Mediante la técnica de bandas C se caracter izó el pat rón de
dist r ibución de los bloques heterocrom át icos en los ejem plares analizados
(Figura 4.10) . Se observó heterocrom at ina C+ cent rom érica en todos los
pares cromosóm icos. Se observaron bandas ext racent rom éricas
polimórficas ( interst iciales) en los pares ACA16 y ACA21. No se observaron
bandas C+ telom éricas en ningún ejem plar analizado.
Figura 4.10: Metafase con bandas C de un macho de Alouat ta caraya (Barra= 10 μm) . Las flechas
indican las bandas C+ interst iciales en los pares ACA16 y ACA21.
4 .2 .1 .2 .1 .2 Alouat ta guar iba clam itans
4 .2 .1 .2 .1 .2 .1 Determ inación del 2 N y NF
Se analizaron bajo m icroscopía ópt ica al m enos 50 m etafases con
t inción uniform e del espécimen analizado. Se confirm ó en m ás de 65% de
las m etafases analizadas, un 2N= 45, X1X2X3Y1Y2 en coincidencia con el 2N
descrito previam ente para la especie (de Oliveira et al. 2002) . La Tabla 4.4
resum e los resultados obtenidos. Se corroboró la fórm ula cromosóm ica
correspondiente a esta especie ya que se observó un com plem ento
autosóm ico com puesto por 5 pares cromosóm icos de m orfología
submetacént r ica, 6 pares m etacént r icos y 9 pares acrocént r icos.

4. Resultados
75
Tabla 4.4: Determ inación del Núm ero Diploide (% 2N) y el Núm ero Fundam ental (NF) en el
especim en m acho de A. guariba clam itans analizado.
4 .2 .1 .2 .1 .2 .2 Pat rones de b andas G y C
El pat rón de bandas G perm it ió corroborar la asignación de especie
de este ejem plar com o correspondiente a Alouat ta guariba clam itans
(Figura 4.11) y sugerir un sistem a de determ inación sexual m últ iple de t ipo
X1X2X3Y1Y2, el cual estaría com puesto por X1 y X2 de morfología
submetacént r ica y crom osom as X3, Y1 e Y2 de morfología acrocéntr ica. El
par AGU6 estaría involucrado en la t ranslocación Y-autosom a en los
m achos.
Figura 4.11: Pat rón de bandas G del ejem plar m acho de A. guariba clam itans (Barra= 10 μm ) .
El par crom osóm ico AGU6 estaría involucrado en la t ranslocación Y-autosom a en los m achos.
I ndividuo
NF
% 2 N
≤42 43 44 4 5 46 ≥47
485 69 4,0 7,0 10,0 6 5 ,0 8,0 6,0

4. Resultados
76
Mediante la técnica de bandas C se observó heterocrom at ina C+
cent rom érica en todos pares crom osóm icos y C+ interst icial polimórfica en 2
pares acrocént r icos (AGU16 y AGU17) . Se observó heterocrom at ina
telom érica en el par 2 (Figura 4.12) .
Figura 4.12: Metafase con bandas C (Barra= 10 μm ) : Las flechas indican las bandas C+
interst iciales polimórficas en los pares 16 y 17. Los asteriscos indican la banda C+ telom érica en el par 2.
4 .2 .1 .2 .1 .3 Alouat ta pigra
4 .2 .1 .2 .1 .3 .1 Determ inación del 2 N y NF
Como una de las cont r ibuciones originales de este t rabajo de Tesis se
describe la pr im er caracterización del car iot ipo del aullador negro Alouat ta
pigra. Para establecer el núm ero diploide (2N) y el núm ero fundam ental
(NF) correspondiente a los individuos de am bos sexos de esta especie, se
analizaron al m enos 50 m etafases con t inción uniform e de cada ejem plar
bajo m icroscopía ópt ica.

4. Resultados
77
El núm ero diploide observado para todos los especím enes, m achos y
hembras fue 2N= 58 en el 69% de las m etafases analizadas. El número
fundam ental (NF) es 78 para las hem bras y 77 para los machos debido a la
presencia de un sistem a sexual m últ iple de t ipo X1X2Y1Y2 (ver sección
4.2.1.2.1.3.3) . En la Tabla 4.5 se resum en los resultados obtenidos.
Tabla 4.5: Determ inación del Núm ero Diploide (% 2N) y el Núm ero Fundam ental (NF) en los
especím enes de A. pigra analizados.
I ndividuo
Sexo
NF
% 2 N
≤55 56 57 5 8 59 ≥60
526 F 78 4,0 6,0 4,0 7 6 ,0 4,0 6,0
527 F 78 16,0 10,0 2,0 6 4 ,0 8,0 0,0
528 M 77 8,0 12,0 8,0 7 0 ,0 2,0 0,0
529 M 77 8,0 28,0 2,0 6 0 ,0 0,0 0,0
530 F 78 10,0 6,0 0,0 7 4 ,0 8,0 2,0
531 F 78 5,9 9,8 3,9 8 0 ,4 0,0 0,0
532 M 77 15,0 4,0 11,0 6 6 ,0 4,0 0,0
533 M 77 24,0 8,0 4,0 6 0 ,0 2,0 2,0
534 H 78 10,0 16,0 4,0 6 0 ,0 10,0 0,0
535 M 77 15,0 5,0 10,0 6 2 ,0 8,0 0,0
536 M 77 10,0 10,0 8,0 7 2 ,0 0,0 3,0
4 .2 .1 .2 .1 .3 .2 Medidas crom osóm icas
Al ser la pr imera caracterización del car iot ipo de esta especie de
aullador, para la corroboración de la fórm ula crom osómica se tom aron las
m edidas crom osómicas a part ir de m etafases de igual elongación y tam año
de los ejem plares estudiados para este t rabajo de Tesis (Tabla 4.6) .
Los valores del índice cent rom érico ( I C) , relación de brazos ( r) y el
porcentaje de la Longitud Total del Com plem ento (% LTC) perm it ieron
confirmar el com plem ento crom osóm ico observado en las m etafases con
t inción estándar. El crom osom a X (X1) en A. pigra com prende 4,87% del
genom a, en concordancia con la Ley de Ohno que establece que el
crom osom a X ancest ral de m amíferos comprende aproxim adam ente 5% del
genom a total (Ohno 1967) .
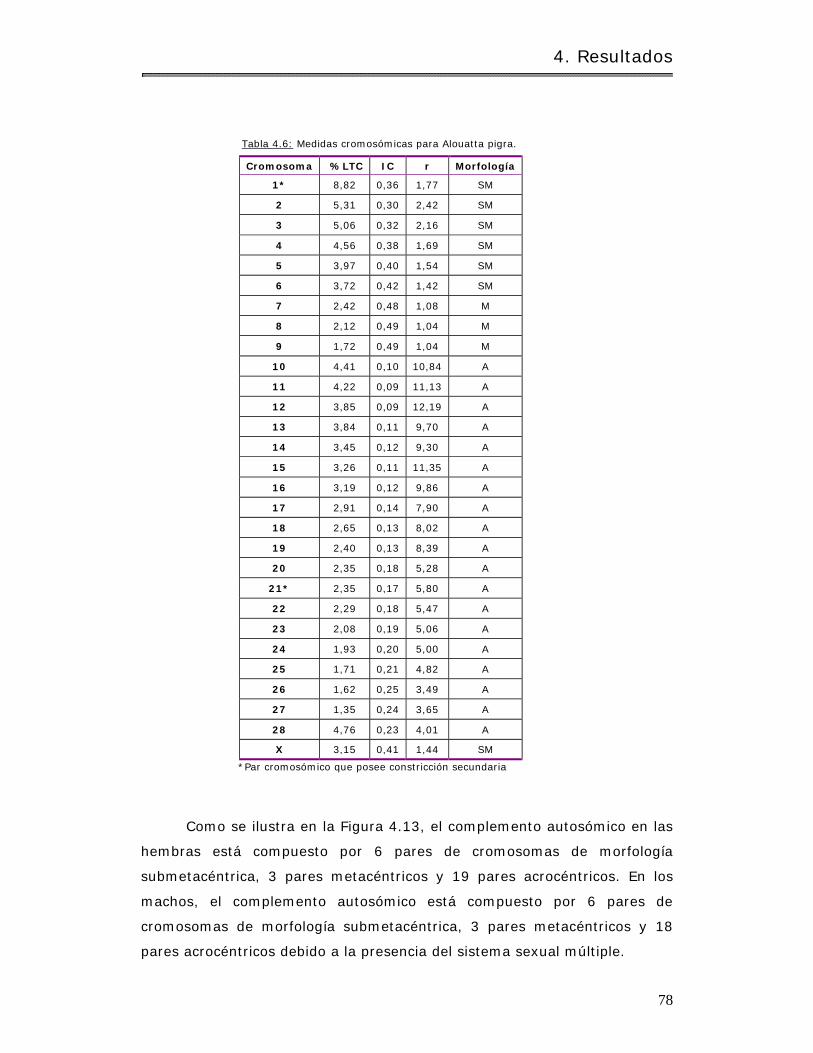
4. Resultados
78
Tabla 4.6: Medidas crom osóm icas para Alouat ta pigra.
Crom osom a % LTC I C r Morfología
1 * 8,82 0,36 1,77 SM
2 5,31 0,30 2,42 SM
3 5,06 0,32 2,16 SM
4 4,56 0,38 1,69 SM
5 3,97 0,40 1,54 SM
6 3,72 0,42 1,42 SM
7 2,42 0,48 1,08 M
8 2,12 0,49 1,04 M
9 1,72 0,49 1,04 M
1 0 4,41 0,10 10,84 A
1 1 4,22 0,09 11,13 A
1 2 3,85 0,09 12,19 A
1 3 3,84 0,11 9,70 A
1 4 3,45 0,12 9,30 A
1 5 3,26 0,11 11,35 A
1 6 3,19 0,12 9,86 A
1 7 2,91 0,14 7,90 A
1 8 2,65 0,13 8,02 A
1 9 2,40 0,13 8,39 A
2 0 2,35 0,18 5,28 A
2 1 * 2,35 0,17 5,80 A
2 2 2,29 0,18 5,47 A
2 3 2,08 0,19 5,06 A
2 4 1,93 0,20 5,00 A
2 5 1,71 0,21 4,82 A
2 6 1,62 0,25 3,49 A
2 7 1,35 0,24 3,65 A
2 8 4,76 0,23 4,01 A
X 3,15 0,41 1,44 SM
* Par cromosóm ico que posee const r icción secundaria
Com o se ilust ra en la Figura 4.13, el com plem ento autosóm ico en las
hem bras está com puesto por 6 pares de crom osomas de morfología
submetacént r ica, 3 pares m etacéntr icos y 19 pares acrocént r icos. En los
m achos, el com plem ento autosóm ico está com puesto por 6 pares de
crom osom as de m orfología subm etacént r ica, 3 pares m etacént r icos y 18
pares acrocént r icos debido a la presencia del sistem a sexual m últ iple.

4. Resultados
79
Figura 4.13: Metafase de una hem bra de Alouat ta pigra con t inción uniform e (Barra= 10 μm ) . El
recuadro infer ior a la derecha muest ra los cromosom as sexuales observados en los m achos.
4 .2 .1 .2 .1 .3 .2 Pat rones de bandas G y C
El pat rón de bandas G obtenido para A. pigra (Figura 4.14) m ost ró la
presencia de un sistem a crom osóm ico de determ inación sexual XX en las
hembras (que se indica por convención com o X1X1X2X2 para hom ogeneizar
la nom enclatura con la ut ilizada para el sistem a sexual en los machos) y
sugir ió un sistem a de determ inación sexual múlt iple X1X2Y1Y2 en los m achos,
que se podría haber producido por una t ranslocación ent re el cromosom a Y
y el par autosóm ico 17. Este sistem a de determ inación sexual múlt iple
parecería estar com puesto por un X1 de m orfología subm etacént r ica y
crom osom as X2, Y1 e Y2 de morfología acrocéntr ica.

4. Resultados
80
Se observó un heteromorfism o en el par crom osómico 1 debido a un
polimorfismo en el tam año de la const r icción secundaria (ver sección
4.2.1.2.1.3.4) .
Figura 4.14: Pat rón de bandas G de una hem bra adulta de A. pigra (Barra= 10 μm ) . El recuadro
muest ra los crom osom as sexuales observados en los m achos. El par API 7 estaría involucrado en la
t ranslocación Y-autosom a en los machos.
En las m etafases con bandas C (Figura 4.15) se observó
heterocrom at ina C+ cent rom érica en todos los pares crom osómicos. En el
par cromosómico API 2 se observaron bandas C+ telom éricas. No se
detectaron bandas C+ interst iciales.

4. Resultados
81
Figura 4.15: Metafase con bandas C de un m acho de Alouat ta pigra m ost rando las bandas
heterocrom át icas C+ centrom éricas en todos los pares crom osóm icos. Las flechas indican las bandas
C+ telom éricas observadas en el par crom osóm ico API 2 (Barra = 10 μm ).
4 .2 .1 .2 .1 .3 .3 Crom osom as m arcadores
Una de las característ icas morfológicas a tener en cuenta en
citotaxonom ía es la const r icción secundaria. Esta const r icción es una región
crom osóm ica acrom át ica que perm ite seguir un crom osom a en las dist intas
especies dent ro de un género o ent re géneros. Para corroborar la presencia
de una región de organización nucleolar ( “Nucleolar Organizer Regions” o
NOR) en las const r icciones secundarias del car iot ipo de A. pigra se realizó la
técnica de t inción diferencial para bandas NOR. Mediante esta técnica se
corroboró la presencia de organizadores nucleolares en las const r icciones
secundarias de los pares API 1 y API 21 (Figura 4.16) . Se observó un
polimorfismo para el tam año de la banda NOR del crom osoma 1 que
explicaría el heteromorfismo observado en este cromosom a en todas las
m etafases analizadas.

4. Resultados
82
Figura 4.16: Metafase con bandas NOR en un m acho de A. pigra. Las líneas indican las bandas
NOR+ en los pares crom osóm icos 1 y 21 (Barra = 10 μm ).
4 .2 .1 .2 .1 .4 Alouat ta pa llia ta
4 .2 .1 .2 .1 .4 .1 Determ inación del 2 N y NF
Para establecer el núm ero diploide (2N) y el núm ero fundam ental
(NF) correspondiente a esta especie, se analizaron al menos 50 m etafases
con t inción uniform e de cada ejem plar bajo m icroscopía ópt ica. En uno de
los ejem plares (APA 537) se confirm ó un 2N= 53, X1X2Y en coincidencia con
el 2N descr ito previam ente para los m achos de la especie (Ma et al. 1975;
Solar i y Rahn 2005) . En el segundo ejemplar (APA 540) , or iginalm ente
caracter izado com o una hem bra, se observó un 2N= 53, X1X2Y, por lo que se
t rataría de un caso de test ículos no descendidos (cr iptorquidia) . Por este
m ot ivo se cam bió la asignación de sexo de hem bra con la que se lo tenía
regist rado en caut iverio a m acho. La Tabla 4.7 resum e los resultados
obtenidos.
Se corroboró la fórm ula crom osómica al observar un com plem ento

4. Resultados
83
autosóm ico com puesto por 8 pares cromosóm icos de m orfología
submetacént r ica, 3 pares m etacént r icos y 14 pares acrocént r icos en los dos
m achos analizados.
Tabla 4.7: Determ inación del Núm ero Diploide (% 2N) y el Núm ero Fundam ental (NF) en los
especim enes de A. palliata analizados.
I ndividuo
NF
% 2 N
≤50 51 52 5 3 54 ≥55
537 76 4 7 10 6 6 8 5
540 76 10 8 6 6 0 4 2
4 .2 .1 .2 .1 .4 .2 Pat ro nes de bandas G y C
El pat rón de bandas G perm it ió corroborar la asignación de especie de
am bos ejem plares a A. palliata (Figura 4.17) y sugir ió un sistem a de
determ inación sexual múlt iple de t ipo X1X2Y, el cual parecería estar
com puesto por un X1 de m orfología subm etacént r ica y cromosom as X2 e Y
de morfología acrocént r ica. El par APA19 sería el involucrado en la
t ranslocación Y-autosom a.
Figura 4.17: Pat rón de bandas G de un m acho adulto de A. palliata (Barra= 10 μm ). El par
APA19 sería el involucrado en la t ranslocación Y-autosoma.

4. Resultados
84
En las m etafases con bandas C (Figura 4.18) se observó
heterocrom at ina C+ cent rom érica en todos los pares cromosóm icos. En los
pares cromosóm icos 3 y 5 se observaron bandas C+ telom éricas. No se
detectaron bandas C+ interst iciales.
Figura 4.18: Metafase de A. palliata con bandas C (Barra = 10 μm ). Las flechas indican las
bandas C+ teloméricas en los pares 3 y 5.
4 .2 .1 .2 .2 Ot ros géneros de Ceboidea ( Cebidae: Platy rr in i)
4 .2 .1 .2 .2 .1 Saim ir i b. boliv iensis
4 .2 .1 .2 .2 .1 .1 Determ inación del 2 N y NF
Se analizaron al m enos 50 m etafases con t inción uniform e de cada
ejem plar bajo m icroscopía ópt ica. En todos los ejemplares, tanto m achos
como hem bras, se confirmó un 2N= 44, XX/ XY en coincidencia con el 2N
descrito previam ente para la especie (Mudry et al. 1982; García et al. 1995) . La
Tabla 4.8 resum e los resultados obtenidos.
Se corroboró la fórm ula crom osómica al observar un com plem ento

4. Resultados
85
autosóm ico com puesto por 5 pares cromosóm icos de m orfología
m etacént r ica (grupo A) , 10 pares subm etacéntr icos (grupo B) y 6 pares
acrocént r icos (grupo C) . El crom osoma X se observa com o subm etacént r ico
y el crom osom a Y com o un acrocént r ico pequeño.
Tabla 4.8: Determ inación del Núm ero Diploide (% 2N) y el Núm ero Fundam ental (NF) en los especím enes de Saim ir i boliviensis boliv iensis analizados.
I ndividuo Sexo
NF
% 2 N
≤41 42 43 4 4 45 46 ≥47
430 M 74 15,23 6,52 5,43 6 7 ,3 9 4,35 1,09 0,00
431 M 74 17,00 8,00 4,00 6 1 ,0 0 4,00 3,00 3,00
548 M 74 8,00 4,00 20,00 6 3 ,0 0 1,00 4,00 0,00
562 H 75 8,00 8,00 14,00 6 0 ,0 0 8,00 2,00 0,00
563 H 75 12,00 12,00 14,00 6 4 ,0 0 16,00 3,00 3,00
564 H 75 14,00 6,00 8,00 7 2 ,0 0 0,00 0,00 0,00
4 .2 .1 .2 .2 .1 .2 Pat rones de bandas G y C
El pat rón de bandas G perm it ió corroborar la asignación de especie
de todos estos ejemplares a Saim ir i boliviensis boliv iensis (Figura 4.19) ,
observándose la presencia de los pares B5 y B2 y la ausencia de los pares
B10 y C1, característ icas del car iot ipo de esta especie.
Figura 4.19: Pat rón de bandas G de una hem bra adulta de Saim ir i b. boliv iensis
(Barra= 10 μm ). El recuadro m uest ra el sistem a sexual XY observado en los m achos.

4. Resultados
86
En las m etafases con bandas C (Figura 4.20) se observó
heterocrom at ina C+ cent rom érica en todos los pares crom osóm icos. En 3
pares crom osómicos (A1, B1, B11) se observaron bandas C+ interst iciales.
En 9 pares crom osómicos (B2, B3, B4, B5, B6, B7, B8, B9, B11) se
observaron bandas C+ telom éricas.
Figura 4.20: Metafase con bandas C (Barra= 10 μm ) : Las flechas indican las bandas C+
interst iciales en los pares A1, B1 y B11. Los aster iscos indican los pares cromosóm icos con bandas C+
telom éricas.
4 .2 .1 .2 .2 .2 Cebus libidinosus
4 .2 .1 .2 .2 .2 .1 Determ inación del 2 N y NF
Para establecer el núm ero diploide (2N) y el núm ero fundam ental
(NF) correspondiente se analizaron al m enos 50 m etafases con t inción
uniform e de cada ejem plar bajo m icroscopía ópt ica. En todos los ejem plares
se confirm ó un 2N= 54, XX/ XY en coincidencia con el 2N descrito
previam ente para la especie (Matayoshi et al. 1987) . El núm ero fundam ental
observado fue NF= 76 para las hem bras y NF= 75 para los m achos. La Tabla
4.9 resum e los resultados obtenidos.

4. Resultados
87
Tabla 4.9: Determ inación del Núm ero Diploide (% 2N) y el Núm ero Fundam ental (NF) en los especím enes de Cebus libidinosus analizados.
I ndividuo Sexo
NF
% 2 N
≤51 52 53 5 4 55 56 ≥57
435 M 75 6 10 6 7 6 0 2 0
547 M 75 12 12 0 6 6 0 5 5
644 H 76 10 8 4 7 2 2 2 2
Se corroboró la fórm ula crom osómica y se observó un com plem ento
autosóm ico com puesto por 8 pares de m orfología subm etacéntr ica, 2 pares
m etacént r icos y 16 pares acrocént r icos. El cromosom a X se presenta com o
un crom osom a subm etacént r ico y el cromosom a Y com o un acrocént r ico
pequeño.
4 .2 .1 .2 .2 .2 .2 Pat rones de bandas G y C
El pat rón de bandas G perm it ió corroborar la asignación de especie
de todos los ejem plares com o Cebus libidinosus (Figura 4.21) .
Figura 4.21: Pat rón de bandas G de una hem bra adulta de Cebus libidinosus (Barra= 10 μm ). El
recuadro abajo a la derecha m uest ra el sistema sexual XY observado en los machos.

4. Resultados
88
En las m etafases con bandas C (Figura 4.22) se observó
heterocrom at ina C+ cent rom érica en todos los pares crom osóm icos. Se
observaron bandas C+ interst iciales en los pares crom osóm icos 4, 6, 11, 12,
13, 17 y 19. No se observaron bandas C+ telom éricas.
Figura 4.22: Metafase de Cebus libidinosus con bandas C (Barra= 10 μm ). Las flechas indican
las bandas C+ interst iciales en los pares 4, 6, 11, 12, 13, 17 y 19.
4 .2 .1 .2 .2 .3 Cebus nigr itus
4 .2 .1 .2 .2 .3 .1 Determ inación del 2 N y NF
En todos los ejem plares se confirm ó un 2N= 54, XX/ XY en
coincidencia con el 2N descrito previam ente para la especie (Mudry 1990) . El
núm ero fundam ental observado fue NF= 76 para las hembras y NF= 75 para
los m achos. La Tabla 4.10 resum e los resultados obtenidos.

4. Resultados
89
Tabla 4.10: Determ inación del Núm ero Diploide (% 2N) y el Núm ero Fundam ental (NF) en los especím enes de Cebus nigr itus analizados.
I ndividuo Sexo
NF
% 2 N
≤51 52 53 5 4 55 56 ≥57
629 M 75 10,0 10,0 8,0 6 6 ,0 4,0 2,0 0,0
645 M 75 11,0 10,0 9,0 6 8 ,0 1,0 1,0 0,0
656 H 76 12,0 8,0 6,0 7 0 ,0 2,0 0,0 2,0
Se corroboró la fórm ula crom osómica al observar un com plem ento
autosóm ico com puesto por 8 pares de m orfología subm etacéntr ica, 2 pares
m etacént r icos y 16 pares acrocént r icos. El crom osom a X es
submetacént r ico y el cromosom a Y es un acrocént r ico pequeño.
4 .2 .1 .2 .2 .3 .2 Pat rones de bandas G y C
El pat rón de bandas G perm it ió corroborar la asignación de especie
de todos los ejem plares a Cebus nigr itus (Figura 4.23) .
Figura 4.23: Pat rón de bandas G de una hem bra adulta de Cebus nigritus (Barra= 10 μm ) . El
recuadro inferior a la derecha indica el sistem a sexual XY observado en los machos.

4. Resultados
90
En las m etafases con bandas C (Figura 4.24) se observó
heterocrom at ina C+ cent rom érica en todos los pares crom osóm icos. Se
observaron bandas C+ interst iciales en los pares crom osóm icos 4, 6, 12, 13
y 19. No se observaron bandas de heterocrom at ina telom érica.
Figura 4.24: Metafase con bandas C (Barra= 10 μm). Las flechas indican las bandas C+
interst iciales en los pares 4, 6, 12, 13 y 19.
4 .2 .1 .2 .2 .4 Aotus azarae
4 .2 .1 .2 .2 .4 .1 Determ inación del 2 N y NF
Para establecer el núm ero diploide (2N) y el núm ero fundam ental
(NF) correspondiente se analizaron al m enos 50 m etafases con t inción
uniform e de cada ejem plar bajo m icroscopía ópt ica. Se observó un 2N= 50,
XX para las hem bras y un 2N = 49, X1X2Y para los m achos en coincidencia
con el 2N descrito previam ente para la especie (Mudry et al. 1984b) . El núm ero
fundam ental observado fue NF= 78 para las hem bras y NF= 76 para los
m achos, en coincidencia con la presencia de un sistem a sexual m últ iple. La
Tabla 4.11 resum e los resultados obtenidos.

4. Resultados
91
Tabla 4.11: Determ inación del Núm ero Diploide (% 2N) y del Núm ero Fundam ental (NF) en los
especím enes de Aotus azarae analizados.
I ndividuo Sexo
NF
% 2 N
≤47 48 4 9 5 0 ≥51
417 H 78 4,0 8,0 16,0 7 0 ,0 4,0
418 H 78 7,8 11,8 7,8 7 0 ,6 1,9
517 M 76 6,0 6,0 7 2 ,0 16,0 0,0
519 M 76 4,0 2,0 7 6 ,0 16,0 2,0
Se corroboró la fórm ula crom osómica y se observó un com plem ento
autosóm ico com puesto, en las hem bras, por 13 pares de morfología
submetacént r ica y 11 pares acrocént r icos y en los m achos, 13 pares de
m orfología subm etacént r ica y 10 pares acrocént r icos en coincidencia con la
presencia de un sistem a sexual m últ iple X1X2Y. El cromosoma X1 es
submetacént r ico.
4 .2 .1 .2 .2 .4 .2 Pat rones de bandas G y C
El pat rón de bandas G perm it ió corroborar la asignación de especie
de todos los ejem plares a Aotus azarae (Figura 4.25) . Se observó un
sistem a sexual m últ iple form ado por la t ranslocación del crom osom a Y con
el autosom a 23 ( t Y: 23) .
El pat rón de bandas C m ost ró heterocrom at ina C+ cent rom érica en
todos los pares crom osómicos. Se observaron dos bandas C+ interst iciales
en los pares 8 y 9 (Figura 4.26) . No se observó heterocrom at ina telom érica.

4. Resultados
92
Figura 4.25: Pat rón de bandas G de una hem bra adulta de Aotus azarae (Barra= 10 μm ). El
recuadro infer ior sobre la derecha, m uest ra los crom osom as sexuales observados en los m achos.
Figura 4.26: Metafase de una hem bra de Aotus azarae con bandas C (Barra= 10 μm ) . Las
flechas indican heterocromat ina interst icial.

4. Resultados
93
4 .2 .1 .2 .3 Hom inoidea
4 .2 .1 .2 .3 .1 Pan t roglodytes
4 .2 .1 .2 .3 .1 .1 Determ inación del 2 N y NF
Para establecer el núm ero diploide (2N) y el núm ero fundam ental (NF)
correspondiente se analizaron al m enos 50 m etafases con t inción uniform e
de cada ejemplar bajo m icroscopía ópt ica. Se observó un 2N= 48, XY para el
ejemplar m acho estudiado, en coincidencia con el 2N descrito previam ente
para la especie (Yunis y Prakash 1982) . La Tabla 4.12 resum e los resultados
obtenidos.
Tabla 4.12: Determ inación del Núm ero Diploide (% 2N) y del Núm ero Fundam ental (NF) en el
espécim en de Pan t roglodytes analizado.
I ndividuo
NF
% 2 N
≤46 47 4 8 49 ≥50
654 83 12 8 8 0 0 0
Se corroboró la fórm ula crom osóm ica, observándose un com plem ento
autosóm ico com puesto por 15 pares de m orfología subm etacént r ica, 2
pares m etacént r icos y 6 pares acrocént r icos. El crom osoma X es
submetacént r ico y el cromosom a Y es un acrocént r ico pequeño.
4 .2 .1 .2 .3 .1 .2 Pat rones de bandas G y C
El pat rón de bandas G perm it ió corroborar la asignación de especie a
Pan t roglodytes (Figura 4.27) .
El pat rón de bandas C m ostró heterocromat ina cent rom érica en todos
los pares crom osómicos (Figura 4.28) . Se observó heterocromat ina C+
term inal en p ter en los pares 8, 10, 11, 16, 19, 18 y 21 y en qter en 16, 18,
20, 21, 22 y 23. Se describió una banda de heterocrom at ina C+ interst icial
en 14q.

4. Resultados
94
Figura 4.27: Pat rón de bandas G del ejem plar analizado de Pan t roglodytes (Barra= 10 μm ).
Figura 4.28: Metafase con bandas C del m acho de Pan t roglodytes (Barra= 10 μm) . Las flechas
indican la banda C+ interst icial en 14q.

4. Resultados
95
4 .2 .1 .2 .3 .2 Hom o sapiens
4 .2 .1 .2 .3 .2 .1 Determ inación del 2 N y NF
Para establecer el núm ero diploide (2N) y el núm ero fundam ental
(NF) correspondiente se analizaron al m enos 50 m etafases con t inción
uniform e de cada ejem plar bajo m icroscopía ópt ica. Se observó un
2N= 46, XY para los individuos de sexo m asculino y un 2N= 46, XX para los
individuos de sexo fem enino, en coincidencia con el 2N descrito
previam ente para la especie (Tj io y Levan 1956) . La Tabla 4.13 resum e los
resultados obtenidos.
Tabla 4.13: Determ inación del Núm ero Diploide (% 2N) y el Núm ero Fundam ental (NF) en los
indiv iduos de Hom o sapiens analizados.
I ndividuo
Sexo
NF
% 2 N
≤43 44 45 4 6 47 ≥48
592 M 91 0 3 5 9 1 1 0
593 H 92 0 0 4 9 3 3 0
594 H 92 1 3 4 8 9 3 0
635 H 92 2 1 5 9 2 0 1
637 M 91 0 4 2 9 4 0 0
Se corroboró la fórm ula crom osóm ica, observándose un com plem ento
autosóm ico com puesto por 12 pares de morfología subm etacént r ica, 5
pares m etacént r icos y 5 pares acrocént r icos. El crom osoma X es
submetacént r ico y el cromosom a Y es un acrocént r ico pequeño.
4 .2 .1 .2 .3 .2 .2 Pat rones de bandas G y C
El pat rón de bandas G obtenido para todos los ejem plares estuvo en
concordancia con el cariot ipo de individuos norm ales (Figura 4.29) .

4. Resultados
96
Figura 4.29: Pat rón de bandas G de un individuo de sexo fem enino de Hom o sapiens (Barra= 10
μm ) . El recuadro m uest ra los crom osom as sexuales en los indiv iduos de sexo m asculino.
El pat rón de bandas C m ost ró heterocrom at ina C+ cent rom érica en
todos los pares crom osóm icos. Se observaron bandas C+ per icent rom éricas
heterom órficas en los pares HSA1, HSA9, HSA16 y telom érica en el
crom osom a HSAYq (Figura 4.30) .
Figura 4.30: Metafase con bandas C de Hom o sapiens (Barra= 10 μm ). a) I ndiv iduo de sexo
masculino. Las flechas indican las bandas C+ per icent roméricas heteromórficas en los pares HSA1, HSA9,
HSA16 y telom érica en el crom osom a HSAYq. b) I ndiv iduo de sexo fem enino. Las flechas indican las
bandas C+ per icent rom éricas heteromórf icas en los pares HSA1, HSA9 y HSA16.

4. Resultados
97
4 .2 .1 .3 Hom eologías y reorganizaci ones crom osóm icas en aulladores
4 .2 .1 .3 .1 Hom o sapiens y Alouat ta caraya
Se analizaron las hom eologías cromosóm icas posibles de detectar
comparando el pat rón de bandas G de Hom o sapiens y Alouat ta caraya. El
análisis se realizó considerando tanto la inform ación provista por el pat rón
de bandas G como por el pat rón de Hibridación in situ Fluorescente (FI SH)
con sondas de crom osom as hum anos (Figura 4.31) (de Oliveira et al. 2002;
Stanyon et al. 2011) . Como resultado de esta com paración se establecieron las
reorganizaciones cromosóm icas que podrían explicar estas hom eologías.
Figura 4.31: Metafase con bandas G de A. caraya con el pat rón de hibridación para las sondas
de crom osom as hum anos indicado a la derecha (Barra= 10 μm ) (Modificada de Stanyon et al. 2011) .
En este t rabajo de Tesis analizando el pat rón de bandas G se
proponen hom eologías cromosóm icas ent re A. caraya y Hom o sapiens y se
corrobora la asignación de hom eologías establecidas por FI SH en los
estudios previos anteriorm ente refer idos.
Dado que no nos fue posible disponer de la batería com pleta de
sondas de los 23 pares crom osóm icos hum anos y que dos de las especies
de aulladores estudiadas, A. pigra (API ) y A. palliata (APA) , no han sido
analizadas hasta la fecha de redacción de este t rabajo de Tesis Doctoral, se

4. Resultados
98
obtendrá el pat rón de hibr idación de API y de APA por FI SH con sondas
hum anas a part ir de la asignación de hom eologías ent re ACA y las especies
m esoam ericanas (API y APA) .
Como antecedente a esta com paración, se tom ó en consideración el
conocim iento aceptado en la literatura de la especialidad que sost iene que,
dent ro de los PNM, el car iot ipo de Cebus apella es el car iot ipo m ás ancest ral
y es el que m uest ra la m ayor conservación cromosómica respecto del
cariot ipo hum ano (Dut r illaux 1979; Clem ente et al. 1990; García et al. 2000) .
Se considera que el cariot ipo de Alouat ta caraya está muy
reorganizado respecto de Cebus apella (de Oliveira et al. 2002) , por lo que para
el estudio de las hom eologías, se ha considerado el cariot ipo de Hom o
sapiens com o cariot ipo de referencia. Ésta es la razón por la que las flechas
que indican las reorganizaciones cromosóm icas estén situadas en el
cariot ipo de HSA y no en el de ACA.
En las Figuras 4.32, 4.33 y 4.34 y en las Tablas 4.14a y 4.14b las
hom eologías se describen para cada cromosom a desde la región p term inal
hasta la q term inal. Las Figuras indican tanto el idiogram a com o el pat rón
de bandas G para los 22 autosom as humanos y para el cromosom a X. El
crom osom a Y no se incluyó en la comparación dado que se ha comprobado
en reiteradas oportunidades en publicaciones internacionales y en
observaciones del equipo de t rabajo donde se desarrolló esta Tesis
Doctoral, que el crom osom a Y hum ano no m uest ra hom eología con el
crom osom a Y de ninguna especie de Platyrrhini. La región hom eóloga de
cada crom osom a de ACA con el respect ivo crom osom a hum ano se indica
m ediante una barra vert ical a la derecha del m ismo.
Se tom aron en consideración los siguientes reordenam ientos:
fisiones/ fusiones ( indicados con la let ra f) , inversiones ( indicados con la
let ra i) y act ivación/ inact ivación cent rom érica ( indicados con la let ra c) . En
el caso de observar más de una inversión en un m ismo crom osoma
hum ano, las m ism as se indicaron con let ras consecut ivas (ej : ia e ib) .

4. Resultados
99
Figura 4.32: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
cromosom as 1 a 6 de HSA (subrayados) y de ACA. Las flechas indican las bandas im plicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de ACA indica
la región homeóloga con el cromosom a hum ano situado a la izquierda. Las homeologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).

4. Resultados
100
Figura 4.33: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
cromosom as 7 a 12 de HSA (subrayados) y de ACA. Las flechas indican las bandas im plicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de ACA indica
la región homeóloga con el cromosom a hum ano situado a la izquierda. Las homeologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).

4. Resultados
101
Figura 4.34: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
cromosom as 13 a X de HSA (subrayados) y de ACA. Las flechas indican las bandas implicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de ACA indica
la región homeóloga con el cromosom a hum ano situado a la izquierda. Las homeologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).
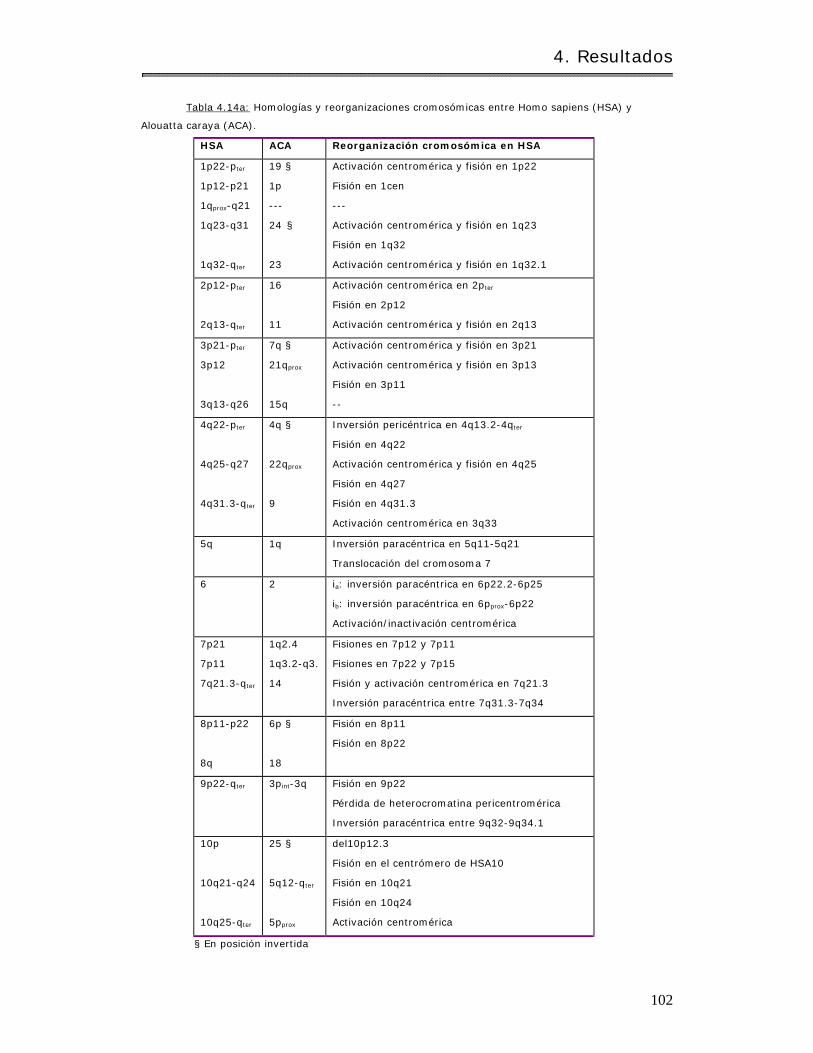
4. Resultados
102
Tabla 4.14a: Hom ologías y reorganizaciones crom osóm icas ent re Hom o sapiens (HSA) y
Alouat ta caraya (ACA) .
HSA ACA Reorganización crom osóm ica en HSA
1p22-p ter
1p12-p21
1qprox-q21
1q23-q31
1q32-q ter
19 §
1p
- - -
24 §
23
Act ivación cent rom érica y fisión en 1p22
Fisión en 1cen
- - -
Act ivación cent rom érica y fisión en 1q23
Fisión en 1q32
Act ivación cent rom érica y fisión en 1q32.1
2p12-p ter
2q13-q ter
16
11
Act ivación cent rom érica en 2p ter
Fisión en 2p12
Act ivación cent rom érica y fisión en 2q13
3p21-p ter
3p12
3q13-q26
7q §
21qprox
15q
Act ivación cent rom érica y fisión en 3p21
Act ivación cent rom érica y fisión en 3p13
Fisión en 3p11
- -
4q22-p ter
4q25-q27
4q31.3-q ter
4q §
22qprox
9
I nversión pericént r ica en 4q13.2-4q t er
Fisión en 4q22
Act ivación cent rom érica y fisión en 4q25
Fisión en 4q27
Fisión en 4q31.3
Act ivación cent rom érica en 3q33
5q 1q I nversión paracént r ica en 5q11-5q21
Translocación del crom osom a 7
6 2 ia: inversión paracént r ica en 6p22.2-6p25
ib: inversión paracént r ica en 6pprox-6p22
Act ivación/ inact ivación cent romérica
7p21
7p11
7q21.3-q ter
1q2.4
1q3.2-q3.
14
Fisiones en 7p12 y 7p11
Fisiones en 7p22 y 7p15
Fisión y act ivación cent romérica en 7q21.3
I nversión paracént r ica ent re 7q31.3-7q34
8p11-p22
8q
6p §
18
Fisión en 8p11
Fisión en 8p22
9p22-q ter 3p int-3q Fisión en 9p22
Pérdida de heterocrom at ina pericent rom érica
I nversión paracént r ica ent re 9q32-9q34.1
10p
10q21-q24
10q25-q ter
25 §
5q12-q ter
5pprox
del10p12.3
Fisión en el cent róm ero de HSA10
Fisión en 10q21
Fisión en 10q24
Act ivación cent rom érica
§ En posición invert ida
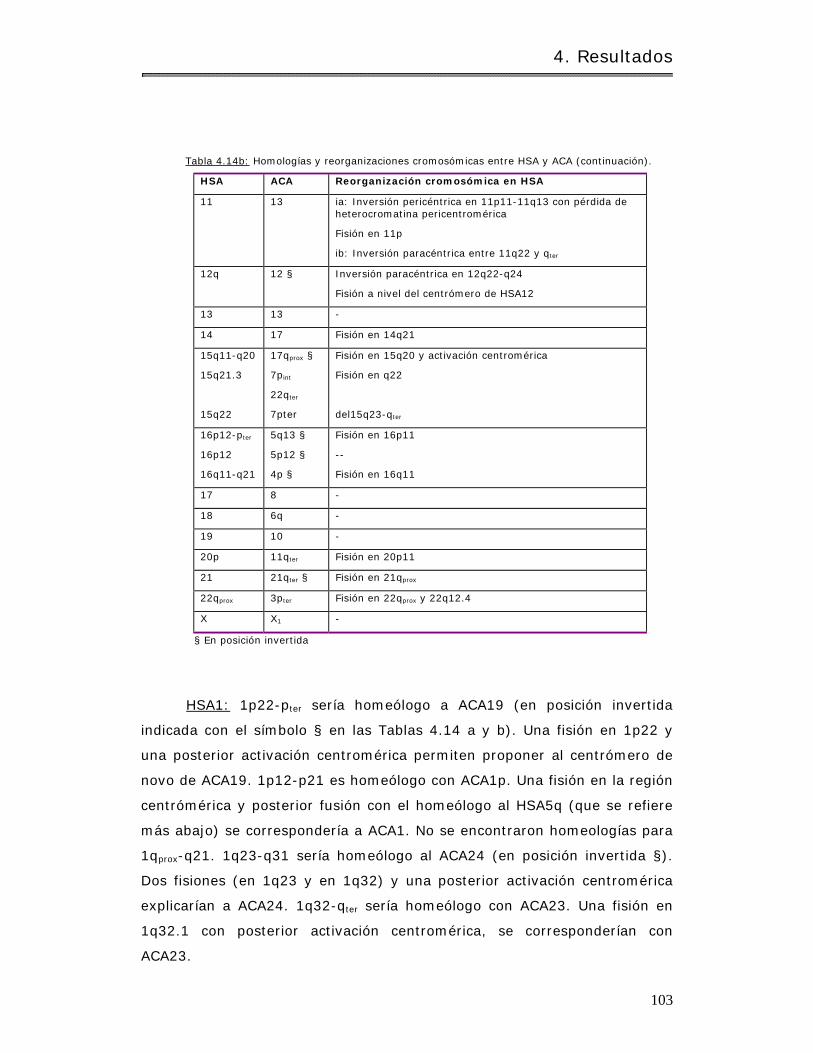
4. Resultados
103
Tabla 4.14b: Hom ologías y reorganizaciones crom osóm icas ent re HSA y ACA ( cont inuación) . HSA ACA Reorganización crom osóm ica en HSA
11 13 ia: I nversión per icént r ica en 11p11-11q13 con pérdida de heterocromat ina per icent rom érica
Fisión en 11p
ib: I nversión paracént r ica ent re 11q22 y q t er
12q 12 § I nversión paracént r ica en 12q22-q24
Fisión a nivel del cent róm ero de HSA12
13 13 -
14 17 Fisión en 14q21
15q11-q20
15q21.3
15q22
17qprox §
7p int
22q ter
7pter
Fisión en 15q20 y act ivación cent rom érica
Fisión en q22
del15q23-q ter
16p12-p ter
16p12
16q11-q21
5q13 §
5p12 §
4p §
Fisión en 16p11
- -
Fisión en 16q11
17 8 -
18 6q -
19 10 -
20p 11q ter Fisión en 20p11
21 21q ter § Fisión en 21qprox
22qprox 3p ter Fisión en 22qprox y 22q12.4
X X1 -
§ En posición invert ida
HSA1: 1p22-p ter sería hom eólogo a ACA19 (en posición invert ida
indicada con el sím bolo § en las Tablas 4.14 a y b) . Una fisión en 1p22 y
una poster ior act ivación cent rom érica perm iten proponer al cent róm ero de
novo de ACA19. 1p12-p21 es hom eólogo con ACA1p. Una fisión en la región
cent róm érica y poster ior fusión con el hom eólogo al HSA5q (que se refiere
m ás abajo) se correspondería a ACA1. No se encont raron hom eologías para
1qprox-q21. 1q23-q31 sería hom eólogo al ACA24 (en posición invert ida §) .
Dos fisiones (en 1q23 y en 1q32) y una poster ior act ivación cent rom érica
explicarían a ACA24. 1q32-q ter sería hom eólogo con ACA23. Una fisión en
1q32.1 con poster ior act ivación cent rom érica, se corresponderían con
ACA23.

4. Resultados
104
HSA2: 2p12-p ter ser ía hom eólogo con ACA16. Una fisión en 2p12 y
una poster ior act ivación cent rom érica en 2p ter se corresponderían con
ACA16. No se encont ró hom eología para 2p11-q12. 2q13-q ter sería
hom eólogo con ACA11 (con excepción de q ter) . Una fisión en 2q13 y
poster ior act ivación cent rom érica explicarían el pat rón de bandas de ACA11.
HSA3: 3p21-p ter sería hom eólogo con ACA7q (en posición invert ida
§) . Una fisión en 3p21 y posterior act ivación cent rom érica explicarían
ACA7q. No se encontró hom eología para 3p20-p13. La banda 3p12 sería
hom eóloga con ACA21qprox. Una fisión en 3p13 y act ivación cent rom érica,
junto con una segunda fisión en 3p11 seguida de la fusión con el hom eólogo
a HSA21 explicarían el pat rón de bandas de ACA21. 3q13-q26 sería
hom eólogo al ACA15q. Una fisión en 3q13 y posterior act ivación
cent rom érica habría dado lugar al ACA15.
HSA4: 4q22-p ter tendría hom eología con ACA4q (en posición invert ida
§) . Una inversión pericént r ica ent re 4q13.2 y 4q ter y una fisión en 4q22
explicarían ACA4q. No se encont ró hom eología para 4q23-q24. 4q25-q27
sería hom eólogo al ACA22qprox. Una fisión en 4q25 y poster ior act ivación
cent rom érica junto con una fisión en 4q27 habrían dado lugar al ACA22qprox.
No se encont ró hom eología para 4q26-q30. 4q31.3-q ter sería hom eólogo a
ACA9. Una fisión en 4q31.3 y una act ivación cent rom érica en 4q33
explicarían la hom eología con ACA9.
HSA5: No se encont ró hom eología para 5p. 5q tendría hom eología
con ACA1q. Una inversión paracént r ica en 5q11-q21 y la t ranslocación no
recíproca de porciones del crom osom a 7 explicarían las bandas hom eólogas
en ACA1q. En ACA1q se observa así la asociación 5/ 7/ 5/ 7.
HSA6: este cromosom a sería hom eólogo con ACA2. Dos inversiones
paracént r icas, una en 6p22.2-p25 (m arcada en la Figura 4.31 com o ia) y
ot ra ent re 6pprox y 6p22 (m arcada en la Figura 4.31 como ib) explicarían el
pat rón de bandas de ACA2.

4. Resultados
105
HSA7: 7p21 tendría hom eología con ACA1q2.4. La banda 7p11
tendría hom eología con ACA1q3.2-q3.3. Fisiones en 7p22, 7p15, 7p12 y
7p11, sufr iendo los fragm entos resultantes una t ranslocación no recíproca
hacia el hom eólogo de HSA5, explicarían los reordenam ientos observados.
7q21.3-q ter sería hom eólogo a ACA14. Una fisión en 7q21.3 seguida de una
act ivación cent rom érica junto con una inversión paracéntr ica en 7q31.3-
7q34 explicarían ACA14.
HSA8: 8p22-p11 sería hom eólogo con ACA6p (en posición invert ida
§) . Dos fisiones (en 8p11 y en 8p22) , seguida de una fusión con el
hom éologo de HSA18 (const ituyendo la asociación 8/ 18 referida en la
literatura) , explicarían ACA6p. 8q sería hom eólogo al ACA18. El cent róm ero
de HSA8 correspondería al cent róm ero de ACA18.
HSA9: este crom osoma sería hom eólogo con ACA3. Una deleción en
9p22, la pérdida de la heterocrom at ina pericent rom érica de HSA9 y una
inversión paracént r ica en 9q32-q34.1 explicarían ACA3.
HSA10: 10p sería hom eólogo con ACA25 (en posición invert ida §) .
Una deleción en 10p12.3 y una fisión en el cent róm ero de HSA10
explicarían ACA25. No se encont ró hom eología para 10qprox. 10q21-q24
tendría hom eología con ACA5q12-q ter. Dos fisiones, una en 10q21 y ot ra en
10q24 explicarían este fragm ento. 10q25-q ter sería hom eólogo a ACA5pprox.
Una fisión seguida act ivación cent rom érica en 10q24 explicaría este
fragm ento. La t ranslocación con segmentos hom eólogos a HSA16 explicaría
ACA5, en dónde se observa la sintenia 16/ 10/ 16/ 10.
HSA11: este cromosom a presentaría hom eología con ACA13. Una
inversión pericént r ica (m arcada ia en la Figura 4.32) en 11p11-q13 (con
pérdida de heterocrom at ina pericent rom érica) y una fisión en 11p12, j unto
con una inversión paracént r ica ent re 11q22 y q ter (m arcada ib en la Figura
4.32) explicarían ACA13. No se encont ró hom eología en ACA para 11p13-
pter.

4. Resultados
106
HSA12: No se encont ró hom eología para 12p. 12q presentaría
hom eología con ACA12. Una fisión a nivel del cent róm ero y una inversión
paracént r ica ent re 12q22-q24 explicarían ACA12.
HSA13: este cromosom a presentaría hom eología con ACA13. La única
reorganización encont rada ent re estos dos crom osom as sería una deleción
en HSA13q34.
HSA14: este crom osoma sería hom eólogo al ACA17q2.1-q ter. Una
deleción en 14q32 y la t ranslocación con el hom eólogo a HSA15q11-q20
(que conform arían la asociación 14/ 15) sería la única reorganización
encont rada ent re estos crom osom as.
HSA15: 15q11-q13 tendría hom eología con ACA17qprox (en posición
invert ida §) . Una fisión en 15q20 y poster ior act ivación cent rom érica, j unto
con la t ranslocación con el hom eólogo a HSA15q11-q20 (que conform arían
la asociación 14/ 15) explicarían ACA17. La banda 15q21.3 sería hom eóloga
tanto al ACA7p int (en posición invert ida §)como a ACA22q ter, m ient ras que la
banda 15q22 tendría hom eología con ACA7p ter. Una fisión en 15q22 y las
poster iores t ranslocaciones con segm entos de HSA3 (en el caso de ACA7) y
con segm entos hom eólogos a HSA4 (en el caso de HSA22) , junto con la
del15q23-q ter, explicarían ACA7 y HSA4, respect ivam ente.
HSA16: 16p12-p ter tendría hom eología con ACA5q13 (en posición
invert ida §) . 16p12 sería hom eólogo al ACA5p12. Una fisión en 16p11 y la
t ranslocación de segm entos hom eólogos a HSA10 (conform ando la sintenia
16/ 10/ 16/ 10) explicarían ACA5. 16q11-q21 tendría hom eología con ACA4p
(en posición invert ida §) . Una fisión en 16q11 y la fusión con un segm ento
hom eólogo al HSA4q (por la sintenia 4/ 16) explicarían ACA4.
HSA17: este crom osom a presentaría hom eología con ACA8. No se
observan reorganizaciones ent re estos crom osom as.

4. Resultados
107
HSA18: este cromosom a sería hom eólogo al ACA6q. Una fusión con
el hom eólogo al 8p11-p22, que se correspondería a la sintenia 8/ 18,
explicaría ACA6.
HSA19: sería hom eólogo al ACA10. No se observan rearreglos ent re
estos crom osom as.
HSA20: este crom osom a presentaría hom eología con ACA11q ter. Una
inact ivación cent rom érica y una fisión en 20p11, seguida de una fusión con
un segm ento hom eólogo al HSA2q13-q ter darían lugar al ACA11.
HSA21: este crom osom a sería hom eólogo al ACA21q ter (en posición
invert ida §) . Se postula una fisión en 21qprox y una fusión con un segm ento
hom eólogo al HSA3p12 (dada la presencia de la sintenia 3/ 21 en ACA21) ,
explicarían ACA21.
HSA22: este cromosom a sería hom eólogo al ACA3p ter. Dos fisiones,
en 22qprox y 22q12.4 explicarían ACA3p ter.
HSAX: este crom osoma sería hom eólogo al ACAX1. No se observan
reorganizaciones ent re estos cromosom as.
Estos resultados nos han perm it ido establecer t res t ipos de relaciones
ent re los cromosom as de HSA y los de ACA:
1. Crom osom as hum anos representados en un único crom osom a de
ACA: 6, 11, 12, 13, 14, 17, 19 y X.
2. Crom osom as hum anos representados en un solo crom osom a de
ACA y acompañados por regiones hom eólogas en m ás de un
crom osom a HSA en el m ism o crom osom a de ACA: 5, 9, 10, 16,
20, 21, 22.

4. Resultados
108
3. Crom osom as hum anos representados en m ás de un crom osoma
de ACA: 2, 7, 8, 10, 16, (en dos crom osom as de ACA) ; 3, 4, 15
(en t res crom osom as de ACA) ; 1 (en cuat ro cromosom as de
ACA) .
Con respecto a las reorganizaciones crom osómicas podem os agrupar
a los crom osom as humanos (HSA) en 2 grupos:
1. Los que no precisan ninguna reorganización crom osómica para ser
hom ologados con alguno de ACA: 13, 14, 17, 19 y X.
2. Los que precisan más de una reorganización para ser
hom ologados con los de ACA: 1, 2, 3, 7, 8, 10, 16, 20, 21, 22
( fusiones/ fisiones y act ivaciones cent rom éricas) , 4, 5, 6, 9, 11,
12, 15, 18 ( fusiones/ fisiones, deleciones, act ivaciones
cent rom éricas e inversiones paracént r icas/ per icént r icas) .
El t ipo de reorganizaciones cromosómicas que encontram os al
com parar los crom osomas HSA con los de ACA son, por orden de
frecuencia: fisiones, act ivaciones cent rom éricas, inversiones
paracént r icas, fusiones e inversiones per icént r icas.

4. Resultados
109
4 .2 .1 .3 .2 Alouat ta caraya y Alouat ta pigra
Se analizaron las hom eologías cromosóm icas posibles de detectar
comparando el pat rón de bandas G de Alouat ta caraya y A. pigra.
Tal com o fuera refer ido en la sección anter ior obtendrá el pat rón de
hibridación con sondas hum anas de API y APA a part ir de la asignación de
hom eologías entre ACA y las especies m esoam ericanas (API y APA) .
Como resultado de esta comparación se establecieron las
reorganizaciones crom osóm icas que podrían explicar estas hom eologías.
Dado que se ut ilizará el cariot ipo de A. caraya para establecer las
hom eologías con el car iot ipo humano, se lo tom ará com o referencia en este
análisis, por lo que las flechas que indican las reorganizaciones
crom osómicas estarán situadas en el cariot ipo de ACA y no en el de API .
Sin embargo, es pert inente aclarar que la polaridad de los cam bios se
podría originar en la dirección opuesta, es decir de API a ACA, lo cual no
puede se determ inado a pr ior i. Es con posterior idad al análisis filogenét ico
que se podrá establecer la polar idad de los cam bios ocurr idos ent re estas
especies.
En las Figuras 4.35, 4.36 y 4.37 se descr iben las hom eologías desde
p term inal hasta q term inal. Para la lectura de estas figuras es necesario
proceder según descrito en la sección 4.2.1.3.1. La Tabla 4.15 resum e la
inform ación obtenida de las Figuras. No se tom aron en cuenta los
crom osom as sexuales resultado de la t ranslocación Y-autosom a (Y1 e Y2) ,
dado que se considera que la hom eología del autosoma que dio or igen al
sexual múlt iple estaría cubierta al analizar a X2.

4. Resultados
110
Figura 4.35: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
cromosom as 1 a 9 de ACA (subrayados) y de API . Las flechas indican las bandas im plicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de API indica
la región hom eóloga con el crom osom a de ACA situado a la izquierda. Las hom eologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).

4. Resultados
111
Figura 4.36: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
cromosom as 10 a 18 de ACA (subrayados) y de API . Las flechas indican las bandas im plicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de API indica
la región hom eóloga con el crom osom a de ACA situado a la izquierda. Las hom eologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).

4. Resultados
112
Figura 4.37: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
crom osom as 19 a X1 de ACA (subrayados) y de API . Las flechas indican las bandas implicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de API indica
la región hom eóloga con el crom osom a de ACA situado a la izquierda. Las hom eologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).
No se pudieron asignar hom eologías con ACA para los pares
API 4pprox, API 14 y API 21 m ediante el nivel de resolución y técnicas
citogenét icas aplicadas en este t rabajo com parat ivo.
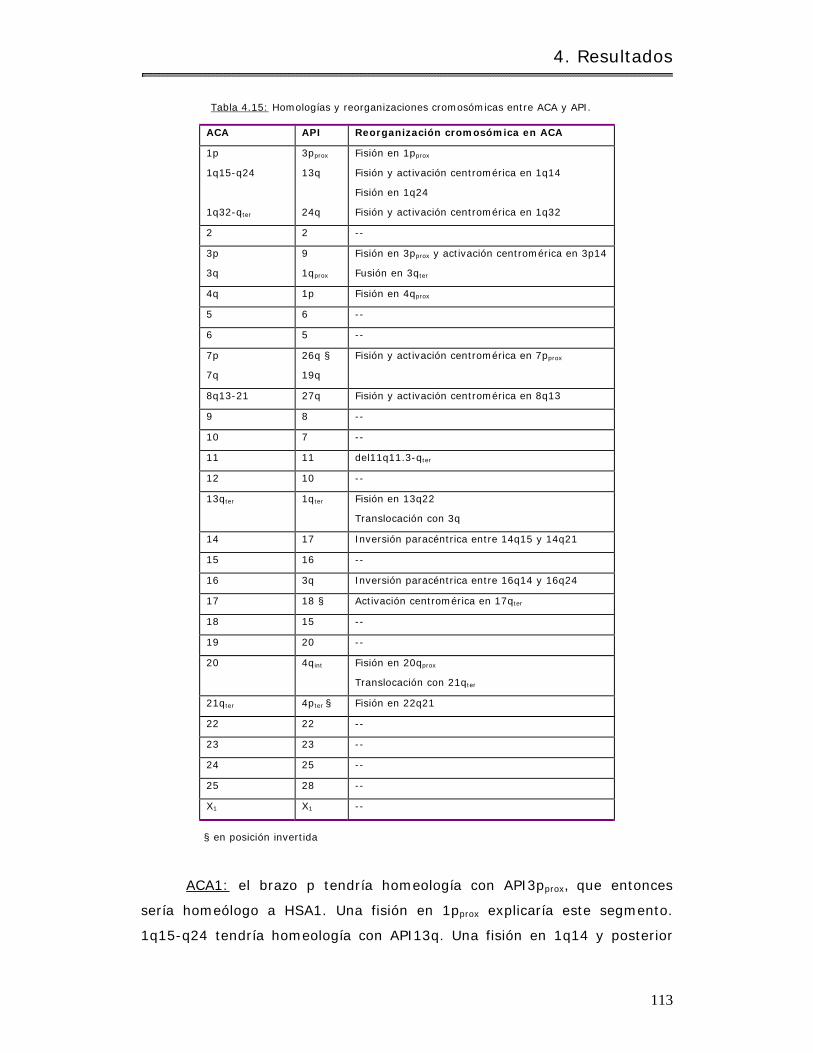
4. Resultados
113
Tabla 4.15: Hom ologías y reorganizaciones crom osóm icas ent re ACA y API .
§ en posición invert ida
ACA1: el brazo p tendría hom eología con API 3pprox, que entonces
sería hom eólogo a HSA1. Una fisión en 1pprox explicaría este segm ento.
1q15-q24 tendría hom eología con API 13q. Una fisión en 1q14 y poster ior
ACA API Reorganización crom osóm ica en ACA
1p
1q15-q24
1q32-q ter
3pprox
13q
24q
Fisión en 1pprox
Fisión y act ivación cent rom érica en 1q14
Fisión en 1q24
Fisión y act ivación cent rom érica en 1q32
2 2 - -
3p
3q
9
1qprox
Fisión en 3pprox y act ivación cent rom érica en 3p14
Fusión en 3q ter
4q 1p Fisión en 4qprox
5 6 - -
6 5 - -
7p
7q
26q §
19q
Fisión y act ivación cent rom érica en 7pprox
8q13-21 27q Fisión y act ivación cent rom érica en 8q13
9 8 - -
10 7 - -
11 11 del11q11.3-q t er
12 10 - -
13q ter 1q ter Fisión en 13q22
Translocación con 3q
14 17 I nversión paracént r ica ent re 14q15 y 14q21
15 16 - -
16 3q I nversión paracént r ica ent re 16q14 y 16q24
17 18 § Act ivación cent rom érica en 17q t er
18 15 - -
19 20 - -
20 4q int Fisión en 20qprox
Translocación con 21q t er
21q ter 4p ter § Fisión en 22q21
22 22 - -
23 23 - -
24 25 - -
25 28 - -
X1 X1 - -

4. Resultados
114
act ivación cent rom érica, junto con una fisión en 1q24 explicarían API 13.
1q32-q ter sería hom eólogo a API 24q. Una fisión y act ivación cent rom érica en
1q32 explicarían API 24. De aquí se desprende que la asociación/ sintenia
5/ 7/ 5/ 7 se conservaría sólo parcialm ente, con API 24 com prendiendo una
asociación 5/ 7.
ACA2: este cromosom a sería hom eólogo a API 2, confirm ándose la
conservación de HSA6 en API . No se observaron reordenam ientos ent re
estos dos crom osom as.
ACA3: el brazo p tendría hom eología con API 9, hom eólogo entonces
a HSA22. Una fisión en 3pprox y una act ivación cent rom érica en 3p14
explicarían API 9. El brazo q tendría hom eología con API 1qprox, que tendría
hom eología con HSA9. Una fusión en 3q ter con ACA13q ter y la form ación de
un organizador nucleolar ent re estos segm entos explicarían API 1q. De estos
datos surge que la sintenia 9/ 22 se encont raría rota.
ACA4: no se encont ró hom eología para ACA4p. El brazo q tendría
hom eología con API 1p, que tendría hom eología con HSA4. Una fisión en
4qprox explicaría API 1p. De esta m anera se infir iere que la sintenia 4/ 16
estaría rota en API .
ACA5: este crom osom a sería hom eólogo a API 6, conservándose así la
asociación 16/ 10/ 16/ 10. No se observaron reordenam ientos ent re estos dos
crom osom as.
ACA6: este cromosom a sería hom eólogo a API 5, estando conservada
la sintenia 8/ 18. No se observaron rearreglos ent re estos dos crom osom as.
ACA7: 7p tendría hom eología con API 26q (en posición invert ida §) ,
que sería hom eólogo a HSA15. Una fisión en 7pprox y una poster ior
act ivación cent rom érica explicarían API 26. 7q sería hom eólogo a API 19q,
con hom eología a HSA3, cuyo cent róm ero correspondería al de ACA7. De
aquí se desprende que la sintenia 3/ 15 estaría rota en API .

4. Resultados
115
ACA8: 8q13-q21 tendría hom eología con API 27q. Una fisión en 8q13
y posterior act ivación cent rom érica explicarían este cromosom a. Habría una
conservación parcial de HSA17.
ACA9: este crom osom a sería hom eólogo a API 8, por lo que este
crom osom a tendría hom eología con HSA4. No se observaron rearreglos
ent re estos dos crom osom as.
ACA10: este cromosom a sería hom eólogo a API 7, por lo que este
crom osom a tendría hom eología con HSA19. No se observaron rearreglos
ent re estos dos crom osom as.
ACA11: este crom osom a sería hom eólogo a API 11. Una
posible del11q11.3-q ter sería el único reordenam iento observado ent re estos
dos crom osom as. Ot ras hipótesis posibles son una t ranslocación a ot ro
crom osom a o una fisión que de lugar a ot ro crom osom a. Se infiere así que
la sintenia 2/ 20 estaría rota.
ACA12: este crom osom a sería hom eólogo a API 10, por lo que API 10
sería hom eólogo a HSA12. No se observaron rearreglos ent re estos dos
crom osom as.
ACA13: 13q ter tendría hom eología con API 1q ter, que así tendría
hom eología con HSA11. Una fisión en 13q22 y poster ior t ranslocación con
ACA3q explicaría API 1q.
ACA14: este crom osom a sería hom eólogo a API 17 y por lo tanto
API 17 tendría hom eología con HSA7. Una inversión paracént r ica ent re
14q15 y 14q21 sería el único reordenam iento observado ent re estos dos
crom osom as.

4. Resultados
116
ACA15: este crom osom a sería hom eólogo a API 16, el cual
m antendría hom eología con HSA3. No se observaron rearreglos ent re estos
dos cromosom as.
ACA16: este crom osom a tendría hom eología con API 3q, el cual sería
hom eólogo a HSA2. Una inversión paracént r ica ent re 16q14 y 16q24 sería
el único rearreglo observado ent re ACA16 y API 3q.
ACA17: este crom osom a sería hom eólogo a API 18 (en posición
invert ida §) . Una act ivación cent rom érica en ACA17q ter sería el único
rearreglo observado ent re estos dos crom osom as. De estos datos se infiere
que la sintenia 14/ 15 estaría conservada en API .
ACA18: este crom osom a sería hom eólogo a API 15, por lo que este
crom osom a tendría hom eología con HSA8. No se observaron
reordenam ientos ent re estos dos crom osom as.
ACA19: este crom osom a sería hom eólogo a API 20, el cual entonces
tendría hom eología con HSA1. No se observaron rearreglos ent re estos dos
crom osom as.
ACA20: este crom osom a tendría hom eología con API 4q int, porción de
API 4 que sería hom eóloga a HSA13. Una fisión en 20qprox y una
t ranslocación con ACA21q ter explicarían API 4q.
ACA21: 21qter tendría hom eología con API 4p ter (en posición invert ida
§) , que sería hom eólogo a HSA21. Una fisión en 22q21 y la posterior
t ranslocación con ACA20 explicarían API 4q. De estos datos se desprende
que la sintenia 3/ 21 se encont raría rota en API .
ACA22: este cromosom a sería hom eólogo a API 22, que tendría
hom eología con HSA4/ HSA15, encontrándose así conservada la asociación
4/ 15 en API . No se observaron rearreglos ent re estos dos crom osomas.

4. Resultados
117
ACA23: este cromosom a sería hom eólogo a API 20, que tendría
hom eología con HSA1. No se observaron rearreglos ent re estos dos
crom osom as.
ACA24: este cromosom a sería hom eólogo a API 25, que tendría
hom eología con HSA1. No se observaron rearreglos ent re estos dos
crom osom as.
ACA25: este crom osom a sería hom eólogo a API 28, por lo que este
crom osom a tendría hom eología con HSA10. No se observaron rearreglos
ent re estos dos crom osom as.
ACAX1: este crom osoma sería hom eólogo a API X1. No se observaron
rearreglos ent re estos dos crom osom as.
Estos resultados nos han perm it ido establecer t res t ipos de relaciones
ent re los crom osom as de ACA y los de API :
1. Cromosom as de ACA representados en un único cromosom a de API :
2, 5, 6, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24,
25 y X1.
2. Cromosom as de ACA representados en un solo crom osom a de API y
acompañados por regiones hom eólogas de m ás de un cromosoma de
ACA en el m ismo cromosom a de API : 4, 13, 20 y 21.
3. Cromosom as de ACA representados en dos o m ás crom osom as de
API : 1, 3 y 7.
Con respecto a las reorganizaciones crom osómicas podem os agrupar
a los crom osom as de ACA en 2 grupos:

4. Resultados
118
1. Los que no precisan ninguna reorganización crom osóm ica para se
homologados con alguno de API : 2, 5, 6, 9, 10, 12, 15, 18, 19,
22, 24, 25 y X1.
2. Los que precisan una o m ás reorganizaciones para ser
hom ologados con API : 1, 3, 4, 7, 8, 11, 13, 14, 16, 17, 20, 21 y
23 ( fusiones/ fisiones, act ivaciones cent rom éricas, deleciones,
inversiones paracént r icas y t ranslocaciones) .
El t ipo de reorganizaciones cromosóm icas que se encont raron al
com parar los crom osom as de ACA con los de API son, por orden de
frecuencia: fisiones, act ivaciones cent rom éricas, inversiones
paracént r icas, fusiones, deleciones y t ranslocaciones.
4 .2 .1 .3 .3 Alouat ta caraya y Alouat ta pa llia ta
Se analizaron las hom eologías cromosóm icas posibles de detectar
comparando el pat rón de bandas G de Alouat ta caraya y A. palliata.
Como resultado de esta comparación se establecieron las
reorganizaciones crom osóm icas que podrían explicar estas hom eologías. El
cariot ipo de ACA se tom ó com o referencia en este análisis, por lo que las
flechas que indican las reorganizaciones cromosóm icas estarán situadas en
el car iot ipo de ACA y no en el de APA. Para este análisis se sigue
m anteniendo com o válida la aclaración realizada en la sección 4.2.1.3.2
respecto a establecer a pr ior i la polaridad de los cam bios.
En las Figuras 4.38, 4.39 y 4.40 se descr iben las hom eologías desde
p term inal hasta q term inal. Para la lectura de estas Figuras es aconsejable
proceder según descrito en la sección 4.2.1.3.1. La Tabla 4.16 resum e la
inform ación obtenida de las Figuras.

4. Resultados
119
Figura 4.38: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
cromosom as 1 a 9 de ACA (subrayados) y de APA. Las flechas indican las bandas implicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de APA indica
la región hom eóloga con el crom osom a de ACA situado a la izquierda. Las hom eologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).

4. Resultados
120
Figura 4.39: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
cromosom as 10 a 18 de ACA (subrayados) y de APA. Las f lechas indican las bandas im plicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de APA indica
la región hom eóloga con el crom osom a de ACA situado a la izquierda. Las hom eologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).

4. Resultados
121
Figura 4.40: Reorganizaciones crom osóm icas propuestas para explicar las hom eologías de los
crom osom as 19 a X1 de ACA (subrayados) y de APA. Las flechas indican las bandas im plicadas en las
reorganizaciones crom osóm icas. La barra vert ical situada a la derecha de los crom osom as de APA indica
la región hom eóloga con el crom osom a de ACA situado a la izquierda. Las hom eologías se descr iben
desde p term inal hasta q term inal. f: fusión/ fisión; i: inversión; c: act ivación/ inact ivación cent romérica
(Barra= 10 μm ).
No se pudieron asignar hom eologías con ACA a los pares APA2p y
APA4q m ediante el nivel de resolución y técnicas citogenét icas aplicadas en
este t rabajo de Tesis.
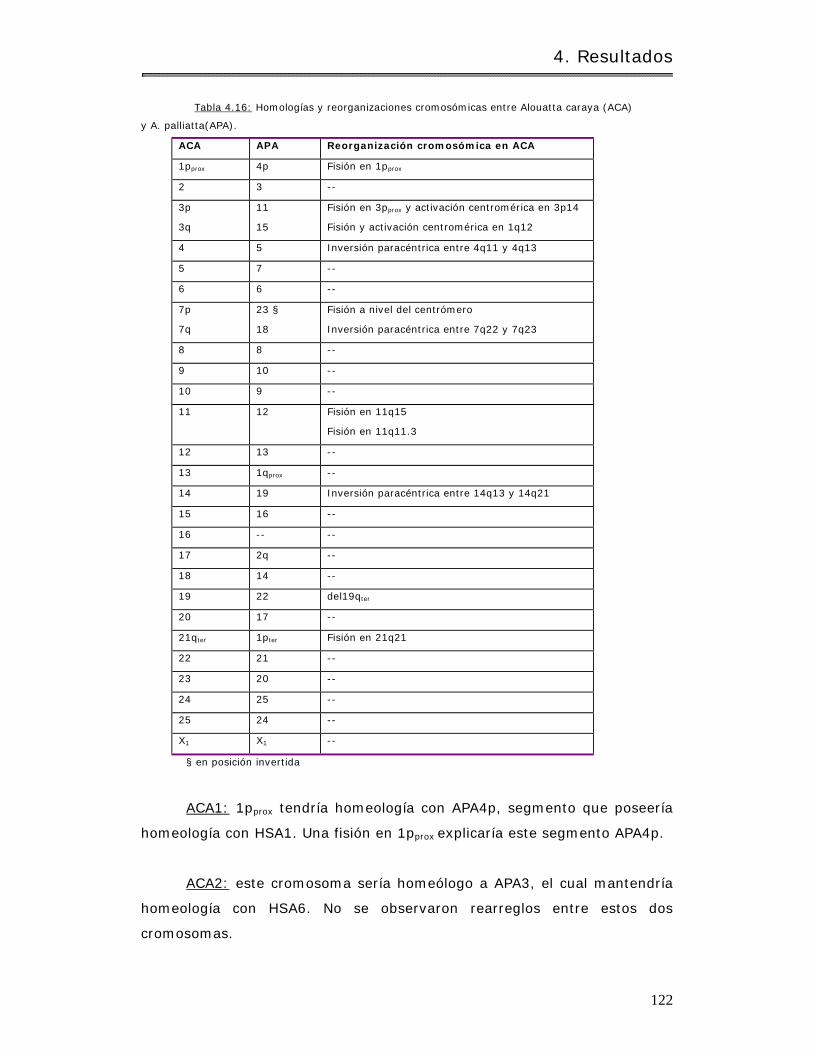
4. Resultados
122
Tabla 4.16: Hom ologías y reorganizaciones crom osóm icas ent re Alouat ta caraya (ACA)
y A. palliat ta(APA) .
ACA APA Reorganización crom osóm ica en ACA
1pprox 4p Fisión en 1pprox
2 3 - -
3p
3q
11
15
Fisión en 3pprox y act ivación cent rom érica en 3p14
Fisión y act ivación cent rom érica en 1q12
4 5 I nversión paracént r ica ent re 4q11 y 4q13
5 7 - -
6 6 - -
7p
7q
23 §
18
Fisión a nivel del cent róm ero
I nversión paracént r ica ent re 7q22 y 7q23
8 8 - -
9 10 - -
10 9 - -
11 12 Fisión en 11q15
Fisión en 11q11.3
12 13 - -
13 1qprox - -
14 19 I nversión paracént r ica ent re 14q13 y 14q21
15 16 - -
16 - - - -
17 2q - -
18 14 - -
19 22 del19q ter
20 17 - -
21q ter 1p ter Fisión en 21q21
22 21 - -
23 20 - -
24 25 - -
25 24 - -
X1 X1 - -
§ en posición invert ida
ACA1: 1pprox tendría hom eología con APA4p, segm ento que poseería
hom eología con HSA1. Una fisión en 1pprox explicaría este segm ento APA4p.
ACA2: este crom osom a sería hom eólogo a APA3, el cual m antendría
hom eología con HSA6. No se observaron rearreglos ent re estos dos
crom osom as.

4. Resultados
123
ACA3: el brazo p sería hom eólogo a APA11, que m antendría
hom eología con HSA22. Una fisión en 3pprox y una act ivación cent rom érica
en 3p14 explicarían APA11. 3q tendría hom eología con APA15, el cual sería
hom eólogo a HSA9. Una fisión y una poster ior act ivación cent rom érica
explicarían APA15. Se infiere así que la sintenia 9/ 22 estaría rota en APA.
ACA4: este crom osom a sería hom eólogo a APA5, el cual m antendría
hom eología con HSA16 en APA5p y con HSA4 en APA5q, con lo que se
conservaría la asociación 16/ 4. El único reordenam iento encont rado ent re
estos dos cromosom as es una inversión paracént r ica ent re 4q11 y 4q13.
ACA5: este crom osoma sería hom eólogo a APA7, el cual conservaría
así la sintenia 16/ 10/ 16/ 10. No se observaron rearreglos ent re estos dos
crom osom as.
ACA6: este crom osom a sería hom eólogo a APA6, el cual m antendría
hom eología con HSA8 en APA6p y con HSA18 en APA6q, conservándose la
asociación 8/ 18. No se observaron rearreglos ent re estos dos crom osom as.
ACA7: 7p tendría hom eología con APA23 (en posición invert ida §) ,
que tendría hom eología con HSA15. Una fisión en 7pprox y posterior
act ivación cent rom érica explicarían APA23. 7q sería hom eólogo a APA18,
que sería hom eólogo a HSA3. Una inversión paracént rica ent re 7q22 y 7q23
sería el único reordenam iento encontrado ent re estos dos cromosom as. Se
infiere así que la sintenia 3/ 15 estaría rota en APA.
ACA8: este crom osoma sería hom eólogo a APA8, el cual conservaría
hom eología con HSA17. No se observaron rearreglos ent re estos dos
crom osom as.
ACA9: este crom osom a sería hom eólogo a APA10, el cual conservaría
hom eología con HSA4. No se observaron rearreglos ent re estos dos
crom osom as.

4. Resultados
124
ACA10: este cromosom a sería hom eólogo a APA9, el cual conservaría
hom eología con HSA19. No se observaron rearreglos ent re estos dos
crom osom as.
ACA11: ACA11qprox tendría hom eología con APA12q (m enos
APA12qprox) , el cual sería hom eólogo a HSA2. Dos fisiones, una en 11q15 y
ot ra en 11q13 explicarían el segm ento hom eólogo de APA12. No se
encont ró hom eología para ACA1q ter. La sintenia 2/ 20 estaría rota en APA.
ACA12: este crom osom a sería hom eólogo a APA13, el cual
conservaría hom eología con HSA12. No se observaron reordenam ientos
ent re estos dos crom osom as.
ACA13: Este cromosom a sería hom eólogo a APA1qprox, segm ento que
conservaría hom eología con HSA13.
ACA14: este crom osom a sería hom eólogo a APA19, el cual
m antendría hom eología con HSA7. El único rearreglo encont rado ent re
estos dos crom osom as es una inversión paracént r ica ent re 14q13 y 14q21.
ACA15: este crom osom a sería hom eólogo a APA16, el cual
conservaría hom eología con HSA3. No se observaron rearreglos ent re estos
dos cromosom as.
ACA16: No se encontraron hom eologías para este crom osom a en
APA.
ACA17: este crom osom a tendría hom eología con APA2q,
conservándose así en esta especie la sintenia 14/ 15. No se encontraron
rearreglos ent re ACA17 y APA2q.
ACA18: este crom osom a sería hom eólogo a APA14, el cual
conservaría hom eología con HSA8. No se observaron rearreglos ent re estos
dos cromosom as.

4. Resultados
125
ACA19: este crom osom a sería hom eólogo a APA22, el cual
conservaría hom eología con HSA1. Una del19q ter sería el único rearreglo
encont rado ent re estos dos cromosom as.
ACA20: este crom osom a sería hom eólogo a APA17, el cual
conservaría hom eología con HSA13. No se observaron reordenam ientos
ent re estos dos crom osom as.
ACA21: 21q ter presenta hom eología con APA1p ter, segm ento que sería
hom eólogo a HSA21. Una fisión en 21q21 explicaría este segm ento de
APA1p. La sintenia 3/ 21 se encont raría rota en APA.
ACA22: este crom osom a sería hom eólogo a APA21, infir iéndose que
la sintenia 4/ 15 estaría conservada en APA. No se observaron rearreglos
ent re estos dos crom osom as.
ACA23: este crom osom a sería hom eólogo a APA20, el cual
conservaría hom eología con HSA1. No se observaron rearreglos ent re estos
dos cromosom as.
ACA24: este crom osom a sería hom eólogo a APA25, el cual
conservaría hom eología con HSA1. No se observaron rearreglos ent re estos
dos cromosom as.
ACA25: este crom osom a sería hom eólogo a APA24, el cual
conservaría hom eología con HSA10. No se observaron rearreglos ent re
estos dos crom osom as.
ACAX1: este cromosom a sería hom eólogo a APAX1. No se observaron
rearreglos ent re estos dos crom osom as.
Estos resultados nos han perm it ido establecer t res t ipos de relaciones
ent re los crom osom as de ACA y los de APA:

4. Resultados
126
1. Cromosom as de ACA representados en un único cromosom a de APA:
todos con excepción de ACA7 y ACA3.
2. Cromosom as de ACA representados en un solo crom osom a de APA y
acompañados por regiones hom eólogas de m ás de un cromosoma de
ACA en el m ismo cromosom a de APA: 13, 17, 20 y 21.
3. Cromosom as de ACA representados en dos cromosom as de APA: 1, 3
y 7.
Con respecto a las reorganizaciones crom osóm icas podem os agrupar a
los crom osom as de ACA en 2 grupos:
1. Los que no precisan ninguna reorganización crom osóm ica para se
hom ologados con alguno de APA: 2, 5, 6, 8, 9, 10, 12, 15, 17, 18,
20, 22, 23, 24, 25 y X1.
2. Los que precisan una o m ás reorganizaciones para ser hom ologados
con APA: 1, 3, 4, 7, 11, 14, 19 y 21 ( fusiones/ fisiones, act ivaciones
cent rom éricas e inversiones paracént r icas) .
El t ipo de reorganizaciones cromosóm icas que se encont raron al
com parar los crom osom as de ACA con los de APA son, por orden de
frecuencia: fisiones/ fusiones, inversiones paracént r icas y act ivaciones
cent rom éricas.
4 .2 .1 .3 .4 Com paración con ot ros aulladores
Los datos de análisis de hom eologías de las restantes especies de
aulladores con ACA realizadas en este t rabajo de Tesis y en la literatura se
volcaron en la Tabla 4.17. Estas homeologías serán ut ilizadas
poster iorm ente para confeccionar la m at r iz básica de datos para los
estudios de filogenias en aulladores.
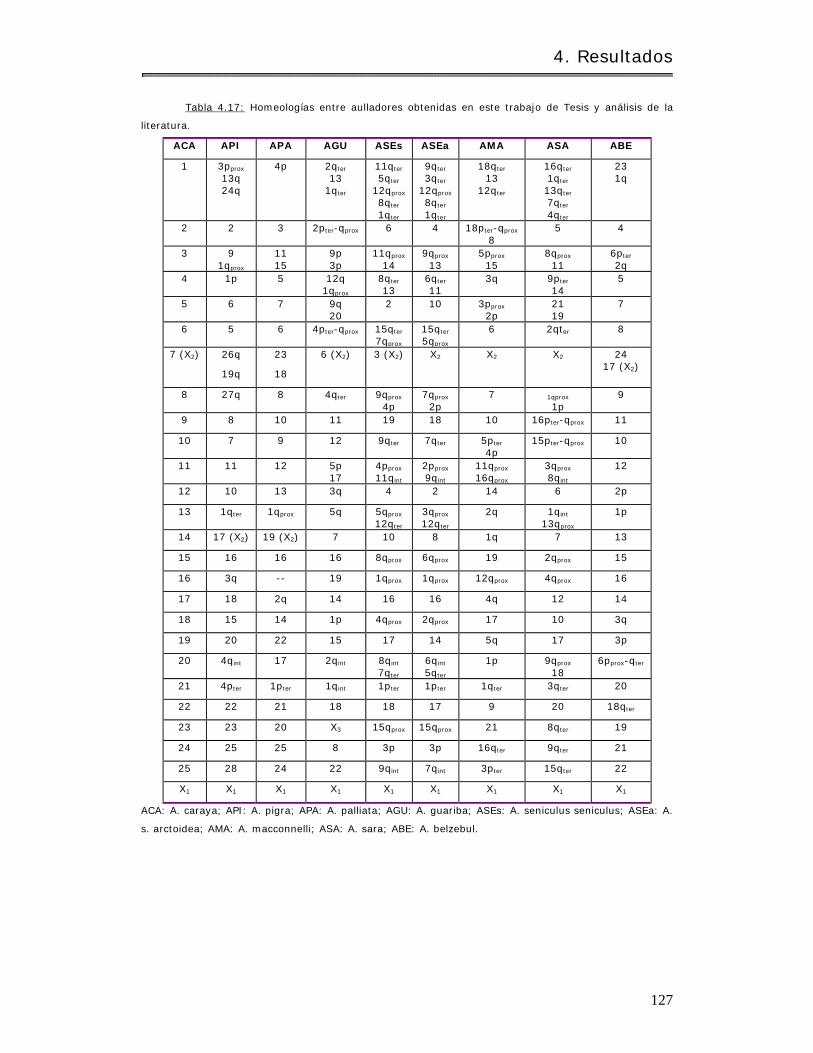
4. Resultados
127
Tabla 4.17: Homeologías ent re aulladores obtenidas en este t rabajo de Tesis y análisis de la
literatura.
ACA API APA AGU ASEs ASEa AMA ASA ABE
1 3pprox 13q 24q
4p
2q ter 13
1q ter
11q ter 5q ter
12qprox 8q ter 1q ter
9q ter 3q ter
12qprox 8q ter 1q ter
18q ter 13
12q ter
16q ter 1q ter
13q ter 7q ter 4q ter
23 1q
2 2 3 2p ter-qprox 6 4 18p ter-qprox 8
5 4
3 9 1qprox
11 15
9p 3p
11qprox 14
9qprox 13
5pprox 15
8qprox 11
6p ter 2q
4 1p 5 12q 1qprox
8q ter 13
6q ter 11
3q 9p ter 14
5
5 6 7 9q 20
2 10 3pprox 2p
21 19
7
6 5 6 4p ter-qprox 15q ter 7qprox
15q ter 5qprox
6 2qt er 8
7 (X2) 26q
19q
23
18
6 (X2) 3 (X2) X2 X2 X2 24 17 (X2)
8 27q 8 4q ter 9qprox 4p
7qprox 2p
7 1qprox
1p 9
9 8 10 11 19 18 10 16p ter-qprox 11
10 7 9 12 9q ter 7q ter 5p ter 4p
15p ter-qprox 10
11 11 12 5p 17
4pprox 11q int
2pprox 9q int
11qprox 16qprox
3qprox 8q int
12
12 10 13 3q 4 2 14 6 2p
13 1q ter 1qprox 5q 5qprox 12q ter
3qprox 12q ter
2q 1q int 13qprox
1p
14 17 (X2) 19 (X2) 7 10 8 1q 7 13
15 16 16 16 8qprox 6qprox 19 2qprox 15
16 3q - - 19 1qprox 1qprox 12qprox 4qprox 16
17 18 2q 14 16 16 4q 12 14
18 15 14 1p 4qprox 2qprox 17 10 3q
19 20 22 15 17 14 5q 17 3p
20 4q int 17 2q int 8q int
7q ter 6q int 5q ter
1p 9qprox 18
6pprox-q ter
21 4p ter 1p t er 1q int 1p ter 1p ter 1q ter 3q ter 20
22 22 21 18 18 17 9 20 18q ter
23 23 20 X3 15qprox 15qprox 21 8q ter 19
24 25 25 8 3p 3p 16q ter 9q ter 21
25 28 24 22 9q int 7q int 3p ter 15q ter 22
X1 X1 X1 X1 X1 X1 X1 X1 X1
ACA: A. caraya; API : A. pigra; APA: A. palliata; AGU: A. guariba; ASEs: A. seniculus seniculus; ASEa: A.
s. arctoidea; AMA: A. m acconnelli; ASA: A. sara; ABE: A. belzebul.
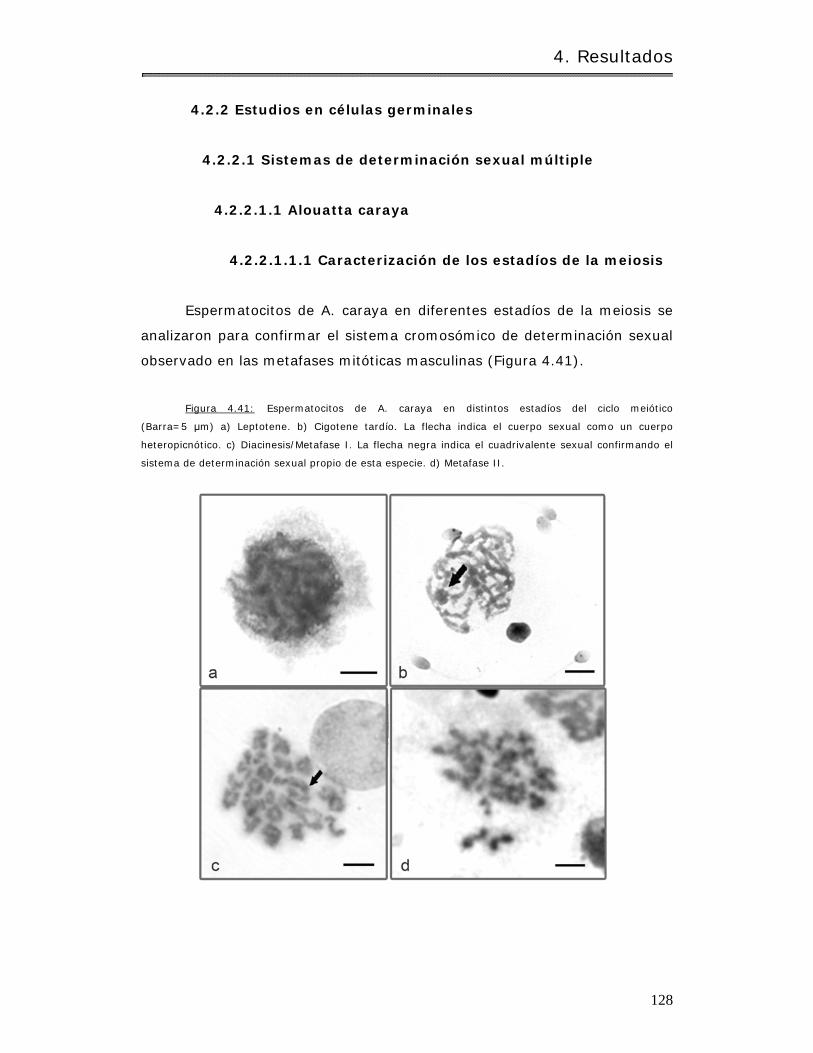
4. Resultados
128
4 .2 .2 Estudios en cé lulas germ ina les
4 .2 .2 .1 Sistem as de determ inación sexu al m últ iple
4 .2 .2 .1 .1 Alouat ta caraya
4 .2 .2 .1 .1 .1 Caracter ización de los estadíos de la m eiosis
Esperm atocitos de A. caraya en diferentes estadíos de la m eiosis se
analizaron para confirm ar el sistem a crom osóm ico de determ inación sexual
observado en las m etafases m itót icas m asculinas (Figura 4.41) .
Figura 4.41: Esperm atocitos de A. caraya en dist intos estadíos del ciclo m eiót ico
(Barra= 5 μm) a) Leptotene. b) Cigotene tardío. La flecha indica el cuerpo sexual como un cuerpo
heteropicnót ico. c) Diacinesis/ Metafase I . La flecha negra indica el cuadrivalente sexual confirmando el
sistem a de determ inación sexual propio de esta especie. d) Metafase I I .

4. Resultados
129
El cuadrivalente sexual en ACA com ienza a ser visible a part ir del
estadío de cigotene, en el que se lo observa como una masa heteropicnót ica
posit iva (Figura 4.41 b)
En Diacinesis/ Metafase I (Figura 4.41c) se observaron 24 bivalentes
autosóm icos y un cuadrivalente, que estaría const ituido por los crom osom as
X1, X2, Y1 e Y2.
En Metafase I I (Figura 4.41d) se observan dos t ipos de células
(am bas con 26 crom osom as) : un t ipo celular con 11 cromosomas
bibraquiados y 15 crom osomas acrocént r icos (conteniendo así a los
crom osom as X1 y X2) y un segundo t ipo celular con 9 crom osom as
bibraquiados y 17 crom osomas acrocént r icos, conteniendo a los
crom osom as Y1 e Y2.
4 .2 .2 .1 .1 .2 El cuadr iva lente sexual en Diacinesis/ M etafase I
Mediante la aplicación de la técnica de bandas C en preparados
m eiót icos (Figura 4.42) se pudo confirm ar inequívocam ente la est ructura
del cuadrivalente sexual en ACA. Se pudieron observar 4 bloques
cent rom éricos (C+ ) correspondientes a una cadena formada por los
crom osom as X1-Y1-X2-Y2 que se disponen en configuración alternada.

4. Resultados
130
Figura 4.42: Sistem a de determ inación sexual m asculino X1X2Y1Y2 (que form a un cuadrivalente
sexual o I V) en A. caraya (Barra= 5 μm) . a) Esperm atocito en m etafase I con bandas C m ost rando la
localización de las regiones heterocromát icas (C+ ) . Las flechas negras indican el cuadr ivalente sexual
X1X2Y1Y2. b) Detalle del cuadr ivalente en el cual se indican con f lechas negras los cent rómeros de los
cuat ro com ponentes crom osóm icos.
4 .2 .2 .1 .2 Alouat ta pigra
4 .2 .2 .1 .2 .1 Caracter ización de los estadíos de la m eiosis
Se analizaron esperm atocitos de A. pigra para confirmar la propuesta
de cuadrivalente sexual X1X2Y1Y2 realizada a part ir de los preparados
m itót icos masculinos (Figura 4.43) .

4. Resultados
131
Figura 4.43: Esperm atocitos de A. pigra en dist intos estadíos del ciclo m eiót ico (Barra= 5 μm ) .
a) Leptotene. b) Cigotene tardío con bivalentes parcialm ente sinapsados. La f lecha negra muest ra el
cuerpo sexual com o un cuerpo heteropicnót ico posit ivo. c) Diacinesis/ Metafase I . La flecha negra indica
el cuadrivalente sexual confirmando el sistem a de determ inación sexual propio de esta especie. d)
Metafase I I .
En Leptotene se observan las m asas crom at ínicas form adas por
autosom as y gonosom as, indist inguibles unas de ot ras. A part ir de Cigotene
se puede observar al cuerpo sexual com o una m asa heteropicnót ica posit iva
(Figura 4.43b) .
En 100% de los esperm atocitos de A. pigra analizados en
Diacinesis/ Metafase I se observaron 27 bivalentes autosóm icos y un
cuadrivalente sexual m asculino X1X2Y1Y2, que se corresponden con el
2N= 58, X1X2Y1Y2 observado en células som át icas m asculinas (Figura 4.43c) .

4. Resultados
132
4 .2 .2 .1 .2 .2 El cuadr iva lente sexual en Diacinesis/ Metafase I
Al aplicarse la técnica de bandas C en células m eiót icas en Metafase
I , se observaron 4 regiones heterocrom át icas cent rom éricas en el
m ult ivalente de A. pigra, correspondientes a una cadena form ada por los
crom osom as X1-Y1-X2-Y2, confirmando la propuesta del cuadrivalente sexual
(Figura 4.44) .
Figura 4.44: Sistem a de determ inación sexual m asculino X1X2Y1Y2 (que form a un cuadrivalente
sexual o I V) en A. pigra. a) Esperm atocito en m etafase I con bandas C m ost rando la localización de las
regiones heterocrom át icas (C+ ) . Las líneas negras indican el cuadrivalente sexual X1X2Y1Y2 superpuesto a
un bivalente autosomal. Los aster iscos indican la presencia de espermatozoides en el m ater ial test icular
(Barra= 5 μm). b) Detalle del cuadrivalente en el cual se indican con líneas negras los cent rómeros de los
cuat ro com ponentes crom osóm icos.

4. Resultados
133
4 .2 .2 .1 .3 Alouat ta pallia ta
4 .2 .2 .1 .3 .1 Caracter ización de los estadíos de la m eiosis
Esperm atocitos de A. palliata en diferentes estadíos de la m eiosis se
analizaron para confirmar el sistem a sexual observado en las metafases
m itót icas masculinas (Figura 4.45) .
Figura 4.45: Esperm atocitos de A. palliata en dist intos estadíos del ciclo m eiót ico
(Barra= 5 μm) a) Leptotene. b) Cigotene tardío. La flecha negra muest ra el cuerpo sexual como un
cuerpo heteropicnót ico posit ivo. c) Diacinesis/ Metafase I . La flecha negra indica el cuadr ivalente sexual
confirm ando el sistem a de determ inación sexual propio de esta especie. d) Metafase I I .
En los esperm atocitos en Diacinesis/ Metafase I (Figura 4.45c) se
observaron 25 bivalentes autosóm icos y un t r ivalente sexual, que estaría
form ado por una cadena conteniendo a los crom osomas X1-Y-X2.

4. Resultados
134
4 .2 .2 .1 .3 .2 El t r iva lente sexu al en Diacinesis/ Metafase I
Al aplicar la técnica de bandas C en células m eiót icas en Metafase I
de A. palliata, se observaron 3 regiones heterocrom át icas cent rom éricas en
el mult ivalente, confirm ando la propuesta de un t r ivalente sexual com o
sistem a sexual múlt iple (Figura 4.46) .
Figura 4.46: Sistem a de determ inación sexual masculino X1X2Y1 en A. palliata. a) Esperm atocito
en m etafase I con bandas C m ost rando la localización de las regiones heterocrom át icas (C+ ) . La f lecha
indica el t r ivalente sexual X1X2Y1 (Barra= 5 μm ). b) Detalle del t r ivalente sexual en el cual se indican con
líneas negras los cent rómeros de los cuat ro componentes crom osóm icos.

4. Resultados
135
4 .2 .2 .2 Sistem as de determ inación sexual XY “ Hum an- like ”
4 .2 .2 .2 .1 Cebus libidinosus
4 .2 .2 .2 .1 .1 Caracter ización de los estadíos de la m eiosis
Esperm atocitos de C. libidinosus en diferentes estadíos de la m eiosis
se analizaron para confirm ar el sistem a sexual observado previam ente en
las m etafases m itót icas m asculinas (Figura 4.47) .
Figura 4.47: Esperm atocitos de C. libidinosus en dist intos estadíos del ciclo m eiót ico
(Barra= 5 μm ) a) Leptotene. b) Cigotene tardío. La f lecha negra m uest ra el cuerpo sexual XY com o un
cuerpo heteropicnót ico posit ivo. c) Diacinesis/ Metafase I . La f lecha negra indica el bivalente XY
confirm ando el sistem a de determ inación sexual propio de esta especie. d) Metafase I I .
El cuerpo XY comenzó a ser v isible en el estadío de Cigotene com o
una m asa heteropicnót ica posit iva en la perifer ia del núcleo (Figura 4.47b) .
En Diacinesis/ Metafase I se observaron 26 bivalentes autosóm icos y
el bivalente sexual XY, most rando una est ructura de t ipo “hum an like” . Los
27 bivalentes se corresponden con el 2N= 54, XY observado en las
m etafases m itót icas m asculinas (Figura 4.47c) .

4. Resultados
136
4 .2 .2 .2 .1 .2 El biva lente sexual en Diacinesis/ Met afase I
En el m aterial obtenido a part ir de las biopsias de C. libidinosus no
fue posible hallar resultados posit ivos mediante la técnica de bandas C en
m eiosis. Si bien no se ha podido caracterizar el bivalente XY m ediante esta
técnica, sí fue posible obtener resultados m ediante la técnica de
inm unofluorescencia (ver sección 4.3.2.1.1) . Cabe com entar que esta
especie ha sido objeto de estudio por análisis m eiót icos en repet idas
ocasiones en el pasado (Seuánez et al. 1983; Mudry et al. 2001) , por lo que la
est ructura del bivalente XY ya ha sido confirm ada en nuest ro equipo de
t rabajo y en la literatura.
4 .2 .2 .2 .2 Saim ir i boliv iensis boliv iensis
4 .2 .2 .2 .2 .1 Caracter ización de los estadíos de la m eiosis
Se analizaron esperm atocitos de S. b. boliviensis en los que se
describió el sistem a de determ inación sexual XY de esta especie (Figura
4.48) .
Figura 4.48: Esperm atocitos de S. boliviensis en dist intos estadíos del ciclo m eiót ico
(Barra= 5 μm ) a) Leptotene. b) Cigotene tardío. La f lecha negra m uest ra el cuerpo sexual XY com o un
cuerpo heteropicnót ico posit ivo. c) Diacinesis/ Metafase I . La f lecha negra indica el bivalente XY
confirm ando el sistem a de determ inación sexual propio de esta especie. d) Metafase I I .

4. Resultados
137
En Leptotene (Figura 4.48a) , el bivalente XY es indist inguible de los
autosom as. Hacia el estadío de Cigotene (Figura 4.48b) , el cuerpo XY
com ienza a ser visible com o una m asa heteropicnót ica posit iva en la
perifer ia del núcleo. Se observa al cuerpo XY com o una est ructura bipart ita,
form ada por dos elem entos que se tocan parcialmente.
En Diacinesis/ Metafase I se observaron 21 bivalentes autosom ales y
el bivalente XY, confirm ando así el sistem a de determ inación sexual
observado previam ente en los análisis m itót icos m asculinos (Figura 4.48c) .
4 .2 .2 .2 .2 .2 El biva lente sexual en Diacine sis/ Metafase I
Mediante la técnica de bandas C se observó la localización de las
regiones heterocrom át icas en células germ inales (Figura 4.49) , y se
confirm ó la presencia de sistem a sexual de t ipo XY.
Figura 4.49: Sistem a de determ inación sexual XY en S. b. boliv iensis. a) Esperm atocito en
Diacinesis/ Metafase I con bandas C m ost rando la localización de las regiones heterocrom át icas (C+ ) . La
flecha indica el bivalente XY (Barra= 5 μm). b) Detalle del bivalente XY en el que se indican con flechas
los cent róm eros de cada com ponente del bivalente (Barra= 2 μm ).

4. Resultados
138
4 .3 Diagnosis por caract er ización citom olecular
4 .3 .1 Estudios en células som át icas
4 .3 .1 .1 Secuencias te lom éricas en Ceboidea
Con el objet ivo de caracter izar la dist r ibución de la secuencia
telom érica (TTAGGG) n en los pr imates ceboideos analizados se aplicó la
técnica de FI SH con una sonda pantelomérica. Este análisis perm it ió
determ inar si las regiones involucradas en los reordenam ientos que dieron
origen a los sistem as sexuales m últ iples en aulladores, conservan
secuencias telom éricas interst iciales.
En las especies estudiadas se observaron señales de hibridación
posit ivas en las regiones term inales de los crom osom as, ident ificadas com o
regiones telom éricas (Figura 4.50 a- f) . Cada crom osom a presentó 4
señales, confirm ando que todos los crom osom as del complem ento son
bibraquiados si bien con diferente m orfología, crom osom as
subm etacént r icos, metacént r icos y acrocént r icos, y diferente tam año. En
concordancia con esto, la intensidad de las señales telom éricas observadas
en las dist intas especies most ró una am plia variabilidad, lo que sugeriría
una longitud de secuencias variable en los dist intos taxa.
Con excepción de Aotus azarae, especie en la que se observó una
señal telom érica interst icial en la región pericent rom érica del crom osom a 5,
en los ceboideos analizados no se observaron ot ras señales interst iciales
(Figura 4.50 e y f) .
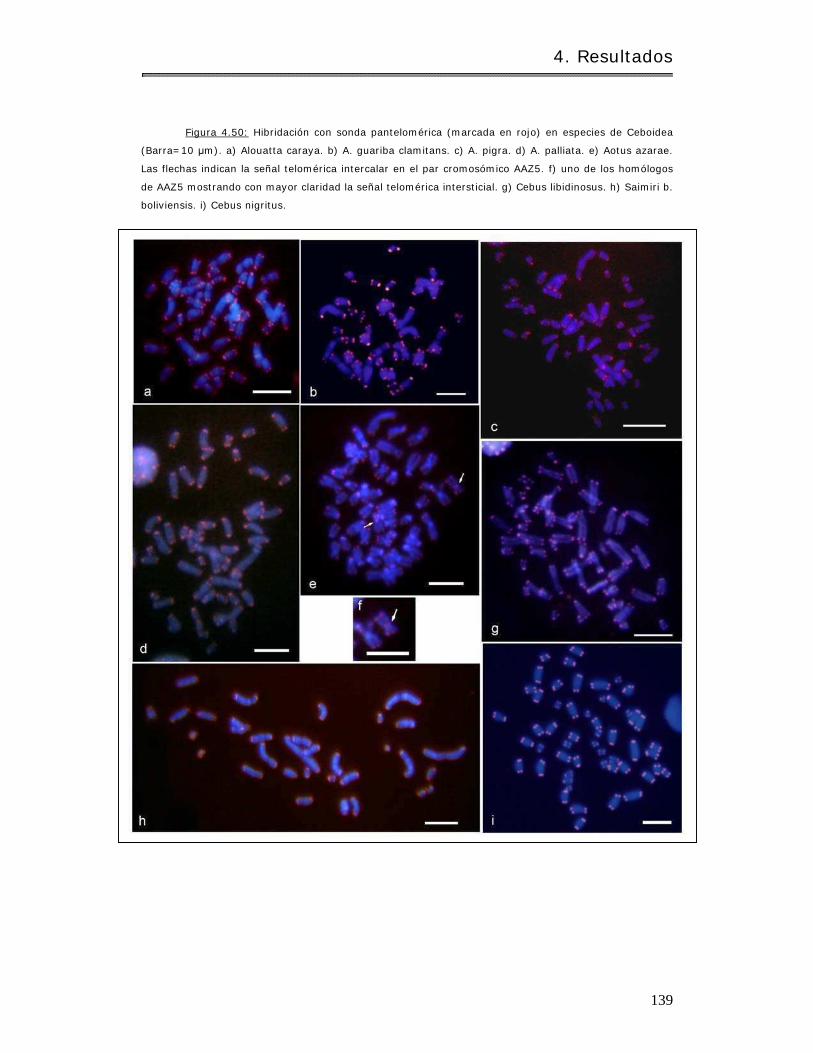
4. Resultados
139
Figura 4.50: Hibridación con sonda pantelom érica (m arcada en rojo) en especies de Ceboidea
(Barra= 10 μm ). a) Alouat ta caraya. b) A. guar iba clam itans. c) A. pigra. d) A. palliata. e) Aotus azarae.
Las flechas indican la señal telom érica intercalar en el par cromosóm ico AAZ5. f) uno de los homólogos
de AAZ5 m ost rando con m ayor claridad la señal telom érica interst icial. g) Cebus libidinosus. h) Saim ir i b.
boliv iensis. i) Cebus nigr itus.
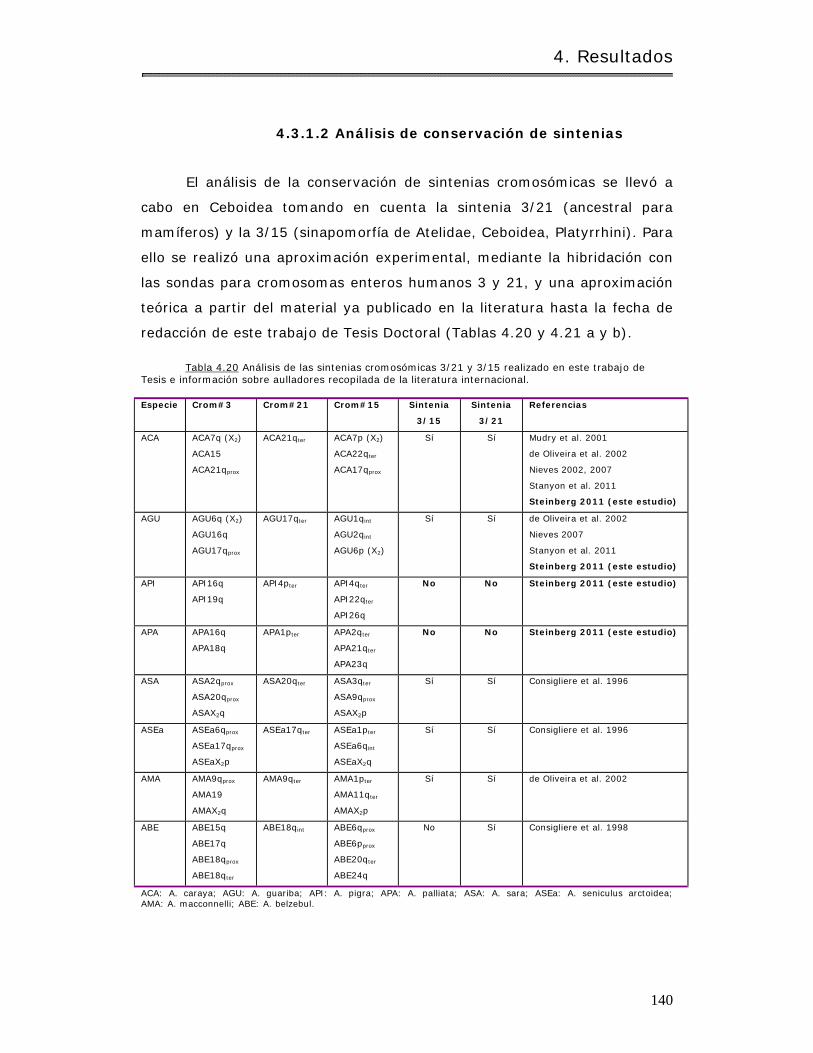
4. Resultados
140
4 .3 .1 .2 Análisis de conservación de sin tenias
El análisis de la conservación de sintenias cromosóm icas se llevó a
cabo en Ceboidea tom ando en cuenta la sintenia 3/ 21 (ancest ral para
m am íferos) y la 3/ 15 (sinapom orfía de Atelidae, Ceboidea, Platyrrhini) . Para
ello se realizó una aproxim ación experim ental, mediante la hibridación con
las sondas para cromosom as enteros hum anos 3 y 21, y una aproxim ación
teórica a part ir del m aterial ya publicado en la literatura hasta la fecha de
redacción de este t rabajo de Tesis Doctoral (Tablas 4.20 y 4.21 a y b) .
Tabla 4.20 Análisis de las sintenias crom osóm icas 3/ 21 y 3/ 15 realizado en este t rabajo de
Tesis e inform ación sobre aulladores recopilada de la literatura internacional. Especie Crom # 3 Crom # 2 1 Crom # 1 5 Sintenia
3 / 1 5
Sintenia
3 / 2 1
Referencias
ACA ACA7q (X2)
ACA15
ACA21qprox
ACA21q ter ACA7p (X2)
ACA22q ter
ACA17qprox
Sí Sí Mudry et al. 2001
de Oliveira et al. 2002
Nieves 2002, 2007
Stanyon et al. 2011
Ste inberg 2 0 1 1 ( este estudio)
AGU AGU6q (X2)
AGU16q
AGU17qprox
AGU17q ter AGU1q int
AGU2q int
AGU6p (X2)
Sí Sí de Oliveira et al. 2002
Nieves 2007
Stanyon et al. 2011
Ste inberg 2 0 1 1 ( este estudio)
API API 16q
API 19q
API 4p ter API 4q ter
API 22q ter
API 26q
No No Ste inberg 2 0 1 1 ( este estudio)
APA APA16q
APA18q
APA1p ter APA2q ter
APA21q t er
APA23q
No No Ste inberg 2 0 1 1 ( este estudio)
ASA ASA2qprox
ASA20qprox
ASAX2q
ASA20q ter ASA3q ter
ASA9qprox
ASAX2p
Sí Sí Consigliere et al. 1996
ASEa ASEa6qprox
ASEa17qprox
ASEaX2p
ASEa17q ter ASEa1p t er
ASEa6q int
ASEaX2q
Sí Sí Consigliere et al. 1996
AMA AMA9qprox
AMA19
AMAX2q
AMA9q ter AMA1p ter
AMA11q ter
AMAX2p
Sí Sí de Oliveira et al. 2002
ABE ABE15q
ABE17q
ABE18qprox
ABE18q ter
ABE18q int
ABE6qprox
ABE6pprox
ABE20q ter
ABE24q
No Sí Consigliere et al. 1998
ACA: A. caraya; AGU: A. guariba; API : A. pigra; APA: A. palliata; ASA: A. sara; ASEa: A. seniculus arctoidea; AMA: A. macconnelli; ABE: A. belzebul.
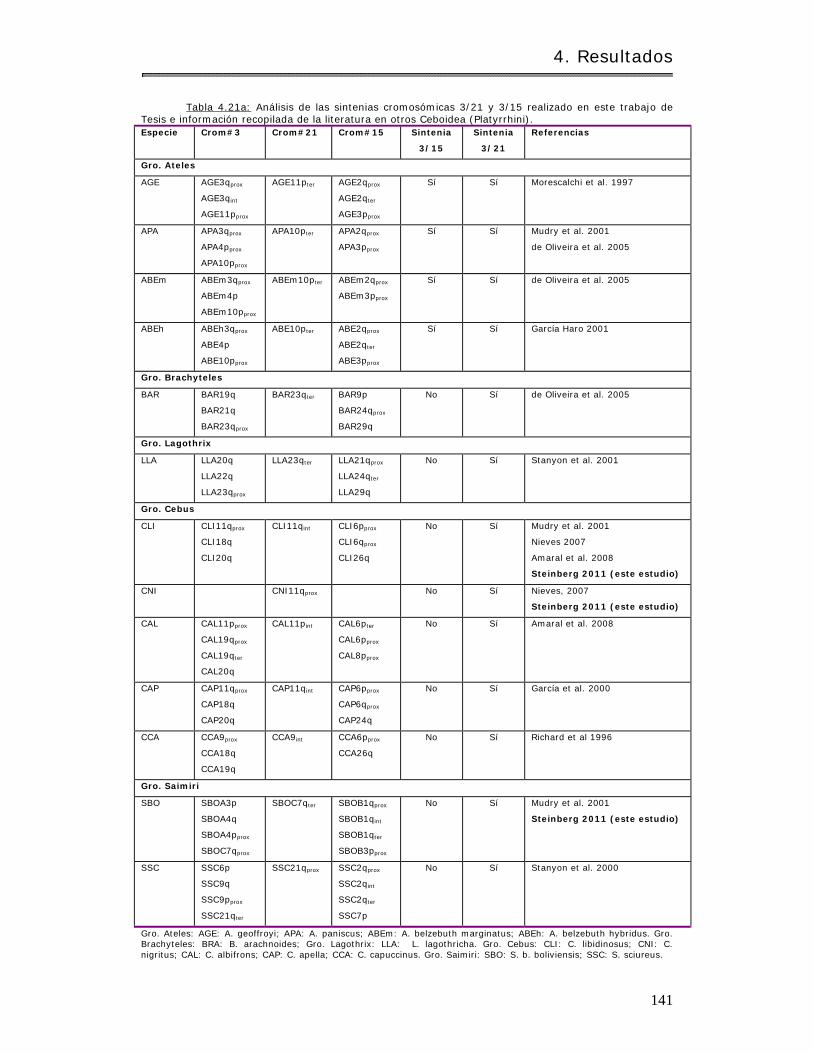
4. Resultados
141
Tabla 4.21a: Análisis de las sintenias cromosóm icas 3/ 21 y 3/ 15 realizado en este t rabajo de Tesis e inform ación recopilada de la literatura en ot ros Ceboidea (Platyrrhini) . Especie Crom # 3 Crom # 2 1 Crom # 1 5 Sintenia
3 / 1 5
Sintenia
3 / 2 1
Referencias
Gro. Ate les
AGE AGE3qprox
AGE3q int
AGE11pprox
AGE11p t er AGE2qprox
AGE2q t er
AGE3pprox
Sí Sí Morescalchi et al. 1997
APA APA3qprox
APA4pprox
APA10pprox
APA10p t er APA2qprox
APA3pprox
Sí Sí Mudry et al. 2001
de Oliveira et al. 2005
ABEm ABEm 3qprox
ABEm 4p
ABEm 10pprox
ABEm 10p ter ABEm 2qprox
ABEm 3pprox
Sí Sí de Oliveira et al. 2005
ABEh ABEh3qprox
ABE4p
ABE10pprox
ABE10p ter ABE2qprox
ABE2q ter
ABE3pprox
Sí Sí García Haro 2001
Gro. Brachyteles
BAR BAR19q
BAR21q
BAR23qprox
BAR23q ter BAR9p
BAR24qprox
BAR29q
No Sí de Oliveira et al. 2005
Gro. Lagothr ix
LLA LLA20q
LLA22q
LLA23qprox
LLA23q t er LLA21qprox
LLA24q ter
LLA29q
No Sí Stanyon et al. 2001
Gro. Cebus
CLI CLI 11qprox
CLI 18q
CLI 20q
CLI 11q int CLI 6pprox
CLI 6qprox
CLI 26q
No Sí Mudry et al. 2001
Nieves 2007
Amaral et al. 2008
Ste inberg 2 0 1 1 ( este estudio)
CNI CNI 11qprox No Sí Nieves, 2007
Ste inberg 2 0 1 1 ( este estudio)
CAL CAL11pprox
CAL19qprox
CAL19q t er
CAL20q
CAL11p int CAL6p ter
CAL6pprox
CAL8pprox
No Sí Amaral et al. 2008
CAP CAP11qprox
CAP18q
CAP20q
CAP11q int CAP6pprox
CAP6qprox
CAP24q
No Sí García et al. 2000
CCA CCA9prox
CCA18q
CCA19q
CCA9 int CCA6pprox
CCA26q
No Sí Richard et al 1996
Gro. Saim ir i
SBO SBOA3p
SBOA4q
SBOA4pprox
SBOC7qprox
SBOC7q ter SBOB1qprox
SBOB1q int
SBOB1q ter
SBOB3pprox
No Sí Mudry et al. 2001
Ste inberg 2 0 1 1 ( este estudio)
SSC SSC6p
SSC9q
SSC9pprox
SSC21q ter
SSC21qprox SSC2qprox
SSC2q int
SSC2q ter
SSC7p
No Sí Stanyon et al. 2000
Gro. Ateles: AGE: A. geoffroyi; APA: A. paniscus; ABEm: A. belzebuth marginatus; ABEh: A. belzebuth hybr idus. Gro. Brachyteles: BRA: B. arachnoides; Gro. Lagothrix: LLA: L. lagothricha. Gro. Cebus: CLI : C. libidinosus; CNI : C. nigr itus; CAL: C. albifrons; CAP: C. apella; CCA: C. capuccinus. Gro. Saim ir i: SBO: S. b. boliviensis; SSC: S. sciureus.
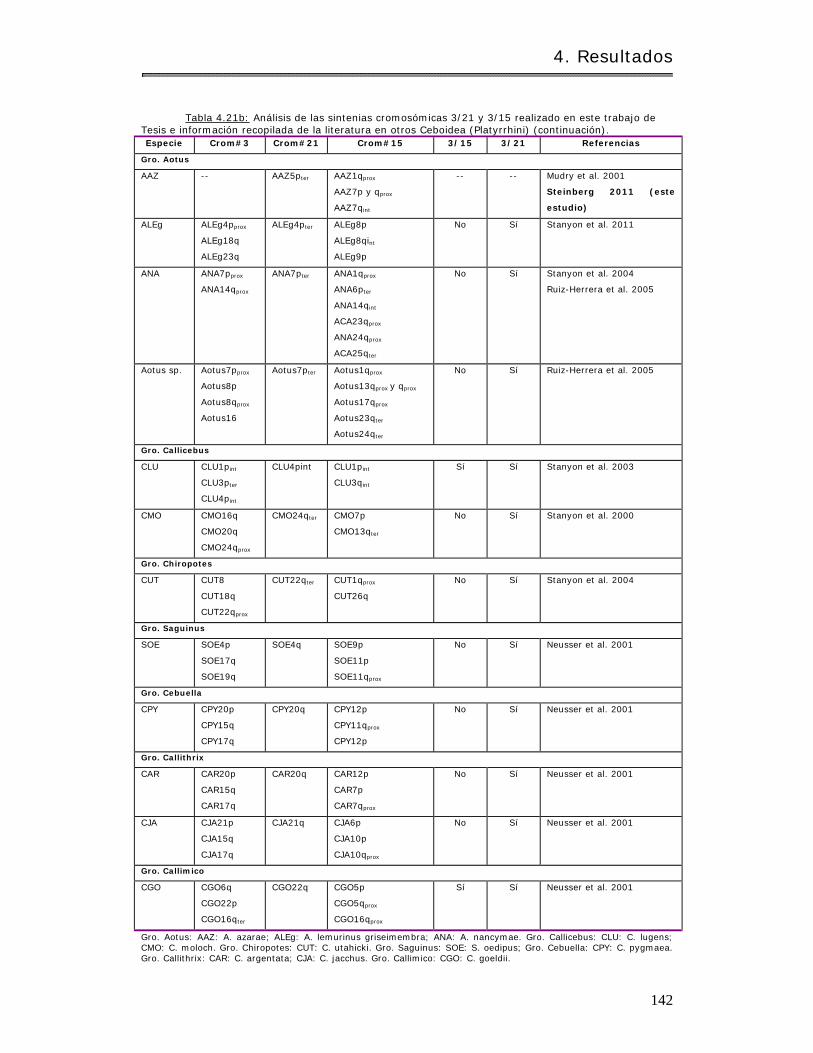
4. Resultados
142
Tabla 4.21b: Análisis de las sintenias crom osóm icas 3/ 21 y 3/ 15 realizado en este t rabajo de
Tesis e inform ación recopilada de la literatura en ot ros Ceboidea (Platyrrhini) (cont inuación) . Especie Crom # 3 Crom # 2 1 Crom # 1 5 3 / 1 5 3 / 2 1 Referencias
Gro. Aotus
AAZ - - AAZ5p ter AAZ1qprox
AAZ7p y qprox
AAZ7q int
- - - - Mudry et al. 2001
Ste inberg 2 0 1 1 ( este
estudio)
ALEg ALEg4pprox
ALEg18q
ALEg23q
ALEg4p ter ALEg8p
ALEg8qint
ALEg9p
No Sí Stanyon et al. 2011
ANA ANA7pprox
ANA14qprox
ANA7p ter ANA1qprox
ANA6p ter
ANA14q int
ACA23qprox
ANA24qprox
ACA25q ter
No Sí Stanyon et al. 2004
Ruiz-Herrera et al. 2005
Aotus sp. Aotus7pprox
Aotus8p
Aotus8qprox
Aotus16
Aotus7p ter Aotus1qprox
Aotus13qprox y qprox
Aotus17qprox
Aotus23q t er
Aotus24q t er
No Sí Ruiz-Herrera et al. 2005
Gro. Ca llicebus
CLU CLU1p int
CLU3pt er
CLU4p int
CLU4pint CLU1p int
CLU3q int
Sí Sí Stanyon et al. 2003
CMO CMO16q
CMO20q
CMO24qprox
CMO24q ter CMO7p
CMO13q ter
No Sí Stanyon et al. 2000
Gro. Ch iropotes
CUT CUT8
CUT18q
CUT22qprox
CUT22q ter CUT1qprox
CUT26q
No Sí Stanyon et al. 2004
Gro. Saguinus
SOE SOE4p
SOE17q
SOE19q
SOE4q SOE9p
SOE11p
SOE11qprox
No Sí Neusser et al. 2001
Gro. Cebuella
CPY CPY20p
CPY15q
CPY17q
CPY20q CPY12p
CPY11qprox
CPY12p
No Sí Neusser et al. 2001
Gro. Ca llith r ix
CAR CAR20p
CAR15q
CAR17q
CAR20q CAR12p
CAR7p
CAR7qprox
No Sí Neusser et al. 2001
CJA CJA21p
CJA15q
CJA17q
CJA21q CJA6p
CJA10p
CJA10qprox
No Sí Neusser et al. 2001
Gro. Ca llim ico
CGO CGO6q
CGO22p
CGO16q ter
CGO22q CGO5p
CGO5qprox
CGO16qprox
Sí Sí Neusser et al. 2001
Gro. Aotus: AAZ: A. azarae; ALEg: A. lem urinus gr iseim em bra; ANA: A. nancym ae. Gro. Callicebus: CLU: C. lugens; CMO: C. m oloch. Gro. Chiropotes: CUT: C. utahicki. Gro. Saguinus: SOE: S. oedipus; Gro. Cebuella: CPY: C. pygmaea. Gro. Callithr ix: CAR: C. argentata; CJA: C. jacchus. Gro. Callim ico: CGO: C. goeldii.
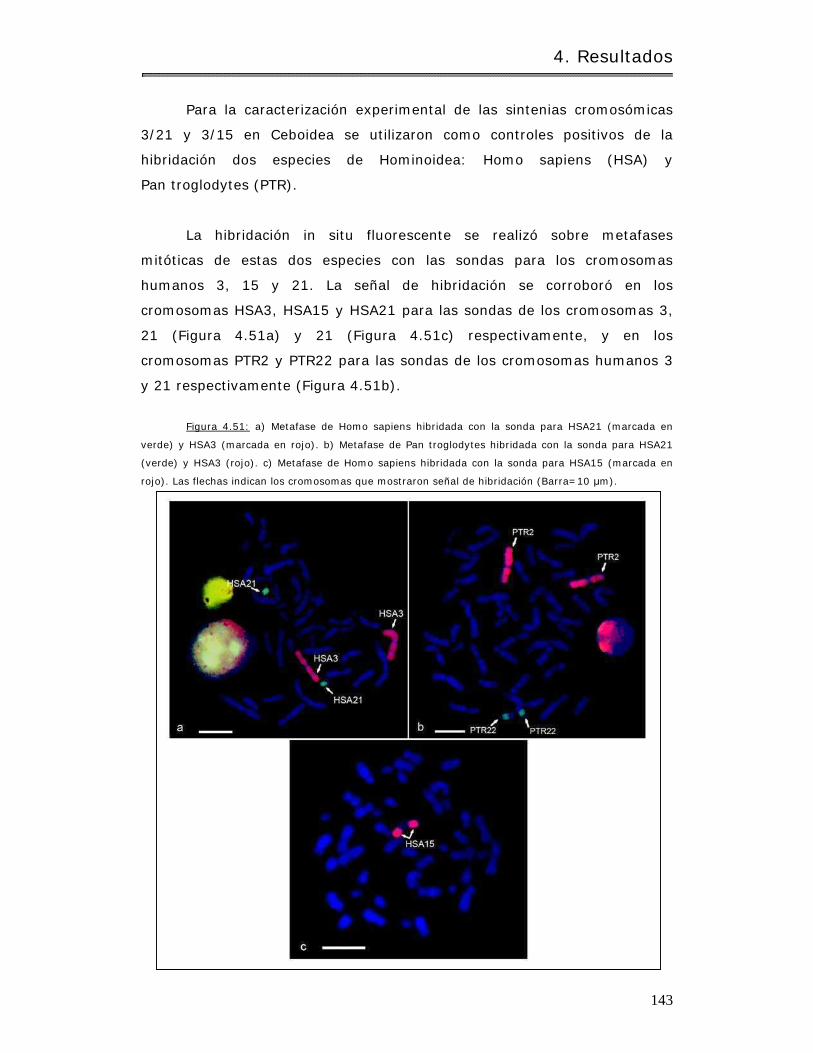
4. Resultados
143
Para la caracterización experim ental de las sintenias cromosóm icas
3/ 21 y 3/ 15 en Ceboidea se ut ilizaron com o cont roles posit ivos de la
hibridación dos especies de Hominoidea: Hom o sapiens (HSA) y
Pan t roglodytes (PTR) .
La hibr idación in situ fluorescente se realizó sobre m etafases
m itót icas de estas dos especies con las sondas para los crom osom as
hum anos 3, 15 y 21. La señal de hibridación se corroboró en los
crom osom as HSA3, HSA15 y HSA21 para las sondas de los crom osom as 3,
21 (Figura 4.51a) y 21 (Figura 4.51c) respect ivam ente, y en los
crom osom as PTR2 y PTR22 para las sondas de los crom osom as hum anos 3
y 21 respect ivam ente (Figura 4.51b) .
Figura 4.51: a) Metafase de Homo sapiens hibr idada con la sonda para HSA21 (marcada en
verde) y HSA3 (m arcada en rojo) . b) Metafase de Pan t roglodytes hibr idada con la sonda para HSA21
(verde) y HSA3 ( rojo) . c) Metafase de Hom o sapiens hibr idada con la sonda para HSA15 (m arcada en
rojo) . Las flechas indican los crom osom as que m ost raron señal de hibr idación (Barra= 10 μm ).

4. Resultados
144
En el género Alouat ta se realizó la hibridación in situ fluorescente con
las sondas 3, 15 y 21 hum anas en las especies A. caraya, A. guariba
clam itans, A. pigra y A. palliata (Figura 4.52 y 4.53) .
Alouat ta caraya mostró hibridación para HSA21 en el par hom eólogo
ACA21. La m arca se ubica en la región term inal del ACA21q. En ACA21, en
la región proxim al ACA21q, se observó la señal para HSA3, confirm ando la
presencia de la sintenia 3/ 21 en ACA (Figura 4.53a) .
La sonda para HSA15 hibr idó en ACA7p (X2) . En ACA7q se observa
señal de hibridación para HSA3, confirm ando la presencia de la sintenia
3/ 15 en esta especie, asociada al sistem a sexual múlt iple X1X2Y1Y2 (Figura
4.53a) .
En ACA22q ter y en ACA17qprox se observaron señales de hibr idación
para HSA15. En ACA15q se observó la señal para HSA3 (Figura 4.53a) .
En Alouat ta guariba clam itans se observó la conservación de la
sintenia 3/ 21 en AGU18q (Figura 4.52b) . La señal para HSA21 se observa
en AGU18q ter y la señal para HSA3 en 18qprox (Figuras 4.52a y b y 4.53b) .
La sintenia 3/ 15 tam bién se encuent ra conservada en esta especie y
asociada al sistem a sexual múlt iple X1X2X3Y1Y2. En AGU6q (X2) se observó
señal para HSA3 y en AGU6p e Y1, la señal para HSA15. La sonda para
HSA15 hibr idó a su vez en AGU1q int y AGU2q int, m ient ras que la sonda para
HSA3 se detectó tam bién en AGU16q e Y2 (Figuras 4.52a y 4.53b) .
En Alouat ta pigra se observó señal para HSA3 en API 16q y API 19q,
m ient ras que la sonda para HSA21, hibridó en API 4p ter, indicando que la
sintenia 3/ 21, ancest ral para m am íferos, no estaría conservada en esta
especie (Figuras 4.52c y Figura 4.53c) .
La sonda para HSA15 hibr idó en las metafases de API en los
crom osom as API 4q ter, API 22q ter y API 26q. De estas observaciones se infiere
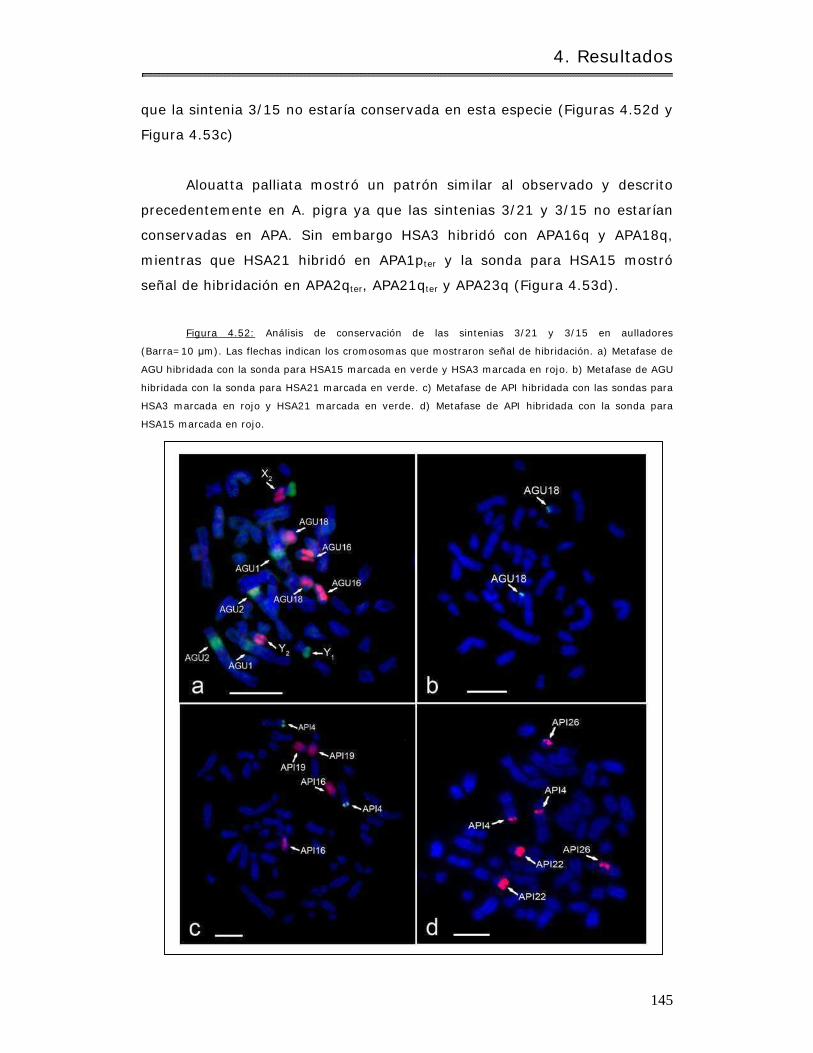
4. Resultados
145
que la sintenia 3/ 15 no estaría conservada en esta especie (Figuras 4.52d y
Figura 4.53c)
Alouat ta palliata m ostró un pat rón sim ilar al observado y descrito
precedentem ente en A. pigra ya que las sintenias 3/ 21 y 3/ 15 no estarían
conservadas en APA. Sin em bargo HSA3 hibridó con APA16q y APA18q,
m ient ras que HSA21 hibr idó en APA1p t er y la sonda para HSA15 m ost ró
señal de hibr idación en APA2q ter, APA21q ter y APA23q (Figura 4.53d) .
Figura 4.52: Análisis de conservación de las sintenias 3/ 21 y 3/ 15 en aulladores
(Barra= 10 μm ). Las flechas indican los crom osom as que most raron señal de hibridación. a) Metafase de
AGU hibr idada con la sonda para HSA15 m arcada en verde y HSA3 m arcada en rojo. b) Metafase de AGU
hibr idada con la sonda para HSA21 m arcada en verde. c) Metafase de API hibr idada con las sondas para
HSA3 m arcada en rojo y HSA21 m arcada en verde. d) Metafase de API hibridada con la sonda para
HSA15 m arcada en rojo.

4. Resultados
146
Figura 4.53: Crom osom as de cada especie de aullador ident if icados por pares al pie de cada
cromosom a con bandas G y en los que se indica, sobre la derecha, el pat rón de hibridación de los
crom osom as HSA3, HSA21 y HSA15. a) A. caraya. b) A. guar iba clam itans. c) A. pigra. d) A. palliata.
La conservación de la sintenia 3/ 21 y la ausencia de conservación de
la sintenia 3/ 15 se observó en los ot ros Ceboidea estudiados
(Cebus libidinosus, C. nigr itus, Saim ir i b. boliviensis) (Figura 4.54a, b y c) .
En ambas especies de Cebus se observó idént ico pat rón de
hibridación ya que la sonda para HSA3 hibridó en los crom osom as 11qprox,
19q y 20q; la sonda para HSA21 most ró señal de hibr idación en 11q int y la
sonda para HSA15 hibr idó con los cromosom as CLI 6 y CNI 6 (en la porción
proxim al de los brazos p y q) así como en CLI 26q y CNI 26q (Figura 4.54a) .

4. Resultados
147
Figura 4.54: Crom osom as de ot ros Ceboidea ident if icados con bandas G y en los que se
indica, sobre la derecha, el pat rón de hibr idación de los crom osom as HSA3, HSA21 y HSA15. a) Cebus
libidinosus (el recuadro de arr iba y a la derecha se m uesta el pat rón de hibr idación de CNI 11) . b) Saim ir i
b. boliviensis. c) Aotus azarae.
En Saim ir i b. boliviensis se observó hibr idación para HSA3 en los
crom osom as SBOA3p y en la porción proxim al de am bos brazos de SBOA4
y en SBOC7qprox. La señal de hibr idación para HSA21 se observó en
SBOC7q ter. La sonda para HSA15 hibridó en SBOB1q y en la región proxim al
de SBOB3p (Figura 4.54b) .
En el caso de Aotus azarae no se logró obtener señal de hibridación
con la sonda de HSA3. Dado que en esta especie, hasta la fecha de este
t rabajo de Tesis y en lo que hace a nuest ro conocim iento, no fue analizada
la conservación de las sintenias 3/ 15 y 3/ 21 nada se puede afirm ar al
respecto. Por el cont rar io, sí se obtuvo señal para la sonda de HSA21 y se
observó su hibr idación en AAZ5p ter (Figura 4.54c) . La sonda para HSA15
hibr ida en AAZ1qprox, en AAZ7p, y en AAZ7qprox y AAZ7q ter (Figura 4.54c) .

4. Resultados
148
4 .3 .1 .3 Análisis de conservación del gen SRY
4 .3 .1 .3 .1 Géneros de Hom inoidea
El análisis de la conservación del gen SRY se realizó con la sonda
para el locus SRY hum ano m ediante hibr idación in situ fluorescente. Se
ut ilizó como cont rol posit ivo de la técnica la sonda del crom osom a 21 entero
(HSA21) y dos especies de Hom inoidea, Pan t roglodytes y Hom o sapiens, se
ut ilizaron como cont roles posit ivos de la hibridación. En am bas especies se
obtuvo hibr idación posit iva para la sonda HSA21 y HSASRY. La señal de
HSA21 se observó en el crom osom a hum ano del m ism o nombre y en PTR22
(Figura 4.55) .
La sonda de HSASRY se observó conservada en el crom osom a Y
hum ano así com o en Pan t roglodytes correspondiendo con PTRY.
Figura 4.55: Hibrididación con las sondas para HSA21 (m arcada en verde) y HSASRY (marcada
en rojo) en Hom inoidea (Barra= 10 μm ). Las flechas indican los crom osom as que m ost raron señal de
hibr idación. a) Metafase de Hom o sapiens hibr idada con la sonda para HSA21 y HSASRY. b) Metafase de
Pan t roglodytes hibr idada con la sonda para HSA21 y HSASRY.

4. Resultados
149
4 .3 .1 .3 .2 Género Alouat ta
Este es el pr imer estudio de la conservación del gen SRY por
hibridación in situ en algunas de las especies de pr im ates con sistem as
sexuales múlt iples. El género Alouat ta se tom a com o ejem plo en part icular,
por ser el Ceboidea que presenta la m ayor variedad de sistem as de
determ inación sexual, m ayor al observado en Aotus o en Cacajao.
En este t rabajo de Tesis Doctoral, la hibridación con las sondas
HSA21 y HSASRY se realizó en las especies de aulladores A. caraya,
A. guariba clam itans, A. pigra y A. palliata (Figura 4.56) .
En ninguna de las especies de aulladores analizadas se observó señal
de hibr idación posit iva para el gen HSASRY. Esta ausencia se vio confirm ada
al observar señal de hibridación posit iva para la sonda cont rol HSA21,
situación que perm it ió descartar artefactos técnicos at r ibuibles a la no
visualización de la hibr idación para HSASRY. Com o cont rol posit ivo de la
hibridación se ut ilizó las sondas HSA21 y HSASRY en m etafases de Hom o
sapiens.
En A. caraya se observó señal posit iva de hibridación para el HSA21
en ACA21q ter (Figura 4.56a) , m ient ras que en A. guariba clam itans, la señal
se observó en AGU18q ter (Figura 4.56b) . Esta es la segunda oportunidad en
que esta sonda perm ite confirm ar la hom eología ACA/ AGU que en este
t rabajo de Tesis fuera descr ipta en 3.2.1.6.
En el caso de las especies mesoam ericanas, es la prim era
oportunidad en que se confirm a por hibr idación la conservación de HSA21
en API y APA al observar la señal en API 4pter y APA1p ter (Figura 4.56c y d)
que en este t rabajo de Tesis se describe en 4.3.2.1,
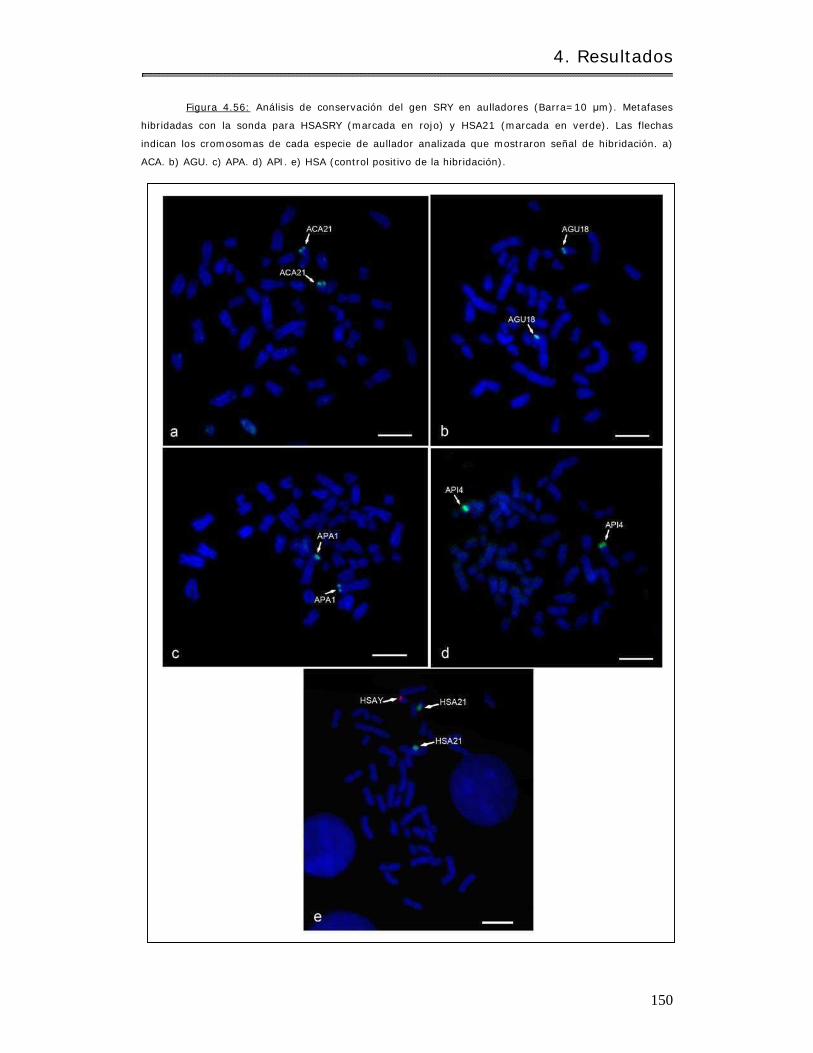
4. Resultados
150
Figura 4.56: Análisis de conservación del gen SRY en aulladores (Barra= 10 μm ). Metafases
hibr idadas con la sonda para HSASRY (m arcada en rojo) y HSA21 (m arcada en verde) . Las flechas
indican los crom osomas de cada especie de aullador analizada que most raron señal de hibr idación. a)
ACA. b) AGU. c) APA. d) API . e) HSA (cont rol posit ivo de la hibr idación) .

4. Resultados
151
4 .3 .1 .3 .1 Ot ros géneros de Ceboidea
En los ot ros Ceboidea estudiados (Cebus libidinosus, C. nigr itus y
Aotus azarae) se observó señal posit iva de hibridación para la sonda cont rol
(HSA21) y no se obtuvo señal para la sonda del SRY (Figura 4.57) . En este
caso tam bién se ut ilizaron como cont rol posit ivo de la hibridación, las
m etafases de Hom o sapiens.
En am bas especies de Cebus (Figura 4.57a y b) se observó señal
posit iva de hibridación en la región proxim al del brazo q del crom osom a 11,
en todas las m etafases analizadas y en ejem plares de am bos sexos
(CLI 11qprox y CNI 11qprox) .
En Aotus azarae la sonda HSA21 hibr idó en la región term inal del
brazo p del crom osom a 5 (AAZ5p ter) en todos los ejem plares adultos de
am bos sexos que fueron objeto de estudio en este t rabajo de Tesis Doctoral
(Figura 4.57c) .
En Saim ir i b. boliv iensis (Figura 4.57d) se observó señal de
hibridación en la región term inal del brazo q del cromosom a C7 (SBOC7q ter) .
Cabe aclarar que si bien el ejem plar de SBO analizado no most ró una señal
de hibr idación nít ida como en los ejem plares de las ot ras especies, esta
hibridación perm it ió verificar la conservación de HSA21 en SBO.
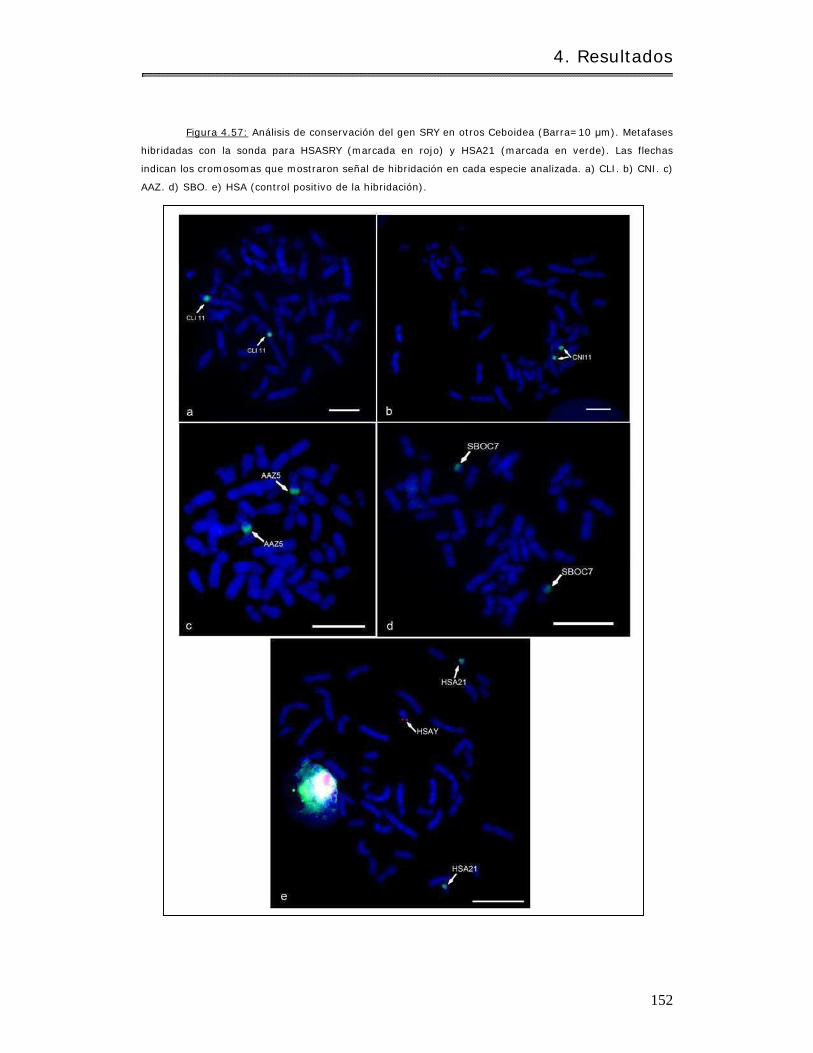
4. Resultados
152
Figura 4.57: Análisis de conservación del gen SRY en ot ros Ceboidea (Barra= 10 μm). Metafases
hibr idadas con la sonda para HSASRY (m arcada en rojo) y HSA21 (m arcada en verde) . Las flechas
indican los crom osomas que most raron señal de hibr idación en cada especie analizada. a) CLI . b) CNI . c)
AAZ. d) SBO. e) HSA (cont rol posit ivo de la hibridación) .

4. Resultados
153
4 .3 .2 Estudios en cé lu las germ inales
4 .3 .2 .1 Caracter izac ión del pat rón de recom binación
4 .3 .2 .1 .1 Cebus libidinosus
4 .3 .2 .1 .1 .1 Apaream iento crom osóm ico y sinapsis
El desarrollo y puesta a punto de estas técnicas se inició con la tom a
de muest ras, m antenim iento en frío en hielo seco para el t raslado a los
laborator ios del GI BE y conservación a –80ºC hasta inicio de su
procesam iento según técnica explicitada en Mater ial y Métodos 3.3.2.1.
Como parte de un t rabajo colaborat ivo con la Unitat de Biologia Cellular i
Genèt ica Mèdica (Facultat de Medicina de la Universidad Autónom a de
Barcelona) las preparaciones m eiót icas se analizaron luego de ser
hibridadas con los ant icuerpos para REC8 (cohesina com ponente del
elem ento lateral del com plejo sinaptoném ico) y SYCP1 (componente de los
elem entos t ransversales del com plejo sinaptoném ico) , a fin de caracter izar
el proceso de apaream iento y sinapsis en C. libidinosus.
Del estudio de inm unofluorescencia realizado, que pusieron en
evidencia las figuras correpondientes a las dist intas etapas m eiót icas
observables en las biopsias test iculares de m achos de CLI , surgen los
resultados descritos a cont inuación.
Las señales ( refer idas com o “core” en inglés) correspondientes a la
inm unolocalización de la proteína REC8 fueron visibles a part ir del estadío
pre-Leptotene.
En el estadío de Leptotene se observó la marcación a modo de finas
hebras, correspondientes a las cohesinas y los elem entos axiales.
En el estadío de Cigotene, apareció por prim era vez la m arca de
SYCP1 en las áreas de sinapsis.

4. Resultados
154
El inicio del apaream iento de los autosom as ocurr ió a nivel
subtelom érico.
Para la visualización del apaream iento de los elem entos axiales y el
análisis de las regiones de sinapsis tardía, se realizaron ensayos de
inm unofluorescencia con ant icuerpos cont ra BRCA1 (proteína reclutada en
la m aquinaria celular ante la presencia de crom osom as no apareados) y
REC8, respect ivam ente.
En la Profase I m eiót ica se analizó un total de 200 núcleos, más
precisamente en el estadío de Paquitene. La mayor proporción de los
núcleos most raron los bivalentes autosóm icos completam ente apareados.
En 12 núcleos los bivalentes autosóm icos se observaron com pletam ente
apareados, con excepción de un bivalente cuya porción term inal se observó
desapareada (Figura 4.58a) . Los elem entos axiales no apareados,
m ost raron señal posit iva para el ant icuerpo BRCA1 (Figura 4.58b) .
La hibr idación in situ f luorescente con la sonda para HSA21 sobre
estos núcleos puso en evidencia que la región no apareada correspondía al
bivalente CLI 11 (Figura 4.58c) . Este es el cromosom a que en CLI , presenta
un bloque heterocrom át ico que ocupa el 75% del brazo q (ver sección
4.2.1.2.2.3.2 página 85. Por arr iba de dicha región es donde se detecta la
hom eología con HSA21, es decir en CLI 11qprox ( ver sección 4.3.1.2 página
147) , región eucrom át ica adyacente a la heterocrom at ina term inal.
En num erosas imágenes de las que se eligieron algunas para ilust rar
el t rabajo realizado, se pudo constatar que los cromosom as que
perm anecían asinapsados se correspondían con CLI 11 (señal posit iva para
HSA21) y CLI X (señal posit iva para HSAX) . En este contexto, se pensó que
el crom osom a asinapsado restante debería corresponder a CLI Y. Sin
embargo, con la m etodología aplicada no se pudo constatar esto últ im o por
FI SH, debido a la falta de hom eología ent re HSAY y el crom osoma Y de los
Ceboidea (Platyrrhini) ( ver I nt roducción, sección 1.2.3.1, página 22) .
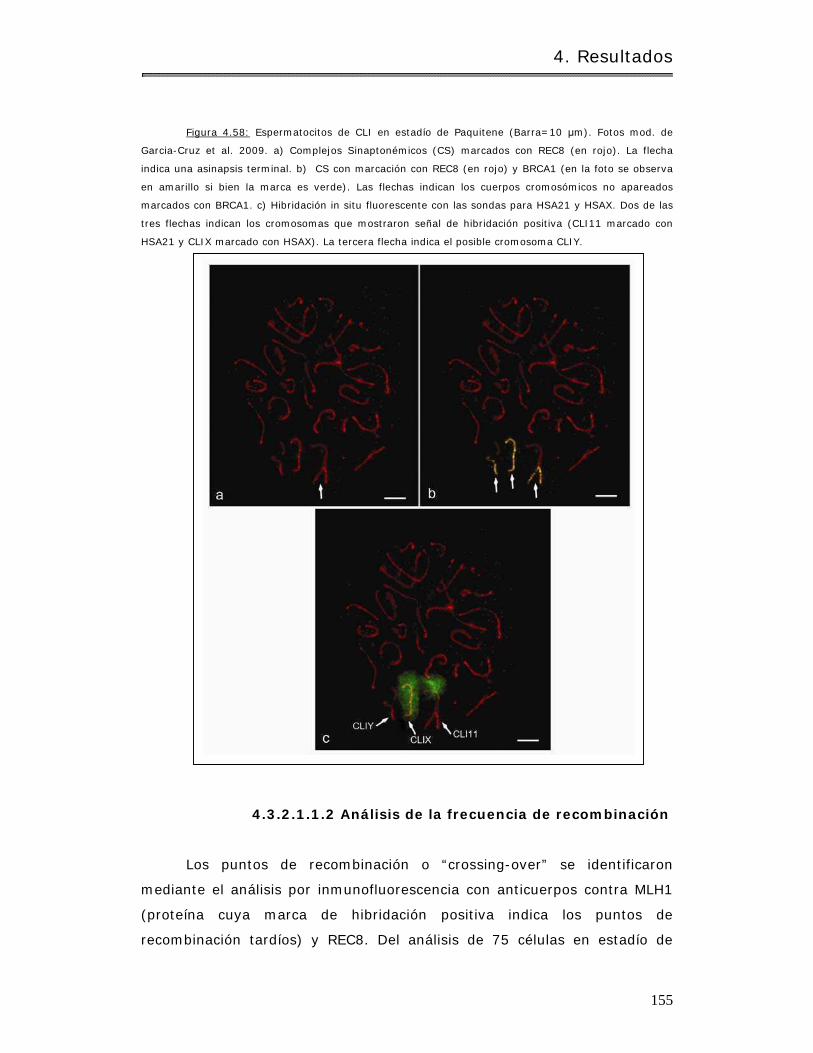
4. Resultados
155
Figura 4.58: Esperm atocitos de CLI en estadío de Paquitene (Barra= 10 μm ) . Fotos m od. de
Garcia-Cruz et al. 2009. a) Com plejos Sinaptoném icos (CS) m arcados con REC8 (en rojo) . La f lecha
indica una asinapsis term inal. b) CS con m arcación con REC8 (en rojo) y BRCA1 (en la foto se observa
en am arillo si bien la m arca es verde) . Las flechas indican los cuerpos cromosóm icos no apareados
marcados con BRCA1. c) Hibr idación in situ fluorescente con las sondas para HSA21 y HSAX. Dos de las
t res flechas indican los cromosom as que m ost raron señal de hibr idación posit iva (CLI 11 m arcado con
HSA21 y CLI X marcado con HSAX) . La tercera flecha indica el posible crom osoma CLI Y.
4 .3 .2 .1 .1 .2 Aná lisis de la frecuencia de recom binación
Los puntos de recom binación o “ crossing-over” se ident ificaron
m ediante el análisis por inmunofluorescencia con ant icuerpos cont ra MLH1
(proteína cuya m arca de hibridación posit iva indica los puntos de
recombinación tardíos) y REC8. Del análisis de 75 células en estadío de

4. Resultados
156
Paquitene (Figura 4.59a) surgió que el núm ero de foci de MLH1 en CLI se
encuent ra en un rango de 33 a 51 por núcleo, con un valor m edio de
43,22± 5,38 por núcleo paquiténico (Coeficiente de variación, CV= 12,45% ) .
En 83 núcleos paquiténicos de CLI se analizó el núm ero de foci de
MLH1 luego de la hibr idación in situ con la sonda para HSA21. En 92,77%
de las células (n= 77) se detectó una única señal de hibr idación posit iva
para MLH1 en el bivalente CLI 11 (Figura 4.59b) , m ient ras que en el resto
de las células analizadas (7,23% , n= 6) se observaron 2 foci en ese
bivalente y así se obtuvo un valor m edio de 1,07 foci de MLH1 por bivalente
CLI 11.
Figura 4.59: Esperm atocitos de CLI en estadío de Paquitene (Barra= 10 μm ) . Fotos m od. de
García-Cruz et al. 2009. a. Esperm atocito m arcado con MLH1 (verde) y REC8 ( rojo) . b. El m ism o
esperm atocito m arcado con la sonda para HSA21 (verde) m anteniendo la m arcación para REC8 ( rojo) .
4 .3 .2 .1 .1 .3 Com portam iento del par sexual XY
Los cromosomas X e Y se observaron asinapsados y m ost raron señal
posit iva para BRAC1 (ver Figura 4.60a) en 6 de los 12 núcleos que
m ost raron crom osom as desapareados en Paquitene (sección 4.3.2.1.1.1,
página 155) . En 4 de esos 12 núcleos se observó un cuerpo XY norm al, con
sinapsis a nivel de la región pseudoautosóm ica, y en los restantes 2
núcleos, el estado de sinapsis de los crom osom as X e Y no pudo ser
determ inado debido a que los cuerpos crom osómicos estaban m uy plegados

4. Resultados
157
y superpuestos y no se podían dist inguir con nit idez y precisión el estado de
apaream iento alcanzado. Sin em bargo, se puede describir que los
crom osom as sexuales X e Y serían los últ imos en iniciar la sinapsis. Esta
inferencia se basa en la observación de núcleos en Paquitene temprano en
los que todos los bivalentes autosóm icos ( incluyendo CLI 11) se encontraban
totalm ente sinapsados a excepción de los crom osom as X e Y (Figura 4.60b) .
Aún así, a pesar de no presentar sinapsis, los crom osom as X e Y
m ost raron regiones de hibridación posit iva para SYCP1. Se puede pensar
que una vez form ado el cuerpo XY, la m orfología del m ism o most raría una
conform ación sim ilar a la del cuerpo XY hum ano. Sin em bargo, al hibr idar
con REC8 y SYCP1 se observó la presencia de m ás de una región posit iva
para SYCP1 en el cuerpo XY, en la región correspondiente al eje del
crom osom a X (Figura 4.60) .
Figura 4.60: Dos esperm atocitos de CLI m arcados con REC8 (verde) y SCYP1 ( rojo) en dist intos
m om entos del estadío de Paquitene (Barra= 10 μm ). Fotos m od. de García-Cruz et al. 2009. a. Los
cromosom as X e Y aún perm anecen desapareados. Se puede observar señal de SYCP1 en los ejes
desapareados del X e Y ( indicados con flechas) . b. El cuerpo XY ya está form ado. Se puede observar
m ás de un focus de SYCP1 en el eje del crom osom a X.

4. Resultados
158
4 .3 .2 .1 .2 Alouat ta caraya
4 .3 .2 .1 .2 .1 Análisis de la frecuencia de recom b inación
Cuando el análisis del proceso de recombinación se llevó a cabo en el
m ono aullador negro y dorado (A. caraya= ACA) , el m ater ial obtenido de la
biopsia no m ost ró el m ism o rendim iento celular que el obtenido con
C. libidinosus. La calidad de los preparados no fue buena y en los
preparados obtenidos no se pudieron observar núcleos en estadios
anteriores o posteriores a Paquitene, por lo que no se pudo caracterizar el
comportam iento m eiót ico en la Profase I .
Dadas las condiciones del m aterial disponible, en esta especie se
ut ilizaron ant icuerpos cont ra las siguientes proteínas: SYCP3 (componente
de los elem entos laterales del com plejo sinaptoném ico o CS) , MLH1 y CenP
(suero CREST, que m arca los cent róm eros) y no se repit ieron los ensayos
previam ente realizados en las biopsias test iculares de CLI .
En esta oportunidad se analizaron 165 esperm atocitos de ACA en
estadío de Paquitene. En los bivalentes autosóm icos se caracter izó por
pr imera vez el núm ero de foci de MLH1, valor que osciló ent re 31 y 50, con
una m edia de 40,56± 4,3 (Figura 4.61a- f) . Asim ism o, se puso en evidencia
la localización y est ructura del CS m ediante la m arcación en rojo de la
proteína SYCP3. Los cent róm eros se pueden contabilizar ya que se
visualizan claram ente en color azul correspondiente a la hibr idación e
inm unolocalización con la sonda de CenP. Nuevam ente la m arca de los foci
de MLH1 pone en evidencia los puntos de recom binación que serían
equivalentes a los quiasmas observables en Diacinesis/ Metafase I .
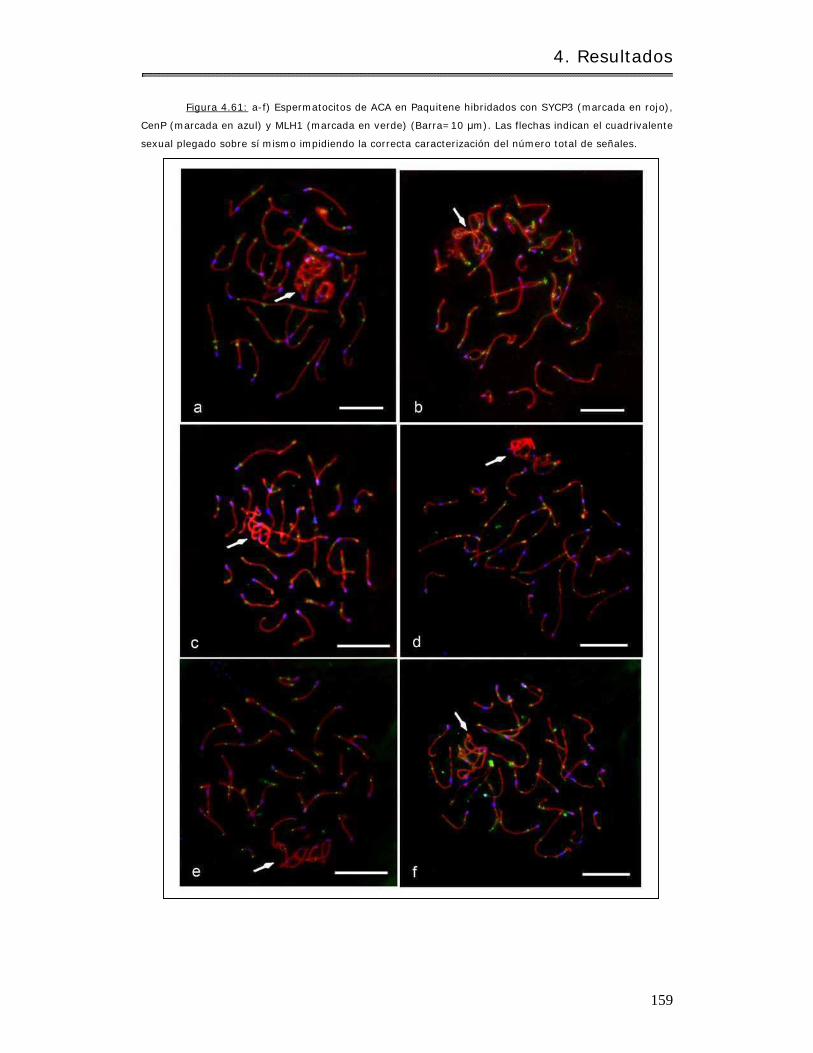
4. Resultados
159
Figura 4.61: a- f) Esperm atocitos de ACA en Paquitene hibr idados con SYCP3 (m arcada en rojo) ,
CenP (m arcada en azul) y MLH1 (marcada en verde) (Barra= 10 μm ) . Las flechas indican el cuadr ivalente
sexual plegado sobre sí m ism o im pidiendo la correcta caracter ización del número total de señales.

4. Resultados
160
4 .3 .2 .1 .2 .2 Análisis del cuadr iva lente sexual
En todos los núcleos analizados, el cuadrivalente sexual de ACA se
observó plegado sobre sí m ism o, de form a que im pidió un análisis
exhaust ivo de este cuerpo sexual por la técnica de inmunofluorescencia. La
Figura 4.62 ejem plifica esperm atocitos en el estadio de Paquitene con
dist intas conform aciones de plegam iento del cuadr ivalente sexual.
Figura 4.62: El cuadr ivalente sexual en esperm atocitos de ACA en Paquitene hibr idados con
SYCP3 (m arcada en rojo) , CenP (m arcada en azul) y MLH1 (m arcada en verde) (Barra= 5 μm ) . Las
flechas indican los foci de MLH1 a. Cuadrivalente sexual plegado de form a que no se puede observar
ningún focus de MLH1. b. Cuadrivalente en el que se observan 2 foci. c. Cuadrivalente en el que se
observan 3 foci. d. Cuadrivalente en el que se pueden observar 4 foci de MLH1.
Sin embargo, a pesar de los plegam ientos, ciertos valores se
pudieron cuant ificar en los núcleos de ACA analizados. Así, en 34% de los
núcleos analizados se pudieron dist inguir dos foci de MLH1 en el
cuadrivalente. En los restantes núcleos, en 23% se observaron 4 foci de
MLH1 en el cuadr ivalente y en 23% se observaron 3 foci en los
cuadrivalentes analizados. En 20% de los núcleos restantes de ACA
analizados no se pudo dist inguir ningún focus debido al plegam iento del
cuadrivalente.

4. Resultados
161
4 .4 Estudios f ilogenét icos
4 .4 .1 Mat r iz Básica de datos
Los datos obtenidos a part ir de las hom eologías analizadas por
bandas G y por FI SH se ut ilizaron como base para realizar un análisis
cladíst ico.
Para la elección de los caracteres cromosóm icos a estudiar se
seleccionaron los caracteres ut ilizados previam ente por Neusser et al. (2001) ,
que fueran m odificados para aulladores por de Oliveira et al. (2002) .
En base a los resultados obtenidos a part ir de las comparaciones
cariot ípicas con bandas G y FI SH con las sintenias 3/ 21 y 3/ 15, se
incorporaron los datos obtenidos para las especies m esoam ericanas
(A. pigra y A. palliata) en la m at r iz de datos descr ita en de Oliveira et al.
(2002) , m odificando la nom enclatura para relacionarla con la nom enclatura
de las bandas G del idiogram a hum ano (ver Materiales y Métodos, sección
3.4.2 página 59) .
A la lista de los 98 caracteres descritos previam ente, se adicionó
com o ext ra el carácter para la asociación Y/ 7, que corresponde a la
t ranslocación Y-autosom a que dio origen a los m ult ivalentes en API y en
APA, const ituyendo así para este análisis, un total de 99 caracteres
crom osóm icos. La asociación 4/ 9/ 11, observada en API 1, no se incluyó en el
análisis dado que al ser una autopom orfía para API , se considera un
carácter no inform at ivo.
En el análisis original realizado por de Oliveira et al. (2002) se tom aron
en cuenta dos cariot ipos diferentes encontrados para A. m acconnelli, uno
proveniente de ejem plares de la zona del r ío Jari (Brasil) y el ot ro de la
zona del r ío Uatum ã, también en Brasil. Ambos car iot ipos difieren por dos
t ranslocaciones Robertsonianas. El cariot ipo de los individuos del r ío Uatum ã
t iene las form as crom osóm icas con hom eología con HSA2p/ HSA2q y HSA16.

4. Resultados
162
Los individuos del r ío Jari t ienen las form as HSA2q y HSA2p/ 16. Dado que
am bas formas crom osómicas const ituyen autopom orfías se decidió incluir
en el análisis sólo uno de los car iot ipos, el del río Jari. Considerando que la
secuencia de Cyt obtenida del genbank para esta especie proviene de la
Guyana Francesa y que la zona del r ío Jari está m ás cercana a ésta, se
decidió ut ilizar este cariot ipo en el análisis (codificado com o A.m acconneJ en
los análisis que se describen a cont inuación) .
En el caso de A. seniculus seniculus y A. seniculus arctoidea, dada la
falta de regist ro de datos en el genbank para A. seniculus seniculus, se
incluye una única subespecie en el análisis.
Una vez realizado el análisis por Máxim a Parsim onia y obtenida la red
o redes resultantes, para conocer las posibles relaciones filogenét icas se
hace necesar io darles polaridad a las m ismas. En este t rabajo de Tesis
Doctoral se ut ilizó el cr iter io del grupo externo (del inglés outgroup) . Por
convención se elige un taxón para el cual los estados de los caracteres
determ inan los estados ancest rales para el grupo de especies en las que se
va a analizar las relaciones de parentesco. En esta oportunidad se tom ó
com o outgroup la especie C. libidinosus ya que a nivel cromosóm ico es
aceptado que presenta el cariot ipo m ás ancest ral dent ro de los Platyrrhini
(Dut r illaux 1979; Clem ente et al. 1990; García et al. 2000) Recordando que para
establecer las relaciones filogenét icas mediante un análisis de t ipo
cladíst ico, siguiendo a Henning (1966) , son inform at ivos los grupos
m onofilét icos, se elige un segundo taxón para analizar la m onofilia del
grupo en estudio. En este caso se tom ó la especie Lagothrix lagot r icha.
En este contexto se obtuvieron 3 m atr ices básicas de datos: una
m at r iz com prendiendo sólo los caracteres crom osómicos, ot ra m at r iz
comprendiendo los caracteres m oleculares disponibles hasta el m om ento de
redacción del presente t rabajo de Tesis Doctoral, para lo cual se tomó en
consideración el gen m itocondrial Citocrom o b (Cytb) , y una tercera m at r iz,
combinando am bos t ipos de datos. Este es el pr im er t rabajo en el que se
realiza un análisis ext ra en aulladores respecto del t radicional análisis por

4. Resultados
163
part iciones, teniendo com o objet ivo cont r ibuir al mejor conocim iento de la
sistem át ica de aulladores dent ro el marco conceptual de “Evidencia total” .
En el Anexo I V se describen las dist intas m at r ices obtenidas.
4 .4 .2 Análisis por PAUP
El análisis de Máxim a Parsim onia se realizó a part ir de las t res
m at r ices antes referidas, ut ilizando la opción de búsqueda exhaust iva del
program a PAUP 4.0. La estabilidad de los nodos se evaluó m ediante el
com ando “bootst rap” ut ilizando 200 iteraciones.
4 .4 .2 .1 Caracteres crom osóm icos
El análisis de los datos crom osómicos produjo 36 caracteres
inform at ivos, 23 caracteres constantes y 40 caracteres no inform at ivos.
Luego de analizar 704 árboles, el program a PAUP retuvo los dos árboles
m ás parsim oniosos (Figuras 4.63 y 4.64) , ambos con un largo de 87
(L= 87) .
Figura 4.63: Uno de los dos árboles m ás parsim oniosos obtenidos para la part ición de datos
crom osóm icos en Ceboidea (Prim ates: Platyrr ini) .

4. Resultados
164
Figura 4.64: Segundo árbol m ás parsim onioso obtenido para la part ición de datos
crom osóm icos en Ceboidea (Prim ates: Plathyrr ini) .
A cont inuación se describe la interpretación de los resultados y valores en
los nodos luego del análisis de “bootst rap” (Figura 4.65) :
Figura 4.65: Árbol consenso de 50% de m ayoría obtenido m ediante el algor itm o de “bootst rap” .

4. Resultados
165
1. Las especies del género Alouat ta form an un grupo independiente de
C. libidinosus y L. lagothr icha con una frecuencia de 65%
2. Las subespecies de A. guariba y A. sara, A. m acconelli y A. s.
arctoidea form an un grupo independiente con una frecuencia de 90%
3. Las subespecies de A. guariba form an un grupo independiente con
una frecuencia de 100% .
4. A. sara, A. m acconelli y A. s. arctoidea form an un grupo
independiente con una frecuencia de 96% .
5. Las especies A. sara y A. seniculus arctoidea form an un grupo
independiente con una frecuencia de 99% .
6. Las especies A. caraya y A. belzebul form an un grupo independiente
con una frecuencia de 86%
7. Las especies A. pigra y A. palliata form an un grupo independiente con
una frecuencia de 79%
El análisis ut ilizando sólo la part ición de datos cromosóm icos no pudo
resolver el nodo ( (APA,API ) ,(ACA,ABE) ,( (AGUc,AGUg) ,(AMAJ,(ASA,ASEa) ) ) ,
ya que se const ituyó en una politom ía.
A part ir de los resultados obtenidos de esta part ición de datos se puede
infer ir que ABE y ACA estarían m ás cercanos ent re sí que respecto al grupo
form ado por ASEa, ASA y AMAJ y que respecto al grupo de las subespecies
de AGU, la ot ra especie de dist r ibución marginal sur del género ( junto con
ACA) . Las relaciones ent re ABE y ACA con los aulladores m esoam ericanos
(APA y API ) no pudieron resolverse con esta part ición.
4 .4 .2 .2 Caracteres m oleculares
El análisis heuríst ico de las secuencias para el gen Cytb, realizado a
part ir de un total de 800 caracteres, produjo 109 caracteres inform at ivos,
551 caracteres constantes y 140 caracteres no inform at ivos. El programa
PAUP luego de analizar 916 árboles, retuvo un único árbol más
parsimonioso (Figura 4.66) , de un largo L= 366.
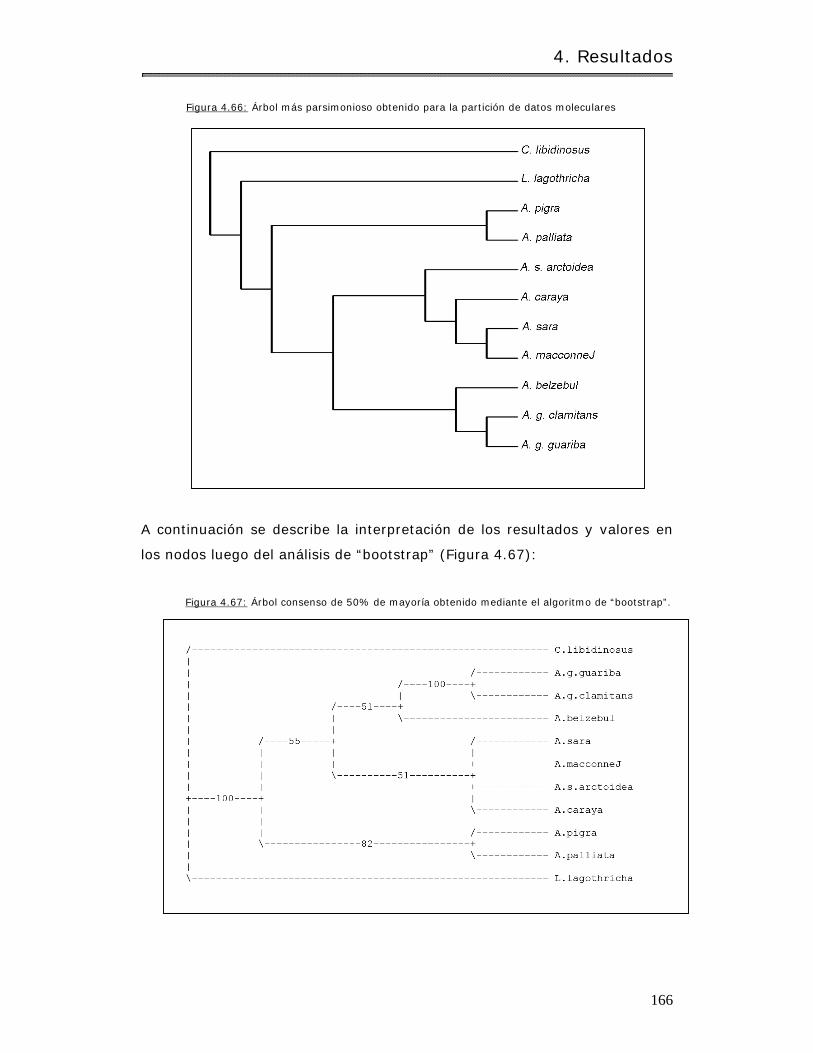
4. Resultados
166
Figura 4.66: Árbol m ás parsim onioso obtenido para la part ición de datos m oleculares
A cont inuación se describe la interpretación de los resultados y valores en
los nodos luego del análisis de “bootst rap” (Figura 4.67) :
Figura 4.67: Árbol consenso de 50% de m ayoría obtenido m ediante el algoritm o de “bootst rap” .

4. Resultados
167
1. Las especies del género Alouat ta form an un grupo independiente de
C. libidinosus y L. lagothr icha con una frecuencia de 100% .
2. Las especies m esoam ericanas (APA y API ) forman un grupo
independiente con una frecuencia de 82% .
3. Las especies sudam ericanas forman un grupo independiente con una
frecuencia de 55% .
4. A. sara, A. m acconelli y A. s. arctoidea y A. caraya form an un grupo
independiente con una frecuencia de 51% .
5. La especie A. belzebul y las subespecies de A. guariba form an un
grupo independiente con una frecuencia de 51% .
6. Las subespecies de A. guariba form an un grupo independiente con
una frecuencia de 100%
El análisis ut ilizando sólo la part ición de datos moleculares no pudo resolver
el nodo (ASA,AMAJ,ASEa,ACA) ya que se const ituyó en una politom ía que si
regresam os en el texto, podem os observar que es diferente de la descr ipta
para los datos crom osóm icos.
A part ir de los resultados obtenidos de esta part ición de datos se puede
inferir que ABE y las subespecies de AGU estarían m ás cercanos ent re sí
que respecto al grupo form ado por ASEa, ASA, ACA y AMAJ y que respecto
al grupo especies m esoam ericanas. Las relaciones ent re ASEa, ASA, ACA y
AMAJ no pudieron resolverse con esta part ición.
Ante estos hallazgos cobró una m ayor im portancia el ya planteado a prior i,
análisis combinado.
4 .4 .2 .3 Análisis com binado
El análisis heuríst ico combinando en una única m atr iz los datos de
hom eologías crom osóm icas y las secuencias para el Cytb puso en evidencia
que de un total de 899 caracteres, 145 caracteres resultaron inform at ivos,
547 caracteres constantes y 180 caracteres no inform at ivos. El programa

4. Resultados
168
PAUP luego de analizar 684 árboles, retuvo un único árbol más
parsimonioso (Figura 4.68) , de un largo L= 460.
Figura 4.68: árbol m ás parsim onioso obtenido durante el análisis com binado.
A cont inuación se describe la interpretación de los resultados y valores en
los nodos luego del análisis de “bootst rap” (Figura 4.69) :
Figura 4.69: Árbol consenso de 50% de m ayoría obtenido m ediante el algor itm o de “bootst rap” .

4. Resultados
169
1. Las especies del género Alouat ta form an un grupo independiente de
C. libidinosus y L. lagothr icha con una frecuencia de 100% .
2. Las especies m esoam ericanas (APA y API ) forman un grupo
independiente con una frecuencia de 90% .
3. Las especies sudam ericanas forman un grupo independiente con una
frecuencia de 61% .
4. A. belzebul y el resto de las especies sudam ericanas form an un grupo
independiente con una frecuencia de 56% .
5. Las subespecies de A. guariba form an un grupo independiente con
una frecuencia de 100%
6. A. sara, A. macconelli, A. s. arctoidea y A. caraya form an un grupo
independiente con una frecuencia de 58% .
7. A. sara, A. m acconelli, A. s. arctoidea form an un grupo independiente
con una frecuencia de 89% .
8. Las especies A. sara y A. s. arctoidea forman un grupo independiente
con una frecuencia de 85% .
Esta m odalidad de análisis perm it ió resolver todos los nodos al resultar en
un arbol libre de politom ías.
A part ir de los resultados obtenidos de esta part ición de datos se puede
infer ir que ACA y el grupo form ado por ASEa, ASA y AMAJ estarían m ás
cercanos ent re sí que respecto a las subespecies de AGU y que respecto a
las especies m esoam ericanas.

DI SCUSI ÓN

5. Discusión
170
5 .1 Citogenét ica y diagnosis taxonóm ica
Tal com o se ha expresado en Materiales y Métodos, como parte de
este t rabajo de Tesis Doctoral se caracter izaron citogenét icamente 37
ejemplares de Ceboidea provenientes tanto de vida silvest re com o de
caut iverio.
Las variables crom osómicas analizadas perm it ieron corroborar el
estatus de especie en todos los ejem plares analizados y, en el caso de uno
de los especim enes de Alouat ta palliata, se m odificó su asignación de sexo
de hem bra a m acho, según fuera previam ente ident ificado en condiciones
de caut iverio.
Como resultado inicial or iginal de este t rabajo cabe comentar que se
realizó la primera caracter ización del cariot ipo del m ono aullador negro,
Alouat ta pigra, estudiando ejem plares de ambos sexos de vida silvest re
m uest reados en áreas de la dist r ibución geográfica de la especie y en
colaboración con nat ivos del lugar. Los datos obtenidos de este análisis se
ut ilizaron poster iorm ente com o pat rón para la diagnosis taxonóm ica de 3
ejemplares alojados en caut iverio en zoológicos del Dist r ito Federal,
México.
Si bien la prim era revisión sistem át ica sobre los aulladores
m esoam ericanos (Lawrence 1933) coloca a A. pigra com o subespecie de
A. palliata, revisiones poster iores recom endaron considerarla com o especie
plena teniendo en cuenta característ icas m orfológicas (Sm ith 1970; Wolfheim
1983) , clasificación m ás recientem ente confirm ada por estudios genét icos
(Cortés-Ort iz et al. 2003) . Los resultados obtenidos en el presente t rabajo de
Tesis Doctoral br indan nuevas evidencias para avalar la asignación de
A. pigra com o especie plena. Mient ras que A. palliata posee un 2N= 53 y un
sistem a sexual m últ iple m asculino de t ipo X1X2Y (Ma et al. 1975; Solar i y Rahn
2005) , A. pigra posee un 2N= 58 y un sistem a sexual m últ iple m asculino de
t ipo X1X2Y1Y2. Su núm ero diploide es el m ás alto descripto para el género
hasta la fecha de escritura de este t rabajo de Tesis. A. pigra posee 2 pares

5. Discusión
171
crom osóm icos submetacént r icos menos y 4 pares acrocént r icos m ás que
A. palliata. De las hom eologías crom osómicas realizadas se desprende que
ent re API y APA exist ir ían 20 crom osom as conservados enteros,
2 reordenam ientos de t ipo fisión/ fusión y 2 inversiones paracént r icas para
completar la interpretación de reordenam ientos cromosóm icos diferenciales
ent re am bas especies.
La caracter ización del cariot ipo de API cobra especial im portancia
debido a que se han ident ificado genét icam ente híbridos ent re APA y API
en una zona de simpat ría en la localidad de Tabasco (México) . El análisis
combinando inform ación obtenida a part ir de m icrosatélites, ADN
m itocondrial y el gen SRY dem ost ró la presencia de individuos híbridos en
esta zona, exist iendo un sesgo en la dirección en que ocurre la hibr idación.
Los cruzam ientos ent re hem bras de APA y m achos de API producen
descendencia inviable o infért il. Los únicos cruzamientos con descendencia
viable y fért il, ocurr ir ían ent re hem bras de API y m achos de APA o ent re
hembras híbridas portadoras del haplot ipo m itocondrial de API y m achos de
API o APA (Cortés-Ort iz et al. 2007) . La descendencia masculina producto de
estos cruzam ientos es inviable o infért il, lo cual coincidir ía con lo postulado
por la regla de Haldane (Haldane 1922) .
Este no es el único caso de hibr idación observado en el género. En la
dist r ibución m arginal sur de Alouat ta se han descrito t ropas com puestas por
individuos de A. caraya y A. guariba clam itans, observando individuos con
pat rón de coloración de pelaje interm edio ent re am bas especies (Aguiar et al.
2007; Agost ini et al. 2008) , si bien aún no se ha confirm ado la sugerida
hibridación m ediante estudios genét icos.
La ocurrencia de hibridación ent re dos especies cariológicam ente
diferentes, no sólo a nivel del com plejo autosóm ico sino tam bién a nivel de
su sistem a de determ inación sexual, enfat iza la im portancia de los estudios
m eiót icos para la m ejor com prensión de la sistem át ica y taxonom ía de los
Platyrrhini. Si bien la literatura combinando m etodologías de valor
diagnóst ico con predicciones aplicables en sistem át ica y citotaxonom ía son

5. Discusión
172
m uy escasas, estudios recientes han dado cuenta de ello en dos especies de
Cebus m antenidas en caut iverio (Nieves et al. 2008; Fant ini 2010) . La realización
de estudios m itót icos y m eiót icos en individuos híbridos y en sus parentales,
perm it irá entender la interacción de los genom as de origen del híbr ido a
nivel cromosómico, perm it iendo explorar potenciales aislam ientos
reproduct ivos bajo el m arco de la evolución cromosómica com o posible
m ecanism o de estudio.
5 .2 Com portam iento m eiót ico de los crom osom as sexua les
5 .2 .1 Diagnosis por citogenét ica clásica
En 5 de las 8 especies analizadas (Alouat ta caraya, A. pigra,
A. palliata, Cebus libidinosus y Saim ir i boliv iensis) se realizaron estudios
m eiót icos que confirmaron el sistem a cromosómico de determ inación sexual
propuesto m ediante los análisis m itót icos previos.
En los aulladores analizados, una comparación de la est ructura de los
m ult ivalentes observados most ró m arcadas diferencias ent re ellos (Figura
5.1) . En API se form a un quiasm a interst icial ent re Y2 y X2 (Figura 5.1a) ,
m ient ras que en ACA el quiasm a interst icial se form a ent re Y1 y X2 (Figura
5.1b) . Más aún, ent re Y1 y X2 en API se form a un quiasm a term inal, con la
característ ica unión ext rem o-ext rem o ( “end- to-end joining” ) de Y1 y X2 a
cont inuación. Esta unión por los ext remos es sim ilar en am bos
cuadrivalentes. En el t r ivalente sexual de APA (Figura 5.1c) el quiasma
interst icial ocurre ent re Y y X2 con la unión ext rem o-ext rem o ent re Y1 y X1
m ost rando una est ructura sim ilar a la de API y ACA.

5. Discusión
173
Figura 5.1: Comparación de los sistem as de determ inación sexual crom osóm icos en Metafase I
en t res especies de aulladores. Las flechas indican los componentes de los mult ivalentes. Las cabezas de
flecha indican la unión ext rem o-ext rem o y los asteriscos los quiasm as interst iciales. a) Alouat ta pigra.
b) Alouat ta caraya. c) Alouat ta palliata.
Esta configuración observada con m icroscopía ópt ica en los
m ult ivalentes de Alouat ta, X1-Y1-X2-Y2 en el caso de los sistem as de t ipo
cuadrivalente y X1-Y1-X2 en el caso de los sistem as de t ipo t r ivalente,
perm ite la segregación alternada en Anafase I , la que asegura la producción
de gam etas balanceadas y la fert ilidad del individuo.
5 .2 .2 Diagnosis por caracter ización citom olecular
5 .2 .2 .1 Cebus libidinosus
Estudios m eiót icos previos en C. libidinosus con la técnica de t inción
por nit rato de plata descr ibieron el com portam iento del par XY como
“hum an like” , es decir , con un com portam iento durante la división m eiót ica
sim ilar al del par XY hum ano (Seuánez et al. 1983; Mudry et al. 2001) .
En el presente t rabajo de Tesis se analizó el comportam iento m eiót ico
m ediante la técnica de Evans et al. (1964) y la técnica de inm unofluorescencia
(Roig et al. 2004) . El uso de esta últ ima perm it ió la visualización de proteínas
específicas involucradas en el proceso de recombinación. Com o una
característ ica a destacar, se observó m ás de una región con m arca posit iva
para SYCP1 en el par XY. Esta situación que no fue observada previam ente
en espermatocitos hum anos (R.García-Cruz y M.García-Caldés, com unicación

5. Discusión
174
personal) , sí se ha descripto en ot ras especies de m amíferos tales com o el
ratón Mus m usculus (Page et al. 2006) , sugir iendo que el comportam iento del
par XY en Cebus no se correspondería en su totalidad al llam ado t ipo
“hum an like” .
Las regiones SYCP1+ se corresponderían con áreas de autosinapsis
ent re secuencias repet it ivas en los ejes desapareados de los cromosomas
X e Y. Este m ecanismo perm it ir ía que dichas secuencias evadan al
reconocim iento por BRCA1 y escapen al silenciam iento (Turner et al. 2005) . Sin
embargo, el significado celular que podría tener este comportam iento
m eiót ico no está claro aún. Recientem ente se ha descrito que los ratones
que carecen de esta proteína (SYCP1 - / -) presentan test ículos anorm ales. Los
esperm atocitos de estos ratones m uest ran defectos en la form ación y
sinapsis del cuerpo XY, sugir iendo que esta proteína tendría un rol en el
ensamblaje del com plejo sinaptoném ico, en la recom binación m eiót ica y en
la form ación del cuerpo XY (de Vries et al. 2005) . Una explicación alternat iva
sería que estas regiones SYCP1+ corresponderían a agregados proteicos a lo
largo de los ejes, coincidiendo con lo previam ente observado por t inción con
nit rato de plata (Mudry et al. 2001) . Más estudios de inm unofluorescencia en
esta y ot ras especies de ceboideos son necesarios para poner a prueba esta
hipótesis.
El núm ero de foci de MLH1 observado en los esperm atocitos de CLI
es cercano al observado en esperm atocitos humanos (43,22± 5,38 en Cebus
vs 48,8± 2,3 en H. sapiens) (Codina-Pascual et al. 2005) . Sin embargo, el
núm ero de bivalentes observables y cuant ificables en am bas especies,
difiere considerablem ente. El complem ento autosóm ico de CLI está
compuesto por 10 pares cromosómicos bibraquiados y 16 pares
acrocént r icos. Por lo tanto, considerando que para asegurar una correcta
segregación en Anafase I los bivalentes bibraquiados deberían tener al
m enos un evento de recom binación por brazo (evidenciado m ediante los
foci de MLH1) y los bivalentes acrocéntr icos, al m enos un focus de MLH1
por bivalente, los esperm atocitos de CLI deberían tener al menos 38 foci de
MLH1. Dado que el núm ero m edio de foci observados es m ayor y que no se

5. Discusión
175
observaron bivalentes sin marca para MLH1 se podría interpretar que el
proceso de recom binación en CLI , ocurre de m anera eficiente a pesar de la
presencia de grandes bloques de heterocrom at ina ext racent rom érica. La
sinapsis de estos bivalentes, si bien se observa ret rasada respecto del resto,
ocurre correctam ente, perm it iendo un adecuado alineam iento de los
bivalentes en la placa m etafísica y una adecuada segregación en Anafase I .
5 .2 .2 .2 Alouat ta caraya
En este aullador no fue posible establecer adecuadam ente el núm ero
de foci de MLH1 en sus crom osom as, debido al plegam iento sobre sí m ism o
del cuadrivalente sexual, si bien se lograron obtener datos cuant itat ivos
parciales (ver sección 4.3.2.1.2.2 página 160) .
El valor para el núm ero de focus de MLH1 obtenido no m uest ra
diferencias estadíst icam ente significat ivas respecto del obtenido en CLI
(p> 0,05 prueba de t -Student ) . Sin em bargo, es pert inente aclarar que, al
no haber sido posible cuant ificar el núm ero de focus de MLH1 en el
cuadrivalente sexual, se estaría subest im ando el valor m edio de los m ismos
para este ejemplar, único de la especie de aullador estudiada.
5 .3 El gen SRY en Ceboidea
En los Ceboidea analizados en este t rabajo de Tesis Doctoral
m ediante FI SH con la sonda para el gen SRY humano, no se observó señal
posit iva de hibridación para el SRY.
Estudios previos en Ceboidea ut ilizando análisis de biología m olecular
(m ediante PCR) (Moreira 2002; Nieves 2007; Cortés-Ort iz et al. 2003) y m ediante
FI SH (Gläser et al. 1998) describieron la conservación del SRY en este grupo
(ver sección 1.2.4.1 página 23) .
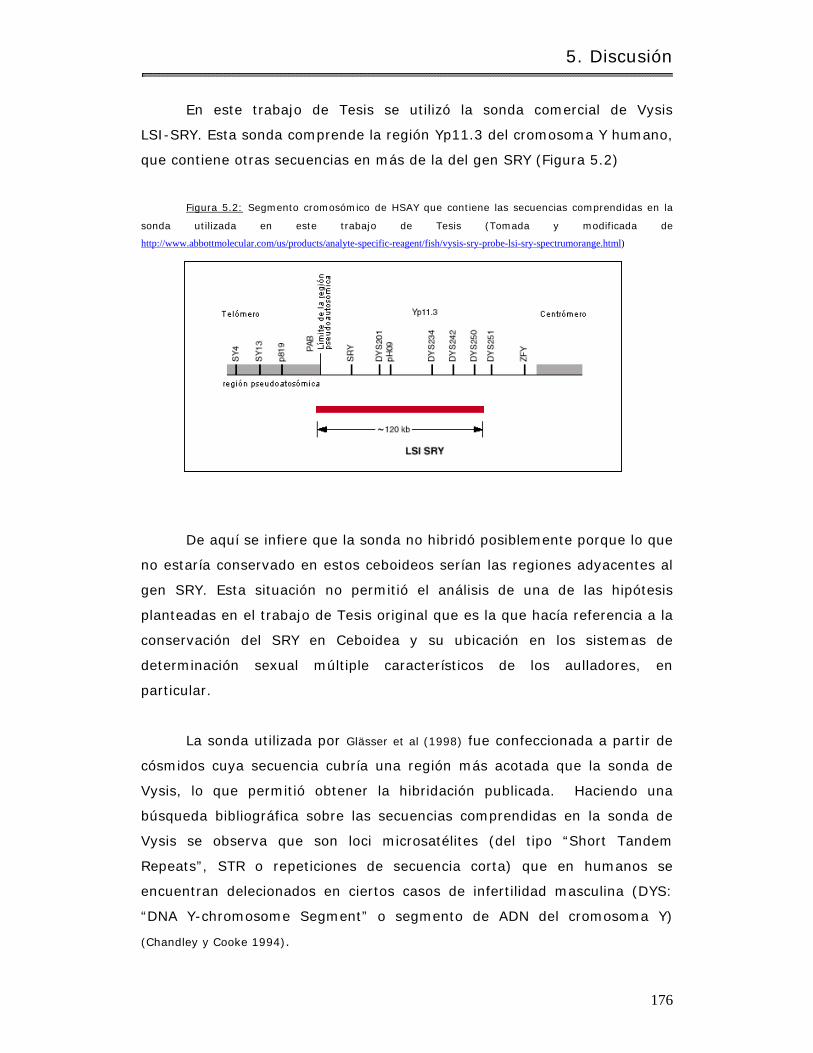
5. Discusión
176
En este t rabajo de Tesis se ut ilizó la sonda com ercial de Vysis
LSI -SRY. Esta sonda com prende la región Yp11.3 del cromosom a Y hum ano,
que cont iene ot ras secuencias en m ás de la del gen SRY (Figura 5.2)
Figura 5.2: Segmento cromosóm ico de HSAY que cont iene las secuencias comprendidas en la
sonda ut ilizada en este t rabajo de Tesis (Tom ada y m odificada de
http://www.abbottmolecular.com/us/products/analyte-specific-reagent/fish/vysis-sry-probe-lsi-sry-spectrumorange.html)
De aquí se infiere que la sonda no hibridó posiblem ente porque lo que
no estaría conservado en estos ceboideos serían las regiones adyacentes al
gen SRY. Esta situación no perm it ió el análisis de una de las hipótesis
planteadas en el t rabajo de Tesis original que es la que hacía referencia a la
conservación del SRY en Ceboidea y su ubicación en los sistem as de
determ inación sexual múlt iple característ icos de los aulladores, en
part icular.
La sonda ut ilizada por Glässer et al (1998) fue confeccionada a part ir de
cósm idos cuya secuencia cubría una región más acotada que la sonda de
Vysis, lo que perm it ió obtener la hibridación publicada. Haciendo una
búsqueda bibliográfica sobre las secuencias com prendidas en la sonda de
Vysis se observa que son loci microsatélites (del t ipo “Short Tandem
Repeats” , STR o repet iciones de secuencia corta) que en hum anos se
encuent ran delecionados en cier tos casos de infert ilidad m asculina (DYS:
“DNA Y-chrom osom e Segm ent ” o segm ento de ADN del crom osom a Y)
(Chandley y Cooke 1994) .

5. Discusión
177
Si bien en el cam po de la biología reproduct iva de los Platyrrhini aún
queda m ucho por estudiar, es interesante considerar que aún con la
presencia de com plejos sistem as de reproducción sexual m últ iple y grandes
bloques de heterocrom at ina ext racent rom érica, la fert ilidad de estos
pr imates parecería no estar afectada.
5 .4 Apor tes a la sistem át ica del género Alouat ta
5 .4 .1 Crom osom as sexu ales en aulladores
La caracterización cariot ípica de A. pigra perm it ió la ident ificación del
par acrocént r ico API 17 como el par autosóm ico involucrado en el sistem a de
determ inación sexual múlt iple (API X2 en los m achos) . Este par crom osómico
tendría hom eología con ACA14 y con APA19 (APAX2 en los machos) y en
consecuencia, tendría hom eología con HSA7q ter (sección 4.2.1.3.2) .
Estudios citogenét icos previos (Ma et al. 1975; Solar i y Rahn 2005)
describieron al par crom osóm ico APA19 como hom eólogo a ABE17 (ABEX2) .
ABE17 tendría hom eología con HSA3 (descrita tanto por bandas G com o por
FI SH con sondas hum anas) . Este es el pr imer estudio que se concreta por
FI SH con sondas hum anas en APA que perm ite confirmar hom eologías
crom osóm icas. En este estudio no se encont ró señal de hibr idación por FI SH
con la sonda de HSA3 en APA19 o en su hom eólogo para la ot ra especie de
aullador antes refer ida, es decir en API 17. La com paración ent re estos pares
crom osóm icos indica sim ilitudes en el pat rón de bandas G (Figura 5.3) , sin
embargo, de acuerdo a los pat rones de bandas G para hablar de
hom eología debería proponerse la ocurrencia de, al m enos, una inversión
paracént r ica. Tal com o se aclaró en la I nt roducción (sección 1.2.3 página
17) los estudios por FI SH han dem ostrado ser un m étodo m ás confiable
para el establecim iento de hom eologías, en part icular en el caso de
cariot ipos muy reordenados, com o es el caso del género Alouat ta.

5. Discusión
178
Figura 5.3: Hom eología por bandas G ent re los pares crom osóm icos ABE17, APA19 y API 17. A
la izquierda se m uest ra el idiogram a de bandas de ABE 17, indicando a la derecha del m ism o la
hom eología con HSA3.
Por las razones anter iorm ente expuestas, en este t rabajo de Tesis se
consideró que APA19 y API 17 tendrían hom eología con ACA14, y en
consecuencia con HSA7.
De la inform ación recabada se puede infer ir que el par autosóm ico
que dio origen a los sistem as sexuales m últ iples en las especies
m esoam ericanas serían diferentes al par autosóm ico que dio or igen a los
m ult ivalentes de las especies sudam ericanas. La formación de m ult ivalentes
habría ocurr ido de form a independiente en am bos grupos de aulladores.
De esta m anera, refir iéndonos a una de las hipótesis postuladas al
inicio de este t rabajo de Tesis Doctoral, la sintenia 3/ 15 no se encont raría
conservada en todas las especies de aulladores y no se encont raría asociada
a los cromosomas sexuales caracterizados en todas ellas.
La sintenia 3/ 21, ancest ral para m am íferos placentar ios y conservada
en los Pr imates Neot ropicales que fueran analizados con anterior idad a este
t rabajo de Tesis Doctoral, no se encont raría conservada en las especies de
aulladores m esoam ericanos, A. pigra y A. palliata. Esta falta de
conservación const ituir ía una sinapom orfía para este grupo.

5. Discusión
179
5 .4 .2 Reconst rucción f ilogenét ica del gé nero
Como aporte or iginal de este t rabajo de Tesis Doctoral se analizaron
las posibles homeologías cromosómicas com parando los pat rones de bandas
G de las dos especies de aulladores m esoam ericanas, A. pigra y A. palliata,
con A. caraya, ésta últ ima cuya dist r ibución geográfica llega a la Argent ina
y que const ituye una de las dos especies de dist r ibución m arginal sur
ext rem a del género. Las hom eologías cromosóm icas propuestas se
ut ilizaron para inferir el pat rón de hibridación in situ fluorescente con
sondas hum anas en API y APA, especies no estudiadas hasta la fecha con
esta m etodología.
Estos datos originales fueron reunidos y ut ilizados para la confección
de una filogenia junto con los datos analizados por de Oliveira et al. (2002) .
Com o se indicó en la I nt roducción (sección 1.3.1.3 página 32) , es
dent ro del marco del concepto de “Evidencia Total” que se pueden lograr
im portantes avances en el conocim iento sistem át ico de una determ inada
especie o grupo de ellas. El ut ilizar el m ayor núm ero posible de var iables en
la reconst rucción filogenét ica perm ite obtener una m ayor congruencia
taxonóm ica.
En este caso se analizaron dos t ipos de variables: crom osóm icas ( las
asociaciones/ sintenias de crom osom as hum anos infer idas m ediante las
hom eologías en el caso de API y APA y obtenidas m ediante FI SH en las
ot ras especies) y m oleculares (secuencias para el gen m itocondrial
Citocrom o b) . Para la elección de este m arcador m olecular se tom ó en
consideración la disponibilidad de secuencias en genbank para las m ism as
especies que com o parte del experim ental de la Tesis Doctoral habían sido
analizadas cromosómicam ente. Si bien en las últ im as dos décadas se han
realizado num erosos estudios filogenét icos en Platyrrhini, estos no
comprenden la totalidad de las especies descriptas hasta el presente y la

5. Discusión
180
disponibilidad de datos moleculares con secuencias publicadas para estos
pr imates es baja.
Con fines com parat ivos se realizaron reconst rucciones filogenét icas
independientes con cada t ipo de part ición, junto con el análisis combinado
de las dos var iables. Los t res t ipos de análisis coincidieron en ubicar a las
especies m esoam ericanas com o un clado separado del resto de las especies
sudam ericanas y, com o era de esperar, a las dos subespecies de A. guariba
en un m ism o grupo.
Ot ra coincidencia se observa en la agrupación de A. sara (ASA) ,
A. s. arctoidea (ASEa) y A. m acconnelli (AMA) com o grupo único, si bien la
part ición de datos m oleculares no pudo resolver las relaciones ent re ellos ya
que se obtuvo una politom ía.
La part ición cromosóm ica agrupó com o taxa herm anos a A. caraya y
A. belzebul (ABE) , relación que no se volvió a repet ir en los ot ros dos
análisis, que agruparon a ACA con ASA, ASEa y AMA (si bien en el caso de
la part ición molecular esta relación se const ituye en una politom ía) . En la
part ición de datos moleculares ABE es agrupado con el clado de A. guariba
(AGU) .
Todos los conjuntos de taxa analizados sólo se pudieron resolver sin
politom ías con el análisis combinado, dem ostrando la ut ilidad de este t ipo
de análisis que incorpora m ás de una fuente de datos con el marco teórico
de “Evidencia Total” para la m ás ajustada elucidación de las relaciones de
parentesco ent re taxa actuales.
5 .4 .3 Biogeografía y crom osom as sexuales en Alouat ta
El agrupam iento de las especies sudam ericanas com o un grupo
separado de las m esoam ericanas coincide con análisis filogenét icos previos
ut ilizando sólo caracteres moleculares (Cortés-Ort iz et al. 2003; Ellsworth y Hoelzer

5. Discusión
181
2006) y con la hipótesis del or igen monofilét ico de los aulladores
m esoam ericanos propuesta por Smith (1970) .
Las teorías actuales sost ienen que los aulladores m esoam ericanos se
originaron por una expansión de la dist r ibución geográfica de los aulladores
sudam ericanos después de la form ación del istm o de Panam á (Fleagle 1999) ,
que se est ima se completó hace aproxim adam ente 3 m illones de años
(Coates et al. 2004) . Sin em bargo, estudios recientes indican que el
levantam iento del istmo const ituyó un proceso más que un evento (Knowlton
y Weigt 1998) , que resultaría en períodos interm itentes con t ierras conectadas
y divididas durante los últ im os 18 m illones de años. Teniendo en cuenta
este últ imo dato, ot ras hipótesis surgen para un análisis más actualizado.
En lugar de un único evento de colonización se propone la ocurrencia de
var ios eventos fundadores, ya sea a t ravés del istmo de Panam á durante
uno de los períodos en que las dos Américas estaban conectadas o t ravés
de islas en el archipiélago caribeño. Esta últ im a hipótesis parecería estar
sustentada por el descubrim iento de fósiles de prim ates en Cuba y Jam aica
(Feagle 1999) .
I ndependiente del escenario que se considere com o cierto, los
aulladores m esoam ericanos t ienen una histor ia evolut iva separada de los
sudam ericanos, afirm ación que es respaldada por la evidencia de la
filogenia con análisis combinado, estudio realizado en este t rabajo de Tesis
Doctoral.
Como se ha refer ido en la I nt roducción (sección 1.2.2.3 página 12) el
sistem a de determ inación sexual considerado m ás ancest ral para los
m am íferos es el sistem a XY. Dent ro de la fam ilia Atelidae, con excepción de
Alouat ta, los géneros presentan determ inación sexual XX/ XY, por lo que la
hipótesis m ás parsim oniosa consist ir ía en hipotet izar que este t ipo de
sistem a de determ inación sexual sería ancest ral para la fam ilia y, por ende,
para Alouat ta.

5. Discusión
182
Como posible explicación para el pat rón de determ inación del sexo
observado en aulladores, teniendo en cuenta las evidencias previas y los
resultados obtenidos en esta Tesis Doctoral, se puede considerar que las
especies de Alouat ta poseían ancest ralm ente un sistem a crom osómico de
determ inación sexual XX/ XY, previo a la separación biogeográfica de los
grupos m eso y sudam ericanos. Luego de esta separación (que según el
escenario biogeográfico asum ido como correcto podría haber ocurr ido como
m ínimo hace 3 m illones años o antes, si tenem os en cuenta los 18 m illones
de años antes refer idos) am bos grupos habrían adquir ido
independientem ente los sistemas sexuales m últ iples que se observan en la
actualidad m ediante el m ecanismo de t ranslocaciones Y-autosom a
independientes.
Una de las hipótesis de t rabajo de esta Tesis Doctoral or ientó el
análisis considerando si el sistema de determ inación sexual ancest ral para
el género fuera de t ipo X1X2Y. De los resultados obtenidos hasta el
m om ento de la redacción del t rabajo de Tesis (presencia o ausencia de
conservación de las sintenias, ausencia de secuencias telom éricas
interst iciales y hom eologías crom osómicas por bandas G y FI SH) se pudo
inferir un posible m ecanismo para la form ación de estos sistemas sexuales.
Los sistem as de determ inación sexual m últ iple de t ipo X1X2Y habrían
surgido de form a independiente en los linajes de aulladores m eso y
sudam ericanos m ediante una t ranslocación Y-autosom a (Figura 5.4) .
Dos fisiones, una en Yp ter y ot ra en la región proxim al del brazo q del
autosom a, seguidas de una t ranslocación de Yq-pprox a la zona proximal del
brazo q, form aron el nuevo crom osom a Y1. El segm ento Yp ter y la región
proxim al del autosoma fisionado se perderían o en ciertas especies darían
origen a m icrocrom osoma. El hom ólogo del par autosóm ico involucrado en
la t ranslocación se t ransform aría en X2 (Figura 5.4) . En el caso de los
aulladores sudam ericanos, el par autosóm ico involucrado tendría
hom eología con HSA3, m ient ras que en los mesoam ericanos presentaría
hom eología con HSA7.
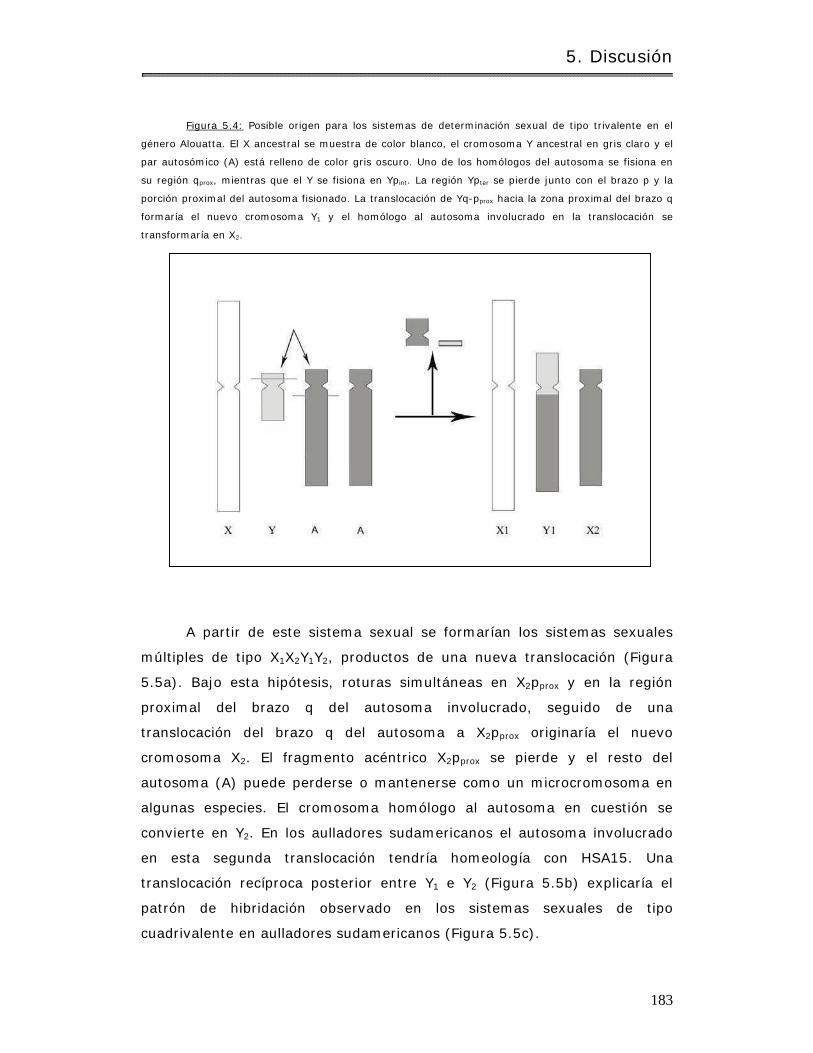
5. Discusión
183
Figura 5.4: Posible or igen para los sistem as de determ inación sexual de t ipo t r ivalente en el
género Alouat ta. El X ancest ral se muest ra de color blanco, el cromosom a Y ancest ral en gr is claro y el
par autosóm ico (A) está relleno de color gr is oscuro. Uno de los hom ólogos del autosoma se fisiona en
su región qprox, m ient ras que el Y se fisiona en Yp int. La región Yp ter se pierde junto con el brazo p y la
porción proxim al del autosom a fisionado. La t ranslocación de Yq-pprox hacia la zona proxim al del brazo q
form aría el nuevo crom osom a Y1 y el homólogo al autosoma involucrado en la t ranslocación se
t ransform aría en X2.
A part ir de este sistem a sexual se form arían los sistem as sexuales
m últ iples de t ipo X1X2Y1Y2, productos de una nueva t ranslocación (Figura
5.5a) . Bajo esta hipótesis, roturas sim ultáneas en X2pprox y en la región
proxim al del brazo q del autosom a involucrado, seguido de una
t ranslocación del brazo q del autosom a a X2pprox or iginaría el nuevo
crom osom a X2. El fragm ento acént r ico X2pprox se pierde y el resto del
autosom a (A) puede perderse o m antenerse com o un m icrocromosom a en
algunas especies. El crom osom a hom ólogo al autosom a en cuest ión se
convierte en Y2. En los aulladores sudam ericanos el autosom a involucrado
en esta segunda t ranslocación tendría hom eología con HSA15. Una
t ranslocación recíproca poster ior ent re Y1 e Y2 (Figura 5.5b) explicaría el
pat rón de hibridación observado en los sistem as sexuales de t ipo
cuadrivalente en aulladores sudam ericanos (Figura 5.5c) .

5. Discusión
184
Figura 5.5: Posible or igen para los sistem as de determ inación sexual de t ipo cuadrivalente en el
género Alouat ta. El X ancest ral se m uest ra de color blanco, el crom osom a Y ancest ral en gris claro y los
pares autosóm icos (A) están rellenos de color negro. a) Roturas simultáneas en X2pprox y en la región
proxim al del brazo q del autosom a involucrado, seguido de una t ranslocación del brazo q del autosom a a
X2pprox or iginaría el nuevo crom osom a X2. El fragm ento acént r ico X2pprox se pierde y el resto del
autosoma (A) puede perderse o m antenerse com o un m icrocrom osoma en algunas especies. El
crom osom a hom ólogo al autosom a en cuest ión se ident if ica com o Y2. b) Una t ranslocación recíproca
posterior ent re Y1 e Y2 explicaría el pat rón de hibr idación observado en los sistem as sexuales de t ipo
cuadrivalente en aulladores sudam ericanos. c) Pat rón de hibr idación en aulladores sudam ericanos
ejem plif icado en ACA.
Sin em bargo, no se puede descartar la posibilidad del origen
independiente de los sistem as sexuales de t ipo t r ivalente y cuadrivalente, a
part ir del m ismo par autosóm ico, dent ro de los grupos m eso y

5. Discusión
185
sudam ericanos. Si bien no es la hipótesis m ás parsim oniosa, en vista de la
evidencia observada respecto al or igen independiente de los m ult ivalentes
ent re estos dos grupos, no se puede descartar esta hipótesis.
5 .4 .4 Perspect ivas
La Sistem át ica del Orden Primates es objeto de cont inuo debate y en
la últ im a década dist intas revisiones han cont r ibuido con nuevos elem entos
de análisis a su posible esclarecim iento en búsqueda de un consenso.
Los resultados obtenidos en este t rabajo de Tesis Doctoral
cont r ibuyen con nueva inform ación de ut ilidad en la sistem át ica del género
Alouat ta, a la vez que respaldan la hipótesis de la evolución cromosómica
com o est rategia especiogénica en Prim ates.
Com o se m encionó en la I nt roducción (sección 1.2.3 página 17) , las
hom eologías propuestas m ediante pat rones de bandas G se ven lim itadas
en el caso de cariot ipos m uy reordenados. En este t rabajo de Tesis se logró
caracterizar m ediante FI SH, en APA y API , la ausencia de conservación de
dos sintenias, la 3/ 15 y la 3/ 21, y, por ende, confirm ar las hom eologías
crom osóm icas asignadas a esos crom osom as hum anos, com plem entando
caracterizaciones previas en ot ros aulladores. El disponer de financiación
suficiente para acceder a la batería com pleta de sondas de crom osom as
enteros humanos perm it irá cont inuar estos t rabajos y refinar la
caracterización de las hom eologías.
Cabe destacar que este es el pr im er estudio filogenét ico ut ilizando
análisis com binado de caracteres crom osóm icos y m oleculares en
ceboideos. A la luz de los datos obtenidos se postularon hipótesis para
explicar el pat rón de sistem as de determ inación sexual en el género
Alouat ta.

5. Discusión
186
Sin em bargo, aún existen especies no caracter izadas
citogenét icam ente por análisis m eiót icos (como por ejem plo A. guariba) .
Una reconst rucción de la posible secuencia de reordenam ientos Y-autosom a
en m onos aulladores requiere un m ayor núm ero de estudios citogenét icos
que abarquen las dist intas especies del género y que incluyan estudios
m eiót icos confirm ator ios de la cariología descrita por m itosis.
La realización de estudios m eiót icos en especies de éste y ot ros
géneros posibilitaría una m ejor com prensión del pat rón de evolución de los
crom osom as sexuales en Prim ates ceboideos, a la vez que cont r ibuiría a su
esclarecim iento sistem át ico colaborando en un avance del conocim iento
taxonóm ico de estos mamíferos superiores.

CONCLUSI ONES

6. Conclusiones
187
6 . Conclusiones
Teniendo en cuenta los objet ivos generales y específicos planteados para el
desarrollo de esta Tesis Doctoral, los hallazgos reunidos a la fecha
perm it ieron arr ibar a las presentes conclusiones:
Ü La caracter ización citogenét ica mediante análisis m itót icos y
m eiót icos perm it ió la correcta asignación de especie en todos los
ejemplares de prim ates analizados.
Ü Se descr ibió por prim era vez el cariot ipo del mono aullador negro,
Alouat ta pigra.
Ü La sintenia 3/ 21, ancest ral para m amíferos, se encontraría rota en
API y APA.
Ü La sintenia 3/ 15, asociada a los sistem as de determ inación sexual en
los aulladores sudam ericanos (con excepción de ABE) , se encontraría
rota en API y APA.
Ü Con excepción de Aotus azarae no se encont raron secuencias
telom éricas interst iciales en los ceboideos analizados.
Ü La diagnosis m eiót ica em pleando la técnica de inmunofluorescencia
perm it ió dem ostrar que el com portam iento del bivalente XY de
C. libidinosus, previam ente considerado com o de t ipo “hum an like” ,
presenta m arcadas diferencias respecto al XY hum ano.
Ü La frecuencia m edia de foci de MLH1 en CLI y en ACA no difiere sin
em bargo, el valor m edio obtenido para ACA está subest im ado al no
analizar el cuadrivalente sexual por su com plejo plegam iento
característ ico.

6. Conclusiones
188
Ü El análisis con la sonda com ercial para el gen SRY no m ost ró señal
posit iva de hibridación en Ceboidea, perm it iendo sugerir que las
secuencias adyacentes al gen SRY en la sonda no estarían
conservadas en estos pr im ates.
Ü Los autosom as involucrados en las t ranslocaciónes Y-autosoma que
dieron or igen a los m ult ivalentes sexuales en las especies
sudam ericanas y m esoam ericanas serían dist intos, indicando un
origen independiente de estos sistem as de determ inación sexual en
am bos grupos de aulladores.
Ü El análisis com binado de var iables m oleculares y cromosóm icas
perm it ió resolver las relaciones de parentesco ent re las especies de
aulladores meso y sudam ericanas, dem ostrando su ut ilidad en los
estudios sistem át icos y evolut ivos.
Ü Las dist intas var iables analizadas apoyan la necesidad de emplear
análisis filogenét icos com binados dent ro de un m arco de “Evidencia
Total” .

BI BLI OGRAFÍ A

7. Bibliografía
189
Agost ini, I . ; Holzm ann, I . ; Di Bitet t i, M.S. 2008. I nfant hybr ids in a newly form ed m ixed-species group of howler m onkeys (Alouat ta guariba clam itans and Alouat ta caraya) in northeastern Argent ina. Prim ates 49 (10) : 304-307.
Aguiar,L.M.; Pie, M.R.; Passos, F.C. 2007. Wild m ixed groups of howler species (Alouat ta caraya and Alouat ta clam itans) and new evidence for their hybridizat ion. Prim ates 49 (10) : 149-152.
Am aral, P.J.S.; Finotelo, L.F.M; de Oliveira, E.H.C.; Pissinat t i, A.; Nagam achi, C.Y.; Pieczarka J.C. 2008. Phylogenet ic studies of the genus Cebus (Cebidae-Prim ates) using chromosom e paint ing and G-banding. BMC Evolut ionary Biology 8: 169.
Archidiacono, N.; Storlazzi, C.T.; Spalluto, C.; Ricco, A.S.; Marzella, R.; Rocchi, M. 1998. Evolut ion of chrom osom e Y in prim ates. Chrom osom a 107 (9) : 241-246.
Arm ada, J.L.A.; Barroso, C.M.L.; Lim a, M.M.C.; Muniz, J.A.P.C.; Seuánez, H. 1987. Chrom osom e Studies in Alouat ta belzebul. Am erican Journal of Pr im atology 13: 283-296.
Baker, S.M.; PlugAnnem ieke, W.; Prolla, T.A.; Bronner, C.E.; Harr is, A.C.; Yao, X.; Christ ie, D.M. et al. 1996. I nvolvem ent of m ouse Mlh1 in DNA m ism atch repair and m eiot ic crossing over. Nature Genet ics 13: 336-342.
Barroso, C.M.L.; Schneider, H.; Schneider, M.P.C. Sam paio, I . ; Harada, M.L.; Czelusniak, J.; Goodm an, M. 1997. Update on the Phylogenet ic System at ics of New World Monkeys: Further DNA Evidence for Placing the Pygm y Marmoset Cebuella within the Genus Callithr ix . I ntenat ional Journal of Pr im atology 18: 651-674.
Barske, L.A.; Capel, B. 2008. Blurr ing the edges in vertebrate sex determ inat ion. Current Opinion in Genet ics & Developm ent 18: 499-505.
Bianchi, N.O.; Cont reras, J.R. 1967. The Chrom osom es of the Field Mouse Akodon azarae (Cr icet idae, Rodent ia) with Special Reference to Sex Chrom osom e Anom alies. Cytogenet ic and Genom e Research 6: 306-313.
Bigoni, F.; Koehler, U.; Stanyon, R.; I shida, T.; Wienberg, J. 1997. Fluorescence in situ hybridizat ion establishes hom ology between human and silvered leaf monkey chromosom es, reveals reciprocal t ranslocat ions between chrom osom es homologous to hum an Y/ 5, 1/ 9, and 6/ 16, and delineates an X1X2Y1Y2/ X1X1X2X2 sex-chrom osom e system . Am erican Journal of Physical Anthropology 102 (3) : 315-327.
Boehringer Mannheim . 1996. Nonradioact ive in situ Hybr idizat ion Applicat ion Manual, 2nd Edit ion. 214 pags.
Bonvicino, C.R.; Lem os, B.; Seuánez, H. 2001. Molecular Phylogenet ics of howler m onkeys (Alouat ta, Platyrrhini) a comparison with karyotypic data. Chrom osom a 110: 241-246.
Bourne, G.H. 1969. The chimpanzee; a series of volum es on the chim panzee, edited by G. H. Bourne. Basel, New York,: S. Karger,

7. Bibliografía
190
Basel, New York, S. Karger.
Boveri, T.H. 1904. Ergebnisse über die Konst itut ion der chrom at ischen Substanz des Zellkerns. Fisher, Jena.
Bull, J.J. 1983. Evolut ion of Sex Determ ining Mechanism s. Benjam in-Cum mings Pub Co. 316 pags.
Cabrera, A. 1958. Catálogo de los mamíferos de Am érica del Sur. Revista del Museo Argent ino de Ciencias Naturales I V, no. 1. Ciencias Zoológicas.
Calderón, P.L.; Pigozzi, M.I . 2006. MLH1- focus m apping in birds shows equal recombinat ion between sexes and diversity of crossover pat terns. Chrom osom e Research 14 (9) : 605-612.
Carvalho, A.B.; Koerich, L.B.; Clark, A.G. 2009. Origin and evolut ion of Y chrom osom es: Drosophila tales. Trends in Genet ics 25: 270-277.
Casado, F.; Bonvicino, C.R.; Nagle, C.; Com as, B.; Manzur, T.D.; Lahoz, M.M.; Seuánez, H.N. 2010. Mitochondrial Divergence Between 2 Populat ions of the Hooded Capuchin, Cebus (Sapajus) cay (Platyrrhini, Prim ates) . Journal of Heredity 101: 261 -269.
Chandley, A.C.; Cooke, H.J. 1994. Hum an male fert ilit y- -Y- linked genes and sperm atogenesis. Hum an Molecular Genet ics 3 Spec No: 1449-1452.
Charlesworth, D. 2002. Plant sex determ inat ion and sex chrom osom es. Heredity 88: 94-101.
Charlesworth, D.; Mank, J.E. 2010. The Birds and the Bees and the Flowers and the Trees: Lessons from Genet ic Mapping of Sex Determ inat ion in Plants and Anim als. Genet ics 186: 9-31.
Chu, E.H.Y.; Bender, M.A. 1962. Cytogenet ics and Evolut ion of Prim ates. Annals of the New York Academ y of Sciences 102: 253-266.
Clem ente, I .C.; Ponsà, M.; García, M.; Egozcue, J. 1990. Evolut ion of the Sim iiform es and the phylogeny of hum an chrom osom es. Hum an Genet ics 84 (5) : 493-506.
Coates, A.G.; Collins, L.S.; Aubry, M.P.; Berggren, W.A. 2004. The Geology of the Darien, Panam a, and the late Miocene-Pliocene collision of the Panam a arc with northwestern South Am erica. Geological Society of Am erica Bullet in 116: 1327-1344.
Codina-Pascual, M.; Cam pillo, M.; Kraus, J.; Speicher, M.R.; Egozcue, J.; Navarro, J.; Benet , J. 2006. Crossover frequency and synaptonem al com plex length: their variabilit y and effects on hum an m ale m eiosis. Molecular Hum an Reproduct ion 12: 123 -133.
Codina-Pascual, M.; Oliver-Bonet , M.; Navarro, J.; Cam pillo, M.; García, F.; Egozcue, S.; Abad, C.; Egozcue, J.; Benet J. 2005. Synapsis and m eiot ic recom binat ion analyses: MLH1 focus in the XY pair as an indicator. Hum an Reproduct ion 20: 2133 -2139.
Consigliere, S.; Stanyon, R.; Koehler, G.; Agoramoorthy, G; Wienberg, J. 1996. Chrom osom e paint ing defines genom ic rearrangem ents between red howler monkey subspecies. Chrom osom e Research 4: 264-270.

7. Bibliografía
191
Consigliere, S.; Stanyon, R.; Koehler, U.; Arnold, N.; Wienberg, J. 1998. I n situ hybridizat ion (FI SH) m aps chrom osomal hom ologies between Alouat ta belzebul (Platyrrhini, Cebidae) and other pr im ates and reveals extensive interchromosom al rearrangem ents between howler m onkey genom es. Am erican Journal of Prim atology 46: 119-133.
Cook, J.M.; I an, C.W. 2002. Sex determ inat ion in I nvertebrates. En Sex rat ios: concepts and research m ethods, ed. Hardy, I . I .C.W. Cam bridge University Press. 438 pags.
Cortés-Ort iz, L.; Berm ingham, E.; Rico, C.; Rodríguez-Luna, E.; Sam paio, I . ; Ruiz-García, M. 2003. Molecular systemat ics and biogeography of the Neot ropical m onkey genus, Alouat ta. Molecular Phylogenet ics and Evolut ion 26: 64-81.
Cortés-Ort iz, L.; Duda, T.F.; Canales-Espinosa, D.; García-Orduña, F.; Rodríguez-Luna, E.; Berm ingham , E. 2007. Hybridizat ion in Large-Bodied New World Primates. Genet ics 176: 2421–2425.
Coyne, J.A.; Orr, H.A. 1989. Pat terns of Speciat ion in Drosophila. Evolut ion 43: 362-381.
———. 2004. Speciat ion. 1º ed. Sinauer Associates, I nc., Mayo 1.
Crocket t , C.; Eisenberg, J. 1987. Howlers: var iat ions in group size and dem ography. En Prim ate societ ies. CHicago I L, USA: University of Chicago Press.
Deuve, J.L.; Bennet t , N.C.; Ruiz-Herrera, A.; Waters, P.D.; Br it ton-Davidian, J.; Robinson, T.J. 2007. Dissect ion of a Y-autosom e t ranslocat ion in Cryptom ys hot tentotus (Rodent ia, Bathyergidae) and implicat ions for the evolut ion of a m eiot ic sex chromosom e chain. Chrom osom a 117 (12) : 211-217.
de Oliveira, E.H.C. 1995. Chromosom al variat ion in Alouat ta fusca. Neotropical Prim ates 3 (4) : 181-183.
de Oliveira, E.H.C.; de Lim a, M.M.C.; Sbalqueiro, I .J.; Pissinat i, A. 1998. The karyotype of Alouat ta fusca clam itans from Rio de Janeiro, Brazil: Evidence for a y-autosom e t ranslocat ion. Genet ics and Molecular Biology 21, no. 3 (9) .
de Oliveira, E.H.C.; Neusser, M.; Figueiredo, W.B.; Nagam achi, C.; Pieczarka, J.C.; Sbalqueiro, I .J.; Wienberg, J.; Müller, S. 2002. The phylogeny of howler m onkeys (Alouat ta, Platyrrhini) : Reconst ruct ion by m ult icolor cross-species chrom osom e paint ing. Chrom osom e Research 10: 669-683.
de Oliveira, E.H.C.; Neusser, M.; Pieczarka, J.C.; Nagam achi, C.; Sbalqueiro, I .J.; Müller, S. 2005. Phylogenet ic inferences of Atelinae (Platyrrhini) based on mult i-direct ional chromosom e paint ing in Brachyteles arachnoides, Ateles paniscus paniscus and Ateles b. m arginatus. Cytogenet ic and Genom e Research 108: 183-90.
de Vries, F.A.T.; de Boer, E.; van den Bosch, M.; Baarends, W.M.; Ooms, M.; Yuan, L.; Liu, J.G.; van Zeeland, A.A.; Heyt ing, C.; Past ink, A. 2005. Mouse Sycp1 funct ions in synaptonem al com plex assem bly, m eiot ic recombinat ion, and XY body form at ion. Genes & Developm ent

7. Bibliografía
192
19: 1376 -1389.
Devlin, R.H.; Nagaham a, Y. 2002. Sex determ inat ion and sex different iat ion in fish: an overview of genet ic, physiological, and environm ental influences. Aquaculture 208: 191-364.
Dobigny, G.; Ducroz, J.F.; Robinson, T.J.; Volobouev, V. 2004. Cytogenet ics and Cladist ics. System at ic Biology 53: 470 -484.
Dum as, F.; Stanyon, R.; Sineo, L.; Stone, G.; Bigoni, F. 2007. Phylogenom ics of species from four genera of New World m onkeys by flow sort ing and reciprocal chrom osom e paint ing. BMC Evolut ionary Biology 7, Suppl 2: S11.
Dut r illaux, B. 1979. Very large analogy of chrom osom e banding between Cebus capucinus (Platyrrhini) and m an. Cytogenet ics and Cell Genet ics 24: 84-94.
Egozcue J. 1969. Aneuploidía cromosóm ica: nuevas observaciones sobre la separación precoz de bivalentes en m eiosis I . Sangre XI V, no. 4: 442-452.
Egozcue, J.; de Egozcue, V. 1966. The Chrom osom e Com plem ent of the Howler Monkey (Alouat ta caraya, HUMBOLDT 1812) . Cytogenet ic and Genom e Research 5: 20-27.
Ehrlich, J.; Sankoff, D.; Nadeau, J.H. 1997. Synteny Conservat ion and Chrom osom e Rearrangem ents During Mamm alian Evolut ion. Genet ics 147: 289-296.
Ellsworth, J.A.; Hoelzer, G.A. 2006. Genet ic Evidence on the Historical Biogeography of Cent ral Am erican Howler Monkeys. En Prim ate biogeography: progress and prospects, ed. Shawn M. Lehm an y John G. Fleagle, 81-103. Springer.
Erickson, J.W.; Quintero, J.J. 2007. I ndirect Effects of Ploidy Suggest X Chrom osom e Dose, Not the X: A Rat io, Signals Sex in Drosophila. PLoS Biol 5 (12) : e332.
Evans, E.P.; Breckon, G.; Ford, C.E. 1964. An Air-Drying Method for Meiot ic Preparat ions from Mamm alian Testes. Cytogenet ic and Genom e Research 3: 289-294.
Ezaz, T.; St iglec, R.; Veyrunes, F.; Marshall Graves, J.A. 2006. Relat ionships between Vertebrate ZW and XY Sex Chrom osom e System s. Current Biology 16: R736-R743.
Fant ini, L. 2010. Hibr idación Genóm ica Com parada en el estudio evolut ivo de Cebus (Primates, Platyrrhini) : Análisis de una progenie híbrida. Tesis de Grado, Buenos Aires - Argent ina: Universidad de Buenos Aires.
Fleagle, J. G. 1999. Prim ate Adaptat ion and Evolut ion. Academ ic Press. 596 pags.
Ferguson-Sm ith, M.A.; Trifonov, V. 2007. Mam m alian karyotype evolut ion. Nature Reviews in Genet ics 8: 950-962.
Forej t , J. 1996. Hybrid ster ility in the m ouse. Trends in Genet ics 12: 412-417.

7. Bibliografía
193
Fredga, K. 1970. Unusual Sex Chrom osom e I nheritance in Mam m als. Philosophical Transact ions of the Royal Society of London. Series B, Biological Sciences 259 (828) : 15-36.
García, M.; Borrell, A.; Mudry, M.D.; Egozcue, J.; Ponsá, M. 1995. Prom etaphase Karyotype and Rest r ict ion-Enzym e Banding in Squirrel Monkeys, Saim ir i boliv iensis boliviensis (Pr imates: Platyrrhini) . Journal of Mamm alogy 76: 497-503.
García-Haro, F. 2001. Evolución crom osómica en Sim iiformes. Hom ologías, Reorganizaciones y Heterocrom at ina. Tesis Doctoral, Universidad Autónom a de Barcelona, España.
García, F.; Nogués, C.; Ponsà, M.; Ruiz-Herrera, A.; Egozcue, J.; Garcia Caldés, M. 2000. Chrom osom al hom ologies between hum ans and Cebus apella (Prim ates) revealed by ZOO-FI SH. Mamm alian Genom e 11: 399-401.
García, F.; Ruiz-Herrera, A.; Egozcue, J.; Ponsà, M.; Garcia, M. 2002. Chrom osomal hom ologies between Cebus and Ateles (Primates) based on ZOO-FI SH and G-banding comparisons. Am erican Journal of Pr im atology 57: 177-188.
Garcia-Cruz, R.; Robles, P.; Steinberg, E.R.; Cam ats, N.; Brieño, M.A.; Garcia-Caldés, M.; Mudry, M.D. 2009. Pair ing and recom binat ion features during m eiosis in Cebus paraguayanus (Prim ates: Platyrrhini) . BMC Genet ics 10: 25.
Gläser, B.; Grützner, F.; Willm ann, U.; Stanyon, R.; Arnold, N.; Taylor, K.; Rietschel, W.; Zeit ler, S.; Toder, R.; Schempp, W. 1998. Sim ian Y Chrom osom es: species-specific rearrangem ents of DAZ, RBM, and TSPY versus cont iguity of PAR and SRY. Mamm alian Genom e 9 (3) : 226-231.
Graves, J.A.M., O'Neill, R.J.W. 1997. Sex Chromosom e Evolut ion and Haldane's Rule. Journal of Heredity 88, no. 5: 358 -360.
Graves, J.A.M. 2006. Sex Chrom osom e Specializat ion and Degenerat ion in Mam m als. Cell 124: 901-914.
———. 2008. Weird Animal Genom es and the Evolut ion of Vertebrate Sex and Sex Chrom osom es. Annual Review of Genet ics 42 (12) : 565-586.
Graves, J.A.M.; Westerm an, M. 2002. Marsupial genet ics and genom ics. Trends in Genet ics 18: 517-521.
Gregorin, R. 2006. Taxonom ia e variação geográfica das espécies do gênero Alouat ta Lacépède (Prim ates, Atelidae) no Brasil. Revista Brasileira de Zoologia 23 (1) : 64-144
Groves, C.P. 2001. Prim ate Taxonom y . USA & England: Sm ithsonian I nst itut ion Press.
Groves, C. 2005. Order Prim ates. En Mam m al species of the world: a taxonom ic and geographic reference, ed. D. E. Wilson y DeeAnn M. Reeder. Vol. 1. 2142 pags.
Haldane, J.B.S. 1922. Sex rat io and unisexual ster ility in hybrid anim als. Journal of Genet ics 12 (10) : 101-109.

7. Bibliografía
194
Harada, M.L.; Schneider, H.; Schneider, M.P.C.; Sam paio, I . ; Czelusniak, J.; Goodm an, M. 1995. DNA Evidence on the Phylogenet ic System at ics of New World Monkeys: Support for the Sister-Grouping of Cebus and Saim ir i from Two Unlinked Nuclear Genes. Molecular Phylogenet ics and Evolut ion 4: 331-349.
Harr is, E.E.; Gifalli- I ughet t i, C.; Hirano Braga, Z.; Koiffm ann, C.P. 2005. Cytochrom e B Sequences Show Subdivision between Populat ions of the Brown Howler Monkey (Alouat ta guar iba) from Rio de Janeiro and Santa Catar ina, Brazil. Neot ropical Prim ates 13 (2) : 16.
Hassold, T.; Judis, L.; Chan, E.R.; Schwartz, S.; Seftel, A.; Lynn, A. 2004. Cytological studies of m eiot ic recombinat ion in hum an males. Cytogenet ic and Genom e Research 107: 249-255.
Hennig, W. 1966. Hennig's Phylogenet ic System at ics. 1966 Edit ion. Univ of I llinois.
Hershkovitz, P. 1977. Living New World Monkeys (Platyrrhini) with an I nt roduct ion to Pr im ates. Chicago, USA: Chicago University Press. 1132 pags.
Hershkovitz, P. 1984. Taxonom y of squirrel m onkeys genus Saim ir i (Cebidae, Platyrrhini) : A prelim inary report with descript ion of a hitherto unnam ed form . Am erican Journal of Pr im atology 7: 155-210.
Hill, W.C.O. 1962. Prim ates. Comparat ive Anatom y and Taxonom y V. Cebidae Part B. Edinburgo: Edinburgh University Press, Edinburgh. 537 pags.
Hirsch, A.; Dias, L.G.; Mart ins, L. de O.; Cam pos, R.F.; Resende, N.A.T.; Landau, E.C. 2002. “Database of Georreferenced Occurrence Localit ies of Neotropical Prim ates” . Departm ent of Zoology / UFMG, Belo Horizonte. ht tp: / / www.icb.ufm g.br/ ~ prim atas/ hom e_bdgeoprim .htm
Howell, W.M.; Black, D.A. 1980. Cont rolled silver-staining of nucleolus organizer regions with a protect ive colloidal developer: a 1-step m ethod. Experient ia 36 (8) : 1014-1015.
Howell, E.C.; Armstrong, S.J.; Filatov, D.A. 2009. Evolut ion of Neo-Sex Chrom osom es in Silene diclinis. Genet ics 182: 1109-1115.
Hsu, T.C.; Arr ighi, F.E.; Klevecz, R.R.; Br inkley, B.R. 1965. The nucleoli in m itot ic divisions of m am m alian cells in vit ro. The Journal of Cell Biology 26: 539-553.
Janzen, F.J.; Phillips, P.C. 2006. Explor ing the evolut ion of environm ental sex determ inat ion, especially in rept iles. Journal of Evolut ionary Biology 19 (11) : 1775-1784.
Just , W.; Baum stark, A.; Ham eister, H.; Schreiner, B.; Reisert , I . ; Hakhverdyan, M.; Vogel, W. 2002. The sex determ inat ion in Ellobius lutescens rem ains bizarre. Cytogenet ic and Genom e Research 96: 146-153.
Kaiser, V.B.; Bacht rog, D. 2010. Evolut ion of Sex Chrom osom es in I nsects. Annual Review of Genet ics 44 (12) : 91-112.
Kasahara, S.; Dut r illaux, B. 1983. Chromosom e banding pat terns of four species of bats, with special reference to a case of X-autosom e

7. Bibliografía
195
t ranslocat ion. Annales De Génét ique 26 (4) : 197-201.
Kawai, A.; Nishida-Um ehara, C.; I shij im a, J.; Tsuda, Y.; Ota, H.; Matsuda, Y. 2007. Different origins of bird and rept ile sex chrom osom es inferred from com parat ive mapping of chicken Z- linked genes. Cytogenet ic and Genom e Research 117: 92-102.
King, M. 1987. Chrom osom al rearrangem ents, speciat ion and the theoret ical approach. Heredity 59: 1-6.
King, M. 1993. Species Evolut ion: The Role of Chromosom e Change. Cam bridge University Press. 339 pags.
Kingswood, S.C.; Kum am oto, A.T.; Sudm an, P.D.; Fletcher, K.C.; Greenbaum, I .F. 1994. Meiosis in chrom osom ally heteromorphic goitered gazelle, Gazella subgut turosa (Art iodactyla, Bovidae) . Chrom osom e Research 2 (1) : 37-46.
Kluge, A.G. 1989. A Concern for Evidence and a Phylogenet ic Hypothesis of Relat ionships Am ong Epicrates (Boidae, Serpentes) . System at ic Zoology 38: 7-25.
Knowlton, N.; Weigt , L.A. 1998. New dates and new rates for divergence across the I sthm us of Panam a. Proceedings of the Royal Society B: Biological Sciences 265: 2257-2257.
Koerich, L.B.; Wang, X.; Clark, A.G.; Carvalho, A.B. 2008. Low conservat ion of gene content in the Drosophila Y chrom osom e. Nature 456: 949-951.
Koiffm ann, C.P.; Saldanha, P.H. 1974. Cytogenet ics of Brazilian monkeys. Journal of Hum an Evolut ion 3: 275-282.
Lacadena, J.R. 1996. Citogenét ica. Editorial Com plutense. 936 pags.
Lander, et al. 2001. I nit ial sequencing and analysis of the hum an genom e. Nature 409 (6822) : 860-921.
Laurie, C.C. 1997. The Weaker Sex is Heterogam et ic: 75 Years of HALDANE` s Rule. Genet ics 147: 937-951.
Lawrence, B. 1933. Howler m onkeys of the palliata group. Bullet in of the Museum of Comparat ive Zoology (Harvard University) 75: 314–354.
Levan, A.; Fredga, K.; Sandberg, A.A. 1964. Nom enclature for cent rom eric posit ion on chrom osom es. Hereditas 52: 201-220.
Lim a M.M.C.; Seuánez H. 1991. Chrom osome studies in the red Howler Monkey, Alouat ta seniculus st ram ineus (Platyrrhini, Prim ates) : descript ion of an X1X2Y1Y2/ X1X1X2X2 sex chrom osom e system and karyological com parasions with other subspecies. Cytogenet ics and Cell Genet ics 57: 151-156.
Lorenz, J.G.; Jackson, W.E.; Beck, J.C.; Hanner, R. 2005. The problem s and prom ise of DNA barcodes for species diagnosis of pr im ate biom aterials. Philosophical Transact ions of the Royal Society B: Biological Sciences 360: 1869 -1877.
Lynn, A.; Koehler, K.E.; Judis, L.; Chan, E.R.; Cherry, J.P.; Schwartz, S.; Seftel, A.; Hunt , P.A.; Hassold, T.J. 2002. Covariat ion of Synaptonem al Com plex Length and Mam m alian Meiot ic Exchange

7. Bibliografía
196
Rates. Science 296: 2222 -2225.
Ma N.S.F.; Jones, T.C.; Thorignton, R.W.; Miller, A.; Morgan L. 1975. Y-Autosom e Translocat ion in the Howler Monkey, Alouat ta palliata. Journal of Medical Pr im atology 4: 299-307.
Ma, N.S.F.; Elliot t , M.W.; Morgan, L.; Miller, A.; Jones, T.C. 1976. Translocat ion of Y chromosom e to an autosom e in the Bolivian owl m onkey, Aotus. Am erican Journal of Physical Anthropology 45 (9) : 191-201.
Matayoshi, T.; Seuánez, H.N.; Nasazzi, N. 1987. Heterochrom at ic variat ion in Cebus apella (Cebidae, Platyrrhini) of different geographic regions. Cytogenet ic and Cell Genet ics 44: 158-162.
Matsubara, K.; Tarui, H.; Toriba, M.; Yam ada, K.; Nishida-Um ehara, C.; Agata, K.; Matsuda, Y. 2006. Evidence for different or igin of sex chrom osom es in snakes, birds, and m am m als and step-wise different iat ion of snake sex chrom osom es. Proceedings of the Nat ional Academ y of Sciences 103: 18190 -18195.
Mayr, E. 1942. System at ics and the or igin of species. New York: Columbia University Press.
Meireles C.M.; Czelusniak J.; Ferrari S.; Cruz M.P.; Schneider H.; Goodm an, M. 1999. Phylogenet ic relat ionships am ong brazilian howler m onkeys, genus Alouat ta (Platyrrhini, Atelidae) , based on gam m a-globin pseudogene sequences. Genet ics and Molecular Biology 22: 337-344.
Mikkelsen, et al. 2005. I nit ial sequence of the chim panzee genom e and com parison with the hum an genom e. Nature 437 (9) : 69-87.
Minezawa, M.; Harada, M.; Jordan, O.C.; Borda, C.J.V. 1985. Cytogenet ics of Bolivian endem ic red howler m onkeys (Alouat ta seniculus sara) : accessory chromosom es and Y-autosom e t ranslocat ion- related num erical var iat ions. Research Report New World Monkeys 5: 7–16.
Ming, R.; Wang, J.; Moore, P.H.; Paterson, A.H. 2007. Sex chrom osom es in flowering plants. Am erican Journal of Botany 94: 141-150.
Moens, P.B.; Kolas, N.K.; Tarsounas, M.; Marcon, E.; Cohen, P.E.; Spyropoulos, B. 2002. The t im e course and chrom osomal localizat ion of recombinat ion- related proteins at m eiosis in the m ouse are com pat ible with models that can resolve the ear ly DNA-DNA interact ions without reciprocal recombinat ion. Journal of Cell Science 115: 1611-1622.
Mola, L.M.; Papeschi, A.G. 2006. Holokinet ic chrom osom es at a glance. Journal of Basic and Applied Genet ics 17: 17-33.
Moreira, M.A.M. 2002. SRY evolut ion in Cebidae (Platyrrhini: Prim ates) . Journal of Molecular Evolut ion 55: 92-103.
Morescalchi, M.; Schempp, W.; Consigliere, S.; Bigoni, F.; Wienberg, J. ; Stanyon, R. 1997. Mapping chromosom al homology between hum ans and the black-handed spider monkey by fluorescence in situ hybridizat ion. Chrom osom e Research 5: 527-536.
Moses, M.J. 1968. Synaptonemal Com plex. Annual Review of Genet ics 2 (12) : 363-412.

7. Bibliografía
197
Moura-Pensin, C.; Pieczarka, J.C.; Nagam achi, C.Y.; Pereira Carneiro Muniz, J.A.; do Carm o de Oliveira Br igido, M.; Pissinat i A.; Marinho, A.; de Souza Barros, R.M. 2001. Cytogenet ic relat ions among the genera of the subfam ily Pitheciinae (Cebidae, Prim ates) . Caryologia 54: 385-391.
Mudry, M.D.; Brieux, S.; Colillas, O.J. 1981. Estudio citogenét ico del m ono aullador Alouat ta caraya de la República Argent ina. Physis 40: 63-70.
Mudry de Pargam ent , M.D.; Slavustky, I . ; Br ieux de Salum , S. 1982. Estudios citogenét icos en Saim ir i sciureus de Boliv ia. Mendeliana 5 (2) : 81-90.
Mudry, M.D.; Labal de Vinuesa, M.L.; Colillas, O.J.; Brieux, S. 1984a. Banding pat terns of Alouat ta caraya. Revista Brasilera de Genét ica 7, no. 2: 373-379.
Mudry de Pargam ent , M.D.; Colillas, O.J.; Brieux de Salum , S. 1984b. The Aotus from northern Argent ina. Prim ates 25: 530-537.
Mudry, M.D. 1990. Cytogenet ic variabilit y within and across populat ions of Cebus apella in Argent ina. Folia Prim atologica 54: 206-16.
Mudry, M.D.; Ponsà, M.; Borrell, A.; Egozcue, J.; García, M. 1994. Prom etaphase chrom osom es of the howler monkey (Alouat ta caraya) : G, C, NOR, and rest r ict ion enzym e (RES) banding. Am erican Journal of Prim atology 33: 121-132.
Mudry, M.D.; Rahn, M.I .; Gorost iaga, M.; Hick, A.; Merani, M.S.; Solar i, A.J. 1998. Revised Karyotype of Alouat ta Caraya (Prim ates: Platyrrhini) Based on Synaptonem al Com plex and Banding Analyses. Hereditas 128: 9-6.
Mudry, M.D.; Rahn, M.I .; Solar i, A.J. 2001. Meiosis and chrom osom e paint ing of sex chrom osom e system s in Ceboidea. Am erican Journal of Prim atology 54: 65-78.
Mudry, M.D.; Nieves, M; Steinberg, E.R.; Zunino, G.E. 2006. Prim ates de Argent ina. En Mam íferos de Argent ina, 86-89. Argent ina: SAREM.
Müller, S. 2006. Pr imate Chrom osom e Evolut ion. En Genom ic Disorders, ed. Jam es R. Lupski y Pawel Stankiewicz, 133-152. Totowa, NJ: Hum ana Press.
Müller, S.; Stanyon, R.; Finelli, P.; Archidiacono, N.; Wienberg, J. 2000. Molecular cytogenet ic dissect ion of hum an chrom osom es 3 and 21 evolut ion. Proceedings of the Nat ional Academ y of Sciences 97: 206-211.
Nagarajan, S.; Rens, W.; Stalker, J.; Cox, T.; Ferguson-Sm ith, M. 2008. Chrom hom e: A r ich internet applicat ion for accessing comparat ive chromosom e hom ology m aps. BMC Bioinform at ics 9: 168.
Nakamura, M. 2009. Sex determ inat ion in amphibians. Seminars in Cell & Developm ental Biology 20: 271-282.
Nascim iento, F.F.; Bonvicino, C.R.; Da Silva, F.C.D.; Schneider, M.P.C.; Seuánez, H. 2005. Cytochrom e b polymorphisms and populat ion st ructure of two species of Alouat ta (Prim ates) . Cytogenet ic and Genom e Research 108: 106-111.

7. Bibliografía
198
Navajas-Pérez, R.; de la Herrán, R.; López González, G.; Jam ilena, M.; Lozano, R.; Ruiz Rejón, C.; Ruiz Rejón, M.; Garr ido-Ram os, M.A. 2005. The Evolut ion of Reproduct ive System s and Sex-Determ ining Mechanisms Within Rumex (Polygonaceae) I nferred from Nuclear and Chloroplast idial Sequence Data. Molecular Biology and Evolut ion 22: 1929 -1939.
Neusser, M.; Stanyon, R.; Bigoni, F.; Wienberg, J.; Müller, S. 2001. Molecular cytotaxonomy of New World m onkeys (Platyrrhini) - com parat ive analysis of five species by mult i-color chrom osom e paint ing gives evidence for a classificat ion of Callim ico goeldii within the fam ily of Callit r ichidae. Cytogenet ics and Cell Genet ics 94: 206-15.
Nieves, M. 2002. Cont r ibución de la Citogenét ica para la ident ificación de especies anim ales en caut iver io: el ejem plo de los Prim ates Neotropicales. Tesis de Grado, Buenos Aires - Argent ina: Universidad de Buenos Aires. 77 pags.
Nieves, M.; Mudry, M.D.; Copelli, S. 2003. Conservación del cromosoma Y humano en Ceboidea (Prim ates: Platyrrhini) . En Anales I V Jornadas Argent ino-Chilenas, SAG. Córdoba, p.52.
Nieves, M.; Ascunce, M.S.; Rahn, M.I .; Mudry, M.D. 2005. Phylogenet ic relat ionships among som e Ateles species: the use of chrom osom ic and molecular characters. Prim ates 46: 155-164.
Nieves, M.; Mendez, G.; Ort iz, A.; Mühlm ann, M.; Mudry, M.D. 2008. Karyological diagnosis of Cebus (Prim ates, Platyrrhini) in capt ivity: Detect ion of hybrids and m anagem ent program applicat ions. Anim al Reproduct ion Science 108: 66-78.
Nieves, M. 2007. Heterocrom at ina y Evolución Cromosóm ica en Prim ates Neotropicales. Tesis Doctoral, Universidad de Buenos Aires.
O'Brien, S.J.; Menot t i-Raym ond, M.; Murphy, W.J.; Nash, W.G.; Wienberg, J.; Stanyon, R.; Copeland, N.G.; Jenkins, N.A.; Wom ack, J.E.; Marshall Graves, J.A. 1999. The Prom ise of Comparat ive Genomics in Mam m als. Science 286: 458-481.
Ogata, M.; Ohtani, H.; I garashi, T.; Hasegawa, Y.; I chikawa, Y.; Miura, I . 2003. Change of the Heterogam et ic Sex From Male to Fem ale in the Frog. Genet ics 164: 613-620.
Ohno, S. 1967. Sex Chrom osom es and Sex-Linked Genes. Springer-Verlag. 192 pags.
Olsen, G. J., P. J. Waddell, D. M. Hillis, y Swofford. 1996. Molecular System at ics: 407-514.
Orr, H.A. 1997. HALDANE'S RULE. Annual Review of Ecology and System at ics 28 (11) : 195-218.
Papeschi, A.G.; Bressa, M.J. 2006. Evolut ionary cytogenet ics in Heteroptera. Journal of Biological Research 5: 3-21.
Pathak, S.; Lin, C.C. 1981. Synaptonem al com plex of the sex-autosom e t r ivalent in a m ale I ndian munt jac. Chrom osom a 82 (4) : 367-376.
Page, J.; Fuente, R.; Góm ez, R.; Calvente, A.; Viera, A.; Parra, M.T.;

7. Bibliografía
199
Santos, J.L.; Berríos, S.; Fernández-Donoso, R.; Suja, J.A.; Rufas, J.S. 2006. Sex chrom osom es, synapsis, and cohesins: a complex affair. Chrom osom a 115 (3) : 250-259.
Pigozzi, M.I . ; Solari, A.J. 1999. Equal frequencies of recom binat ion nodules in both sexes of the pigeon suggest a basic difference with eutherian mamm als. Genom e 42: 315-321.
———. 2000. Los crom osom as sexuales y la evolución de las aves. Ciencia Hoy 10 (56) .
———. 2005. The germ- line- rest r icted chromosom e in the zebra finch: recom binat ion in fem ales and elim inat ion in m ales. Chrom osom a 114 (10) : 403-409.
Presgraves, D.C. 2008. Sex chrom osom es and speciat ion in Drosophila. Trends in Genet ics 24: 336-343.
Radder, R.S., Quinn, A.E.; Georges, A.; Sarre, S.D.; Shine, R. 2008. Genet ic evidence for co-occurrence of chrom osom al and therm al sex-determ ining systems in a lizard. Biology Let ters 4: 176-178.
Rahn M.I .; Mudry, M.D.; Merani, M.S.; Solar i, A.J. 1996. Meiot ic behavior of the X1X2Y1Y2 quadrivalent of the Prim ate Alouat ta caraya Chrom osom e Research 4: 350-356.
Reig, O.A. 1984. Significado de los m étodos citogenét icos para la dist inción y la interpretación de las especies, con especial referencia a los mamíferos. Zoología XI I I , no. 3: 19-44.
Rens, W.; O'Brien, P.C.M.; Grützner, F.; Clarke, O.; Graphodatskaya, D.; Tsend-Ayush, E.; Tr ifonov, V.A. et al. 2007. The m ult iple sex chrom osom es of platypus and echidna are not com pletely ident ical and several share homology with the avian Z. Genom e Biology 8: R243-R243.
Renwick, J.H. 1971. Assignm ent and m ap-posit ioning of hum an loci using chrom osom al var iat ion. Annals of Hum an Genet ics 35 (7) : 79-97.
Richard, F.; Lom bard, M.; Dutr illaux, B. 1996. ZOO-FI SH Suggests a Complete Homology between Hum an and Capuchin Monkey (Platyrrhini) Euchrom at in. Genomics 36: 417-423.
Richard, F.; Dut r illaux, B. 1998. Origin of hum an chromosom e 21 and its consequences: a 50-m illion-year-old story. Chromosom e Research 6: 263-268.
Robinson, T.J. 2001. The comparat ive cytogenet ics of Afr ican sm all Mam m als in perspect ive. Status, t rends, and bibliography. En Proceedings of the Afr ican Sm all Mam m als Sym posium , 185-214. París: I RD Edit ions.
Robinson, T.J.; Ruiz-Herrera, A. 2008. Defining the ancest ral eutherian karyotype: A cladist ic interpretat ion of chrom osom e paint ing and genom e sequence assem bly data. Chrom osom e Research 16 (12) : 1133-1141.
———. 2010. Mam m alian Chromosom al Evolut ion: From Ancest ral States to Evolut ionary Regions. En Evolut ionary Biology - Concepts, Molecular and Morphological Evolut ion, ed. Pierre Pontarot t i, 143-158. Berlin,

7. Bibliografía
200
Heidelberg: Springer-Verlag.
Rodrigues Norohna R.C.; Nagam achi, C.Y.; Pieczarka, J.C.; Marques-Aguiar, S., Lim a de Asis, M.F.; de Souza Barros, R.M. 2004. Meiot ic analyses of the sex chrom osom es in Carolliinae-Phyllostom idae (Chiroptera) : NOR separates the XY1Y2 into two independent parts. Caryologia 57: 1-9.
Roig, I . ; Liebe, B.; Egozcue, J.; Cabero, Ll. ; García, M.; Scherthan, H. 2004. Fem ale-specific features of recom binat ional double-st randed DNA repair in relat ion to synapsis and telom ere dynam ics in hum an oocytes. Chromosom a 113 (7) : 22-33.
Rosenberger A.L.; Tejedor, M.F. 2011. The Misbegot ten: Long Lineages, Long Branches and the I nterrelat ionships of Aotus, Callicebus and the sakiuacaris. En Evolut ionary Biology and Conservat ion of t it is, sakis and uakaris. A.A. Barnet t , L.M. Veiga, S.F. Ferrar i, and M.A. Norconk. Cam bridge University Press. En prensa.
Ross, J.A.; Peichel, C.L. 2008. Molecular Cytogenet ic Evidence of Rearrangem ents on the Y Chromosom e of the Threespine St ickleback Fish. Genet ics 179: 2173-2182.
Rowe, N. 1996. The Pictorial Guide to the Living Prim ates. East Ham pton, New York: Pogonias Press.
Ruiz-Herrera, A; García, F.; Mora, L.; Egozcue, J.; Ponsà, M.; García, M. 2005. Evolut ionary conserved chrom osom al segm ents in the hum an karyotype are bounded by unstable chrom osom e bands. Cytogenet ic and Genom e Research 108: 161-74.
Rylands, A.B.; Schneider, H.; Langguth, A.; Mit term eier, R.A.; Groves, C.P.; Rodrìguez-Luna, E. 2000. An assessm ent of the diversity of New World Primates. Neot ropical Prim ates 8 (2) : 61-93
Rylands, A.B.; Mit term eier, R.A. 2009. The Diversity of the NewWorld Prim ates (Platyrrhini) : An Annotated Taxonom y. En South Am erican Prim ates: Comparat ives Perspect ives in the study of behavior ecology, and conservat ion, 23-54.
Sanchez, L. 2008. Sex-determ ining m echanisms in insects. The I nternat ional Journal of Developm ental Biology 52 (7) : 837-856.
Sankoff, D. 2003. Rearrangem ents and chrom osom al evolut ion. Current Opinion in Genet ics & Developm ent 13: 583-587.
Schneider, H.; Rosenberger, A.L. 1996. Molecules, morphology and Platyrrhine system at ics. En Adapt ive radiat ions of neot ropical pr im ates, 3-19. Springer.
Schneider, H.; Schneider, M.P.; Sam paio, M.I .; Montoya, E.; Tapia, J.; Encarnación, F.; Anselm o, S.P.; Salzano, F.M. 1993. Divergence between biochem ical and cytogenet ic differences in three species of the Callicebus m oloch group. Am erican Journal of Physical Anthropology 90: 345-50.
Schneider, H. 2000. The current status of the New World m onkey phylogeny. Anais Da Academ ia Brasileira De Ciências 72 (2) : 165-72.
Schneider, H.; Canavez, F.C.; Sam paio, I . ; Moreira, M.A.M.; Tagliaro, C.H.;

7. Bibliografía
201
Seuánez, H.N. 2001. Can m olecular data place each neot ropical monkey in it s own branch? Chrom osom a 109: 515-523.
Schwarzacher, Dr. 2000. Pract ical in situ Hybridizat ion . Garland Science.
Seabright , M. 1971. A rapid banding technique for hum an chrom osom es. Lancet 2 (7731) : 971-2.
Seuánez, H.N.; Arm ada, J.L.; Barroso, C.; Rezende, C.; da Silva, V.F.. 1983. The m eiot ic chrom osom es of Cebus apella (Cebidae, Platyrrhini) . Cytogenet ic and Genom e Research 36: 517-524.
Seuánez, H.N.; Forman, L.; Matayoshi, T.; Fanning, T.G. 1989. The Callim ico goeldii (Pr im ates, Platyrrhini) genom e: Karyology and m iddle repet it ive (LI NE-1) DNA sequences. Chrom osom a 98: 389-395.
Sharp, P. 1982. Sex chromosom e pair ing during m ale m eiosis in marsupials. Chrom osom a 86: 27-47.
Simpson, G.G. 1945. The pr inciples of classificat ion and a classificat ion of mamm als. Bullet in of the Am erican Museum of Natural History 85: 1-272.
Sinclair, A.H.; Berta, P.; Palm er, M.S.; Ross Hawkins, J.; Griffiths, B.L.; Sm ith, M.J.; Foster, J.W.; Frischauf, A.M.; Lovell-Badge, R.; Goodfellow, P.N. 1990. A gene from the hum an sex-determ ining region encodes a protein with hom ology to a conserved DNA-binding mot if. Nature 346: 240-244.
Sm ith, J.D. 1970. The System at ic Status of the Black Howler Monkey, Alouat ta pigra Lawrence. Journal of Mam m alogy 51: 358-369.
Solari, A.J.; Rahn, M.I . 2005. Fine st ructure and m eiot ic behaviour of the male mult iple sex chromosom es in the genus Alouat ta. Cytogenet ic and Genom e Research 108: 262-267.
Solar i, A.J.; Pigozzi, M.I . 1994. Fine st ructure of the XY body in the XY1Y2 t r ivalent of the bat Art ibeus lituratus. Chrom osom e Research 2: 53-58.
Solar i, A.J. 1993. Sex chrom osom es and sex determ inat ion in vertebrates. CRC Press. 308 pags.
Solar i, A.J. 2004. Genét ica hum ana: fundam entos y aplicaciones en m edicina. Ed. Médica Panam ericana, Agosto.
Stanyon, R.; Tofanelli, S.; Morescalchi, M.A.; Agoram oorthy, G.; Ryder, O.A.; Wienberg, J. 1995. Cytogenet ic analysis shows extensive genom ic rearrangem ents between red howler (Alouat ta seniculus, Linnaeus) subspecies. Am erican Journal of Prim atology 35 (3) : 171-183.
Stanyon, R.; Consigliere, S.; Müller, S.; Morescalchi, A.; Neusser, M.; Wienberg, J. 2000. Fluorescence in situ hybridizat ion (FI SH) m aps chrom osom al hom ologies between the dusky t it i and squirrel m onkey. Am erican Journal of Prim atology 50: 95-107.
Stanyon, R.; Consigliere, S.; Bigoni, F.; Ferguson-Sm ith, M.; O'Brien, P.C.M.; Wienberg, J. 2001. Reciprocal chrom osom e paint ing between

7. Bibliografía
202
a New World prim ate, the woolly m onkey, and hum ans. Chrom osom e Research 9: 97-106.
Stanyon, R.; Bonvicino, C.R.; Svartm an, M.; Seuánez, H.N. 2003. Chrom osom e paint ing in Callicebus lugens, the species with the lowest diploid num ber (2n= 16) known in pr im ates. Chromosoma 112: 201-206.
Stanyon, R.; Bigoni, F.; Slaby, T.; Muller, S.; Stone, G.; Bonvicino, C.R.; Neusser, M.; Seuánez, H.N. 2004. Mult i-direct ional chrom osom e paint ing m aps hom ologies between species belonging to three genera of New World m onkeys and hum ans. Chrom osom a 113 (6) : 305-315.
Stanyon, R.; Rocchi, M.; Capozzi, O.; Roberto, R.; Misceo, D.; Ventura, M.; Cardone, M.F.; Bigoni, F.; Archidiacono, N. 2008. Pr imate chromosom e evolut ion: ancest ral karyotypes, m arker order and neocent rom eres. Chrom osom e Research 16: 17-39.
Stanyon R.; Garofalo, F.; Steinberg, E.R.; Capozzi, O.; Di Marco, S.; Nieves, M.; Archidiacono, N.; Mudry, M.D. 2011. Chrom osom e paint ing in two genera of South Am erican Monkeys: Species ident ificat ion, Conservat ion, and Managem ent . Cytogenet ic & Genom e Research en prensa.
Steinberg, E.R.; Nieves, M.; Mudry, M.D. 2007. Meiot ic characterizat ion and sex determ inat ion system of neot ropical pr im ates: Bolivian squirrel monkey Saim ir i boliviensis (Pr im ates: Cebidae) . Am erican Journal of Pr im atology 69: 1236-1241.
Steinem ann, M.; Pinsker, W.; Sperlich, D. 1984. Chrom osom e hom ologies within the Drosophila obscura group probed by in situ hybr idizat ion. Chrom osom a 91 (12) : 46-53.
Sumner, A.T. 1972. A simple technique for demonst rat ing cent rom eric heterochromat in. Exper im ental Cell Research 75: 304-6.
Sutou, S.; Mitsui, Y.; Tsuchiya, K. 2001. Sex determ inat ion without the Y Chrom osom e in two Japanese rodents Tokudaia osim ensis osim ensis and Tokudaia osim ensis spp. Mamm alian Genom e 12 (1) : 17-21.
Sut ton, W.S. 1902. On the Morphology of the Chromosom e Group in Brachystola m agna. Biological Bullet in 14: 24-39.
Swofford, D.L. 2002. PAUP* . Phylogenet ic analysis using parsimony ( * and other m ethods) . Version 4. Sinauer Associates, Sunderland, Massachuset ts.
Takehana, Y.; Naruse, K.; Ham aguchi, S.; Sakaizum i, M. 2007. Evolut ion of ZZ/ ZW and XX/ XY sex-determ inat ion systems in the closely related m edaka species, Oryzias hubbsi and O. dancena. Chromosom a 116 (5) : 463-470.
Thorington, R.W. 1977. The Biology and Conservat ion of the Callit r ichidae. Washington D.C.: Sm ithsonian University Press.
Thom pson, J.D.; Higgins, D.G.; Gibson, T.J. 1994. CLUSTAL W: im proving the sensit ivit y of progressive mult iple sequence alignm ent through sequence weight ing, posit ion-specific gap penalt ies and weight m at r ix choice. Nucleic Acids Research 22: 4673 -4680.

7. Bibliografía
203
Tj io, J.H.; Levan, A. 1956. The chrom osom e num ber of m an. Hereditas 42: 1-6.
Torres, O.M.; Leibovici, M. 2001. Caracterización del cariot ipo del m ono aullador colorado Alouat ta seniculus que habita en Colombia. Caldasia 23 (2) : 537-548.
Torres, O.M,; Ram írez, C. 2003. Estudio citogenét ico de Alouat ta palliata (Cebidae) . Caldasia 25 (1) : 193-198.
Traut , W.; Sahara, K.; Marec, F. 2007. Sex Chrom osom es and Sex Determ inat ion in Lepidoptera. Sexual Developm ent 1: 332-346.
Turner, J.M.A., Mahadevaiah, S.K.; Fernandez-Capet illo, O.; Nussenzweig, A.; Xu, X.; Deng, C.X.; Burgoyne, P.S. 2005. Silencing of unsynapsed m eiot ic chrom osom es in the m ouse. Nature Genet ics 37: 41-47.
Uno, Y.; Nishida, C.; Oshim a, Y.; Yokoyam a, S.; Miura, I . ; Matsuda, Y.; Nakam ura, M. 2008. Comparat ive chrom osom e mapping of sex-linked genes and ident if icat ion of sex chrom osom al rearrangem ents in the Japanese wrinkled frog (Rana rugosa, Ranidae) with ZW and XY sex chromosom e systems. Chrom osom e Research 16 (5) : 637-647.
Vassart , M.; Guédant , A.; Vlé, J.C.; Kéravec, J.; Séguéla, A.; Volobouev, V.T. 1996. Chromosom es of Alouat ta seniculus (Platyrrhini, Prim ates) From French Guiana. Journal of Heredity 87: 331 -334.
Veit ia, R.A.; Salas-Cortés, L.; Ot tolenghi, C.; Pailhoux, E.; Cot inot , C.; Fellous, M. 2001. Test is determ inat ion in mamm als: m ore quest ions than answers. Molecular and Cellular Endocrinology 179: 3-16.
Venter, J.C.; Adam s, M.D.; Myers, E.W.; Li, P.W.; Mural, R.J.; Sut ton, G.G.; Sm ith, H.O. et al. 2001. The Sequence of the Hum an Genom e. Science 291: 1304 -1351.
Verhulst , E.C.; van de Zande, L.; Beukeboom , L.W. 2010. I nsect sex determ inat ion: it all evolves around t ransform er. Current Opinion in Genet ics & Developm ent 20: 376-383.
Veyrunes, F.; Watson, J.; Robinson, T.J.; Brit ton-Davidian, J. 2007. Accum ulat ion of rare sex chrom osom e rearrangem ents in the Afr ican pygm y m ouse, Mus (Nannom ys) m inutoides: a whole-arm reciprocal t ranslocat ion (WART) involving an X-autosom e fusion. Chrom osom e Research 15: 223-230.
Viegas-Péquignot , E.; Benazzou, T.; Dut r illaux, B.; Pet ter, F. 1982. Com plex evolut ion of sex chrom osom es in Gerbillidae (Rodent ia) . Cytogenet ic and Genom e Research 34: 158-167.
Villalobos, F.; Valer io, A.A.; Retana, A.P. 2004. A phylogeny of howler monkeys (Cebidae: Alouat ta) based on m itochondrial, chrom osom al and morphological data. Revista De Biología Tropical 52: 665-677.
Vyskot , B.; Hobza, R. 2004. Gender in plants: sex chrom osom es are em erging from the fog. Trends in Genet ics 20: 432-438.
Wallis, M.C.; Waters, P.D.; Delbridge, M.L.; Kirby, P.J.; Pask, A.J.; Grützner, F.; Rens, W.; Ferguson-Sm ith, M.A.; Graves, J.A.M. 2008. Sex determ inat ion in platypus and echidna: autosom al locat ion of

7. Bibliografía
204
SOX3 confirm s the absence of SRY from monot rem es. Chrom osom e Research 15 (1) : 949-959.
White, M.J.D. 1977. Anim al cytology and evolut ion. Cam bridge University Press. 468 pags.
Wienberg, J. 2005. Fluorescence in situ hybridizat ion to chrom osom es as a tool to understand hum an and prim ate genom e evolut ion. Cytogenet ic and Genom e Research 108: 139-60.
Wienberg, J. 2004. The evolut ion of euther ian chromosom es. Current Opinion in Genet ics & Developm ent 14: 657-666.
Wienberg, J.; Jauch, A.; Stanyon, R.; Crem er, T. 1990. Molecular cytotaxonom y of pr im ates by chrom osom al in situ suppression hybridizat ion. Genom ics 8: 347-350.
Wienberg, J.; Stanyon, R. 1995. Chromosom e paint ing in m am m als as an approach to com parat ive genom ics. Current Opinion in Genet ics & Developm ent 5: 792-797.
———. 1997. Comparat ive paint ing of m am m alian chrom osom es. Current Opinion in Genet ics & Developm ent 7: 784-791.
Wolfheim , J.H. 1983. Prim ates of the World: Dist r ibut ion, Abundance and Conservat ion. Rout ledge.
Yi, S.; Charlesworth, B. 2000. Contrast ing Pat terns of Molecular Evolut ion of the Genes on the New and Old Sex Chromosom es of Drosophila m iranda. Molecular Biology and Evolut ion 17: 703-717.
Yoshido, A.; Yam ada, Y.; Sahara, K. 2006. The W Chrom osom e Detect ion in Several Lepidopteran Species by Genomic in situ Hybridizat ion (GI SH) . Journal of I nsect Biotechnology and Sericology 75: 147-151.
Yu, Q.; Navajas-Pérez, R.; Tong, E.; Robertson, J.; Moore, P.H.; Paterson, A.H.; Ming, R. 2008. Recent Origin of Dioecious and Gynodioecious Y Chrom osom es in Papaya. Tropical Plant Biology 1: 49-57.
Yunis, E.J.; Torres De Caballero, O.M.; Ramírez, C.; Ramírez, E.Z. 1976. Chromosomal Variat ions in the Prim ate Alouat ta seniculus seniculus. Folia Prim atologica 25: 215-224.
Yunis, J.J.; Prakash, O. 1982. The or igin of m an: a chrom osom al pictorial legacy. Science 215: 1525 -1530.
Zunino, G.E.; González, V.; Kowalewski, M.M.; Bravo, S.P. 2001. Alouat ta caraya. Relat ions am ong habitat , density and social organizat ion. Prim ate Report 61: 37-46.

ANEXOS
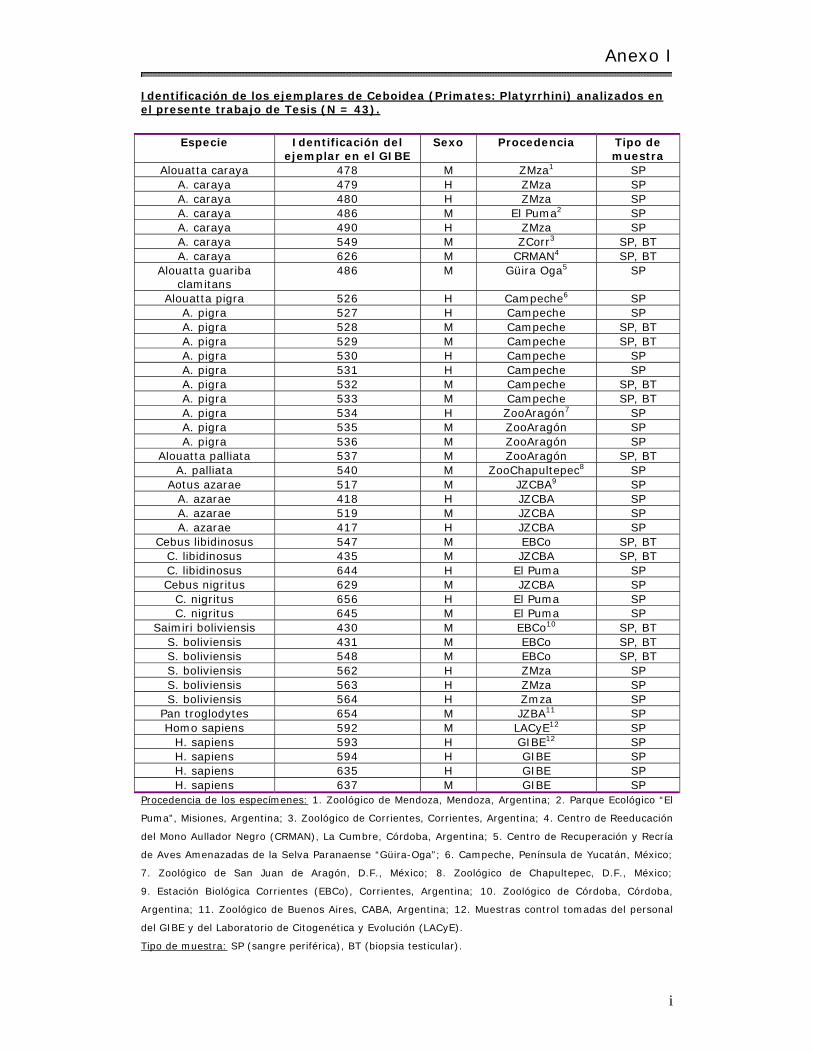
Anexo I
i
I dent if icación de los e jem plares de Ceboidea ( Pr im ates: Pla tyr rhin i) ana lizados en e l presente t raba jo de Tesis ( N = 4 3 ) .
Especie I dent if icación del e j em plar en e l GI BE
Sexo Procedencia Tipo de m uest ra
Alouat ta caraya 478 M ZMza1 SP A. caraya 479 H ZMza SP A. caraya 480 H ZMza SP A. caraya 486 M El Pum a2 SP A. caraya 490 H ZMza SP A. caraya 549 M ZCorr3 SP, BT A. caraya 626 M CRMAN4 SP, BT
Alouat ta guariba clam itans
486 M Güira Oga5 SP
Alouat ta pigra 526 H Cam peche6 SP A. pigra 527 H Cam peche SP A. pigra 528 M Cam peche SP, BT A. pigra 529 M Cam peche SP, BT A. pigra 530 H Cam peche SP A. pigra 531 H Cam peche SP A. pigra 532 M Cam peche SP, BT A. pigra 533 M Cam peche SP, BT A. pigra 534 H ZooAragón7 SP A. pigra 535 M ZooAragón SP A. pigra 536 M ZooAragón SP
Alouat ta palliata 537 M ZooAragón SP, BT A. palliata 540 M ZooChapultepec8 SP
Aotus azarae 517 M JZCBA9 SP A. azarae 418 H JZCBA SP A. azarae 519 M JZCBA SP A. azarae 417 H JZCBA SP
Cebus libidinosus 547 M EBCo SP, BT C. libidinosus 435 M JZCBA SP, BT C. libidinosus 644 H El Pum a SP Cebus nigr itus 629 M JZCBA SP
C. nigr itus 656 H El Pum a SP C. nigr itus 645 M El Pum a SP
Saim ir i boliviensis 430 M EBCo10 SP, BT S. boliviensis 431 M EBCo SP, BT S. boliviensis 548 M EBCo SP, BT S. boliviensis 562 H ZMza SP S. boliviensis 563 H ZMza SP S. boliviensis 564 H Zm za SP
Pan t roglodytes 654 M JZBA11 SP Hom o sapiens 592 M LACyE12 SP
H. sapiens 593 H GI BE12 SP H. sapiens 594 H GI BE SP H. sapiens 635 H GI BE SP H. sapiens 637 M GI BE SP
Procedencia de los especím enes: 1. Zoológico de Mendoza, Mendoza, Argent ina; 2. Parque Ecológico “El
Pum a” , Misiones, Argent ina; 3. Zoológico de Corr ientes, Corrientes, Argent ina; 4. Cent ro de Reeducación
del Mono Aullador Negro (CRMAN), La Cum bre, Córdoba, Argent ina; 5. Cent ro de Recuperación y Recr ía
de Aves Am enazadas de la Selva Paranaense “Güira-Oga” ; 6. Cam peche, Península de Yucatán, México;
7. Zoológico de San Juan de Aragón, D.F., México; 8. Zoológico de Chapultepec, D.F., México;
9. Estación Biológica Corrientes (EBCo) , Corr ientes, Argent ina; 10. Zoológico de Córdoba, Córdoba,
Argent ina; 11. Zoológico de Buenos Aires, CABA, Argent ina; 12. Muest ras cont rol tom adas del personal
del GI BE y del Laborator io de Citogenét ica y Evolución (LACyE) .
Tipo de m uest ra: SP (sangre perifér ica) , BT (biopsia test icular) .

Anexo I I
ii
Soluciones de t rabajo para citogenét ica clásica
3 .2 .1 .2 Cult ivo de Sangre Perifér ica
Solución de cult ivo
En cada frasco de cult ivo se m ezclaron los siguientes react ivos en las
proporciones indicadas:
-7,5 m l m edio F10 (Gibco BRL)
-1,5 m l Suero Fetal Bovino (Bioser)
-0,2 m l ant ibiót ico (Penicilina-Est reptom icina – Gibco BRL 5000 U/ m l)
-0,2 m l Fitohem aglut inina (Gibco BRL)
Solución hipotónica
Cloruro de Potasio (KCl, Merck) 0,075 M
Fijador
Se ut ilizó como fij ador la solución de Carnoy, com puesta por Metanol
(Merck) y Ácido acét ico glacial (Merck) en proporción 3: 1, respect ivam ente.
3 .2 .1 .4 .2 Bandas G con t inción de W right
Soluciones
a) Preparación del Colorante de Wright
1. Poner en agitador 100 m l de m etanol y agregarle lentam ente 0,25 g
de colorante de Wright (Sigm a W3000) . Dejar agitando durante
m edia hora en agitador m agnét ico (DeCaLab S.R.L.) .
2. Filt rar la solución con papel de filt ro (110 mm Ø, Whatm an) .
3. Guardar la solución resultante en una botella color caram elo o
recubierta de papel alum inio para evitar el efecto de la luz directa.
Dejar a 37º C en estufa (San Jor) por 2 a 4 días. Después de ese
t iem po conservar a tem peratura ambiente en frasco recubierto con
papel alum inio por un período de hasta 3 m eses.
b) Solución 2xSSC (Solución Salina Cit rato) :
Para preparar un lit ro de esta solución se m ezclan:

Anexo I I
iii
NaCl (Merck) ... . . . .. . . . .. . . .. . . .. . .. . ... . . .. . . .. . .. . ... .17,53 gr
Cit rato de sodio dihidratado (Merck) .. . . . . ... . . ..8,82 gr
Agua bidest ilada... . . . . . . . .. . . .. . .. . ... . . . . . . .. . .. . ... . . .1,00 l.
c) Solución colorante
Se debe preparar en el m om ento de teñir el preparado.
“Buffer” Sorensen
Solución A: KH2PO4 (Merck) M/ 15.... . . . . . . .. . .4,53 gr/ 500 m l agua bidest ilada
Solución B: Na2HPO4 (Merck) M/ 15.... . . . .. . . .9,47 gr/ 500 m l agua bidest ilada
La solución de t rabajo se prepara con un 60% de la solución A y un 30% de
la solución B.
Para arm ar la solución de t inción se m ezclan 1,5 m l de “buffer” Sorensen y
0,75 m l de colorante de Wright (Ver 3.2.4.1) .
3 .2 .1 .4 .3 Bandas G con t r ipsina
Soluciones
a) Tripsina
Solución Madre: se prepara una solución de t r ipsina al 1% en agua
bidest ilada a part ir de la presentación com ercial en polvo (Difco 1: 250) .
Esta solución se agita durante 2 horas en agitador m agnét ico (DeCaLab
S.R.L.) . Se filt ra con papel de filt ro (110 m m Ø, Whatman) . Se conserva
en freezer, fraccionado en alícuotas de 3 o 5 m l por no m ás de dos años.
Solución de Tr ipsina a l 0 ,1 %
i. Tr ipsina (Difco 1: 250) 1% .... . . . ... . . . . . . .. . .. . .. 1,00 m l
ii. Na2HPO2 (Merck) 0,2N……………………………….. 4,5 m l
iii. NaCl (Merck) 0,9% en agua dest ilada.... . . .. 4,5 m l
Solución de t raba jo : Tripsina 0,05% en PBS sin Ca2+ y Mg2+
Para preparar la t r ipsina 0,05% la m ism a se diluyó a part ir de la solución
al 0,1% .

Anexo I I
iv
PBS (sin Ca2+ ni Mg2+ ) :
NaCl (Merck)………………………………………………8,00 gr
KCl (Merck)………………………………………………..0,20 gr
Na2HPO4 (Merck,anhidro) * ……………...………1,15 gr
KH2PO4 (Merck) ... . . . ... . . .. . . .. . . . . ... . . .. . . .. . .. .0,20 gr
Agua bidest ilada.... . . .. . . .. . . .. . .. . ... . . .. . . .. . .. . .1,00 lt r
* Na2HPO4.H2O = 1,3 gr ; Na2HPO4.7H2O= 2,15 gr ; Na2HPO4.12H2O= 2,9 gr
Filt rar por filt ro de 0,2 µm (STERI VET-Millipore) .
Cont rolar la esterilidad incubando una alícuota 72 horas en estufa (San Jor)
a 37º C.
3 .2 .1 .4 .4 Bandas C ( Sum ner, 1 9 7 2 ) .
a) HCl 0,2 N: 1,6 m l de ácido clorhídrico (HCl, Merck) en 100 m l de agua
dest ilada.
b) Solución de Ba(OH) 2 (Merck) al 5% en agua bidest ilada (preparar
siem pre en el día) : 2,5 gr en 50 m l (queda sobresaturado) .
c) Solución 2xSSC:
NaCl (Merck) ... . . . .. . . . .. . . .. . . .. . .. . ... . . .. . . .. . .. . ... .17,53 gr
Cit rato de sodio dihidratado (Merck) ... . . .. . .. . . .8,82 gr
Agua bidest ilada... . . . .. . . .. . . .. . .. . ... . . . . . . .. . .. . ... . .1,00 lt rs
d) Coloración Giemsa 2% en “buffer” fosfato Sorensen M/ 15 ( pH 7,1)
Sol. A: KH2PO4 (Merck) M/ 15... . . . .. . . .. . .. . .4,53 gr/ 500 m l agua bidest ilada
Sol. B: Na2HPO4 (Merck) M/ 15... . . . .. . . .. . .. .9,47 gr/ 500 m l agua bidest ilada
Se prepara la solución “buffer" m ezclando 70 m l de la solución A y 30 m l de
la solución B.

Anexo I I
v
3 .2 .1 .4 .5 Bandas NOR ( “Nucleolar Organizer Regions” ) o Bandas
de Regiones Organizadoras Nucleolares.
-Solución de nit rato de plata al 50%
Agua bidest ilada…………………….. 500,00 μl
Nit rato de plata (Sigm a)……………. 0,25 gr
Preparar alícuotas en tubos cónicos de 1 m l (Eppendorf) . Envolver el tubo
cónico con un papel de alum inio para evitar la exposición a la luz.
-Solución de gelat ina al 1%
Gelat ina (Sigm a G2500)…………...... . . .. . .. .0,1 gr
Agua bidest ilada………………………………….. 10,0 μl
Acido fórm ico (Merck) * 100% ..... . . . . . .. . .. .50,0 μl
* Tam bién se puede ut ilizar ácido fórm ico al 85% , en cuyo caso se ut ilizan 54 μl.
1. Disolver la gelat ina en agua a 60-70º C en baño térm ico (Vicking) .
2. Dejar enfr iar y agregar el ácido fórm ico (Merck) . Guardar en frasco color
caram elo.

Anexo I I I
vi
Soluciones de t raba jo e infraest ructura para estudi os
citom oleculares
1 . Soluciones de t raba jo
3 .3 .1 .1 H ibr idación “ in situ” Fluorescente ( FI SH)
3 .3 .1 .1 .4 .1 .1 Desnatura lización por separado con Fo rm am ida
-Form am ida al 70% :
28 m l de Form amida (Carlo Erba) + 12 m l de 2xSSC
3 .3 .1 .1 .4 .2 Desnatura lización con Hidróx ido de sodi o ( NaOH)
-NaOH 5M
Disolver 2 gr de NaOH (Merck) en 10 m l de agua m iliQ
-NaOH 100m M (Volum en final 20 m l)
NaOH 5M.... . . . . . .. . . .. . .. . ... . . . . . . .. . . . . ... . . .. . . . . . .. . ..0,4 m l
Etanol 100% ..... . . . . . .. . ... . . .. . . .. . . . . ... . . .. . . .. . .. . .14,0 m l
Agua m iliQ……………………………………………………... 5,6 m l
3 .3 .1 .1 .5 Revelado
Enjuague 1: 0,4% 2xSSC/ 0,3% Tween
Para arm ar 90 m l de solución (equivalente al volumen del recipiente “ Joplin”
a ut ilizar) se m ezclaron:
18,00 m l 2xSSC
0, 27 m l Tween 20 (Prom ega)
71, 73 m l Agua dest ilada
Enjuague 2: 2xSSC/ 0,1% Tween
Para arm ar 90 m l de solución (equivalente al volumen del recipiente “ Joplin”
a ut ilizar) se m ezclaron:
90,00 m l 2xSSC
90,00 µl Tween 20 (Prom ega)

Anexo I I I
vii
Solución “Ant ifade” :
1. Diluir 50 m g de p- fenilendiam ida dihidroclorhídico (Sigm a) en 5 m l de
PBS.
2. Llevar a pH= 8 con CO3Na (Merck) .
3. Llevar a 50 m l con glicerol (Merck) .
4. Filt rar con papel de filt ro (110 m m Ø, Whatm an) .
5. Conservar en freezer a -20ºC.
Solución m adre de DAPI (20 ug/ m l, Sigm a) :
Diluir 10 µl de solución com ercial (2 ng/ m l) en 1m l de agua m iliQ.
Solución de t rabajo de DAPI :
Diluir 5 µl de solución m adre de DAPI (20 ug/ m l) en 1 m l de “ant ifade” .
3 .3 .2 .2 I nm unofluorescencia ( I F)
“Buffer” PBTG (0,2% BSA, 0,05% Tween, 0,2% gelat ina en PBS
BSA (Sigm a)……………………………………………………180 µl
Tween 20 (Prom ega)……………………………………… 45 µl
Gelat ina (Sigm a)…………………………………………….180 µl
PBS…………………………………………………………………..90 m l
Solución fijadora
1% de paraform aldehído (Merck) , 10% de borato de sodio (Merck) 50m M y
0,15% de Triton X-100 (Prom ega) en agua (pH= 9.8) .
2 . Confección de los preparados
3 .3 .1 .2 .1 Envejecim iento de los extendidos
Para el envejecim iento de los extendidos se ut ilizaron diferentes
est rategias aplicadas según el éxito obtenido con cada una de ellas, en cada
individuo y/ o en cada especie que es objeto de análisis en este Trabajo de
Tesis.
Se ut ilizaron extendidos:

Anexo I I I
viii
o Envejecidos 1 día en estufa (San Jor) a 45º C e incubados 30 m inutos
en un Koplin con 2xSSC en baño térm ico (Vicking) a 39º C.
o Envejecidos 1 hora en estufa (San Jor) a 45º C e incubados 30
m inutos en un Koplin con 2xSSC en baño térm ico (Vicking) a 39º C.
o Envejecidos 5 días en estufa (San Jor) a 56º C y luego colocados 3
días en estufa (San Jor) a 37º C.
o Envejecidos 2 días en estufa (San Jor) a 56º C y luego colocados 1 día
en estufa (San Jor) a 37º C. El día de la hibridación se los incubó 30
m inutos en un Koplin con 2xSSC en baño térm ico (Vicking) a 39º C.
o Envejecidos 1 hora en estufa (San Jor) a 56º C
o Envejecidos 1 día a temperatura am biente ( se considera 25 º C)
o Envejecidos 7 días en freezer a -20º C.
o Envejecidos 4 días en freezer a -20º C.
o Envejecidos 5 días en freezer a -20º C.
o Envejecidos 2 días en Koplin con alcohol 70% a tem peratura
am biente ( se considera 25 º C) .
o Extendidos sin envejecer, preparados en el día.
3 . Filt ros de f luorescencia insta lados en e l m icroscopio Leica
DMLB ( GI BE – lab. 4 6 – EGE – FCEyN – UBA)
En el listado siguiente se resaltan con negrita y subrayado los fluorocromos ut ilizados en los procedim ientos de FI SH implem entados en el GI BE para los análisis citomoleculares comparat ivos correspondientes a este t rabajo de Tesis Filt ro Leica A • Fluorocrom os visibles m ediante este filt ro: Acr idine blue AMCA (Am inocoum arin) Aniline blue BisAm inophenyl Oxdiazol (BAO) Bisbenzim ide (Hoechst ) Blancophor BA

Anexo I I I
ix
Blancophor SV Calcein blue Calcofluor White standard solut ion Cascade Blue DANS (diam ino-naphtyl sulphonic acid) Dansyl chlor ide DAPI DI PI Dopam ine DPH (diphenyl hextariene) Fast Blue FI F ( form aldehyde induced fluorescence) Fluoro Gold Fluram ( fluorescam ine) Granular Blue Hoechst dye no. 33258 Hoechst dye no. 33342 Hydroxy-4-m ethylcoum arin Maleim ide MPS (m ethyl Green Pyronine st ilbene) Nuclear Yellow Serot in SI TS (st ilbene isothiosulphonic acid) Sulphaflavine Thioflavine TCN Thiolyte (brom obim ane) True Blue Uvitex 2 B Filt ro Leica I 3 • Fluorocrom os visibles m ediante este filt ro: Acridine orange Acridine yellow Ast razone Orange R Auram ine Aurophosphine, Aurophosphine G BODI PY FL Calcein Calcium Green Carboxyfluorescein diacetate C-FDA Coriphosphine O DiO FDA ( fluorescein diacetate) FI TC ( f luorescein isothiocyanate) FI TC/ ethidium brom ide Fluo 3 Phosphine 3 R Rhodamine 123 Filt ro Leica N21

Anexo I I I
x
• Fluorocrom os visibles m ediante este filt ro: Acid fuchsin Acridine red Alizarin com plexion Alizarin red Am inoact inomycin D-AAD Ast razone Brilliant Red 4G Ast razone Red 6B Calcium Orange Eosin B Ethidium brom ide Evans Blue Fast Green FC G Feulgen FI TC/ ethidium brom ide Genacryl Brilliant Red B Haem atoporphyr in Lissam ine- rhodam ine B (RB 200) Magdala Red Merocyanin 540 Nuclear Fast Red Pararosaniline (Feulgen) Phycoerythr in (PE) Propidium iodide Pyronine B Resorufin Rhodamine B Spect rum Orange Thiazin red R TRI TC ( tet ram ethyl rhodam in isothiocyanate) XRI TC Xylene orange

Anexo I V
xi
Mat r ices obtenidas para cada par t ición de datos -Matr iz básica de datos crom osóm icos #Nexus Begi n dat a; di mensi ons nt ax=11 nchar =99; mat r i x
A. guariba guariba 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0
A. guariba clamitans 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0
A. sara 0 0 0 0 0 0 1 0 1 0 0 0 0 0 1 0 0 0 1 0 0 1 1 1 1 1 0 0 1 0 1 儀0 0 0 0 0 0 1 0
A. macconnelli J 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1
A. s.arctoidea 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0
A. caraya 1 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Alouatta belzebul 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. pigra 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. palliata 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lagothrix lagothricha 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cebus libidinosus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. guariba guariba 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 0 0
A. guariba clamitans 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 0
A. sara 0 0 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0
A. macconnelli J 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1
A. s.arctoidea 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0
A. caraya 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Alouatta belzebul ? 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. pigra 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. palliata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lagothrix lagothricha 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cebus libidinosus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. guariba guariba 0 0 0 0 0 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 0
A. guariba clamitans 0 0 0 0 0 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 0
A. sara 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 0
A. macconnelli J 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 0
A. s.arctoidea 0 0 0 0 0 0 0 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 0
A. caraya 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0
Alouatta belzebul 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 ? 1 1 0 1
A. pigra 0 1 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 0 0
A. palliata 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 1 1 0 0 1 0 0
Lagothrix lagothricha 0 0 0 1 0 0 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 0 0
Cebus libidinosus 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 0 1 1 0 0 1 1 0 0
Anexo I V
xii
-Matr iz básica de datos m oleculares #NEXUS BEGI N DATA; di mensi ons nt ax=11 nchar =800; f or mat mi ssi ng=? symbol s=" ABCDEFGHI KLMNOPQRSTUVWXYZ" i nt er l eave dat at ype=DNA gap= - ; mat r i x A. g. guar i ba TACTCCCCGCAAAACACATCCACTAGCAAAAATCATCAACAACTCATTTA A. g. c l ami t ans TACTCCCCGCAAAACACATCCACTAGCAAAAATCATCAACAACTCATTTA A. sar a TACCCCCCGCAAAACACATCCACTAGCAAAAATCATTAACAACTCATTTA A. macconneJ TACCCCCCGCAAAACACATCCACTAGCAAAAATCATTAACAACTCATTCA A. s. ar ct oi dea TACCCCCCGCAAAACACACCCACTAGCAAAAATCATTAACAACTCATTTA A. car aya TACCCCCCGCAAGACACATCCACTAACAAAAATCATTAACAACTCACTCA A. bel zebul TACCCCCCGCAAAACACACCCACTAGCAAAAATCATCAACAACTCATTCA A. pi gr a TACCCCCCGCAAAACTCACCCACTAGCAAAAATCATCAACAATTCATTCA A. pal l i at a TACCCCCCGCAAAACTCACCCACTAGCAAAAATCATCAACAATTCATTCA L. l agot hr i cha CACCCCTCGCAA- ACACACCCACTAGCAAAAATCATTAACAACTCACTCA C. l i bi di nosus CTCTTCCCGCAAAACACATCCACTAATAAAAATTATTAATAACTCACTTT A. g. guar i ba TTGATCTCCCTACACCATCCAACATCTCCGCCTGATGAAATTTCGGCTCA A. g. c l ami t ans TTGATCTCCCTACACCATCCAACATCTCCGCCTGATGAAATTTCGGCTCA A. sar a TTGATCTTCCCACACCATCCAACATCTCCGCCTGATGAAACTTCGGCTCA A. macconneJ TTGATCTCCCCACACCATCCAACATCTCCGCCTGATGAAATTTCGGCTCA A. s. ar ct oi dea TTGACCTTCCCACACCATCCAACATCTCCGCTTGATGAAATTTCGGCTCG A. car aya TTGATCTCCCCACACCATCCAACATTTCCGCCTGATGAAATTTCGGCTCA A. bel zebul TTGACCTTCCTACACCATCCAACATCTCCGCCTGATGAAATTTTGGCTCA A. pi gr a TTGACCTCCCTACACCATCCAACATCTCCGCCTGATGAAATTTCGGCTCA A. pal l i at a TTGACCTCCCTACACCATCCAACATCTCCGCCTGGTGAAATTTCGGCTCA L. l agot hr i cha TTGACCTACCCTCACCATCCAATATTTCTGCTTGATGAAATTTTGGTTCA C. l i bi di nosus TTGACCTCCCTACACCATCCAACATCTCCTCCTGATGAAACTTCGGATCA A. g. guar i ba CTCCTAGGTATTTGCCTAATTATTCAAATCACTACAGGTCTATTCTTAGC A. g. c l ami t ans CTCCTAGGTATTTGCCTAATTATTCAAATCACTACAGGTCTATTCTTAGC A. sar a CTTCTAGGTATTTGCCTAATTATCCAAATCACTACAGGCCTATTCTTAGC A. macconneJ CTCCTAGGTATTTGCCTGATTATCCAAATTACTACAGGTCTATTCTTAGC A. s. ar ct oi dea CTCCTAGGTATCTGCCTGATTATCCAAATCACTACAGGTCTATTCTTAGC A. car aya CTCCTAGGTATTTGCCTAATTATCCAAATCACTACAGGTCTATTCCTAGC A. bel zebul CTCCTAGGTATTTGCCTAATTATTCAAATCACCACAGGTCTATTCTTAGC A. pi gr a CTCCTAGGCATTTGCCTCATTATTCAAATTACTACAGGCCTATTCTTAGC A. pal l i at a CTCCTAGGTATTTGCCTAATTATTCAAATCACTACAGGTCTATTCTTAGC L. l agot hr i cha CTCTTAGGCATTTGTTTAATTATTCAAATCGCCACAGGCCTATTCCTAGC C. l i bi di nosus CTTCTAGGCGCCTGCCTAATAATTCAAATCACCACAGGCCTATTCTTAGC A. g. guar i ba CATACACTATACACCAGACACTTCAACTGCCTTCTCCTCAGTCGCCCACA A. g. c l ami t ans CATACACTATACACCAGACACTTCAACTGCCTTCTCCTCAGTCGCCCACA A. sar a CATACACTACACACCAGACACCTCAACTGCCTTCTCCTCAGTCGCCCACA A. macconneJ CATACACTACACACCAGACACCTCAACTGCCTTCTCCTCAGTCGCCCACA A. s. ar ct oi dea TATGCACTACACACCAGACACCTCAACTGCCTTCTCCTCAGTCGCCCACA A. car aya CATACATTATACACCAGACACTTCAACTGCCTTCTCCTCGGTCGCCCACA A. bel zebul CATACATTATACACCAGACACTTCAACTGCCTTCTCCTCAGTTACCCATA A. pi gr a CATACACTATACACCAGATACTTCAACCGCCTTCTCTTCAGTCGCTCACA A. pal l i at a CATACACTATACACCAGACACTTCAACCGCCTTCTCCTCAGTCGCCCACA L. l agot hr i cha CATACACTATACACCAGACACTTCAACCGCCTTCTCTTCAGTTGCCCATA C. l i bi di nosus AATACACTACACGCCAGACACCTCAACCGCCTTCTCCTCAGTAGCTCACA A. g. guar i ba TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAT A. g. c l ami t ans TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAT A. sar a TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAT A. macconneJ TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAT A. s. ar ct oi dea TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAC A. car aya TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAC A. bel zebul TCACCCGAGACGTCAATTACGGCTGAATAATCCGCTACCTACACGCCAAT A. pi gr a TCACCCGAGACGTCAACTATGGCTGAATAATCCGCTACCTACACGCCAAT A. pal l i at a TCACCCGAGACGTCAACTATGGCTGAATAATCCGCTATCTACATGCCAAC L. l agot hr i cha TTGCCCGAGACGTAAACTACGGATGAATAATCCGCTACCTACACGCCAAC C. l i bi di nosus TCACCCGAGATATCAACTATGGCTGAATAATCCGCCTCCTACACGCCAAT A. g. guar i ba GGCGCCTCCATATTCTTCATCTGCCTCTTCCTTCACATTGGCCGAGGCCT A. g. c l ami t ans GGCGCCTCCATATTCTTCATCTGCCTCTTCCTTCACATTGGCCGAGGCCT A. sar a GGCGCCTCCATATTCTTCATCTGCCTCTTCCTCCACATTGGCCGAGGCTT A. macconneJ GGCGCCTCCATATTCTTCATCTGCCTCTTCCTCCACATTGGCCGAGGCTT A. s. ar ct oi dea GGCGCCTCCATATTTTTCATCTGCCTCTTCCTCCACATTGGCCGAGGCTT A. car aya GGCGCTTCCATATTCTTCATCTGCCTATTCCTCCACATTGGCCGAGGCTT A. bel zebul GGCGCCTCCATATTCTTTATCTGCCTCTTTCTCCACATTGGCCGAGGCTT A. pi gr a GGCGCCTCCATATTCTTTATCTGTCTCTTTCTCCACATTGGCCGAGGCTT A. pal l i at a GGCGCCTCCATATTCTTTATCTGCCTCTTTCTCCACATTGGCCGAGGCTT L. l agot hr i cha GGTGCTTCCATATTCTTTATTTGCCTCTTCCTACACGTCGGCCGAGGCTT C. l i bi di nosus GGTGCCTCCGTATTTTTTGCATGCTTATTCCTCCACATCGGCCGAGGCCT A. g. guar i ba ATATTATGGATCATTCCTTTTTCTGAAGACCTGAAACATCGGTATTATCC A. g. c l ami t ans ATATTATGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. sar a ATATTACGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTTTCC A. macconneJ ATATTATGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC
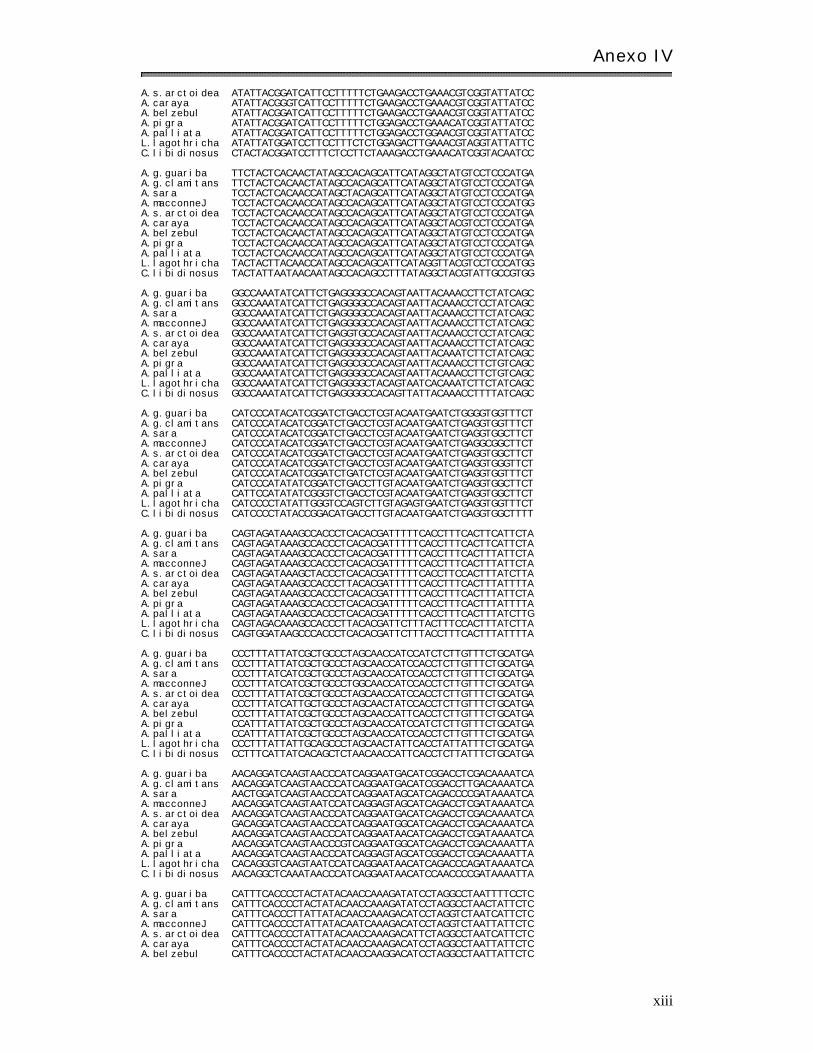
Anexo I V
xiii
A. s. ar ct oi dea ATATTACGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. car aya ATATTACGGGTCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. bel zebul ATATTACGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. pi gr a ATATTACGGATCATTCCTTTTTCTGGAGACCTGAAACATCGGTATTATCC A. pal l i at a ATATTACGGATCATTCCTTTTTCTGGAGACCTGGAACGTCGGTATTATCC L. l agot hr i cha ATATTATGGATCCTTCCTTTCTCTGGAGACTTGAAACGTAGGTATTATTC C. l i bi di nosus CTACTACGGATCCTTTCTCCTTCTAAAGACCTGAAACATCGGTACAATCC A. g. guar i ba TTCTACTCACAACTATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. g. c l ami t ans TTCTACTCACAACTATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. sar a TCCTACTCACAACCATAGCTACAGCATTCATAGGCTATGTCCTCCCATGA A. macconneJ TCCTACTCACAACCATAGCCACAGCATTCATAGGCTATGTCCTCCCATGG A. s. ar ct oi dea TCCTACTCACAACCATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. car aya TCCTACTCACAACCATAGCCACAGCATTCATAGGCTACGTCCTCCCATGA A. bel zebul TCCTACTCACAACTATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. pi gr a TCCTACTCACAACCATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. pal l i at a TCCTACTCACAACCATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA L. l agot hr i cha TACTACTTACAACCATAGCCACAGCATTCATAGGTTACGTCCTCCCATGG C. l i bi di nosus TACTATTAATAACAATAGCCACAGCCTTTATAGGCTACGTATTGCCGTGG A. g. guar i ba GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTATCAGC A. g. c l ami t ans GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTCCTATCAGC A. sar a GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTATCAGC A. macconneJ GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTATCAGC A. s. ar ct oi dea GGCCAAATATCATTCTGAGGTGCCACAGTAATTACAAACCTCCTATCAGC A. car aya GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTATCAGC A. bel zebul GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAATCTTCTATCAGC A. pi gr a GGCCAAATATCATTCTGAGGCGCCACAGTAATTACAAACCTTCTGTCAGC A. pal l i at a GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTGTCAGC L. l agot hr i cha GGCCAAATATCATTCTGAGGGGCTACAGTAATCACAAATCTTCTATCAGC C. l i bi di nosus GGCCAAATATCATTCTGAGGGGCCACAGTTATTACAAACCTTTTATCAGC A. g. guar i ba CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGGGGTGGTTTCT A. g. c l ami t ans CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGTGGTTTCT A. sar a CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGTGGCTTCT A. macconneJ CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGCGGCTTCT A. s. ar ct oi dea CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGTGGCTTCT A. car aya CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGTGGGTTCT A. bel zebul CATCCCATACATCGGATCTGATCTCGTACAATGAATCTGAGGTGGTTTCT A. pi gr a CATCCCATATATCGGATCTGACCTTGTACAATGAATCTGAGGTGGCTTCT A. pal l i at a CATTCCATATATCGGGTCTGACCTCGTACAATGAATCTGAGGTGGCTTCT L. l agot hr i cha CATCCCCTATATTGGGTCCAGTCTTGTAGAGTGAATCTGAGGTGGTTTCT C. l i bi di nosus CATCCCCTATACCGGACATGACCTTGTACAATGAATCTGAGGTGGCTTTT A. g. guar i ba CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTCATTCTA A. g. c l ami t ans CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTCATTCTA A. sar a CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATTCTA A. macconneJ CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATTCTA A. s. ar ct oi dea CAGTAGATAAAGCTACCCTCACACGATTTTTCACCTTCCACTTTATCTTA A. car aya CAGTAGATAAAGCCACCCTTACACGATTTTTCACCTTTCACTTTATTTTA A. bel zebul CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATTCTA A. pi gr a CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATTTTA A. pal l i at a CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATCTTG L. l agot hr i cha CAGTAGACAAAGCCACCCTTACACGATTCTTTACTTTCCACTTTATCTTA C. l i bi di nosus CAGTGGATAAGCCCACCCTCACACGATTCTTTACCTTTCACTTTATTTTA A. g. guar i ba CCCTTTATTATCGCTGCCCTAGCAACCATCCATCTCTTGTTTCTGCATGA A. g. c l ami t ans CCCTTTATTATCGCTGCCCTAGCAACCATCCACCTCTTGTTTCTGCATGA A. sar a CCCTTTATCATCGCTGCCCTAGCAACCATCCACCTCTTGTTTCTGCATGA A. macconneJ CCCTTTATCATCGCTGCCCTGGCAACCATCCACCTCTTGTTTCTGCATGA A. s. ar ct oi dea CCCTTTATTATCGCTGCCCTAGCAACCATCCACCTCTTGTTTCTGCATGA A. car aya CCCTTTATCATTGCTGCCCTAGCAACTATCCACCTCTTGTTTCTGCATGA A. bel zebul CCCTTTATTATCGCTGCCCTAGCAACCATTCACCTCTTGTTTCTGCATGA A. pi gr a CCATTTATTATCGCTGCCCTAGCAACCATCCATCTCTTGTTTCTGCATGA A. pal l i at a CCATTTATTATCGCTGCCCTAGCAACCATCCACCTCTTGTTTCTGCATGA L. l agot hr i cha CCCTTTATTATTGCAGCCCTAGCAACTATTCACCTATTATTTCTGCATGA C. l i bi di nosus CCTTTCATTATCACAGCTCTAACAACCATTCACCTCTTATTTCTGCATGA A. g. guar i ba AACAGGATCAAGTAACCCATCAGGAATGACATCGGACCTCGACAAAATCA A. g. c l ami t ans AACAGGATCAAGTAACCCATCAGGAATGACATCGGACCTTGACAAAATCA A. sar a AACTGGATCAAGTAACCCATCAGGAATAGCATCAGACCCCGATAAAATCA A. macconneJ AACAGGATCAAGTAATCCATCAGGAGTAGCATCAGACCTCGATAAAATCA A. s. ar ct oi dea AACAGGATCAAGTAACCCATCAGGAATGACATCAGACCTCGACAAAATCA A. car aya GACAGGATCAAGTAACCCATCAGGAATGGCATCAGACCTCGACAAAATCA A. bel zebul AACAGGATCAAGTAACCCATCAGGAATAACATCAGACCTCGATAAAATCA A. pi gr a AACAGGATCAAGTAACCCGTCAGGAATGGCATCAGACCTCGACAAAATTA A. pal l i at a AACAGGATCAAGTAACCCATCAGGAGTAGCATCGGACCTCGACAAAATTA L. l agot hr i cha CACAGGGTCAAGTAATCCATCAGGAATAACATCAGACCCAGATAAAATCA C. l i bi di nosus AACAGGCTCAAATAACCCATCAGGAATAACATCCAACCCCGATAAAATTA A. g. guar i ba CATTTCACCCCTACTATACAACCAAAGATATCCTAGGCCTAATTTTCCTC A. g. c l ami t ans CATTTCACCCCTACTATACAACCAAAGATATCCTAGGCCTAACTATTCTC A. sar a CATTTCACCCTTATTATACAACCAAAGACATCCTAGGTCTAATCATTCTC A. macconneJ CATTTCACCCCTATTATACAATCAAAGACATCCTAGGTCTAATTATTCTC A. s. ar ct oi dea CATTTCACCCCTATTATACAACCAAAGACATTCTAGGCCTAATCATTCTC A. car aya CATTTCACCCCTACTATACAACCAAAGACATCCTAGGCCTAATTATTCTC A. bel zebul CATTTCACCCCTACTATACAACCAAGGACATCCTAGGCCTAATTATTCTC
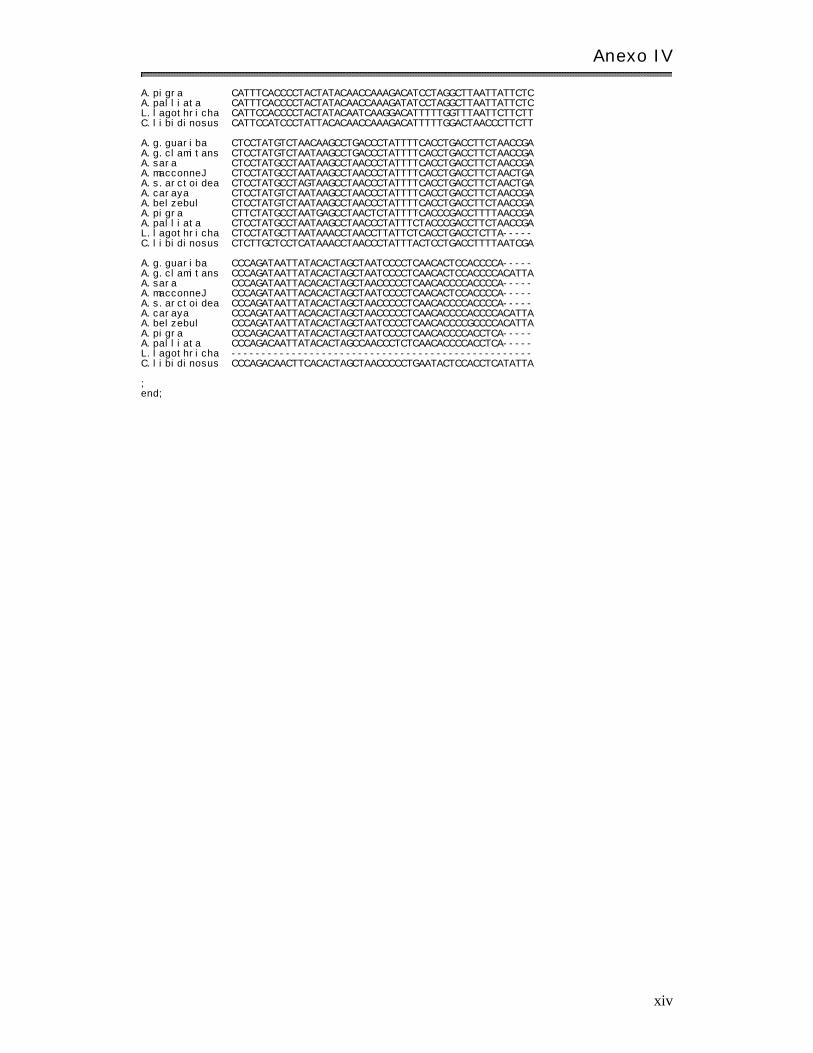
Anexo I V
xiv
A. pi gr a CATTTCACCCCTACTATACAACCAAAGACATCCTAGGCTTAATTATTCTC A. pal l i at a CATTTCACCCCTACTATACAACCAAAGATATCCTAGGCTTAATTATTCTC L. l agot hr i cha CATTCCACCCCTACTATACAATCAAGGACATTTTTGGTTTAATTCTTCTT C. l i bi di nosus CATTCCATCCCTATTACACAACCAAAGACATTTTTGGACTAACCCTTCTT A. g. guar i ba CTCCTATGTCTAACAAGCCTGACCCTATTTTCACCTGACCTTCTAACCGA A. g. c l ami t ans CTCCTATGTCTAATAAGCCTGACCCTATTTTCACCTGACCTTCTAACCGA A. sar a CTCCTATGCCTAATAAGCCTAACCCTATTTTCACCTGACCTTCTAACCGA A. macconneJ CTCCTATGCCTAATAAGCCTAACCCTATTTTCACCTGACCTTCTAACTGA A. s. ar ct oi dea CTCCTATGCCTAGTAAGCCTAACCCTATTTTCACCTGACCTTCTAACTGA A. car aya CTCCTATGTCTAATAAGCCTAACCCTATTTTCACCTGACCTTCTAACCGA A. bel zebul CTCCTATGTCTAATAAGCCTAACCCTATTTTCACCTGACCTTCTAACCGA A. pi gr a CTTCTATGCCTAATGAGCCTAACTCTATTTTCACCCGACCTTTTAACCGA A. pal l i at a CTCCTATGCCTAATAAGCCTAACCCTATTTCTACCCGACCTTCTAACCGA L. l agot hr i cha CTCCTATGCTTAATAAACCTAACCTTATTCTCACCTGACCTCTTA- - - - - C. l i bi di nosus CTCTTGCTCCTCATAAACCTAACCCTATTTACTCCTGACCTTTTAATCGA A. g. guar i ba CCCAGATAATTATACACTAGCTAATCCCCTCAACACTCCACCCCA- - - - - A. g. c l ami t ans CCCAGATAATTATACACTAGCTAATCCCCTCAACACTCCACCCCACATTA A. sar a CCCAGATAATTACACACTAGCTAACCCCCTCAACACCCCACCCCA- - - - - A. macconneJ CCCAGATAATTACACACTAGCTAATCCCCTCAACACTCCACCCCA- - - - - A. s. ar ct oi dea CCCAGATAATTATACACTAGCTAACCCCCTCAACACCCCACCCCA- - - - - A. car aya CCCAGATAATTACACACTAGCTAACCCCCTCAACACCCCACCCCACATTA A. bel zebul CCCAGATAATTATACACTAGCTAATCCCCTCAACACCCCGCCCCACATTA A. pi gr a CCCAGACAATTATACACTAGCTAATCCCCTCAACACCCCACCTCA- - - - - A. pal l i at a CCCAGACAATTATACACTAGCCAACCCTCTCAACACCCCACCTCA- - - - - L. l agot hr i cha - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C. l i bi di nosus CCCAGACAACTTCACACTAGCTAACCCCCTGAATACTCCACCTCATATTA ; end;

Anexo I V
xv
-Matr iz básica de datos com binados #Nexus Begi n dat a; di mensi ons nt ax=11 nchar =899; f or mat dat at ype=st andar d i nt er l eave symbol s=" 01ACGT" gap=- ;
A. guariba guariba 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0
A. guariba clamitans 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0
A. sara 0 0 0 0 0 0 1 0 1 0 0 0 0 0 1 0 0 0 1 0 0 1 1 1 1 1 0 0 1 0 1 0 0 0 0 0 0 0 1 0
A. macconnelli J 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1
A. s.arctoidea 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0
A. caraya 1 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Alouatta belzebul 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. pigra 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. palliata 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lagothrix lagothricha 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cebus libidinosus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. guariba guariba 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 0 0
A. guariba clamitans 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 0
A. sara 0 0 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0
A. macconnelli J 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1
A. s.arctoidea 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0
A. caraya 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Alouatta belzebul ? 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. pigra 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. palliata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lagothrix lagothricha 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cebus libidinosus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
A. g. guar i ba TACTCCCCGCAAAACACATCCACTAGCAAAAATCATCAACAACTCATTTA A. g. c l ami t ans TACTCCCCGCAAAACACATCCACTAGCAAAAATCATCAACAACTCATTTA A. sar a TACCCCCCGCAAAACACATCCACTAGCAAAAATCATTAACAACTCATTTA A. macconneJ TACCCCCCGCAAAACACATCCACTAGCAAAAATCATTAACAACTCATTCA A. s. ar ct oi dea TACCCCCCGCAAAACACACCCACTAGCAAAAATCATTAACAACTCATTTA A. car aya TACCCCCCGCAAGACACATCCACTAACAAAAATCATTAACAACTCACTCA A. bel zebul TACCCCCCGCAAAACACACCCACTAGCAAAAATCATCAACAACTCATTCA A. pi gr a TACCCCCCGCAAAACTCACCCACTAGCAAAAATCATCAACAATTCATTCA A. pal l i at a TACCCCCCGCAAAACTCACCCACTAGCAAAAATCATCAACAATTCATTCA L. l agot hr i cha CACCCCTCGCAA- ACACACCCACTAGCAAAAATCATTAACAACTCACTCA C. l i bi di nosus CTCTTCCCGCAAAACACATCCACTAATAAAAATTATTAATAACTCACTTT
A. guariba guariba 0 0 0 0 0 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 0
A. guariba clamitans 0 0 0 0 0 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 0
A. sara 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1 1 0
A. macconnelli J 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 0
A. s.arctoidea 0 0 0 0 0 0 0 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 0
A. caraya 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0
Alouatta belzebul 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 ? 1 1 0 1
A. pigra 0 1 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 1 1 0 1 1 0 0
A. palliata 0 0 0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 0 1 1 1 1 1 1 1 0 0 1 0 0
Lagothrix lagothricha 0 0 0 1 0 0 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 0 0
Cebus libidinosus 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 0 1 1 0 0 1 1 0 0
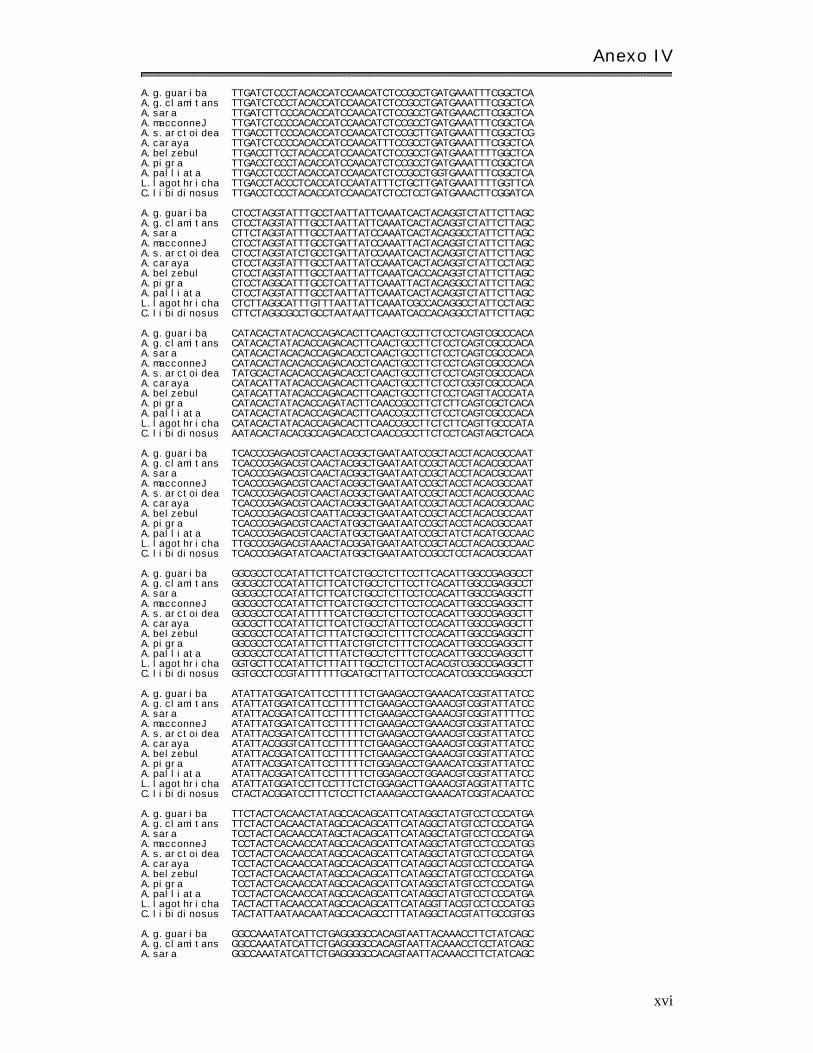
Anexo I V
xvi
A. g. guar i ba TTGATCTCCCTACACCATCCAACATCTCCGCCTGATGAAATTTCGGCTCA A. g. c l ami t ans TTGATCTCCCTACACCATCCAACATCTCCGCCTGATGAAATTTCGGCTCA A. sar a TTGATCTTCCCACACCATCCAACATCTCCGCCTGATGAAACTTCGGCTCA A. macconneJ TTGATCTCCCCACACCATCCAACATCTCCGCCTGATGAAATTTCGGCTCA A. s. ar ct oi dea TTGACCTTCCCACACCATCCAACATCTCCGCTTGATGAAATTTCGGCTCG A. car aya TTGATCTCCCCACACCATCCAACATTTCCGCCTGATGAAATTTCGGCTCA A. bel zebul TTGACCTTCCTACACCATCCAACATCTCCGCCTGATGAAATTTTGGCTCA A. pi gr a TTGACCTCCCTACACCATCCAACATCTCCGCCTGATGAAATTTCGGCTCA A. pal l i at a TTGACCTCCCTACACCATCCAACATCTCCGCCTGGTGAAATTTCGGCTCA L. l agot hr i cha TTGACCTACCCTCACCATCCAATATTTCTGCTTGATGAAATTTTGGTTCA C. l i bi di nosus TTGACCTCCCTACACCATCCAACATCTCCTCCTGATGAAACTTCGGATCA A. g. guar i ba CTCCTAGGTATTTGCCTAATTATTCAAATCACTACAGGTCTATTCTTAGC A. g. c l ami t ans CTCCTAGGTATTTGCCTAATTATTCAAATCACTACAGGTCTATTCTTAGC A. sar a CTTCTAGGTATTTGCCTAATTATCCAAATCACTACAGGCCTATTCTTAGC A. macconneJ CTCCTAGGTATTTGCCTGATTATCCAAATTACTACAGGTCTATTCTTAGC A. s. ar ct oi dea CTCCTAGGTATCTGCCTGATTATCCAAATCACTACAGGTCTATTCTTAGC A. car aya CTCCTAGGTATTTGCCTAATTATCCAAATCACTACAGGTCTATTCCTAGC A. bel zebul CTCCTAGGTATTTGCCTAATTATTCAAATCACCACAGGTCTATTCTTAGC A. pi gr a CTCCTAGGCATTTGCCTCATTATTCAAATTACTACAGGCCTATTCTTAGC A. pal l i at a CTCCTAGGTATTTGCCTAATTATTCAAATCACTACAGGTCTATTCTTAGC L. l agot hr i cha CTCTTAGGCATTTGTTTAATTATTCAAATCGCCACAGGCCTATTCCTAGC C. l i bi di nosus CTTCTAGGCGCCTGCCTAATAATTCAAATCACCACAGGCCTATTCTTAGC A. g. guar i ba CATACACTATACACCAGACACTTCAACTGCCTTCTCCTCAGTCGCCCACA A. g. c l ami t ans CATACACTATACACCAGACACTTCAACTGCCTTCTCCTCAGTCGCCCACA A. sar a CATACACTACACACCAGACACCTCAACTGCCTTCTCCTCAGTCGCCCACA A. macconneJ CATACACTACACACCAGACACCTCAACTGCCTTCTCCTCAGTCGCCCACA A. s. ar ct oi dea TATGCACTACACACCAGACACCTCAACTGCCTTCTCCTCAGTCGCCCACA A. car aya CATACATTATACACCAGACACTTCAACTGCCTTCTCCTCGGTCGCCCACA A. bel zebul CATACATTATACACCAGACACTTCAACTGCCTTCTCCTCAGTTACCCATA A. pi gr a CATACACTATACACCAGATACTTCAACCGCCTTCTCTTCAGTCGCTCACA A. pal l i at a CATACACTATACACCAGACACTTCAACCGCCTTCTCCTCAGTCGCCCACA L. l agot hr i cha CATACACTATACACCAGACACTTCAACCGCCTTCTCTTCAGTTGCCCATA C. l i bi di nosus AATACACTACACGCCAGACACCTCAACCGCCTTCTCCTCAGTAGCTCACA A. g. guar i ba TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAT A. g. c l ami t ans TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAT A. sar a TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAT A. macconneJ TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAT A. s. ar ct oi dea TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAC A. car aya TCACCCGAGACGTCAACTACGGCTGAATAATCCGCTACCTACACGCCAAC A. bel zebul TCACCCGAGACGTCAATTACGGCTGAATAATCCGCTACCTACACGCCAAT A. pi gr a TCACCCGAGACGTCAACTATGGCTGAATAATCCGCTACCTACACGCCAAT A. pal l i at a TCACCCGAGACGTCAACTATGGCTGAATAATCCGCTATCTACATGCCAAC L. l agot hr i cha TTGCCCGAGACGTAAACTACGGATGAATAATCCGCTACCTACACGCCAAC C. l i bi di nosus TCACCCGAGATATCAACTATGGCTGAATAATCCGCCTCCTACACGCCAAT A. g. guar i ba GGCGCCTCCATATTCTTCATCTGCCTCTTCCTTCACATTGGCCGAGGCCT A. g. c l ami t ans GGCGCCTCCATATTCTTCATCTGCCTCTTCCTTCACATTGGCCGAGGCCT A. sar a GGCGCCTCCATATTCTTCATCTGCCTCTTCCTCCACATTGGCCGAGGCTT A. macconneJ GGCGCCTCCATATTCTTCATCTGCCTCTTCCTCCACATTGGCCGAGGCTT A. s. ar ct oi dea GGCGCCTCCATATTTTTCATCTGCCTCTTCCTCCACATTGGCCGAGGCTT A. car aya GGCGCTTCCATATTCTTCATCTGCCTATTCCTCCACATTGGCCGAGGCTT A. bel zebul GGCGCCTCCATATTCTTTATCTGCCTCTTTCTCCACATTGGCCGAGGCTT A. pi gr a GGCGCCTCCATATTCTTTATCTGTCTCTTTCTCCACATTGGCCGAGGCTT A. pal l i at a GGCGCCTCCATATTCTTTATCTGCCTCTTTCTCCACATTGGCCGAGGCTT L. l agot hr i cha GGTGCTTCCATATTCTTTATTTGCCTCTTCCTACACGTCGGCCGAGGCTT C. l i bi di nosus GGTGCCTCCGTATTTTTTGCATGCTTATTCCTCCACATCGGCCGAGGCCT A. g. guar i ba ATATTATGGATCATTCCTTTTTCTGAAGACCTGAAACATCGGTATTATCC A. g. c l ami t ans ATATTATGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. sar a ATATTACGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTTTCC A. macconneJ ATATTATGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. s. ar ct oi dea ATATTACGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. car aya ATATTACGGGTCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. bel zebul ATATTACGGATCATTCCTTTTTCTGAAGACCTGAAACGTCGGTATTATCC A. pi gr a ATATTACGGATCATTCCTTTTTCTGGAGACCTGAAACATCGGTATTATCC A. pal l i at a ATATTACGGATCATTCCTTTTTCTGGAGACCTGGAACGTCGGTATTATCC L. l agot hr i cha ATATTATGGATCCTTCCTTTCTCTGGAGACTTGAAACGTAGGTATTATTC C. l i bi di nosus CTACTACGGATCCTTTCTCCTTCTAAAGACCTGAAACATCGGTACAATCC A. g. guar i ba TTCTACTCACAACTATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. g. c l ami t ans TTCTACTCACAACTATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. sar a TCCTACTCACAACCATAGCTACAGCATTCATAGGCTATGTCCTCCCATGA A. macconneJ TCCTACTCACAACCATAGCCACAGCATTCATAGGCTATGTCCTCCCATGG A. s. ar ct oi dea TCCTACTCACAACCATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. car aya TCCTACTCACAACCATAGCCACAGCATTCATAGGCTACGTCCTCCCATGA A. bel zebul TCCTACTCACAACTATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. pi gr a TCCTACTCACAACCATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA A. pal l i at a TCCTACTCACAACCATAGCCACAGCATTCATAGGCTATGTCCTCCCATGA L. l agot hr i cha TACTACTTACAACCATAGCCACAGCATTCATAGGTTACGTCCTCCCATGG C. l i bi di nosus TACTATTAATAACAATAGCCACAGCCTTTATAGGCTACGTATTGCCGTGG A. g. guar i ba GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTATCAGC A. g. c l ami t ans GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTCCTATCAGC A. sar a GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTATCAGC
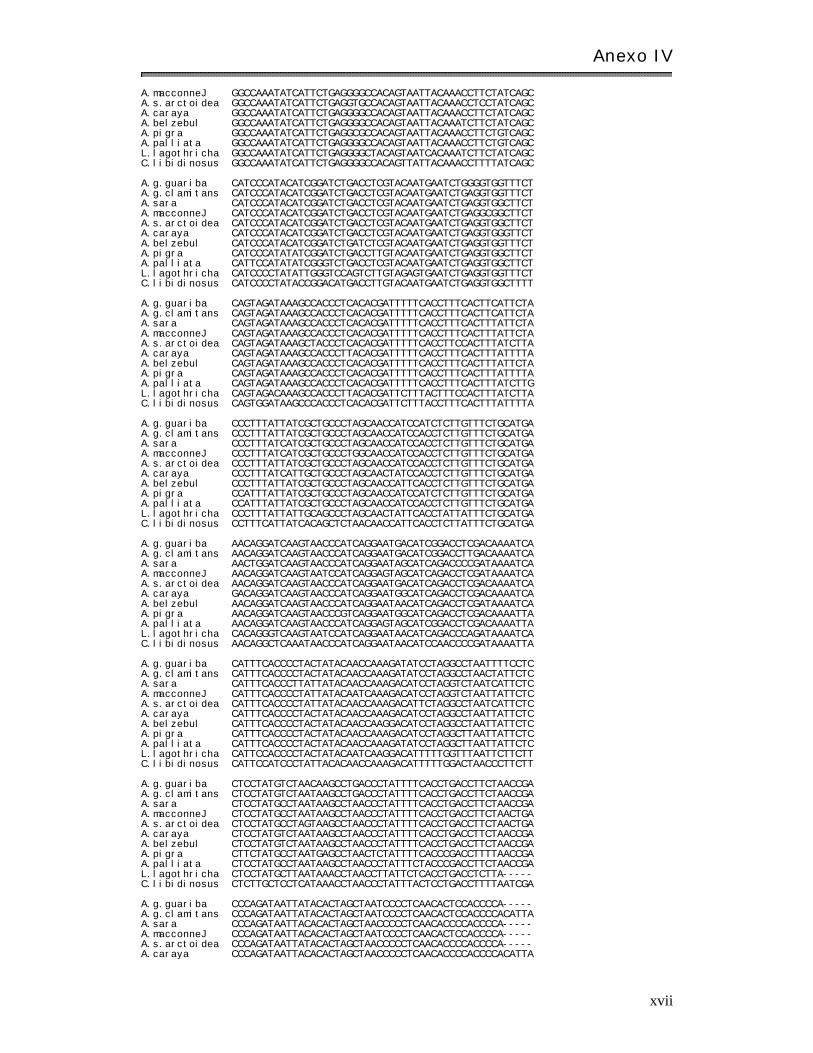
Anexo I V
xvii
A. macconneJ GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTATCAGC A. s. ar ct oi dea GGCCAAATATCATTCTGAGGTGCCACAGTAATTACAAACCTCCTATCAGC A. car aya GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTATCAGC A. bel zebul GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAATCTTCTATCAGC A. pi gr a GGCCAAATATCATTCTGAGGCGCCACAGTAATTACAAACCTTCTGTCAGC A. pal l i at a GGCCAAATATCATTCTGAGGGGCCACAGTAATTACAAACCTTCTGTCAGC L. l agot hr i cha GGCCAAATATCATTCTGAGGGGCTACAGTAATCACAAATCTTCTATCAGC C. l i bi di nosus GGCCAAATATCATTCTGAGGGGCCACAGTTATTACAAACCTTTTATCAGC A. g. guar i ba CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGGGGTGGTTTCT A. g. c l ami t ans CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGTGGTTTCT A. sar a CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGTGGCTTCT A. macconneJ CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGCGGCTTCT A. s. ar ct oi dea CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGTGGCTTCT A. car aya CATCCCATACATCGGATCTGACCTCGTACAATGAATCTGAGGTGGGTTCT A. bel zebul CATCCCATACATCGGATCTGATCTCGTACAATGAATCTGAGGTGGTTTCT A. pi gr a CATCCCATATATCGGATCTGACCTTGTACAATGAATCTGAGGTGGCTTCT A. pal l i at a CATTCCATATATCGGGTCTGACCTCGTACAATGAATCTGAGGTGGCTTCT L. l agot hr i cha CATCCCCTATATTGGGTCCAGTCTTGTAGAGTGAATCTGAGGTGGTTTCT C. l i bi di nosus CATCCCCTATACCGGACATGACCTTGTACAATGAATCTGAGGTGGCTTTT A. g. guar i ba CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTCATTCTA A. g. c l ami t ans CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTCATTCTA A. sar a CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATTCTA A. macconneJ CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATTCTA A. s. ar ct oi dea CAGTAGATAAAGCTACCCTCACACGATTTTTCACCTTCCACTTTATCTTA A. car aya CAGTAGATAAAGCCACCCTTACACGATTTTTCACCTTTCACTTTATTTTA A. bel zebul CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATTCTA A. pi gr a CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATTTTA A. pal l i at a CAGTAGATAAAGCCACCCTCACACGATTTTTCACCTTTCACTTTATCTTG L. l agot hr i cha CAGTAGACAAAGCCACCCTTACACGATTCTTTACTTTCCACTTTATCTTA C. l i bi di nosus CAGTGGATAAGCCCACCCTCACACGATTCTTTACCTTTCACTTTATTTTA A. g. guar i ba CCCTTTATTATCGCTGCCCTAGCAACCATCCATCTCTTGTTTCTGCATGA A. g. c l ami t ans CCCTTTATTATCGCTGCCCTAGCAACCATCCACCTCTTGTTTCTGCATGA A. sar a CCCTTTATCATCGCTGCCCTAGCAACCATCCACCTCTTGTTTCTGCATGA A. macconneJ CCCTTTATCATCGCTGCCCTGGCAACCATCCACCTCTTGTTTCTGCATGA A. s. ar ct oi dea CCCTTTATTATCGCTGCCCTAGCAACCATCCACCTCTTGTTTCTGCATGA A. car aya CCCTTTATCATTGCTGCCCTAGCAACTATCCACCTCTTGTTTCTGCATGA A. bel zebul CCCTTTATTATCGCTGCCCTAGCAACCATTCACCTCTTGTTTCTGCATGA A. pi gr a CCATTTATTATCGCTGCCCTAGCAACCATCCATCTCTTGTTTCTGCATGA A. pal l i at a CCATTTATTATCGCTGCCCTAGCAACCATCCACCTCTTGTTTCTGCATGA L. l agot hr i cha CCCTTTATTATTGCAGCCCTAGCAACTATTCACCTATTATTTCTGCATGA C. l i bi di nosus CCTTTCATTATCACAGCTCTAACAACCATTCACCTCTTATTTCTGCATGA A. g. guar i ba AACAGGATCAAGTAACCCATCAGGAATGACATCGGACCTCGACAAAATCA A. g. c l ami t ans AACAGGATCAAGTAACCCATCAGGAATGACATCGGACCTTGACAAAATCA A. sar a AACTGGATCAAGTAACCCATCAGGAATAGCATCAGACCCCGATAAAATCA A. macconneJ AACAGGATCAAGTAATCCATCAGGAGTAGCATCAGACCTCGATAAAATCA A. s. ar ct oi dea AACAGGATCAAGTAACCCATCAGGAATGACATCAGACCTCGACAAAATCA A. car aya GACAGGATCAAGTAACCCATCAGGAATGGCATCAGACCTCGACAAAATCA A. bel zebul AACAGGATCAAGTAACCCATCAGGAATAACATCAGACCTCGATAAAATCA A. pi gr a AACAGGATCAAGTAACCCGTCAGGAATGGCATCAGACCTCGACAAAATTA A. pal l i at a AACAGGATCAAGTAACCCATCAGGAGTAGCATCGGACCTCGACAAAATTA L. l agot hr i cha CACAGGGTCAAGTAATCCATCAGGAATAACATCAGACCCAGATAAAATCA C. l i bi di nosus AACAGGCTCAAATAACCCATCAGGAATAACATCCAACCCCGATAAAATTA A. g. guar i ba CATTTCACCCCTACTATACAACCAAAGATATCCTAGGCCTAATTTTCCTC A. g. c l ami t ans CATTTCACCCCTACTATACAACCAAAGATATCCTAGGCCTAACTATTCTC A. sar a CATTTCACCCTTATTATACAACCAAAGACATCCTAGGTCTAATCATTCTC A. macconneJ CATTTCACCCCTATTATACAATCAAAGACATCCTAGGTCTAATTATTCTC A. s. ar ct oi dea CATTTCACCCCTATTATACAACCAAAGACATTCTAGGCCTAATCATTCTC A. car aya CATTTCACCCCTACTATACAACCAAAGACATCCTAGGCCTAATTATTCTC A. bel zebul CATTTCACCCCTACTATACAACCAAGGACATCCTAGGCCTAATTATTCTC A. pi gr a CATTTCACCCCTACTATACAACCAAAGACATCCTAGGCTTAATTATTCTC A. pal l i at a CATTTCACCCCTACTATACAACCAAAGATATCCTAGGCTTAATTATTCTC L. l agot hr i cha CATTCCACCCCTACTATACAATCAAGGACATTTTTGGTTTAATTCTTCTT C. l i bi di nosus CATTCCATCCCTATTACACAACCAAAGACATTTTTGGACTAACCCTTCTT A. g. guar i ba CTCCTATGTCTAACAAGCCTGACCCTATTTTCACCTGACCTTCTAACCGA A. g. c l ami t ans CTCCTATGTCTAATAAGCCTGACCCTATTTTCACCTGACCTTCTAACCGA A. sar a CTCCTATGCCTAATAAGCCTAACCCTATTTTCACCTGACCTTCTAACCGA A. macconneJ CTCCTATGCCTAATAAGCCTAACCCTATTTTCACCTGACCTTCTAACTGA A. s. ar ct oi dea CTCCTATGCCTAGTAAGCCTAACCCTATTTTCACCTGACCTTCTAACTGA A. car aya CTCCTATGTCTAATAAGCCTAACCCTATTTTCACCTGACCTTCTAACCGA A. bel zebul CTCCTATGTCTAATAAGCCTAACCCTATTTTCACCTGACCTTCTAACCGA A. pi gr a CTTCTATGCCTAATGAGCCTAACTCTATTTTCACCCGACCTTTTAACCGA A. pal l i at a CTCCTATGCCTAATAAGCCTAACCCTATTTCTACCCGACCTTCTAACCGA L. l agot hr i cha CTCCTATGCTTAATAAACCTAACCTTATTCTCACCTGACCTCTTA- - - - - C. l i bi di nosus CTCTTGCTCCTCATAAACCTAACCCTATTTACTCCTGACCTTTTAATCGA A. g. guar i ba CCCAGATAATTATACACTAGCTAATCCCCTCAACACTCCACCCCA- - - - - A. g. c l ami t ans CCCAGATAATTATACACTAGCTAATCCCCTCAACACTCCACCCCACATTA A. sar a CCCAGATAATTACACACTAGCTAACCCCCTCAACACCCCACCCCA- - - - - A. macconneJ CCCAGATAATTACACACTAGCTAATCCCCTCAACACTCCACCCCA- - - - - A. s. ar ct oi dea CCCAGATAATTATACACTAGCTAACCCCCTCAACACCCCACCCCA- - - - - A. car aya CCCAGATAATTACACACTAGCTAACCCCCTCAACACCCCACCCCACATTA

Anexo I V
xviii
A. bel zebul CCCAGATAATTATACACTAGCTAATCCCCTCAACACCCCGCCCCACATTA A. pi gr a CCCAGACAATTATACACTAGCTAATCCCCTCAACACCCCACCTCA- - - - - A. pal l i at a CCCAGACAATTATACACTAGCCAACCCTCTCAACACCCCACCTCA- - - - - L. l agot hr i cha - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - C. l i bi di nosus CCCAGACAACTTCACACTAGCTAACCCCCTGAATACTCCACCTCATATTA ; end;

Anexo V
xix
Fuente de las fotografías ut ilizadas en las cará tul as I nt roducción Hem bra de Alouat ta caraya con cría (Foto gent ileza de Gabriela Bruno) Objet ivos e Hipótesis Macho de Alouat ta guar iba (modificada de la página Web: ht tp: / / www.arkive.org/ brown-howling-monkey/ alouat ta-guar iba/ im age-G34701.htm l) Mater ia les y Métodos Macho de Alouat ta seniculus seniculus (modificada de la página Web: ht tp: / / www.arkive.org/ colombian- red-howler-m onkey/ alouat ta-seniculus/ im age-G80103.htm l) Resultados Macho de Alouat ta pigra (m odificada de la página Web: ht tp: / / www.arkive.org/ guatem alan-black-howler/ alouat ta-pigra/ image-G19263.htm l) Discusión Macho de Alouat ta palliata (m odificada de la página Web: ht tp: / / www.arkive.org/ m ant led-howler-monkey/ alouat ta-palliata/ im age-G20750.htm l) Conclusiones Ejem plares de Alouat ta belzebul (m odificada de la página Web: ht tp: / / www.arkive.org/ red-handed-howler-monkey/ alouat ta-belzebul/ im age-G63267.htm l) Bibliografía Ejem plar de Alouat ta sara ( tom ada de la página Web: ht tp: / / big.dp.ua/ index.php?page= news&type= 60&p= -2) Anexos Ejem plar de Alouat ta m acconnelli ( tomada de la página Web: ht tp: / / big.dp.ua/ index.php?page= news&type= 60&p= -2)





























